postsettlement patterns of habitat use by sciaenid … patterns of habitat use by sciaenid fishes in...
TRANSCRIPT

Estuaries Vol. 21, No. 2, p. 318-327 June 1998
Postsettlement Patterns of Habitat Use by Sciaenid
Fishes in Subtropical Seagrass Meadows
JAY R. ROOKER~,~ SCOTT A. HOLT MANUEL A. SOT0 G. TOAN HOLT ~-J--- Matine Science Institute The University of Texas at Austin 750 Channelview Drive Port Aransas, Texas 78373
ABSTRACT: Spatial and temporal patterns of distribution and abundance were examined for postsettlement sciaenids collected from seagrass meadows in the Aransas Estuary, Texas. Overall, 5443 sciaenid larvae and early juveniles were identified from biweekly epibenthic sled collections taken from August 1994 to August 1995. Eight species were present in seagrass meadows, witb five accounting for over 99.9% of sciaenids collected: silver perch (B&die& chrysoura), spotted seatrout (Cynoscion nebulosus), spot (Lekxtomus xonth~r~~), Atlantic croaker (Micropogonias undulutus), and red drum (Sckenops ocellati). Settlement to seagrass meadows was partitioned temporally with little overlap among the five species. Postsettlers from inshore spawners (B. chrysoura, C. nebulosus, S. ocellatw) inhabited seagrass meadows during the spring aud s- er, while individuals from offshore spawners (L. xanthurus, M. undulatus) were present in the late fall and winter. Densities of B. chrysouru, C. nebulosus, S. ocellatus were highest for small individuals (4-8 mm SL) and these taxa remained in seagrass sites through the early juvenile stage. Conversely, L. xanthurus and M. undulatus main- tained longer pelagic periods and generally entered seagrass meadows at larger sizes (lo-14 mm SL). Moreover, these taxa were only temporary residents of selected seagrass meadows, apparently migrating to alternative habitats shortly after arrival. During peak settlement, mean and maximum densities among species ranged from 0.1 m-r to 0.8 m-* and 0.7 m-r to 23.8 rne2, respectively. Density and mean size of postsettlement sciaenids differed significantly between seagrass species (Halodule wright& Thahzssia testudinum) and among sites within the estuary.
Introduction Seagrass meadows provide critical nursery habi-
tat for a diversity of fishes during their early life stages (Olney and Boehlert 1988; Bell and Pollard 1989). Increased abundance and diversity of fishes associated with seagrass meadows have been relat- ed to the physical complexity of the seagrass can- opy. Increased complexity provides such benefits as refuge from predators (Heck and Orth 1980; Stoner 1982; Ryer 1988; Savino and Stein 1989) and increased food supply (Orth 1992). In addi- tion, the physical nature of the seagrass canopy may affect the hydrodynamics of larval recruitment (Eckman 1983) and influence the chance of settle- ment by pelagic larvae (Bell et al. 1987).
Larvae of several estuarine fishes settle into sea- grass meadows following a dispersive pelagic phase (Bell and Pollard 1989; Day et al. 1989; Leis 1991). Since postsettlement mortality is typically high and variable, recruitment success or failure can be
1 Corresponding author. * Present address: University of Maryland, Center for Environ-
mental Science, Chesapeake Biological Laboratory, P.O. Box 38, Solomons, Maryland 20688-0038.
0 1998 Estuarine Research Federation 318
linked to differential survival during this period (Jones 1991; Carr and Hixon 1995; Rooker 1997). As a result, studies focusing on the postsettlement phase are essential for understanding the dynamics of early life. However, detailed ecological assess- ments of habitat requirements during the postset- tlement phase are incomplete for many estuarine species. This information is required for effective conservation and management of the estuarine fish-habitat complex and will provide the founda- tion for future studies addressing the causes and consequences of early life mortality.
Species from the family Sciaenidae (drums) are vital to the recreational fishery in the Gulf of Mex- ico, and several sciaenids exploit seagrass meadows during early life (e.g., Stoner 1983; Tolan et al. 1997). From limited data, it appears that certain sciaenids immigrate to seagrass meadows following a pelagic interval of approximately 15-30 d and remain in these habitats for weeks to months (Holt et al. 1983; Peters and McMichaell987; McMichael and Peters 1989; Rooker and Holt 1997). To date, few studies have examined postsettlement patterns of habitat use in seagrass meadows and important life history data (e.g., duration and timing of set-

Postsettlement Sciaenids in Seagrass Meadows 319
tlement, size-at-settlement, and spatial distribu- tion) are lacking for many species.
The purpose of the present study was to examine sciaenid utilization of seagrass meadows during the postsettlement phase. Specific objectives of this re- search were to describe quantitatively the sciaenids inhabiting seagrass meadows in the Aransas Estu- ary, Texas, determine the temporal patterns of set- tlement, including the timing and duration of events, and determine the consistency of habitat use over different spatial scales.
Methods %XJDlY hEA
The study was conducted in the Aransas Estuary, _._ 0c ” 1..--9 L_..l-..:lL __.__ f~_ _. _ PA_ ~_ ~~ _ an oo.* Km', uar-Dullr eslcudryconasting 01 two ma-
jor bays (Aransas Bay, Redfish Bay) oriented par- allel to the coast. The estuary is located in the vi- cinity of the Aransas Pass tidal inlet (Fig. l), which is approximately 400 m wide and connects Gulf of Mexico waters to inshore habitats. Seagrass mead- ows within the estuary are dominated by shoal ~T~PEQ / Unln&110 w&rrhtl’i\ 2nd tllrtlP CTT>CC / Thnlnrrin w-“” \‘LW”“W~yY w,“~‘““y”, %+*-La GUI LA.. b’““” \ 11 ,Y..“vIyUYYW
testudinum). Both species occur primarily in shal- low water (ca. <l m), low-energy areas and are considered the major primary producers in these areas (Zieman and Wetzel 1980). Water tempera- ture, salinity, and turbidity are variable within the estuary; however, our study sites were in close prox- imitv to the tidal inlet (<lo km) and maintained con&tions relatively similar to nearshore waters in the Gulf of Mexico.
