potential of extremophiles for biotechnological and ... and metals interaction/pdf/potential... ·...
TRANSCRIPT

Energy Sources, 24:543–559, 2002Copyright © 2002 Taylor & Francis0090-8312 /02 $12.00 C .00DOI: 10.1080/0090831029008655 4
Potential of Extremophiles for Biotechnological andPetroleum Applications
M. S. A. TANGOM. R. ISLAM
Department of Civil EngineeringDalhousie UniversityHalifax, Canada
The ability of microorganisms to survive under harsh conditions has recently promptedresearchers to study these microbes to better understand their characteristics andeventually utilize them in various applications. Extremophilic microorganisms with-stand one or more extreme condition constraints due to cold, heat, high acidity, highalkalinity, high salinity, and high pressure levels. These constraints would destroy nor-mal microorganisms. Extremophiles can be classi� ed according to the environment inwhich they survive. Many extremophilic microorganisms possess properties suitablefor biotechnological and commercial uses. This article discusses the characteristicsand future potentials of extremophiles.
Keywords biotechnological, biohydrometallurgy, extromophiles, extromozymes,growth environments, heavy metal recovery, petroleum containment
Introduction
Over the last two decades, scientists have been intrigued by the microbes that thrivein extreme environments. The microorganisms that have the ability to resist or with-stand extreme environmental conditions are known as extremophiles. Extremophiles maybe divided into 5 categories—thermophiles (high temperature), acidophiles (low pH),alkaliphiles (high pH), halophiles (high salinity), and psychrophiles (low temperature)—clearly indicating the nature of the habitats favored by these microbes.
These habitats include hot springs, shallow submarine hydrothermal systems, orhot-vent systems, where microorganisms can be found at temperatures above 100° C. Ex-tremophiles are also found in highly saline lakes or salterns, sometimes at salt conditionsnear that of saturation, and in environments with extreme pH values, either acidic (acidic� elds and acidic sulfur pyrite areas) or alkaline (freshwater, alkaline hot springs, car-bonate springs, alkaline soils, and soda lakes). The sources of psychrophilic organismsinclude the cold polar seas and soils and Alpine glaciers, as well as deep-sea sedi-ments, which are cold and are also at high pressures. Figures 1–4 illustrate images ofextremophilic organisms found in deep ocean (about 3.7 km below the ocean surface) en-vironments. Often extremophiles live in situations combining several stress factors, for in-stance, a high temperature and acidic conditions or a high pressure and low pH conditions.
Received 4 June 2001; accepted 12 July 2001.Address correspondence to Dr. R. Islam, Department of Civil Engineering, Dalhousie Univer-
sity, 1360 Barrington Street, Halifax, Canada, NS B3J 2X4. E-mail: Ra� [email protected]
543

544 M. S. A. Tango and M. R. Islam
Figure 1. Environmental scanning electron microscopy (ESEM) showing chains of cocci bacteria(extremophiles) detected in the rusticles. (Courtesy: Wells and Mann, 1997.)
Figure 2. Filamentous (Leptothrix sp.) extremophilic bacteria in the rusticles formed basket-shapedstructures. (Courtesy: Wells and Mann, 1997.)
Extremophiles for Biotechnological and Petroleum Uses 545
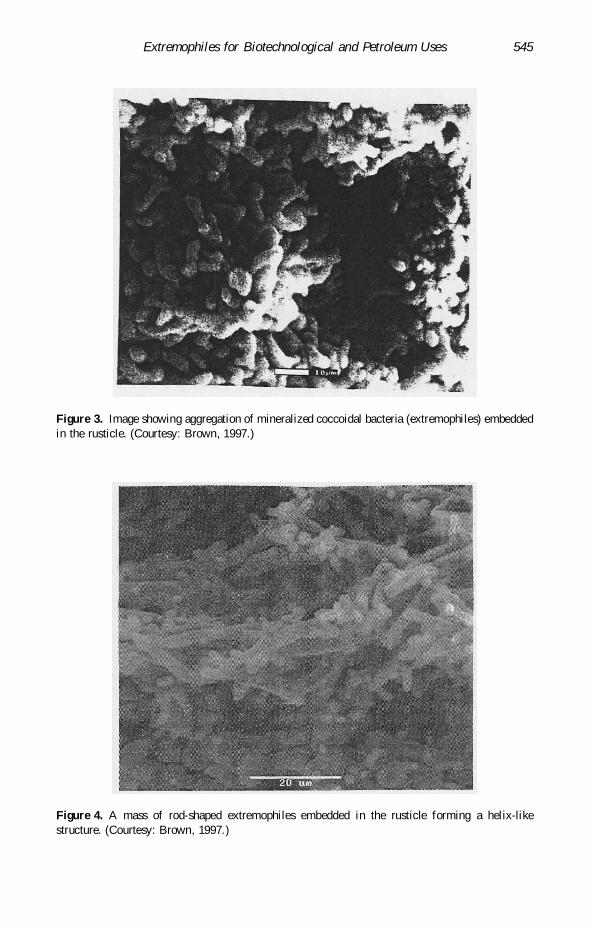
Figure 3. Image showing aggregation of mineralized coccoidal bacteria (extremophiles) embeddedin the rusticle. (Courtesy: Brown, 1997.)
Figure 4. A mass of rod-shaped extremophiles embedded in the rusticle forming a helix-likestructure. (Courtesy: Brown, 1997.)
Tab
le1
Bio
dive
rsity
ofex
trem
ophi
les
Ext
rem
egr
owth
Met
abol
icC
ondi
tion
Mic
roor
gani
sms
Sour
ces
cond
ition
sch
arac
teri
stic
sR
efer
ence
s
Low
tem
pera
ture
Bac
illu
sTA
41C
old
sea
wat
er4°
CA
erob
iche
tero
trop
hPo
laro
mon
asva
cuol
ata
4°C
Rin
ker
etal
.,19
99
Hig
hte
mpe
ratu
reP
yroc
occu
sfu
rios
usG
eoth
erm
alm
arin
ese
dim
ents
85°C
Ana
erob
iche
tero
trop
hSc
hich
oet
al.,
1993
The
rmot
oga
neap
olit
ana
Geo
ther
mal
mar
ine
sedi
men
ts10
0°C
Ana
erob
iche
tero
trop
h
Hig
hte
mpe
ratu
rean
dhi
ghpr
essu
reM
etha
noco
ccus
jann
asch
iiD
eep
sea
Hyd
roth
erm
alve
nts
85°C
250
atm
Met
hane
/gro
wth
pro-
duct
ion
Rin
ker
etal
.,19
99
Hig
hpH
Clo
stri
dium
para
doxu
mSe
wag
esl
udge
pH10
.3A
naer
obic
hete
rotr
oph
Wie
gel,
1999
Low
pHan
dhi
ghte
mpe
ratu
reM
etal
losp
haer
ase
dula
Hot
acid
pool
spH
2.0
75°C
Facu
ltativ
ech
emo-
litho
trop
hC
lark
etal
.,19
93,
and
Pee
ples
and
Kel
ly,
1995
Hig
hsa
ltH
alob
acte
rium
halo
bium
Hyp
ersa
line
wat
ers
4–5
MN
aCl
Aer
obic
hete
rotr
oph
Hal
obac
teri
umsa
lina
rum
Hyp
ersa
line
wat
ers
4–5
MN
aCl
Taya
let
al.,
1997
546

