polypeptide hormone regulation of gene expression · the journal of biological chemistry vol. 258,...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 258, No. 24, Issue of December 25, pp. 15329-15335,1983 Prmtedm U.S.A.
Polypeptide Hormone Regulation of Gene Expression THYROTROPIN-RELEASING HORMONE RAPIDLY STIMULATES BOTH TRANSCRIPTION OF THE PROLACTIN GENE AND THE PHOSPHORYLATION OF A SPECIFIC NUCLEAR PROTEIN*
(Received for publication, July 5, 1983)
Geoffrey H. Murdochz, Rodrigo Francoz, Ronald M. Evans$, and Michael G. Rosenfeldt From the $Eukaryotic Regulaton, Biology Program (M-OlJ), University of California, San Diego, La Jolla, California 92093 and the §Molecular Biology and Virology Laboratory, The Salk Institute, San Diego, California 92138
The tripeptide hypothalamic releasing factor, thy- rotropin-releasing hormone (TRH), rapidly increases prolactin gene transcription 7- to 12-fold in the GH4 rat pituitary cell line. The maximal rates of transcrip- tion are achieved within minutes and begin to atten- uate within 1 h following addition of TRH. This tran- scriptional response appears to account for the ob- served effects of TRH on the accumulation of prolactin mRNA and the stimulation of prolactin biosynthesis. The rapid transcriptional effects of TRH include a large number of polymerase II-catalyzed transcription units. Addition of TRH to GH4 cell cultures also rapidly induces the phosphorylation of a 23,000-dalton chro- matin-associated basic protein, distinct from known high mobility group or histone proteins and referred to as basic regulated phosphoprotein. The time course of TRH-stimulated prolactin transcription and basic regulated phosphoprotein phosphorylation demon- strates that the signal generated by TRH binding to its plasma membrane receptor rapidly reaches the cell nucleus.
There are two functional classes of regulatory signals in the neuroendocrine system, those which bind to an intracellular receptor and those which initially bind to a receptor located on the plasma membrane. Specific intracellular receptors for steroid and thyroid hormones have been demonstrated to interact directly with chromatin when occupied by an appro- priate ligand (1). The structural similarity between this sys- tem and the well characterized prokaryotic transcriptional regulatory systems, such as the CAMP receptor protein regu- lation of catabolite repression (2, 3), suggested that these hormones probably acted at a transcriptional level. As pre- dicted by this model, steroid and thyroid hormones were proven to regulate protein synthesis in many model systems by directly modulating the transcription of specific genes (4- 10) apparently by interacting with specific DNA recognition signals (11-15) analogous to those present in bacterial ope- rons.
In contrast, there is not a comparable framework to explain the mechanisms by which the interaction of a hormone with a receptor on the surface of the cell can influence the flow of information from gene to protein. The interaction of hor- mones on the surface of the cell can be transduced into a
* This work was supported by grants from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "aduertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
variety of intracellular second messengers which activate spe- cific protein kinases and, through the resultant phosphoryla- tions, regulate various cytoplasmic events (16, 17). Recently polypeptide hormones have been shown to stimulate the syn- thesis of specific proteins in several model systems, and in each case subjected to detailed analysis an accumulation of the corresponding specific mRNA has been demonstrated (18- 20). The regulation of a nuclear event, gene transcription, by an interaction at the surface of the cell is therefore an exciting possibility. To address this issue, we have studied the regu- lation of prolactin biosynthesis by the tripeptide hypotha- lamic releasing factor, thyrotropin-releasing hormone. TRH' has previously been shown to stimulate the accumulation of cytoplasmic prolactin mRNA sequences in a line of pituitary cells (GH) (20). We now demonstrate that TRH acts within minutes to increase prolactin gene transcription. The conse- quence of this effect is the rapid accumulation of nuclear prolactin precursors and a progressive increase in cytoplasmic prolactin mRNA. TRH also rapidly stimulates the phospho- rylation of a novel basic chromatin protein, BRP, distinct from described histone or HMG proteins. It is postulated that the regulation of gene expression by TRH is prototypic of regulation of other genes by peptide hormones in that the initial events at the plasma membrane rapidly lead to a series of nuclear events, including the phosphorylation of a basic chromatin-bound protein and increased transcription of spe- cific genes.
EXPERIMENTAL PROCEDURES
Materials Proteinase K, RNase TI, and DNA polymerase I were obtained
from Boehringer Mannheim. RNase-free DNase (PPRF) and RNase A were obtained from Worthington. ["'SS]Methionine (600-1400 Ci/ mmol), [~u-"P]UTP (>400 Ci/mmol), and [wX2P]dCTP (2000-4000 Cilmmol) were obtained from Amersham Corp. ["P]H,PO, (carrier- free) was obtained from ICN. TRH was purchased from Calbiochem- Behring. GH4 cells were originally established by Dr. Armen Tashjian. Jr.
