precursors of molecules related to mammalian opioid peptides in brain of a marine worm
TRANSCRIPT

Int. J . Peptide Protein Res. 39, 1992, 300-307
Precursors of molecules related to mammalian opioid peptides in brain of a marine worm
THIERRY DUGIMONT, SANAE GUISSI-KADRI and JEAN-JACQUES CURGY
Laboratory of Invertehrare Eidocriiiologj., CNRS UA 148, C'nirersig. cf Lille-Flrriirlres-Artois, Villeizeuve d'Ascq, France
Received 15 July, accepted for publication 10 November 1991
Total mRNA were extracted from brain of Nereis diversicolor (Aniielida. Polychaeta) and were translated in vitro or in o w . The newly synthesized polypeptides were analyzed through electrophoresis of immunoprecipitated products or the Western blotting technique using polyclonal antibodies raised against mammalian dynorphin 1-17 and mammalian r-neo-endorphin. Among thc products translated in vitro, orzlj, one class of polypeptidc of 70 kDa was recognized by anti-dynorphin 1-17 antibodies. Furthermore, some in ovo translated products as well as proteins extracted from brain of worms showed identical immunoreactivity. These polypeptides, 60-70 kDa, reacted with anti-dynorphin 1 - 17 and anti r-neo-endorphin antibodies. These results suggest the existence of epitopes common to in ow and in r i m translated products, to polypeptides extracted from the brain and to some mammalian opioid peptides of the prodynorphin family. We postulate the presence, in the brain of N. diversicolor, of precursors of peptides related to mammalian dynorphin 1- 17 and a-neo-endorphin. Data reported in this investigation do not allow us to propose or even postulate the presence, in the brain of the worm, of one precursor molecule common to polypeptides related to mammalian dynorphin 1-17 and r-neo-endorphin. Furthermore, the Nereis precursor molecules exhibit a clear-cut difference in molecular mass with the mammalian prodynorphin: 70 kDa versus 30 kDa.
Kej' wordst brain; hormone precursors: marine \vorm; neuropeptides; opioid peptides
Various neuropeptides are present in vertebrate and invertebrate brain and may modulate a wide variety of nervous activities. Most of these neuropeptides appear to be initially synthesized as large precursor proteins which undergo proteolytic processing.
The endogenous opioid system includes three major families of peptides: endorphins, enkephalins and dynorphins (1-3). Molecular genetics has shown that these three families of opioid peptides, present in dif- ferent neuronal pathways within the central nervous system, are derived from three distinct precursors which contain multiple biologically active peptides (4). Pro- opiomelanocortin gives rise to the endorphins as well as to adrenocorticotropic hormone (ACTH) and mel- anotropic hormones (5, 6). Met-enkephalin, Lcu- enkephalin and the related heptapeptide Met-enke- phalin-Arg6-Phe7 and octapeptide Met-enkephalin- Arg6-Gly7-Leu8 are derived from proenkcphalin (7). The third family is derived from prodynorphin and in- cludes dynorphin A, dynorphin B and J - and p-neo- endorphin (8). The amino-acid structure of each of these precursors was deduced from sequenced cDNA clones
300
(9-12). The structure of the genes coding for these precursors is similar, suggesting one common ancestral gene (3, 13).
In mammalian brain, opioid peptides may regulate responses to pain and stress (14), as well as modulate reproductive functions (15) and seizure activity (16).
In insects, studies on the opioid like peptide of Cul- liphom were initiated by the finding that a-endorphin immunoreactive neurons were found in median neuro- secretory cclls (17). Since the latter evidence, the pres- ence of r - , /?-endorphin, enkcphalins and ACTH has been disclosed by imniunocytochemistry and radioim- munoassay ( 1 8). Some of these opioid peptides have been isolated from the central nervous system of Mytilus ediilis (mollusc) (19). Receptors of enkephalins have also been demonstrated in the latter animal (20). Stud- ies including several mollusc species indicate that, as in vertebrates, opioid systems are implicated in the regu- lation and modulation of behavioral and physiological functions, such as stress and feeding (21, 22).
Previous immunocytochemical studies have shown that substances immunologically related to opioids are

Hormone precursors in marine worm
Protein extraction Proteins from Mereis brains and Xenopus oocytes, and from the incubation medium of oocytes were recovered following the procedure described in Guissi-Kadri et al. (32).
synthesized in the brain of the annelid Nereis diversicolor (23) and the reactions were very clear for dynorphin and a-neo-endorphin (24). Some of these opioid pep- tides were also detected by several authors in the cen- tral nervous system of other annelids (25-29). Un- doubtedly, most of these peptides may act as neuromodulators (23). This hypothesis is supported by the results of Golding & May (30), which reveal exo- cytosis of secretory granules in Polychaete neuropiles. Elsewhere, Leu-enkephalin and dynorphin-like immu- noreactivities in Nereis oocytes and spermatocytes (Dhainaut-Courtois et al., unpublished data) indicate a potential role for opioids in reproductive events, as has been shown in the mollusc Helix aspersa (31).