Extremophiles for Biotechnological and Petroleum Uses 547
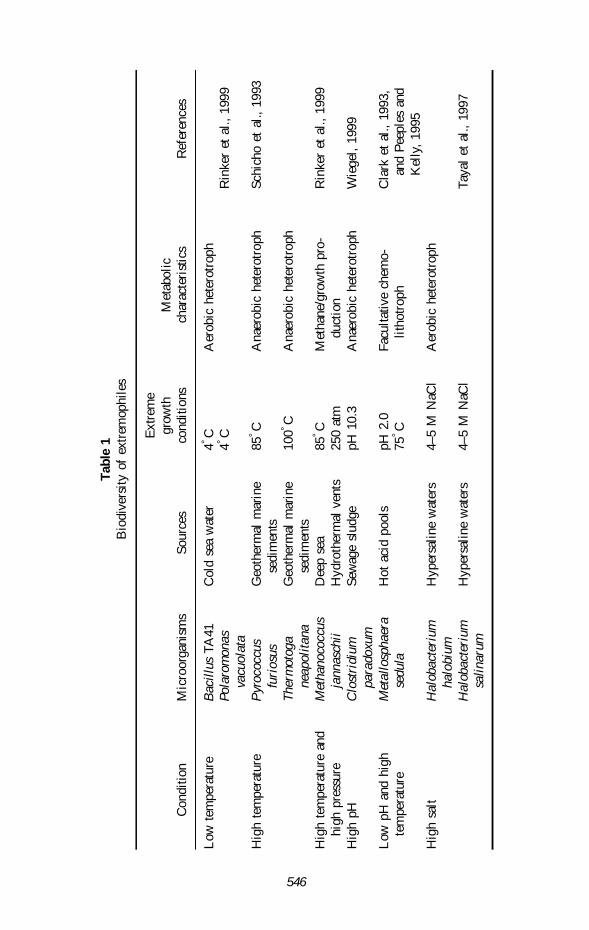
These instances suggest that wherever there is an extreme environment, there are verylikely to be extremophiles. Table 1 shows the diversity of some extremophiles.
The ability of microorganisms to survive under harsh conditions has recently promptedresearchers to study these organisms to better understand their characteristics and even-tually utilize them in various applications. Many extremophilic microorganisms possessproperties suitable for biotechnological and commercial uses.
This paper reviews the pertinent role of extremophiles and their potential in biotech-nological and petroleum industries.
Growth Environments
Cold Environments
Cold environments are often more common than hot ones. More than half of the earth’ssurface is ocean and has an average temperature of 1–3° C. The majority of land areasin the Arctic and Antarctic are permanently frozen and are thawed for only a few weeksduring summer. These areas are favored by and support the life of psychrophile microbes.
Some studies have reported on microbial communities in Antarctic sea ice-oceanwater that remains frozen for much of the year (Irgens et al., 1996; Rinker et al., 1999).These communities include photosynthetic eukarya, algae, and diatoms, as well as a va-riety of bacteria. One specie of bacterium found was Polaromonas vacuolata, a typicalrepresentative of a psychrophile. Its optimal temperature for growth is 4° C, and temper-atures above 12° C were found to be too warm for reproduction. Cold-loving organismshave started to interest manufacturers for the development of enzymes that work at re-frigerator temperatures. Food processing companies whose products often require coldtemperatures to avoid spoilage, producers of fragrances that evaporate at high tempera-tures, and manufacturers of cold-wash laundry detergents are the potential bene� ciariesof this group of microbes.
Acidic Environments
The majority of natural environments on the earth are essentially neutral, having pH valuesbetween 5 and 9. Acidophiles survive in the habitats having a pH <5 and at temperaturesbetween 50 and 79° C (Brock et al., 1972). These organisms are the subject of increasinglyintense studies due to their potential for in situ metal leaching of geothermally heatedore deposits. Clark et al. (1993) observed highly ef� cient ore-leaching capabilities usingMetallosphaera sedula.
Highly acidic environments can result naturally from geochemical activities suchas the production of sulfurous gases in hydrothermal vents, some hot springs, and themetabolic activities of certain acidophiles themselves (Peeples and Kelly, 1995). Acido-philes are also found in the debris left over from coal mining. Apparently, acidophilescannot tolerate great acidity inside their cells, where it would destroy important moleculessuch as DNA. They survive by keeping the acid out. The molecules that protect the cell,as well as others that come into contact with the environment, must be able to operatein extreme acidity. Indeed, extremozymes that are able to work at a pH below 1, moreacidic than even vinegar or stomach � uids, have been isolated from the cell wall andunderlying cell membrane of some acidophiles.
Potential applications of acid-tolerant extremozymes range from catalysts for thesynthesis of compounds in an acidic solution to additives for animal feed, which areintended to work in the stomachs of animals. The enzymes that are selected are those

