population biology of the salt marsh annual salicornia europaea agg
TRANSCRIPT

Journal of Ecology (1981), 69, 17-3 1
POPULATION BIOLOGY OF THE SALT MARSH ANNUAL SALICORNIA EUROPAEA agg.
R. L. JEFFERIES*, A. J. DAVYt AND T. RUDMIK*
Department of Botany*, University of Toronto, Toronto, Ontario, Canada M5S 1A 1, and School of Biological Sciencest, University ofEastAnglia, Norwich NR4 7TJ
SUMMARY
(1) The biology of two populations of Salicornia europaea agg. from the upper and lower levels of Stiffkey salt marsh on the north Norfolk coast was examined. Although seeds of both populations germinate in spring, upper-marsh seedlings grow little until July. In contrast, individuals from the lower marsh show continuous growth throughout the summer.
(2) These differences in phenology are maintained when reciprocal transplants are made, or when nitrate or ammonium salts, or sea water, are added to permanent field plots. The delayed growth of individuals is interpreted as a genetic response of the popula- tion to the adverse effects of hypersalinity in the upper marsh.
(3) The demography of the two populations was examined. Permanent quadrats were mapped or photographed to follow the fate of seedlings; the reproductive and growth performances of individuals in relation to plant density were measured.
(4) Overall, demographic trends in the two populations were similar. The probability of a seed giving rise to an adult plant was very low; most seeds died without germinating. Mortality of plants appeared to be density-independent, but there was a significant negative density-dependent relationship between number of seeds per plant and the density of Salicornia plants.
(5) By midsummer the seed bank in the sediments was exhausted, so each generation of Salicornia at both sites appears to be distinct.
(6) A model is given which describes the population dynamics of Salicornia in terms of density-dependent regulation of seed number and density-independent mortality. The reasons for the prevailing densities are discussed.
INTRODUCTION
Salicornia europaea agg. is widespread in the coastal salt marshes of north-west Europe. Plants grow as pure stands on open tidal mudflats and are also present in the upper levels of salt marshes, where they are associated with other halophytes (Wiehe 1935; Ball & Brown 1970; Brereton 1971). Although several closely-related species have been recognized within the S. europaea group in the British Isles (Ball & Tutin 1959; Dalby 1962; Ball 1964; Ball & Brown 1970; Scott 1978), the specific limits are obscured both by considerable phenotypic plasticity and by the local differentiation of populations. Much of this local variation is probably the result of inbreeding, as cleistogamy is frequent (Ball 1964).
In this study the behaviour of two populations of Salicornia europaea agg., from the upper marsh and from the open sandy mudflats of the lower marsh, respectively, has been examined in relation to environmental heterogeneity at Stiffkey salt marsh on the north Norfolk coast, England. In cultivation, plants from the two populations exhibit different
0022-0477/81/0300-0017 $02.00 (? 1981 Blackwell Scientific Publications
17

18 Population biology of Salicornia europaea agg.
growth rates, and preliminary evidence has indicated that under field conditions their phenology differs (Jefferies 1977; Jefferies, Davy & Rudmik 1979). The upper marsh, unlike the lower marsh, is not tidal during much of the summer, and, as a result of evaporation, hypersaline conditions develop in most years. Salicornia shows delayed growth until late in the summer on the upper marsh, and it has been suggested that this is a genetically determined response to adverse conditions (Jefferies, Davy & Rudmik 1979). In late summer, when tides are large and the sea floods the upper marsh, conditions ameliorate. Growth rate increases, and is associated with flowering and seed-set.
Stearns (1977) has suggested criteria necessary to interpret differences in life-history traits between populations. They include the following: the growth of plants under controlled conditions in order to isolate the genetic component of variability observed in the field; measurement of environmental conditions associated with differences in reproductive traits; degree of density-dependent or density-independent regulation; year-to-year variability in mortality; and measurement of reproductive effort. As far as practicable these criteria have been adopted for this study. Evidence of genetic differentiation between populations, and of the underlying differences in environmental conditions between the upper and lower marshes, has been reported previously (Jefferies 1977; Jefferies, Davy & Rudmik 1979). This paper describes experimental studies on the phenology of the two populations of Salicornia, and presents demographic data relating to the natural regulation of population size.
MATERIALS AND METHODS
The study sites
The upper marsh and the lower marsh at Stiffkey (National Grid reference TF 964441) are separated by an old sand dune which runs parallel to the coast. Species which are at high frequency on the flat areas of the lower marsh away from the creeks include Aster tripolium*, Halimione portulacoides and Salicornia europaea agg. There is a relatively rich plant community in the upper marsh at sites adjacent to old pools. It consists mainly of Limonium vulgare, Puccinellia maritima, Salicornia europaea agg. and Triglochin maritima. The vegetation of the Norfolk coastal salt marshes has been described by Chapman (1938, 1939, 1960), and Jefferies (1976) has provided a recent general account of the coastline.
Chromosome number
Both tetraploid and diploid populations of Salicornia europaea agg. occur in salt marshes (Dalby 1962; Ball 1964). In order to determine the somatic chromosome number of plants from the two populations at Stiffkey, developing embryos from individuals of the two populations were fixed in acetic alcohol (4: 1 v/v, glacial acetic acid and ethanol) in August 1978. The chromosomes were stained with a solution of propionic orcein, before tissue squashes were prepared and the chromosomes counted.