POSTSETTLEMENT SURVEY Densities were estimated by enumerating sciaen-
ids in biweekly samples taken from August 1994 to August 1995 (postsettlers were defined as individ- uais 140 mm standard iength collected in seagrass meadows). Collections were taken at six sites with- in the estuary (Fig. 1): Aransas Bay 1, 2, and 3 (ABl, AB2, Al33); Redfish Bay 1 and 2 (RBl, RB2); and Corpus Christi Bay (CBl). At each of the six locations, H. wrightii meadows were sampled. At one site within each bay (ABl, RBl, CBl), co-oc- curring, monotypic stands (>90% single species) of H. wrightii and x testwdinum were present (des- ignated paired sites) and sampled separately. Sea- grass meadows at paired sites were adjacent to one another and less than 50 m apart.
Postsettlement sciaenids were collected from sea- grass meadows with an epibenthic sled. The sled opening measured 0.75 m (length) X 0.50 m (height) and was equipped with a 505~pm conical plankton net. The sled was towed by hand at a con- stant speed (-0.7 m s-l) through seagrass mead- ows. Triplicate sled tows were made at each sam- pling site on each trip. Sled tows were 20 m in
length and the area of seagrass sampled per tow was 15.0 m2. Individuals were preserved in 95% ethanol immediately after capture. Standard lengths (SL) were measured in the laboratory to the nearest 1.0 mm.
Throughout the postsettlement survey, environ- mental conditions were recorded in the Aransas Pass at the University of Texas, Marine Science In- stitute Pier Laboratory. In addition, depth, salinity, and water temperature were measured from each site on all sampling trips. Estimates of seagrass complexity were also made from samples collected with a 7.5-cm diameter coring device. Shoot den- sity, blade height, and number of blades per shoot were measured from three replicate cores taken K,-,.- ,,,.1, n:,, ,e ?z n -_ :,c-,.,“l” l?,c:-..C,.” ,c II”lll C;aL.ll JILr; aL J1 ill” IIILC1 “al>. IjJUllldLCJ “I blade height and blades per shoot for each core were mean values based on 10 blades and shoots, respectively.
STATISTICAL ANALYSIS Three-way factorial analysis of variance (ANO-
VA) was applied to postsettlement density data (main effects: habitat, site, date). This analysis was performed separately on each taxon and based only on samples from paired sites (ABl, RBl, CBl) . Two-way factorial ANOVA (main effects: site, date) was used to examine variation in density among the six H. wrightii sites (ABl, AB2, AB3, RBl, RB2, CBl). Prior to ANOVA testing, density data were log, (x + 1) transformed to minimize heteroscedasticity. Since seasonal patterns of re- cruitment were pronounced, many samples con- tained zero values. As a result, the assumption of multivariate normality (normality of error terms) was not met in some cases. ANOVA is robust to most types and magnitudes of departure from nor- maiity and, therefore, non-normaiity is uniitiy to compromise results (Underwood 1981). Tukey’s HSD test was used to find a posteriori differences (CY = 0.05) among sample means. Mean sizes of sciaenids in H. wrightii and ?: testudinum were com- pared using a one-way ANOVA. Due to heterosce- dasticity and unequal sample sizes, means were weighted by the reciprocal of the sample variances of the group means (JMP 1989).
Results ENVIRONMENTAL CONDITIONS AND
SITE CHARACTERISTICS Temporal variation in water temperature and sa-
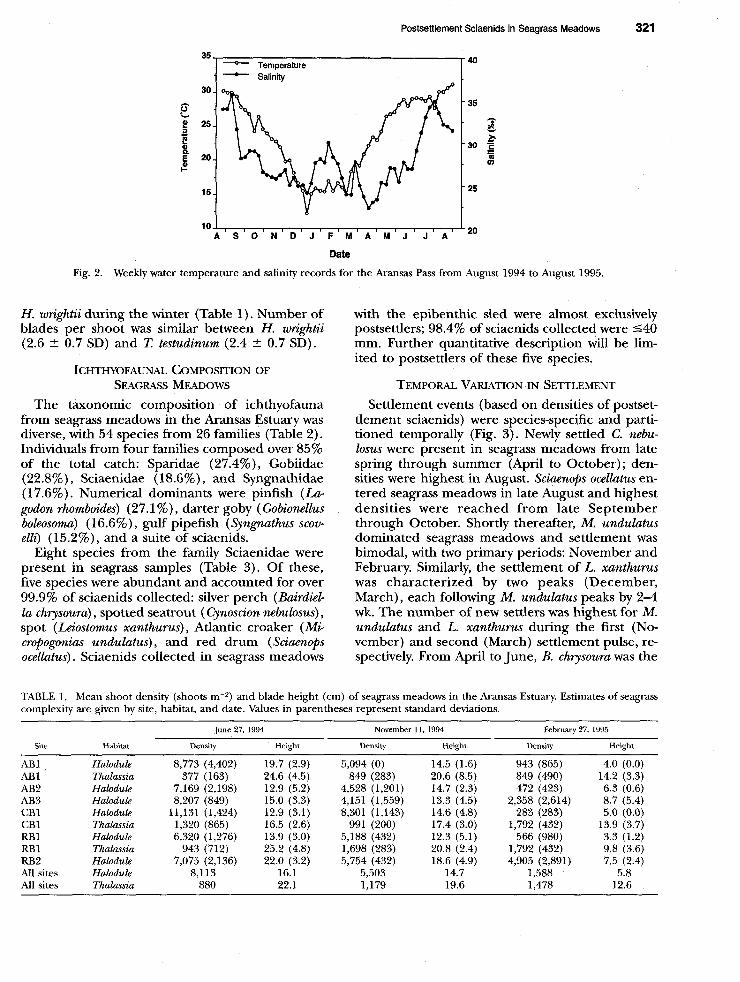
linity in the Aransas Pass were pronounced (Fig. 2) ; temperature and salinity ranged from 12.2”C to 31.1”C and 22.5%0 to 35.7%0, respectively. Mea- surements were also recorded each trip at the dif- ferent sampling sites within the Aransas Estuary; however, it is difficult to compare temperature and

320 J. FL Rooker et al.
n
^^____^ CiJHPiJS 0
CHRIST/ BA Y
,27’54’
.27”50
97'06'
Fig. 1. Map of Aransas Estuary showing location of sampling sites in Aransas Bay (ABl, AB2, AB3), Redfish Bay (RBI, RB2), and Corpus Christi Bay (CB-1).
salinity profiles among sites due to the time-of-day effect. In general, temperature changes in the Aransas Estuary closely matched recordings at the Aransas Pass, but larger fluctuations were apparent in the shallower waters of the estuary. Salinity mea- surements at the tidal pass were representative of conditions at study sites within the estuary.