548 M. S. A. Tango and M. R. Islam
that microbes usually secrete into the environment to break food into sizes suitable foringestion. When added to feed, these enzymes improve the digestibility of inexpensivegrains, thereby avoiding the need for more expensive food.
Alkaline Environments
Alkaliphiles live in soils rich with carbonates and in so-called soda lakes, such as thosefound in Africa (such as Egypt and the Rift Valley in Kenya and Tanzania) and the westernU.S. At a pH above 8, certain molecules, especially those made of ribonucleic acid (RNA),break down. Alkaliphiles, just like acidophiles, maintain neutral pH in their interior, andtheir extremozymes are located on or near the cell surface and in external secretions. Thisfeature makes alkaliphiles suitable for producing detergents. Manufacturers of detergentsin the U.S. and Japan are particularly excited by alkaliphilic enzymes. The Horikoshigroup at Japan Marine Science and Technology has embraced extremozymes with en-thusiasm and pioneered the research into alkaliphilic extremozymes (Horikoshi, 1999).
To work effectively, detergents must be able to cope with stains from food and othersources of grease. This task can easily be accomplished by such enzymes as proteases(protein degraders) and lipases (grease degraders). However, laundry detergents tend tobe highly alkaline and thus destructive to standard proteases and lipases. Alkaliphilicversions of these enzymes can solve the problem, and several types that can operateef� ciently in heat or cold are now in use or being developed. Alkaliphilic extremozymesare also earmarked to replace standard enzymes that produce better aesthetics on fabric.Over the past 2 decades, many studies have focused on the enzymology, physiology,ecology, taxonomy, molecular biology, and genetics of alkaliphilic microorganisms toestablish a new microbiology of alkaliphilic microbes (Horikoshi, 1999).
Saline Environments
The group of halophilic extremophiles inhabit highly saline environments, especiallynatural salt lakes and solar salt evaporation ponds. The solar salt ponds are constructedpools where seawater collects and evaporates, leaving behind high concentrations ofsalt that can be harvested and utilized as deicing and antiicing agents. Some salineenvironments are also extremely alkaline, due to weathering of sodium carbonate, andcertain other salts can release ions that produce alkalinity. In such cases, microorganismsin these environments are adapted to both high alkalinity and high salinity.
Halophiles are able to live in salty conditions through a fascinating adaptation. Be-cause water tends to � ow from areas of high solute concentration to areas of lowerconcentration, a cell suspended in a very salty solution will lose water and becomedehydrated unless its cytoplasm contains a higher concentration of salt (or some othersolute) than its environment. Halophiles contend with this problem by producing largeamounts of an internal solute or by retaining a solute extracted from outside (Garabitoet al., 1998). For instance, an archaean known as Halobacterium salinarum concentratespotassium chloride in its interior. As might be expected, the enzymes in its cytoplasmwill function only if a high concentration of potassium chloride is present, but proteins inHalobacterium salinarum cell structures that are in contact with the environment requirea high concentration of sodium chloride.
The potential applications for salt-tolerant enzymes are steadily growing, followingthe trends for other extremozymes. For example, investigators are now exploring incor-porating halophilic extremozymes into procedures used to increase the amount of crude

Extremophiles for Biotechnological and Petroleum Uses 549
extracted from oil wells. Tayal et al. (1997) reported promising laboratory tests throughenzymatic hydrolysis of this aspect while studying the viscosity of hydraulic fracturing� uids.
Scientists rarely � nd large quantities of a single species of microbe in nature. Adesired organism must be puri� ed, usually by isolating single cells, and then grownin a laboratory culture. For organisms with extreme lifestyles, isolation and large-scaleproduction can prove both dif� cult and expensive.
Harvesting Extremozymes
Fortunately, extremozymes can be produced through recombinant DNA technology with-out massive culturing of the source extremophiles (Adams and Kelly, 1998). Genes, whichconsist of DNA, specify the composition of the enzymes and other proteins made by cells.These proteins carry out most cellular activities. As long as microbial prospectors canobtain sample genes from extremophiles in nature or from small laboratory cultures, theycan generally clone those genes and use them to make the corresponding proteins. Thatis, by using the recombinant DNA technologies, they can insert the genes into ordinary,or domesticated, microbes, which will often use the genes to produce unlimited, puresupplies of the enzymes (Robertson et al., 1996).
Two approaches have been exploited to identify potentially valuable extremozymes.The more traditional route requires scientists to grow at least small cultures of an ex-tremophile obtained from a particular environment. For instance, if the scientists areinterested in protein-degrading enzymes, they test to see whether extracts of the culturedcells break down selected proteins. Once the activity is detected, the scientists resort tostandard biochemical methods to isolate the enzymes responsible for the activity andto isolate the genes encoding the enzymes. They then must hope that the genes can beinduced to produce their corresponding proteins in a host gene.
In the other approach, researchers avoid growing any cultures of extremophiles.They isolate the DNA from all living things in a sample of water, soil, mud, rocks, orother material from an extreme environment. Then, using recombinant DNA technologyonce again, they deliver random stretches of DNA into a domesticated host. Often theyperform one insert per host cell without knowing the identities of the genes in thosefragments. Eventually they screen the colonies that grow out, looking for evidence ofactivity by new novel enzymes. If they � nd such evidence, they conclude that an insertedgene is responsible and that it will work in the domesticated host. In this way they avoidmany bottlenecks in the traditional process. Thus scientists can extract the genes for theenzymes from mixed populations of microbes without needing to culture extremophilesthat might have trouble growing outside their native habitat.
Although the microbial world is diverse, scientists rarely succeed in extracting theperfect enzyme for a particular task. Therefore the leading scientists of industrial enzymetechnology have begun to modify extremozymes, tailoring them to meet speci� c needs.For example, after � nding an extremozyme that degrades proteins fairly ef� ciently athigh temperatures, researchers might modify the enzyme so that it functions across awider range of acidity and salinity. So far most extremozymes in commercial use arealtered little from their original state. But the enzyme modi� cation approaches promise toenhance extremozymes. They may also help to convert enzymes from ordinary microbesinto arti� cial extremozymes.
Discovery of extremophiles opens new opportunities for the development of enzymeshaving superior catalytic capabilities. However, for any new enzyme to gain commercial