In the present work, we report findings concerning in OIJO and in vitro translated polypeptides specific to mRNA extracted from the brain of M. diversicolor. Some of these polypeptides were recognized by polyclonal antibodies raised against two mammalian opioid pep- tides (dynorphin 1 - 17 and a-neo-endorphin). These results suggest the presence, in worm brain, of precur- sors sharing epitopes homologous with mammalian opioid peptides.
MATERIALS AND METHODS
Animals Worms (Nereis diversicolor, Annelida Polychaeta) were collected in the mouth of the river Aa (Petit-or Grand- Fort-Philippe, Nord, France). Animals were kept in seawater at 4" until used.
Total RNA extraction and RNA treatments Total RNA extraction and RNA treatments were per- formed as previously described (32).
Selection of mRNA by afinity chromatography on oligo (dT)-cellulose This procedure was a modified version of that of Aviv & Leder (33) and Maniatis et al. (34). Details of the procedure are available in Benouareth et al. (35).
mRNA translation systems In ovo mRMA trunslation was carried out following the procedure described in Guissi-Kadri et al. (32).
In vitro mRNA translation. Rabbit reticulocyte 1ysate:The reaction mixture containing nuclease-treated rabbit re- ticulocyte lysate N-90 (Amersham) was used. Experi- mental conditions were as recommended by Amer- sham. Details of the procedure were reported by Benouareth et al. (35).
Wheat germ extract: The reaction mixture containing nuclease-treated wheat germ extract (amino-acid de- pleted, code RPNl, Amersham) was used. Experimen- tal conditions were as recommended by Amersham and as described previously (36).
Antibody preparation and control of specificity Antibodies elaborated against mammalian dynorphin 1-17 and a-neo-endorphin were used. Antisera were raised in the rabbit after injection of a synthetic antigen (Serva, Heidelberg) coupled by glutaraldehyde with human serum albumin, consequently antibodies were confronted to the latter protein before used. Antibod- ies were a generous gift of Professor G. Tramu.
Specificity of antisera used in the present work was controlled by the radioimmunoassay technique (RIA). Cross-reactions between anti a-neo-endorphin and var- ious antigens were as follows: with Met-enkephalins, Leu-enkephalins, dynorphin 1-13, dynorphin €3: 0% ; with a-endorphin: 0.017 % and with P-endorphin: 0.048 %. Cross-reactions between anti-dynorphin 1- 17 antibody and various antigens were as follows: with dynorphin 1 - 18, P-endorphin, Met-enkephalin, Leu- enkephalin, a-endorphin and GnRH: 0%; with a-neo- endorphin: 0.00275%; and with dynorphin B: 0.0041 % (all these data were generously communicated by Dr. D. Croix). Other results obtained with these antibod- ies have already been published (37, 38).
In our experiments, specificity of immunoreactions with polyclonal antibodies was controlled by using blocked antisera. Antibodies were treated as follows: 24 h gentle shaking at 4" of a mixture containing the antiserum and its relevant or non-relevant antigen (anti- dynorphin 1- 17ldynorphin 1- 17 or a-neo-endorphin; anti-a-neo-endorphinla-neo-endorphin or dynorphin 1-17). Proportions were 5 mg antigen/l mL pure an- tiserum. Some batches of preadsorbed antibodies were used immediately, while others were used after a treat- ment using dextran-coated charcoal which removed the excess of non cross-reacting molecules. Indeed, the lat- ter step was performed in order to avoid artefactual inhibition of any immunoreaction when an excess of non-relevant antigen remained in the solution of treated antibodies.
Immunoprecipitation and SDS polyacrylamide gel electrophoresis A 900 pL sample of a diluted mixture containing either the proteins synthesized in ovo or in vitro was mixed with 100 pL of anti-dynorphin 1-17 antibody and 5 mg of protein A-Sepharose CL 4B (Pharmacia). Immune complexes were analyzed by electrophoresis. Urea polyacrylamide gel electrophoresis in SDS was per- formed as described by Burr & Burr (39). Polyacryla- mide gel electrophoresis in SDS was performed as de- scribed by Laemmli (40) using 5-25?' acrylamide gradients gel. After electrophoresis, proteins were stained with Coomassie blue and Crocein Scarlet and
301

T. Dugimont et ai.
the stained gel processed for autoradiography. After developing, the films were scanned with a densitometer (A = 595 nm).