Methods Cell Culture and Quantitation of Prolactin Biosynthesis"GH4 cells
were routinely cultured on 100-mm plates in Ham's F10 (Gibco) with 12.5% horse serum and 2.5% fetal calf serum. Cells were split 1:4 weekly and fed every other day. Experiments were typically performed to terminate on the 6th day following the split. Prolactin accumula- tion in the media was quantitated by radioimmunoassay (21). Prolac- tin synthesis was measured by pulse-labeling cells with ["S]methio- nine and immunoprecipitation of Triton X-100-soluble protein with
' The abbreviations used are: T R H , thyrotropin-releasing hor- mone; BRP, basic regulated phosphoprotein; HMG, high mobility group; SDS, sodium dodecyl sulfate; CHO, Chinese hamster ovary; MLV, murine leukosis virus; kb, kilobase pairs.
15329
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15330 TRH Stimulates Prolactin Gene Transcription
anti-rat prolactin as described previously (22). usingprotein A instead of second antibody (Pansofbin, Calbiochem-Behring). The radioac- tivity in purified prolactin was quantitated by densitometric scanning of radiographs of the SDS-polyacrylamide gels.
Quantitation of Prolactin mRNA-Cytoplasmic RNA was prepared by phenol/chloroform extraction after lysing the cells in 0.4% Nonidet P-40, 10 mM NaC1, 5 mM MgCI,, 10 mM Tris (pH 8.0). Prolactin sequences were quantitated by hybridizing equal aliquots of this RNA immobilized to squares of diazotized paper with nick-translated cloned prolactin cDNA as previously described (23). Nuclear prolactin RNA was prepared and quantitatively analyzed by DNA-excess hy- hridization to RNA fractionated under denaturing conditions, as previously reported (24). Analysis of serial dilutions of prolactin mRNA standard confirmed the linearity of this assay over a 50-fold concentration range.
Transcriptional Assays-Nascent nuclear RNA transcripts were laheled and quantitated as we have previously described (23,25) using a modification of the trichloroacetic acid precipitation method of McKnight and Palmiter (6.26). Briefly, washed nuclei (approximately 6 X 10S/reaction) were incubated for 20-45 min in the presence of 200 pCi of [‘”PIUTP in 100 pI of buffer containing 150 mM KCI, 5 mM MgCL 1 mM MnCI2, 10 mM Tris (pH 7.8), 10% glycerol, and 1 mM ATP, CTP, and GTP. Labeled RNA was purified by DNase and proteinase K digestion and phenol/chloroform extraction followed by 5% trichloroacetic acid precipitation. The trichloroacetic acid precip- itate was collected on Millipore filters and subjected to a second DNase digestion. Following serial ethanol precipitations, purified labeled RNA was then hybridized to 2 pg of cloned DNA bound to a 3-mm disk of nitrocellulose (500 mM NaCI, 5 mM EDTA, 15 mM NaHPO,, 0.4% SDS, 75 mM PIPES (pH 7.8), and 33% formamide for 60 h). Typically, 25-80 X lo6 cpm of labeled RNA were obtained from each reaction. Background hybridizations to a pBR322 filter (2 p g ) were less than 1.5 ppm when corrected for a machine background of‘ 35 cpm. The use of an intervening sequence as the DNA probe prevented potential variable efficiencies of hybridization produced by accumulating prolactin mRNA, as verified by the constant fractional hybridization with increasing labeled RNA and constant total hybrid- ization with decreasing DNA per filter. Quantitated relative prolactin transcription rates were identical in reactions incubated 10 to 45 min, indicating that transcription, rather than stability, was in fact quan- titated by this assay. For each gene examined in these studies, the inclusion of cu-amanitin (0.8 p M ) in the nascent chain elongation assay reduced gene-specific transcription to the background level, demonstrating that RNA polymerase I1 is exclusively involved in the observed transcriptional effects. Each point is the mean -+ S.E. of the transcription rate (specific counts/min hound/106 counts/min added) of triplicate hybridizations. The group of Chinese hamster ovary random cDNA clones (CHO A to F) and MLV gap/pol and env clones were the generous gift of Michael Wilson (Scripps Institute) and lnder Verma (Salk Institute), respectively.
In experiments designed to measure total transcriptional effects, RNA transcripts were labeled for 10 min under the same conditions except that each reaction contained 100 p M [‘”PIUTP (1-10 Ci/mM) and only 1-2 X 10‘ nuclei. Following purification, the RNA was precipitated with 5% trichloroacetic acid, 1% sodium pyrophosphate and collected on GFC filters. Each point is the mean f S.E. of triplicate reactions.
HRP Phosphorylation-100-mm plates of GH4 cells were preincu- hated with [‘2P]P042- (0.5 mCi/ml) in phosphate-free Ham’s F12 with serum for 5 h prior to addition of hormones. After the appropri- ate incubations, the cells were lysed (1% Triton, 50 mM Na bisulfite, 10 MgCI,, 10% sucrose, 10 mM Tris, pH 7.0) and sulfuric acid-soluble chromatin proteins were prepared as described previously (25). Equiv- alent amounts of trichloroacetic acid-precipitable counts/min were analyzed on two-dimensional gel acid/urea/Triton (5% acetic acid, 5 M urea, 0.38% Triton X-100) in the first dimension and SDS-poly- acrylamide in the second (25). In Fig. 6 the phosphorylation of BRP was quantitated by densitometric scanning of the autoradiographic spot corresponding to BRP. The integrated area was normalized to the density of the H2A spot, which does not vary with TRH additions.