Water and nutrient experiments on the growth of Salicornia
Lack of growth of individuals in the upper-marsh population during much of the summer may have been either a phenotypic or a genotypic response. If the lack of growth is a phenotypic response, additions of water or nitrogen or both should alleviate
* Nomenclature follows Clapham, Tutin & Warburg (1962).

R. L. JEFFERIES, A. J. DAVY AND T. RUDMIK 19
hypersalinity or a deficiency of nitrogen and provide conditions suitable for growth. Accordingly, in 1976 two latin-square experimental plots, of similar design to those described by Jefferies & Perkins (1977), were set up in the upper marsh to examine the effects of additions of sea water and nitrogen on the growth of Salicornia.
At each site care was taken to select by subjective judgement a plot of sufficient size in which the distribution of Salicornia was uniform, and in which the difference in height of sediment surface across the plot was less than 0.5 cm. At each site the experimental plots were laid out in the form of a latin square, 5 x 5 m. The plot was designed to give five rows and five columns, and each of the twenty-five sub-plots was 50 x 50 cm. Between the rows and columns were strips 50 cm wide which allowed access to the sub-plots. Throughout the season from early May, inorganic salts were applied to designated sub-plots once every 10-14 days, depending on the tidal regime. When necessary, additions of salts were made immediately following the time of spring tides so that salts would not be washed away. Amounts of sodium nitrate or ammonium sulphate added on each occasion were equivalent to those used by Willis (1963), and were 10-05 g and 7.95 g per sub-plot, respectively. Two sub-plots in each row or column received sodium nitrate and two plots received ammonium sulphate; the remaining plot served as a control. Salts were crushed and spread carefully across plots by hand. One set of sub-plots receiving a particular nitrogen treatment was used for measurments of standing crop. Plants in the other set of sub-plots, receiving an identical treatment, were analysed for nutrients. Results of the latter study will be reported elsewhere.
One of the experimental plots in the upper marsh in 1976 received approximately 600 cm3 of sea water three times a week, in addition to the salts supplied. The sea water was added throughout the experimental period (May to August), except during periods in July and August when the sea, at spring tides, flooded the upper marsh. Although the sea did not flood the site between May and early July, it did nearly fill the creeks, which were approximately 10 m from the experimental plots. Thus, sea water could be pumped mechanically from a creek to water the plots. Additions of salts were not made on days when water was added to plots. Turfs were cut from the different sub-plots at intervals of 10-14 days in order to estimate the standing crop of Salicornia. On each sampling occasion, one turf of diameter 10 cm was taken at random from each of five sub-plots receiving a particular treatment. Plants of Salicornia in the turfs were harvested, washed in tap water for 2 min, dried at 80 0C and weighed. The results are expressed as g m-2.
The study of the effect of additions of nutrients on the growth of Salicornia was extended in 1977 to include populations in both the upper and lower marshes. The summer was wet, and the salinity of the interstitial water in the rooting zone of sediments in the upper marsh was only just above that of sea water (Jefferies, Davy & Rudmik 1979), hence plots were not watered with sea water. Experimental plots of similar design to those described above were laid out in the upper and lower marshes. Additions of nitrogen salts were made to the plots once every 10-14 days. In both plots there was a uniform distribution of Salicornia. Measurements of standing crop were made on turfs as described previously.
Reciprocal transplant experiment Reciprocal transplants were made, in order to establish whether the differences in
growth between the populations were maintained when plants were transplanted between the upper and lower marshes. Seedlings of Salicornia at the post-cotyledon stage were transplanted in mid-May 1978. Seedlings were dug up, washed free of sediment as far as

20 Population biology of Salicornia europaea agg.
possible, and transplanted on the same day to the other site; 140 seedlings of each population were transplanted to the alternative site and an additional 140 seedlings of the same population were transplanted within a site, to serve as the control population. The seedlings were planted in groups of 14, consisting of 2 parallel rows of 7 plants. The seedlings were spaced approximately 1 cm apart. Sites for planting the seedlings were chosen subjectively, as typical of the area in which each local population of Salicornia grew. Before planting, all vegetation was cleared from these small plots, which were at least 50 cm apart. Successive harvests were made throughout the season from late May to late August, at intervals of 10 days. On each occasion, fourteen plants were harvested from each of the two control populations and the two transplanted populations; two rows of plants within each population were selected at random. Plants were washed free of sediment, separated into shoot and root, dried at 80 0 C and weighed. It was not possible to obtain the dry weights of roots and shoots of individual plants in the early harvests in May and June, as the plants were too small; consequently plants were bulked and weighed as a group.
Demographic studies
A series of permanent plots was chosen subjectively to include a range of population densities at which populations of Salicornia are commonly found in both the lower marsh and the upper marsh. Five and six permanent plots (5 x 5-cm) were marked out in the lower and upper marsh, respectively, during late April, at the time of germination. For the first three sampling dates the plots were mapped, but subsequently they were photographed. By this technique, recruitment to and loss from the population could be followed by making observations at intervals of 10-21 days from late April to late August 1978. From these results, survivorship curves for the two populations were constructed, and the relationship between mortality and density of Salicornia examined.
