potential impact of deep seabed mining on the larvae of tunas
TRANSCRIPT

JULY 1884
POTENTIAL IMPACT OF DEEP SEABED MINING ON THE LARVAE OF TUNAS AND BILLFISHES
Walter M. Matsumoto
NOAA-TM-NMFS-SWFC-44
U.S. DEPARTMENT OF COMMERCE National Oceanic and Atmospheric Administration National Marine Fisheries Service Southwest Fisheries Center

NOAA Technical Memorandum NMFS
The National Oceanic and Atwospheric Administration (NOAA), organized in 1970, has evolved into an agency which establishes national policies and manages and conserves our oceanic, coastal, and atrno- spheric resources An organizational element within NOAA, the Office of Fisheries is responsible for fisheries policy and the direction of the National Marine Fisheries Service (NMFS)
In addition to its formal publications. the NMFS uses the NOAATechnical Memorandum series to issue informal scientific and technical publications when complete formal revtew and editorial processing are not appropriate or feasible Documents within this series, however, reflect sound professional work and may be referenced in the fornial scientific and technical literature
.

NOAA Technical Memorandum NMFS
This TM Series 1s llsed lor documentation and timely communication of preliminary resulfs imfelim reports. 01
special purpose information and have not received complete formal review. editorial control. or detailed editing
JULY 1984
POTENTIAL IMPACT OF DEEP SEABED MINING ON THE LARVAE OF TUNAS AND BILLFISHES
Walter M. Matsumoto Southwest Fisheries Center Honolulu Laboratory
National Marine Fisheries Service, NOAA Honolulu, Hawaii 968 12
Prepared for NOAA Division of Ocean Minerals and Energy
Washington, D.C. 20235
NOAA-TM-NMFS-SWFC-44
U.S. DEPARTMENT OF COMMERCE Malcolm Baldr ige. S e c r e t a r y National Oceanic and Atmospheric Administration John V. Byrne. Administrator
National Marine Fisheries Service William G. Gordon, Assistant Administrator for Fisheries

TABLE OF CONTENTS
Page
I n t r o d u c t i o n . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
i n t h e P a c i f i c Ocean . . . . . . . . . . . . . . . . . . . . . . . . . 1 D i s t r i b u t i o n . . . . . . . . . . . . . . . . . . . . . . . . . . 1
. d u l t s . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 L a r v a e . e . . . . . . . . . . . . . . . . . . . . . . . . . 6
Foodand f eed ing . . . . . . . . . . . . . . . . . . . . . . . . 9 Adults . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 Larvae and j u v e n i l e 8 .................... 11
Spawning . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 2 Development and growth of 13
15 Environment . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
Oceanographic f e a t u r e 8 . . . . . . . . . . . . . . . . . . . 15 B i o l o g i c a l f e a t u r e s . . . . . . . . . . . . . . . . . . . . 16
Tunas and b i l l f i s h e s i n t h e DOMES area . . . . . . . . . . . . . 18 Movemen to f tunas . . . . . . . . . . . . . . . . . . . . . . . . 19
E f f e c t s of mining . . . . . . . . . . . . . . . . . . . . . . . . . . 20 Impact on environment . . . . . . . . . . . . . . . . . . . . . . 20 Impact on p r i m a r y product ion . . . . . . . . . . . . . . . . . . 24 Impact on microzooplankton . . . . . . . . . . . . . . . . . . . 25 Impact o n f i s h . . . . . . . . . . . . . . . . . . . . . . . . . . 25
Adults . . . . . . . . . . . . . . . . . . . . . . . . . . . 25 Tuna and b i l l f i s h larvae . . . . . . . . . . . . . . . . . . 27
Sedimentat ion . . . . . . . . . . . . . . . . . . . . . 27 T u r b i d i t y . . . . . . . . . . . . . . . . . . . . . . . 28 Environmental f a c t o r s . . . . . . . . . . . . . . . . . 29
Other p o s e i b l e effects . . . . . . . . . . . . . . . . . . . . . 32 E f f e c t s on f i s h e r i e s . . . . . . . . . . . . . . . . . . . . . . 34
Summary and conc lus ions . . . . . . . . . . . . . . . . . . . . . . . 35 Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . 38 L i t e r a t u r e c i t e d . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
D i s t r i b u t i o n and biology of t unas and b i l l f i s h e e
larvae . . . . . . . . . . . . . . . . . The Deep Ocean Mining Environmental Study (DOMES) Area . . . . . . . .

TABLE
.
1.
1.
2.
3.
4.
5 .
6.
7 .
8.
9.
F ish spec ie s caught ( i n pounds) by pole-and-line f i s h i n g i n 1978 dur ing t h e Hawaiian f i s h aggrega t ing device experiment (Matsumoto e t a l . 1981) *
LIST OF FIGURES
D i s t r i b u t i o n and a reas of abundance of ye l lowf in tuna i n t h e P a c i f i c Ocean. Hatched a r e a r e p r e s e n t s range of d i s t r i b u t i o n ; d o t s r e p r e s e n t ca t ches of mare than 1 f i s h pe r 100 hooks ( [ Japan . ] F i s h e r i e s Agency, Research Divi.sion 1970, 1972, 1973, 1981; Shingu e t a l . 1974; Calk ins 1975; [Korea.] F i s h e r i e s Research and Development Agency 1981).
D i s t r i b u t i o n and a reas of abundance of bigeye tuna i n t h e P a c i f i c Ocean. ca t ches of more than 0.8 f i s h per 100 hooks ( [ Japan . ] Research Div i s ion 1970, 1972, 1973, 1981; Shingu e t a l . 1974; [Korea.] F i s h e r i e s Research and Development Agency 1981).
Hatched a r e a r e p r e e e n t s range of d i s t r i b u t i o n ; d o t s r e p r e s e n t F i s h e r i e s Agency,
Skip jack tuna d i s t r i b u t i o n (hatched a r e a s ) and f i s h e r i e s (c ross -ha tch a r e a s ) i n t h e major oceans. l a te summer poleward displacement (Matsumoto e t a l . 1984) . The mean 15°C isotherms r e p r e s e n t maximum
D i s t r i b u t i o n and a r e a s of abundance of b lue mar l in i n t h e P a c i f i c Ocean. Hatched a r e a r e p r e s e n t s range of d i s t r i b u t i o n ; d o t s r e p r e s e n t ca t ches of more than 0.3 f i s h pe r 100 hooks ([Japan.] Research Div i s ion 1970, 1972, 1973, 1981; Shingu e t a l . 1974) .
F i s h e r i e s Agency,
D i s t r i b u t i o n and a r e a s of abundance of s t r i p e d mar l in i n t h e P a c i f i c Ocean. Hatched area r e p r e s e n t s range of d i s t r i b u t i o n ; d o t s r e p r e s e n t ca t ches of more than 0.4 f i s h per 100 hooks ([Japan.] Research Div i s ion 1970, 1972, 1973, 1981; Shingu e t a l . 1974) .
F i s h e r i e s Agency,
Ocean c u r r e n t sys t ems i n t h e P a c i f i c Ocean.
D i s t r i b u t i o n of ye l lowf in tuna l a r v a e i n t h e P a c i f i c Ocean (Klawe 1963; Matsumoto 1966; Ueyanagi 1967, 1969; H i r o t a 1977; Nishikawa e t a l . 1978).
D i s t r i b u t i o n of bigeye tuna l a r v a e i n t h e P a c i f i c Ocean (Matsumoto 1966; Ueyanagi 1969; Nishikawa e t al. 1978).
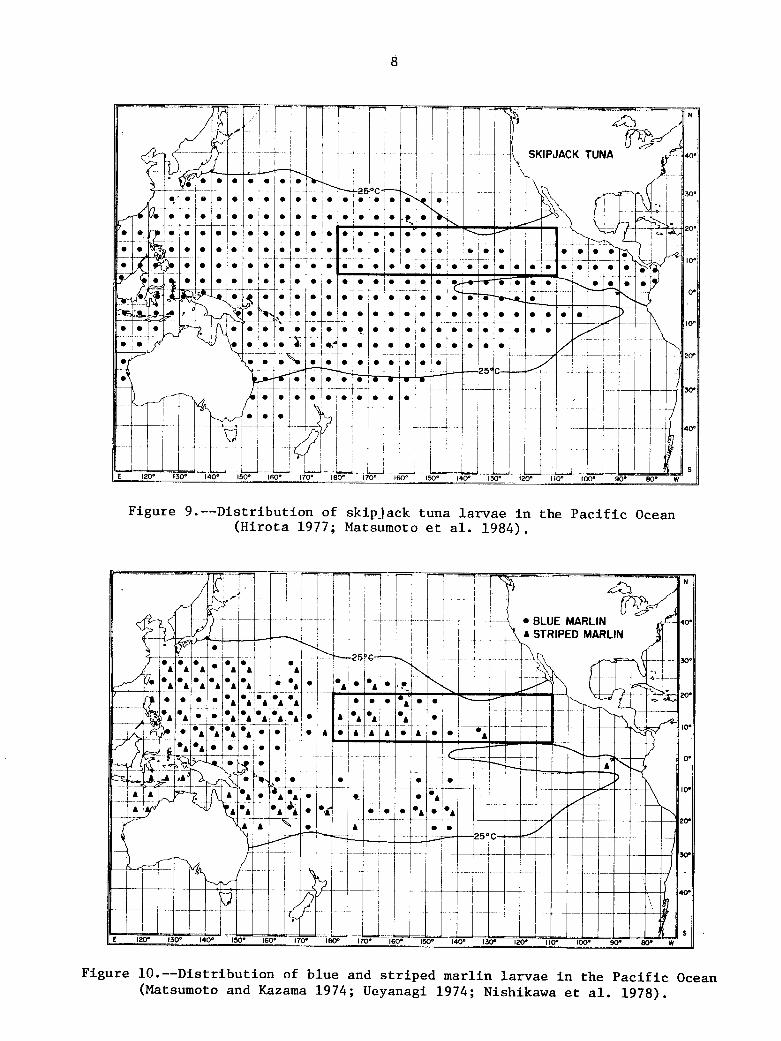
D i s t r i b u t i o n of s k i p j a c k tuna l a r v a e i n t h e P a c i f i c Ocean ( H i r o t a 1977; Matsumoto e t a l . 1984).
10. D i s t r i b u t i o n of blue and s t r i p e d mar l in l a r v a e i n t h e P a c i f i c Ocean (Matsumoto and Kazama 1974; Ueyanagi 1974; Nishikawa e t a l . 1978).
11. General s u r f a c e c i r c u l a t i o n scheme i n t h e e a s t e r n t r o p i c a l P a c i f i c w i th p r i m manganese nodule mining a r e a (ha tched) and DOMES s t a t i o n s A, B, and C.

12. Assumed movement of t h e v a r i o u s s tocks of s k i p j a c k tuna i n t h e P a c i f i c Ocean. The numerals along the migratory r o u t e s r e p r e s e n t q u a r t e r s and l o c a t i o n s of high CPUE c e l l s of s k i p j a c k tuna taken by t h e Japanese tuna l o n g l i n e f i s h e r y , 1964-67. Stock d e s i g n a t i o n s are shown i n pa ren theses (Matsumoto 1975).
13. Assumed r o u t e s followed by tagged s k i p j a c k tuna from t h e e a s t e r n t o t h e c e n t r a l P a c i f i c . The l e t t e r s r e p r e s e n t tagged f i s h and t h e numerals a long t h e r o u t e s r e p r e s e n t q u a r t e r s of high c a t c h rates of s k i p j a c k tuna t aken by t h e Japanefie tuna l o n g l i n e f i s h e r y , 1964-67 (Matsumoto 1975).
14. P l ana r view of t h e plume recons t ruc t ed from nephelometer t r a c e s . Contour l e v e l s are nephelometer v o l t a g e s . Background v o l t a g e is approximately 3.3 (Burns e t a l . 1980).
15. Calcu la t ed s u r f a c e plume of a commercial mining s h i p t r a v e r s i n g a r e c t a n g u l a r p a t t e r n o r i e n t e d normal t o t h e advec t ion d i r e c t i o n a t advec t ion v e l o c i t y of 2 5 c m / s and mining s h i p speed of 1 m / s . Concentrati.ons of p a r t i c u l a t e s i n suspension are given i n ug/R ( L a v e l l e and Ozturgut 1981).
iii

ABSTRACT
Assessment of t h e probable impact of deep seabed mining on t h e eggs and larvae of t u s a s and b i l l f i s h e s i s made, based l a r g e l y on s t u d i e s of t h e e f f e c t s of sedimentat ion on eggs, l a r v a e , and a d u l t s of f r e s h w a t e r , e s t u a r i n e , and a few marine f i s h e s . Most of t h e changes t o t h e environment r e s u l t i n g from mining, such a a i nc reased suspended sediment p a r t i c l e s , reduced i l l u m i n a t i o n due t o t u r b i d i t y , mixing of co ld bottom water wi th s u r f a c e water, and changes i n t r a c e m e t a l s , s a l i n i t y , and oxygen levels of t h e s u r f a c e water, a re n o t e x t e n s i v e enough t o endanger tuna and b i l l f i s h eggs and larvae. One probable e f f e c t , temperature of d i s c h a r g e , i s acknowledged, but i t s e f f e c t s a re expected t o be i n s i g n i f i c a n t . Cold bottom water ( 4 " - 1 O o C ) discharged ove r t h e s i d e of t h e mining s h i p and f a l l i n g d i r e c t l y o n t o tuna eggs and larvae a t t h e s u r f a c e could cause thermal shock, r e s u l t i n g i n t h e t e r m i n a t i o n of embryonic development, development of abnormal larvae, and l o s s of e q u i l i b r i u m b y , t h e larvae. Losses from thermal shock, e s t ima ted annual ly a t 15.3 m e t r i c t o n s (MT) of s k i p j a c k tuna and 10.7 MT of ye l lowf in t u n a , r e p r e s e n t i n g 0.16 and 0.04% of t h e t o t a l annual ca t ches of t h e r e s p e c t i v e s p e c i e s i n t h e e a s t e r n P a c i f i c no r the rn f i s h e r y ( t h e f i s h e r y no r th of Mexico), a r e i n s i g n i f i c a n t and are no t l i k e l y t o have any n o t i c e a b l e impact on t h e f i s h e r y . These l o s s e s could p o s s i b l y be inc reased f o u r f o l d , i f t h e mining s h i p were t o a c t as f i s h aggrega t ing d e v i c e s , but even a t t hese h ighe r l e v e l s , t h e impact on t h e f i s h e r y would be n e g l i g i b l e .
iv

INTRODUCTION
I n t h e Programmatic Environmental Impact Statement prepared by t h e Nat iona l Oceanic and Atmospheric Adminis t ra t ion (NOAA 1981) pursuant t o t h e Deep Seabed Hard Mineral Resources Act and t h e Na t iona l Environmental Po l i cy Act, several unresolved impacts from seabed mining of manganese nodules i n t h e e a s t e r n P a c i f i c Ocean ( l a t . 5"-20"N, long. ll0"W-180") are i d e n t i f i e d as having t h e p o t e n t i a l t o cause adverse e f f e c t on t h e b i o t a . One p o t e n t i a l unreso lved impact is t h e e f f e c t of p a r t i c u l a t e s d i scharged a t t h e s u r f a c e on f i s h larvae.
Recent s t u d i e s on t h e a l t e r a t i o n of t h e environment and t h e e f f e c t s on phytoplankton, macrozooplankton, and c e r t a i n f i s h s p e c i e s r e s u l t i n g from mining o p e r a t i o n s i n t h e area have been completed (Ozturgut e t a l . 1988; Chan and Anderson 1981; Hirota 1981; Lavelle and Ozturgut 1981; L a v e l l e e t a l . 1982) . These s t u d i e s inc lude d a t a on t h e d i s c h a r g e c h a r a c t e r i s t i c s , s u r f a c e plume, and e f f e c t s of suspended p a r t i c u l a t e matter on phytoplankton and macrozooplankton of t h e a r e a . of t unas exposed t o seawater con ta in ing suspended p a r t i c u l a t e matter i n c o n c e n t r a t i o n s similar t o t h a t expected from mining o p e r a t i o n s have been done a l s o (Barry 1978). a d u l t t unas would n o t be a f f e c t e d s i g n i f i c a n t l y by d i scha rges from mining ope ra t ions . growth of t h e larvaeo however, have n o t been addressed adequate ly .
Laboratory experiments w i th t w o s p e c i e s
Based on t h e s e experiments , i t was determined t h a t
The p o s s i b l e e f f e c t s of p a r t i c u l a t e s on t h e s u r v i v a l and
Th i s r e p o r t d e s c r i b e s t h e b io logy of t unas and b i l l f i s h e s i n t h e P a c i f i c Ocean, w i th emphasis on t h e prime mining areas, examines t h e probable e f f e c t s of mining on eggs, larvae and a d u l t s , and d i s c u s s e s t h e s i g n i f i c a n c e of adverse e f f e c t s on t h e f i s h e r i e s .
DISTRIBUTION AND BIOLOGY OF TUNAS AND BILLFISHES I N THE PACIFIC OCEAN
D i s t r i b u t i o n
Adul t s
The d i s t r i b u t i o n of t h e tunas i n t h e P a c i f i c Ocean has been desc r ibed O f t hose s p e c i e s of a t l e n g t h by Blackburn (1976) and Sund e t a l . (1981).
tunas and b i l l f i s h e s which occur more or less cont inuous ly a c r o s s t h e ocean, t h e fo l lowing , due t o t h e i r commercial importance, are of concern i n t h e area of prime i n t e r e s t f o r commercial deep seabed mining: ye l lowf in tuna , Thunnus a l b a c a r e s ; b igeye tuna , r. obesus; s k i p j a c k tuna , Katsuwonus pe lamis ; b l u e mar l in , Makaira n i n r i c a n s ; and s t r i p e d mar l in , Te t r aP tu rus audax.
The areal d i s t r i b u t i o n of t h e tunas does n o t v a r y much by s p e c i e s (Figs. 1-31, but t h e r e are d i f f e r e n c e s i n abundance w i t h i n t h e d i s t r i b u - t i o n a l range. t h e wes tern P a c i f i c west of t h e 180th mer id ian , between la t . 40"N and 40"s i n t h e c e n t r a l P a c i f i c west of long. 1 4 O o W , and from Cen t ra l America t o l a t . 35"s i n t h e e a s t e r n P a c i f i c . The d i s t r i b u t i o n i s narrowed t o l a t . 20"N and 30"s i n t h e e a s t e r n c e n t r a l P a c i f i c a t long. 115"-120"W. abundance g e n e r a l l y are r e s t r i c t e d t o w i t h i n 20" of t h e Equator. t una , a more tempera te s p e c i e s , i s found i n c o o l e r waters than t h e y e l l o w f i n tuna.
Yel lowfin tuna are d i s t r i b u t e d between l a t . 45"N and 45"s in
Areas of Bigeye
It i s d i s t r i b u t e d from let . 45"N t o n e a r l y 50"s i n t h e

2
Figure 1.--Distribution and areas of abundance of yellowfin tuna in the Pacific Ocean. Hatched area represents range of distribution; dots represent catches of more than 1 fish per 100 hooks ([,Japan.] Fisheries Agency, Research Division 1970, 1972, 1973, 1981; Shingu et al. 1974; Calkins 1975; [Korea.] Fisheries Research and Development Agency 1981).
western P a c i f i c , from r a t . 40"N t o 45'5 i n t h e c e n t r a l P a c i f i c , and Ut. 35'N t o 40'5 i n t h e e a s t e r n P a c i f i c e a s t of long. 130'W. Unlike t h a t of ye l lowf in tuna , t h e d i s t r i b u t i o n of bigeye tuna i s no t c o n s t r i c t e d i n t h e e a s t e r n c e n t r a l P a c i f i c . The a r e a s of ,abundance a r e skewed f u r t h e r t o t h e no r th i n t h e c e n t r a l and e a s t e r n c e n t r a l P a c i f i c (between long. 165'E and 125'W) and t o t h e south i n t h e e a s t e r n c e n t r a l and e a s t e r n P a c i f i c ( e a s t of long. 140"W). Skip jack tuna a r e d i s t r i b u t e d between l a t . 45"N and 45'5 west of t h e 180th mer id ian . The d i s t r i b u t i o n narrows g r a d u a l l y t o t h e e a s t and i s c o n s t r i c t e d o f f Baja C a l i f o r n i a and o f f s h o r e from t h e South American Continent . The a reas of abundance a r e i n d i c a t e d by t h e e x t e n t of presen t f i s h e r i e s . I n t h e wes tern P a c i f i c , west of t h e 180th mer id ian , t h e a r e a of abundance extends from l a t . 45"N t o about 20"s and narrows r a p i d l y t o t h e a r e a s l i g h t l y e a s t of t h e Hawaiian I s l ands . I n t h e e a s t e r n P a c i f i c , t h e a r e a of abundance extends from Baja C a l i f o r n i a t o c e n t r a l C h i l e a long t h e coas t and narrows seaward t o about long. 1 4 O " W . The apparent gap i n t h e a r e a of abundance between long. 140" and 160"W i s due t o t h e absence of f i s h i n g e f f o r t .
The d i s t r i b u t i o n of b i l l f i s h e s does no t d i f f e r g r e a t l y from t h a t of t h e tunas . Blue m a r l i n occurs between la t . 45"N and 40"s i n t h e western P a c i f i c , l a t . 40"N and 35"s i n t h e c e n t r a l P a c i f i c , and l a t . 35"N and 35'5 i n t h e e a s t e r n P a c i f i c (F ig . 4). The a r e a of abundance l i e s between t h e Equator and l a t . 25"N i n t h e western h a l f of t h e ocean and between la t . 5"N and 20"s i n t h e e a s t e r n h a l f . The d i s t r i b u t i o n of s t r i p e d m a r l i n i s n e a r l y

3
Figure 2.--Distr ibut ion and areas of abundance of bigeye tuna i n t h e P a c i f i c Ocean. Hatched area r e p r e s e n t s range of d i s t r i b u t i o n ; dots r e p r e s e n t ca t ches of more than 0.8 f i s h pe r 100 hooks ( [ Japan . ] F i s h e r i e s Agency, Research Div is ion 1970, 1972, 1973, 1981; Shingu e t a l . 1974; [Korea.] F i s h e r i e s Research and Development Agency 1981) .
OF OCCURRENCE
RARE OCCURREN
Figure 3.--Skipjack tuna d i s t r i b u t i o n (hatched a r e a s ) and f i s h e r i e s (c ross - ha tch a r e a s ) i n t h e major oceans. l a te summer poleward displacement (Matsumoto e t a l . 1984).
The mean 15°C i so therms r e p r e s e n t maximum

h
Figure 4 . - -Dis t r ibu t ion and areas of abundance of b lue marlin i n t h e P a c i f i c Ocean. Hatched area r e p r e s m t s range of d i s t r i b u t i o n ; d o t s r ep resen t ca t ches of more than 0.3 f i s h p e r 100 hooks ( [ Japan . ] F i s h e r i e s Agency, Research Div is ion 1970, 1972, 1973, 1981; Shingu e t a l . 1974) .
s i m i l a r t o t h a t of b lue m a r l i n i n t h e c e n t r a l and e a s t e r n P a c i f i c . I n t h e western s i d e , however, t h e poleward l i m i t s a re a t l a t . 50"N and 50"s (F ig . 5). The area of abundance l i e s i n a northwestward d i r e c t i o n from t h e nor thwes tern coas t of South America t o Japan, and i s s i t u a t e d t o t h e n o r t h of t h e c e n t e r of b lue m a r l i n abundance. The southern edge of s t r i p e d m a r l i n abundance o v e r l a p s t h e n o r t h e r n edge of b lue m a r l i n abundance i n t h e east e r n Pac i f ic .
The d i s t r i b u t i o n of tunas and b i l l f i s h e s i s in f luenced by s u r f a c e cur- r e n t s and tempera ture . I n the P a c i f i c Ocean, w a r m c u r r e n t s (North and South E q u a t o r i a l Cur ren t s ) f low westward a c r o s s t h e ocean (F ig . 6 ) . A t t h e west- e r n end, t h e c u r r e n t s are d e f l e c t e d poleward and carry w a r m water t o h igher l a t i t u d e s . Cool c u r r e n t s f low eastward a c r o s s t h e ocean a t about l a t . 45"N and 45"s. A t t h e e a s t e r n end, t h e main body of bo th c u r r e n t s t u r n toward t h e Equator and carry co ld water t o lower l a t i t u d e s . The c u r r e n t flow a f f e c t s t h e c l i m a t i c s u r f a c e tempera tures so t h a t t h e wes tern s i d e of t h e ocean i s c h a r a c t e r i z e d by a ve ry broad b e l t of t r o p i c a l water, a very narrow b e l t of tempera te water, and a r a p i d t r a n s i t i o n t o t h e co ld wa te r s of h ighe r l a t i t u d e s . The e a s t e r n s i d e i s c h a r a c t e r i z e d by a ve ry broad b e l t o f tempera te water and a v e r y narrow b e l t of t r o p i c a l water (Brock 1959). Th i s resu l t s i n t h e wider d i s t r i b u t i o n of tunas and b i l l f i s h e s i n t h e wes tern s i d e and t h e narrower d i s t r i b u t i o n i n t h e e a s t e r n s i d e .
Temperature ranges i n t h e e n t i r e d i s t r i b u t i o n a l a r e a and i n areas where abundance i s high enough f o r commercial f i s h i n g have been publ i shed

5
N
u
u!
'C
IC
C
IC
EC
x
K
3 - Figure 5.--Distr ibut ion and areas of abundance of s t r i p e d m a r l i n i n t h e P a c i f i c Ocean. Hatched area r e p r e s e n t s range of d i s t r i b u t i o n ; d o t s r e p r e s e n t ca t ches of more than 0.4 f i s h p e r 100 hooks ( [ Japan . ] F i s h e r i e s Agency, Research Div i s ion 1970, 1972, 1973, 1981; Shingu e t al. 1974) .
for t h e tunas on a worldwide b a s i s (Laevastu and Rosa 1963). These are , r e s p e c t i v e l y , 11"-28" and 18"-22°C f o r bigeye tuna , 18"-31" and 20"-28°C f o r y e l l o w f i n tuna , and 17"-28" and 19"-23°C f o r s k i p j a c k tuna. r e p o r t s (Robins 1952; Uda 1957; Jones and S i l a s 1963; Broadhead and Barrett 1964; Williams 1970; Sharp 1978) i n d i c a t e an even wider tempera ture range f o r s k i p j a c k tuna: 14"-30" and 17"-30°C. A t t h e no r the rn and southern extremes of t h e i r d i s t r i b u t i o n , t h e tunas almost c e r t a i n l y respond d i r e c t l y t o tempera ture changes (Uda 1957; Schaefer 1961) . The d i r e c t i n f l u e n c e of s u r f a c e temperature on d i s t r i b u t i o n i s less obvious i n t h e t r o p i c s where tempera ture i s r e l a t i v e l y uniform throughout t h e year (Laevastu and Rosa 1963). by t h e d i s t r i b u t i o n and abundance of s u i t a b l e food , such as i n areas of upwell ing and convergence (Blackburn 19651, by ve r t i ca l thermal s t r u c t u r e (Green 1967; Sharp 19781, and by oxygen levels (Hanamoto 1975; Sharp 1978). I n t h e e a s t e r n P a c i f i c y e l l o w f i n tuna f i s h e r y , Green showed t h a t pu r se s e i n i n g w a s more s u c c e s s f u l where t h e t o p of t h e thermocl ine d i d n o t exceed 18 q and where t h e mean temperature g r a d i e n t was s h a r p (>0.5S°C/m) w i t h i n t h e thermocl ine. Sharp (1978) noted t h a t t h e optimum f i s h i n g f o r t h i s s p e c i e s appeared t o occur where t h e 23°C isotherm was s h o a l e r than 50 m and t h e 15°C isotherm was above 80 m.
Other
Here t h e d i s t r i b u t i o n and abundance of t unas are c o n t r o l l e d mainly
Tunas and b i l l f i s h e s a re d i s t r i b u t e d v e r t i c a l l y t o g r e a t depths . The v e r t i c a l d i s t r i b u t i o n of t unas and some b i l l f i s h e s has been e s t i m a t e d by v a r i o u s methods. depth of 380 m , based on experimental v e r t i c a l l o n g l i n e ca t ches ( S a i t o
Although b igeye and y e l l o w f i n tunas were d i s t r i b u t e d t o a

6
Figure 6.--Ocean c u r r e n t systems in t h e P a c i f i c Ocean.
1973; S a i t o and Sasak i 19741, t h e depths of g r e a t e s t concen t r a t ions were s t r a t i f i e d w i t h some ove r l ap among s p e c i e s . mostly a t 150-290 m, yel lowf in tuna a t 150-300 m, and bigeye tuna a t 290- 380 m . because t h e y a r e l a r g e l y taken by s u r f a c e f i s h i n g gea r , were observed a t dep ths of 152 m from submarines (S t r a sburg e t a l . 1968). They have been d e t e c t e d a t 235 q w i t h echo sounders (Yamanaka e t a l . 19661, and a t 263 w i t h u l t r a s o n i c devices (Dizon e t a l . 1978).
S t r i p e d mar l in were taken
Skip jack tuna , which have been cons idered s u r f a c e dwe l l e r s , mainly
Larvae
The d i s t r i b u t i o n a l range of tuna l a r v a e i s l e s s e x t e n s i v e than t h a t of t h e a d u l t s . Yel lowfin tuna l a r v a e (F ig . 7) range from la t . 35"N t o 25'5 i n t h e western P a c i f i c and from l a t . 30"N t o 2 5 " s i n t h e c e n t r a l P a c i f i c . The range narrows t o about l a t . 10"N and 20"-25"s i n t h e e a s t c e n t r a l P a c i f i c ( long . 130" t o 140"W) and t o l a t . 26"N and 10"s i n t h e e a s t e r n P a c i f i c . The north-south d i s t r i b u t i o n of bigeye tuna l a r v a e (F ig . 8) is l e s s e x t e n s i v e than t h a t of ye l lowf in tuna l a rvae . It ranges from l a t . 35"N t o 25"s i n t h e western P a c i f i c , from l a t . 25"N t o 10" o r 15"s i n t h e c e n t r a l P a c i f i c and l a t . 20"N t o 10"s i n t h e e a s t e r n P a c i f i c . The d i s t r i b u t i o n c o n s t r i c t s t o about 10" no r th and south of t h e Equator a t about long. 135"- 155"W and l a t . 15"N t o t h e Equator a t about long. 125"W. Skip jack tuaa l a r v a e a r e d i s t r i b u t e d over t h e wides t north-south range among tuna s p e c i e s i n t h e western h a l f of t h e ocean (F ig . 9). 35"N t o 40"s i n t h e wes tern s e c t o r , and la t . 30"N t o 35"s i n t h e c e n t r a l s e c t o r , but c o n s t r i c t s sha rp ly t o la t . 10"-15"N and 20'5 e a s t of long. 145"W and l a t . 15"N e a s t of long. 1OO"W. l i m i t s co inc ide q u i t e w e l l w i th t h e 25°C s u r f a c e isotherm.
The range ex tends from l a t .
I n a l l s p e c i e s t h e poleward

7
Figure i ' .--Distri.bution of ye l lowf in tuna l a r v a e i n t h e P a c i f i c Ocean (Klawe 1963; Matsumoto 1966; Ueyanagi 1967, 1969; H i ro t a 1977; Nishikawa et a l . 1978).
I
N
D
D
0
IO
0
IO
0
K
K
S - Figure 8.--Distr ibut ion of bigeye tuna l a r v a e i n t h e Pac i f ic Ocean
(Matsumoto 1966; Ueyanagi 1969; Nishikawa e t a l . 1978).
8
Yet. I.
Figure 9.--Distribution of skipjack tuna larvae in the Pacific Ocean (Hirota 1977; Matsumoto et al. 1984).
Figure 10.--Distribution of blue and striped marlin larvae in the Pacific Ocean (Matsumoto and Kazama 1974; Ueyanagi 1974; Nishikawa et al. 1978).