Monotypic stands of H. wrightii and ?: testudinum differed in physical structure and complexity. On
average, lY testudinum was present at depths lo-20 cm > H. wrightii (0.58 m and 0.44 m, respectively). Shoot densities in H. tightii were approximately 5-10 times higher than ‘T testudinum during the summer and fall, while blade heights were gener- ally lower. Shoot densities and blade heights varied among sites and seasonal patterns were observed (Table 1). The most salient feature was the marked decline in the shoot density and blade height of
Postsettlement Sciaenids in Seagrass Meadows 321
- Temperature - Salinity
A S 0 N D J F M A M J J A 2o
Date
Fig. 2. Weekly water temperature and salinity records for the Aransas Pass from August 1994 to August 1995.
H. wrightii during the winter (Table 1). Number of blades per shoot was similar between H wr;,ghtii (2.6 + 0.7 SD) and ?: testudinum (2.4 2 0.7 SD).
ICHTHYOFAUNAL COMPOSITION OF SEAGRASS MEADOWS
The taxonomic composition of ichthyofauna from seagrass meadows in the Aransas Estuary was diverse, with 54 species from 26 families (Table 2). Individuals from four families composed over 85% of the total catch: Sparidae (27.4%), Gobiidae (22.8%)) Sciaenidae (18.6%)) and Syngnathidae (17.6%). Numerical dominants were pinfish (Lu- godon rhomboides) (27.1%) , darter goby (Co&one&s bokosoma) (16.6%)) gulf pipefish (Syngrzathus scov- eZZi) (15.2%)) and a suite of sciaenids.
Eight species from the family Sciaenidae were present in seagrass samples (Table 3). Of these, five species were abundant and accounted for over 99.9% of sciaenids collected: silver perch (BairdieZ- la chrysowu) , spotted seatrout (Cynoseion nebulosus) , spot (Leiostomus xunthurus) , Atlantic croaker (Mi- cqbogonias undulatus) , and red drum (Sciaenops ocellatus) . Sciaenids collected in seagrass meadows
with the epibenthic sled were almost exclusively postsettlers; 98.4% of sciaenids collected were 540 mm. Further quantitative description will be lim- ited to postsettlers of these five species.
TEMPORAL VARIATION IN SETTLEMENT
Settlement events (based on densities of postset- tlement sciaenids) were species-specific and parti- tioned temporally (Fig. 3). Newly settled C. nebu- Zosus were present in seagrass meadows from late spring through summer (April to October); den- sities were highest in August. Sciaeno@ ocellatus en- tered seagrass meadows in late August and highest densities were reached from late September through October. Shortly thereafter, M. undulatus dominated seagrass meadows and settlement was bimodal, with two primary periods: November and February. Similarly, the settlement of L. xunthurus was characterized by two peaks (December, March), each following M. undulatus peaks by 2-4 wk. The number of new settlers was highest for M. undulatus and L. xanthurus during the first (No- vember) and second (March) settlement pulse, re- spectively. From April to June, B. chrysoura was the
TABLE 1. Mean shoot density (shoots m-2) and blade height (cm) of seagrass meadows in the Aransas Estuary. Estimates of seagrass complexity are given by site, habitat, and date. Values in parentheses represent standard deviations.
Site
ABl. Al31 AB2 AB3 CBl CBl RBl RBl RB2 All sites All sites
June 27, 1994 November 11, 1994 February 27, 1995
Density Height Density Height D.ZlSi~ Height
8,773 (4,402) 19.7 (2.9) 5,094 (0) 14.5 (1.6) 943 (865) 4.0 (0.0) 377 (163) 24.6 (4.5) 849 (283) 20.6 (8.5) 849 (490) 14.2 (3.3)
7,169 (2,198) 12.9 (5.2) 4,528 (1,201) 14.7 (2.3) 472 (423) 6.3 (0.6) 8,207 (849) 15.0 (3.3) 4,151 (1,559) 13.3 (4.5) 2,358 (2,614) 8.7 (5.4)
11,131 (1,424) 12.9 (3.1) 8,301 (1,143) 14.6 (4.8) 283 (283) 5.0 (0.0) 1,320 (865) 16.5 (2.6) 991 (200) 17.4 (3.0) 1,792 (432) 13.9 (3.7) 6,320 (1,276) 13.9 (3.0) 5,188 (432) 12.3 (5.1) 566 (980) 3.3 (1.2)
943 (712) 25.2 (4.8) 1,698 (283) 20.8 (2.4) 1,792 (432) 9.8 (3.6) 7,075 (2,136) 22.0 (3.2) 5,754 (432) 18.6 (4.9) 4,905 (2,891) 7.5 (2.4)
8,113 16.1 5,503 14.7 1,588 5.8 880 22.1 1,179 19.6 1,478 12.6
Habitat
Halodule Thalussia Haloduk Halodule Halodule Thalmsia Haloduk Thalassia Halodule Halodule Thalassia

322 J. R. Rooker et al.
TABLE 2. Number and percent composition of fishes (by fam- TABLE 3. Number and percent composition of sciaenids col- ily) collected from seagrass meadows in the Aransas Estuary. lected from seagrass meadows in the Aransas Estuary.
Family Number PelX.Zllt
Sparidae 8,014 27.37 Gobiidae 6,686 22.83 Sciaenidae 5,443 18.59 Syngnathidae 5,149 17.59 Cyprinodontidae 1,046 3.57 Clupeidae 894 3.05 Haemulidae 693 2.37 Mugilidae 465 1.59 Gerriidae 390 1.33 Atherinidae 179 0.61 Engraulidae 116 0.40 Cynoglossidae 38 0.13 Bothidae 38 0.13 Lutjanidae 33 0.11 Blenniidae 32 0.11 Ophichthidae 15 0.05 Bauachoididae 12 0.04 Synodontidae 11 0.04 Other taxa 26 0.08 Total 29,280 100
Species Common Name Number Percent=
Leiostomus xanthurus spot 2,015 37.0 Bairdiellu chrysoura silver perch 1,271 23.4 Sciaenops ocellatus red drum 1,170 21.5 Micropogonias undulatus Atlantic croaker 723 13.3 Cynoscion nebubsus spotted seatrout 258 4.7 Pogonias cromis black drum 4 0.1 Cynosn’on arenarius sand seatrout 1 co.1 Menticirrhus ama’canus southern kingfish 1 co.1
a Based on total number of sciaenids collected.
50.
40.
30.
Cynoscion nebutosus
dominant sciaenid in seagrass meadows with peak densities occurring in May.