Western blotting technique After electrophoretic separation of the proteins ex- tracted either from oocytes or brains, gels were trans- ferred onto nitrocellulose according to Towbin et al. (41). Nitrocellulose sheets were then incubated with the primary antibody diluted to ljl00 in TBS buffer con- taining 2% BSA overnight at 4'. After washing steps, nitrocellulose sheets were incubated with the second antibody (goat anti-rabbit IgG conjugated to peroxi- dase, Diagnostic Pasteur), diluted 1/2000 in TBS buffer containing 0.5% dry milk for 2 h/ Peroxidase activity was visualized with the following mixture: 50 mL TBS, 3 mL of 3% 4-chloro-1-naphthol in cold methanol, 10 pL 30% H202. The reaction was stopped with bi- distilled water and nitrocellulose sheets were stocked in the dark.
Enzyme-linked irnmunosorbent assq* (ELISA) The 96-well plates were coated with 50 pL of an oocyte homogenate [ oocytes injected by distilled water (con- trols) or by brain mRNA solution] and incubated over- night at 4". The plates were saturated by TBS: 50 mM Tris pH 7.4, 150 mM NaC1, 0.29; BSA. The plates were washed with TBS, 0.1 % BSA. The first antibody (anti-dynorphin 1- 17 antiserum, anti r-neo-endorphin antiserum, or antisera preadsorbed by the relevant or non-relevant antigen) diluted at lj200 was added at 50 pL/well and incubated at 37' for 2 h. After washing steps with TBS buffer 0.101~ BSA, the plates were in- cubated with the secondary antibody (goat anti-rabbit IgG conjugated to peroxidase, Diagnostic Pasteur). The peroxidase reaction was developed using orthophe- nylene diamine (Sigma USA) as substrate. The reading of optical density of each well at 490 nm was done using a microplate reader MR 5000 Dynatech.
RESULTS
In vitro translation of nzRNA
Electrophoretic analysis of total in vitro translated prod- ucts. Rabbit reticulocyte lysate cell-free system used for in vitro translation: Electrophoretic analysis in SDS- polyacrylamide gcl, using 5-25 0; acrylamide gradient, of products obtained from the cell-free translation of total brain mRNA, revealed numerous newly synthe- sized proteins with a molecular mass between 15 and 120 kDa (graph E, Fig. 1).
The control experiment, corresponding to the addi- tion to the reticulocyte lysate of distilled water with- out mRNA, gave electrophoretic data shown in Fig. 1, graph A. No well defined polypeptide was rec- ognizable.
302
Wheat germ extract cell-free system used for in vitro translation: Electrophoretic analysis under the same conditions as those reported in the above paragraph, of polypeptides translated on total brain mRNA added to the wheat germ extract displayed several products with a molecular mass between 12 and 140 kDa (graph F, Fig. 1).
No obvious translatcd molecules appeared in the control experiment, which was performed by addition of distilled water without mRNA in the wheat germ extract (graph B, Fig. 1).
Electrophoretic analysis of imrnunoprecipitates obtained .from CI reaction between mammalian dynorphin 1-1 7 an- tibodies and in vitro translated products. Translation in rabbit reticulocyte lysate cell-free system: In vitro trans- lated polypeptides were immunoprecipitated by anti- dynorphin 1-17 antibodies and analyzed by SDS- polycrylamide gel electrophoresis. The electrophoretic pattern revealed only one class of polypeptides with a 70 kDa molecular mass, reacting with anti-dynorphin 1-17 antibodies (arrow, graph G, Fig. 1). Graph C in Fig. 1 shows the absence of an immune reaction in the control experiment (addition of distilled water to rab- bit reticulocyte lysate).
Translation in wheat germ extract cell-free system: Immunodetection similar to that described in the above paragraph 2 was performed with polypeptides trans- lated in wheat germ extract. Anti-dynorphin 1-17 an- tibodies reacted with only one class of products of 70 kDa (arrow, graph H, Fig. 1). Graph D in Fig. 1 shows the control experiment obtained after the addi- tion of distilled water without mRNA to the wheat germ extract. The absence of the immune reaction was sim- ilar to that reported in graph C, Fig. 1.
In ovo translation of mRNA
Eiq*me irnrnunosorbent assay (ELISA) results. 96-well plates were coated with brain-mRNA-injected oocyte homogenate or with distilled water oocyte homogenate (controls). Oocyte homogenates were used at different dilutions ( l / lO, ljl00, ljl000, l/lOOOO). Incubations were performed with all batches of antisera: pread- sorbed (or not) with relevant or non relevant antigens, with (or without) elimination of excess antigens by using dextran-coated charcoal. From ELISA results, it was noted: 1) that control oocyte homogenates were signif- icantly less immunoreactive than oocytes injected by brain mRNA. These results express genuine translation of mRNA of interest in Xenopus oocytes; 2) that an excess of non-relevant antigen could reduce the im- mune reaction, but when this excess was removed by dextran-coated charcoal treatment, immunoreactivity was recovered.