9
The d i s t r i b u t i o n of blue and s t r i p e d m a r l i n l a r v a e (Fig. 10) somewhat resembles t h a t of t h e bigeye tuna l a r v a e i n t h e western and c e n t r a l P a c i f i c . With t h e excep t ion of two c a p t u r e s i tes shown i n t h e f i g u r e , none has been r e p o r t e d east of long. 140"W.
Information on t h e v e r t i c a l d i s t r i b u t i o n of tuna larvae has been ob ta ined e n t i r e l y from zooplankton and l a r v a l f i s h n e t tows. There i s s u f f i c i e n t evidence t o i n d i c a t e t h a t tuna l a r v a e are d i s t r i b u t e d v e r t i c a l l y t o dep ths of 130 m o r more (S t r a sburg 19601, 'bu t most are confined t o t h e upper 50 o r 60 m (Matsumoto 1958; S t r a sburg 1960; K l a w e 1963; Ueyanagi 1969; H i r o t a 1977). S t r a sburg found no d i f f e r e n c e i n t h e ca t ches of tuna l a r v a e made a t t h e s u r f a c e ( h o r i z o n t a l n e t tows) and a t 0-60 m (ob l ique n e t tows). Ueyanagi a l s o found no d i f f e r e n c e i n t h e c a t c h rates of larvae sampled i n h o r i z o n t a l tows made a t t h e s u r f a c e (0-2 m) and i n subsu r face (20-30 m) layers. Th i s was a l s o t r u e f o r larvae of a l b a c o r e , s k i p j a c k , ye l lowf in and bigeye tunas. No d i f f e r e n c e was apparent i n t h e c a t c h rates of and ye l lowf in tuna l a r v a e taken i n neuston n e t tows a t t h e s u r f a c e and i n 1-75 m ob l ique bongo n e t tows, but much h ighe r c a t c h rates were ob ta ined f o r s k i p j a c k tuna larvae a t t h e s u r f a c e t h a n ' i n t h e 1-75 m depth l a y e r ( H i r o t a 1977, t a b l e s 21-24).
There i s some evidence of v e r t i c a l d i u r n a l m i g r a t i o n by s k i p j a c k tuna larvae, but t h i s may not be t r u e of ye l lowf in and bigeye tuna larvae. Ueyanagi (1969) r epor t ed t h a t t h e mean s u r f a c e c a t c h ra te of s k i p j a c k tuna larvae p e r s u c c e s s f u l n i g h t tow was f i v e t i m e s g r e a t e r than t h a t of day tows; whereas t h e mean c a t c h rates of ye l lowf in and bigeye tuna larvae were about equa l f o r day and n i g h t tows. Par t of t h e h ighe r n i g h t c a t c h e s of s k i p j a c k tuna l a r v a e could have been due t o n e t dodging du r ing d a y l i g h t , but t h i s e f f e c t should be small i n view of t h e absence of similar day-night d i f f e r e n c e s i n t h e c a t c h rates of ye l lowf in tuna and bigeye tuna l a r v a e .
Food and Feeding
Adults
Tunas a re broadly ca rn ivo rous , n o n s e l e c t i v e , o p p o r t u n i s t i c f e e d e r s , and w i l l l i k e l y prey upon any fo rage organisms p r e s e n t i n t h e i r v i c i n i t y . S tud ie s of stomach con ten t s of tunas i n t h e e a s t e r n , c e n t r a l , and western P a c i f i c ( R e i n t j e s and King 1953; Ronquillo 1953; King and Ikehara 1956; Watanabe 1958, 1960; Ive r sen 1962; Alverson 1963; Waldron and King 1963) a l l i n d i c a t e t h a t t h e d i e t of tunas i s q u i t e v a r i e d , but on ly a few kinds of t h e more abundant organisms p r e s e n t i n a given area are taken i n q u a n t i t y .
The food of tunas c o n s i s t s of t h r e e major c a t e g o r i e s : c r u s t a c e a n s , molluscs (almost e n t i r e l y cephalopods) , and f i s h i n va ry ing p r o p o r t i o n s , depending upon t h e tuna s p e c i e s and t h e area i n which they occur. c e n t r a l P a c i f i c , f i s h and cephalopods make up t h e bulk of t h e food consumed. The p r o p o r t i o n s of t h e major food c a t e g o r i e s range from 1 t o 6% c r u s t a c e a n s , 20 t o 39% cephalopods, and 62 t o 75% f i s h . I n t h e e a s t e r n P a c i f i c , however, c rus t aceans and f i s h are t h e major source of food, and cephalopods a r e only of minor importance. 592 c r u s t a c e a n s , 3 t o 8X cephalopods, and 37 t o 47% f i s h (King and Ikehara 1956; Ive r sen 1962; Alverson 1963; Waldron and King 1963).
I n t h e
The p r o p o r t i o n s range from 45 t o

10
The d i v e r s i t y of food e a t e n by tunas i s r e f l e c t e d i n t h e l a r g e number of i n v e r t e b r a t e o r d e r s and f i s h f a m i l i e s r ep resen ted i n t h e stomach con ten t s . Among t h e c rus t aceans , food items consumed by ye l lowf in and s k i p j a c k tunas i n t h e c e n t r a l and e a s t e r n P a c i f i c inc luded organisms i n t h e Orders Isopoda, Amphipoda, Euphausiacea, Decapoda, Mysidncea, and Stomatopoda. Copepods were consumed on ly by s k i p j a c k tuna i n t h e central P a c i f i c , and b igeye tuna u t i l i z e d t h e fewest c rus t acean Orders: Isopoda, Amphipoda, Euphausiacea, and Decapoda.
The g r e a t e s t d i v e r s i t y i n t h e food of t unas i s i n t h e l a r g e number of f i s h f a m i l i e s r ep resen ted . King and Ikehara (1956) l i s t 47 f i s h f a m i l i e s i n t h e food e a t e n by ye l lowf in tuna and 35 f i s h f a m i l i e s f o r b igeye tuna i n t h e c e n t r a l P a c i f i c , Alverson (1963) l i s ts 22 f i s h f a m i l i e s f o r s k i p j a c k tuna and 41 f o r ye l lowf in tuna i n t h e e a s t e r n P a c i f i c , and Waldron and King (1963) l i s t 41 f i s h f a m i l i e s f o r s k i p j a c k tuna i n t h e c e n t r a l P a c i f i c . Of t h e t o t a l f i s h f a m i l i e s g iven f o r each tuna species, 14 were common t o a l l fou r s p e c i e s , 22 were common t o b igeye , ye l lowf in , and s k i p j a c k t u n a s i n t h e c e n t r a l P a c i f i c , 33 were common t o b igeye and y e l l o w f i n tunas i n t h e c e n t r a l P a c i f i c , and 13 were common t o ye l lowf in and s k i p j a c k t u n a s i n t h e east e r n P a c i f i c .
Desp i t e t h e d i v e r s i t y of t h e f i s h s p e c i e s e a t e n by t h e tunas , f i s h of only a few f a m i l i e s make up t h e bulk of t h e f i s h p o r t i o n of t h e d i e t , and t h e major f i s h f a m i l i e s d i f f e r e d f o r some s p e c i e s by a r e a . I n t h e c e n t r a l P a c i f i c , t h e Fami l i e s Scombridae, Gempylidae, and Bramidae were t h e most important f i s h food items of ye l lowf in and b igeye t u n a s and Scombridae, Carangidae, and Mul l idae of s k i p j a c k tuna. I n t h e e a s t e r n P a c i f i c , t h e f i s h e s of t h e Fami l ies Scombridae, O s t r a c i i d a e , Te t r aodon t idae , and Exocoet idae were impor tan t food items f o r t h e ye l lowf in tuna and those of Gonostomatidae, Exocoet idae, T r i c h i u r i d a e , Myctophidae, and Engraul idae were impor tan t food items of s k i p j a c k tuna. Other s t u d i e s by Ronqui l lo (1953) and Watanabe (1958) show s i m i l a r d i v e r s i t y and s imi la r i t i es i n f i s h f a m i l i e s r e p r e s e n t e d i n t h e stomach c o n t e n t s of t unas from t h e wes tern e q u a t o r i a l P a c i f i c .
Tunas f eed p r i m a r i l y dur ing t h e hours o f d a y l i g h t from t h e s u r f a c e t o dep ths of 380 m ( S a i t o 1973; S a i t o and S a s a k i 1974). s u r f a c e u s u a l l y occurs i n compact s choo l s , c o n s i s t i n g of f i s h t h a t are g e n e r a l l y smaller than 100 cm. These schoo l s a re o f t e n composed of mixed s p e c i e s , such as s k i p j a c k tuna wi th sma l l y e l l o w f i n tuna , small ye l lowf in tuna wi th small b igeye tuna , o r a l l t h r e e s p e c i e s t o g e t h e r , and a r e g e n e r a l l y encountered i n t h e v i c i n i t y of land masses, seamounts, banks, or around f l o a t i n g o b j e c t s (Alverson and Pe te r son 1963). The l a r g e r ye l lowf in and b igeye tunas f eed a t g r e a t dep ths (135 m o r more), but may ascend t o sha l lower waters t o f eed a t n i g h t (Watanabe 1958).
Feeding a t t h e
The m a r l i n s a l s o a r e ca rn ivo rous o p p o r t u n i s t i c f e e d e r s and f e e d on p e l a g i c f i s h , squid , and c r u s t a c e a n s , Members of s i x o r d e r s of inve r t e - b r a t e s (Decapoda, Amphipoda, Stomatopoda, Heteropoda, Pteropoda, and Cephalopoda) and 36 f i s h f a m i l i e s have been r e p o r t e d from stomachs of 6 s p e c i e s of b i l l f i s h e s from t h e western P a c i f i c (Watanabe 1960). Although t h e l i s t of food i t e m s may n o t be r e p r e s e n t a t i v e of any s i n g l e s p e c i e s , ( a l l items were l i s t e d under m a r l i n ) , it does i n d i c a t e t h a t t h e var ie ty of food items e a t e n by b i l l f i s h e s as a group i s comparable t o t h a t of tunas . S t r a sburg (1970) i n d i c a t e d t h a t t h e d i e t a r y h a b i t s of m a r l i n s v a r i e d between s p e c i e s and areas. He found t h a t t h e s t r i p e d mar l in f e d m o s t l y on
c

squ ids , s a u r i e s , and o t h e r f i s h e s o f f New Zealand, s a u r i e s o f f C a l i f o r n i a , squ ids o f f Peru and C h i l e , anchovies o f f no r the rn C h i l e , snake mackerel and l a n c e t f i s h o f f t h e Bonin I s l a n d s , and tunas i n t h e c e n t r a l P a c i f i c . The b lue mar l in f e d mostly on squ ids i n the P h i l i p p i n e Sea, t u n a l i k e f i s h o f f New Zealand and i n t h e c e n t r a l P a c i f i c , dolphin and t u n a l i k e f i s h o f f t h e Bahamas, and smal l tunas (young a lbacore ; f r i g a t e tuna , A u x i s t haza rd ; and kawakawa, Euthynnus a f f i n i s ) i n Hawaii. O t h e r f i s h e s consumed by s t r i p e d mar l in inc lude mackerels , ca rang ids , s a r d i n e s , h e r r i n g , c o r n e t f i s h e s , and boxf i shes (Ueyanagi and Wares 1975).
Larvae and J u v e n i l e s
There i s only l imi t ed informat ion on t h e food organisms consumed by tuna and b i l l f i s h larvae. S tudies of stomach con ten t s provide only a p a r t i a l l i s t of t h e food e a t e n , s i n c e much of t he d e l i c a t e food items presumed t o comprise t h e d i e t of t h e l a r v a e quick ly become unrecognizable due t o d i g e s t i o n . Nishikawa (1975) noted only smal l numbers of copepods and r e s i d u e s of f i s h l a rvae and c rus t aceans i n t h e stomachs of s k i p j a c k tuna larvae (3-9.5 mm s tandard l eng th (SL)) , and f i s h ' larvae, Euphausia, l a r v a l Decapoda, Amphipoda, and Octopoda i n stomachs of s k i p j a c k tuna j u v e n i l e s (17.5-26 .O mm SL). Lipskaya and Gorbunova (1977) recorded 4 kinds of copepods and l a r v a e of 2 k inds of f l y i n g f i s h i n stomachs of s t r i p e d mar l in l a r v a e and j u v e n i l e s (6.4-29.0 mm S L ) , and 7-9 d i f f e r e n t s p e c i e s of copepods and larvae of 12-15 f i s h s p e c i e s , inc luding those of t h e Fami l ies I s t i o p h o r i d a e , Gempylidae, Scombridae, Coryphaenidae, Holocent r idae , Mull idae, Myctophidae, Engraul idae, and Exocoetidae, i n t h e stomachs of b lue mar l in l a r v a e and j u v e n i l e s (6.0-22.0 mm SL).
Uotani e t a l . (1981) examined stomach con ten t s of numerous (>4,000) l a r v a e of a lbacore , ye l lowf in , bigeye, southern b l u e f i n , Thu_n_nu_g mgno_pii, s k i p j a c k , and f r i g a t e tunas , ranging from 2.21 t o 22.5 mm t o t a l l eng th (TL), from t h e a r e a northwest of A u s t r a l i a . copepods, copepod naup l ius and eggs, Ostracoda, 1 s p e c i e s of Cladocera, and o t h e r Crus tacea ; po lychae te l a r v a e ; f i s h l a rvae ; arrow-worm; and Appendi- c u l a r i a . The stomach con ten t s of four s p e c i e s of tunas ( a lbacore , ye l lowf in , bigeye, and southern b l u e f i n ) c o n s i s t e d mainly of small itans 0.21-0.80 mm body l eng th , but t h a t of s k i p j a c k and f r i g a t e tunas conta ined numerous items exceeding 0.81 mm body length . Feeding s t u d i e s of larval f i s h such as h e r r i n g (B lax te r 1965; Bjoerke 1978; Uotani e t 81. 1978) and anchovy (Ciechomski 1967b; Lasker 19751, show t h a t t h e d i e t a t i n i t i a l feed ing c o n s i s t s l a r g e l y of mollusc l a r v a e , and copepod eggs and n a u p l i i . Other c rus tacean l a r v a e such a s t h e n a u p l i i of ba rnac le s , mollusc larvae t h a t g e n e r a l l y hatch as t rochophores , t rochophores of s a b e l l i d polychae te and larvae of echinoderms a l l may se rve as i n i t i a l food of f i s h l a r v a e (May 1970). I t i s conce ivable t h a t l a r v a e of tunas and b i l l f i s h e s a l s o u t i l i z e s i m i l a r food items a t f i r s t f eed ing , a s evidenced by t h e copepod n a u p l i i found by Uotani e t 01. (1981) and by t h e s u c c e s s f u l experimental r e a r i n g i n tanks of var ious tuna l a r v a e t h a t had been i n i t i a l l y fed r o t i f e r s o r o y s t e r l a r v a e (Hnrada e t a l . 1971; Harada, Murata, and Furu tan i 1973; Harada, Murata, and Miyashi ta 1973; Harada e t a l . 1980).
They recorded 10 spec ie s of
Although l a r v a e of tunas and b i l l f i s h e s may r e q u i r e d e l i c a t e food items a t i n i t i a l feed ing , t h e i r dependence on such food may be f o r a b r i e f per iod only. The unusual ly l a r g e mouth of t hese l a r v a e , which i s approximately 20 t o 32% of body length i n l a r v a e l a r g e r than about 4.0 n u (Gorbunova 1 9 7 5 , f i g u r e s from Ueyanagi 1963) permi ts thun t o f eed on f i s h l a r v a e and o t h e r l a r g e r orgnnisnls much sooner than l a r v a e of o t h e r f i s h e s .

1 2
Vision i s t h e most important sensory f u n c t i o n used i n f eed ing by marine f i s h e s . For s u c c e s s f u l f eed ing and s u r v i v a l , t h e l a r v a e must pe rce ive t h e prey. f i r s t - f e e d i n g larvae of h e r r i n g , p l a i c e , p i l c h a r d , and anchovy, which a l l have r a t h e r small eyes, r e a c t t o prey a t d i s t a n c e s of 0.2-1.0 body l eng th , and t h a t t h e p e r c e p t i v e range i n c r e a s e s wi th growth (Hunter 1980). obse rva t ions have no t been made f o r tuna o r b i l l f i s h l a r v a e , but t h e i r p r o p o r t i o n a t e l y l a r g e r eyes, which range from 10 t o 15% of body l eng th i n l a r v a e 3.2-16.5 mm SL (Gorbunova 1975) , should enab le them t o p e r c e i v e prey a t g r e a t e r d i s t a n c e s . Other t es t s measuring t h r e s h o l d s of l i g h t i n t e n s i t y f o r l a r v a e and j u v e n i l e s of s e v e r a l s p e c i e s of salmon, w h i t e f i s h , p ike , h e r r i n g , and p l a i c e i n d i c a t e a f a l l o f f i n t h e r a t e of f eed ing corresponding wi th l i g h t condit . ions a t dusk and dawn ( B l a x t e r 1969). per iod thus must vary cons ide rab ly with season, l a t i t u d e , c l a r i t y of wa te r , and cloud cover f o r t hese f i s h , and t h i s should apply t o tuna and b i l l f i s h larvae a s w e l l .
S tud ie s on v i s u a l pe rcep t ion by l a r v a e i n d i c a t e t h a t
S i m i l a r
The d a i l y f eed ing
Spawning
Spawning by tunas and b i l If i s h e s have been determined from examina- t i o n of gonads and from t h e c a p t u r e of l a rvae . F igu res 7-10 show t h e maximum area i n which spawning has occurred i n t h e P a c i f i c Ocean, based on t h e c a p t u r e of larvae. throughout t h e y e a r i n t r o p i c a l wa te r s and du r ing t h e summer a t h ighe r l a t i t u d e s . Consequently, s easona l v a r i a t i o n s are ev iden t mainly a t t h e no r the rn and sou the rn e x t r e m e t i e s of t h e spawning area.
Spawning by tunas and b i l l f i s h e s g e n e r a l l y occur
I n t h e western P a c i f i c , spawning by ye l lowf in and bigeye tunas occur s between l a t . 20"N and t h e Equator i n t h e f i r s t q u a r t e r of t h e y e a r , between l a t . 30"N and 10"s i n t h e second q u a r t e r , between la t . 35" and 15"N i n t h e t h i r d q u a r t e r , and between l a t . 15"N and 20"s i n t h e f o u r t h q u a r t e r (Nishikawa e t a l . 1978). I n t h e c e n t r a l P a c i f i c ( long. 140"W t o 180">, yel lowfin tuna spawn throughout t h e yea r between 1st. 14"N and 5"s and i n t h e f i r s t q u a r t e r between l a t . 6 " and 14"s (Matsumoto 1958). e a s t e r n P a c i f i c ye l lowf in tuna spawn throughout t h e yea r between la t . 25" and 7 " N ( K l a w e 1963). The annual spawning p o t e n t i a l of ye l lowf in tuna has been e s t ima ted , on t h e b a s i s of gonad i n d i c e s (Kikawa 19661, as being s l i g h t l y g r e a t e r i n t h e c e n t r a l and e a s t e r n e q u a t o r i a l P a c i f i c t han i n t h e western P a c i f i c , and t h a t of bigeye tuna between long. 100" and 140"W as being double t h a t i n t h e western P a c i f i c west of t h e 180th meridian.
I n t h e
Sk ip jack tuna a l s o spawn a c r o s s t h e e n t i r e P a c i f i c but spawning i n t e n s i t y i s s h a r p l y reduced i n t h e e a s t e r n e q u a t o r i a l P a c i f i c east of long. 12O"W (Klawe 1963). Most of t h e spawning by s k i p j a c k tuna occurs i n e q u a t o r i a l waters between l a t . 1 O " N and 1O"S, and long. 130" t o 145"W, and between l a t . 20"N and 10"s from long. 145"W t o t h e Asian Continent. The c e n t e r of h ighes t spawning l i e s between long. 140"W and 160"E (Matsumoto 1975). Other spawning areas n o r t h of l a t . 20"N inc lude t h e waters around t h e Hawaiian I s l a n d s (Seckel 1972) and south of Japan between l a t . 25" and 30"N (Ueyanagi 1969). A s i n ye l lowf in and bigeye tunas , spawning occur s throughout t h e y e a r i n e q u a t o r i a l waters and du r ing summer a t h ighe r l a t i t u d e s .
c
Tunas spawn i n a f a i r l y wide range of temperatures , va ry ing by species and l o c a l i t y . I n t h e western P a c i f i c , ye l lowf in tuna spawn i n temperatures between 24" and 29°C; bigeye tuna , between 27" and 29°C; and s k i p j a c k tuna , between 23.5" and 30°C (Ueyanagi 1969). I n t h e e a s t e r n P a c i f i c , t h e

13
t empera ture range of ye l lowf in tuna spawning i s between 24" and 32"C, w i t h a marked i n c r e a s e i n spawning as t h e tempera ture i n c r e a s e s above 26" o r 27°C (Klawe e t a l . 1970). The spawning temperatures i n t h e c e n t r a l P a c i f i c around t h e Hawaiian I s l a n d s range from 21.7" t o 27°C f o r ye l lowf in tuna (unpubl ished d a t a , Southwest F i s h e r i e s Center Honolulu Laboratory, Nat ional Marine F i s h e r i e s Se rv ice ) and 22" t o 27°C f o r s k i p j a c k tuna (Matsumoto e t a l . 1984). Hawaiian waters appears t o be c l o s e t o t h e minimum spawning tempera ture f o r t h i s spec ie s . l a r v a e hatched a t tempera tures of 17.2"-20.8"C dur ing l a b o r a t o r y r e a r i n g experiments .
The lower end of t h e tempera ture range f o r ye l lowf in tuna i n
Harada e t a l e (1978) r epor t ed t h a t no normal ye l lowf in tuna
The known spawning a r e a s f o r s t r i p e d and b lue m a r l i n s a r e i n d i c a t e d i n Figure 10. waters e a s t of long. 130"W may be due t o t h e absence of l a r v a l s t u d i e s of t hese f i s h e s from t h a t a r ea . A r e p o r t on t h e sexual m a t u r i t y of b i l l f i s h caught i n t h i s a r e a (Kume and Joseph 1969) shows t h a t females of both s t r i p e d and b lue m a r l i n s i n spawning c o n d i t i o n were p re sen t i n t h e area bounded by la t . 20"N and 3OoS, and long. 1100 and 130"W, wi th t h e southern d i s t r i b u t i o n l i m i t e d t o t h e lower l a t i t u d e s (10"s ) east of ' long. 110"W. i n t h e tunas , spawning presumably occurs throughout t h e yea r a long t h e Equator and du r ing summer, n o r t h and south of t h e Equator.
The n e a r l y complete absence of b i l l f i s h l a r v a e i n e q u a t o r i a l
As
In a l l t h e s p e c i e s mentioned above, t h e mature female r e l e a s e s l a r g e numbers of eggs i n a s i n g l e spawning. The e s t i m a t e s f o r bigeye tuna ranged from 2.9 t o 6.3 m i l l i o n eggs (Yuen 1955); f o r ye l lowf in tuna , 2 t o 8 m i l l i o n eggs (June 1953); f o r s k i p j a c k tuna , 0.1 t o 2 m i l l i o n eggs (Yoshida 1966); and f o r s t r i p e d mar l in , 11 t o 29 m i l l i o n eggs (E ld r idge and Wares 1974). No informat ion i s a v a i l a b l e on t h e f e c u n d i t y of blue. mar l in .
Spawning by tuna i n c a p t i v i t y has been r epor t ed by Kaya e t a l . (1982). Female s k i p j a c k tuna weighing between 1.4 and 2.2 kg spawned i n a tank w i t h i n 8 h a f t e r cap tu re . The spawned ova measured 1.0 mm, t h e same a s a r i p e ova, as compared wi th ovar ian ova of 0.59-0.74 mm i n f i s h taken from t h e same school . The r a p i d development of t h e ova--normal development i s e s t ima ted a t 2-3 months (Matsumoto e t a l . 1984)--was be l i eved t o have been induced by stresses experienced by t h e f i s h dur ing t h e c a p t u r e , t r a n s p o r t , and confinement processes . The v i a b i l i t y of t h e ova was demonstrated by a r t i f i c i a l f e r t i l i z a t i o n and s u c c e s s f u l ha tch ing of t h e l a rvae . The r a p i d ma tu ra t ion and spawning of s k i p j a c k tuna i n c a p t i v i t y i s c u r r e n t l y being e x p l o i t e d t o produce l a r v a l s k i p j a c k tuna f o r experimental s t u d i e s .
Development and Growth of Larvae
A number of tuna s p e c i e s have been r ea red s u c c e s s f u l l y from a r t i f i - c i a l l y f e r t i l i z e d eggs. 1980) , and s k i p j a c k tuna (Inoue e t a l . 1974; Ueyanagi e t a l . 19741, and a l s o b u l l e t t una , &cjt~ ggchgi (= 4. &~&sg=a; Harada, Murata, and Furu tan i 1973) , f r i g a t e tuna , 4. ~l~gggiid (Harada, Murata, and Miyashi ta 19731, and s t r i p e d bon i to , Sgxdg gr_ie_r~&_a&ig (Harada e t a l . 19741, which a r e of minor commercial importance .
These inc lude ye l lowf in tuna (Harada e t a l . 1971,
Yel lowfin and s k i p j a c k tuna embryos developed and hatched i n 23 t o 35 h a f t e r f e r t i l i z a t i o n i n wa te r temperaturea of 24.5O-29"C; whereas those of b u l l e t and f r i g a t e tunas and s t r i p e d b o n i t o developed and hatched in 34 t o 72 h i n water tempera tures of 20°-24"C. Yellowfin and s k i p j a c k tuna l a r v a e