RESULTS
TRH Regulates Prolactin Biosynthesis at the Level of Tran- scription-The regulation of prolactin and growth hormone synthesis in GH4 cells has proved to be a fruitful model system in which to elucidate the sites at which peptide and steroid hormones regulate gene expression (27, 28). TRH
A
300 - -
260 - *
220 - -
180 - - 140 -
- 100 -
1 I 1 I 1 I
o “ 10-11 1 0 - 1 0 10-9 I O - * 1 0 - 7 1 0 - 6
[TRH] M
B
= 1 9 o c
j \ \ \ \
I e a q( I , l I , I I I I I y;:
100 100
0 2 4 6 8 IO 12 14 16 18 20 22 24 2 6 28 30
TIME (hours) FIG. 1. Stimulation of prolactin synthesis and mRNA accu-
mulation by TRH. A, GH4 cell cultures were treated with various concentrations of TRH. Cytoplasmic prolactin mRNA levels (W) were measured after 24 h. The accumulation of prolactin in the media was determined after 48 h (.”-.I. Each point is the mean o f triplicate determinations. E, TRH ( 3 x lo” M) was added to cultures at various times before either preparation and quantita- tion of prolactin mRNA (a”-o) or determination of prolactin synthesis by pulse-labeling and immunoprecipitation (see “Methods”) (0- - -0). Prolactin mRNA was determined in triplicate, prolactin synthesis in duplicate.
stimulates accumulation of prolactin mRNA sequences in these cells (29). The data in Fig. 1 provide additional docu- mentation of the close correlation between the stimulation of prolactin mRNA levels and prolactin biosynthesis. An iden- tical concentration of TRH produces half-maximal stimula- tion of cytoplasmic prolactin mRNA and accumulation of prolactin in the media of GH4 cultures. The kinetic course of stimulation and rate of decay from peak levels are also iden- tical. The correspondence of prolactin mRNA levels and prolactin biosynthesis suggest that, while other factors may influence either the efficiency of prolactin mRNA translation or the stability of newly synthesized prolactin molecules, TRH does not exert effects at these levels.
Because TRH does exert rapid nuclear effects (24), regula- tion at the level of transcription was the most likely mecha- nism of prolactin mRNA accumulation. Fig. 2 shows a more
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from
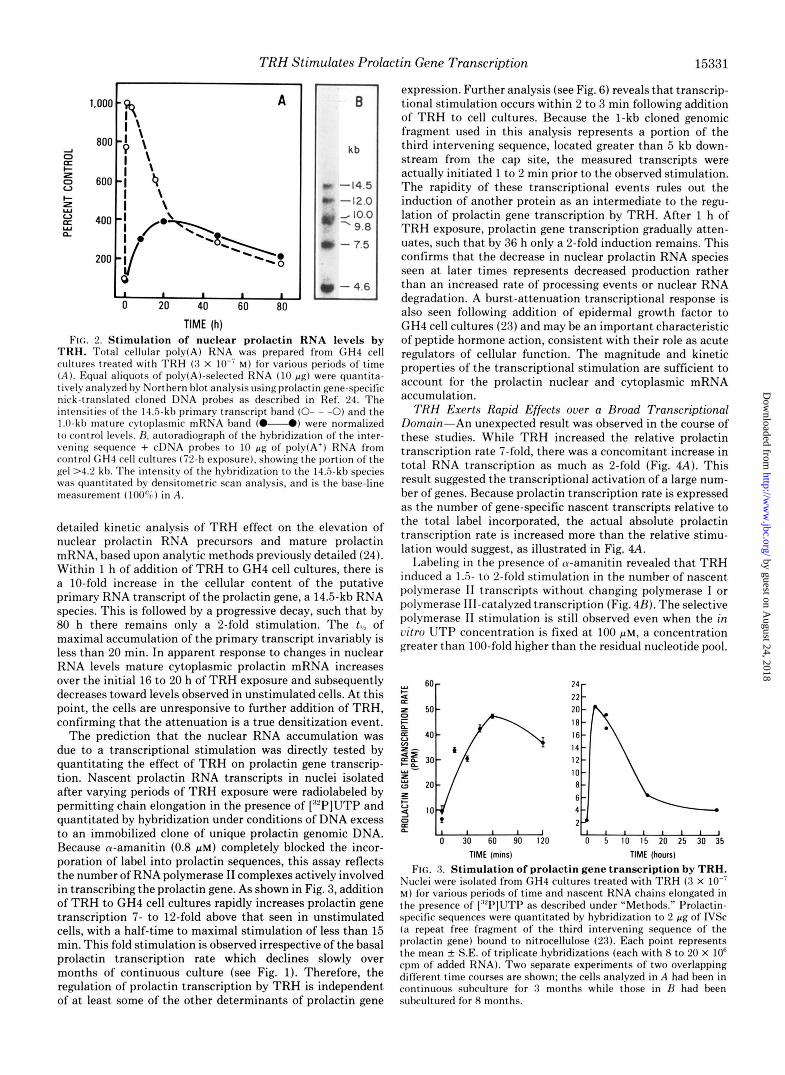
TRH Stimulates Prolactin Gene Transcription 15331
4 0 c z 0 V c
a
V
w a
s a
1.000
800
600
400
200
A
J I I 1 I 0 20 40 60 80
TIME (h)
~
8
kb
I -14.5 c -12.0 - 9.8 0 -7.5
(c' d 10.0
- 4.6
FIG. 2. Stimulation of nuclear prolactin RNA levels by TRH. Total cellular poly(A) RNA was prepared from GH4 cell cultures treated with TRH ( 3 X lo-' M ) for various periods of time ( A ) . Equal aliquots of poly(A)-selected RNA ( 1 0 pg) were quantita- tively analyzed hv Northern blot analysis usingprolactin gene-specific nick-translated cloned DNA probes as described in Ref. 24. The intensities o f t he 14.5-kb primary transcript band (0- - -0) and the 1.0-kl) mature cytoplasmic mRNA band (."-.) were normalized to control levels. H. autoradiograph of the hybridization of the inter- vening sequence + cDNA probes to 1 0 pg of poly(A+) RNA from rontrol GH4 cell cultures (72-h exposure), showing the portion o f the gel >4.2 kl). The intensity of the hybridization to the 14.5-kb species was quantitated by densitometric scan analysis, and is the base-line measurement (100%) in A.