In order to investigate the relationship between the density of plants of Salicornia and fecundity, thirty individuals from each of the five plots in the lower marsh and from four of the plots in the upper marsh were collected on 15 October 1978, and the number of seeds produced by each plant counted. A wide range of density of Salicornia occurred on the different plots at each site. The densities of individuals in the different plots were recorded, and the height of one-hundred plants from each plot was measured and the number of side branches recorded.
Parallel experiments (see below) showed there is no persistent seed bank of Salicornia in the sediments. Each generation is discrete, and there is virtually no overlap between generations. Hence it is possible to calculate the probability of a seed producing an adult plant. The number of seedlings in twelve 5 x 5-cm plots at the two sites was counted in May and June 1979, in order to estimate the probability of a seed producing a plant.
Germination studies
In order to determine that there was no persistent seed bank in these populations, turfs were collected on a number of occasions from both areas in the upper and lower marshes between February 1979 and October 1979. Subsequently each was divided into four 10 x 10-cm units. These smaller turfs were placed in a growth cabinet at 20 OC, with a photosynthetically-active photon flux density of 100,uEinstein m-2 s-1 for 16 h of each 24-h cycle; for the remainder of the time the cabinets were kept dark. The number of seedlings of Salicornia which emerged over a period of 6 weeks was counted, and the results expressed on a unit-area basis.
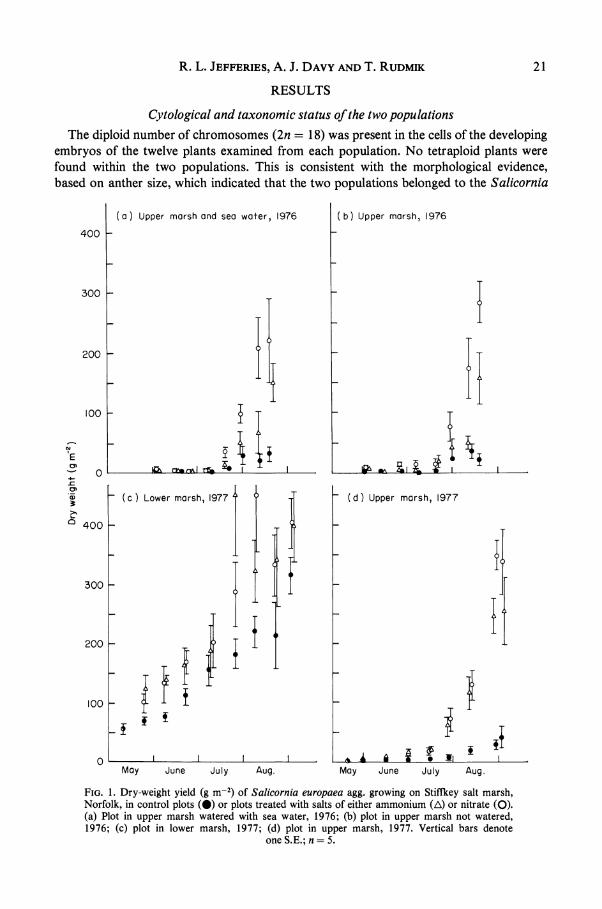
R. L. JEFFERIES, A. J. DAVY AND T. RUDMIK 21
RESULTS
Cytological and taxonomic status of the two populations The diploid number of chromosomes (2n = 18) was present in the cells of the developing
embryos of the twelve plants examined from each population. No tetraploid plants were found within the two populations. This is consistent with the morphological evidence, based on anther size, which indicated that the two populations belonged to the Salicornia
(a) Upper marsh and sea water, 1976 (b) Upper marsh, 1976
400
0 0r
(c) Lower marsh, 1977 (d) Upper marsh, 1977
o400
I ~~~~~~~~~~~Ii 200_
20 I I, II t 2Ig
May June July Aug. May June July Aug.
FIG. 1. Dry-weight yield (g m-2) of Salicornia europaea agg. growing on Stiffkey salt marsh, Norfolk, in control plots (0) or plots treated with salts of either ammonium (A) or nitrate (0). (a) Plot in upper marsh watered with sea water, 1976; (b) plot in upper marsh not watered, 1976; (c) plot in lower marsh, 1977; (d) plot in upper marsh, 1977. Vertical bars denote
one S.E.; n = 5.

22 Population biology of Salicornia europaea agg.
europaea group (Ball 1964; Scott 1978) in which all the micro-species are diploid. The width of the scarious margins to the fertile segments, and the size of the lower fertile segments of the terminal spikes most closely resembled the descriptions of Salicornia ramosissima, J. Woods for the upper-marsh population and Salicornia europaea (L.) (sensu stricto) for the low-marsh population (Ball 1964; Scott 1978).