Sciaenops ocektus
SIZE-AT-SETTLEMENT
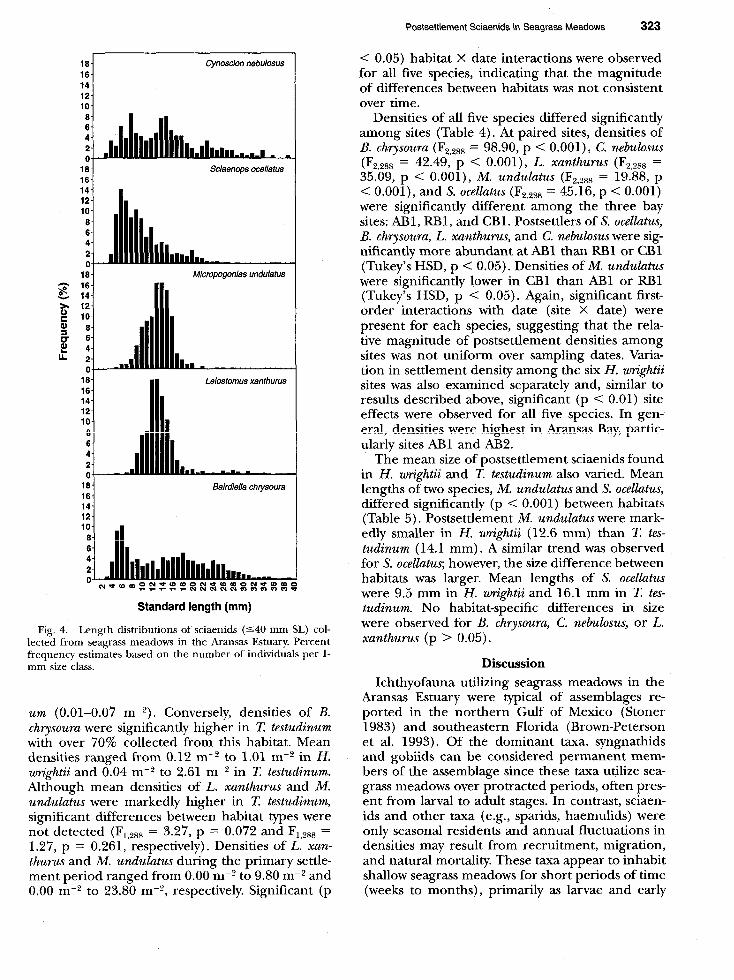
Bairdiella chrysoura, C. nebulosus, and S. ocellatus were first detected at approximately 3-4 mm and peak numbers of settlers entered seagrass meadows at 4-8 mm (Fig. 4). Individuals from these species were present at collection sites throughout much of the early juvenile stage (540 mm). In contrast, newly settled L. xanthurus and M. undulatus were considerably larger. Although individuals <lo mm were collected in seagrass meadows, densities were highest for individuals lo-14 mm. Moreover, L. xanthurus and M. undulatus remained in these hab- itats for short periods of time before moving to other habitats; individuals >20 mm were caught infrequently.
Micropogonias undulatus
Leiostomus xanthurus
SPATIAL DISTRIBUTION AND ABUNDANCE
Densities of postsettlers varied markedly be- tween habitats Andy among sites at paired sites (ABl, RBl, CBl). A significant habitat effect was observed for three species: B. chrysoura (F1,*s8 = 9.48, p = O.OOl), C. nebulosus (F1,288 = 4.07, p = 0.045), and S. ocellatus (F1,288 = 18.11, p < 0.001). Densities of C. nebulosus and S. ocellatus were ap- proximately 2-3 times higher in H. wrightii than ?: testudinum. Mean densities of S. ocellatus during the primary settlement period (defined as trips with >50% of samples with target species) were 0.34- 1.79 mm2 and 0.04.0.73 mP2 in H. wrightii and ?: testudinum, respectively (Table 4). Although mean densities of C. nebulosus were lower, trends were similar: H. wrightii (,0.04-0.21 mm2) and X testudin-
Bairdiella chrysoura
1994 1995 Date
Fig. 3. Relative abundance of sciaenids (540 mm SL) col- lected from seagrass meadows during biweekly sampling trips in the Aransas Estuary. Percent frequency estimates based on the number of individuals per sampling date.
Sciaenops ocellatus
12 10
6- 6- 4- 2- O-
Micmpogonias undulatus
16. 16. 14.
12. lo-
6-
6-
Leiostomus xanthurus
_lll I
16 16 14 12
Bairdiella chrysoura
Standard length (mm)
Fig. 4. Length distributions of sciaenids (540 mm SL) col- lected from seagrass meadows in the Aransas Estuary. Percent frequency estimates based on the number of individuals per l- mm size class.
urn (0.01-0.07 mm2). Conversely, densities of B. chrysoura were significantly higher in T. testudinum with over 70% collected from this habitat. Mean densities ranged from 0.12 m-* to 1.01 m-* in H. wrightii and 0.04 m-* to 2.61 mm2 in T testudinum. Although mean densities of L. xanthurus and M. undulatus were markedly higher in lY testudinum, significant differences between habitat types were not detected (F,,,,, = 3.27, p = 0.072 and F1,288 = 1.27, p = 0.261, respectively). Densities of L. xan- thurus and M. undulatus during the primary settle- ment period ranged from 0.00 mm2 to 9.80 m-* and 0.00 m-* to 23.80 m-*, respectively. Significant (p
Postsettlement Sciaenids in Seagrass Meadows 323
< 0.05) habitat X date interactions were observed for all five species, indicating that the magnitude of differences between habitats was not consistent over time.
Densities of all five species differed significantly among sites (Table 4). At paired sites, densities of B. chrysoura ( F2,,,, = 98.90, p < O.OOl), C. nebulosus F2,266 = 42.49, p < O.OOl), L. xanthurus (F2,288 = 35.09, p < O.OOl), M. undulatus (F,,,,, = 19.88, p < O.OOl), and S. ocellatus (F2,288 = 45.16, p < 0.001) were significantly different among the three bay sites: ABl, RBl, and CBl. Postsettlers of S. ocellatus, B. chrysoura, L. xanthurus, and C. nebulosus were sig- nificantly more abundant at ABl than RBl or CBl (Tukey’s HSD, p < 0.05). Densities of M. undulatus were significantly lower in CBl than ABl or RBl (Tukey’s HSD, p < 0.05). Again, significant first- order interactions with date (site X date) were
l?_- ___I- --__:__ _..---_Ll._- LI^_L Al-^ -_l_ pr-esenr I01 eacr1 spc’c1cs, suggcsLIIlg LIldL LIIC lCI&
tive magnitude of postsettlement densities among sites was not uniform over sampling dates. Varia- tion in settlement density among the six H. wrightii sites was also examined separately and, similar to results described above, significant (p < 0.01) site effects were observed for all five species. In gen- eml. &nsiti_e!: ycye higheg in_ Aransas l&q partic- _- --, _ - -__-__- ularly sites ABl and AB2.