detailed kinetic analysis of TRH effect on the elevation of nuclear prolactin RNA precursors and mature prolactin mRNA, based upon analytic methods previously detailed (24). Within 1 h of addition of TRH to GH4 cell cultures, there is a 10-fold increase in the cellular content of the putative primary RNA transcript of the prolactin gene, a 14.5-kb RNA species. This is followed by a progressive decay, such that by 80 h there remains only a 2-fold stimulation. The t8,2 of maximal accumulation of the primary transcript invariably is less than 20 min. In apparent response to changes in nuclear RNA levels mature cytoplasmic prolactin mRNA increases over the initial 16 to 20 h of TRH exposure and subsequently decreases toward levels observed in unstimulated cells. At this point, the cells are unresponsive to further addition of TRH, confirming that the attenuation is a true densitization event.
The prediction that the nuclear RNA accumulation was due to a transcriptional stimulation was directly tested by quantitating the effect of TRH on prolactin gene transcrip- tion. Nascent prolactin RNA transcripts in nuclei isolated after varying periods of TRH exposure were radiolabeled by permitting chain elongation in the presence of ["'PIUTP and quantitated by hybridization under conditions of DNA excess to an immobilized clone of unique prolactin genomic DNA. Because cu-amanitin (0.8 p ~ ) completely blocked the incor- poration of label into prolactin sequences, this assay reflects the number of RNA polymerase I1 complexes actively involved in transcribing the prolactin gene. As shown in Fig. 3, addition of TRH to GH4 cell cultures rapidly increases prolactin gene transcription 7- to 12-fold above that seen in unstimulated cells, with a half-time to maximal stimulation of less than 15 min. This fold stimulation is observed irrespective of the basal prolactin transcription rate which declines slowly over months of continuous culture (see Fig. 1). Therefore, the regulation of prolactin transcription by TRH is independent of a t least some of the other determinants of prolactin gene
expression. Further analysis (see Fig. 6) reveals that transcrip- tional stimulation occurs within 2 to 3 min following addition of TRH to cell cultures. Because the 1-kb cloned genomic fragment used in this analysis represents a portion of the third intervening sequence, located greater than 5 kb down- stream from the cap site, the measured transcripts were actually initiated 1 to 2 min prior to the observed stimulation. The rapidity of these transcriptional events rules out the induction of another protein as an intermediate to the regu- lation of prolactin gene transcription by TRH. After 1 h of TRH exposure, prolactin gene transcription gradually atten- uates, such that by 36 h only a 2-fold induction remains. This confirms that the decrease in nuclear prolactin RNA species seen at later times represents decreased production rather than an increased rate of processing events or nuclear RNA degradation. A burst-attenuation transcriptional response is also seen following addition of epidermal growth factor to GH4 cell cultures (23) and may be an important characteristic of peptide hormone action, consistent with their role as acute regulators of cellular function. The magnitude and kinetic properties of the transcriptional stimulation are sufficient to account for the prolactin nuclear and cytoplasmic mRNA accumulation.
TRH Exerts Rapid Effects over a Broad Transcriptional Domain-An unexpected result was observed in the course of these studies. While TRH increased the relative prolactin transcription rate 7-fold, there was a concomitant increase in total RNA transcription as much as %fold (Fig. 4A). This result suggested the transcriptional activation of a large num- ber of genes. Because prolactin transcription rate is expressed as the number of gene-specific nascent transcripts relative to the total label incorporated, the actual absolute prolactin transcription rate is increased more than the relative stimu- lation would suggest, as illustrated in Fig. 4.4.
Labeling in the presence of tu-amanitin revealed that TRH induced a 1.5- to 2-fold stimulation in the number of nascent polymerase I1 transcripts without changing polymerase I or polymerase 111-catalyzed transcription (Fig. 4B). The selective polymerase I1 stimulation is still observed even when the in vitro UTP concentration is fixed a t 100 pM, a concentration greater than 100-fold higher than the residual nucleotide pool.