These results were confirmed by the following exper- iments: immunoprecipitations and Western Blotting techniques using the mentioned antibodies.

Hormone precursors in marine worm
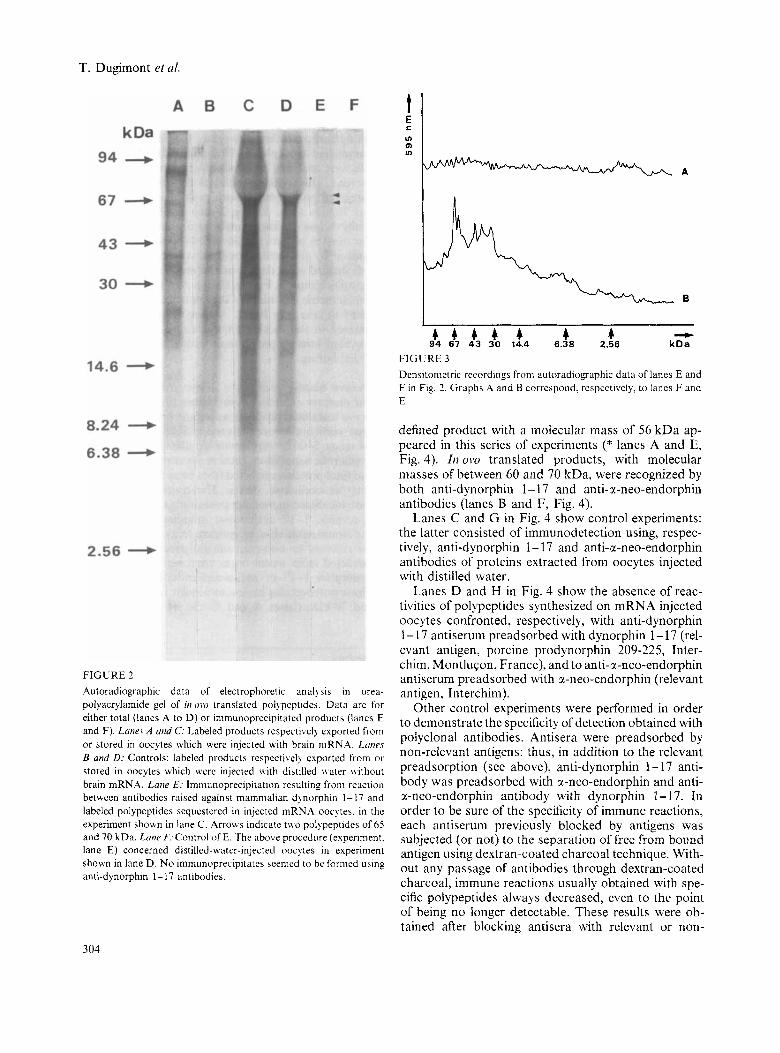
Electrophoretic analysis of irnmunoprecipitates obtained after reaction between anti-dynorphin I - I 7 antibodies and in ovo translated polypeptides. Immune complexes, re- sultingfrom areaction between antibodies raised against mammalian dynorphin 1-17 and in ovo translated prod- ucts were analyzed by urea-polyacrylamide gel electro- phoresis. This technique showed evidence of two major translated polypeptides, with molecular masses of 65 and 70 kDa (lane E, Fig. 2 and graph B, Fig. 3). A family of less obvious translated products with molec- ular masses of between 30 and 44 kDa was also rec- ognized by the antibodies used; these products were well defined after densitometric recording of autorad- iography (graph B, Fig. 3). Graph A in Fig. 3 shows the densitometric recording of the control experiment illus- trated in lane F in Fig. 2. Among translated products released in the incubation medium of oocytes, none provided immunoprecipitates with anti-dynorphin 1 - 17 antibodies.
Electrophoretic analysis of in ovo translated products. 35S-methionine-labeled polypeptides, in ovo translated, were analyzed by urea-polycrylamide gel electrophore- sis, followed by autoradiography (Fig. 2). Several prod- ucts, with molecular masses of between 4.5 and 95 kDa, were detected both in Xenopus oocytes and in the in- cubation medium (lanes A to D, Fig. 2). Comparisons between electrophoretic patterns obtained from poly A + -mRNA injected oocytes and distilled-water- injected oocytes (control) indicated that, in the mRNA recipient cells, as in their incubation medium, proteins specific to worm brain mRNA were recovered (com- pare lanes A and B, C and D, Fig. 2).