1 4
were 2.6-2.7 mm TL a t ha tch ing , b u l l e t and f r i g a t e tunas 3.3-3.6 mm TL, and s t r i p e d b o n i t o 4.1-4.3 mm TL. As i n o t h e r f i s h e s (Ciechomski 1967a; Fonds e t a l . 19741, t h e r a t e of embryonic development increased wi th temperature . The yolk was absorbed 2 days a f t e r ha tch ing i n s t r i p e d b o n i t o and s k i p j a c k tuna , i n 3 days i n ye l lowf in tuna , and from 2 t o 4 days i n mackerel tuna. The eyes were completely pigmented, t h e mouth was f u l l y formed, and f eed ing began a t t h e t i m e of yolk absorp t ion .
Feeding by tuna l a r v a e r ea red i n tank8 s t a r t e d 2-3 days a f t e r ha tch ing , a t about t h e t i m e of yo lk absorp t ion . The i n i t i a l food of ye l lowf in tuna l a r v a e cons i s t ed of r o t i f e r s , _Bganc_h_ipu_g ~ h g g & l i b , and o y s t e r l a r v a e , Q_aspg&~g_eg gipp5la_ (Harada e t a l . 1971). The larvae were fed s e v e r a l kinds of marine copepods (unspec i f i ed s p e c i e s ) 8 days a f t e r hatching when they were 5-6 mm TL. I n one feeding s tudy (Harada e t a l . 19801, ye l lowf in tuna l a r v a e were fed a d i e t of r o t i f e r s , copepods, and larvae of p a r r o t f i s h , QElegngghgg fggc_iggug, 2.5-4.0 mm TL. Due l a r g e l y t o feeding on f i s h larvae, l a r v a l growth was found t o be e s p e c i a l l y r ap id a f t e r t h e 1 0 t h day. I n examining stomach con ten t s of larval and j u v e n i l e s k i p j a c k tuna taken i n p lankton n e t s , Nishikawa (1975) noted t h a t of 1 5 l a r v a e wi th i d e n t i f i a b l e food remains, 5 l a r v a e (3.0 t o 9.2 mm TL) had consumed copepods and 1 l a r v a (7.0 mm TL) had e a t e n a f i s h larva; t h a t most (73%) of t h e j u v e n i l e s (17.5-26.0 mm TL) had consumed f i s h l a r v a e , whereas o t h e r s had consumed euphaus i id s , copepods, decapods, amphipods, and octopods; and t h a t j u v e n i l e s , 48.0-120 mm, fed almost e n t i r e l y on f i s h l a rvae . Based on t h e s e s t u d i e s , it appears t h a t l a r v a l tunas begin feeding on copepods a t about 3.7 mm body length and cont inue t o do so u n t i l they are a t l e a s t 26 mm i n body l eng th ; l a r v a l f i s h appear i n t h e i r d i e t a t about 6 o r 7 mm TL o r approximately 1 0 days a f t e r ha tch ing; and f i s h larvae and euphaus i ids a r e t h e dominant food items i n j u v e n i l e s 17 t o 26 mm i n TL. S i m i l a r l y , stomach content ana lyses by Lipskaya and Gorbunova (1977) and Gorbunova and Lipskaya (1975) i n d i c a t e t h a t b i l l f i s h larvae i n i t i a l l y feed on copepods and feed on f i s h larvae when they a r e about 6 mm TL.
Growth was extremely r a p i d f o r l a rvae of a l l s p e c i e s from about t h e 1 0 t h day. days ( i n d i c a t i n g a l i n e a r growth r a t e of 1.6 mm/day), i n water tempera tures d e c l i n i n g from 28.6' t o 25.6'C. The t o t a l growth was from 2.7 mm a t ha tch ing t o 51 mm i n 38 days. B u l l e t and f r i g a t e tunas grew even f a s t e r , 3.2-3.5 mm/day i n temperatures r i s i n g from 25.4' t o 28.3'C. F r i g a t e tuna grew from 3.3 mm a t ha tch ing t o 120 mm i n 33 days , and b u l l e t tuna from 3.3 t o 157 mm i n 52 days. S t r i p e d b o n i t o showed t h e f a s t e s t growth r a t e , 6.4 mm/day from t h e 1 0 t h t o 42d day, i n tempera tures i n c r e a s i n g from about 20" t o 28°C. days. Fonds e t a l . (1974) i n d i c a t e d t h a t the growth r a t e of g a r f i s h , B_g&gg bglogg, l a r v a e w a s l i n e a r l y c o r r e l a t e d w i t h temperature . I f t h i s ho lds f o r tuna l a r v a e as w e l l , the growth r a t e of ye l lowf in tuna l a r v a e should have been g r e a t e r than r epor t ed above. larvae i s not known s i n c e l a r v a e of t h i s s p e c i e s have not been r ea red beyond 5 days.
A 10-day-old ye l lowf in tuna l a r v a grew from 5 . 8 t o 51 mm i n 28
The t o t a l growth was from 4.1 mm a t ha tch ing t o 290 mm i n 99
Growth of s k i p j a c k tuna
15
THE DEEP OCEAN M I N I N G ENVIRONMENTAL STUDY (DOMES) AREA
Env i ro nme n t
Oceanographic F e a t u r e s
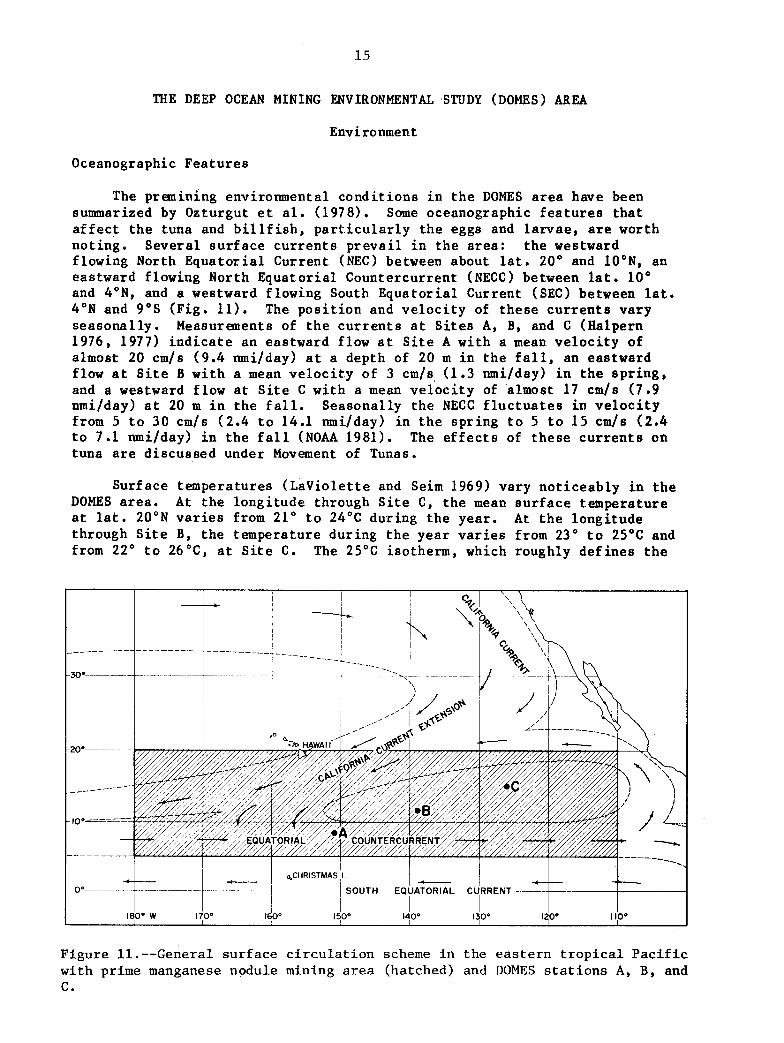
The premining environmental c o n d i t i o n s i n t h e DOMES area have been summarized by Ozturgut e t a l . (1978). Some oceanographic f e a t u r e s t h a t a f f e c t t h e tuna and b i l l f i s h , p a r t i c u l a r l y t h e eggs and l a r v a e , are worth no t ing . Seve ra l s u r f a c e c u r r e n t s p r e v a i l i n t h e a r e a : t h e westward flowing North E q u a t o r i a l Current ( N E C ) between about l a t . 20" and 10"N, an eastward flowing North Equa to r i a l Countercurrent (NECC) between la t . 10" and 4"N, and a westward flowing South E q u a t o r i a l Current (SEC) between l a t . 4"N and 9 " s (F ig . 11). The p o s i t i o n and v e l o c i t y of t h e s e c u r r e n t s va ry s e a s o n a l l y . Measurements of t h e c u r r e n t s a t Si tes A, B, and C (Halpern 1976, 1977) i n d i c a t e an eastward flow a t S i t e A w i t h a mean v e l o c i t y of almost 20 cm/s (9.4 nmi/day) a t a depth of 20 m i n t h e f a l l , an eastward flow a t S i t e B w i t h a mean v e l o c i t y of 3 cm/s (1.3 nmi/day) i n t h e s p r i n g , and a westward f low a t S i t e C w i th a mean v e l o c i t y of almost 17 cm/s (7.9 nmi/day) a t 20 m i n t h e f a l l . Seasonal ly t h e NECC f l u c t u a t e s i n v e l o c i t y from 5 t o 30 cm/s (2.4 t o 14.1 nmi/day) i n t h e s p r i n g t o 5 t o 1 5 cm/s (2.4 t o 7.1 nmi/day) i n t h e f a l l (NOAA 1981). The e f f e c t s of t h e s e c u r r e n t s on tuna are d i scussed under Movement of Tunas.
Surface temperatures ( L a V i o l e t t e and Seim 1969) vary n o t i c e a b l y i n t h e DOMES area. a t l a t . 20"N v a r i e s from 21" t o 24°C during t h e yea r . A t t h e long i tude through S i t e B, t h e temperature du r ing t h e yea r v a r i e s from 23" t o 25°C and from 22" t o 26"C, a t S i t e C.
A t t h e long i tude through S i t e C , t h e mean s u r f a c e temperature
The 25°C isotherm, which roughly d e f i n e s t h e
.9-- I ~CHRISTMAS,l ~~ ~ SOUTH EQUATORIAL i - CURRENT I +-+. I I
0"- ~
1800 w I 70° 150° 140° 130' 130' I100
Figure 11.--General s u r f a c e c i r c u l a t i o n scheme i n the e a s t e r n t r o p i c a l P a c i f i c with prime manganese nodule mining area (hatched) and DOMES s t a t i o n s A, B, and C.

1 6
l i m i t of l a r v a l c a p t u r e , l i e s t o t h e n o r t h of S i t e s A and B dur ing t h e e n t i r e year, but only f o r 6 months a t S i t e C. The thermal s t r u c t u r e i n t h e DOMES area i s t y p i c a l of t h e t r o p i c a l P a c i f i c w i t h a w e l l - d e f i n e d . s u r f a c e mixed l a y e r o v e r l y i n g a s t r o n g thermocl ine (F ig . 1 2 ) . The mean depth of t h e mixed l a y e r was 35532 m du r ing summer and 55218 dur ing w i n t e r (Ozturgut e t a l . 1978).
Measurements of suspended p a r t i c u l a t e m a t t e r (SPM) i n t h e DOMES reg ion made du r ing t h e summer of 1975 i n d i c a t e t h a t SPM i s most abundant i n s u r f a c e waters (mean c o n c e n t r a t i o n of 42220 ~ . l g / R ) , and uniformly low, 1053 w/R, below 200 m (Baker e t a l . 1979). The concen t r a t ion i n s u r f a c e waters i s considered low but t y p i c a l of open ocean waters. The g e n e r a l f e a t u r e s of SPM concen t r a t ions change ve ry l i t t l e between summer and w i n t e r . Most of t h e p a r t i c l e s (9122.9%) a re smaller than 4 p n w i t h mean medium d iame te r s of 2.020.4 pm.
RTHWEST (NW 1 NORTHEAST (NE 1
__________-----_---
SOUTHEAST (SE 1
- N
10
IC
!C
IC
C
IC
?(
x
K
- Figure 12.--Assumed movement of t h e v a r i o u s s t o c k s of s k i p j a c k tuna i n t h e P a c i f i c Ocean. The numerals a long t h e mig ra to ry r o u t e s r e p r e s e n t q u a r t e r s and l o c a t i o n s . of h igh CPUE cells of s k i p j a c k tuna taken by t h e Japanese tuna l o n g l i n e f i s h e r y , 1964-67. Stock d e s i g n a t i o n s are shown i n pa ren theses (Matsumoto 1975).
B i o l o g i c a l F e a t u r e s
Zooplankton r e s e a r c h i n t h e DOMES a r e a a t Si tes A , B, and C was done i n t h e summer of 1975 and w i n t e r of 1976 ( H i r o t a 1977). Information on microzooplankton was l i m i t e d t o v e r t i c a l d i s t r i b u t i o n . The major f e a t u r e s of d i s t r i b u t i o n of microzooplankton (0.183-0.035 mm d i ame te r ) were: n a u p l i i , t i n t i n n i d s , and Micgggktgllg (copepod) tended t o have maxima shal lower than 40-60 m ; ( 2 ) gpcgag (copepod) tended t o have a much deeper
( 1 )

1 7
maximum, o f t e n a t 100 m o r more; and ( 3 ) r a d i o l a r i a and f o r a m i n i f e r a showed r e l a t i v e l y smaller dep th v a r i a t i o n than ( 1 ) o r ( 2 ) above. t i n t i n n i d s , and M.cgegg&glJg were t h e most l i k e l y t o be a s s o c i a t e d w i t h t h e c h l o r o p h y l l maximum, which occurred a t dep ths of 69224 m i n summer and 54230 m i n w i n t e r . The approximate ranges of numerical abundance of each of t h e f i v e t a x a were 3,000-15,000/m3 f o r n a u p l i i , 1,000-15,000/m3 f o r t i n t i n n i d s , 1 ,000-10,000/m3 f o r r a d i o l a r i a n s and f o r a m i n i f e r s , 500-5,000/m3 f o r Qgcggg, and 100-500/m3 f o r l$cg~ggtgl lg . about a t h i r d of those i n t h e eastern P a c i f i c (EASTROPAC) area (Beers and Stewart 1971) and overlapped those i n Hawaiian waters (Gundersen e t a l . 1976). t h e upper 160 m was 145/11 of c i l i a t e s and 5/R of copepod n a u p l i i and f o r a m i n i f e r s . The microplankton s t a n d i n g s t o c k as " t i s s u e volumes'' i n t e g r a t e d over t h e 0-150 m layer showed a range of 423-852 mm3/m2. volumes were approximately one-half of t h e volumes r e p o r t e d f o r Hawaiian waters (Gundersen e t a l . 1976) and one-third of t hose f o r t h e EASTROPAC area along long. 105"W (Beers and Stewart 1971).
The n a u p l i i ,
The abundance estimates were
The average abundance of microzooplankton smaller than 0.035 mm i n
These
The average macrozooplankton s t and ing s t o c k s ( d r y weight) i n t e g r a t e d 2 over t h e 0-200 m depth l a y e r ranged from 1 ,048 t o 2,900 mg/m a t t h e t h r e e
s i t e s t a t i o n s ( H i r o t a 1977). The h ighes t concen t r a t ions of macrozooplank- t o n were i n t h e upper 150 m and moderate c o n c e n t r a t i o n s between 200 and 900 m. The areal d i s t r i b u t i o n of t h e s t and ing s t o c k s (neuston n e t c a t c h e s ) showed a g r a d i e n t from sou theas t t o no r thwes t , approximately normal t o t h e DOMES s t a t i o n s i tes . B and C and t h e lowest a long S i t e A and t h e t o p h a l f of S i t e C.
The h i g h e s t s t o c k s were a t t h e sou the rn h a l f of S i t e s
The major t a x a of macrozooplankton were copepods, euphaus i id s , chae togna ths , l a rvaceans , o s t r a c o d s , pteropods, and larval f i s h e s ( H i r o t a 1977). The mean abundance of copepods i n the upper 100 m ranged from 152,000/1,000 m3 a t S i t e A t o 296,000/1,000 m3 a t S i t e B i n 1975, and 171,000/1,000 m3 a t S i t e A t o 205,000/1,000 m3 a t S i t e C i n 1976. The most common s p e c i e s were t h e small ( c a . 1 mm TL) cyclopoids (Qgcggg and _O&Bpng> and c a l a n o i d s ( E g g g c g l g g g g , cglecglg~gg, and Clausocalanus) , whose abundance8 ranged from 1,000 t o 59,000/1,000 m . euphaus i id s i n the neuston l a y e r ranged from 1,400/1,000 m3 a t S i t e A t o 2,900/1,000 m3 a t S i t e 8. a t S i t e B ; whereas copepods were n e a r l y equa l ly abundant a t S i t e s A and C, and euphaus i id s least abundant a t S i t e A.
i n t h e neuston layer and 20,000 t o 33,000/1,000 m3 i n t h e 0- t o 200-01 depth layer ( H i r o t a 1977).
18,000/1,000 m3 f o r o s t r a c o d s i n t h e two depth layers. of chaetognaths ranged from 2,000 t o 9,000/1,000 m3. abundance were noted between si tes and years.
-3-----------
Both copepods and euphaus i id s were most abundant
The main abundance of
The mean abundance of l a rvaceans ranged from 18,000 t o 250,000/1,000 m 3
Corresponding va lues f o r pteropods were 3 000- 16,000/1,000 m3 and 2,000-5,000/1,000 m3, and 200-1,500/1,000 m 3 and 5,000-
The mean abundance Large d i f f e r e n c e s i n
F i s h l a r v a e from t h e neuston layer were predominantly Hemiramphidae and Exocoetidae with a few midwater forms, mainly Gonostomatidae, and a number of n e a r s u r f a c e f i s h (Carangidae, Scombridae, Bramidae, and Scomberesocidae). midwater forms (Gonostomatidae and Myctophidae) and low c o n c e n t r a t i o n s of n e a r s u r f a c e f i s h (Scombridae, Coryphaenidae, Hemiramphidae, and Exocoet idae) . Three t a x a , Hemiramphidae, Exocoetidae, and Gonostomatidae, were abundant a t a l l s i tes whereas Scombridae were w e l l r e p r e s e n t e d a t S i t e
Larvae from t h e 0- t o 100-m depth layer included mainly

18
A, Carangidae and Bramidae a t S i t e B, and Scomberesocidae a t S i t e C. The mean abundance of a l l f i s h larvae i n t h e 0- t o 100-m depth l a y e r ranged from 700 t o 900/1,000 m3 i n 1975 and from 300 t o 400/1,000 m3 i n 1976. The families of commercial i n t e r e s t included t h e Scombridae ( t u n a s and mackerels) and I s t i o p h o r i d a e ( b i l l f i s h e s ) , t h e l a t t e r r e p r e s e n t e d by a s i n g l e specimen t aken a t S i t e A ( H i r o t a 1977).
Tuna8 and B i l l f i s h e s i n t h e DOMES Area
Information on f i s h e s in t h e DOMES area has been reviewed by Blackburn (1976). s k i p j a c k t u n a s and s t r i p e d and b l u e mar l in s . small q u a n t i t i e s i n c l u d e t h e s a i l f i s h , I s t i o p h o r u s p l a t y p t e r u s , s h o r t b i l l s p e a r f i s h , T e t r a p t u r u s a n g u s t i r o s t r i s , and b l a c k mar l in , Makaira ind ica . There a re f i v e commercial f i s h e r i e s f o r t unas or b i l l f i s h e s i n t h e area: (1) t h e Japanese l o n g l i n e f i s h e r y , which f i s h e s throughout t h e area wi th t h e except ion of t h e n o r t h e a s t e r n c o r n e r ( l a t . 10"-15"N, long. 115"-130"W); ( 2 ) t h e Japanese l i v e - b a i t and pu r se s e i n e f i s h e r i e s f o r s k i p j a c k and small ye l lowf in tunas i n t h e South P a c i f i c , which i n r e c e n t y e a r s ( p a r t i c u l a r l y t h e pu r se s e i n e r e ) have extended eastward along t h e E q u a t o r i a l Counter- c u r r e n t t o long. 169"W (Tanaka 1978); (3) t h e United S t a t e s l i v e - b a i t and pu r se s e i n e f i s h e r i e s f o r s k i p j a c k and small y e l l o w f i n tunas , which i n r e c e n t years ( p a r t i c u l a r l y by t h e s e i n e r s ) have expanded westward t o long. 150"W between l a t . 15"N and 5"s (Matsumoto e t a l . 1984); (4) t h e Hawaiian l i v e - b a i t f i s h e r y f o r s k i p j a c k tuna , which f i s h e s a t or j u s t o u t s i d e t h e n o r t h e r n edge of t h e DOMES area; and ( 5 ) t h e Hawaiian l o n g l i n e f i s h e r y f o r ye l lowf in and b igeye tunas and b i l l f i s h e s . c a t c h e s i n metric t o n s (MT) i n 1973 (Blackburn 1976) were approximately:
Commercial f i s h e r i e s i n t h e area e x i s t f o r ye l lowf in , bigeye, and Other b i l l f i s h e s t aken i n
The DOMES area commercial
Yel lowfin tuna 62,700 MT (57,400 MT United S t a t e s pu r se s e i n e and l o n g l i n e ; 5,300 MT Japanese l o n g l i n e )
Bigeye tuna 16,600 MT (Japanese l o n g f i n e )
B i l If ishes 5,500 MT (Japanese l o n g l i n e )
Bigeye tuna and b i l l f i s h e s 300 MT (Hawaiian l o n g l i n e )
Skip jack tuna 6,700 MT (United S t a t e s purse s e i n e and l i ve b a i t )
T o t a l 91,800 MT
The t o t a l c a t c h would b e much g r e a t e r today i f t h e Japanese s k i p j a c k tuna c a t c h east o f t h e 180 th meridian--the eastward expansion t o long. 160"-165"W began i n 1977 (Tanaka 1978)--and t h e c a t c h of t h e Hawaiian r e c r e a t i o n a l f i s h e r y were included. tuna f i s h e r y and t h e Hawaiian r e c r e a t i o n a l f i s h e r y i n t h e DOMES area are n o t r e a d i l y a v a i l a b l e ; however, it was e s t ima ted (Honolulu Laboratory s t a f f ) t h a t t h e l a t t e r caught 996 MT of b i l l f i s h e s and 2,501 MT of tunas , mos t ly y e l l o w f i n and s k i p j a c k , i n 1976.
Catches of bo th t h e Japanese s k i p j a c k
Larvae of ye l lowf in , bigeye, and s k i p j a c k t u n a s and s t r i p e d and b l u e m a r l i n s are g e n e r a l l y p r e s e n t i n t h e DOMES area (F igs . 7-10). a re larvae of s p e c i e s of f i s h t aken i n smaller q u a n t i t i e s , i .e. , f r i g a t e tuna , s a i l f i s h , and s h o r t b i l l s p e a r f i s h . Larvae of ye l lowf in and s k i p j a c k
Also p r e s e n t

19
t unas are most abundant around Hawaii i n summer and a b s e n t i n winter, In o t h e r sectors of t h e DOMES area n o r t h of la t . 15"N, l a r v a l abundance is also seasonal . I n t h e s e c t o r west of long. 150"W between la t . 14" and. 5"N, larvae are abundant throughout t h e yea r and t h e i r abundance peaks d u r i n g w i n t e r (Matsumoto 1958). Between long. 150" and 130"W, larvae of ye l lowf in tuna are g e n e r a l l y r e s t r i c t e d t o sou th of l a t . 10"N. Th i s i s more pronounced f o r s k i p j a c k tuna l a r v a e (F ig . 9 ) . A s mentioned p rev ious ly , t h e d i p i n t h e d i s t r i b u t i o n t o l a t . 10°N i n t h i s s e c t o r may be due t o a s i m i l a r southward s h i f t of t h e s u r f a c e i so therms, bu t it i s a180 due p a r t l y t o t h e low level of sampling n o r t h of l a t . 10"N. East of long. 130°W, larvae of ye l lowf in tuna a re taken as f a r n o r t h as l a t . 20"N dur ing summer (Matsumoto 1958; S t r a sburg 1960; Klawe 19631, b u t l a r v a e of s k i p j a c k tuna occur only occae iona l ly and in small numbers (Klawe 1963). i n a similar manner as those of ye l lowf in tuna bu t a t lower levels of abundance. s k i p j a c k tuna l a r v a e are t h e most abundant: t h e catches are twofold t o t h r e e f o l d g r e a t e r than t h a t of ye l lowf in tuna l a r v a e (Matsumoto 1958; Higgins 1970).
Larvae of b igeye tuna occur
In Hawaii and t h e e q u a t o r i a l P a c i f i c , i nc lud ing t h e DOMES area,
Larvae of s t r i p e d and blue marlins a re found throughout t h e nor th- sou th range of t h e DOMES area west o f long. 145"W (F ig . 10). The appa ren t absence of larvae east of long. 145"W may be t h e r e s u l t of low p r i o r i t y g iven t o s t u d i e s of b i l l f i s h l a r v a e i n t h a t reg ion . s t u d i e s of la rva l f i s h i n t h e EASTROPAC area have been made (Ahlstrom 1971, 19721, t h e s t u d i e s were based s o l e l y on c o l l e c t i o n s ob ta ined from o b l i q u e h a u l s t o 200 m. C o l l e c t i o n s ob ta ined from s u r f a c e h a u l s , which produce t h e most larval b i l l f i s h , have no t been analyzed. Thus f a r , larvae of on ly one s p e c i e s of lesser commercial importance, s h o r t b i l l s p e a r f i s h , have been taken a t one s t a t i o n i n t h e DOMES area a t approximately l a t . 19"N, long. 137"W (Howard and Ueyanagi 1965).
Although d e t a i l e d
Movement of Tunas
Hypotheses and obse rva t ions on tuna movement i n t h i s r eg ion i n c l u d e those of Rothschi ld (19651, Blackburn and Laurs (19721, Seckel (1972) , Williams (19721, and Matsumoto (1975). These a u t h o r s v a r i o u s l y s t r e s s e d t h e probable importance of food sources , ocean c u r r e n t s , and tempera ture i n t h e migra tory behavior of s k i p j a c k tuna.
It i s g e n e r a l l y accepted t h a t t unas , mainly s k i p j a c k , mig ra t e eastward from t h e e a s t e r n h a l f of t h e DOMES area along t h e NECC. These f i s h e n t e r t h e e a s t e r n P a c i f i c no r the rn f i s h e r y o f f c e n t r a l Mexico i n t h e s p r i n g , proceed northward a long Baja C a l i f o r n i a as t h e season p rogres ses , and r e t u r n sou th a long t h e coast i n t h e f a l l . t h e e q u a t o r i a l c e n t r a l P a c i f i c t o spawn (Fink and B a y l i f f 1970).
The major p o r t i o n of t h e f i s h r e t u r n t o
Williams (1972) proposed t h r e e mig ra t ion models i nvo lv ing t h e movement of larvae, young and a d u l t s k i p j a c k tuna from t h e e q u a t o r i a l P a c i f i c west of long. 130"W t o t h e e a s t e r n P a c i f i c no r the rn f i s h e r y i n s p r i n g and t h e i r r e t u r n t o t h e c e n t r a l P a c i f i c a t t h e end of t h e season. According t o t h e model, t h e NECC provides t h e eastward t r a n s p o r t of l a r v a e and young, and t h e C a l i f o r n i a Curren t Extension p rov ides t r a n s p o r t f o r t h e i r r e t u r n as a d u l t s t o t h e e q u a t o r i a l spawning grounds west of long. 130"W. proposed t h e e x i s t e n c e of a number of semi-independent s k i p j a c k tuna stocks i n t h e P a c i f i c Ocean by fo l lowing t h e q u a r t e r l y movement of t h e a r e a e of h igh c a t c h rates i n t h e Japanese l o n g l i n e f i s h e r y over a per iod of 4 years
Matsumoto (1975)