E V a IO 4 tl 0 30 60 90 120 0 5 IO 15 20 25 30 35
TIME (mins) TIME (hours)
FIG. 3. Stimulation of prolactin gene transcription by TRH. Nuclei were isolated from GH4 cultures treated with TRH (3 X lo-' M) for various periods of time and nascent RNA chains elongated in the presence of ['"'PJUTP as described under "Methods." Prolactin- specific sequences were quantitated by hybridization to 2 pg of IVSc (a repeat free fragment of the third intervening sequence of the prolactin gene) bound to nitrocellulose (23). Each point represents the mean k S.E. of triplicate hybridizations (each with 8 to 20 X IO6 cpm of added RNA). Two separate experiments of two overlapping different time courses are shown; the cells analyzed in A had been in continuous subculture for 3 months while those in H had been subcultured for 8 months.
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15332 TRH Stimulates Prolactin Gene Transcription
FIG. 4. Stimulation of RNA polymerase template activity by TRH. A, GH4 cell cultures were treated with TRH (0.3 p ~ ) for various periods of time and isolated nuclei were incubated with [32P]UTP as described. Shown are the total counts/min incorporated (U), the relative prolactin gene-specific transcription in parts per million (U), and the absolute prolactin gene transcription, the product of the relative transcription rate, and the total transcription (M). All values have been normalized to the control level. B, GH4 cultures were treated with TRH (0.3 p ~ ) for various periods of time and total RNA polymerase template activity was measured in the presence of 100 PM UTP (1 Ci/mmol in B, 10 Ci/mmol in C) as described under “Methods.” Total activity (U) and activity in the presence of 0.8 PM cr-amanitin (U) were determined. Each point is the mean & S.E. of triplicate determinations. C, GH4 cultures were incubated with various concentrations of TRH for 30 min and RNA polymerase template activity was measured in the presence (U) or absence (W) of 0.8 PM cr- amanitin. Each point is the mean k S.E. of triplicate determinations.
This suggests that the effect cannot be attributed to changes in nucleotide pools. The maximal effect of TRH occurred within 30 min, followed by a rapid attenuation. As shown in Fig. 4C, this effect of TRH is half-maximal when cells are treated with a concentration of TRH which half-maximally stimulates prolactin biosynthesis, suggesting that both effects could be due to the same receptor-mediated process. The observed transcriptional effects could be the result of a 1.5- to 2-fold stimulation of the transcription of all genes catalyzed by polymerase I1 or the stimulation of a transcription of a restricted set of genes by a proportionally larger extent. The fact that the stimulation affects only polymerase I1 transcripts argues against a general in vitro stabilization of DNA template or RNA transcripts, by a mechanism such as altering DNase or RNase activity.
It was necessary to distinguish between a nonspecific stim- ulation of polymerase activity affecting all genes and the specific stimulation of transcription of a selected subset of genes. T h e effect of TRH on the transcription rate of nine representative polymerase I1 transcription units was therefore studied using rat growth hormone cDNA, a group of six cDNA clones prepared from Chinese hamster ovary cells (30), which are highly homologous to mature messenger RNAs produced in a wide variety of rat tissues,2 and portions of the murine leukosis virus genome. TRH does not stimulate the accumu- lation of the mRNAs encoded by these genes. As shown in Table I, although TRH stimulates prolactin transcription approximately 10-fold, no dramatic changes are seen in the transcription of the other genes. When corrections are made for the TRH stimulation of total polymerase I1 transcription, most of these nine genes show no alteration in transcription rate at all, although one gene (CHO C) did show a small (75%) increase in transcription. Therefore, the effect of TRH on transcription must exhibit gene specificity and is not a general effect on RNA polymerase I1 activity. TO account for the magnitude of the TRH-induced increase in polymerase 11-catalyzed transcription, a large number of transcription units must be stimulated. If each affected polymerase I1 transcription unit was stimulated by TRH to the same extent as the prolactin gene (10-fold), then approximately 10% of
’ R. M. Evans and M. G. Rosenfeld, unpublished observations.
TABLE I TRH effect on specific gene transcription rates
Nuclei were isolated from control and TRH-treated (3 X M, 40 min) GH4 cells and nascent RNA transcripts labeled as described above. 10’ cpm of purified RNA was hybridized to 2 gg of each of the listed cDNAs bound to individual filters; hybridization to each cDNA, expressed as parts per million, is the average of triplicate determi- nations differing less than 10%. Because TRH stimulated the incor- poration of label into total RNA transcripts by 3096, the ratio of the relative gene specific transcription rates (ppmTRH/ppmcontrol) is less than the actual increase in transcription. The final column is the actual fold transcriptional stimulation after correcting for the 30% increase in RNA transcription in the TRH-treated cells. The tran- scription of these genes is entirely inhibited by warnanitin (0.8 p M ) . These results have been confirmed in three separate experiments of similar design, examining early (20 min) and late (10 h) time points.
Prolactin Growth hormone CHO A
B C D E F
MLV env MLV gag-pol
Relative gene specific transcription PPmTRd transcriptional
Actual fold
PPrnmntm’ stimulation Control TRH-treated
PPm 3.36 30.9 9.2 12.0 3.5 3.1 0.9 1.1
19.3 21.0 1.1 1.4 40.0 36.9 0.9 1.1 8.2 10.7 1.3 1.7 4.6 3.4 0.7 1 .o
13.6 10.0 0.7 1.0 4.0 3.6 0.9 1.1
12.6 10.6 0.7 1 .o 8.4 6.9 0.8 1.0
active polymerase I1 templates would be stimulated. Although a complete characterization of these TRH-regulated genes is potentially interesting, the more important question is the identification of the biochemical mechanism(s) responsible for the effects of TRH on gene transcription. One possibility is that the modification of a relatively widely distributed chromatin-associated protein could mediate, in part, the ob- served enhancement of transcription of a large number of transcription units.