Experimental studies on the growth of Salicornia populations In Fig. 1 is shown the yield of plants (g m-2) of Salicornia in the upper marsh and
lower marsh, in response to different treatments, during the summers of 1976 and 1977. The growth of Salicornia in the upper marsh under all treatments appears to be strongly determinate. Seedling emergence occurs in late April or early May, and the plants undergo a period of slow vegetative growth until mid-July, when there is an increase in growth rate associated with flowering. The vegetative stage coincides with the period when the sediments are most likely to be hypersaline, before the July spring tides cause the sea to flood the upper marsh and to reduce the hypersalinity. Additions of sea water or nitrogen salts or both to treated plots completely failed to modify the growth patterns of the upper-marsh population in both 1976 and 1977.
In the untreated plots (Fig. 1 (b) and (d)), the yield of Salicornia in the upper marsh was between 30 and 40 g m-2 in 1976 and 1977; most of the dry matter accumulated during late July and August. The yield at the final harvest in the plots treated with nitrogen was appreciably greater than the yield of untreated plots, indicating that growth was limited by a deficiency of nitrogen in the latter plots. The enhanced growth in the presence of added nitrogen was not manifest, however, before flowering began. Additions of sea water alone (Fig. 1(a)) not only failed to modify the growth pattern but resulted in a final yield similar to that of untreated plots (Fig. 1 (b)).
( (a) Lower _ lower (b) Lower _ upper
40
~ 0
(c) Upper _ lower (d) Upper _ upper
40
20
May June July Aug May June July Aug.
FIG. 2. Growth of control and reciprocal transplants of two populations of Salicornia europaea agg., growing in the upper and lower levels of Stiffkey salt marsh, respectively, between May and August 1978. (a) Lower-marsh population transplanted in lower marsh (control); (b) Lower-marsh population transplanted in upper marsh; (c) upper-marsh population transplanted
in lower marsh; (d) upper-marsh population transplanted in upper marsh (control).

R. L. JEFFERIES, A. J. DAVY AND T. RUDMIK 23
The growth pattern of Salicornia in the lower marsh was very different from that in the upper marsh (Fig. 1 (c)). In both the treated and untreated plots, growth of individuals was continuous from May to September, and there was no increase in growth rate late in summer. Although additions of nitrogen resulted in higher yields of Salicornia, the percentage increases were relatively small, about 30%, compared with more than 500% for the upper-marsh population of Salicornia. Except at low neap tides, sea water covered the low marsh at high tide throughout the experimental period.
The growth patterns characteristic of each population were maintained when plants were reciprocally transplanted (Fig. 2). Salicornia plants from the low marsh showed continuous growth in both the upper and lower marshes, although the final mean weight of plants was lower in the population transplanted to the upper marsh. The phase of slow vegetative growth was evident whether plants of Salicornia from the upper marsh were transplanted in the upper marsh or the lower marsh. At both sites the period of slow growth was followed by one in which the growth rate increased.
These results, which were obtained over three growing seasons from 1976 to 1978, indicate that the phenology of Salicornia plants from the upper marsh is different from that of plants of Salicornia which grow in the lower marsh.
Survivorship Survivorship curves for the two sites are shown in Fig. 3. These curves are based on
counts of the number of seedlings, and take no account of death before or at germination. In addition, because initial densities of seedlings were high, death of young emergent seedlings may have been masked to a small extent by additional recruitment. The survivorship curves of the populations are different at the two sites. The population of Salicornia in the lower marsh had a similar initial density to that in the upper marsh (50 plants per 25 cm2), but the numbers declined steadily throughout the season. In the upper marsh, few seedlings died before the end of June, but thereafter there was a decline in the density of Salicornia plants from 50 to 22 plants per 25 cm2. At this stage in the season the slopes of the survivorship curves of both populations approached one another.
60 -
50 -
-T 40-
30 E
C\j ~~~~~~~Lower-marsh
u-) 20-
10 11 11 l April May June July August
FIG. 3. Survivorship curves in 1978 for two populations of Salicornia europaea agg. from Stiffkey salt marsh, Norfolk. Vertical bars denote one S.E.; n = 5 (lower marsh) or 6 (upper
marsh).

24 Population biology of Salicornia europaea agg.
For plants in the individual plots in the upper marsh, the probability of dying ranged between 0.37 and 0.71; corresponding values for plants for the lower marsh were 0.57 and 0 91. Some causes of death were identified. Seedlings in the lower marsh were washed free of the sediments by wave action; seedlings frequently became enmeshed in algal mats of Enteromorpha prolifera (O. F. Mull.) J. Ag., which then floated as the water rose with the tide and pulled seedlings from the sediments. A number of plants died in both the upper and lower marshes during the period of inflorescence development probably from nitrogen deficiency or desiccation or both.
The survivorship curves mirror (i.e. are the inverse of) the growth curves (Fig. 2). In the upper-marsh population, before late summer few plants died on the plots, and the mean weight of plants altered little from early May to late June. Loss of individuals occurred in late summer, when growth and development of Salicornia resulted in a marked increase in the mean weight of individuals (Fig. 2). On plots in the lower marsh, plants died continually from late April to late August. This loss was correlated with a steady increase in the mean weight of the surviving plants throughout the period of study.