20
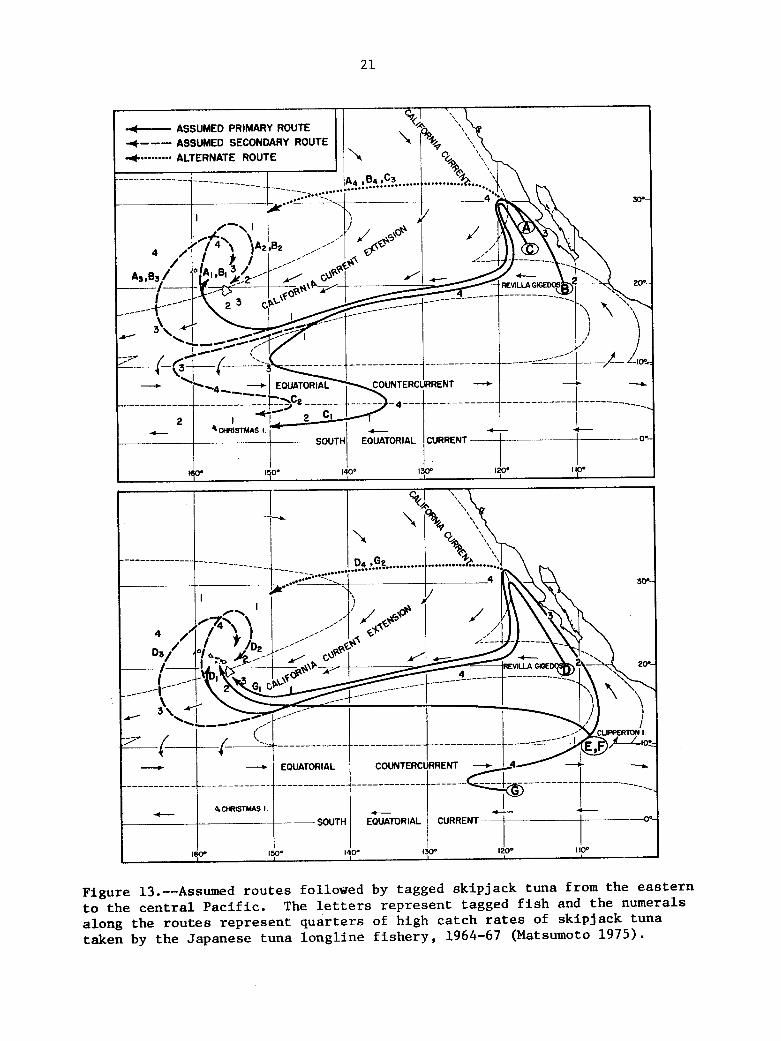
(1964-67). (Northern and Sou the rn ) , w i th t h e migratory pa ths of t h e s t o c k s fol lowing a clockwise d i r e c t i o n i n t h e c e n t r a l and western North P a c i f i c and a counterclockwise d i r e c t i o n i n t h e e a s t e r n North P a c i f i c and Southern Hemisphere, corresponding wi th t h e s u r f a c e c u r r e n t f low of t h e major water masses (Fig. 12 ) . taken by s k i p j a c k tuna tagged i n t h e e a s t e r n P a c i f i c and cap tu red i n Hawaii and Christmas I s l a n d by p r o j e c t i n g t h e d u r a t i o n of l i b e r t y of t h e s e f i s h t o t h e time s c a l e and d i r e c t i o n of movement i n d i c a t e d i n t h e proposed model (F ig . 13, primary and assumed secondary r o u t e s ) . The most l i k e l y pa ths a l l pas s through t h e DOMES a r e a .
H e recognized approximately seven s tocks i n each hemisphere
Matsumoto f u r t h e r a t tempted t o f i t t h e assumed routes
EFFECTS OF M I N I N G
Impa c t on Environment
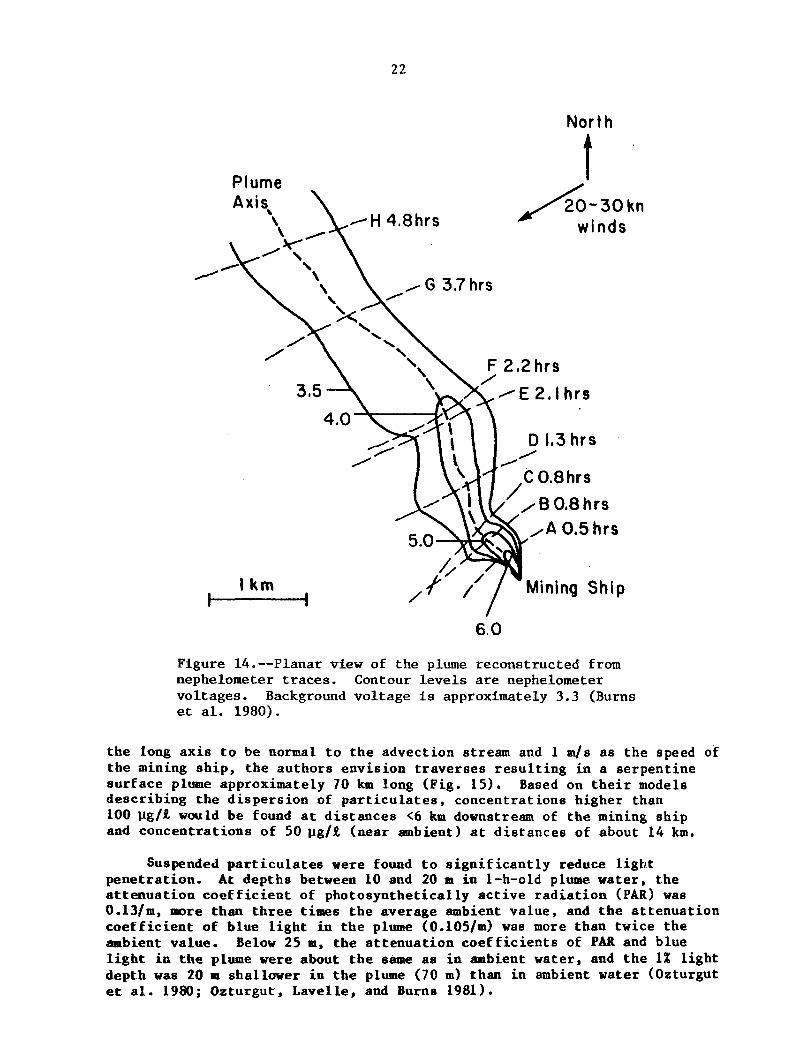
Most of t h e concern on seabed mining relates t o t h e s u r f a c e and b e n t h i c d i s c h a r g e s and t h e i r p o s s i b l e e f f e c t s on the b i o t a . d i s c h a r g e and surf ace plume r e s u l t i n g from tes t mining o p e r a t i o n s i n d i c a t e d t h a t t he d i s c h a r g e c o n s i s t e d of sediment, bottom water, nodule fragments, and b e n t h i c b i o t a . Sediment was t h e major component of t h e p a r t i c u l a t e s d i scha rged , of which 68-87% by weight was p e l a g i c c l a y , and t h e r e s i d u a l was p r i m a r i l y calcium ca rbona te , sea s a l t , and s i l i c o n d iox ide (Bischoff e t a l . 1979; Ozturgut , L a v e l l e , and Erickson 1981). Measurements of t h e d i s c h a r g e were made du r ing mining tests conducted by Ocean Mining Assoc ia t e s (OMA) and Ocean Management, Inc . (OMI). I n t h e OMA tes ts , t h e t o t a l p a r t i c u l a t e c o n c e n t r a t i o n i n t h e shipboard d i scha rge averaged 5.8 g/R. The d i scha rge passed ove r t h e s i d e , f e l l t o t h e s e a , and mixed r a p i d l y wi th t h e s u r f a c e water. A l a r g e percentage of t h e sediment p a r t i c u l a t e s were found t o s e t t l e ou t of t h e eupho t i c zone w i t h i n a few hours , but t h e f i n e f r a c t i o n of t h e d i s c h a r g e was presumed t o be long-lived i n t h e mixed l a y e r . near s u r f a c e c o n c e n t r a t i o n of p a r t i c u l a t e s a t t h e t r a n s e c t c l o s e s t t o t h e mining s h i p (15 min a f t e r i n i t i a l mixing o r 75 m behind t h e s h i p ) was 900 %/R (Ozturgut e t a l . 1980). mixed l a y e r averaged 443 p g / R a f t e r 1 h, 1 7 7 w/R a f t e r 2 h, and 69 yg/R a f t e r 3 h, w i t h ambient c o n c e n t r a t i o n being 28.5 p g / R . I n t h e OM1 tes ts , t h e p a r t i c u l a t e c o n c e n t r a t i o n s were 598 w/R a f t e r 1 h, 55 ~ . l g / R a f t e r 5 h, 45 p g / R a f t e r 28 h , and 34 pg/k a f t e r 37 h, and t h e ambient c o n c e n t r a t i o n was 37 p g / t ( L a v e l l e e t a l . 1982). The s u r f a c e plume measured du r ing t h e OM1 tests (Burns e t a l . 1980) , had t r a v e l e d about 4.5 km from t h e mining s h i p i n <5 h and was 1 km wide a t t h a t po in t (F ig . 14 ) .
I n v e s t i g a t i o n s on the
The maximum
The p a r t i c u l a t e concen t r a t ions i n t h e
During f u l l - s c a l e commercial mining, t he miner w i l l d i s cha rge t h r e e t i m e s t h e volume of t h e t e s t miner and a t h ighe r bulk d e n s i t i e s , but t h e mining s h i p speed ( e s t ima ted a t 1 m / s ) w i l l be f o u r t i m e s t h a t of t h e average tes t miner speed, so t h a t d i scha rge volumes pe r u n i t ocean area w i l l be s l i g h t l y decreased over t ha t of t h e mining tests ( L a v e l l e and Ozturgut 1981 1.
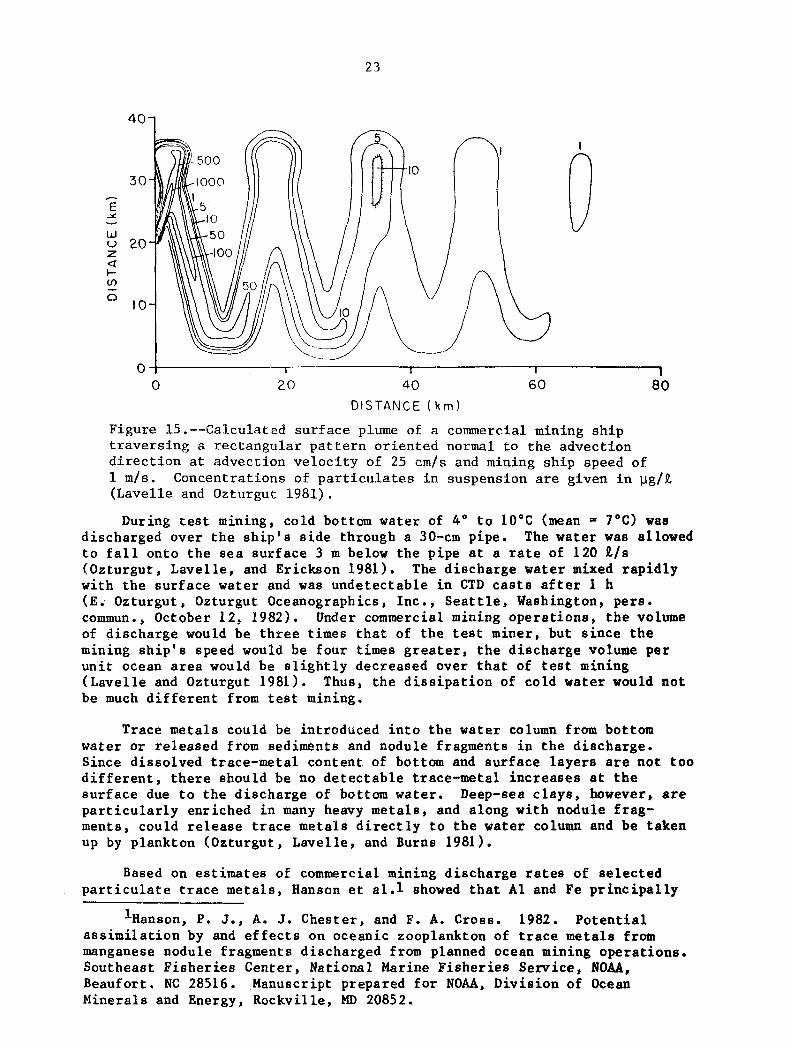
The p a t t e r n t h a t commercial mining s h i p s make a c r o s s a mining area w i l l be one t h a t maximizes t h e sea f l o o r recovery of nodules. Thus Lave l l e and Ozturgut (1981) e n v i s i o n t h a t 30- by 30-km mining t r a c t s w i l l be s u c c e s s f u l l y and i n t e n s i v e l y mined, and t h a t w i t h i n t h i s a r e a , t h e mining s h i p w i l l t r a v e r s e a r e c t a n g u l a r p a t t e r n 30 km on t h e long s i d e and 3.6 km on t h e s h o r t e r s i d e , w i th such r e c t a n g u l a r p a t t e r n s o f f s e t by 40 m. Taking
2 1
I 150. I b I200 I io- I
4 1
Ds,'
4) 3'..
--/*I*----' /- - \ 4
77- -6- --t
c-
Figure 13.--Assumed routes followed by tagged skipjack tuna from the eastern to the central Pac i f i c . along the routes represent quarters of high catch rates of skipjack tuna taken by the Japanese tuna longline f i shery , 1964-67 (Matsumoto 1975).
The letters represent tagged f i s h and the numerals
22
North
t Plume A x i s \
I krn H
/*-3Okn winds
6.8
Figure 14.--Planar v i e w of t h e plume r econs t ruc t ed from nephelometer traces. vo l t ages . Background v o l t a g e is approximately 3.3 (Burns et al. 1980).
Contour l e v e l s are nephelometer
t h e tong axis t o be normal t o t h e advec t ion stream and 1 m / s as t h e speed of t h e mining s h i p , t h e a u t h o r s e n v i s i o n t r a v e r s e s r e s u l t i n g i n a s e r p e n t i n e s u r f a c e plume approximately 7 0 km long (F ig . 15) . d e s c r i b i n g t h e d i s p e r s i o n of p a r t i c u l a t e s , concen t r a t ions h ighe r than 100 and c o n c e n t r a t i o n s of 50 pg/% (nea r ambient) a t d i s t a n c e s of about 14 km.
Eased on t h e i r models
would be found a t d i s t a n c e s <6 km downstream of t h e mining s h i p
Suspended p a r t i c u l a t e s were found t o s i g n i f i c a n t l y reduce l i g h t p e n e t r a t i o n . A t dep ths between 10 and 20 m in l-h-old plume wa te r , t h e a t t e n u a t i o n c o e f f i c i e n t of p h o t o s y n t h e t i c a l l y a c t i v e r a d i a t i o n (PAR) wa8 0.13/m, more than t h r e e times t h e average ambient va lue , and t h e a t t e n u a t i o n c o e f f i c i e n t of b lue l i g h t i n t h e plume (0.105/m) was more than twice t h e ambient va lue . Below 25 m, t h e a t t e n u a t i o n c o e f f i c i e n t s of PAR and b lue l i g h t i n t h e plume were about t h e saute as i n ambient water , and t h e 1% l i g h t dep th w a s 20 IU sha l lower i n t h e plume (70 m) than i n ambient water (Ozturgut e t al. 1980; Ozturgut , Lave l l e , and Burns 1981).
23
o l I I 1 1 0 20 40 60 80
DISTANCE ( k m )
Figure 15.--@alculated s u r f a c e plume of a commercial mining s h i p t r a v e r s i n g a r e c t a n g u l a r p a t t e r n o r i e n t e d normal t o t h e advec t ion d i r e c t i o n a t advec t ion v e l o c i t y of 25 c m / s and mining s h i p speed of 1 m / s . (EavelPe and Ozturgut 1981).
Concentrat ions of p a r t i c u l a t e s i n suspension are g iven i n p g / t
During test mining, cold bottom water of 4" t o 10°C (mean = 7 ° C ) was discharged ove r t h e s h i p ' s s i d e through a 30-cm pipe. t o f a l l on to t h e sea s u r f a c e 3 m below t h e p ipe a t a ra te of 120 f i l s (Ozturgut , Lave l l e , and Erickson 1981). The d i scha rge water mixed r a p i d l y wi th t h e s u r f a c e water and was u n d e t e c t a b l e i n CTD c a s t s a f t e r 1 h ( E . Ozturgut , Ozturgut Bceanographics, Inc . , S e a t t l e , Washington, p e r s b commun., October 12 , 1982). Under commercial mining o p e r a t i o n s , t h e volume of d i scha rge would be t h r e e times t h a t of t h e tes t miner , but s i n c e t h e mining s h i p ' s speed would be fou r times g r e a t e r , t h e d i scha rge volume p e r u n i t ocean area would be s l i g h t l y decreased over t h a t of t es t mining ( L a v e l l e and Ozturgut 1981). Thus, t h e d i s s i p a t i o n of co ld water would n o t be much d i f f e r e n t from tes t mining.
The water was allowed
Trace metals could be introduced i n t o t h e water column from bottom water o r r e l e a s e d from sediments and nodule fragments i n t h e d i scha rge . Since d i s so lved trace-metal con ten t of bottom and s u r f a c e l a y e r s a re n o t t o o d i f f e r e n t , t h e r e should be no d e t e c t a b l e trace-metal i n c r e a s e s a t t h e s u r f a c e due t o t h e d i scha rge of bottom water. Deep-sea c l a y s , however, are p a r t i c u l a r l y enr iched i n many heavy metals, and along w i t h nodule f r a g - ments, could release trace metals d i r e c t l y t o t h e water column and be t aken up by p l ank ton (Ozturgut , Lave l l e , and Burns 1981).
Based on estimates of commercial mining d i s c h a r g e rates of s e l e c t e d
laanson, P. J., A. J. Ches te r , and F. A. Cross.
p a r t i c u l a t e t r a c e metals, Hanson e t a l .1 showed t h a t A 1 and Fe p r i n c i p a l l y
1982. P o t e n t i a l a s s i m i l a t i o n by and e f f e c t s on ocean ic zooplankton of t r a c e metals from manganese nodule fragments d i scha rged from planned ocean mining ope ra t ions . Southeast F i s h e r i e s Center , Nat ional Marine F i s h e r i e s Se rv ice , NOM, Beaufort . NC 28516. Manuscript prepared f o r NOAA, D iv i s ion of Ocean Minerals and Energy, Rockvi l le , MD 20852.

24
from sediments and Mn from nodule fragments were t h e most abundant (d i scha rge rates of 1,100-1,700 g / s ) ; followed by Ba, N i , Cu, and Co (12-77 g / s > ; and Zn, Pb, Ma, V, and C r (1.4-9.5 g / s ) . By i n t e g r a t i n g t h e d i scha rge rates over t h e mining y e a r and by comparing them wi th t h e annual atmospheric d e p o s i t i o n ra tes , which were t h e p r i n c i p a l s teady s t a t e i n p u t s of t r a c e metals t o t h e mixed l a y e r , t h e author8 demonstrated t h a t t h e d i scha rge rates exceeded t h e d e p o s i t i o n r a t e s by s e v e r a l o r d e r s of magnitude and concluded t h a t t h e mining d i scha rge could s i g n i f i c a n t l y inc rease t h e l e v e l s of some metals and a l t e r t h e t r a c e metal concen t r a t ions .
Experiments w i t h resuspended bottom sediments by Rosenbauer and Bigchoff (19781, however, i n d i c a t e d t h a t a l l concen t r a t ions of Ni, Zn, C r , Fe, Cu, Mn, and A1 were below d e t e c t i o n l imi t s i n oxygenated seawater and only Mn and N i were r e l e a s e d i n i n s i g n i f i c a n t l y small q u a n t i t i e s (0.03 ppm Ni, 0.03-2 ppm Mn) i n deoxygenated seawater. a s s e s s t h e amount and d i r e c t i o n of metal exchange between a r t i f i c i a l seawater and crushed Mn nodules (Benjamin and Felmy 1981) i n d i c a t e d t h a t i n t h e pH range of seawater, t h e amounts of Co, Cu, and Cd r e l e a s e d were below t h e a n a l y t i c a l l y d e t e c t a b l e l i m i t , even i n t h e most concen t r a t ed s l u r r y t e s t e d (20 g/R).
Other experiments conducted t o
Impact on Pr imary Product i on
Impact of mining on phytoplankton pho tosyn thes i s and primary product ion was examined by Chan and Anderson (1981) i n terms of l i g h t r e d u c t i o n , n u t r i e n t enr ichment , and t o x i c chemical e f f e e t s . The s tudy i n d i c a t e d t h a t l o c a l r e d u c t i o n i n primary product ion due t o l i g h t a t t e n u a t i o n i n s i d e t h e plume was s i g n i f i c a n t ; however, because of r a p i d s e t t l i n g of a l a r g e percentage of t h e sediment p a r t i c u l a t e s , t h e shading e f f e c t on phytoplankton wae expected t o be only temporary, on t h e o r d e r of s e v e r a l hours. Since t h e a d a p t a t i o n of phytoplankton t o a new l i g h t regime t a k e s 1 t o 2 days, s h o r t - term shading i n t he d i s c h a r g e plume was n o t expected t o apprec i ab ly a f f e c t phy t op l a n k t on phot osyn thee i s .
Chan and Anderson (1981) a s s e s s e d t h e e f f e c t of mining on n u t r i e n t enrichment from information i n the l i t e r a t u r e . Based on t h e average d i scha rge r a t e and t h e r a t e of mixing between d i scha rge and ambient water i n t es t mining, t h e a u t h o r s c a l c u l a t e d t h a t n i t r a t e enrichment would be no more than 0.03 pg-atm/J?, w i t h i n t h e f i r s t few minutes and 0.003 yg-atrnlg w i t h i n t h e f i r s t hour, These i n c r e a s e s were expected t o be below a n a l y t i c a l d e t e c t a b i l i t y and n o t t o have any a p p r e c i a b l e immediate e f f e c t on t h e n i t r a t e - u p t a k e ra tes and primary product ion. n u t r i e n t s i n t roduced i n t o t h e eupho t i c zone were expected e v e n t u a l l y t o be inco rpora t ed i n t o primary p roduc t ion , but even a t f u l l - s c a l e mining rates t h e i n c r e a s e i n primary product ion due t o one or even a dozen mining v e s s e l 8 was not expected t o c r e a t e any s i g n i f i c a n t change i n t h e marine food c h a i n ,
I n t h e long tern t h e
Experiments conducted by Chan and Anderson (1981) t o s tudy t h e e f f e c t s of heavy me ta l s and o t h e r t o x i c subs t ances i n d i c a t e d t h a t t h e mining d i scha rge had no e f f e c t on t h e short- term pho tosyn the t i c ra te , even when t h e s e subs t ances were p r e s e n t a t a f a i r l y h igh concen t r a t ion . Due t o t h e i n s i g n i f i c a n t amount of heavy metal8 from bottom sediment and t h e r a p i d mixing and d i s p e r s a l of t h e mining d i s c h a r g e , any long-term t o x i c e f f e c t was considered n e g l i g i b l e .

25
Impact on Hicrozooplankton
.
The e f f e c t s of mining d i scha rge on microzooplankton have not been i n v e s t i g a t e d . Any e f f e c t on t h e s e organisms could be c a t a s t r o p h i c s i n c e they a r e e s s e n t i a l f o r f i r s t - f e e d i n g f i s h l a rvae . Although f i s h larvae are dependent on t h e s e food items f o r a b r i e f period on ly , they r e p r e s e n t an e s s e n t i a l f i r s t s t e p i n ensu r ing t h e s u r v i v a l of t h e l a r v a e .
The p o t e n t i a l e f f e c t s of mining on macrozooplankton have been examined by H i r o t a (1981) from l imi t ed f i e l d d a t a ( t h r e e r e p l i c a t e neuston n e t tows each i n ambient water and d i scha rge plume a t two t e s t mining s i tes) and i n l a b o r a t o r y experiments. The s t and ing s tocks of macrozooplankton, based on neuston n e t tows, were s i g n i f i c a n t l y h ighe r i n ambient water a t one mining s i t e and i n plume water a t t h e o t h e r . macrozooplankton t a x a i n ambient and plume l o c a l i t i e s were e q u a l l y h i g h a t both mining s i t e s . were appa ren t ly una f fec t ed by t h e mining plume, a t least ove r a short- term pe r iod b
The s i m i l a r i t y of occurrences of
Stock abundance and taxonomic composition a t t h e s u r f a c e
Laboratory experiments with 1 2 s p e c i e s of copepods exposed t o low-to- m e d i m p a r t i c u l a t e c o n c e n t r a t i o n s , i nc lud ing t h e range of c o n c e n t r a t i o n s observed du r ing test mining, have been made t o determine m o r t a l i t y rates ( H i r o t a 1981). was ( 5 % f o r most s p e c i e s a f t e r 24 h and only s l i g h t l y h ighe r ( w i t h i n 10%) a f t e r 48 h. The r e s u l t s i n d i c a t e d t h a t mining d i scha rge a t t h e c o n c e n t r a t i o n s used i n t h e experiments and observed i n tes t mining plumes d i d not cause unusual ly h igh m o r t a l i t y .
The d i f f e r e n c e i n m o r t a l i t y between t r ea tmen t s and c o n t r o l s
The s tudy by H i r o t a (1981) a l s o included t h e examination of f e c a l p e l l e t s from copepods and measurements of ra tes of f e c a l p e l l e t product ion, p e l l e t s i z e , and s i n k i n g speeds. On t h e b a s i s of t h i s s tudy as wel l as s t u d i e s by o t h e r workers, Hanson e t a l . (see f o o t n o t e 1 ) suggested t h a t zooplankton w i l l i n g e s t s i g n i f i c a n t q u a n t i t i e s of p a r t i c u l a t e matter i n j e c t e d i n t o s u r f a c e w a t e r s , which could l ead t o a s s i m i l a t i o n of trace metals. From r e s u l t s of f eed ing experiments i n t h e l i t e r a t u r e , t hey concluded t h a t a s s i m i l a t i o n of t r a c e metals from food through t h e gu t wal ls of zooplankton was no t p a r t i c u l a r l y e f f i c i e n t and t h a t normal p rocesses of e l i m i n a t i o n , such a s e x c r e t i o n (bo th s o l u b l e release and f e c a l p e l l e t p roduc t ion ) , mo l t ing , and egg l ay ing , appeared t o r a p i d l y reduce t h e levels of t r a c e metals t h a t were i n i t i a l l y inco rpora t ed i n t o t i s s u e .
Impact on F i s h
Adults
There have been r e l a t i v e l y few s t u d i e s on t h e e f f e c t s of s u s p n d e d p a r t i c u l a t e s and t u r b i d i t y on p e l a g i c marine f i s h e s . have focused on t h e phys io log ica l and p a t h o l o g i c a l e f f e c t s of high t u r b i d i t y on f r e shwa te r and e s t u a r i n e s p e c i e s . Many of t h e s e f i s h e s t h a t are normally exposed t o high t u r b i d i t i e s i n n a t u r e a re no t bothered by high test t u r b i d i t i e s (Wallen 1951; Herbert and Merkens 1961; A l a b a s t e r 1972, 1977; Sherk 1972; Neumann e t a l . 1975). Wallen (1951) and A l a b a s t e r (1972, 1977) r e p o r t e d t h a t p a r t i c u l a t e concen t r a t ions below 60 mg/R were n o t harmful t o t r o u t , B&Q g4if;dng~i and 8. &u,&&g; however, p h y s i o l o g i c a l e f f e c t s have been observed i n some f i s h e s a t much h ighe r t u r b i d i t i e s . Prolonged exposure t o e x c e s s i v e l y h igh t u r b i d i t i e s have produced clogged g i l l c a v i t i e e
Most of t h e q t u d i e s