550 M. S. A. Tango and M. R. Islam
acceptance, its producers will have to reduce the costs of production. In addition, theyhave to ensure that the domesticated microbes used as sources of extremozymes willreliably generate large quantities of the protein. The dif� culties of perfecting manufac-turing techniques, and the reluctance of industries to change systems that already workreasonably well, could slow the entry of new extremozymes into commerce. It seemsinevitable, however, that their many advantages will eventually prove irresistible.
Biotechnological Applications of Extremophiles
Extremozymes
The use of extremophilic microorganisms in industrial processes has grown rapidly overthe last 2 decades. Every category of these microbes has unique characteristics that can beharnessed for use in biotechnological industries. Enzymes from these microbes, namelyextremozymes, possess high stability and reduced risk of contamination of the organismsthat produce them. Other useful features of these enzymes during the production processinclude improved transfer rates and lower viscosity. Due to the superior properties ofthese enzymes, they are expected to form the bridge between biological and chemicalprocesses (Adams and Kelly, 1998).
Recent developments of extremozymes indicate that they are good novel catalystsof great industrial interest (Adams and Kelly, 1995). Typical examples are polymer-degrading enzymes like amylases, proteases, cellulases, pullulanases, and xylanases.These enzymes have signi� cant roles in chemical, food, pharmaceutical, paper, pulp, andwaste-treatment industries (Adams et al., 1995). In the pharmaceutical industry, mem-branes of some extremophiles have been found to contain surfactants bearing a uniquestability. Other important innovative products are cyclodextrins, compatible solutes, andpolyunsaturated fatty acids.
Heavy Metal Removal
Heavy metal contamination is a problem in many environments as a result of increasingindustrial activities. Currently, the risk associated with toxic heavy metals is not wellknown. Microorganisms can interact with heavy metals in a variety of ways that resultin decreased metal mobility and solubility. Two biogeochemically important groups ofmicrobes, the metal and sulfate-reducing bacteria, have suitable physiology for metalprecipitation and immobilization. The activities of these microbes provide metabolicproducts such as iron and hydrogen sul� de, which lead to mineral formation. These min-erals can react with heavy metals, forming precipitation and hence detoxi� cation. Currentunderstanding of how these microbes work is limited, and research has to be conductedto investigate how these metal and sulfate-reducing organisms behave in contaminatedsites. In order to understand the removal of these toxins, we must explore the effects ofheavy metals on the metabolism of anaerobic bacteria.
Toxic waste sites can poison or kill more commonly found organisms—those thatrequire oxygen for survival. However, microbes that can live anaerobically (without oxy-gen) can tolerate the extreme conditions found in underground waste sites. Such microbesmight be useful in keeping contamination from spreading away from already contam-inated radioactive and metal-saturated soil. This application would be highly useful inmany countries searching for ways to clean up contaminated soil. The research team led

Extremophiles for Biotechnological and Petroleum Uses 551
by T. C. Onstott of Princeton University went to the deep mines of the Transvaal regionof South Africa looking for microbes that live at high temperatures and considerable pres-sures and that can handle an environment rich in uranium and organic carbon. The teamfound anaerobic microbes that survive in deep mine environments, possess high metalcontent, and have a higher than average amount of radiation. In particular, they werelooking for microbes inhabiting an organic-rich vein that is highly mineralized, contain-ing high concentrations of uranium and pyrite and one of the richest gold deposits in theworld. The isolation process of extremophiles found in such areas need special attention.Rock samples were processed at the mine to remove the outer, potentially contaminatedsurfaces. Crushed rock fragments and groundwater collected from boreholes were usedto inoculate the growth media in the sampling containers before the samples were added.The team also collected rock samples for further analyses on chemical and biologicalprocesses that occur around the microbes themselves, nucleic acids and membrane lipids,and to identify their types and metabolism.
Waste uranium in groundwater, streams, and ponds may be in the oxidized state andposes a potential health problem. Such uranium is soluble and mobile, increasing the riskof being spread into other areas. Regulatory guidelines require uranium to be con� ned toits sites, thus the need to change the chemical state of uranium. The main aim is to makeuranium attach to soil, rocks, and stream sediments to ensure that it is retained on site.Bacteria (Shewanella putrefaciens MR-1 bacterium) that prevent uranium from dispersinginto groundwater has been isolated (Palumbo and Zhou, 1996). This microbe is intriguingbecause it not only reduces metals (and thus is ideal for treating contaminated waters),but it also produces hydrogen and magnetite. Further studies on the genes from thismicrobial genome might be used to design microorganisms for advanced bioremediationof contaminated sites and streams. As a result, the uranium should precipitate out of thewater, drop into the sediments, and remain within the con� ned site.
Energy Production
Biological hydrogen production is another interesting area. Extremozymes are also em-ployed in the production of hydrogen gas. Hydrogen may be the fuel of the future oncehydrogen fuel cells for propelling cars are perfected (McAlister, 1998). When oxygenand hydrogen are combined in a fuel cell, they provide electricity and a little heat, giv-ing water as the only waste product. The hydrogen car will be clean because it willnot discharge nitrogen oxides and carbon dioxide. Hydrogen can be obtained cheaplyusing special enzymes (extremozymes) by the transformation of cellulose into glucosesugar, then converting the glucose product and its byproduct , gluconic acid, into hydrogen(Woodward et al., 2000). The Woodward team has identi� ed, isolated, puri� ed, and char-acterized extremozymes that might be useful for bioprocessing and energy production.
The process of converting glucose to hydrogen is more ef� cient at higher temper-atures, and thus it makes sense to replace some standard enzymes with extremozymes(hydrogenase). This hydrogenase is produced by the extremophile Pyrococcus furiosus, astrain of bacteria from a deep-sea hydrothermal vent (Raven et al., 1992). It works mostef� ciently at a temperature of 85° C and is also one of only two enzymes known to acceptelectrons from NADPH to produce hydrogen. Standard enzymes will stop working whenexposed to higher temperatures unless special, costly measures are taken to protect theseproteins. Some extremozymes might eliminate the need for protective steps, increasingef� ciency and reducing costs. In addition, extremozymes may be more stable and react

552 M. S. A. Tango and M. R. Islam
faster than their mesophilic counterparts. Similar studies in this � eld showed that hy-drogen can be obtained enzymatically from other biomass substrates, such as lactose,sucrose, xylan, starch, and steam-exploded aspen wood (Miyamoto, 1997).
Biological hydrogen production is the most challenging area of biotechnology withrespect to environmental problems. The future of biological hydrogen production dependsnot only on research advances, but also on economic considerations (the cost of fossilfuels), social acceptance, and the development of hydrogen energy systems.
Compatible Solutes
These are osmotically active substances that maintain the halophilic cell in positive wa-ter balances and are compatible with the cellular metabolism. They have low molecularweight and are highly water-soluble sugars, alcohols, amino acids, and their derivatives(Galinski, 1995; Ventosa et al., 1998). Compatible solutes are increasingly being uti-lized in biotechnological applications as stabilizers of biomolecules (DNA, enzymes, andmembranes) and whole cells or as stress-protective agents. Galinski (1995) reported on anovel biotechnological process called “bacterial milking,” which deals with production ofectoines and hydroxyectoines using extremely halotolerant Halomonas elongata. Ectoineand ectoine derivatives have also been patented as moisturizers in cosmetics for the careof aged, dry, or irritated skin (Motitschke et al., 2000). Another promising applicationis the use of ectoine as stabilizers in the polymerase chain reaction (PCR) (Sauer andGalinski, 1998).
Biopolymers
Several biopolyners have been developed from halophilic microorganisms. These includebiosurfactants, exopolysaccharides, and bioplastics.
Biosurfactants enhance the remediation of oil-contaminated soil and water. By de-creasing surface tension, they increase the solubility and thus the mobility of hydrophobi chydrocarbons, which may promote degradation. Biosurfactant-producing halophilic mi-croorganisms may therefore play a signi� cant role in the accelerated remediation ofoil-polluted saline environments. Hack et al. (2000) reported promising results in hydro-carbon pollution control in marine biotopes in closed systems, such as oil storage tanks,and several studies indicated potential for pollution treatment in marine environmentsor coastal areas. New biosurfactants (trehalose lipids), produced by marine rhodococciduring cultivation on n-alkanes, could become promising surface-active agents for in situbioremediation of cold marine environments (Yakimov et al., 1999). Unfortunately, theproduction cost of biosurfactants is about 3–10 times higher than that of the conven-tional chemical compounds, which is too expensive for microbial enhanced oil recovery(MEOR) (Desai and Banat, 1997).
Exocellular polymeric substances are also called exopolysaccharides (EPS) (Weineret al., 1995). These substances can also be derived from halophytic microorganismsand are of interest for MEOR. The desirable properties of EPS include high meltingtemperatures, pseudoplasticity, and resistance to salt, color, and thermal degradation. EPSare often used as emulsi� ers and mobility controllers. Active emulsi� cation of petroleumhas been noted for 6 strains, close to Halobacterium salinarum, Haloferax volcanii, andHalobacterium distributum (Kulichevskaya et al., 1992).
Polyhydroxyalkanoate s (PHA) are intracellularly accumulated bacterial storage com-pounds. These biodegradable plastics that have properties comparable to those of poly-