Density
In the permanent plots in both the upper marsh and the lower marsh the proportion of plants which died before flowering appeared to depend upon conditions prevailing in each plot, and not upon the density of Salicornia seedlings per se (Fig. 4). Within these plots, where there was a fifty-fold range in plant density, there was no evidence of density-dependent mortality of plants. Birth rate, however, was strongly density- dependent. The number of seeds produced per plant depended upon plant density in both the upper and lower marshes (Fig. 5), although this dependence was more marked in the low-marsh population. The number of seeds per plant fell from 936 to 36 in plots where the density increased from 130 to 8900 plants m-2. Corresponding results for the upper marsh
100 _
_ ~~~~A
80 A 1978
LA 0 0
60 A
tz 00 0 @0
E 40 0
20 A A &A A
20 0^ *^-0 1979
0 20 40 60 80 100 Initial density (plants per 25 cm2
FIG. 4. Relationship between mortality of individuals of Salicornia europaea agg. from seedling emergence to flowering and the density of seedlings (number per 25 cm2) in permanent plots in the upper and lower levels of Stiffkey salt marsh in 1978 and 1979. Symbols: A, lower-marsh population 1978; A, lower-marsh population 1979; 0, upper-marsh population 1978; 0,
upper-marsh population 1979.

R. L. JEFFERIES, A. J. DAVY AND T. RUDMIK 25
1000 (a) Lower - marsh
800
0~
O 600
a)
an
0 400 40 ( b) Upper -marsh
nQ * \30
200 - 20
l0
I I I I I 100 1000 10 000 500 1000 5000 10 000
Density of plants (m2) [log scale]
FIG. 5. Semi-logarithmic plot of the relationship between number of seeds per plant and the density of the two populations of Salicornia europaea agg. from the lower (a) and upper (b)
levels of Stiffkey salt marsh.
showed a fall from 29 to 12 seeds per plant as the density increased from 800 plants to 1300 plants m-2.
The different morphology of plants from the two populations accounts for these differences. In Fig. 6 are shown the relationships between plant density, plant height and degree of branching of plants. Not only are individual plants from the lower-marsh
24 - (b)
A 0 (0)
200 - 20
v Lower-morsh
160 - 16 Lea
E _ o 2 \Lower-morsh E 0
c 120 - 12 a) -m orsh aUppe-mars
r - mo rhUpper - marsh A
0 I I I 100 1000 10 000 100 1000 10 000
Density of plonts (m-2) [log scale]
FIG. 6. Relationship between (a) plant height (mm), (b) number of primary and secondary branches, and the logarithm of the density of plants of two populations of Salicornia europaea agg. from the upper and lower levels of Stiffkey salt marsh. 0, Mean plant height, upper marsh; 0, mean plant height, lower marsh; A, mean number of primary branches on plants from lower marsh; V, mean number of primary branches on plants from upper marsh; A, mean number of secondary branches on plants from lower marsh. (Plants from the upper marsh did not produce
secondary branches.)

26 Population biology of Salicornia europaea agg.
population taller, but primary and secondary branches are more numerous at all densities, compared with plants from the upper marsh. Many plants from the upper marsh popula- tion have no branches, particularly at high densities. The steep fall in seed production with increasing plant density shown by plants in the low marsh is caused by a decrease in the number of primary and secondary branches per plant. At high density secondary branches are not produced, and the mean number of primary branches is only 4 per plant compared with 22 at low density.
Bank of viable seeds, and germination
Almost no seedlings of either population of Salicornia appeared when turfs collected between May and October were incubated, either in a laboratory or in a growth cabinet (Table 1). At other times, incubation of turfs resulted in the appearance of numerous seedlings. After the first flush of seedlings very little further germination took place. The
TABLE 1. Mean viable seed bank (m-2), ?S.E., of Salicornia europaea agg. in sediments from the upper and lower levels of Stiffkey salt marsh; values in
parentheses are the number of samples collected
Date (in 1979) Upper marsh Lower marsh
Before seedling emergence in field: 28 February 19 170 + 1007 (30) 2720 + 345 (30) 12 March 23 500 ? 1470 (15) 2730 + 737 (15)
After seedling emergence in field: 26 June 0 (30) 0 (30)
5 September 0 (20) 5 + 5* (20) 16 October 0 (20) 5 + 5* (20) * This represents the germination of a single seed in one sample.
absence of seedlings in turfs collected during late summer and early autumn, and incubated under suitable conditions, indicated that the seed bank was virtually exhausted. The bank was replenished in late autumn when seed was shed from the parent plants. Additional evidence of the absence of a seed bank in late summer came from germination studies and from an examination of sediment. Seed collected in the autumn of 1977 germinated readily in Petri dishes in July 1979, which strongly suggested that if seed had been present in sediments, germination would have occurred under the experimental conditions used. In three turfs in which seedlings were absent, the upper 1 cm was cut from the turf and soaked in sea water. Subsequently, this layer was gently broken into small pieces, and a binocular microscope used to examine the sediment for seeds of Salicornia. None was found.
A relatively small number of seedlings germinated in turfs collected from the lower marsh in February and March 1979 because during the winter an area of the lower marsh was eroded by about 3 cm, and seeds present in the upper layers of sediment were removed.