The mean size of postsettlement sciaenids found in H. wrightii and T. testudinum also varied. Mean lengths of two species, M. undulatus and S. ocellatus, differed significantly (p < 0.001) between habitats (Table 5). Postsettlement M. undulatus were mark- edly smaller in H. wrightii (12.6 mm) than T. tes- tudinum (14.1 mm). A similar trend was observed for S. ocellatus; however, the size difference between habitats was larger. Mean lengths of S. ocellatus were 9.5 mm in H. wrightii and 16.1 mm in T tes- tudinum. No habitat-specific differences in size were observed for B. chrysoura, C. nebulosus, or L. xanthurus (p > 0.05).
Discussion Ichthyofauna utilizing seagrass meadows in the
Aransas Estuary were typical of assemblages re- ported in the northern Gulf of Mexico (Stoner 1983) and southeastern Florida (Brown-Peterson et al. 1993). Of the dominant taxa, syngnathids and gobiids can be considered permanent mem- bers of the assemblage since these taxa utilize sea- grass meadows over protracted periods, often pres- ent from larval to adult stages. In contrast, sciaen- ids and other taxa (e.g., sparids, haemulids) were only seasonal residents and annual fluctuations in densities may result from recruitment, migration, and natural mortality. These taxa appear to inhabit shallow seagrass meadows for short periods of time (weeks to months), primarily as larvae and early

Site
ABl ABl AB2 AB3 CB 1 CB 1 RB1 RB1 RB2 All sites % Frequency na
324 J. Ft. Rooker et al.
TABLE 4. Mean and maximum densities (individuals mm*) of sciaenids (540 mm SL) from different sampling sites in the Aransas Estuary during the primary recruitment episode (defined as trips with >50% of samples with target species). Percent frequency of occurrence estimates (based on presence or absence of individuals in sled tows) are shown. Values in parentheses are standard errors.
Bairdi& chrpmra Cynos&l nebuimus Leiostomus xanthuw Mzcropopnias undulattu saaaops ocelbtw
Seagrass MeaIl MZIX MGUI MU Mean MtlX MeaIl MZtX MeaIl Ma
Halodule 1.01 (0.24) 3.46 0.21 (0.05) 0.47 1.31 (0.34) 5.33 0.38 (0.22) 2.07 1.79 (0.20) 2.53 Thalessia 2.61 (0.55) 7.67 0.07 (0.03) 0.27 4.00 (1.63) 23.80 2.62 (1.27) 9.80 0.73 (0.20) 2.07 Haloduk 1.11 (0.36) 4.53 0.31 (0.08) 0.73 0.23 (0.09) 1.47 0.26 (0.08) 0.80 1.66 (0.23) 2.87 Halodule 0.01 (0.01) 0.20 0.11 (0.04) 0.40 0.31 (0.12) 1.80 0.07 (0.04) 0.33 0.90 (0.35) 2.87 Haladuk 0.12 (0.02) 0.33 0.07 (0.03) 0.27 0.22 (0.09) 1.33 0.03 (0.01) 0.07 0.34 (0.07) 0.67 Thalassia 0.04 (0.02) 0.20 0.01 (0.01) 0.01 0.29 (0.15) 2.40 0.00 (0.00) 0.00 0.04 (0.04) 0.33 Haladub 0.06 (0.02) 0.33 0.04 (0.02) 0.20 0.20 (0.08) 1.33 0.33 (0.10) 0.80 1.11 (0.64) 4.73 Thakzssia 0.11 (0.03) 0.33 0.04 (0.02) 0.13 0.06 (0.02) 0.27 0.31 (0.15) 1.47 0.33 (0.09) 0.93 Halodule 0.36 (0.18) 2.46 0.16 (0.04) 0.33 0.39 (0.14) 2.20 0.25 (0.09) 0.80 0.71 (0.12) 1.20
0.62 (0.11) 0.11 (0.04) 0.78 (0.21) 0.47 (0.16) 0.84 (0.09) 64.3 59.3 67.3 66.7 82.7
135 81 135 81 81
a Represents number of sled tows (27 per trip).
juveniles. In this study, sciaenids >40 mm were caught in limited numbers, suggesting that these individuals may migrate to different habitats. How- ever, Rooker (1997) showed that larger sciaenids (>40 mm) are more capable of avoiding the epi- benthic sled and thus low collection numbers may be attributed to both sampling bias and migration activity.
Temporal variation in settlement of sciaenids to seagrass meadows was pronounced and habitat use was staggered over time. Each of the five sciaenids inhabiting seagrass meadows showed distinct settle- ment periods with little temporal overlap among species. Similarly, Deegan and Thompson (1985) found that in Louisiana estuaries peak occurrences of sciaenids were segregated temporally with lim- ited overlap. Moreover, Chao and Musick (1977) suggested that the coexistence of young sciaenids in a Virginia estuary was made possible in part be- cause recruits entered nursery habitats at different times of the year. If resources within seagrass mead-
ows are in limited supply (e.g., food, space), it is possible that newly settled sciaenids are exposed to reduced interspecific competition through differ- ences in the timing of settlement.
Variability in size-at-settlement was observed among sciaenids and appears related to the loca- tion of adult spawning grounds (i.e., distance from estuary). Bairdiella chrysoura and C. nebulosus spawn primarily in estuarine habitats (e.g., bays, chan- nels), often in close proximity to nursery areas (Johnson 1978; Overstreet 1983a; Brown-Peterson et al. 1988). Settlers from these taxa arrive in peak numbers at relatively small sizes (4-8 mm). Con- versely, M. undulatus and L. xunthurus generally spawn offshore, several kilometers from tidal pass- es (Johnson 1978; Warlen and Chester 1985; Cow- an and Shaw 1988). Individuals were markedly larger (lo-14 mm) than settlers from inshore spawners, supporting the assumption that the source population was far removed from the tidal pass. The spawning strategy of S. ocellatus differs
TABLE 5. ANOVA comparisons of mean lengths of sciaenids (540 mm SL) between different seagrass habitats in the Aransas Estuary. Estimates based on length estimations from the three sites containing monotypic stands of both Halodule wrightii and Thakzssia testu- dinum (ABl, RBl, CBl).