26
r e s u l t i n g i n s u f f o c a t i o n (Everhar t and Duchrow 1970, c i t e d by Swenson and Matson 1976) and abnormal changes i n t h e g i l l s (Herber t and Merkens 1961; A labas t e r 1972). A labas t e r (1972) found th ickening and f u s i o n o f . t h e e p i t h e l i a l c e l l s of t h e secondary lamellae i n some 5. trgttg t aken from r i v e r s c a r r y i n g p a r t i c u l a t e concen t r a t ions of 1 ,000 and 5,000 mg/R. and Merkens (19611 observed t h a t s u r v i v a l of rainbow t r o u t , S. gghrdgggi, was adve r se ly a f f e c t e d i n l abora to ry tes ts a t concen t r a t ions of 270-810 mg/k
Herber t
The t u r b i d i t y caused by suspended p a r t i c u l a t e s a f f e c t f i s h i n o t h e r ways, most impor t an t ly i n t h e i r f eed ing behavior , by making it more d i f f i c u l t f o r t h e f i s h t o see t h e i r prey. observed tha t a t cons t an t l i g h t i n t e n s i t y t h e r e a c t i v e d i s t a n c e s of b l u e g i l l s , &gg@g mcgpcoiggg, t o h $ n _ ~ g diminished as t u r b i d i t y approached 30 Jackson T u r b i d i t y Uni t s (JTU) (ca. 44 mg/R). noted a s i g n i f i c a n t r e d u c t i o n i n t h e f eed ing rates of t h e s e f i s h a8 t u r b i d i t y was inc reased from 60 t o 120 and 190 Nephelometric T u r b i d i t y Uni t s ("TU) (ca. 88, 177 , and 280 ug/R, r e s p e c t i v e l y ) . Reduction i n f eed ing rate due t o t u r b i d i t y could u l t i m a t e l y l ead t o a r e d u c t i o n i n growth (Sykora e t a l . 1972) . Heimstra e t a l . (1969) found a gene ra l dec rease i n a c t i v i t y of j u v e n i l e largemouth bass , kf4cge~rgggg sglgg_idgg, exposed t o t u r b i d i t i e s of 14-16 JTU ( c a . 21-24 p g / t ) and S h i r a i s h i and Shimada (1972) r e p o r t e d t h a t a c t i v i t y of downstream migra t ing salmonid f i s h was reduced by inc reased t u r b i d i t y .
Vinyard and O'Brien (1976)
Gardner (1981)
Informat ion on pure ly marine s p e c i e s i s l i m i t e d . R i t c h i e (1970) d e t e c t e d no ill e f f e c t s of overboard dredge d i s p o s a l on 44 s p e c i e s of f i s h i n upper Chesapeake Bay. f l o r a and fauna , i nc lud ing a lbacore and salmonids, f l o u r i s h o f f s h o r e from t h e mouth of t h e Columbia River i n wa te r con ta in ing p a r t i c u l a t e concentra- t i o n s o f 10-100 mg/R. Moore and Moore (1976) showed t h a t t u r b i d i t y reduced t h e a b i l i t y of European f lounde r , P l a t i c h t h v s f l e s u s , t o see e p i b e n t h i c prey and inc reased t h e time r e q u i r e d t o cap tu re pursued prey. Connor (1976) found t h a t wh i t eba i t ( i m m a t u r e c lupeo ids ) avoided t h e edge of advancing "white water" mass p o l l u t e d wi th ch ina c l a y , whereas mackerel , Scomber scombrus, d i d no t . It seemed t h a t sma l l numbers of mackerel e n t e r e d t h e p o l l u t e d water and, a l though be ing v i s u a l f e e d e r s , were a b l e t o l o c a t e prey ( f i s h i n g l u r e ) .
P r u t e r and Alverson (1972) i n d i c a t e d t h a t abundant
Wilson and
The only s tudy of t u r b i d i t y and tuna was done by Barry (19781, who observed t h e r e a c t i o n of c a p t i v e ye l lowf in tuna and kawakawa t o a t u r b i d i t y c loud of suspended p a r t i c u l a t e s of deep-sea mud from t h e DOMES area. No ill ef fec ts were observed as a r e s u l t of s h o r t exposure t o t u r b i d i t i e s ranging from 6 to 40 NTU (ca. 9-59 m g / R ) . Behavioral responses were mixed. Although tunas were sometimes observed t o avoid t h e t u r b i d cloud, they r e a d i l y passed through i t , as t h e c loud c i r c l e d t h e tank . Maximum t u r b i d i t i e s a t t h e p o i n t where tunas passed through t h e cloud ranged from 1.6 t o 3.0 NTU (2.7-4.7 mg/R) f o r ye l lowf in tuna and 1.7 t o 4.43 NTU (2.28-6.8 rng/R) f o r kawakawa, and f eed ing occurred i n t u r b i d i t i e s t o 7.4 NTU (11.2 mg/R). experiments by Barry i n d i c a t e d a l s o t h a t i n DOMES mud t u r b i d i t i e s g r e a t e r t han 2.5 NTU (4 mg/R), t h e t e s t tunas sometimes became s u f f i c i e n t l y upse t t o s t o p f e e d i n g and e x h i b i t e d coughing behavior . Barry poin ted o u t t h a t it would be d i f f i c u l t t o ex tend t h e s e f i n d i n g s t o tunas i n t h e wi ld , since (1) t h e c a p t i v e tunas may have developed a h ighe r t o l e r a n c e t o t u r b i d i t y because of t h e i r exposure t o algae-produced t u r b i d i t y i n t h e tank , and (2 ) t h e a b i l i t y of t unas t o d e t e c t t u r b i d i t y c louds i n t h e wi ld might be d i f f e r e n t from t h a t of c a p t i v e tunas .
The p re l imina ry

27
Reduction i n t h e f eed ing r a t e of a d u l t tunas i n t h e mining plume need no t be a l a r g e concern, however, because of t h e low c o n c e n t r a t i o n of p a r t i c u l a t e s , the t r a n s i t o r y n a t u r e of t h e plume, t h e a b i l i t y of t u n a s t o f eed i n t u r b i d waters, and most impor t an t ly , t h e c a p a b i l i t y of t h e tunas t o s w i m through and ou t of plume waters . T h e only p o s s i b l e e f f e c t t o tunas could be t h e r e d u c t i o n of food organisms i n p lume waters as a r e s u l t of a r e d u c t i o n i n primary p r o d u c t i v i t y . product ion (Chan and Anderson 19811, however, have been shown t o be l i t t l e a f f e c t e d by t h e d i scha rge plume. of a d u l t b i l l f i s h e s t o t u r b i d wa te r s . Based on t h e s i m i l a r i t y of t h e i r f eed ing h a b i t s ( t y p e s of food e a t e n ) w i t h t h a t of a d u l t tunas and on t h e i r g r e a t m o b i l i t y , they t o o should be una f fec t ed by t h e d i s c h a r g e plume.
Food organisms ( H i r o t a 1981) and p r i m a r y
No s t u d i e s have been done on t h e behavior
Tuna and B i l l f i s h Larvae
There have been some s t u d i e s on t h e e f f e c t s of suspended s o l i d s and t u r b i d i t y on f i s h l a r v a e , but ha rd ly any on l a r v a e of tunas and b i l l f i s h e s . The e f f e c t s of plume water on l a r v a e of tunas and b i l l f i s h e s must, t h e r e f o r e , be i n f e r r e d from s t u d i e s made of o t h e r f i s h l a rvae . The normal development of embryos, l a r v a e , and p o s t l a r v a e of f i s h e s i n t h e mining area could be a f f e c t e d by sed imen ta t ion , l o s s of i l l u m i n a t i o n due t o t u r b i d i t y , and changes i n t h e p r o p e r t i e s of wa te r , such a s temperature .
------------- Sedimentation.--There have been only l i m i t e d s t u d i e s on t h e d i r e c t e f f e c t s of sediment on t h e e a r l y developmental s t a g e s of f i s h e s . Rosenthal (1971) observed t h a t t h e r e were major decreases i n egg hatching success and s u r v i v a l of h e r r i n g l a r v a e , Clg~e_a h_a_ggn_ggg, a t high c o n c e n t r a t i o n s of "red mud" (600-7,000 mg/R), and t h a t t h e development of t h e embryos were r e t a r d e d . i nc reased wi th i n c r e a s i n g c o n c e n t r a t i o n s , due t o t h e adherence of mud p a r t i c l e s t o t h e e g g s h e l l s and thereby p o s s i b l y i n t e r f e r i n g w i t h gaseous and o t h e r exchanges between t h e eggs and surrounding medium. H e f u r t h e r noted t h a t t h e i n g e s t i o n of r ed mud p a r t i c l e s by t h e larvae e v e n t u a l l y r e s u l t e d i n blocked food in t ake . Sherk e t a l . (1975) noted t h a t exposure of e s t u a r i n e f i s h e s t o s u b l e t h a l c o n c e n t r a t i o n s of suspended s o l i d s s i g n i f i c a n t l y inc reased hematocrit v a l u e , hemoglobin c o n c e n t r a t i o n , and e r y t h r o c i t e number, i n d i c a t i n g r e s p i r a t o r y stress, and t h a t t h e e a r l y l i f e s t a g e s of t h e s e f i s h e s were more s e n s i t i v e t o suspended s o l i d s than t h e a d u l t s . They r e p o r t e d a l s o t h a t 18-h exposure t o f u l l e r ' s e a r t h i n c o n c e n t r a t i o n s of 0.8 g/R r e s u l t e d i n 100% m o r t a l i t y of j u v e n i l e b l u e f i s h , Pomatomidae. Swenson and Matson (1976) r e p o r t e d t h a t growth and s u r v i v a l of l ake h e r r i n g , C Q ~ _ ~ ~ Q I N I ~ gr_&gd_i, l a r v a e held f o r up t o 62 d a y s i n experimental tanks were not a f f e c t e d by red c l a y c o n c e n t r a t i o n s ranging from 0 t o 4 8 Formazon T u r b i d i t y Units ca. 1-28 mg/R). No d i f f e r e n c e s were found i n m o r t a l i t y nor average weekly growth r a t e s over t h e observed range of red c l a y c o n c e n t r a t i o n s .
He r e p o r t e d a l s o t h a t t h e number of embryonic malformations
Di rec t e f f e c t s of sediment on e s t u a r i n e and f r e shwa te r f i s h e s , such as
While these f i s h e s those mentioned above, g e n e r a l l y occurred a f t e r prolonged exposure t o h igh sediment c o n c e n t r a t i o n s ranging from 270 t o 7,000 mg/t. have g r e a t e r t o l e r a n c e than ocean s p e c i e s t o suspended p a r t i c u l a t e s , t h e lower l e v e l s of c o n c e n t r a t i o n s and t h e r a p i d d i s s i p a t i o n of sediments a t t h e mining s i t e s should l e s s e n t h e impact on eggs and l a r v a e of t h e l a t t e r spec ie s . I n t h e t es t mining a r e a , t h e maximum c o n c e n t r a t i o n of p a r t i c u l a t e s 15 min a f t e r i n i t i a l mixing w a s ( ca . 900 ug/R) Ozturgut , L a v e l l e , and Burns 1981) and t h e c o n c e n t r a t i o n of p a r t i c u l a t e s was found t o d i s s i p a t e r a p i d l y

2 8
to- nea r ambient i n (5 h (Burns e t a l . 1980). sediment c o n c e n t r a t i o n and t h e r a p i d development of tuna embryos (ha t ch ing occur s i n about 1 day a f t e r f e r t i l i z a t i o n ) would allow t h e eggs minimal exposure t i m e t o s ed imen ta t ion and thus reduce o r nega te embryonic malformation. 2 t o 3 days a f t e r hatching. By t h i s t i m e , of course, t h e c o n c e n t r a t i o n of sediment p a r t i c u l a t e s should be a t o r near ambient and thus not pose any danger t o t h e larvae i n causing blocked food i n t a k e or i n a f f e c t i n g t h e i r growth. The o n l y l i k e l y e f f e c t of sediment would be t o those l a r v a e a l r e a d y i n t h e f eed ing s t a g e s . However, i n view of t h e r a p i d d i s s i p a t i o n of t h e d i s c h a r g e , t h e exposure t i m e would be confined t o a matter of hours and, t h e r e f o r e , should n o t n o t i c e a b l y a f f e c t t h e growth of t h e larvae. Under commercial mining c o n d i t i o n s , where d i s c h a r g e volumes per u n i t ocean a r e a a r e expected t o be s l i g h t l y less than t h a t of mining tes ts ( L a v e l l e and Ozturgut 19811, t h e chances of sediment p a r t i c u l a t e s d i r e c t l y a f f e c t i n g tuna eggs and l a r v a e should be n e g l i g i b l e .
The r a p i d d i s s i p a t i o n of
As f o r t h e newly hatched l a r v a e , f eed ing g e n e r a l l y begins i n
'&gh&!&y.--Feeding i s t h e prime a c t i v i t y of a l l f i s h l a r v a e f o r s u r v i v a l . s ea rch ing power and r e a c t i v e d i s t a n c e of t h e l a r v a e , t h e i r f eed ing e f f i c i e n c y , and t h e abundance of s u i t a b l e food organisms p r e s e n t ( B l a x t e r 19651, bu t a l s o on i l l umina t ion . B lax te r (19801, i n h i s review of v i s i o n and f eed ing of f i s h e s , r e p o r t e d t h a t t h e a b i l i t y of f i s h t o f e e d f e l l p r o g r e s s i v e l y du r ing dusk o r a s t h e i l l u m i n a t i o n w a s a r t i f i c i a l l y reduced. The t h r e s h o l d i l l u m i n a t i o n f o r f eed ing by more than 20 s p e c i e s of f i s h averaged about 0.1 lux. and t o f e e d s u c c e s s f u l l y f e l l p r o g r e s s i v e l y f o r larvae of some s p e c i e s ( h e r r i n g , p l a i c e , and n o r t h e r n p i k e , _Esgx hc_iu_g) dur ing dusk o r as i l l u m i n a t i o n was reduced. Kawamura and Hara (1980) noted t h a t m i l k f i s h , C_h_ag_og &nos, l a r v a e a l s o could n o t t a k e food i n t h e da rk , but t h a t t h e j u v e n i l e s were a b l e t o do so. For these l a r v a e , feeding thus would be l i m i t e d t o d a y l i g h t hours , and any r e d u c t i o n i n i l l u m i n a t i o n due t o t u r b i d i t y w i l l i n e f f e c t dec rease t h e hour6 a v a i l a b l e f o r f e e d i n g o r l i m i t f eed ing a c t i v i t y t o less than optimum l e v e l s .
The success o r f a i l u r e i n f eed ing w i l l depend n o t only upon t h e
He repor t ed a l s o t h a t t h e a b i l i t y t o p e r c e i v e prey
To compensate f o r reduced i l l u m i n a t i o n , t h e l a r v a e w i l l be r e q u i r e d t o depend even more g r e a t l y upon t h e i r swimming power t o ensu re s u f f i c i e n t prey c o n t a c t s and upon t h e i r r e a c t i v e d i s t a n c e and f eed ing e f f i c i e n c y . B i s h a i (1960) found t h a t h e r r i n g l a r v a e 6-8 mm long could s u s t a i n a water c u r r e n t of 0.57-1.03 cm/s f o r a t least 1 h, g i v i n g them the p o s s i b i l i t y of swimming 20-30 m i n 1 h. Presumably l a r g e r and s t r o n g e r l a r v a e w i l l have g r e a t e r swimming c a p a b i l i t i e s and enab le them t o cover g r e a t e r d i s t a n c e s w i t h i n d a y l i g h t hours. For l a r v a e sma l l e r than 6 mm, however, d i s t a n c e covered and r e a c t i v e d i s t a n c e , as w e l l as f eed ing e f f i c i e n c y , would be g r e a t l y reduced i n most s p e c i e s . I n most f i s h l a r v a e , t h e d i s t a n c e a t which they r e a c t t o prey i s q u i t e s h o r t . Herring l a r v a e were found t o r e a c t t o prey a t d i s t a n c e s of 0.7-10 body l eng ths when they f i r s t s t a r t e d f eed ing (Rosenthal and Hempel 1970). B lax te r and S t a i n e s (1971) r e p o r t e d r e a c t i v e d i s t a n c e s of 0.4 body l e n g t h f o r h e r r i n g , 0.5 body l e n g t h f o r p l a i c e , P&gggggcQg ~ l g p g s g , and s o l e , soleg w l g g , and 0.2 body l eng th f o r p i l c h a r d , _Sg:di.g ~ilcqggd~g. Hunter (1972) found t h e r e a c t i v e d i s t a n c e of n o r t h e r n anchovy, _Eggggg_lig umgugx, t o be 0.4 body l eng th . The l a r v a e of a l l s p e c i e s , except p l a i c e , were r e p o r t e d a s having low feed ing success (2-10%) a t i n i t i a l f eed ing . P l a i c e had high success r a t e s (32-62%) due t o t h e i r i n c r e a s e d maneuverab i l i t y and a b i l i t y t o s w i m backward. The s h o r t r e a c t i v e d i s t a n c e and low feed ing e f f i c i e n c y a t f i r s t f e e d i n g would

29
be lessened even more by reduced i l l umina t ion , which i n e f f e c t would reduce t h e a v a i l a b i l i t y of food. Thus, growth and s u r v i v a l would be a f f e c t e d , as i t has been observed i n feeding s t u d i e s of l a rvae of p l a i c e (R i l ey 1966; Wyatt 19721, sea bream, kcmsgggug ghkmhsiPglig (Houde 19751, bay anchovy, Ag$_hg_a gi&$billi (Saksena and Houde 1972; Houde 1977) , s a r d i n e , Hgggngulg ~egggc~llg (Saksena and Houde 19721, and l i ned s o l e , &_hjggs lin_egtge (Houde 1977). To compensate f o r reduced r e a c t i v e d i s t a n c e r e s u l t i n g from lower i l l u m i n a t i o n , it w i l l be necefisary f o r t h e l a r v a e t o i n c r e a s e prey con tac t s . Hunter and Thomas (1974) observed such an adjustment i n l a r v a e of anchovy, which increased t h e i r swimming speed and a r e a of s ea rch a t low prey d e n s i t i e s .
Although l i g h t i n t e n s i t y i s important f o r t h e s u r v i v a l of l a r v a e , l i g h t i n t e n s i t y a lone may not d i r e c t l y in f luence embryonic development. Ciechomski (1967a) observed t h a t eggs of anchovy, E. mi_ho.&g, kept i n complete darkness developed and hatched i n a normal way. I n o t h e r t ank experiments , Houde and Palko (1970) noted t h a t m o r t a l i t y of s a rd ine larvae was h ighe r i n a tank wi th poor l i g h t i n g (one 75-W incandescent bu lb) t han i n one wi th b e t t e r l i g h t i n g (two 40-W f l u o r e s c e n t bu lbs ) . Thacker (1963) a l s o r epor t ed h igh m o r t a l i t i e s f o r l a rvae of p l a i c e r e a r e d under poor l i g h t cond i t ions ( t o t a l l i g h t a t t h e su r face was below 30-f t candles ) .
Ri ley and
Undoubtedly, t h e feeding of tuna and b i l l f i s h larvae w i l l be a f f e c t e d by reduced i l l umina t ion . however, by t h e i r g r e a t e r swimming power, feeding e f f i c i e n c y , and b e t t e r v i s i o n because of t h e i r l a r g e r eyes and mouth, and ear l ie r development of f i n s than most o t h e r f i s h la rvae . The r a p i d growth e x h i b i t e d by these larvae should f u r t h e r a i d them t o s u r v i v e and pass througn t h e c r i t i c a l e a r l y l i f e s t ages .
They should be a b l e t o compensate f o r t h i s ,
Turb id i ty a l s o a f f e c t s l a rvae i n o t h e r ways. Swenson and Matson (1976) noted t h a t h e r r i n g l a r v a e had a tendency t o c o n c e n t r a t e c l o s e r t o t h e s u r f a c e when sub jec t ed t o high t u r b i d i t y . r e a c t i o n t o reduced l i g h t l e v e l s a t t h e bottom of t h e t u r b i d water chambers and suggested t h a t such a r e a c t i o n i n the wild could i n d i r e c t l y in f luence s u r v i v a l by exposing t h e l a r v a e to g r e a t e r preda t ion . Miller (1974) , i n h i s survey of l a r v a l f i s h i n wa te r s off sugar m i l l s , sewer o u t f a l l s , o i l r e f i n e r i e s , thermal o u t f a l l s , urbanized a reas wi th a s s o c i a t e d r u n o f f s , and i n harbors i n the Hawaiian I s l a n d s , noted t h a t s p e c i e s of f i s h larvae, inc luding tunas , i n t u r b i d water were lower i n d e n s i t y by 75% and i n number by 55%, and t h a t t u r b i d i t y , whether n a t u r a l o r a r t i f i c i a l , was nega t ive ly c o r r e l a t e d wi th l a r v a l f i s h abundance. This s tudy impl ies t h a t f i s h l a r v a e i n t h e wild avoided t u r b i d waters . I f t h i s i s so, i t does no t seem l i k e l y t h a t m o r t a l i t y by p reda t ion would be s i g n i f i c a n t l y increased i n plume wa te r s , a s suggested by Swenson and Matson (1976) .
They a t t r i b u t e d t h i s
E~vig~ggggtgl fgctpyg.--Sal ini ty , oxygen, and tempera ture a l s o a f f e c t t h e e a r l y development of f i s h e s , p a r t i c u l a r l y i n t h e embryonic and la rva l s t a g e s . occurred normally a t s a l i n i t i e s of 25.8-50'/00, but s a l i n i t i e s above o r below t h i s range produced pronounced abnorma l i t i e s i n t h e embryos. Fonds e t a l . (1974) found t h a t s u r v i v a l of g a r f i s h , _ B g l ~ g e b_&gg, eggs was inf luenced predominantly by tempera ture , but t h e s u r v i v a l of larvae from ha tch ing t o 30-40 mm TL was inf luenced by both s a l i n i t y and temperature. Su rv iva l of eggs and l a r v a e was found t o be maximized a t s a l i n i t i e s of
Ciechomski (1967a) noted t h a t t h e embryonic development of anchovy

30
31.4-37.1"/00 and temperatures of 17.4'-19.4"C, and t h e l i m i t s f o r s u r v i v a l of eggs were e s t ima ted a t s a l i n i t i e s of < l o t o > 4 5 ' / 0 0 , and temperatures of 12O-24"C. s a l i n i t y of 5 " / o 0 . oxygen on s t r i p e d b a s s , &_r_ogg g g x g & _ i l i g , eggs and l a r v a e , Turner and F a r l e y (1971) observed t h a t r e d u c t i o n s i n d i s so lved oxygen from s a t u r a t i o n (7.3-8.8 mg/R) t o 4 and 5 mg/R adve r se ly a f f e c t e d ha tch ing success and s u r v i v a l of t h e l a r v a e . Egg s u r v i v a l was found t o dec rease wi th an i n c r e a s e i n temperature (from 18.3' t o 22.2'C) o r exposure t i m e (from 6 t o 56 h ) , o r a dec rease i n oxygen l e v e l . A t an oxygen l e v e l of 4 mg/&, s u r v i v a l was c o n s i s t e n t l y reduced f o r a l l exposure times, t h e l e n g t h of time f o r ha t ch ing was longe r , and a number of l a r v a e hatched with v a r i o u s a b n o r m a l i t i e s .
I n o t h e r experiments , g a r f i s h eggs f a i l e d t o develop a t a I n l abora to ry experiments on t h e e f f e c t s of d i s so lved
Based on t h e s e s t u d i e s , i t seems t h a t d e t r i m e n t a l e f f e c t s occur a t e i t h e r extremely high o r low s a l i n i t y levels--normal development t a k e s p l a c e over a f a i r l y wide s a l i n i t y range--and a f t e r some hours of exposure t o reduced d i s s o l v e d oxygen levels . Although t h e l e v e l s a t which these e f f e c t s occur i n eggs and l a r v a e of b i l l f i s h e s and t u n a s could d i f f e r from t h e s p e c i e s noted above, t h e o v e r a l l e f f e c t s may be s imilar . area, however, ill e f f e c t s from s a l i n i t y and d i s s o l v e d oxygen i n t h e d i scha rge water should be minimal because of t h e absence of extreme c o n c e n t r a t i o n s , such a s those used i n experimental t e s t s , and because of t h e r a p i d mixing of t h e d i scha rge water. of t h e water a t dep ths g r e a t e r than 5,000 m was 34.72 ~ O . O l o / o ~ (Burns e t ill, 19801, which was w e l l w i th in t h e s easona l range of 33-35"/00 i n t h e s u r f a c e mixed layer (Ozturgut e t a l . 1978). From monitoring tests (Ozre t i ch 19811, t h e mean oxygen concen t r a t ion i n 1-, 2-, and 3-h-old plume was r e p o r t e d as 4.68850.035 m l / & , n e a r l y t h e same a s t h e ambient mean c o n c e n t r a t i o n of 4.69520.05 m l / R , and t h e maximum d i l u t i o n of t h e d i s c h a r g e water was r e p o r t e d t o occur wi th in t h e f i r s t few minutes of mixing w i t h s u r f a c e water.
I n t h e mining
During t e s t mining, t h e s a l i n i t y
Temperature appears t o have a predominant i n f l u e n c e i n the develop- ment of embryos and l a r v a e . Most f i s h eggs and l a r v a e were found t o s u r v i v e a wide range of temperatures: a l . 19811, 10"-17°C by anchovy (Ciechomski 1967a), ll"-2OoC by t i u l k a , Clupeonel la d e l i c a t u l a d e l i c a t u l a (Pinus 1974) and 23 "-30°C by ye l lowf in tuna (Harada e t a l . 19781, t o mention a few. A t temperatures below o r above t h e normal r anges , however, t h e development of embryos and l a r v a e could be a f f e c t e d d r a s t i c a l l y . Ciechomski (1967a) observed t h a t a temperature of 4°C water i n t h e early b l a s t u l a s t a g e showed a complete h a l t i n t h e i r development, d e a t h occurred i n 5 t o 6 days, and eggs which had been kep t f o r 1 t o 2 days a t 4°C and t r a n s f e r r e d t o normal temperature (14°C) d i e d more q u i c k l y , sugges t ing t h a t lower temperature may cause i r r e v e r s i b l e damages. A t low temperatures (1Oo-12"C1, l a r v a e of s t r i p e d b a s s were found t o hatch wi th a depressed c e p h a l i c r eg ion (Morgan e t a l . 1981) and t h e l a r v a e of ye l lowf in tuna were found t o be deformed when hatched i n temperatures below 20°C (Harada e t a l . 1978). Larvae of P a c i f i c cod became so i n a c t i v e a t very low temperatures t h a t t h e y were unable t o c a t c h and consume food (Alde rd ice and F o r r e s t e r 1971). higher than t h e normal range, hatched larvae of s t r i p e d b a s s showed pugheaded f e a t u r e s , small yolk s a c s , and deformed i n t e r n a l organs (Morgan e t a l . 19811, and larvae of anchovy e x h i b i t e d an i n c r e a s e i n h e a r t b e a t from 65-70 beats /min t o about 100 (Ciechomski 1967a).
14"-22"C by s t r i p e d b a s s (Morgan e t
A t temperatures