Extremophiles for Biotechnological and Petroleum Uses 553
ethylene and polypropylene (Rodriguez-Valera and Lillo, 1992; Steinbücel et al., 1997).Such biodegradable plastics could replace oil-derived thermoplastics in some applica-tions. Haloferax mediterranei accumulates large amounts (up to 60% of cell dw) ofpolyhydroxy butyric acid (PHB). PHB production can be enhanced to about 6 g/l usingphosphate limitation and starch, a cheap substrate, as the carbon source (Rodriguez-Valeraand Lillo, 1992). PHB recovery is simpli� ed because the exposure of the halophile tolow salt concentrations causes cell lysis (Nieto et al., 2000). In addition, this type ofhalophile can be cultivated easily in a simple saline open pond without risk of contami-nation. Recently, an extremely halophilic archaeon, isolated from an Egyptian saline soil,was reported to accumulate PHB as intracellular granules with an amount of about 53%of its cell dry weight (Hezayen et al., 2000). The substrates used were n-butyric acid andsodium acetate.
Microbial Leaching (Biohydrometallurgy)
Biohydrometallurgy using autotrophs extremophiles has made signi� cant advances indevelopment as a commercially viable technology for processing sulphide ores. Due toits environmentally friendly nature, biohydrometallurgy should come to replace manypyrometallurgical ore extraction methods in the near future.
Bioleaching has future potential for remediation of heavy metals contaminated ma-terials (Krebs et al., 1997; Haddadin et al., 1995). The process has no routine treat-ment, hence leaching processes using autotrophic or heterotrophic extremophiles maybe considered for environmental clean ups. Biohydrometallury’s advantages over othertechniques include low capital and energy costs, high � exibility, ability to be used onsite, lack environmental pollution, and a shorter set-up time frame than competing ap-proaches. Bioprocesses have been determined as being one-third to one-half the cost ofconventional chemical and physical remediation technologies.
Chirpa et al. (1998) has described the new technology of electrobioleaching employ-ing the principles of applying DC potentials to promote mineral dissolution and growthof Thiobacillus ferrooxidans. Some metals such as copper, uranium, and gold can berecovered from the ores by the process called biomining. This process is based on theactivity of chemoautolithotrophic bacteria. These bacteria can grow in highly acidic envi-ronments and are able to utilize either sulfur or iron as their energy source. For example,Thiobacillus ferrooxidans has been employed to enhance copper and gold recovery fromthe ores (Rawlings, 1998). One of the limitations of this process is that of achieving highmicrobial growth rates. However, ferric iron (Sugio et al., 1985) and essential growthrequirements (Touvinen et al., 1971) have been used to improve ef� ciency of the metalrecovery from ores.
Microbial leaching is considered to be a simple and effective process for extractingprecious metals from low-grade ores and mineral concentrates. Remediation using bio-leaching has an economic advantage because it involves low capital and energy costs,high � exibility, and on-site application, and it does not cause environmental pollution. Inaddition, microbial leaching serves as a means for remediation of mining sites, treatmentof mineral industrial waste products, detoxi� cation of sewage sludge, and recovery ofheavy metals from soils and sediments (Bosecker, 2001).
Chirpa et al. (1998) has suggested that fundamental research should be directed to-ward heterotrophic leaching and/or bene� ciation of silicate, carbonate, and oxide ores.Additional research is required in microbiology and the interaction of biologists withmetallurgical engineers. There should be more comprehensive study of microbial com-

554 M. S. A. Tango and M. R. Islam
position bioheap systems and stirred-tank bioreactors. There is a need to de� ne andunderstand potential interactions among components of micro� ora in bioreactors to im-prove bioleaching and mineral biooxidation. Rapid, accurate, and simple techniques formonitoring the microbial activity in bioleach/mineral biooxidation systems are needed forcontrol of these processes by operators. New types of reactors, such as vat-type systemsthat permit bioleaching and effective handling of whole ore particles in the 10-mesh sizerange, are required. New materials for high temperature, highly corrosive conditions thatare relatively low cost are required. Industrially applicable technology for heterotrophicleaching of metal oxides, carbonates, and silicates should be explored (Brierley andBrierley, 2001).
Petroleum Contaminant Removal
Petroleum is an important energy source in industry and daily life. The annual con-sumption of petroleum has been increasing in the last several years. With an increasingfrequency of petroleum transportation and use, the probability of potential accidentalrelease of petroleum into the environment is also increasing. Nowadays, the occurrenceof oil spills on soil and in oceans is a matter of concern. In this regard, researchersare continuously investigating the fate of petroleum hydrocarbons in the marine environ-ment and on land with an overall aim of developing effective biotechnologies for thebioremediation of spilled oil.
Hydrocarbons
The application of bioremediation as an alternative remediation technology is becomingthe technique of choice for environmental professionals. This method offers superior ben-e� ts not found in other remediation processes. Biodegradation of petroleum hydrocarbonsvaries with the chemical structure and molecular weight of hydrocarbon molecules. Thechemical structure of organic pollutants has signi� cant in� uence on the extent and rateof their biodegradation (Alexander, 1981).
Extremophiles have also been utilized for the microbial degradation of crude oil andre� ned petroleum pollutants. Foght and McFarlane (1999) have discussed the growth ofextremophiles on petroleum hydrocarbon. Some potential applications are those related tomolecular genetics of polycyclic aromatic hydrocarbon degradation by bacteria. Speci� cstudies have focused on the factors that control degradation and methods to enhance theability of bacteria to degrade such pollutants in the environment. Foght and Westlake(1996) have isolated bacterial strains from the environment and studied the mutationof their plasmid-borne and/or chromosomal genes. April et al. (2000) have studied theprocess of crude oil degradation by mixed populations, pure cultures, and genetic mutants.They demonstrated the loss of parent compounds and analyzed the products of bacterialattack on crude oils using gas chromatography, mass spectrometry, and radiorespirometry.New studies in this area involve biodegradation of hydrocarbon contaminated under coldAntarctica conditions (Aislabie et al., 2000).
Most bacteria that are capable of degrading petroleum hydrocarbons have been iso-lated from deep ocean environments. However, their roles in the bioremediation processare virtually unknown. Yakimov et al. (1999) isolated the Alcanivorax group from theNorth Sea as biosurfactant-producing and n-alkane-degrading marine bacteria. Thesebacterial strains were isolated from a variety of marine environments, including oil spillcontaminated sites. During growth on n-alkanes, Alcanivorax borkmensis produces bio-surfactants that are shown to be glucose lipids (Abraham et al., 1998). Genus Alcanivorax