Species Habitat Mean Length
” (mm) F Ratio” 0
Bairdiella chrysoura
cynosnon nebulosus
Lziostomus xanthurus
Micropogonias undulatus
SGiaenops ocellatus
Halodule Thalassia H&dub Thalassia Halodule Thalassia Halodule Thalassia Halodule Thakzssia
279 15.3 (0.53) 1.543 0.215”s 537 14.5 (0.38)
71 17.7 (1.39) 0.028 0.866”’ 43 18.1 (1.79)
528 15.3 (0.30) 0.304 0.581”’ 525 15.1 (0.30) 106 12.6 (0.23) 27.971 <O.OOl 376 14.1 (0.13) 441 9.5 (0.34) 49.855 <O.OOl 163 16.1 (0.56)
a ANOVA based on means weighted by the reciprocal of the sample variances of the group means; ns nonsignificant comparisons (p > 0.05).

Postsettlement Sciaenids in Seagrass Meadows 325
from both inshore and offshore spawners. Sciaenops ocelkztus spawning locations range from coastal (i.e., near tidal passes) to offshore waters (Johnson 1978; Overstreet 1983b; Comyns et al. 1991). The pattern of size-at-settlement observed for S. ocella- tus was similar to inshore spawners with peak set- tlement densities occurring at 6-8 mm. Recent work using hydrophones to monitor sound pro- duction associated with courtship and spawning of S. ocellatus has shown that activity is particularly high in the confines of the Aransas Pass (S. A. Holt unpublished data). Thus, it appears that some spawning sites for S. ocellatus are in coastal waters, probably in or near the Aransas Pass.
Although conspicuous differences in size-at-set- tlement were reported in this study, our estimates were based on size-at-capture data. As a result, the frequency of collections may have biased size-at-set- tlement estimates to some degree. For example, with biweekly collections there is a limit to the amount of postsettlement growth that could have occurred in seagrass meadows. Still, estimates of size-at-settlement are going to be overestimated to some degree depending on the amount of time individuals spend in the seagrass meadows prior to capture. Nevertheless, length-frequency profiles of each sciaenid examined in this study are based on a series of cohorts, which enter seagrass beds over a number of months. Since estimates of size-at-set- tlement are averaged across many cohorts for each taxon, bias associated with length-frequency pro- files should be minimized.
Densities of newly settled sciaenids were influ- enced by seagrass spedes. Some sciaenids were sig- nificantly more abundant in one seagrass species or the other, suggesting that settlers may discrimi- nate between habitats,, either during or after the initial settlement event. These data support studies which have shown that patterns of distribution for larvae and early juveniles are seagrass-specific (To- lan et al. 1997; Stoner 1.983; Middleton et al. 1984), but appear to challenge the “settle-and-stay” hy- pothesis (Bell and Westoby 1986; Bell et al. 1987). According to this hypothesis, larval recruits do not discriminate between meadows of different com- plexity when they settle, and redistribution only oc- curs within the original meadow since individuals that re-enter the plankton or migrate across un- vegetated substrata will be more vulnerable to predators. It should b’e noted, however, that pro- ponents of the “settle-and-stay” hypothesis recog- nize that it may not hold for species capable of safely migrating to different habitats. In this study, paired sites were composed of adjoining H. wrightii and 7: testudinum meadows. As a result, postsettle- ment migration activity may have been more likely since individuals were not required to cross areas
of bare substrate to reach different seagrass spe- cies.
Patterns in length-frequency data support the view that postsettlement migration occurs for cer- tain taxa. Inshore-spawned B. chrysoura and C. ne- buulosus were common in seagrass meadows from larval through early juvenile stages, suggesting that these taxa settle and stay in seagrass meadows for extended periods. In contrast, individuals larger than 20 mm from offshore-spawned M. undulutus and L. xanthurus were rarely observed and appar- ently migrate to different habitats shortly after ar- rival. Since migratory behaviors were only conspic- uous for species utilizing seagrass meadows in the late fall and winter, it is possible that activities are related to changes in environmental conditions (e.g., temperature, food supply).
Postsettlement densities of sciaenids varied be- tween seagrass habitats in close proximity (~50 m) and patterns of this nature are often attributed to either food supply and/or predation pressure. Since recent studies have shown that growth and foraging conditions for certain sciaenids in the Aransas Estuary are similar between H. wrightii and ?: testudinum (Rooker et al. 1997; Rooker and Holt 1997; Soto 1997), it seems plausible to assume that patterns of abundance were directly or indirectly related to predation. Predator efficiency is often correlated with macrophyte complexity (e.g., shoot or blade density, biomass; Coen et al. 1981; Stoner 1982; Stoner and Lewis 1985; Ryer 1988; Heck and Crowder 1991). As complexity increases, the visi- bility of prey (encounter rate) declines and pred- ator movements are restricted (Stoner 1980; Savi- no and Stein 1989), resulting in lower predator efficiency. Although H. wrightii and X testudinum differ in complexity (i.e., shoot density) and mor- phology (blade width and height), Rooker et al. (1998) demonstrated that the vulnerability of post- settlement red drum to predatory fish did not dif- fer between the seagrass species. Consequently, other factors related to predation may be respon- sible for observed patterns. For example, habitat complexity (e.g., physical structure of seagrass can- opy) and predator fields vary over spatial and tem- poral scales and may influence settlement patterns and postsettlement survival.
On larger scales, significant correlations be- tween density of settlers and habitat complexity are rare (Stoner 1983; Bell and Westoby 1986; Sogard et al. 1987) and recruitment variability is often at- tributed to hydrodynamic processes (Miller et al. 1984; Boehlert and Mundy 1988; Shaw et al. 1988). Recent studies have elucidated the importance of hydrodynamic processes in estuarine systems (i.e., tidal flux) and suggested that such processes are responsible for spatial variability in larval supply

326 J. R. Rooker et al
and settlement (Boehlert and Mundy 1988; Pietra- fesa and Janowitz 1988; Jenkins et al. 1996). Tidal flow rates in the three channels leading from the Aransas Pass (see Fig. 1) were measured in 1995 (R. Solis, Texas Water Development Board person- al communication), and it appears that densities of sciaenids in the Aransas Estuary were positively as- sociated with tidal flow rates. On average, flow rates in Lydia Ann Channel (Aransas Bay) were 4- 6 times higher than Aransas Channel (Redfish Bay) and post-settlement density was 3 times high- er in Aransas Bay. There tvas no correiation be- tween density and flow rate in Corpus Christi Bay; however, our only sampling site was relatively far removed and indirectly connected to the water conduit from the Arzmsas Pass. Although variation in tidal flow rates is probably a key component in the estuarine-ocean exchange mechanism, a vari- ety of nont;dai factors, both_ biological (P cr hP- y-.6’, -- havior of fish larvae) and physical (e.g., meteoro- logical forcing meclhanisms) are also important and likely to influence large-scale patterns of set- tlement within the Aransas Estuary.