t E C v) ul v)
t E C v) ul v)
4 4 4 4 4 4 -
- kDa
4 4 4 4 4 4 94 67 43 30 20 14.4
Western blotting analysis of in ovo translated products and proteins extracted from Nereis diversicolor brain. In ovo translated products as well as proteins extracted from worm brain were analyzed by SDS-polyacrylamide gel using 5-25% acrylamide gradient. After the elec- trophoretic separation, total in ovo translated products and total proteins extracted from brain were transferred onto nitrocellulose sheets. The transferred products were incubated in medium containing mammalian anti- dynorphin 1 - 17 antibodies, mammalian anti-sr-neo- endorphin antibodies, or immune sera preadsorbed with the relevant or non-relevant antigens. Some proteins extracted from worm brain were recognized by anti- dynorphin 1- 17 or anti-a-neo-endorphin antibodies. These polypeptides have a molecular mass between 60 and 70 kDa (lanes A and E, Fig. 4). Furthermore, a
FIGURE I Graphs A , B , E and F: Densitometric recordings from autoradio- graphic data of electrophoretic analysis in SDS-polyacrylaniide gel of cell-free translated polypeptides in rabbit reticulocyte lysate or in wheat germ extract. Graphs A and B are respectively controls of E and F: incubation medium was added with distilled water. Gruph ,!?:Total products translated in rabbit reticulocyte lysate added with distilled water containing brain poly(A) + -mRNA. Polypeptides of 15 to 120 kDa were synthesized. Graph F: Total products translated in wheat germ extract added with distilled water containing brain mRNA. Polypeptides of 12 to 140 kDa molecular mass were syn- thesized. Graphs C, D, G and H : Densitometric recording of autoradiographic data after electrophoretic separation of immunoprecipitates resulting from a reaction between antibodies raised against mammalian dynor- phin 1-17 and polypeptides translated in rabbit reticulocyte lysate or in wheat germ extract. Graphs C and D are respectively controls of G and H: incubation medium was added with distilled water. Graphs G and H : Experimental conditions are those shown respectively in legend of graphs E and F. Only one class of 70 kDa polypeptides precipitated (arrows).
303
T. Dugimont et ai.
FIGURE 2 Autoradiographic data of electrophoretic analysis in urea- polyacrylamide gel of iir uru translated polypeptides. Data are for either total (lanes A to D) or immunoprecipitated products (lanes E and F). Lanes A and C: Labeled products respectivcl) exported from or stored in oocytes which were injected with brain mRNA. L m e s B and D: Controls: labeled products respectivel? cxported froiii or stored in oocytes which wci-c injected \vith distilled \vater without brain mRNA. Lane Ec Immunoprecipitation resulting from reaction betwccn antibodies raised against matnmalian dlnorphin 1- 17 and labeled polypeptides sequestered in injected mRNA oocytes. in the experiment shown in lane C. Arrows indicate t\vo polypeptides of 65 and 70 kDa. Ltrne F: Control of E. The above procedure (experiment. lane E) concerned distilled-water-injected ooc) tes in experiment shown in lane D. No irnmunoprecipitatcs aeemed to be formed using anti-dynorphin 1 - 17 antibodies.
I - kDa
4 4 4 4 4 4 4 94 67 43 30 14.4 6.38 2.56
FIGURE 3 Densitometric recordings from autoradiographic data of Iancs E and F i n Fig. 2. Graphs A and B correspond, respcctively, to lanes F and E.
defined product with a molecular mass of 56 kDa ap- peared in this series of experiments (* lanes A and E, Fig. 4). Irz ovo translated products, with molecular masses of between 60 and 70 kDa, were recognized by both anti-dynorphin 1- 17 and anti-x-neo-endorphin antibodies (lanes B and F, Fig. 4).
Lanes C and G in Fig. 4 show control experiments: the latter consisted of immunodetection using, respec- tively, anti-dynorphin 1- 17 and anti-a-neo-endorphin antibodies of proteins extracted from oocytes injected with distilled water.
Lanes D and H in Fig. 4 show the absence of reac- tivities of polypeptidcs synthesized on mRNA injected oocytes confronted, respectively, with anti-dynorphin 1- 17 antiserum preadsorbed with dynorphin 1- 17 (rel- evant antigen, porcine prodynorphin 209-225, Inter- chim, MontluCon, France), and to anti-a-neo-endorphin antiserum preadsorbed with a-neo-endorphin (relevant antigen, In terchim).