DISCUSSION
In north-west Europe, populations of the micro-species Salicornia ramossisima are widespread in the upper parts of salt marshes (Ball & Tutin 1959). They grow on the strand line, in the 'general' salt marsh and along the sides of channels. In contrast,

R. L. JEFFERIES, A. J. DAVY AND T. RUDMIK 27
populations of Salicornia europaea (sensu stricto) are restricted to open or sandy mud. Morphological, cytological and ecological evidence given in this paper indicates that the populations of Salicornia at the sites in the upper and lower parts of the salt marsh at Stiffkey most closely resemble the descriptions of Salicornia ramosissima and Salicornia europaea (sensu stricto) respectively.
Evidence based on growth of plants in a glasshouse (Jefferies 1977), and the results from the experiments described here (Figs 1 & 2), indicate that the two populations show genetic differentiation with respect to growth responses. Two important questions arise from these studies: how are these genetic differences maintained, and what selective advantages do they confer upon the respective populations? As pointed out by Ball (1964), inbreeding probably predominates, and this may account for much of the local variation between populations. Dalby (1962) examined the breeding systems in different populations of Salicornia, including some populations from the north Norfolk coast, and concluded that many of the annual forms were self-pollinated.
Flowers of plants of both populations examined at Stiffkey were strongly cleistogamous. Anthers were rarely exserted, and developing seed was present in early August, buried in tissue. Although some pollen may have been transferred from flower to flower and possibly from plant to plant, the high density of Salicornia plants in both the upper and lower marshes made it unlikely that pollen flow between populations would have resulted in cross-pollination of flowers of individuals in adjacent populations.
Besides pollen flow between populations there is the possibility that seeds move between the upper and lower marshes. A number of mechanisms ensure, however, that most of the seed produced remains in the vicinity of the parent plant. Seeds reach maturity from late-September onwards, and usually fall from the dead or dying parent plant (Ball & Brown 1970). It is not unusual to find in spring, however, that seeds still have not been shed. They may germinate in situ on the parent plant, which, by that time, has fallen over (Dalby 1962; Ball & Brown 1970; Ungar 1978). Most seeds appear to fall from the parent plant in late November or December. The seeds, which bear an indumentum of mucilaginous hairs, many of them hooked, apparently sink in sea water (Petch & Swann 1968). These hairs may assist in anchoring the seed to the sediment. Besides these biological features restricting seed movement, the sea rarely floods the upper marsh during November or December, so that in any event the number of seeds transported over long distances is likely to be small. Experiments are in progress to determine the ability of Salicornia to colonize sediments in different parts of the marsh.
The results indicate that there are at least two biologically distinct populations of Salicornia europaea agg., within which inbreeding predominates, and that each population possesses a series of characteristics that either restricts, or reduces the effects of, gene flow.
The growth pattern of plants in the upper marsh appears to be strongly determinate (Harper & White 1974), in that there is a long period of slow vegetative growth before an increased rate of growth associated with sexual reproduction. Additions of sea water or nitrogen salts or both almost completely failed to modify the growth rates during this period of vegetative growth (Fig. 1). The results of the reciprocal transplant experiment show that the seasonal growth pattern is maintained when plants are transplanted to the lower marsh (Fig. 2). The lag phase is characteristic of organisms living in an environment exhibiting cyclic stability (Orians 1975), where there is selection for organisms which can time their growth and reproduction to coincide with favourable environmental conditions. Harper & White (1974) conclude that this determinate growth is the optimal behaviour in an environment of high predictability. The period of rapid growth of the upper-marsh

28 Population biology of Salicornia europaea agg.
population in late summer approximately coincides with the flooding of the upper marsh and an amelioration of conditions. We do not know the environmental triggers which initiate active growth.
The growth response of the population from the lower marsh is different. The growth of individuals is continuous from May to early September. With the exception of neap tides, all tides cover this site, so that the sediments do not become extremely hypersaline. Much of the continuous growth of plants is associated with the development of primary and secondary branches (Fig. 6), many of which bear flowers.
As discussed by Jefferies, Davy & Rudmik (1979) and Stewart et al. (1979) soluble nitrogen and carbon compounds are involved in osmoregulation in plants growing under saline conditions. It may be that individuals growing in the upper layers of the sediment of the upper marsh, where hypersaline conditions prevail, utilize the limited supplies of soil nitrogen for osmoregulation at the expense of growth. There appears to have been selection for individuals which do not show a growth response to additions of nitrogen; instead there is accumulation as soluble nitrogen in the plant tissues during early summer, when the soil-water is often hypersaline. It is only later in the season, when the sea floods the upper marsh, that there is rapid growth of individuals, and the concentration of soluble nitrogen in the tissues falls (R. L. Jefferies, unpublished data).
As Watkinson & Harper (1978) point out, the population dynamics of a species in different habitats and in similar habitats in different areas have not been well studied. The number of births, deaths, immigrants and emigrants determines the size of a population. In some species of non-weedy annuals, such as Minuartia uniflora (Sharitz & McCormick 1973), there is very high mortality during the seed and seedling establishment phases of the life cycle. At the other extreme, Vulpia fasciculata, a winter annual, shows a peak of mortality during inflorescence development (Watkinson & Harper 1978). Relatively few seeds are produced by each plant of Vulpia, in contrast with Minuartia, but the probability of a seed surviving to maturity is high. Mortality of plants of Vulpia is not density-dependent, although the number of spikelets produced per plant is negatively correlated with the density of plants. Each generation is virtually discrete, as there is no bank of viable seed in the soil.