The Stimulation of Prolactin Gene Transcription Correlates with the Phosphorylation of a Specific Nuclear Protein, BRP- While the mechanism by which the binding of TRH to i ts plasma membrane receptor results in effects on a nuclear
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

TRH Stimulates Prolactin Gene Transcription 15333
event, gene transcription, is entirely undefined, the phospho- rylation of several cytoplasmic and membrane proteins in response to TRH addition has been well documented (31,32). Presumably, then, TRH either directly or via the formation of a second rnessengeds) activates a protein kinase(s). There- fore, studies were initiated to define whether any rapid phos- phorylation of a nuclear protein(s) occurred in response to TRH. Nuclei were isolated from control and TRH-treated cells preincubated with :“PO4 for 4 to 24 h and examined by two dimensional gels. There is a single detectable nuclear protein whose phosphorylation is stimulated 5- to 10-fold by TRH (Fig. 5 ) . This protein is an acid-soluble, chromatin- associated 23,000-dalton phosphoprotein that co-migrates with H1 on SDS-polyacrylamide gels, but has slower mobility on acid/urea/Triton polyacrylamide gels. Subcellular frac- tionation suggests that most, if not all, of this basic amino acid-rich regulated protein (BRP) is bound to chromatin in the nucleus. As estimated by staining of two-dimensional gels
7 - A
p“
FIG. 5. St imulat ion of BRP phosphoryla t ion of TRH. 100- mm plates o f (;HI cells were incuhated with [:“I’]PO:- for 5 h prior to addition of TRH (0.3 PM) or vehicle alone for an additional 30 min. Equal amounts of trichloroacetic acid-precipitable counts per min of acid-soluble chromatin protein were analyzed on two-dimen- sional polyacrylamide gels (acid/urea/Triton X-100-SDS) as de- scribed under “Methods.” Shown are autoradiographs of control ( A ) and TRH-treated ( R ) cultures. The results of this experiment were confirmed in seven experiments of similar design; while other proteins do occasionally show small, variable alterations in phosphorylation following addition of TRH, the effect of TRH on BRP phosphoryla- tion is highly reproducihle.
1 2 3 4 5 Time Following TRH Addition (min)
FIG. 6. Ea r ly t ime cour se of TRH-st imulated prolactin gene t ranscr ip t ion and BRP phosphoryla t ion . Parallel sets of GH4 cells were treated with TRH (0.3 PM) for various periods of time. One set (A-A) was used to measure prolactin gene transcription as descrihed under “Methods.” The other set (M), which had been preincuhated with [:“P]PO:- was used to prepare acid-soluble chro- matin proteins which were analyzed on two-dimensional gels as in the legend to Fig. 5 . The BRP spot was quantitated by densitometry and standardized (7; stnd) as described under “Met,hods.”
of acid-soluble chromatin proteins, the relative mass of this protein is less than 5% that of a major H1 histone subspecies and is comparable to that of HMG 14 and HMG 17. However, RRP appears to a novel basic amino acid-rich chromatin- associated protein, clearly distinguishable from known his- tone and HMG proteins by its migration in this two-dimen- sional gel system and its salt extractability and insolubility in low concentrations of perchloric acid. The TRH-stimulated phosphorylation appears to exclusively involve serine resi- dues.” Fig. 6 summarizes the acute effects of TRH addition on BRP phosphorylation, as quantitated by densitometry, and on prolactin gene transcription. There is a remarkably rapid onset of the hormone-stimulated phosphorylation and transcription, and both events have similar kinetics. Thus, RRP phosphorylation is significantly increased after 2 min of TRH treatment, while prolactin gene transcription is simi- larly increased within a few minutes. Since, as previously discussed, the probe used for the transcriptional assay imposes an obligatory 1- to 2-min lag prior to recording any increased initiation of transcription, the two curves shown in Fig. 6 would reflect nearly simultaneous events. With continuing exposure to TRH, BRP phosphorylation gradually decreases, reaching levels only slightly above control by 36 h. In almost all cases, TRH produces a 7- to 12-fold increase in transcrip- tion and a comparable increase in BRP phosphorylation; however, in those experiments where there is a markedly diminished transcriptional effect there remains a close quan- titative correlation between the two events. The similarity in the rapid kinetic responses of hormone-stimulated BRP phos- phorylation and increased prolactin gene transcription is con- sistent with the possibility that both events represent primary effects of the TRH-receptor interaction.
DISCUSSION
In this manuscript, we have examined the regulation of prolactin biosynthesis by TRH in the rat pituitary cell line,
:’ M. Waterman, G. H. Murdoch, R. M. Evans, and M. G. Rosenfeld, manuscript in preparation.
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15334 TRH Stimulates Prolactin Gene Transcription
GH4, and document that it exerts rapid, specific effects on gene transcription. Within minutes following binding to plasma membrane receptors on the surface of the cell, TRH produces a 7- to 12-fold stimulation of prolactin gene tran- scription. This transcriptional effect could entirely account for the observed actions of TRH on prolactin biosynthesis and indicates that the mechanisms responsible for the nuclear effect are closely linked with the binding of TRH to its receptor.