Extremophiles for Biotechnological and Petroleum Uses 555
seems to play a major role in the � rst step of crude oil biodegradation in the marineenvironment (Harayama et al., 1999). However, information on the growth character-istics of Alcanivorax are still very limited. Therefore researchers are currently inves-tigating the nutrient requirements of Alcanivorax coupled with investigations into thealkane-degradation pathways and gene analysis. They use phylogenetic identi� cation andquanti� cation by the technique called � uorescence in situ hybridization (FISH) (Amannet al., 1995). These investigations are performed with the overall aim to achieve com-plete and rapid biodegradation of alkanes. Harayama et al. (1999) observed that bacteriabelonging to the genus Alcanivorax are important for the biodegradation of petroleum,especially under bioremediation conditions.
Al-Maghrabi et al. (1998) reported rapid degradation of the crude oil using ther-mophilic bacteria. The same specie of these thermophilic bacteria was found to survivein saline environments. Oil spills have been successfully bioremediated in marine, Arctic,and Antarctic environments (Delille et al. 1998; Margesin and Schinner, 1999). A newhalo- and thermotolerant Streptomyces albaxialis was found that degraded crude oil andpetroleum products even in the presence of 30% (w/v) NaCl (Kuznetsov et al. 1992).Zvyagintseva et al. (1995) isolated an extremely halophilic Archaea (Haloferax mediter-ranei) able to grow at 10–25% (w/v) NaCl; the same specie was also reported to utilizeoil as the sole carbon source (Kulichevskaya et al. 1992) obtained for the � rst time asan isolate of the Halobacterium group from salt-rich stratum � uids of an oil deposit.The strain degraded n-alkanes with a C10-C30 composition in the presence of 30% (w/v)NaCl.
However, the use of bioremediation is still unproven and has some limitations whenit is employed for speci� c problems. Few of the previous studies have reported that somechemical compounds are not biodegradable, need extensive monitoring, may involvepotential production of toxic byproducts, and require strong qualitative scienti� c support.
New Findings
Heavy Metal Recovery
Recent � ndings at Princeton University indicate a newly discovered form of the bac-terium Thermus sp. isolated from a groundwater sample (Onstott, 2001). This microbeis moderately tolerant to radiation effects, grows anaerobically, and in the mine environ-ment interacts with forms of iron, chromium, cobalt, and uranium, forms of elementsthat often occur in contaminated soils. None of the other forms previously studied haveshown this metabolic versatility, despite previous studies on the physiology and geneticsof the microbe for nearly 3 decades. Two strains of thermophilic bacteria belonging tothe Bacillus family were isolated from a hot water stream and used to remove strontiumfrom aqueous stream systems (Chalaal and Islam, 2001). These bacteria were able toconcentrate strontium in one side of a two-compartment bioreactor. Immobilization ofheavy metals using sul� de-producing microorganisms has been reported as an effectivemeans of treating some metals-contaminated sites (Crawford and Crawford, 1996).
Future Research Interests
Although several research � ndings have explored some of the unique features of ex-tremophiles, some physiological and metabolic characteristics of these microbes in rela-tion to their growth environment still need further study.

556 M. S. A. Tango and M. R. Islam
Extremophiles such as anaerobic bacteria possess metabolic activities for biode-grading a range of pollutant chemicals, including aromatic, halogenated aromatic, andhalogenated alkene compounds. In order to utilize anaerobic bacteria in extreme envi-ronments, it is necessary to understand their physiology and effects the environment mayhave on the metabolism of the bacteria of interest.
Presently, a majority of commercial applications of bioremediation depend uponindigeneous microorganisms, and most are employed for hydrocarbon-contaminated sites.The use of genetically engineered microorganisms for remediation of some selectedindustrial pollutants such as trichloroethane (TCE) will gain an increased role in thecoming years.
Impending bioremediation development programs should continue to dwell on isola-tion and characterization of superior and novel microbes to degrade halocarbons, geneticengineering of hard to degrade compound pathways, optimization of conditions for growthand activity of such microbes, and design and testing of � eld pilot systems.
At present, most of the biotechnological potential applications of metal/microorganisminteractions are in the bioleaching of ores, bioremediation of metal-polluted sites, andmineralization of polluting organic matter. Further development on the microbial interac-tions with metals need some attention for ef� cient future exploitation of this technology.Genetic improvement of metal solubilizing microorganisms, whether by mutation and se-lection or by genetic engineering, will permit bioremediation processes to be improved.Intensive inter-disciplinary collaborations in basic and applied research will be bene� cialfor development of more effective applications.
Halophilic extremozymes have a potential in applications that is expected to increasethe amount of crude extracted from oil wells. The shortcomings in perfecting manufac-turing techniques, and the reluctance of industries to change systems that are in place andwork reasonably well, could limit the promotion of new extremozymes into commercialuse. However, future use looks inevitable because there are numerous advantages thatwill eventually bring about acceptance.
Extremophiles are not cultivatable under conventional laboratory culture conditions,but may offer a wealth of valuable bioproducts, ranging from bioactive small molecules tounique biopolymers and enzymes. To overcome the limitation of cultivating extremophileson a production scale, researchers have developed a unique production-scale bioreactorcapable of continuous operation at extreme temperatures and pressures. Further devel-opmental work in this direction needs to be done because the demand is growing at anexponential rate.
References
Abraham, W. R., H. Meyer, and M. Yakimov. 1998. Novel glycine containing glucolipids from thealkane using bacterium Alcanivorax borkumensis. Biochim. Biophys. Acta 1393:57–62.
Adams, M. W. W., and R. M. Kelly. 1995. Enzymes isolated from microorganisms that grow inextreme environments. Chemical and Engineering News 73(51):32–42.
Adams, M. W. W., and R. M. Kelly. 1998. Finding and using hyperthermophilic enzymes. TIBTECH,16:329–332.
Adams, M. W. W., F. B. Perler, and R. M. Kelly. 1995. Extremozymes. Expanding the limits ofbiocatalysis. Bio/technology 13:662–668.
Aislabie, J., J. Foght, and D. Saul. 2000. Aromatic hydrocarbon-degrading bacteria isolated fromsoil near Scott Base, Antarctica. Polar Biol. 23:183–188
Al-Maghrabi, I. M. A., A. A. Bin Aqil, M. R. Islam, and O. Chalaal. 1998. Use of thermophilicbacteria for bioremediation of petroleum contaminants. Energy Sources 21:17–29.