The probability of an individual of Salicornia europaea agg. surviving different stages of the life cycle from seed maturation to maturity is given in Table 2. The risk of death before maturity is extremely high in both populations. The probability of a seed producing a
TABLE 2. Probabilities of individuals of two populations of Salicornia europaea agg. from the upper and lower levels of Stiffhey salt marsh surviving from seed
maturation to maturity
Period Lower marsh Upper marsh
Seed maturation to germination 0.040 to 0-086* 0.131 to 0.401 Germination to flowering stage 0.081 to 0.044 0-291 to 0.639 Complete life cycle 0.001 to 0.028 0.035 to 0.083
* Range of values.
mature flowering plant in either population is less than 0.081. High mortality occurs between the production of ripe seed and seedling establishment. Although seed production is strongly density-dependent in populations of Salicornia, the number of seeds per plant is similar to that of Minuartia uniflora, whereas Vulpiafasciculata on average produces only 1 7 mature seeds per plant, of which 90% germinate.

R. L. JEFFERIES, A. J. DAVY AND T. RUDMIK 29
All plants respond to an increase in density, either through an increase in mortality or a plastic response in the growth and reproductive capacity of individuals, or both. The density-dependent regulation of number of seeds produced per plant was the only regulation of this kind recorded for the Salicornia populations (Fig. 5). Although mortality was high throughout the life cycles, it appeared to be density-independent (Fig. 4). The causes of death include the effects of drought, hypersaline conditions, wave action and the smothering of seedlings by algal mats. Recently, Ungar, Benner & McGraw (1979) have reported high mortality of seedlings of an inland population of Salicornia europaea agg. during drought.
Williamson (1972) and Watkinson & Harper (1978) have developed a model which shows that an interaction between a density-independent death rate and a negatively density-dependent birth rate will lead to an equilibrium population density. The flux of population size, however, will also be affected by the number of immigrants and emigrants. As mentioned earlier, most Salicornia seeds are dispersed for short distances only, and the rates of immigration and emigration are probably low and equal. Where areas have been cleared of vegetation in the lower and upper marshes, the initial level of recruitment of seedlings is less than five per square metre, which is very small in comparison with the number of seedlings in an undisturbed area. As the two populations of Salicornia had, in 1979 at least, no bank of viable seed, the model of Watkinson & Harper (1978) of non-overlapping generations may be used to predict the size of the equilibrium population.
The average seed output per flowering plant for densities observed in the field can be calculated from the linear relationship apparent in Fig. 5.
Ns/N=-clogloN+k (1)
where NS is the number of seeds produced per square metre, and N is the number of flowering plants per square metre; c and k are both constants of the regression equation. At densities beyond the range observed in the field this relationship may not be valid. It can be shown (cf. Watkinson & Harper 1978) that if the probability, P, of an individual surviving from seed production to maturity is constant from year to year, then N, the equilibrium density, is given by
~k 1 log1o N= -- p (2)
c cP(2
Substituting the values for c and k into Equation 2 gives an equilibrium population density for an area of 1 m2 for any value of P, where P is greater than the minimum value of P necessary to maintain the population. The equilibrium population density becomes increasingly sensitive to a decrease in the probability of a seed producing a mature plant. If it is assumed that the population density in the upper and lower marshes is at equilibrium, then the estimated values of P (calculated from Equation 2) which correspond to the observed densities may be calculated (Table 3). These values of P are similar to the estimates of P derived from observations in the field (Table 2).
Although it is unsatisfactory to state that there is never any overlap between generations and that the populations are close to their equilibrium density on the basis of data for one year only, it can be concluded that the density of the Salicornia populations is largely a function of a density-dependent birth rate and a density-independent death rate.
Environmental factors which account for the density-independent death rate are being investigated, but it is already evident that seedling-emergence is a critical phase for the

30 Population biology of Salicornia europaea agg.
TABLE 3. Demographic data for two populations of Salicornia europaea agg. from the upper and lower levels of Stiffkey salt marsh, 1978
Lower marsh Upper marsh -c k -c k
* Parameters of the regression of number 498 1982 9.07 52.9 of seeds per plant on density of plants
Equilibrium density of population when 9480 525 000 P= lt
Ratio of above density to highest observed 1.09 27-1 in the field
Estimated values of P which correspond 0-001 to 0.050 0.036 to 0.071 to observed densities
Estimates of P derived from observations 0.001 to 0.028 0-035 to 0.083 in the field * Regression equation is: Ns/N = -c log1o N + k, where NS is the number of seeds per
plant, and N is the density of plants (m-2). t Probability of a seed giving rise to a flowering plant.
survival of individuals. Overall, the demographic patterns of the two populations are similar. The density of the upper-marsh populations, in particular, appears to be close to equilibrium. This result, together with results which show that the growth patterns of individuals are determinate, is characteristic of organisms growing in a predictable and constant environment. Although equilibrium density may be attained in the lower-marsh population, conditions at this site are less predictable, as shown by the erosion of sediment during the winter of 1978-79, so the density may frequently not be at equilibrium. The high fecundity of individuals and the low probability of a seed forming an adult plant are characteristic of populations growing in a more unpredictable environment than exists in the upper marsh.