Ackmtowledgments We thank Kathy Binney, Patti Pickering, and Cameron Pratt
for providing assistance in the laboratory and field. Lee Fuiman, Sharon Herzka, Chris Onuf, and two anonymous reviewers pro- vided valuable comments on the manuscript. This study was funded in part by grants from the Texas Higher Education Co- ordinating Board Advanced Research Program (grant no. 003658-392), Texas A&M Sea Grant Program (grant no. NBR: RIF65), Sid W. Richardson Foundation, and fellowships to J. R. R. (E. J. Lund, Julian C. Barton). This is contribution no. 1026 of The University of Texas at Austin, Marine Science Institute.
Literature Cited BELL, J. D. AND D. A. POLLO. 1989. Ecology of fish assemblages
and fisheries associated with seagrasses, p. 569-609. In A. W. D. Larkum, A. J. McComb, and S. A. Shepard (eds.), Biology of Seagrasses. Elsevier, New York.
BELL, J. D. AND M. WESTOBY. 1986. Variation in seagrass height and density over a wide spatial scale: Effects on common fish and decapods. Journal of Expektental Marine Biology and Ecology 104:275-295.
BELL, J. D., M. WESTOBY, AND A. S. STEFFE. 1987. Fish larvae settling into seagrass: Do they discriminate between beds of different leaf density? JmAmal ofExperimental Man’ne Biology and Ecology 111:133-144.
BOEHLERT, G. W. AND B. C. MUNDY. 1988. Roles of behavior and physical factors in larval and juvenile fish recruitment to es- tuarine nursery areas. A*merican Fisheries Society Symposium 3:51- 67.
BROWN-PETERSON, N. J., M. S. PETERSON, D. A. R~ENE, AND R. W. EAMES. 1993. Fish assemblages in natural versus wellestab lished recolonized seagrass meadows. Estuaries 16177-189.
BROWN-PETERSON, N., P. THOMAS, AND C. R. ARNOLD. 1988. Re- productive biology of the spotted seatrout, Cynoscion nebulosus, in south Texas. Fishery Bulletin, United States 86:373-388.
CARR, M. H. AND M. A. HIXON. 1995. Predation effects on early post-settlement survivorship of coral-reef fishes. MarineEcology Progress Series 124:31-42.
CHAO, L. N. AND J. A. MUSI~CK. 1977. Life history, feeding habits,
and functional morphology of juvenile sciaenid fishes in the New York River Estuary, Virginia. Fishery Bulletin, United States 4~657-702.
COEN, L. D., K. L. HECK, JR., AND L. G. ABELE. 1981. Experiments on competition and predation among shrimps of seagrass meadows. Ecology 62:1484-1493.
COMYNS, B. H., J. LYCZKOWSKI-SHULTZ, D. L. NIEL.AND, AND C. A. WILSON. 1991. Reproduction of red drum, Sciaenops ocellatus, in the north central Gulf of Mexico: Seasonal+ and spawner biomass. National Oceanic and Atmospheric Administration Technical Report NMFS 95:17-26.
COWAN, J. H. AND R. F. SHAW. 1988. The distribution, abun- dance, and transport of larval sciaenids collected during the winter and early spring from the continental shelf waters off west Louisiana. Fishery Bulletin, United States 86129-142.
DAY, J. W., JR., C. A. S. HALL, W. M. KEMP, AND A. Y~Ez-ARAN- CIBLA. 1989. Estuarine Ecology. John Wiley and Sons, Inc. New York.
DEEGAN, L. A. AND B. A. THOMPSON. 1985. The ecology of fish communities in the Mississippi River Del&c Plain, p. 35-56. In A. Y&nez-Arancibia (ed.), Fish Community Ecology in Es- tuaries and Coastal Lagoons: Toward an Ecosystem Integra- tinn Erlitnrial Tlnivercitaria nR IR) TTNAM Pwcc M&C-Q. _ __--. -- -____- _ - ---._-I_ -__, --- \__, -_._-._ ___“I)
ECKMAN, J. E. 1983. Hydrodynamic processes affecting benthic recruitment. Limnology and Oceanography 28:241-257.
HECK, K. L., JR. AND L. V. CROWDER. 1991. Habitat structure and predator-prey interactions in vegetated aquatic systems, p. 281-289. In S. S. Bell, E. D. McCoy, and H. R. Mushinsky (eds.), Habitat Complexity: The Physical Arrangement of Ob-
jects in Space. Chapman and Hall, New York. HECK, K. L., JR. AND R. J. ORTH. 1980. Seagrass habitats: The
_^1_. ,.c L-I.:&-& ,.---1,...:&. ^^--^L.:-- _-A -__A_&:_- . ~U~C;J UI ll*~llal ~ulllplc~lly, CVIII~~UUU~~ d1~ ylcuarlull ii?
structuring associated fish and motile macroinvertebrate as- semblages, p. 449-464. In V. S. Kennedy (ed.), Estuarine Per- spectives. Academic Press, New York.
HOLT, S. A., C. L. KITTING, AND C. R. ARNOLD. 1983. Distribution of young red drums among different sea-grass meadows. Transactions of the American Fisheries Society 112:267-271.
JENKINS, G. P., M. J. WHEATLEY, AND A. G. B. POORE. 1996. Spatial variation in recruitment, growth, and feeding of postsettle- __. ment Kmg George whiting, Biiaginodes punctata, associated with seagrass beds of Port Phillip Bay, Australia. Canadian Jour- nal of Fisheries and Aquatic Sciences 531350-359.
JONES, G. P. 1991. Postrecruitment processes in the ecology of coral reef fish populations: A multifactorial perspective, p. 294-328. In P. F. Sale (ed.), The Ecology of Fishes on Coral Reefs. Academic Press, San Diego, California.
JOHNSON, G. D. 1978. Development of fishes of the mid-Atlantic Bight: An atlas of egg, larval and juvenile stages. Volume IV. Carangidae through Ephippidae. United States Department of the Interior, Biological Services Program. 9.