Extremophiles for Biotechnological and Petroleum Uses 557
Alexander, M. 1981. Biodegradation of chemicals of environmental concern. Science 211:132–138.Amann, R., W. Ludwig, and K. H. Schleifer. 1995. Phylogenetic identi� cation and in situ detection
of individual microbial cells without cultivation. Microbiological Reviews 59:143–169.April, T. M., J. M. Foght, and R. S. Currah. 2000. Hydrocarbon-degrading fungi isolated from � are
pit soils in northern and western Canada. Can. J. Microbiol. 46:38–49.Bosecker, K. 2001. Microbial leaching in environmental clean-up programmes. Hydrometallurgy
59:245–248.Brierley, J. A., and C. L. Brierley. 2001. Present and future commercial applications of biohy-
drometallurgy. Hydrometallurgy 59:233–239.Brock, T. D., K. M. Brock, R. T. Belly, and R. L. Weiss. 1972. Sulfolobus: A new genus of sulfur-
oxidizing bacteria living at low pH and high temperature. Arch. Microbiology 84:54–68.Brown, M. R. 1997. The identi� cation of bacteria pure culture from and analysis of the elemental
and structural composition of a nestide taken from the R. M. S. Titanic. B.Sc (Hans) thesis,Dalhousie University, Halifax, Canada.
Chalaal, O., and M. R. Islam. 2001. Integrated management of radiactive strontium contaminationin aqueous stream systems. J. Environ. Management 61:51–59.
Chirpa, S., J. M. Mudak, and K. A. Natarajan. 1998. Electrobioleaching of sphalerite � oatationconcentrate. Minerals Engineering 11:783–788.
Clark, T. R., F. Baldi, and G. J. Olson. 1993. Coal depyritization by the thermophilic archaeonMetallosphaera sedula. Appl. Environ. Microbiol. 59:2375–2379.
Crawford, R. L., and D. L. Crawford (ed.). 1996. Bioremediation: Principles and Applications,Cambridge: University Press.
Delille, D., A. Bassères, and A. A. Dessommes. 1998. Effectiveness of bioremediation for oil-polluted Antarctic seawater. Polar Biol 19:237–241.
Desai, J. D., and I. M. Banat. 1997. Microbial production of surfactants and their commercialpotential. Microbiol. Mol. Biol. Rev. 61:47–64.
Ehrlich, H. L. 2001. Past, present, and future of biohydrometallurgy. Hydrometallurgy 59:127–134.Foght, J. M., and D. M. McFarlane. 1999. Growth of Extremophiles on Petroleum. In Enigmatic
Microorganisms and Life in Extreme Environments, J. Seckbach (ed.). Dordrecht, Netherlands:Kluwer Academic Publishers, pp. 527–538.
Foght, J. M., and D. W. S. Westlake. 1996. Transposon and spontaneous deletion mutants ofplasmid-borne genes encoding polycyclic aromatic hydrocarbon degradation by a strain ofPseudomonas � uorescens. Biodegradation 7:353–366.
Galinski, E. A. 1995. Osmoadaptation in bacteria. Advances in Microbiology and Physiology37:273–328.
Galinski, E. A., and B. J. Tindall. 1992. Biotechnological Prospects for Halophiles and HalotolerantMicro-organisms. In Molecular Biology and Biotechnology of Extremophiles, R. H. Herbertand R. J. Sharp (eds.). Glasgow: Blackie, pp. 76–114.
Garabito, M. J., M. C. Marquez, and A. Ventosa. 1998. Halotolerant Bacillus diversity in hypersalineenvironments. Canadian Journal of Microbiology 44:95–102.
Hack, C. J., Woodley, J. M., Lilly, M. D., and Liddell, J. M. 2000. Design of a control system forbiotrasis formation of toxis substrates: Toluene hydroxylation by pseudomonas putida UV4.Enzyme and Microbial Technology 26:530–536.
Haddadin, J., C. Dagot, and M. Fick. 1995. Models of bacterial leaching. Reviews: Enzyme andMicrobial Technology 17:290–305.
Harayama, S., H. Kishira, Y. Kasai, and K. Shutsubo. 1999. Petroleum biodegradation in marineenvironments. Journal of Molecular Microbiology and Biotechnology 1:63–70.
Hezayen, F. F., B. H. A. Rehm, R. Eberhardt, and A. Steinbüchel. 2000. Polymer production bytwo newly isolated extremely halophilic archaea: Application of a novel corrosion-resistantbioreactor. Appl. Microbiol. Biotechnol. 54:319–325.
Horikoshi, K. 1999. Alkaliphiles, Tokyo: Harwood Academic Publishers.Irgens, R. L., J. J. Gosink, and J. T. Staley. 1996. Polaromonas vacuolata, nov. gen et sp., a
psychrophilic, marine, gas vacuolate bacterium from Antarctica. Int. J. Syst. Bacteriol. 46:822–826.