One well described mechanism whereby the peptide-recep- tor interaction on the cell surface influences intracellular processes is the activation of protein kinases. The results presented in this manuscript demonstrate a similar time course of the phosphorylation of a novel nuclear protein, BRP, and the transcription regulation of a specific gene by TRH. The physiochemical properties of BRP are clearly distinct from all known H1 subtypes or HMG proteins; how- ever, it may be similar to other nuclear proteins (33). It is therefore important to investigate the possibility that phos- phorylation of BRP itself by an as yet unidentified hormone- activated protein kinase could enhance the transcription of specific genes.
The regulation of prolactin gene expression by a mechanism involving a reversible chemical modification of a protein, (e.g. such as BRP phosphorylation) would allow acute alterations in transcriptional rate. The transcriptional stimulation is so rapid that receptor-hormone internalization and translation to the nucleus becomes an unlikely explanation of the tran- scriptional effect. The responsiveness of prolactin gene tran- scription to membrane-bound TRH is more dramatically il- lustrated during the process of desensitization. Although the mechanism of desensitization is not known, the half-time for the decay of the transcriptional stimulation, 7 h, is very similar to that reported for the down-regulation of the TRH receptor (34). These data are consistent with a close linkage between the occupancy of TRH receptors on the surface of the cell and the rate of prolactin gene transcription, alt,hough it is presently unclear exactly which of the many putative second messenger systems activated by TRH (30, 31, 35-37) is responsible for the transcriptional effects. Although stim- ulation of BRP phosphorylation and of prolactin transcription is produced by administration of CAMP analogs (25), TRH appears to produce phosphorylation of BRP via a CAMP- independent mechan i~m,~ predicting a potentially complex regulation of BRP phosphorylation.
In this manuscript, TRH is documented to have a transcrip- tional effect on a large number of genes by directly measuring the amount of polymerase I1 complexes active after hormone treatment by labeling nascent transcripts with [32P]UTP. Through analyzing the nascent transcripts by hybridization to random cDNA probes, it is apparent that the observed stimulation is not simply the result of a general activation of polymerase I1 activity, but rather represents a stimulation of transcriptional activity of a large number of individual genes. In fact, analysis of chromatin structure by techniques such as Escherichia coli RNA polymerase binding sites (38) has indi- rectly suggested that other hormones might modify transcrip- tion of a large number of genes. There are probably multiple reasons why the number of genes which are transcriptionally stimulated by a hormone have been underestimated by ana- lyzing changes in cellular protein synthesis. These include the fact that the RNA polymerase I1 transcripts of many genes are not quantitatively processed into cytoplasmic mRNAs (30), that the magnitude of the transcriptional stimulation
G. H. Murdoch, M. Waterman, R. M. Evans, and M. G. Rosenfeld, manuscript in preparation.
appears to be gene-specific and small changes in many pro- teins may be overlooked, and that most analyses have been performed a t times long after transcriptional effects have attenuated. However, in one detailed study using two-dimen- sional gel analysis of pulse-labeled proteins from nerve growth factor-treated PC12 neuroblastoma cells, quantitated by com- puter-assisted scanning densitometers, the synthetic rate of up to 30% of observed proteins were measurably increased (39). A rapid attenuation of a transcriptional stimulation produced by nerve growth factor would account for the small stimulations (30% increases) observed, because the cells were labeled 72 h after addition of nerve growth factor.
The regulation of prolactin gene expression by peptide hormones appears to be an interesting model system in which to study the nuclear effects of agents which initially interact with the surface of the cell. Prolactin gene transcription is stimulated by alterations of intracellular CAMP, epidermal growth factor, thyrotropin-releasing factor, and several other peptide hormones (23, 25)2 and inhibited by dopamine (40). Each of the positive transcriptional regulators of the prolactin gene is believed to activate distinct second messenger systems. In order to appreciate the potential for complex transcrip- tional regulation produced by interactions between different polypeptide hormones, it will be necessary to determine whether there is a final common molecular event which influ- ences transcriptional activity, and to evaluate the role, if any, of phosphorylation of BRP in enhancing prolactin gene tran- scription.
A~knoruled~ments-We thank Dr. Tony Hunter, Dr. Geoffrey Wahl, and Dr. Gordon Gill for their critical suggestions during the preparation of this manuscript.
1.
:i . 2.
4.
5.
6.
r I .
8.
9.
10.
11. 12.
19.
14.
1s. 16. 17
18
19 20
REFERENCES
Yamamoto, K. R., and Alberts, B. M. (1976) Annu. Rev. Biochem.