JMP. 1989. JMP User’s Guide. SAS Institute Inc., Gary, North Carolina.
LEIS, J. M. 1991. The pelagic stage of reef fishes: The larval biology of coral reef fishes, p. 183-230. In P. F. Sale (ed.), The Ecology of Fishes on Coral Reefs. Academic Press, San Diego, California.
MCMICHAEL, R. H., JR. AND K. M. PETERS. 1989. Early life history of spotted seatrout, Cynoscion nebulosus (Pisces: Sciaenidae), in Tampa Bay, Florida. Estuaries 12:98-110.
MIDDLETON, M. J., J. D. BELL, J. J. BURCHMORE, D. A. POLLARD, AND B. C. PEASE. 1984. Structural differences in the fish com- munities of Zostera capricorni and Posidonia australis seagrass meadows in Botany Bay, New South Wales. Aquatic Botany 18: 89-109.
MILLER, J. M., J. P. REED, AND L. J. PIETRAFESA. 1984. Patterns, mechanisms and approaches to the study of migration of es- tuarinedependent fish larvae and juveniles, p. 209-255. In J. D. McCleave, G. P. Arnold, J. J. Dodson, and W. H. Neil1

Postsettlement Sciaenids in Seagrass Meadows 327
(eds.), Mechanisms of Migration in Fishes. Plenum, New York.
tween fish predators and their prey: Effects of plant density. Animal Behavior 32311-321.
OLNEY, J. E. AND G. W. BOEHLERT. 1988. Nearshore ichthyo- plankton associated with seagrass beds in the lower Chesa- peake Bay. Matine Ecology Progress Series 45:33-43.
ORTH, R. J. 1992. A perspective on plant-animal interactions in seagrasses: Physical and biological determinants influencing plant and animal abundance, p. 147-164.1n D. M. John, S. J. Hawkins, and J. H. Price (eds.), Plant-Animal Interactions in the Marine Benthos. Claredon Press, Oxford, England.
OVERSTREET, R. M. 1983a. Aspects of the biology of the spotted seatrout, Cynostion nebulosus, in Mississippi. Gulf Research Re- ports, Supplement l:l-43.
SHAW, R. F., B. D. ROGERS, J. H. COWAN, JR., AND W. H. HERKE. 1988. Ocean-estuarine coupling of ichthyoplankton and nek- ton in the northern Gulf of Mexico. Amtican Fisheries Society Symposium 3177-89.
SOGARD, S. M., G. V. N. POWELL, AND J. G. HOLMQUIST. 1987. Epibenthic fish communities on Florida Bay banks: Relations with physical parameters and seagrass cover. Matine Ecology Progress Series 40:25-39.
OVERSTREET, R. M. 1983b. Aspects of the biology of the red drum, Sciaeno~s ocellatus, in Mississippi. Gulf&search Reports, Supplement 1:45-68.
SOTO, M. A. 1997. Food habits and resource partitioning of lar- val and juvenile red drum (Sciaenops ocellatus) and Atlantic croaker (Micrqogonias undulatus). M.S. Thesis, Texas A&M University-Kingsville, Kingsville, Texas.
STONER, A. W. 1980. The role of seagrass biomass in the orga- nization of benthic macrofaunal assemblages. Bulletin of Ma- rine Science 30:537-551.
PETERS, K. M. AND R. H. MCMICHAEL, JR. 1987. Early life history of the red drum, Sciaaops ocellatus (Pisces: Sciaenidae), in Tampa Bay, Florida. Estuaries 10:92-107.
PIETRAFESA, L. J. AND G. S. JANOWITZ. 1988. Physical oceano- graphic processes affecting larval transport around and through North Carolina inlets. American Fishties Society Sym- posium 3:34-50.
STONER, A. W. 1982. The influence of benthic macrophytes on the foraging behavior of pinfish, Lagodon rhomboides (Linnae- us). Journal of Experimental Marine Biology and Ecology 58:271- 284.
ROOKER, J. R. 1997. Early life history of red drum (Sciaenops ocellatus) in subtropical seagrass meadows: Patterns of condi- tion, growth, and mortality. Ph.D. Dissertation, The University of Texas at Austin, Austin, Texas.
ROOKER, J. R., G. J. HOLT, AND S. A. HOLT. 1997. Condition of larval and juvenile red drum (Sciaenops ocellatus) from estua- rine nursery habitats. Marine Biology 127:387-394.
ROOKER, J. R., G. J. HOLT, AND S. A. HOLT. 1998. Vulnerability of newly settled red drum (Stiaenops ocellatus) to predatory fish: Is early life survival enhanced by seagrass meadows? Ma- rine Biology 131:145-151.
STONER, A. W. 1983. Distribution of fishes in seagrass meadows: Role of macrophyte biomass and species composition. Fishery Bulletin, United States 81:837-846.
STONER, A. W. AND F. G. LEWIS, III. 1985. The influence of quan- titative and qualitative aspects of habitat complexity in tropi- cal sea-grass meadows. Journal of Expekmental Matine Biology and Ecology 94:19-40.
TOLAN, J. M., S. A. HOLT, AND C. P. ONUF. 1997. Distribution and community structure of ichthyoplankton in Laguna Ma- dre Seagrass Meadows: Potential impact of seagrass species succession. Estuaries 20:450-464.
ROOKER, J. R. AND S. A. HOLT. 1997. Utilization of subtropical seagrass meadows by newly settled red drum (Stiaenops ocel- latus) : Patterns of distribution and growth. Matine Ecoloa Prog- Tess Series 158:139-149.
RYER, C. H. 1988. Pipefish foraging: Effects of fish size, prey size and altered habitat complexity. Marine Ecology Progress Series 48: 37-45.
UNDERWOOD, A. J. 1981. Techniques of analysis of variance in experimental marine biology and ecology. Oceanography and Marine Biology Annual Review 19:513-605.
WARLEN, S. M. AND A. J. CHESTER. 1985. Age, growth, and dis- tribution of larval spot, Leiostomus xanthurus, off North Caro- lina. Fishery Bulletin, United States 83:587-599.
ZIEMAN, J. C. AND R. G. WETZEL. 1980. Productivity of seagrasses: Methods and rates, p. 87-118. In D. C. Phillips and C. P. McRoy (eds.), Handbook of Seagrass Biology: An Ecosystem Perspective. Garland STPM Press. New York.
SAVINO, J. F. AND R. A. STEIN. 1989. Behavioral interactions be- Received for consideration, March 17, I997
Accepted for publication, October 14, I997