31
.
Fish larvae can be a f f e c t e d a l s o by thermal shock when sub jec t ed t o sudden changes i n temperature . Hoss e t a l . (1974) noted t h a t l a r v a e of A t l a n t i c menhaden, B r g y ~ ~ g t i g tygpgggg; s p o t , &gio_sto_g~g IK~Q~~Q~IJ~; p i n f i s h , Egg_o&q ghhmqipclg; and t h r e e s p e c i e s of f lounde r s , &rgdjcc&_hyg &g&g&ug, e . lg&hpgtigma, and 1) . db_6gu_t&_a, acc l ima t i zed a t 5 " , l o " , and 15°C and sub jec t ed t o 15"-18°C i n c r e a s e s i n temperature showed immediate r e a c t i o n s , such as l o s s of equ i l ib r ium, e r r a t i c swimming, v i o l e n t jumping, and convuls ions. This i n i t i a l per iod of r e a c t i o n was o f t e n followed by a r e t u r n t o r e l a t i v e l y normal behavior. These f i s h a l s o r ece ived a thermal shock when they were r e t u r n e d t o the o r i g i n a l temperature. This second shock was o f t e n more v i o l e n t than t h e f i r s t and was more pronounced a t t h e lower a c c l i m a t i z a t i o n temperatures . According t o t h e a u t h o r s , t h e second shock may be a d i r e c t cause of m o r t a l i t y , o r i f t h e f i s h are n o t k i l l e d , they may be more v u l n e r a b l e t o predat ion. G r i f f i t h (1978) noted t h a t t h r e a d f i n shad, Qo_g~gega pxz&gggggg, acc l ima t i zed a t 15°C and exposed t o sudden coo l ing (2"-5"C dec reases i n 4 h ) l o s t e q u i l i b r i u m i n p r o p o r t i o n t o t h e magnitude of temperature dec rease , as t h e temperature approached 4"- 6"C, and tha t as much as 32% of t h e cold-shocked f i s h t h a t had l o s t e q u i l i b r i u m f o r a s s h o r t a t i m e as 12 s b e f o r e being r e tu rned t o warmer wacer could n o t recover and experienced m o r t a l i t y .
Eggs and l a r v a e of tunas a l s o could be a f f e c t e d by temperature as noted above. Because of r a p i d d i s s i p a t i o n of t h e cold d i scha rge water soon a f t e r i n i t i a l mixing, such e f f e c t s would be l i m i t e d t o t h e area immediately beneath t h e o u t f a l l . With t h e ambient s u r f a c e temperature ranging from 21" t o 26°C i n t h e mining area and t h a t of t h e d i scha rge ranging from 4 " t o 1 O " C , t he d i f f e r e n c e i n the temperatures (17°C) would be s imilar i n magnitude t o t h a t t e s t e d by Hoss e t a l . (1974) f o r o t h e r f i s h s p e c i e s . With thermal shock l i k e l y t o produce an immediate r e a c t i o n , t h e r e i s r eason t o b e l i e v e t h a t eggs and larvae coming i n t o d i r e c t con tac t w i th t h e c o l d d i scha rge water would be a f f e c t e d adve r se ly .
Assuming t h a t a l l tuna eggs and l a r v a e coming i n t o d i r e c t c o n t a c t w i t h t h e cold d i s c h a r g e water were t o s u f f e r m o r t a l i t y , a rough estimate of t h e t o t a l number of larvae t h a t could be k i l l e d can be made and t h e l o s s e s evaluated i n terms of t h e magnitude of t h e annual c a t c h e s (see s e c t i o n on E f f e c t s on F i s h e r i e s ) .
Under f u l l - s c a l e mining o p e r a t i o n , bottom water w i l l be discharged a t t h e s u r f a c e through a 30-cm pipe. The discharged water w i l l f a l l t o t h e s u r f a c e from a he igh t of approximately 5 m and impact a swath of s u r f a c e water approximately 1-m wide, l - m deep, and 89.4-km long (mining s h i p speed of 1 m / s p e r day. Thus, t h e volume of water a f f e c t e d p e r day would be
w i t h i n t h e planned mining area (Matsumoto 1958; S t r a sburg 1960; and unpublished data, Honolulu Laboratory f i l e s ) , t h e c o n c e n t r a t i o n s of tuna larvae were approximately 0.89 s k i p j a c k and 0.30 yel lowfin tunas p e r 1,000 m3. c o n c e n t r a t i o n t i m e s 86,400/1,000), could be a f f e c t e d by t h e d i s c h a r g e water each day. These numbers, however, r e p r e s e n t on ly l a r v a e p a s t t h e i n i t i a l f eed ing s t a g e and do n o t i nc lude eggs and yo lk s a c larvae which, a l though p r e s e n t , were n o t analyzed i n t h e larval s t u d i e s . Size-frequency d a t a from S t r a s b u r g (1960) i n d i c a t e t h a t about 33% of t h e s k i p j a c k tuna larvae and 29% of t h e y e l l o w f i n tuna l a r v a e taken i n plankton n e t tows were a t t h e i n i t i a l f eed ing s t a g e (3.0-3.9 mm TL). Assuming t h a t eggs and 1- and 2- day-old y o l k sac larvae were p r e s e n t a t t h e t i m e n e t tows were made and
86,400 m 4 . Based on surface day and n i g h t plankton n e t c a t c h d a t a from
Hence, 77 s k i p j a c k tuna and 26 ye l lowf in tuna l a r v a e ( l a r v a l

32
each s t a g e surv ived t o t h e f i r s t feed ing s t a g e i n 1 t o 3 days i n l i k e numbers, then an a d d i t i o n a l 76 s k i p j a c k tuna (77 x 0.33 X 3) and 23 ye l lowf in tuna (26 X 0.29 X 3) l a rvae would be a f f e c t e d , b r inging . t h e t o t a l eggs and larvae a f f e c t e d per day t o 153 s k i p j a c k tuna and 49 ye l lowf in tuna. annual product ion of 1.5 m i l l i o n tons of ore (Ozturgut , Lave l l e and Erickson 19811, ca . 45,900 s k i p j a c k tuna l a r v a e and 14,700 ye l lowf in tuna l a r v a e could be s u s c e p t i b l e t o thermal shock each yea r .
I n 300 mining days , t h e e f f o r t r equ i r ed t o achieve t h e es t imated
These estimates rep resen t t h e maximum number of l a rvae t h a t could be a f f e c t e d i f mining were done e n t i r e l y i n a r e a s where spawning by both spec ie s occur . q u a r t e r ( n o r t h of l a t . 10"N and e a s t of long. f40"W) of t h e p ro jec t ed mining area. a f f e c t e d by mining w i l l be reduced i n p ropor t ion t o the time (days) mining i s done i n t h i s s e c t o r .
Skip jack tuna , however, do no t spawn i n t h e n o r t h e a s t e r n
Consequently, t h e number of s k i p j a c k tuna l a r v a e l i k e l y t o be
Other Poss ib l e E f f e c t s
Tunas, as w e l l as o t h e r p e l a g i c f i s h e s , have been known t o congrega te under d r i f t i n g o b j e c t s , such a s logs , seaweed, r a f t s , and o t h e r f lo t sam. Fishermen have u t i l i z e d t h i s behavior of f i s h e s by seeking and f i s h i n g around such o b j e c t s , o f t e n wi th huge success (Uda 1933; Kimura 1954; McNeeley 1961; Inoue e t a l . 1963, 1968). I n r ecen t years manmade f i s h aggrega t ing dev ices (FAD's) have been devised and anchored i n c o a s t a l and deep open ocean waters i n t h e P h i l i p p i n e s (Murdy 1980; Apr ie to 1981, de Je sus 1982, Matsumoto2), Hawaii (Matsumoto e t a l . 19811, many c e n t r a l and South P a c i f i c I s l a n d s , New Zealand, A u s t r a l i a , and i n t h e Indian Ocean (Shomura and Matsumoto 1982), and i n t h e e a s t e r n P a c i f i c (Gu i l l en and Brat ten3) (Fig. 16 ) . i n s u f f i c i e n t l y l a r g e concen t r a t ions t o e n t i c e commercial tuna purse s e i n e r s and pole-and-line v e s s e l s t o f i s h around them. I n H a w a i i , pole- and-line f i s h i n g v e s s e l s caught over 471 MT (1 m i l l i o n l b ) of s k i p j a c k tuna and 45.4 MT (100,000 l b ) of ye l lowf in tuna o f f fou r FAD's i n 1978 (Table 1 ) . t unas , i nc lud ing s k i p j a c k , ye l lowf in , bigeye, kawakawa, f r i g a t e , and b u l l e t , have been taken i n a s i n g l e purse s e i n e set (see f o o t n o t e 2) .
These FAD's have a t t r a c t e d f i s h , predominantly tunas ,
I n t h e P h i l i p p i n e s e s t ima tes of up t o 200 MT of v a r i o u s s p e c i e s of
Tunas of v a r i e d s i z e s ranging from (1 t o >45 kg have been r epor t ed from beneath and around FAD's (Matsumoto e t a l . 1981). Small tunas ((1 o r 2 kg) g e n e r a l l y r m a i n e d c l o s e t o t h e FAD's du r ing t h e day and n i g h t , but l a r g e r t unas ( > 3 k g ) , p a r t i c u l a r l y t h e s k i p j a c k tuna , roamed over d i s t a n c e s of 5 km o r more away from t h e FAD's du r ing d a y l i g h t . The l a t t e r depar ted from t h e FAD's a t s u n r i s e and r e tu rned t o i t a t sunse t . The a b i l i t y of t h e s k i p j a c k tuna t o depa r t and r e t u r n t o a r e fe rence p o i n t has been demonstrated by Yuen (1970). By t r a c k i n g bank-associated small (40-44 cm)
2Matsumoto, W. M. Manuscr. i n prep. Payao f i s h i n g i n t h e P h i l i p p i n e s . Southwest F i s h e r i e s Center , Nat ional Marine F i s h e r i e s Se rv ice , NOAA, P. 0. Box 3830, Honolulu, H I 96812.
3Guil len, R . , and D. A. Bra t ten . 1981. Anchored r a f t experiment t o aggrega te tunas i n t h e eastern P a c i f i c Ocean. Inter-Am. Trop. Tuna Comm., I n t e r n a l Rep. 14, 1 0 p. Inter-American Tropica l Tuna Commission, c /o Sc r ipps I n s t i t u t i o n of Oceanography, P. 0. Box 1529, La J o l l a , CA 92093.
.

3 3
Figure 16.--Locations where f i s h aggrega t ing dev ices have been deployed, 1979-81, o r where deployment i s planned i n t h e P a c i f i c and Ind ian Oceans (Shomura and Matsumoto 1982).
Table I.--Fiah s p e c i e s caught ( i n pounrm/ uy pole-and-line Eiahing i n 1978 during the Hawaiian f i e h aggregating dev ice experiment (Matnumot6 et a t . 1981).
L
Skipjack tuna Yel lowfin tuna Rawakawa Dolphin Tot81
Catch per Catch per Catch per Catch per Catch per Buoy V i a i t s Catch v i s i t Catch v i s i t Catch v i a i t Catch v i s i t . Catch v i s i t
A 92 357,044 3,880.3 22,682 246.5 1,479 16.0 854 9.3 382,031 4,152.5
B 1 5,110 5,110.0 0 0 .o 0 0 .o 0 0 .o 5,110 5,110.0
C 14 103,037 7,359.8 1,475 105.4 4,218 301.3 0 0.0 108,730 7,766.4
D 139 573,106 4,k23.1 80,183 576.9 1,706 12.3 3,034 22.6 658,029 4,734.0
Tota l 246 1,038,297 4,220.7 104,340 424.1 7,403 30.0 3,888 15.8 1,153,900 4,690.6
Percent of t o t 8 1 Catch 89.73 9.28 0.64 0.34 99.99
s k i p j a c k tuna tagged wi th an u l t r a s o n i c d e v i c e , Yuen showed t h a t t h e f i s h was capab le of making n i g h t l y journeys of 25-106 km (maximum s t r a i g h t l i n e d i s t a n c e s of up t o 35 km from t h e bank). The tagged f i s h was observed t o l eave t h e bank a t n i g h t f a l l , r e t u r n i n g t o i t a t s u n r i s e , and remaining over t h e bank throughout t h e day as a member of a school . This p a t t e r n of d a i l y movement w a s observed f o r 3 days and a g a i n f o r 3 more days a f t e r an i n t e r r u p t i o n of 2 days, when t h e v e s s e l was r e q u i r e d t o r e t u r n t o p o r t . The r epea ted d a i l y p a t t e r n from and r e t u r n t o t h e bank l e d Yuen t o conclude t h a t t h e s k i p j a c k tuna was a b l e t o n a v i g a t e and was aware of t i m e .
The a b i l i t y of t h e s k i p j a c k tuna t o d e p a r t and r e t u r n t o a FAD can s i g n i f i c a n t l y a l t e r t h e normal d i s t r i b u t i o n a l p a t t e r n of schools w i t h i n an area. A f t e r having made i n i t i a l con tac t w i th a FAD, a school may prolong i t s 8 t a y . h t h e area. schoo l s around t h e FAD, as observed by Matsumoto e t a l . (1981).
I n due t i m e t h i s would r e s u l t i n a bui ldup of During t h e

3 4
peak f i s h i n g season , t h e FAD's a t t r a c t e d more tuna schools than normally would have been found i n t h e a r e a . The schools a t t r a c t e d t o t h e FAD's remained i n t h e v i c i n i t y or were rep laced by o t h e r s choo l s on a cont inuing b a s i s , a l lowing t h e pole-and-line f i s h i n g v e s s e l s t o f i s h around t h e FAD's f o r pe r iods of up t o 18 consecut ive days.
I n t h e Hawaiian FAD's s tudy , Matsumoto e t a l . (1981) r epor t ed t h a t two o r more schoo l s were p resen t around a FAD a t t h e same t i m e du r ing A p r i l and May. The long-term (1950-61) s i g h t i n g s of s k i p j a c k tuna schoo l s i n Hawaiian waters f o r a comparable per iod (March-May) is g iven as 0.87 schoo l s p e r 10 h of s cou t ing (Waldron 1964). Based on these s i g h t i n g s , FAD's could i n c r e a s e t h e number of schools w i t h i n i t s v i c i n i t y by a f a c t o r of t h r e e o r fou r . I n nodule mining, t h e s h i p , moving s lowly a t a speed of 1 m / s , could a c t a s a FAD and accumulate tuna schools . Should spawning occur a t t h i s t i m e , t h e concen t r a t ion of spawned eggs around and in t h e path of t h e mining s h i p would be h ighe r than u s u a l , and t h e number of eggs t h a t would e v e n t u a l l y be exposed t o d i r e c t con tac t wi th t h e co ld d i scha rge water a l s o would be increased . A t h r e e f o l d o r f o u r f o l d i n c r e a s e , as noted above, may n o t be unreasonable .
E f f e c t s on F i s h e r i e s
M o r t a l i t y of tuna eggs and larvae caused by mining i s bound t o have some e f f e c t on t h e ca t ches i n t h e e a s t e r n P a c i f i c su r face f i s h e r y and t h e Hawaiian s k i p j a c k tuna f i s h e r y . The e a s t e r n P a c i f i c f i s h e r y i s a major f i s h e r y : The c a t c h of s k i p j a c k tuna reached a high of 170,000 MT i n 1978 and averaged 128,300 MT annual ly i n t h e 5-year per iod p r i o r t o 1980 (Forsbergh 1980) . i n 1976 and averaged 201,000 MT annual ly over t h e same pe r iod (Cole 1980). The Hawaiian s k i p j a c k tuna f i s h e r y i s a minor f i s h e r y , wi th annual ca t ches f o r t h e same pe r iod averaging 3,100 MT (unpubl ished d a t a , Honolulu Laboratory f i l e s 1.
The c a t c h of ye l lowf in tuna reached a h igh of 236,400 MT
A crude estimate of t h e probable impact mining w i l l have on t h e r ec ru i tmen t of 1-year-old f i s h can be made from a v a i l a b l e length-frequency and c a t c h d a t a . The e a s t e r n P a c i f i c f i s h e r y can be sepa ra t ed a t l a t . 15"- 16"N o f f t h e coas t of Mexico i n t o a no r the rn and southern f i s h e r y . Nearly a l l of t h e f i s h e n t e r i n g t h e no r the rn f i s h e r y o r i g i n a t e i n e q u a t o r i a l waters from an area west of long. 13OoW, w i t h i n and n o r t h of t h e NECC (Rothschi ld 1965; Fink and B a y l i f f 1970; Williams 1972). I n t h e per iod 1971-74, t h e mean annual c a t c h i n t h e no r the rn f i s h e r y was 9,747 MT of s k i p j a c k tuna and 24,833 MT of ye l lowf in tuna (Calk ins 1975). Based on e a r l i e r publ i shed d a t a (Davidoff 1963; Broadhead and Barrett 19641, 1-year- o ld (below 50 cm) s k i p j a c k tuna averaged 24.0% by weight of t h e c a t c h i n t h e no r the rn f i s h e r y and 1-year-old (below 60 cm) ye l lowf in tuna averaged 0.9% of t h e ye l lowf in tuna ca t ch . Assuming t h a t t h e p ropor t ion of 1-year- o l d f i s h remained t h e same i n t h e 1971-74 per iod , t h e average annual c a t c h of t h i s age group would r e p r e s e n t approximately 2,336 MT f o r s k i p j a c k tuna and 2,235 MT f o r ye l lowf in tuna.
From ea r l i e r estimates, roughly 49,500 s k i p j a c k tuna and 14,700 ye l lowf in tuna larvae were expected t o be a f f e c t e d by mining each year . a nonmining s i t u a t i o n , t h e s e l a r v a e would be expected t o s u f f e r a n a t u r a l m o r t a l i t y of 80% (Murphy and Sakagawa 1977) du r ing t h e year. n a t u r a l m o r t a l i t y , 9,900 s k i p j a c k tuna and 2,940 ye l lowf in tuna larvae should s u r v i v e and e n t e r t h e f i s h e r y as 1-year-old f i s h .
I n
Adjus t ing f o r
These numbers

34
r e p r e s e n t approximately 15.3 MT of s k i p j a c k tuna and 10.7 MT of ye l lowf in tuna , based on an average weight of 1.85 kg f o r 40-50 cm s k i p j a c k tuna and 3.17 kg f o r 50-60 cm ye l lowf in tuna (length-weight r e l a t i o n s h i p t a b l e s , Honolulu Laboratory f i l e s ) . Adjust ing f o r p o s s i b l e FAD-like e f f e c t s of t h e mining s h i p would i n c r e a s e t h e t o t a l s t o 65.2 and 42.8 W. f o r s k i p j a c k tuna and ye l lowf in tuna , r e s p e c t i v e l y . These t o t a l s r e p r e s e n t only an i n s i g n i f i c a n t amount (0.64 and 0.16%, r e s p e c t i v e l y ) of t h e annual ca t ches of t h e two s p e c i e s i n t h e e a s t e r n P a c i f i c no r the rn f i s h e r y . The a c t u a l impact on t h e annual ca t ches would be f a r less, i f t h e e f f i c i e n c y of t h e g e a r , which i s w e l l below loo%, were considered.
I n t h e Hawaiian f i s h e r y , t h e s k i p j a c k tuna ca t ch c o n s i s t s of f i s h mig ra t ing from t h e Japanese o f f s h o r e and t h e e a s t e r n P a c i f i c f i s h e r i e s , as i n d i c a t e d from r e c a p t u r e s of tagged f i s h (Matsumoto e t a l . 1984) and ve ry l i k e l y from t h e c e n t r a l e q u a t o r i a l wa te r s t o t h e south of t h e i s l a n d s (Matsumoto 1975). a r e a t o t h e south. The e f f e c t of mining on eggs and larvae could a f f e c t t h e even tua l r ec ru i tmen t of 1-year-olds, but u n l i k e t h e s e c t o r east of long. 130°W, where tuna spawning does no t occur n o r t h of l a t . 10"-15"N, heavy spawning by s k i p j a c k and yel lowfin tunas occur i n Hawaiian waters n o r t h of t h e mining a r e a . The l a r v a e spawned could remain i n t h e area and e n t e r t h e f i s h e r y as 1-year-old f i s h a year l a t e r o r mig ra t e sou th i n t o t h e mining a r e a as j u v e n i l e s du r ing w i n t e r , a t s i z e s l a r g e enough t o be una f fec t ed by mining d i scha rge waters, and r e t u r n as 1-year-old r e c r u i t s . This should lessen t h e e f f e c t s of mining i n s o f a r a s t h e r ec ru i tmen t of 1- year-olds i n t o t h e IIawaiian f i s h e r y i s concerned.
Recruitment of 1-year-olds may be most ly from t h e mining
SUMMARY AND CONCLUSIONS
O f t h e many changes t o t h e environment r e s u l t i n g from nodule mining, i nc reased sediment and t u r b i d i t y i n s u r f a c e water and t h e d i scha rge of co ld bottom water a t t h e s u r f a c e were considered most l i k e l y t o a f f e c t tuna and b i l l f i s h eggs and l a r v a e . The p o t e n t i a l e f f e c t s on t h e b i o t a were examined i n terms of small-scale t e s t mining r e s u l t s and p r o j e c t i o n s t o f u l l - s c a l e mining e f f e c t s were attempted whenever such information was a v a i l a b l e .
I n t e s t mining s t u d i e s , t h e maximum concen t r a t ion of sediment p a r t i c u l a t e s soon a f t e r mixing of t he d i scha rge and s u r f a c e wa te r s was g iven a s 900 1lg/2 (Ozturgut e t a l . 1980). from mining extended downstream wi th t h e c o n c e n t r a t i o n of p a r t i c u l a t e s d i s s i p a t i n g r a p i d l y t o nea r ambient ( 5 5 p g / R ) i n 5 h a t a d i s t a n c e of approximately 5 km from t h e s h i p and t o ambient (34 1ig/2) i n about 37 h. Light a t t e n u a t i o n i n 1-h-old plume a t dep ths of 10-20 m was 0.13/m, more than t h r e e times t h e average ambient va lue . (mean - 7"C), was found t o mix r a p i d l y wi th s u r f a c e water and was unde tec t ab le i n chlor ini ty- temperature-depth c a s t s a f t e r 1 h.
The s u r f a c e plume t h a t r e s u l t e d
Cold d i s c h a r g e water, 4"-10"C
Under commercial mining c o n d i t i o n s , w i t h t h e s h i p t r a v e r s i n g t h e mining a r e a i n a r e c t a n g u l a r p a t t e r n 30 km on t h e long s i d e and 3.6 km on t h e s h o r t s i d e , p a r t i c u l a t e concen t r a t ion of 100 ygl9, was expected a t d i s t a n c e s of (6 km downstream of t h e mining s h i p and c o n c e n t r a t i o n of 50 vg/R ( n e a r ambient) a t d i s t a n c e s of about 1 4 km. W B R expected t o be reached i n about 1 5 h. was expected t o be t h r e e t imes t h a t of t h e t e s t miner , but because t h e s h i p ' s speed would be fou r times g r e a t e r , t h e d i scha rge volume p e r u n i t of ocean was expected t o be s l i g h t l y less than t h a t of t e s t mining ( L a v e l l e
The l a t t e r c o n c e n t r a t i o n The volume of discharged water

and Ozturgut 1981) . Consequently, t h e d i s s i p a t i o n of cold water was expected t o be no t much d i f f e r e n t from t h a t of t h e t e s t mining.
Pas t s t u d i e s on t h e e f f e c t s of suspended s o l i d s and t u r b i d i t y on f i s h eggs and l a r v a e have d e a l t mainly wi th f reshwater and e s t u a r i n e s p e c i e s . D i rec t e f f e c t s of suspended s o l i d s on t h e e a r l y developmental s t a g e s of f i s h e s inc luded such th ings a s r e t a r d e d embryonic development, development of malformed larvae, blocked food in t ake leading t o undernourishment and reduced growth, and clogged g i l l s r e s u l t i n g i n r e s p i r a t o r y stress and m o r t a l i t y . These e f f e c t s g e n e r a l l y occurred a t r e l a t i v e l y h igh t u r b i d i t i e s of 270 t o 7,000 mg/R and a f t e r prolonged exposures of 8 o r 9 weeks (Rosenthal 1971; Sherk e t a l . 1975; Swenson and Matson 1976). The combined e f f e c t s of low concen t r a t ion and r ap id d i s s i p a t i o n of p a r t i c u l a t e s i n t es t mining and t h e extremely rapid development of t h e eggs and larvae of tunas (embryonic development and ha tch ing of ye l lowf in tuna i n about 1 day i n t r o p i c a l tempera tures , yo lk abso rp t ion i n 2-3 days , and growth of larvae from 2.6 mm a t ha tch ing t o 51 mm i n 38 days) were expected t o reduce t h e r i s k of prolonged exposure and thus prevent any i l l e f f e c t s .
Reduced i l l umina t ion caused by t u r b i d i t y has been r epor t ed t o a f f e c t
These e f f e c t s have been overcome by some s p e c i e s (e .g . , h e r r i n g ) feeding by reducing the r e a c t i v e d i s t a n c e and feeding e f f i c i e n c y of f i s h l a rvae . which inc rease t h e i r swimming speed and area of s ea rch f o r prey (Hunter and Thomas 1974). Because of t h e e x c e p t i o n a l l y l a r g e eyes and mouth and t h e r a p i d development of t h e f i n s i n l a r v a e of tunas and b i l l f i s h e s , it was expected t h a t t h e s e l a rvae could compensate f o r reduced i l l u m i n a t i o n b e t t e r than most f i s h l a rvae . Thus, it was concluded t h a t increased t u r b i d i t y r e s u l t i n g from mining would n o t a f f e c t t h e s u r v i v a l of t hese l a rvae .
Other p r o p e r t i e s of seawater , such as s a l i n i t y , oxygen, and tempera ture , a l s o were found t o a f f e c t t h e development and s u r v i v a l of f i s h eggs and larvae, but only a t extremely low or high l e v e l s and a f t e r prolonged exposures . 111 e f f e c t s on tuna eggs and l a r v a e i n t h e mining s i t e should be minimal, however, because of i n s i g n i f i c a n t d i f f e r e n c e s i n t h e s a l i n i t y of bottom and su r face water and t h e r a p i d d i l u t i o n of t h e low oxygen bottom water a f t e r mixing w i t h s u r f a c e water .
The most l i k e l y , and perhaps t h e on ly , e f f e c t of tempera ture on t h e eggs and larvae of tunas w i l l occur a t t h e d i scha rge p o i n t of bottom water. I n nodule mining, t h e co ld d i scha rge water (4°-100C, average 7°C) f a l l i n g on to t h e sea s u r f a c e can come i n t o d i r e c t contac t wi th t h e developing eggs and newly hatched l a r v a e d r i f t i n g a t t h e s u r f a c e . Such c o n t a c t s could cause t h e c e s s a t i o n o f - embryonic development, t h e development of deformed larvae, o r r e s u l t i n thermal shock t o t h e l a r v a e , caus ing them t o lo se equ i l ib r ium and become easy t a r g e t s t o p reda to r s o r even r e s u l t i n dea th .
Supe r sa tu ra t ion of a tmospheric gases could be a problem, but i n t h e d i scha rge of an a i r l i f t mining ope ra t ion , s u p e r s a t u r a t i o n las t s only minutes due t o r a p i d d i l u t i o n . Exposure of f i s h t o supe r sa tu ra t ed gases , t hus , i s no t expected t o be a concern (Ozre t i ch 1981) .
Based on t h e premining c o n c e n t r a t i o n of tuna larvae i n t h e s u r f a c e l a y e r ( 1 m deep) wi th in t h e mining a r e a and on t h e e s t ima ted volume of s u r f a c e water t h a t would be a f f e c t e d by t h e d i scha rge water i n 300 days of mining each y e a r , it was e s t ima ted t h a t approximately 45,900 s k i p j a c k tuna and 14,700 ye l lowf in tuna larvae could be l o s t annual ly due t o mining.

37
These f i g u r e s would be about f o u r times l a r g e r i f t h e mining s h i p were t o act a s a FAD by concen t r a t ing tuna schools i n t h e immediate v i c i n i t y of t h e mining s h i p . Spawning a c t i v i t i e s by these schoo l s could i n c r e a s e . t h e concen t r a t ion of eggs n e a r t h e mining s h i p . t h r e e f o l d o r f o u r f o l d g r e a t e r than t h e concen t r a t ion i n nonmining s i t u a t i o n s .
Such inc reases can e a s i l y be
The impact of t h e m o r t a l i t y es t imated above on t h e e a s t e r n P a c i f i c nor thern f i s h e r y ( n o r t h of l a t . 15"N) and on t h e Hawaiian sk ip j ack tuna f i s h e r y can be es t imated . I n t h e e a s t e r n P a c i f i c , p r a c t i c a l l y a l l of t h e f i s h e n t e r i n g t h e no r the rn f i s h e r y o r i g i n a t e i n e q u a t o r i a l waters west of long. 13O"W. Larvae from t h i s r eg ion a re t r anspor t ed eastward i n t h e NECC and e n t e r t h e f i s h e r y as 1-year-old f i s h . F i s h of t h i s age group comprise 24% by weight (2,336 MT) of t h e s k i p j a c k tuna and 0.9% (2,235 MT) of t h e ye l lowf in tuna taken annual ly i n t h e no r the rn f i s h e r y (average annual ca t ches es t imated f o r t h e per iod 1971-74). O f t h e es t imated number of larvae t h a t would l i k e l y be k i l l e d by mining (45,700 s k i p j a c k tuna and 14,700 ye l lowf in t u n a ) , 80% are expected t o d i e of n a t u r a l causes , l eav ing 9,900 s k i p j a c k tuna and 2,940 yel lowf in tuna ( equ iva len t t o 15.3 and 10.7 MT, r e s p e c t i v e l y ) t o su rv ive and enter t h e f i s h e r y as 1-year-old f i s h . Adjus t ing f o r p o s s i b l e FAD-like e f f e c t s of t h e mining s h i p would inc rease t h e t o t a l s t o 65.2 and 42.8 MT f o r s k i p j a c k and ye l lowf in tunas , r e s p e c t i v e l y . These t o t a l s , r e p r e s e n t i n g approximat.ely 0.64 and 0.16% of t h e annual ca t ches of sk ip j ack and ye l lowf in t u n a s , r e s p e c t i v e l y , should have no n o t i c e a b l e e f f e c t on t h e t o t a l annual ca tches of both s p e c i e s taken i n t h e e a s t e r n P a c i f i c no r the rn f i s h e r y .
I n t h e Hawaiian f i s h e r y , t h e impact of mining on t h e s k i p j a c k tuna f i s h e r y would be even l e s s , s i n c e t h e spec ie s a l s o spawns i n t h e f i s h i n g grounds, o u t s i d e of t h e p ro jec t ed mining a r e a ( i . e . , n0rt.h of l a t . 20"N). Larvae from these spawnings could remain i n t h e area and be sub jec t ed t o f i s h i n g a year l a t e r ( a s 1-year-olds) o r mig ra t e south i n t o t h e mining area i n win te r as j u v e n i l e s , a t s i z e s l a r g e enough t o wi ths tand t h e impact of mining, and r e t u r n as 1-year-old f i s h i n t h e summer, t o g e t h e r w i th f i s h spawned w i t h i n t h e mining a rea .
Based on t h i s s tudy , t h e probable e f f e c t s of mining on tuna and b i l l f i s h eggs and l a r v a e appear t o be n e g l i g i b l e ; however, t h e premises on which c e r t a i n deduct ions have been made, r e q u i r e v e r i f i c a t i o n . Laboratory s t u d i e s t o a c c u r a t e l y d e s c r i b e t h e e f f e c t s of sediment and tempera ture on tuna eggs and larvae and on t h e spawning behavior of a d u l t tunas need t o be made. I n t h e f i e l d , more d e t a i l e d obse rva t ions on t h e tempera ture of t h e water a t t h e po in t of d i scha rge a re needed and t h e e f f e c t of t h e d i scha rge water on tuna eggs and l a r v a e a t t h e p o i n t of mixing need t o be determined. The abundance of tuna and b i l l f i s h eggs and l a r v a e along t h e pa th of t h e mining s h i p needs t o be determined r o u t i n e l y , so t h a t t h e ex ten t of t h e damage t o these l i f e s t a g e s can be monitored. I f damage t o t h e eggs and larvae by t h e d i scha rge i s found t o be excess ive , t hen a l t e r a t i o n of t h e d i scha rge p o i n t , perhaps t o subsur face l e v e l s , may be necessary .
Aside from these s t u d i e s concerning d i r e c t e f f e c t s on eggs and larvae, a d d i t i o n a l s t u d i e s a re needed on t h e pe l ag ic food web. It i s recommended t h a t t h e e f f e c t s of sediment and tempera ture on a l l components i n t h e food web be s t u d i e d , p a r t i c u l a r l y those organisms which a r e consumed dur ing i n i t i a l feed ing by t h e larvae.