Other control experiments were performed in order to demonstrate the specificity of detection obtained with polyclonal antibodies. Antisera were preadsorbed by non-relevant antigens: thus, in addition to the relevant preadsorption (see above), anti-dynorphin 1- 17 anti- body was preadsorbed with r-neo-endorphin and anti- x-neo-endorphin antibody with dynorphin 1- 17. In order to be sure of the specificity of immune reactions, each antiserum previously blocked by antigens was subjected (or not) to the separation of free from bound antigen using dextran-coated charcoal technique. With- out any passage of antibodies through dextran-coated charcoal, immune reactions usually obtained with spe- cific polypeptides always decreased, even to the point of being no longer detectable. These results were ob- tained after blocking antisera with relevant or non-
304

Hormone precursors in marine worm
mammalian anti-dynorphin 1- 17 antibodies demon- strated that polypeptides of 70 kDa, translated in both in vitro and in ovo systems, reacted with these antibod- ies. In addition, among worm brain-specific proteins translated only in ow, a 65 kDa species and a block of poorly defined polypeptides with 30-44 kDa, were rcc- ognized by anti-dynorphin 1- 17 antibody.
Western blots performed on polypeptides translated in oocytes, with mammalian anti-dynorphin 1-17 and mammalian anti-a-neo-endorphin antibodies, demon- strated the presence of a block of rather well defined polypeptides with a molecular mass betwcen 60 and 70 kDa. Polypeptides of similar high molecular mass, 60-70 kDa, were detected among proteins extracted from the total worm brain. These data suggest that polypeptides translated on isolated brain mRNA and in cerebral cells have epitopes in common. In other words, these immunological reactions suggest that nereidian 60-70 kDa polypeptides share homologous molecular substructures with the previously mentioned mamma- lian opioid peptides.
The polypeptides excreted by mRNA-injected oocytes failed to exhibit any immunoreactivity with the antibodies used.
Immunocytochemical studies performed with the above mentioned antibodies reported the presence, in the nereidian brain, of molecules related to mammalian dynorphin 1-17 and to mammalian a-neo-endorphin (23, 24). The cytological approach used in these stud- ies revealed a strong immune reaction with both anti- bodies. Bearing in mind the fundamental results of Dhainaut-Courtois’ group, we postulate that polypep- tides with a molecular mass of 60-70 kDa are proba- bly the precursors of molecules related to mammalian dynorphin 1- 17 and a-neo-endorphin.
It should be borne in mind that control experiments carried out by replacing the antisera with the antisera preincubated with relevant antigen with or without pas- sage on dextran-coated charcoal indicated that poly- peptides recognized by the non-blocked antisera were no longer detectable.
Kakidani et al. (1 1) showed that, in mammals, dynor- phin 1- 17 and a-neo-endorphin are opioid peptides which are processed from a common precursor, named prodynorphin or proenkephalin B. In the present work, we report the recognition by each of the antibodies used (anti-dynorphin 1- 17 and anti-a-neo-endorphin) of similar polypeptides by means of their molecular mass; this was verified for polypeptides in ovo translated or directly extracted from the brain. These data are argu- ments in favor of the existence in nereidian worms of large precursors from which some molecules related to peptides of prodynorphin can be processed. Among the in vitro translated products, oniyone class, with a 70 kDa molecular mass, reacted with mammalian anti- dynorphin 1-17 antibodies, even though, among in ovo translated products, others with molecular lower than 70 kDa exhibited immunoreactivity with the antibodies.
305
FIGURE 4 Western blot corresponding to electrophoretic analysis in SDS- polyacrylamide gel of polypeptides stocked in oocytes (lanes B, C, D and F, G, H) and proteins extracted from worm brain (lanes A and E). Lanes A and E: Proteins extracted from worm brain. Immuno- detection with antibodies raised against mammalian dynorphin 1-17 (A) and mammalian cc-neo-endorphin (E) indicates several proteins with a molecular mass of between 60 and 70 kDa (60, 62, 65 and 70 kDa). Lanes B and F: Proteins extracted from mRNA-injected oocytes. lmmunodetection with antibodies raised against mamma- lian dynorphin 1-17 (B) and mammalian a-neo-endorphin (F). Four polypeptides of 60, 62, 65 and 70 kDa reacted with tested antibod- ies. Lanes C and G: Controls of B and F. Proteins extracted from oocytes injected with distilled water. Lane D: Control of immuno- blotting experiment performed with anti-dynorphin 1- 17 preadsorbed with dynorphin 1-17. Lane H: Control of immunoblotting experiment performed with anti-cc-neo-endorphin preadsorbed with a-neo- endorphin.
relevant antigens (data not shown). On the contrary, when blocked antisera were subjected to the action of dextran-coated charcoal, polypeptides with a molecu- lar mass of between 60 and 70 kDa remained undetect- able after relevant saturation of antibodies, but immu- noreactivity was recovered when antibodies were preadsorbed with non-relevant antigens (data not shown).
Experiments involving translated products exported in the incubation medium of oocytes showed no prod- ucts reacting with antibodies.