558 M. S. A. Tango and M. R. Islam
Krebs, W., C. Brombacher, P. P. Bosshard, R. Bachofen, and H. Brandl. 1997. Microbial recoveryof metals from solids. FEMS Microbiology Reviews 20:605–617.
Kulichevskaya, I. S., E. I. Milekhina, I. A. Borzenkov, I. S. Zvyagintseva, and S. S. Belyaev. 1992.Oxidation of petroleum hydrocarbons by extremely halophilic archaebacteria. Microbiology60:596–601.
Kuznetsov, V. D., T. A. Zaitseva, L. V. Vakulenko, and S. N. Filippova. 1992. Streptomyces albi-axalis sp. nov.: A new petroleum hydrocarbon-degrading species of thermo- and halotolerantStreptomyces. Microbiology 61:62–67.
Madigan, M. T., J. M. Martinko, and J. Parker. 1997. Brock Biology of Microorganisms, 8th ed.,Englewood Cliffs, NJ: Prentice Hall.
Margesin, R., and F. Schinner. 1999. Biological decontamination of oil spills in cold environments.J. Chem. Technol. Biotechnol. 74:381–389.
McAlister, R. 1998. Manufacturing Sustainable Prosperity. In Hydrogen Today, American HydrogenAssociation. Vol. 8, No. 1. pp. 1–34.
Miyamoto, K. 1997. Renewable Biological Systems for Alternative Sustainable Energy Production,FAO Agricultural Services Bulletin 128, Osaka University, Japan.
Motitschke, L., H. Driller, and E. Galinski. 2000. Ectoin and Ectoin derivatives as moisturizers incosmetics. US Patent 060071.
Nieto, J. J., C. Vargas, and A. Ventosa. 2000. Osmoprotection mechanism in the moderatelyhalophilic bacteria Halomonas alongata. Recent Research and Development in Microbiology4:43–54.
Onstott, T. C. 2001. Microbial Diversity in Hot Zone: South African Microbiology Project, De-partment of Geosciences, Geomicrobiology Research Group’s website, Princeton University,Princeton, NJ. http://geoweb.princeton.edu/people/faculty/enstott/research.html
Palumbo, J., and J. Zhou. 1996. Amazing Microbes, URL www.ornl.govPeeples, T. L., and R. M. Kelly. 1995. Bioenergetic response of the extreme thermoacidophile
Metallosphaera sedula to thermal and nutritional stress. Appl. Environ. Microbiol., 61:2314–2321.
Raven, N., N. Ladwa, D. Cossar, and R. Sharp. 1992. Continuous culture of the hyperthermophilicarchaeum Pyrococcus furiosus. Appl. Microbiol. Biotech. 38:263–267.
Rawlings, D. E. 1998. Industrial practice and the biology of leaching of metals from ores: The1997 Pan labs lecture. J. Indust. Microbiol. Biotech. 20:268–274.
Rinker, K. D., C. J. Han, M. W. W. Adams, and R. M. Kelly. 1999. Cultivation of Hyperthermophilicand Extremely Thermoacidophilic Microorganisms. In Manual of Industrial Microbiology andBiotechnology, 2nd ed. A. L. Demain and J. E. Davis (eds.), Washington, D.C.: ASM Presspp. 119–136.
Robertson, D. E., E. J. Mathurs, R. V. Swarsy, B. L. Marrs, and J. M. Short. 1996. The discoveryof new biocatalysts from microbial diversity. SIM News 46:3–8.
Rodriguez-Valera, F., and J. G. Lillo. 1992. Halobacteria as producers of polyhydroxyalkanoates .FEMS Microbiol. Rev. 103:181–186.
Sauer, T., and E. A. Galinski. 1998. Bacterial milking: A novel bioprocess for production ofcompatible solutes. Biotechnology Bioengineering 57:306–313.
Schicho, R. N., K. Ma, M. W. W. Adams, and R. M. Kelly. 1993. Bioenergetics of sulfur reductionin the hyperthermophilic archaeon Pyrococcus furiosus. Journal of Bacteriology, 175:1823–1830.
Steinbüchel, A., B. Füchtenbusch, V. Goren� o, S. Hein, R. Jossek, S. Langenbach, and B. H. A.Rehm. 1997. Biosynthesis of polyesters in bacteria and recombinant organisms. Polymer De-grad. Stabil. 59:177–182.
Sugio, T., C. Domatsu, O. Munakata, T. Tano, and K. Imai. 1985. Role of ferric ion-reducingsystem in sulfur oxidation of Thiobacillus ferrooxidans. Appl. Environ. Microbiol. 49:1401–1406.
Tayal, A., R. M. Kelly, and S. A. Khan. 1997. Viscosity reduction of hydraulic fracturing � uidsthrough enzymatic hydrolysis. Journal of Society of Petroleum Engineering 2(2):204–212.

Extremophiles for Biotechnological and Petroleum Uses 559
Touvinen, O. H., S. I. Niemela, and H. G. Gyllenberg. 1971. Effect of mineral nutrients andorganic substances on the development of the Thiobacillus ferrooxidans. Biotechnology andBioengineering 13:517–527.
Ventosa, A., J. J. Nieto, and Oren. 1998. Biology of moderately halophilic aerobic bacteria. Mi-crobiology and Molecular Biology Reviews 62:504–544.
Weiner, R., S. Langille, and E. Quintero. 1995. Structure, function and immunochemistry of bac-terial exopolysaccharides. Journal of Industrial Microbiology 15:339–346.
Wells, W. S., and H. Mann. 1997. Microbiology and Formation of Microbially Assisted Rusticlesfrom the R.M.S. Titanic, B.Sc. (Hons) thesis, St. Mary’s University, Halifax, Canada.
Wells, W. S., and H. Mann. 1997. Microbiology information of rustides from the R. M. S. Titanic.Resource and Environmental Biotechnology 1:271–281.
Wiegel, J. 1999. Microbial Biosystems: New Frontiers. Proceedings of the 8th International Sym-posium on Microbial Ecology, C. R. Bell, M. Brylinsky, and P. Johnson-Green (eds.), AtlanticCanada Society for Microbial Ecology, Halifax, Canada.
Woodward, J., M. Orr, K. Cordray, and E. Greenbaum. 2000. Biotechnology: Enzymatic productionof biohydrogen. Nature, 405:1014.
Yakimov, M. M., L. Guiliano, V. Bruni, S. Scar� , and P. N. Golyshin. 1999. Characterization ofAntarctic hydrocarbon-degrading bacteria capable of producing bioemulsi� ers. Microbiologica(Pavia) 22:249–256.
Zvyagintseva, I. S., S. S. Belyaev, I. A. Borzenkov, N. A. Kostrikina, E. I. Milekhina, and M. V.Ivanov. 1995. Halophilic archaebacteria from the Kalamkass oil � eld. Microbiology 64:67–71.