38
ACKNOWLEDGMENTS
The author wi shes t o thank William F . Hettler and Jean Snider for t h e i r h e l p f u l s u g g e s t i o n s and c r i t i c a l reviews. NOAA's Deep Ocean Mining Environmental Study conducted by t h e Nat iona l Ocean S e r v i c e , D i v i s i o n of Ocean Minerala and Energy.
Thi s work i e p a r t of
.

39
LITERATURE CITED
Ahlstrom, E. He 1971.
P a c i f i c , based on c o l l e c t i o n s made on EASTROPAC I. U.S. 69:3-77.
Kinds and abundance of f i s h larvae i n t h e e a s t e r n t r o p i c a l Fish. Bull . ,
1972. Kinds and abundance of f i s h larvae i n t h e e a s t e r n t r o p i c a l P a c i f i c on t h e second m u l t i v e s s e l EASTROPAC survey, and o b s e r v a t i o n s on t h e annual c y c l e of l a r v a l abundance. F i sh . Bul l . , U. S. 70 : 11 53-1242.
A l a b a s t e r , J. S. 1972. Suspended s o l i d s and f i s h e r i e s . Proc. R. SOC. Lond., B Biol .
S c i . 180:395-406.
1977. B i o l o g i c a l monitor ing of i n l and f i s h e r i e s . In Proceedings of a Symposium of European In l and F i s h e r i e s Advisory Commission, 226 p. Appl. Sc i . Publ., Lond.
A lde rd ice , D. F., and C. R. F o r r e s t e r . 1971. E f f e c t s of s a l i n i t y , temperature , and d i s s o l v e d oxygen on early
J. Fish. Res, development of t h e P a c i f i c cod (Gadus macrocePhalus). Board Can. 28:883-902.
Alverson, F. G. 1963. The food of ye l lowf in and s k i p j a c k tunas i n t h e e a s t e r n
t r o p i c a l P a c i f i c Ocean. [ I n Engl. and Span.] Inter-Am. Trop. Tuna Comm. Bull . 7:293-396.
Alverson, F. G. , and C. L. Pe t e r son . 1963. Synopsis of b i o l o g i c a l d a t a on bigeye tuna Parathunnus s i b i
(Temminck and Sch lege l ) 1844. FA0 F i sh . Rep. No. 6 , Vol. 2:482-514.
Apr i e to , V. L. 1981. F i s h e r y management and extended maritime j u r i s d i c t i o n : The
P h i l i p p i n e tuna f i s h e r y s i t u a t i o n . East-West Center , East-West Environ. P o l i c y I n s t . , Res. Rep. 4 , 7 8 p.
Baker, E., R. A. Feeley, and K. Takahashi. 1979. Chemical composition, s i z e d i s t r i b u t i o n , and p a r t i c l e
morphology of suspended p a r t i c u l a t e matter a t DOMES S i t e s A, B , and C: R e l a t i o n s h i p wi th l o c a l sediment composition. J. L. Bischoff and D. Z . P i p e r ( e d i t o r s ) , Marine geology and oceanography of t h e P a c i f i c manganese nodule province, p. 163-201. Plenum P r e s s , N.Y.
B a r r y , M. 1978. Behavioral response of ye l lowf in tuna , Thunnus a l b a c a r e s , and
kawakawa, Euthynnus a f f i n i s , t o t u r b i d i t y . U.S. Dep. Commer . , Natl. Tech. I n f . Serv., S p r i n g f i e l d , Va., NTIS Rep. No. PB/297106/AS, 31 p. + 21 t a b l e s .
Beers, 9. R., and G. L. Stewar t . 1971. Microzooplanktons i n t h e p lankton communities of t h e upper
waters of t h e eastern t r o p i c a l P a c i f i c . Deep-sea Res. 18:861-883.

40
Benjamin, M. M e , and A. Felmy. 1981. Trace metal exchange between ferromanganese nodules and
a r t i f i c i a l seawater . Mar. Min. 3:151-183.
Bischoff , J. L. , G. R. Heath, and M. Leinen. 1979. Geochemistry of deep-sea sediments from t h e P a c i f i c manganese
nodule province : DOMES S i t e s A , 8 , C. &n J. L. Bischoff and D. Z. Pipe r ( e d i t o r s ) , Marine geology and oceanography of t h e P a c i f i c manganese nodule province , p. 397-436. Plenum Press, N . Y .
Bisha i , H. M. 1960. The e f f e c t of water c u r r e n t s on t h e s u r v i v a l and d i s t r i b u t i o n
of f i s h l a rvae . J. Cons. Cons. I n t . Explor. Mer 25:134-146.
Bjoerke, H. 1978. Food and feeding of young h e r r i n g l a r v a e of Norwegian s p r i n g
spawners. F i s k e r i d i r . Skr. Ser . Havunders. 16:405-422.
Blackburn, H. 1965. Oceanography and t h e ecology of tunas. Oceanogr. Mar. Biol .
Annu. Rev. 3 : 299-322.
1976. Review of e x i s t i n g informat ion on f i s h e s i n t h e deep ocean mining environmental s tudy (DOMES) a r e a of t h e t r o p i c a l P a c i f i c . I n s t . Mar. Resour. Univ. C a l i f . San Diego, La J o l l a , IMR Ref. No. 76-1, 77 p.
Blackburn, M., and R. M. Laurs. 1972. D i s t r i b u t i o n of forage s k i p j a c k tuna (EgLhyn_ngg ~glg&) i n t h e
e a s t e r n t r o p i c a l P a c i f i c . U.S. Dep. Commer., NOAA Tech. Rep. NMFS SSRF-649, 16 p .
Blax te r , J. H. S. 1965. The feeding of h e r r i n g l a r v a e and t h e i r ecology i n r e l a t i o n t o
feeding . C a l i f . Coop. Oceanic F ish . I n v e s t . Rep. 10:79-88.
1969. Development: Eggs and l a rvae . &g W. S. Hoar and D. J. Randal l ( e d i t o r s ) , F i sh physiology, Vol. 111, p. 177-252. Acad. P res s , N.Y.
1980. Vision and t h e feeding of f i s h e s . &g J. E. Bardach, J. J. Magnuson, R. C. May, and J . M. Reinhart ( e d i t o r s ) , F i sh behavior and i t s use i n t h e c a p t u r e and c u l t u r e of f i s h e s , p. 32-56. I n t . Cent. Living Aquat. Resour. Manage., Conf. Proc. 5 , Manila.
B lax te r , J. H. S., and M. E. S t a i n e s . 1971. Food searching p o t e n t i a l i n marine f i s h la rvae . & D. J. Cr i sp
( e d i t o r ) , 4 t h European Marine Biology Symposium, p. 467-485. Cambridge Univ. Press, Cambridge.
Broadhead, G. C., and I. Barrett. 1964. Some f a c t o r s a f f e c t i n g t h e d i s t r i b u t i o n and apparent abundance
of ye l lowf in and s k i p j a c k tuna in t h e e a s t e r n P a c i f i c Ocean. Engl. and Span.] Inter-Am. Trop. Tuna Comm. B u l l . 8:417-473.
[In

4 1
Brock, V. E. 1959. The tuna r e source i n r e l a t i o n t o oceanographic f e a t u r e s . &
Tuna Indus t ry Conference Papers, May 1959, p. 1-11. U.S. F i s h Wildl . Serv., Circ. 65.
Burns, R. E . , B. H. Erickson, J. W. Lave l l e , and E. Ozturgut. 1980. Observat ions and measurements du r ing t h e monitor ing of deep
ocean manganese nodule mining tes ts i n t h e North P a c i f i c , March-May 1978. U.S. Dep, Comer . , NOAA Tech. Memo. ERL-MESA-47, 6 3 p.
Ca lk ins , T. P. 1975. Geographical d i s t r i b u t i o n of ye l lowf in and s k i p j a c k tuna
c a t c h e s i n t h e e a s t e r n P a c i f i c Ocean, and f l e e t and t o t a l c a t c h s t a t i s t i c s , 1971-74. [ I n Engl. and Span.] Inter-Am. Trop. Tuna COUUII. B u l l . 17:l-116.
Chan, A. T . , and G . C. Anderson. 1981. Environmental i n v e s t i g a t i o n of t h e e f f e c t s of deep-sea mining
on marine phytoplankton and primary p r o d u c t i v i t y i n t h e t r o p i c a l e a s t e r n North P a c i f i c Ocean. Mar. Min. 3:121-149.
Ciechomski, J. Dz. de. 1967a. I n f l u e n c e of some environmental f a c t o r s upon t h e embryonic
development of t h e Argent ine anchovy Engrau l i s a n c h o i t a (Hubbs, Marini) . C a l i f . Coop. Oceanic F i sh . I n v e s t . Rep. 11:67-71.
1967b. I n v e s t i g a t i o n s of food and f eed ing h a b i t s of larvae and j u v e n i l e s of t h e Argent ine anchovy Ennrau l i s ancho i t a . C a l i f . Coop. Oceanic F i sh . Inves t . Rep. 11:72-81.
Cole, J. S. 1980. Synopsis of b i o l o g i c a l d a t a on t h e ye l lowf in tuna , Thunnus
a l b a c a r e s (Bonnaterre , 17881, i n t h e P a c i f i c Ocean. JXJ W , H. B a y l i f f ( e d i t o r ) , Synopses of b i o l o g i c a l d a t a on e i g h t s p e c i e s of scombrids, p. 71-150. Inter-Am. Trop. Tuna Comm., Spec. Rep. 2.
Davidoff , E. B. 1963. S i z e and year c l a s s composition of c a t c h , age and growth of
ye l lowf in tuna i n t h e e a s t e r n t r o p i c a l P a c i f i c Ocean, 1951-1961. [ I n Engl. and Span.] Inter-Am. Trop. Tuna Comm. Bull . 8:199-251.
De J e s u s , A. S. 1982. Tuna f i s h i n g g e a r s of t h e P h i l i p p i n e s . United Nations
Development Programme, South China Seas F i s h . Dev. Coord. Prog., SCS/82-WP/lll: 1-47
Dizon, A. E., R. W. B r i l l , and 8. S. H. Yuen. 1978. 111. C o r r e l a t i o n s between environment, physiology, and
a c t i v i t y and t h e e f f e c t s on thermoregulat ion i n s k i p j a c k tuna. G. D. Sharp and A. E . Dizon ( e d i t o r s ) , The p h y s i o l o g i c a l ecology of t unas , p. 233-259. Acad. P r e s s , N.Y.
Eldr idge , M. B. , and P. G. Wares. 1974. Some b i o l o g i c a l obse rva t ions of b i l l f i s h e s t aken i n t h e e a s t e r n
P a c i f i c Ocean, 1967-1970. R . S. Shomura and F. Williams ( e d i t o r s ) , Proceedings of t h e I n t e r n a t i o n a l B i l l f i s h Symposium,

42
Kailua-Kona, Hawaii, 9-12 August 1972. P a r t 2 , Review and Contr ibuted Papers , p. 89-101. U .S . Dep. Commer., N O M Tech . Rep. NMFS SSRF-675
Everha r t , W. H., and R. M. Duchrow. 1970. E f f e c t s of suspended sediment on a q u a t i c environment. U.S.
Bur. Reclam. Proj. , Colorado S t a t e Univ., Completion Rep. No. 14-86-D-6596, 106 p .
Fink, B. D . , and W. H. B a y l i f f . 1970. Migrat ions of yel lowfin and s k i p j a c k tuna i n t h e eastern
P a c i f i c Ocean ae determined by tagging experiments , 1952-1964. [In Engl. and Span.] Inter-Am. Trop. Tuna Comm. B u l l . 15: l -227 .
.
Fonds, H., B . Rosenthal , and D . F . Alderdice. 1974. In f luence of temperature and s a l i n i t y on embryonic development,
l a r v a l growth and number of v e r t e b r a e of t h e g a r f i s h , Belone belone. - I n J. H. S. Blax te r ( e d i t o r ) , The ear ly l i f e h i s t o r y of f i s h , p a 509-525. Springer-Verlag, N.Y.
Forsbergh, E. D. 1980. Synopsis of b i o l o g i c a l d a t a on t h e s k i p j a c k t u n a , Katsuwonus
pelamis (Linnaeus, 17581, i n t he P a c i f i c Ocean. Inter-Am. Trop. Tuna Comm., Spec. Rep. 2 ~ 2 9 5 - 3 6 0 . 3 W . H . Bayl i f f ( e d i t o r ) , Synopses of b i o l o g i c a l d a t a on e i g h t s p e c i e s of scombrids, p. 295- 360 . Inter-Am. Trop. Tuna Comm., Spec. Rep. 2 .
Gardner, M . B. 1981. E f f e c t s of t u r b i d i t y on f eed ing ra tes and s e l e c t i v i t y of
b l u e g i l l s . Trans. Am. F i s h . SOC. 110:446-450.
Gorbunova, N. N. 1975. A review of t h e l a r v a e of scombrid f i s h e s ( P i s c e s , Scombridae)
(Obzor l i c h i n o k skumbrievykn ryb (Pisces, Scombridae)) . -- In T. S. Rass ( e d i t o r ) , Ikh t iop lank ton , glubokovodnye ryby i kal 'mary t r o p i c h e s k i k h vod Mirovogo okeana, 96:23-75. T r . Ins t . Okeanol. Akad. Nauk SSSR. (Engl. t r a n s l . f o r Lang. S e n . B r . , Natl. Mar. F i sh . Serv. , N O M , Wash. D.C., by Amerind Publ. Co. f i t . , Ltd., New De lh i , 1977 , 7 4 p.)
[ In Russ . 1
Gorbunova, N. N., and N. Ya. Lipskaya. 1975. Feeding of l a r v a e of t h e blue m a r l i n , M_akglrg giggicggg
(Pisces, I s t i o p h o r i d a e ) . J. I c h t h y o l . 15:95-101.
Green, R. E. 1967. R e l a t i o n s h i p of t h e thennoc l ine t o success of t r o p i c a l tuna
v e s s e l s based i n C a l i f o r n i a . F i sh . Ind. Res. 3:29-45.
G r i f f i t h , J. S. 1978. E f f e c t s of low temperature on t h e s u r v i v a l and behavior of
t h r e a d f i n shad, &rgggrn_a ggtgqgpg. Trans. Am. Fish. SOC. 107 : 63-70.
Gundersen, K. R., J. S. Corbin, C. L. Hanson, M. L. Hanson, R. B. Haneon, D e J. R u s s e l l , A. S t o l l a r , and 0. Yamada.
1976, S t r u c t u r e and b i o l o g i c a l dynamics of t h e o l i g o t r o p h i c ocean p h o t i c zone off t h e Hawaiian I s l a n d s . Pac. Sc i . 30:45-68.

43
Halpern, D. 1976. Upper ocean c i r c u l a t i o n s t u d i e s i n t h e e a s t e r n t r o p i c a l North
P a c i f i c du r ing September and October 1975. Technology Conference, OTC Pap. No, 2457, p. 357-372.
8 t h Annual Offehore
1977. DOMES upper water phys i ca l oceanography; a component of t h e DOMES Program. F i n a l r e p o r t t o t h e DOMES P r o j e c t . U.S* Dep. Commer., Natl. Tech. I n f , Serv. , S p r i n g f i e l d , Va., NTIS Rep. No. PB283195/AS, 42 p.
Hanamoto, E. 1975. F i she ry oceanography of bigeye tuna. 11. Thermocline and
d i s s o l v e d oxygen content i n r e l a t i o n t o tuna l o n g l i n e f i s h i n g grounds i n t h e e a s t e r n t r o p i c a l P a c i f i c Ocean. [ I n Jpn., Engl. eumm.] La Mer (Bu l l . SOC. Franco-Japonaise d 'oceanographie) 13( 2): 58-71 e
Harada, T., S o Miyashi ta , and €I, Yoneshima. 1978. E f f e c t of water tempera ture on ye l lowf in tuna ha tch ing . [In
Jpn., Engl. synop.] Mem. Fac. Agric. Kinki Univ. 13:29-32.
Harada, T., IC. Mizuno, 0 . Murata, S. Miyashi ta , and H, Hurutani . 1971. On t h e a r t i f i c i a l f e r t i l i z a t i o n and r e a r i n g of l a r v a e i n
ye l lowf in tuna . [ I n Jpn., Engl. synop.] Mem. Fac. Agric . Kinki Univ. 4:145-151 ,
Harada, T., 0 . Murata, and H. Furu tan i . 1973. On t h e a r t i f i c i a l f e r t i l i z a t i o n and r e a r i n g of larvae i n
marusoda, &gig y g ~ g & g g q g g . [In Jpn. , Engl. summ.1 M e m . Fac. Agric. Kinki Univ. 6:113-116.
Harada, T., 0 , Murata, and S. Miyashi ta . 1973. On t h e a r t i f i c i a l f e r t i l i z a t i o n and r e a r i n g of l a r v a e in
h i ra soda , &~rjg &frgge&. [ I n Jpn., Engl. synop.] M e m . Fac. Agric . Kinki Univ. 6:109-112.
1974, On t h e a r t i f i c i a l f e r t i l i z a t i o n and r e a r i n g of larvae i n boni to . [ I n Jpn., Engl. synop.] Mem. Fac. Agric. Kinki Univ, 7:l-4.
Harada, T., 0 . Murata, and S. Oda. 1980. Rearing of and morphological changes i n l a r v a e and j u v e n i l e s of
ye l lowf in tuna . [ I n Jpn., Engl. synop.] Mem. Fac. Agric. Kinki Univ. 13:33-36. (Engl. t r a n s l . by T. Otsu, 1980, 8 p. , Trans l . No. 51; a v a i l a b l e Southwest F ish . Cent., Natl. Mar. Fish. Serv. , N O M , Honolulu, HI 96812.)
Hehstra, N. W., D. K. Damkot, and N. G . Benson. 1969. Some e f f e c t s of s i l t t u r b i d i t y on behavior of j u v e n i l e
largemouth bass and green sunf i sh . U.S. F i s h Wildl. Serv., Bur. Sport F ish . Wildl. , Tech. Pap. No. 20, 9 p.
Herbe r t , D. W. M., and J. C. Merkens. The e f f e c t of suspended minera l s o l i d s on t h e s u r v i v a l of 1961.
t r o u t . I n t . J. A i r Water P o l l u t . 5 : 4 6 - 5 5 .

44
Higgins, B. E. 1970. Juveniic turms c n l l e r t e a i y pielwater t r a w l i n g i n Hawaiian
waters, July-September 1967. Trans. Am. Fish. SOC. 99:60-69.
H i r o t a , J. 1977. DOMES zooplankton. U.S. Dep. Cmmer., Natl. Tech. I n f . S e n . ,
S p r i n g f i e l d , Va., NTIS Rep. No. PB274662/AS, 247 p.
1981. P o t e n t i a l e f f e c t s of deep-sea mine ra l s mining on macrozooplankton i n t h e North E q u a t o r i a l P a c i f i c . 3 : 19-57.
Mar, Min.
HOBS, D. E., W. F. Hettler, Jr., and L. C , Coston. 1974. E f f e c t s of thermal shock on l a r v a l e s t u a r i n e f i sh - - eco log ica l
i m p l i c a t i o n s wi th r e s p e c t t o entrainment i n power p l a n t c o o l i n g systems. I n J. H , S. Blaxter ( e d i t o r ) , The e a r l y l i f e h i s t o r y of f i s h , p. 337-371. Springer-Verlag, N.Y.
Houde, E. D. 1975. E f f e c t s of s tock ing d e n s i t y and food d e n s i t y on s u r v i v a l ,
growth and y i e l d of l abora to ry - rea red larvae of sea bream Agrhgggmgyg ghgghgidhllig ( L . ) (Spa r idae ) . J. F i sh Biol . 7:115-127.
1977. Food c o n c e n t r a t i o n and s tock ing d e n s i t y e f f e c t s on s u r v i v a l and growth of l abora to ry - rea red l a r v a e of bay anchovy Agehog gng&il&i and l i n e d s o l e &higyg linggags. Mar, Biol. 43:333-341,
Houde, E. D., and B. J. Palko. 1970. Laboratory r e a r i n g of t h e c l u p e i d f i s h €Ilgeg&g ~gng_ay_o&g
from F e r t i l i z e d eggs. Mar. Biol. 5:354-358.
Howard, J. K, , and S. Ueyanagi. 1965. D i s t r i b u t i o n and r e l a t i v e abundance of b i l l f i s h e s
( I s t i o p h o r i d a e ) of t he P a c i f i c Ocean. Stud, Trop. Oceanogr. ( M i a m i ) , I n s t . Mar. Sci . , Univ. M i a m i Press, 134 p e
Hunter, J. B. 1972. Swimming and f eed ing behavior of l a r v a l anchovy hgggy1ig
ggrdgx. Fish. Bul l . , U.S. 70: 821-838.
1980. The f eed ing behavior and ecology of marine f i s h l a rvae . J. E. Bardach, J. J. Magnussn, R. C. May, and J. M. Reinhart ( e d i t o r s ) , F i s h behavior and i t s ,use i n t h e c a p t u r e and c u l t u r e of f i s h e s , p. 287-330. I n t . Cent. Living Aquat. Resour. Manage., Conf. Proc. 5 , Manila.
Hunter, J. R., and G. L. Thomas. 1974. E f f e c t of prey d i s t r i b u t i o n and d e n s i t y on t h e sea rch ing and
f eed ing behaviour of larval anchovy hgggylig g ~ f d g g Girard. H. B lax te r ( e d i t o r ) , The e a r l y l i f e h i s t o r y of f i s h , p a 559-57;': Springer-Verlag, N.Y.
I n J.
Pnoue, M., R. Amano, and Y. Iwasaki. 1963. S tud ie s on environments a l l u r i n g s k i p j a c k and o t h e r tunas--I.
On t h e oceanographical c o n d i t i o n of Japan ad jacen t waters and t h e d r i f t i n g subs t ances accompanied by s k i p j a c k and o t h e r tunas. [ I n Jpn., EngP. summ.] Rep. Fish. Res. Lab., Tokai Univ. 1:12-23.

45
Inoue, M. , R. Amano, Y. Iwasaki, and M. Yamauti. 1968. S t u d i e s on environments a l l u r i n g s k i p j a c k and o t h e r tunas--11.
On t h e dr i f twoods accompanied by s k i p j a c k and tunas. Bul l i Jpn. SOC. Sc i . F i s h , 34:283-287.
Inoue, M., K. Tutumi, H. Nagaoka, and T, Nagata. 1974. Some n o t e s on t h e a r t i f i c i a l f e r t i l i z a t i o n and r e a r i n g of
larvae i n t h e s k i p j a c k tuna. S c i . Tech,, Tokai Univ. 8:37-42.
[ I n Jpn., Engl. a b s t r . ] J. Fac. Mar.
I v e r s e n , R. T. B. 1962. Food of a l b a c o r e t u n a , Thunnus germo ( L a d p s d e ) , i n t h e central
and n o r t h e a s t e r n P a c i f i c . U.S. F i s h Wildl. Serv. , F i sh . Bul l . 62 :459-481.
[Japan.] F i s h e r i e s Agency o f , Research Div i s ion . 1970. Annual r e p o r t of e f f o r t and c a t c h s t a t i s t i c s by area on
Japanese tuna l o n g l i n e f i s h e r y , 1968. Fish . Agency Jpn., Res. Div,, 283 p.
1972. Annual r e p o r t o f e f f o r t and c a t c h s t a t i s t i c s by area on Japanese tuna l o n g l i n e f i s h e r y , 1970. Fish . Agency Jpn., Res. Div., 326 p.
1973. Annual r e p o r t o f e f f o r t and c a t c h s t a t i s t i c s by area on Japanese tuna l o n g l i n e f i s h e r y , 1971. 319 p.
F i sh . Agency Jpn., Res. Div.,
1981. Annual r e p o r t of e f f o r t and c a t c h s t a t i s t i c s by area on Japanese tuna l o n g l i n e f i s h e r y , 1979. Fish . Agency Jpn., Res. Div., 243 p.
Jones, S. , and E. G. S i l a s . 1963. Synopsis of b i o l o g i c a l d a t a on s k i p j a c k , Katsuwonus pelamis
(Linnaeus) 1758 ( I n d i a n Ocean). FA0 F i sh . Rep. No. 6 , V o l , 2:663-694.
June, F. C. 1953. Spawning of ye l lowf in tuna i n Hawaiian waters. [U.S.] F i s h
Wildl. Serv., F i sh . Bul l . 54:47-64.
Kawamura, G., and S . Hara. 1980. On t h e v i s u a l f eed ing of m i l k f i s h larvae and j u v e n i l e s i n
c a p t i v i t y . B u l l . Jpn. SOC. S c i . F i s h 46:1297-1300.
Kaya, C. M., A. E. Dizon, S . D. Hendrix, T. K. Kazama, and M. K. K. Queenth.
1982. Rapid and spontaneous ma tu ra t ion , o v u l a t i o n , and spawning of ova by newly cap tu red s k i p j a c k tuna , Katsuwonus Pelamis. F i sh . B u l l . , U.S. 80:393-396.
Kikawa, S. 1966. The d i s t r i b u t i o n of maturing b igeye and ye l lowf in and an
e v a l u a t i o n of t h e i r spawning p o t e n t i a l i n d i f f e r e n t areas i n t h e tuna l o n g l i n e grounds i n t h e P a c i f i c . Rep. Nanlcai Reg. F i s h . Res. Lab. 23:131-208.