DISCUSSION AND CONCLUSION
Poly(A) + -mRNA of worm brain were translated in vit- YO or in ovo. Immunoprecipitation of polypeptides using

T. Dugimont et al.
It is known that rabbit reticulocyte lysate and wheat germ extract are acellular systems in which no normal or natural post-translational processing occurs. Thus we infer that the 70 kDa molecular mass polypeptides are the primary elaborated form of precursors which are also found among proteins synthesized itz ovo or in brain cells. Thus, polypeptides with a molecular mass lower than 70 kDa (60, 62, 65 kDa) could originate by a normal maturation process in living cerebral cells, in the large 70 kDa molecules (lanes A and E, Fig. 4). These arguments are in favor of the hypothesis of the existence of large 70 kDa precursors from which some peptides of one opioid family are derived. The matu- ration of these precursors could occur in several steps, giving intermediate molecules as exemplified by mam- malian pancreatic endocrine cells (42). The precursor molecules, which we characterized, exhibit an obvious difference in molecular mass with the mammalian pro- dynorphin: 70 kDa versus 30 kDa. Furthermore, our results do not permit us to propose that in our mate- rial only one precursor molecule is proteolytically ma- tured in order to liberate two peptides related to mam- malian r-neo-endorphin and dynorphin 1- 17.
In other respects, a class of poorly defined inovo translated products (30-44 kDa) precipitated in the presence of anti-dynorphin 1-17 antibodies. We hy- pothesize that these molecules are the consequence of in ovo enzymatic degradation o f higher polypeptides (60-70 kDa). This interpretation is supported by the demonstration of the activity of proteolytic enzymes occurring at specific sites of newly translated polypep- tides in mRNA-injected oocytes (43). The absence of this class of molecular species among cellular extracts from brain lends support to the above hypothesis.
Finally, we postulate that, in a lower invertebrate such as Nereis, there may exist precursors of neuropep- tides with a large molecular mass.
ACKNOWLEDGMENTS
We waould like to thank Professor G. Tramu (Laboratoire de Phys- iologie des Interactions cellulaires, Universite de Bordeaux I. Tal- ence, France) for his generous gift of antibodies. We thank Dr. D. Croix (Laboratoire d’Endocrinologie cellulaire. INSERM U. 156. Lille, France) for critical and helpful comments in the preparation of the manuscript. She generously communicated important infornia- tion about the specificity of aniibodies used in this Lvork. We are indebted to F. Fournier (Laboratoirc dc Phqsiologie Cellulaire, Lille I ) for mRNA injection into Xenopus oocytes. We thank M. Vennin for skillful technical assistance and N. Masse for preparing the nianu- script.
This work was supported by the Conseil Regional Nord Pas-de- Calais (France).
REFERENCES
I . Numa, S. (1984) in The Peprides: 84no!~~si .~, Biolog~.. S>.ruheti\
2. Khachaturian, H., Lewis. M.E., Schafer. M.G.11. & Watson. (Udenfriend, S . & Meienhofer. J.. eds.). vol. 6 , pp. 1-73
S.J. (1984) Trendy Jvertrmci. 7, 111-119
306
3. Patey, G. 8: Rossier, J. (1986) Ann. Endocrinol. 47, 71-87 4. Douglass, J., Civelli, 0. & Herbert, E. (1986)Ann. Rev. Biochem.
53, 665-715 5. Mains, R.E., Eipper, B.A. & Ling, N. (1977) Proc. Natl. Acad.
Sci. C’SA 74, 4 17 1-4 174 6. Noel, G., Zollinger, L., Lariviere, N., Nault, C., Crine, P. &
Boileau. G. (1987) J . B i d . Chem. 262, 1876-1881 7 . Davis, T.P.. Porreca, F., Burks, T.F. & Dray, A. (1985) Euro-
peait J . Pltarmacol. 111, 177-183 8 . Christensson-Nylander, I., Nyberg, F., Ragnarsson, U. & Ter-
emiub, L. (1985) Reg. Peprides 11, 65-76 9. Nakanishi. S., Inoue, A,, Kita, T., Nukamura, M., Chung,
A.C.Y., Cohcn, S.N. & Numa, S. (1979) Nature 278, 423-427 10. Comb, M.. Seeburg, P.H., Adelman, J., Eiden, 1. & Herbert, E.