Adhya, S., and Garges, S. (1982) Cell 7, 6628-6632 Peterkofsky, A. (1976) Adu. Cyclic Nucleotide Res. 7, 1-35 Ringold, G. M., Yamamoto, K. R., Bishop, J. M., and Varmus,
H. (1977) Proc. Natl. Acad. Sci. U. S. A. 74, 2879-2883 Roop, D. R., Nordstrom, J. L., Tsai, S. Y., Tsai, M., and O’Malley,
B. (1978) Cell 15, 571-685 McKnight, G. S., and Palmiter, R. D. (1979) J. Bid. Chem. 254,
9050-9058 Swaneck, G. E., Nordstrom, J. L., Kreuzalek, F., Tsai, M. J., and
O’Malley, B. W. (1979) Proc. Natl. Acad. Sci. U. S. A. 76, 1049-1053
Mayo, K. E., and Palmiter, R. D. (1981) J. Biol. Chem. 256, 2621-2624
Evans, R. E., Birnberg, N. C., and Rosenfeld, M. G . (1982) Proc. Natl. Acad. Sci. U. S. A. 79, 7659-7663
Spindler, S. R., Mellon, S. H., and Baxter, J. D. (1982) J. Bid. Chem. 257, 11627-11632
Kurtz, D. T. (1981) Nature (Lond.) 291,629-631 Lee, F., Mulligan, R., Berg, P., and Ringold, G. (1982) Nature
Doehmer, J., Barinaga, M., Vale, W., Rosenfeld, M. G., Verma, I. M.. and Evans. R. M. (1982) Proc. Natl. Acad. Sci. U. S. A.
45,721-746
(LO&.) 294,228-232
79,2268-2272 ’
son, J.. and Yamamoto, K. (1981) Proc. Natl. A c ~ . s~i. u. s. Payvar, F., Wrange, 0.. Carlstedt-Duke, J., O’Kret, S., Gustafa-
A . 75,6628-6632 Compton, d . G., Shrader, W. T., and OMalley, B. (1983) Proc.
Roach, P. J. (1981) Curr. Top. Cell. Regul. 20, 45-106 Dedman, J. R., Brinkley, B. R., and Means, A. R. (1979) Adu.
Matusik, R. J., and Rosen, J. M. (1978) J. Biol. Chem. 253,
Natl. Acad. Sci. U. S. A. 80, 16-20
Cyclic Nucleotide Res. 11, 131-174
2343-2347 Lynch, K. R. (1982) Cell 28, 185-189 Evans, G. A,, and Rosenfeld, M. G. (1979) J . Biol. Chem. 254,
8023-8030
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

TRH Stimulates Prolactin Gene Transcription 15335
21. Murdoch, G. H., and Rosenfeld, M. G . (1981) J . Bid. Chem. 256,
2 2 . Dannies, P. S., and Tashjian, A. H., Jr. (1973) J. Biol. Chem.
23. Murdoch, G. H., Potter, E., Nicolaisen, A. K., Evans, R. M., and Rosenfeld, M. G. (1982) Nature (Lond.) 300, 192-194
24. Potter, E., Nicolaisen, A. K., Ong, E. S., Evans, R. M., and Rosenfeld, M. G. (1981) Proc. Natl. Acad. Sci. U. S. A. 78, 6662-6668
25. Murdoch, G. H., Rosenfeld, M. G., and Evans, R. M. (1982) Science (Wash. D. C.) 218, 1315-1317
26. Compere, S. J., McKnight, G. S., and Palmiter, R. D. (1981) J. Biol. Chem. 256, 6341-6347
27. Bancroft, F. C. (1981) Functionally Differentiated CellLines (Sato, et al. eds), pp. 47-59, Alan R. Liss, New York
28. Tashjian, A. H., Jr . (1979) Methods Enzymol. 58, 527-535 29. Evans, G. A,, David, D. N., and Rosenfeld, M. G. (1978) Proc.
30. Harpold, M. M., Evans, R. M., Salditt-Georgieff, M., and Darnell,
4050-4055
248,6174-6179
Natl. Acad. Sci. U. S. A. 75, 1294-1298
J. E. (1979) Cell 17, 1025-1035
Chem. 257,3306-3312
7566-7573
31. Drust, D. S., Sutton, C. A., and Martin, T. F. J. (1982) J. Bid.
32. Drust, D. S., and Martin, T. F. J. (1982) J. Biol. Chem. 257,
33. Yu, M. W., Tolson, N. W., and Guroff, G. (1980) J . Bid. Chem.
34. Hinkle, P. M., and Tashjian, A. H., Jr. (1975) Biochemistry 14,
3 . 5 . Gershengorn, M. C. (1980) J. Biol. Chem. 255, 1801-1803
255, 10481-10492
3845-3850
36. Sutton, C. A., and Martin, T. F. J. (1982) Endocrinology 110, 1273-1280
X . Tan, K.-N., and Tashjian, A. H., Jr. (1981) J. Biol. Chem. 256,
38. dohnson, L. K., Baxter, J. D., Vlodavsky, I., and Gospodarowicz,
39. Garrels, J. I., and Schubert, D. (1979) J. Biol. Chem. 254, 7978-
40. Maurer, R. A. (1981) Nature (Lond.) 294,94-97
8994-9002
D. (1980) Proc. Natl. Acad. Sci. U. S. A . 77, 394-398
7985
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from

G H Murdoch, R Franco, R M Evans and M G Rosenfeldphosphorylation of a specific nuclear protein.
hormone rapidly stimulates both transcription of the prolactin gene and the Polypeptide hormone regulation of gene expression. Thyrotropin-releasing
1983, 258:15329-15335.J. Biol. Chem.
http://www.jbc.org/content/258/24/15329Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/258/24/15329.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on August 24, 2018
http://ww
w.jbc.org/
Dow
nloaded from