ACKNOWLEDGMENTS
We thank Dr P. W. Ball and Dr A. R. Watkinson for helpful criticism of the manuscript. Part of this work was supported by a grant to R.L.J. from the Natural Sciences and Engineering Research Council of Canada. We thank the Nature Conservancy and the National Trust for permission to conduct experimental studies on Stiffkey salt marsh. Gail Richardson and Mrs N. Nolan helped prepare the manuscript.
REFERENCES
Ball, P. W. (1964). Salicornia. Flora Europaea, Vol. 1 (Ed. by T. G. Tutin, V. H. Heywood, N. A. Burges, D. H. Valentine, S. M. Walters & D. A. Webb.), pp. 101-102. Cambridge University Press, London.
Ball, P. W. & Tutin, T. G. (1959). Notes on annual species of Salicornia. Watsonia, 4, 193-205. Ball, P. W. & Brown, K. G. (1970). A biosystematic and ecological study of Salicornia in the Dee estuary.
Watsonia, 8,27-40. Brereton, A. J. (1971). The structure of the species populations in the initial stages of salt-marsh succession.
Journal of Ecology, 59, 321-338. Chapman, V. J. (1938). Studies in salt marsh ecology I-III. Journal of Ecology, 26, 144-179. Chapman, V. J. (1939). Studies in salt marsh ecology IV-V. Journal of Ecology, 27, 160-201. Chapman, V. J. (1960). Salt Marshes and Salt Deserts of the World. Leonard Hill, London. Clapham, A. R., Tutin, T. G. & Warburg, E. F. (1962). Flora of the British Isles, 2nd edn. Cambridge
University Press, London. Dalby, D. H. (1962). Chromosome number, morphology and breeding behaviour in the British Salicorniae.
Watsonia, 5, 150-161.

R. L. JEFFERIES, A. J. DAVY AND T. RUDMIK 31
Deevey, E. S. (1947). Life tables for natural populations of animals. Quarterly Review of Biology, 22, 283-314.
Harper, J. L. & White, J. (1974). The demography of plants. Annual Reviews of Ecology and Systematics, 5, 419-463.
Jefferies, R. L. (1976). The North Norfolk coast. Nature in Norfolk: a Heritage in Trust (The Norfolk Naturalists' Trust), pp. 130-138. Jarrold & Sons, Norwich.
Jefferies, R. L. (1977). Growth responses of coastal halophytes to inorganic nitrogen. Journal of Ecology, 65, 847-865.
Jefferies, R. L. & Perkins, N. (1977). The effects on the vegetation of the additions of inorganic nutrients to salt marsh soils at Stiffkey, Norfolk. Journal of Ecology, 65, 867-882.
Jefferies, R. L., Davy, A. J. & Rudmik, T. (1979). The growth strategies of coastal halophytes. Ecological Processes in Coastal Environments (Ed. by R. L. Jefferies & A. J. Davy), pp. 243-268. Blackwell Scientific Publications, Oxford.
Orians, G. H. (1975). Diversity, stability and maturity in natural ecosystems. Unifying Concepts in Ecology (Ed. by W. H. van Dobben & R. H. Lowe-McConnell), pp. 139-150. Junk, The Hague.
Petch, C. P. & Swann, E. L. (1968). Flora of Norfolk. Jarrold, Norwich. Scott, A. J. (1978). Reinstatement and revision of Salicorniaceae J. Agardh (Caryophyllales). Botanical
Journal of the Linnean Society, 75, 357-3 74. Sharitz, R. R. & McCormick, J. F. (1973). Population dynamics of two competing annual plant species.
Ecology, 54, 723-740. Stearns, S. C. (1977). The evolution of life history traits: a critique of the theory and a review of the data.
Annual Reviews of Ecology and Systematics, 8, 145-17 1. Stewart, G. R., Larher, F., Ahmad, I. & Lee, J. A. (1979). Nitrogen metabolism and salt-tolerance in higher
plant halophytes. Ecological Processes in Coastal Environments (Ed. by R. L. Jefferies & A. J. Davy), pp. 211-227. Blackwell Scientific Publications, Oxford.
Ungar, I. A. (1978). Halophyte seed germination. Botanical Review, 44, 233-264. Ungar, I. A., Benner, D. K. & McGraw, D. C. (1979). The distribution and growth of Salicornia europaea on
an inland salt pan. Ecology, 60,329-336. Watkinson, A. R. & Harper, J. L. (1978). The demography of the sand dune annual Vulpiafasciculata. 1. The
natural regulation of populations. Journal of Ecology, 66, 15-33. Wiehe, P. 0. (1935). A quantitative study of the influence of tide upon populations of Salicornia europaea.
Journal of Ecology, 23, 323-333. Williamson, M. H. (1972). The Analysis of Biological Populations. Arnold, London. Willis, A. J. (1963). Braunton Burrows: the effects on the vegetation of the additions of mineral nutrients to the
dune soils. Journal of Ecology, 51, 353-374.
(Received 26 November 1979)