(1982) Norwe 295, 663-666 1 1 . Kakidani, H., Furutani, Y., Takahashi, H., Noda, M., Morim-
oto, Y., Hirose, T., Asai, M., Inayama, S., Nakanishi, S. & Numa, S. (1982) Nature 298, 245-249
12. Noda, M.. Furutani, Y., Takahashi, H., Toyosato, M., Hirose, T.. Inalama, S. , Nakanishi, S. & Numa, S . (1982) Nature 295, 202-206
13. Rossier, J. (1988) Ann. Endocrinol. 49, 371-373 14. Akil. H., Watson, S.J., Young, E., Lewis, M., Khaehaturian, H.
15, Cicero, T.J. (1980) Fed. Proc. 39, 2551-2554 16. Kananiatsu, T., Mc Ginty, J.F., Mitchell, C.L. & Hong, J.S.
17. Duve, H . & Thorpe, A. (1983) Cell Tissue Res. 233, 415-426 18. Duve, H.. Thorpe, A. & Scott, A.G. (1986) in Comparative As-
peers qf Opioid and Rekited Neuropeptide Mechanisms (Stephano, G.B., ed.). pp. 197-211, CRC Press, Boca Raton, FL
19. Leung, M.K. & Stefano, G.B. (1984) Proc. Nutl. Acad. Sci. USA 81, 955-956
20. Stefano, G.B., Cadet, P. & Scharrer, B. (1989) Proc. Nutl. Acad.
21. Stefano. G.B. (1982) Cell. Mol. Neurobiol. 2, 167-178 22. Kavaliers. M., Hirst, M. & Mathers, A. (1985) Neuroendocrinol.
23. Dhainaut-Courtois, N.. Tramu, G., Beauvillain, J.C. & Masson, M. (1986) Neurochein. In t . 8, 327-338
24. Dhainaut-Courtois, N. & Golding, D.W. (1988) in The Ultra- smrcrure ~f Polrchaera (W’estheide, W. & Hermans, C.O., eds.), vol. 4, pp. 89-1 10, Fisher, Stuttgart
25. Aluniets, J., Hakanson, R., Sundlcr, F. & Thorell, J. (1979) .Varrrre 279. 805-806
26. Remy, C. & Dubois, M.P. (1979) Experientia 35, 137-138 27. Zipser. B. (1980) Nature 283, 857-858 28. Osborne, N.N., Patel, S . & Dockray, G. (1982) Histochemistry
29. Rzasa, P.J., Kavoustian, K.V. & Prokop, E.K. (1984) Comp.
30. Golding, D.W. & May, B.A. (1982) Acta Zoof. 63, 229-238 31. Marchand, C.R. & Dubois, M.P. (1986) Cell Tissue Res. 245,
337-341 32. Guissi-Kadri, S. , Bulet, P. & Curgy, J.J. (1990) Gen. Comp.
Eiidocriitol. 17, 339-347 33. Aviv, H. & Leder. P. (1972) Proc. Natl. Acad. Sci. U S A 69,
1408- 14 1 7 33. hlaniatis. T.. Fritsch, E.F. & Sambrook, J. eds. (1982) Mulec-
ulur Cloning: A kiboraror? Martual, Cold Spring Harbor Labora- Lor), p. 545, Cold Spring Harbor, New York
35. Benouareth, D.E., Dhainaut-Courtois, N., Porchet-HennerC, E. & Curgy. J . J . (1989) Biol. Cell. 67, 167-171
& Walker. J.M. (1984) Aiinu. Rev. Neurosci. 7, 223-255
(1986) J. Neurosci. 16, 644-649
Sci. USA 86, 6307-63 1 1
40, 533-535
75, 573-583
Biocheiit. Physiol. 77C, 345-350

36. Guissi-Kadri, S . , Dhainaut-Courtois, N., Porchet, M. & Curgy, J.J. (1991) Biol. Cell. 71, 81-87
37. Engelhardt, R.P., Croix, D. & Leonardelli, J. (1989) XIX Coll. SOC. Neuroendoc. Exp. Rouen. France. Ann. Endocrinol. 50, 27N
38. Rodriguez-Bello, A,, Croix, D. & Tramu, G. (1990) 2nd Int. Congress Neuroendocrinol. Bordeaux. France. Neuroendocrinol. 52, S1, 91
39. Burr, F.A. & Burr, B. (1983) Methods Enzymol. 96, 239-244 40. Laemmli, U.K. (1970) Nucure 227, 680-685 41. Towbin, H., Staehelin, T. & Gordon, J. (1979) Proc. Nutl. Acad.
Sci. USA 76,4350-4354
Hormone precursors in marine worm
42. Acher, R. (1988) Ann. Etzdocrinol. 49, 369-370 43. Soreq, H. & Miskin, R. (1981) FEBS Lett. 128, 305-310
Address: Dr. Th. Dugimont C.N.R.S. UA 148 Service Ac. Nucleiques SN 3-2 Etage-Porte 214 Endocrinologie des Invertbbres Universitt. de Lille I 59655 Villeneuve d’Ascq Cedex France
307