46
Kimura, K. 1954. Analys is of s k i p j a c k (Katsuwonus pelamis) s h o a l s i n t h e water
of "Tohoku Kaiku" by i t s a s s o c i a t i o n with o t h e r animals and o b j e c t s based on t h e r eco rds by f i s h i n g boa t s . B u l l . Tohoku Reg. Fish . Res. Lab. 3 , 87 p .
[ I n Jpn. , Engl. summ. 1
King, J. E., and I. I. Ikehara . 1956. Comparative s tudy of food of bigeye and ye l lowf in tuna i n t h e
c e n t r a l P a c i f i c . [U.S.] F i s h Wildl. Serv., F i sh . Bul l . 57:61-85.
Klawe, W. L. 1963. Observa t ions on t h e spawning of f o u r s p e c i e s of tuna
(Neotbunnus macropterus, Katsuwonus pelamis, Auxis tbazard and Euthynnus l i n e a t u s ) i n t h e e a s t e r n P a c i f i c Ocean, baeed on t h e d i s t r i b u t i o n of t h e i r l a r v a e and j u v e n i l e s . Inter-Am. Trop. Tuna Comm. B u l l . 6:447-540.
[ I n Engl. and Span.]
Klawe, W. L., 9. J. P e l l a , and W. S. Leet . 1970. The d i s t r i b u t i o n , abundance and ecology of larval tunas from
t h e e n t r a n c e t o t h e Gulf of C a l i f o r n i a . Inter-Am. Trop. Tuna Comm. B u l l . 14:505-544.
[ I n Engl. and Span.]
[Korea.] F i s h e r i e s Research and Development Agency. 1981. Adnual r e p o r t of c a t c h and e f f o r t s t a t i s t i c s and f i s h i n g
grounds on Korean tuna l o n g l i n e f i s h e r y 1979. F i sh . Res. Dev. Agency, Korea, 351 p.
Kume, S., and J. Joseph. 1969. S i z e composi t ion and sexua l m a t u r i t y of b i l l f i s h caught by t h e
Japanese l o n g l i n e f i s h e r y i n t h e P a c i f i c Ocean east of 130'W. F a r Seas F ish . Res. Lab. (Shimizu) 2:115-162.
Bul l .
Laevastu, T., and H. Rosa, Jr . 1963. D i s t r i b u t i o n and r e l a t i v e abundance of tunas i n r e l a t i o n t o
t h e i r environment. FA0 F i sh . Rep. No. 6 , Vol. 3:1835-1851.
Lasker , R, 1975. F i e l d c r i t e r i a f o r s u r v i v a l of anchovy larvae: The r e l a t i o n
between i n s h o r e c h l o r o p h y l l maximum layers and s u c c e s s f u l f i r s t f eed ing . F i sh . B u l l . U.S. 73:453-462.
Lavelle, J. W e , and E. Ozturgut . 1981. Dispers ion of deep-sea mining p a r t i c u l a t e s and t h e i r effect on
l i g h t i n ocean s u r f a c e layers. Mar. Min. 3:185-212.
Lave l l e , J. W . , E. Ozturgut , E. T. Baker, and S . A. S w i f t . 1982. Discharge and s u r f a c e plume measurements d u r i n g manganese
nodule mining tes ts i n t h e North Equa to r i a l P a c i f i c . Mar. Environ. Res. 7 : 51-70.
EaVio le t t e , P. E., and S. E. Seim. 1969. Monthly c h a r t s of m e a n , minimum, and maximum sea s u r f a c e
tempera ture of t h e North P a c i f i c Ocean. Spec. Publ. SP-123, 58 p.
U.S. Nav. Oceanogr. O f f . ,

47
Lipskaya, N. Ya., and N. N. Gorbunova. 1977. Feeding of s a i l f i s h l a r v a e . Oceanology 17:340-344. (T rans l .
from Okeanolog i i a . Matsumoto, W. M.
1958. D e s c r i p t i o n and d i s t r i b u t i o n of larvae of f o u r s p e c i e s of tuna i n c e n t r a l P a c i f i c waters. U.S. F i s h Wildl. Serv., Fish. Bul l . 58: 31-72.
1966. D i s t r i b u t i o n and abundance of tuna l a r v a e i n t h e P a c i f i c Ocean. -- I n T. A. Manar ( e d i t o r ) , Proceedings, Governor's Conference on C e n t r a l P a c i f i c F i she ry Resources, p. 221-120. S t a t e of Hawaii.
1975. D i s t r i b u t i o n , r e l a t i v e abundance, and movement of s k i p j a c k tuna , K_a&gyygggg gglggig, i n t h e P a c i f i c Ocean based on Japanese tuna l o n g l i n e c a t c h e s , 1964-67. U.S. Dep. Canrmer., NOAA Tech, Rep. NMFS SSRF-695, 3 0 p.
Matsumoto, W. M., and T. K. Kazama. 1974. Occurrence of young b i l l f i s h e s i n t h e c e n t r a l P a c i f i c Ocean.
-- I n R. S. Shomura and F. Williams ( e d i t o r s ) , Proceedings of t h e I n t e r n a t i o n a l B i l l f i s h Symposium, Kailua-Kona, Hawaii, 9-12 August 1972. Par t 2. Review and Contr ibuted Papers, p. 238-251. U.S. Dep. Commer . , NOAA Tech. Rep. NMFS SSRF-675.
Matsumoto, W. M., T. K. Kazama, and D. C. Aasted. 1981 . Anchored f i s h aggregat ing dev ices i n Hawaiian waters. Mar.
F i sh . Rev. 43(9):1-13.
Matsumoto, W. M., R. A. Skil lman, and A. E. Dizon. 1984. Synopsis of b i o l o g i c a l d a t a on s k i p j a c k tuna , Kggggyogyg gglag6g. U.S. Dep. Commer., NOAA Tech. Rep., NMFS, Circ. 451, 92 p.
May, R. C. 1970. Feeding larval m a r i n e f i s h e s i n t h e l a b o r a t o r y : A review.
C a l i f . Coop. Oceanic Fish. I n v e s t . Rep. 14:76-83.
McNeeley, R. L. 1961. Purse s e i n e r e v o l u t i o n i n tuna f , i sh ing . Pac. Fisherman
59( 7) : 27-56
Miller, J. M. 1974. Nearshore d i s t r i b u t i o n of Hawaiian marine f i s h l a rvae : E f f e c t s
o f water q u a l i t y , t u r b i d i t y and c u r r e n t s . &! J. €I. S. Blaxter ( e d i t o r ) , The e a r l y l i f e h i s t o r y of f i s h , p. 217-231. Verlag, N.Y.
Springer-
Moore, J. W., and I. A. Moore. 1976. The b a s i s of food s e l e c t i o n i n f l o u n d e r s , Phg&ichg!!yg f&g!29
(L.) i n t h e Severn e s t u a r y . J. F i s h Biol . 9:139-156.
Morgan, R. P., 11, V. J. Rasin, Jr., and R. L. Copp. 1981. Temperature and s a l i n i t y e f f e c t s on development of s t r i p e d baee
eggs and l a rvae . Trans. Am. Fish. SOC. 110:95-99.

48
Murdy, E. 0. 1980. The commercial h a r v e s t i n g of t u n a - a t t r a c t i n g payaos: A
p o s s i b l e boon f o r small-scale fishermen. I n t . Cent. Living Aquat. Resour. Manage. Newsl. 3(1):10-13.
Murphy, T. C . , and G. T. Sakagawa. 1977. A review and e v a l u a t i o n of estimates of n a t u r a l m o r t a l i t y rates
of tunas . C o l l e c t . Vol. Sc i . Pap., I n t . Comm. Conserv. A t l . Tunas (SCRS-1976) 6: 117-123.
Nat ional Oceanic and Atmospheric Admin i s t r a t ion (NOM) . 1981. Deep seabed mining. F i n a l Programmatic Environmental Impact
Statement. U.S. Dep. Commer., N O M , O f f i c e of Ocean Minerals and Energy, Wash. D.C., Vol. 1, 295 p.
Neumann, D. A., J. M. B'Connor, J. A. Sherk, and K. V. Wood. 1975. Resp i r a to ry and hematological responses of o y s t e r t o a d f i s h
(_O~g_agu_g Q g ) t o suspended s o l i d s . Trans. Am. Fish. SOC. 104:775-781 e
Nishikawa, Y e 1975. Feeding of l a r v a l and j u v e n i l e s k i p j a c k tuna i n r e l a t i o n t o t h e
development of t h e i r stomachs. (Shimizu) 12: 221-236.
Bu l l . Far Seas F i sh . Res. Lab.
Nishikawa, Y., S. Kikawa, M. Honma, and S. Ueyanagi. 1978. D i s t r i b u t i o n a t l a s of l a r v a l t unas , b i l l f i s h e s and r e l a t e d
s p e c i e s - R e s u l t s of l a r v a l surveys by R / V _S_hggyg Hg,rg and S&yg Mlgg 1956-1975. [ I n Jpn., Engl. summ.] Far Seas Fish. Res. Lab., S Ser . 9, 99 p .
Ozre t i ch , R. 9. 1981. Dissolved components of d i scha rge . Mar. Min. 3:95-107.
Ozturgut , E., G. C. Anderson, R. E. Burns, J. W. Lave l l e , and S. A. Swift .
Pre-mining environmental c o n d i t i o n s and a n t i c i p a t e d mining e f f e c t s . U.S. Dep. Commer., NOAA Tech. Memo. ERL MESA-33, 133 p.
1978. Deep ocean mining of manganese nodules i n t h e North P a c i f i c :
Ozturgut , E l , J. W. L a v e l l e , and R. E. Burns. 1981. Impacts of manganese nodule mining on t h e environment: R e s u l t s
from p i l o t - s c a l e mining tes ts i n t h e North E q u a t o r i a l P a c i f i c . R. A. Geyer ( e d i t o r ) , Marine environmental p o l l u t i o n , 2. Dumping and mining, p. 437-474. E l s e v i e r Oceanogr. S 27B, E l s e v i e r Sc i . Publ. Go. , E l s e v i e r , Amsterdam.
Ozturgut , E., J. W. L a v e l l e , and €5. H. Erickson. 1981. Estimated d i s c h a r g e c h a r a c t e r i s t i c s of a commercial nodule
mining o p e r a t i o n . Mar. Min. 3:l-17.
Ozturgut , E., J. W. Lave l l e , 0. S t e f f i n , and S. A. Swif t . 1980. Environmental i n v e s t i g a t i o n du r ing manganese nodule mining
tes ts i n t h e North P a c i f i c i n November 1978. U.S. Dep. Commer., NOAA Tech. Memo. ERL MESA-48, 50 p.

49
Pinus , 6. N. 1974. Some f a c t o r s i n f luenc ing e a r l y s u r v i v a l and abundance of --a CluEgpellg i n t h e Sea of Azov. I n J. H. S. Blaxter ( e d i t o r ) , The e a r l y l i f e h i s t o r y of f i s h , p. 81Z86. Springer-Verlag, N . Y .
P r u t e r , A. T., and D. L. Alverson ( e d i t o r s ) . 1972. The Columbia River e s t u a r y and ad jacen t ocean waters:
Bioenvironmental s t u d i e s . Univ. Wash. Press, S e a t t l e , 868 p.
R e i n t j e s , J. W., and J . E. King. 1953. Food of ye l lowf in tuna i n t h e c e n t r a l P a c i f i c . [U.S.] F i sh
Wildl. Serv., Fish. Bul l . 54: 91-110.
R i l ey , J. D. 1966. Marine f i s h c u l t u r e i n B r i t a i n . V I I . P l a i c e (Plkggeggchgg
~ l _ a g g g g g L.) pos t - l a rva l feeding on &&gei_a g_al_in,a L. n a u p l i i and t h e e f f e c t s of vary ing f eed ing l e v e l s . J. Cons. Cons. I n t . Explor. Mer 3O:204-221.
R i l ey , J. D. , and 6. T. Thacker. 1963. Marine f i s h c u l t u r e i n B r i t a i n . 111. P l a i c e (Plgggsng&$g
~dgggegg ( E . ) ) r e a r i n g i n c losed c i r c u l a t i o n a t Loweetoft , 1961. J. Cons. Cons. I n t . Explor. Mer 28:80-90.
R i t c h i e , D. W. 1970. Fish. I n Gross p h y s i c a l and b i o l o g i c a l e f f e c t s of overboard
s p o i l d i s p o s a i i n upper Chesapeake Bay. Rep, 3 , Univ. Maryland, Col lege Park.
Natl. Resour. I n s t . , Spec.
Robins, 9. P. 1952. Fur the r obse rva t ions on t h e d i s t r i b u t i o n of s t r i p e d tuna ,
&_agpyggs pdgg_ig L., i n e a s t e r n A u s t r a l i a n waters, and i t s r e l a t i o n t o s u r f a c e temperature . Aust. J. Mar. Freshwater Res. 3 : 101-1 10.
Ronquil lo , I. A. 1953. Food h a b i t s of tunas and do lph ins based upon t h e examinat ion of
t h e i r stomach con ten t s . Ph i l ipp . J. Fish. 2:71-83. [ Iesued i n 1954.1
Rosenbauer, R. J. and J. L. Bischof f . 1978. Chemical changes i n seawater caused by resuspens ion of deep-sea
sediments from DOMES S i t e s A, B, and C. Mar. Min. 1:283-304.
Rosenthal , B. 1971. E f f e c t s of "red mud" on embryos and l a r v a e of t h e h e r r i n g -a- CluEgg _h_aggnggg. [ I n Ger., Engl. a b s t r . ] Helgol. Wiss. Meeresunters. 22:366-376.
Rosenthal , H., and G. Hempel. 1970. Experimental s t u d i e s i n feeding and food requirements of
h e r r i n g l a r v a e (E&upga _hgmgngug L.) Jg J. H. Steele ( e d i t o r ) , Marine food cha ins , p. 344-364. Univ. C a l i f . P r e s s , Berkeley.

50
Rothschi ld , B. J. 1965. Hypotheses on t h e o r i g i n of e x p l o i t e d s k i p j a c k tuna (K_g&gyy~ngg
@,a&) i n t h e e a s t e r n and c e n t r a l P a c i f i c Ocean. U.S. F i s h Wildl. Serv. , Spec. Sc i . Rep. Fish. 512, 20 p.
S a i t o , S. 1973. S t u d i e s on f i s h i n g of a l b a c o r e , Xhyggng ghlgggg (Bonna te r r e )
by expe r imen ta l deep-sea tuna long-l ine. Mem. Fac. Fish. Hokkaido Univ. 21 : 107-182.
S a i t o , S., and S. Sasaki .
Ocean--11. Vertical d i s t r i b u t i o n of a lbacore c a t c h by an improved v e r t i c a l long-line. [ I n Jpn. , Engl. a b s t r . ] Bull . Jpn. SOC. Sc i . F i sh . 40: 643-649.
1974. Swimming depth of l a r g e s i z e d a l b a c o r e i n t h e South P a c i f i c
Saksena e V. P., and E. D. Houde. 1972. E f f e c t of food l e v e l on t h e growth and s u r v i v a l of
l abora to ry - rea red l a r v a e of bay anchovy (&_hng g&c_hilli Valenciennes) and s c a l e d s a r d i n e (ggg_eggt~l_a ~ggggsglge Goode C Bean). J. Exp. Mar. Biol . Ecol. 8:249-258.
Schaefer , M. B. 1961. Tuna oceanography programs i n t h e t r o p i c a l c e n t r a l and e a s t e r n
P a c i f i c . C a l i f . Coop. Oceanic Fish. I n v e s t . Rep. 8:41-44.
SeckeP, @. R. 1972. Hawaiian-caught s k i p j a c k tuna and t h e i r p h y s i c a l environment.
F i sh . Bul l . , U.S. 70:763-787.
Sharp, G. D. 1978. I. Behavioral and p h y s i o l o g i c a l p r o p e r t i e s of tunas and t h e i r
e f f e c t s on v u l n e r a b i l i t y t o f i s h i n g gear . I n G. D. Sharp and A. E. Dizon ( e d i t o r s ) , The p h y s i o l o g i c a l ecology of t u n a s , p. 397-449. Acad. P r e s s , N.Y.
Sherk, J. A. 1972. Current s t a t u s of t h e knowledge of t h e b i o l o g i c a l e f f e c t s of
suspended and depos i t ed sediments i n t h e Chesapeake Bay. Chesapeake Sc i . 13 (Suppl.):S137-S144.
Sherk. J. A., J. M. O'Connor, a m D. A. Neumann. 1975. E f f e c t s of suspended and d e p o s i t e d sediments on e s t u a r i n e
environments. E s t u a r i n e r e s e a r c h , geology and eng inee r ing , Vol. 11, p. 541-558. Acad. Press, N.Y.
Shingu, C . , P. K. Tomlinson, and C. L. Pe te r son . A review of t h e Japanese l o n g l i n e f i s h e r y for t unas and 1974.
b i l l f i s h e s i n t h e e a s t e r n P a c i f i c Ocean, 1967-70. [In Engl. and Span.] Inter-Am. Trop. Tuna Comm. B u l l . 16:65-230.
S h i r a i s h i , Y., and T. Shimada. 1972. S t u d i e s on t h e e f f e c t i v e s t o c k i n g of salmonid f i s h , 11:
A c t i v i t y of down mig ra t ion of himemasu, Oncorhvnchus nerka, soon a f t e r s t o c k i n g w i t h s p e c i a l r e f e r e n c e t o t h e f a c t o r s of t h e i r mig ra t ion . [In Jpn., Engl. summ.] B u l l . Freshwater F i sh . Res. Lab. (Tokyo) 22:1-12.

51
Shomura, R . S., and W. M. Matsumoto. 1982. S t r u c t u r e d f l o t s a m as f i s h aggrega t ing dev ices . U.S. Dep.
Commer., NOAA Tech. Memo. NMFS, NOAA-TM-NMFS-SWFC-22, 9 p.
S t r a s b u r g , D. W . 1960. Estimates of l a r v a l tuna abundance i n t h e c e n t r a l P a c i f i c .
U.S. F i s h W i l d l . Serv. , F i sh . B u l l . 60:231-255.
1970. A r e p o r t on t h e b i l l f i s h e s of t h e c e n t r a l P a c i f i c Ocean. B u l l . Mar. S c i . 20:575-604.
S t r a s b u r g , D. W . , E . C. Jones , and R . T. B. I ve r sen . 1968. Use of a small submarine f o r b i o l o g i c a l and oceanographic
r e s e a r c h . J . Cons. Cons. I n t . Explor. Mer 31:410-426.
Sund, P. N . , M. Blackburn, and F. Williams. 1981. Tunas and t h e i r environment i n t h e P a c i f i c Ocean: A review.
Oceanogr. Mar. B io l . Annu. Rev. 19:443-512.
Swenson, W. A., and M. 1,. Matson. 1976. I n f l u e n c e of t u r b i d i t y on s u r v i v a l , growth, and d i s t r i b u t i o n of
l a rva l l a k e h e r r i n g (Corenonus a r t e d i i ) . Trans. Am. F i s h . SOC. 105: 541 -545.
Sykora, J. L., R . J. Smith, and M. Synok. 1972. E f f e c t of l i m e n e u t r a l i z e d i r o n hydroxide suspens ions on
j u v e n i l e brook t r o u t ( S a l v e l i n u s f o n t i n a l i s , M i t c h e l l ) . Water Res. 6:935-950.
Tanaka, T. 1978. Atlas of s k i p j a c k tuna f i s h i n g grounds i n sou the rn waters, 1977
f i s h i n g season (June 1977-April 1978). Tohoku Reg. F i s h . Res. Lab., 8 p. t e x t , 16 c h a r t s . (Engl. t r a n s l . by T. O t s u , 1978, 33 p . , T r a n s l . No. 30; a v a i l a b l e Southwest F i sh . Cent., Natl. Mar. F i sh . Serv. , NOAA, Honolulu, H I 96812.)
Turner , J. L., and T. C. F a r l e y . 1971. E f f e c t s of t empera tu re , s a l i n i t y , and d i s s o l v e d oxygen on t h e
s u r v i v a l of s t r i p e d bass eggs and larvae. C a l i f . F i s h G a m e 57 :268-273.
Uda, M. 1933. The s h o a l s of "katuwo" and t h e i r ang l ing . [ I n Jpn., Engl.
s u m . ] Bu l l . Jpn. SOC. S c i . F i sh . 2:107-111. (Engl. t r a n s l . by W. G. Van Campen, 1952, In U.S. F i s h Wildl. Serv. , Spec. S c i . Rep. F i sh . 83:68-78.)
1957. A c o n s i d e r a t i o n on t h e long years t r e n d of t h e f i s h e r i e s f l u c t u a t i o n i n r e l a t i o n t o sea cond i t ions . B u l l . Jpn. SOC. sci. F i sh . 23 :368-372.
Ueyanagi, S. 1963. Methods f o r i d e n t i f i c a t i o n and d i s c r i m i n a t i o n of t h e larvae of
f i v e i s t i o p h o r i d s p e c i e s d i s t r i b u t i n g i n t h e Indo-Pacif ic . Jpn., Engl. synop.] Rep. Nankai Reg. F i sh . Res. Lab. 17:137-150.
[ I n

52
1967. On t h e spawning grounds of tunas. Tuna F i sh . 60:15-20. (Engl. t r a n s l . In Fish. Res. Board Can., T r a n s l . Ser . No. 1100, 1968, 14 p.)
1969. Observat ions on t h e d i s t r i b u t i o n of tuna larvae i n t h e Indo-Pacif ic Ocean with emphasis on t h e d e l i n e a t i o n of t h e spawning areas of a l b a c o r e , Thunnus a l a lunga . [ I n Jpn., Engl. synop.] Bul l . F a r Seas F i sh . Res. Lab. (Shimizu) 2:177-256.
1974. A review of t h e world commercial f i s h e r i e s f o r b i l l f i s b e s . In R. So Shomura and F. Williams ' ( e d i t o r s ) , Proceedings of t h e I n t e r n a t i o n a l B i l l f i s h Symposium, Kailua-Kona, Hawaii, 9-12 August 1972. P a r t 2. Review and Contr ibuted Papers , p. 1-11. U.S. Dep. Commer., NOAA Tech. Rep. NMFS SSRF-675.
Ueyanagi, S., Y. Nishikawa, and T. Matsuoka. 1974. A r t i f i c i a l f e r t i l i z a t i o n and larval development of s k i p j a c k
tuna , Katsuwonus pelamis. [ I n Jpn., Engl. summ,] Bul l . F a r Seas F i sh . Res. Lab. (Shimizu) 10:179-188.
Ueyanagi, S., and P. G . Wares. 1975, Synopsis of b i o l o g i c i l d a t a on s t r i p e d mar l in , T e t r a p t u r u s
audax ( P h i l i p p i ) , 1887. R. S. Shomura and F. Williams ( e d i t o r s ) , Proceedings of t h e I n t e r r i a t i o n a l B i l l f i s h Symposium, Kailua-Kona, Hawaii, 9-12 August 1972. P a r t 3. Species Synopses, p. 132-159. U.S. Dep. Commer., NOAA Tech. Rep. NMFS SSRF-675.
Uotani , I., A. Izuha, and K. Aiai . 1978. Food h a b i t s and s e l e c t i v e f eed ing of anchovy larvae (EnP;raulis
j a p o n i c a ) . Bul l . Jpn. SOC. Sc i . F i s h . 44:427-434.
Uotani , I., K . Matsuzaki, Y. Makino, K. Noda, 0. Inamura, and M. Horikawa.
area northwest of A u s t r a l i a . B u l l . Jpn. SOC. Sc i . F i s h , 47: 1981. Food h a b i t s of lamae of tunas and t h e i r r e l a t e d s p e c i e s i n t h e
1165-1172.
Vinyard, G. L., and W. J. O'Brien. E f f e c t s of l i g h t and t u r b i d i t y on t h e react ive d i s t a n c e of 1976.
b l u e g i l l (Lepomis macrochirus) . J . F i sh . Ses. Board Can. 33:2845-2849.
Waldron, K. D. 1964. F i s h schoo l s and b i r d f l o c k s i n t h e c e n t r a l P a c i f i c Ocean,
1950-61. U.S. F i s h Wildl. Serv. , Spec. S c i . Rep. F i sh . 464, 20 p.
Waldron, K. D., and J. E. King. Food of s k i p j a c k i n t h e c e n t r a l P a c i f i c . 1963. FAQ F i s h . Rep. No. 6 ,
Val. 3: 1431-1457 e
Wallen, I. E. 1951. The d i r e c t e f f e c t of t u r b i d i t y on f i s h e s . B u l l . Qkla . Agric.
Mech. Col l . , Biol. Se r . 2, 48 (2 ) , 27 p.
Watanabe, H. 1958. On t h e d i f f e r e n c e of t h e stomach c o n t e n t s of t h e y e l l o w f i n and
b igeye t u n a s from t h e western e q u a t o r i a l P a c i f i c . [ I n Jpn., Engl.

53
summ.1 Rep. Nankai Reg. F i s h . Res. Lab. 7:72-81. G , Y. Beard, 1958, 15 p.; a v a i l a b l e Southwest F ish . Cen t . , Natl. Mar. F ish . Serv. , N O M , Honolulu, H I 96812.)
(Engl. t ransl . by
1960. Regional d i f f e r e n c e s i n food composition of t h e tuna8 and marlins from s e v e r a l oceanic a r e a s . [ I n Jpn., Engl. summ.] Rep. Nankai Reg. F i sh . Res. Lab. 12:75-84.
Williams, F. 1970. Sea s u r f a c e temperature and t h e d i s t r i b u t i o n and apparent
abundance of sk ip j ack (Katsuwonus uelamis) i n t h e e a s t e r n P a c i f i c Ocean, 1951-1968. [ I n Engl. and Span.] Inter-Am. Trop. Tuna Comm. B u l l . 15:231-281.
1972. Considera t ion of t h r e e proposed models of t h e mig ra t ion of young sk ip j ack t u n a (Katsuwonus uelamis) i n t o t h e e a s t e r n P a c i f i c Ocean. F i sh . B u l l . , U.S. 70:741-762.
Wilson, K, W . , and P. M. Connor. 1976. The e f f e c t of ch ina c l a y on t h e f i s h of S t A u s t e l l and
Mevagissey Bays. J . Mar. Biol . Aesoc. U. K. 56:769-780.
Wyatt, T. 1972. Some e f f e c t s of f o o l d e n s i t y on t h e growth and behaviour of
p l a i c e l a rvae . Marl B i c l . 14:210-216.
Yamanaka, H., Y. K u r o h i j i , and J. Morita. 1966. General r e s u l t s of t h e i n v e s t i g a t i o n i n t h e south wes tern
P a c i f i c Ocean by t h e f i s h - f i n d e r . [ I n Jpn., Engl. surmm.1 Rep. Nankai Reg. Fish. Res. Lab. 24: 115-127.
Yoshida, H. 0. 1966. Skip jack tuna spawning i n t h e Marquesas I s l ands and Tuamotu
Archipelago. U.S. F i s h Wildl. Serv., Fish. Bul l . 65:479-488.
Yuen, H. S. H. 1955. Matur i ty and f ecund i ty of bigeye tuna i n t h e P a c i f i c . [ U . S . ]
F i s h Wildl. Serv. , Spec. Sc i . Rep. F ish . 150, 30 p.
1970. Behavior of sk ip j ack tuna , K_apu_woggg ~ g h g i g , a s determined by t r a c k i n g wi th u l t r a s o n i c devices . J. Fish. Res,, Board Can. 27 : 207 1-207 9.

RECENT TECHNICAL MEMORANDUMS
1
k
4
Copies of this and other NOAA Technical Memorandums are available from the National Technical Information Service, 5285 Port Royal Road, Springfield, VA 22167. Paper copies vary In price. Mlcrofishe copies cost $3.50. Recent issues of NOAA Technical memorandums from the NMFS Southwest Fisheries Center are listed below:
NOAA TM-NMFS SWFC 34
35
36
37
38
39
40
41
42
43
“Some data on dolphin mortality In the eastern tropical Pacific tuna purse seine fishery prior to 1970.” T. D. SMITH and N. C. H. LO (July 1983)
“Precision of age determination of northern offshore spotted dolphins.” S. B. REILLY, A. A. HOHN, and A. C. MYRICK, JR. (August 1983)
“Recovery of adult green turtles observed or originally tagged at French Frigate Shoals, Northwestern Hawaiian Islands.” G. BALAZS (August 1983)
“Report of the workshop on long-range planning for the North Pacific albacore fishery.” D. J. MACKETT, Editor (November 1983)
Distribution of four dolphins (Stenella spp. and Delphhus delphls) In the eastern tropical Pacific, with an annotated catalog of data sources. W. F. PERRIN, M. D. SCOll, G. J. WALKER, F. M. RALSTON and D. W. K. AU (Qecember 1983)
Annotated references to techniques capable of assessing the roles of cephalopods in the eastern tropical Pacific Ocean, with emphasis on pelagic squids. J. B. HEDGEPETH (December 1983)
Summary of environmental and fishing information on Guam and the commonwealth of the Northern Mariana Islands: Historical background, description of the islands, and review of the climate, oceanography, and submarine topography. L. G. ELDREDGE (December 1983)
Diving patterns of the Hawaiian Monk Seal, Lisianski Island, 1982. F. V. SCHLEXER (February 1984) “The Hawaiian Monk Seal of Laysan Island: 1982” D. J. ALCORN (April 1984) Atlas of airborne sea surface temperature observations In nearshore California waters, 1980-1983. P.N. SUND (July 1984)