plant cyanogenic glycosides
TRANSCRIPT

Plant cyanogenic glycosides
Ja nos Vetter
Department of Botany, University of Veterinary Sciences, Budapest, 1400 Pf. 2. Hungary
Received 12 February 1999; accepted 9 March 1999
Abstract
The cyanogenic glycosides belong to the products of secondary metabolism, to thenatural products of plants. These compounds are composed of an a-hydroxynitrile typeaglycone and of a sugar moiety (mostly D-glucose). The distribution of the cyanogenic
glycosides (CGs) in the plant kingdom is relatively wide, the number of CG-containing taxais at least 2500, and a lot of such taxa belong to families Fabaceae, Rosaceae, Linaceae,Compositae and others. Di�erent methods of determination are discussed (including the
indirect classical photometrical and the new direct chromatographic ones). The geneticcontrol of cyanogenesis has no unique mechanism, the plants show variation in the amountof the produced HCN. The production of HCN depends on both the biosynthesis of CGs
and on the existence (or absence) of its degrading enzymes. The biosynthetic precursors ofthe CGs are di�erent L-amino acids, these are hydroxylated then the N-hydroxylaminoacids are converted to aldoximes, these are turned into nitriles. The last ones are
hydroxylated to a-hydroxynitriles and then they are glycosilated to CGs. The generation ofHCN from CGs is a two step process involving a deglycosilation and a cleavage of themolecule (regulated by b-glucosidase and a-hydroxynitrilase). The tissue levelcompartmentalisation of CGs and their hydrolysing enzymes prevents large-scale hydrolysis
in intact plant tissue. The actual level of CGs is determined by various factors bothdevelopmental and ecological ones, which are reviewed too. The last part of the presentwork demonstrates the biological roles of CGs in plant physiological processes and in plant
defence mechanisms as well. The e�ect of CGs (HCN) on di�erent animals, the symptomsof poisonings are discussed to cows, sheep, donkeys, horses and chicks. Finally, thepoisonous e�ects of cassava (Manihot esculenta ) roots are summarised on experimental
animals and on the human organism. # 1999 Elsevier Science Ltd. All rights reserved.
0041-0101/99/$ - see front matter # 1999 Elsevier Science Ltd. All rights reserved.
PII: S0041 -0101 (99)00128 -2
Toxicon 38 (2000) 11±36
www.elsevier.com/locate/toxicon

Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
2. Occurrence, structure, distribution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
3. Estimation assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
4. Cyanogenesis: genetic background . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
5. Cyanogenesis: biosynthesis and transport . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
6. Regulation of CG level in plants. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
7. Biological e�ects of CGs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 177.1. Role in plant physiological processes. . . . . . . . . . . . . . . . . . . . . . . . . . . . 17
7.2. Role in plant defence mechanism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187.3. E�ects on animals . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187.4. E�ects on human organism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
1. Introduction
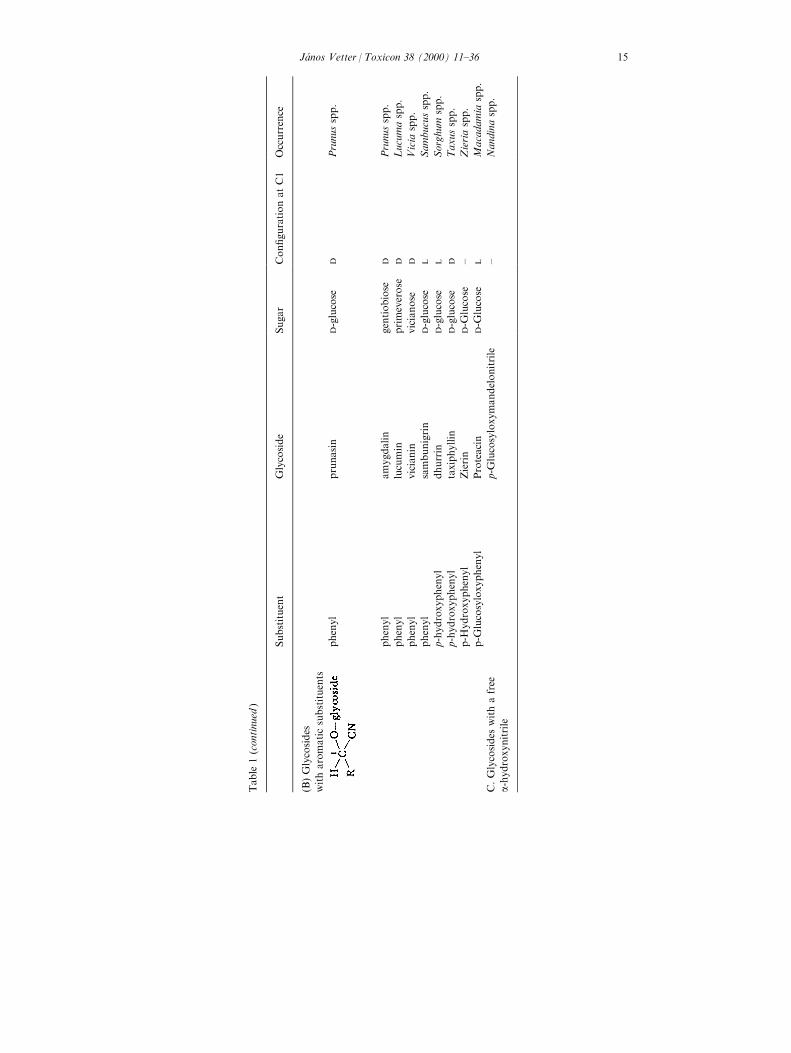
The cyanogenic glycosides (abbreviated: CG) may be de®ned chemically asglycosides of a-hydroxinitriles and belong to the secondary metabolites of plants(to natural products). They are amino acid-derived plants constituents, present inmore than 2500 plant species. Hydrogen cyanide (HCN) as a product of theirhydrolysis was ®rstly isolated from plant in 1802 by Scrade (from bitter almondand from the leaves of peach). The release of HCN by plants was ®rstly ascribedto a particular compound by Robiquet and Charlard who isolated amygdalinfrom bitter almonds. On enzymatic hydrolysis cyanogen glycosides yield theaglycone (that is an a-hydroxynitrile) and the sugar moiety. The aglycones can begrouped into aliphatic and aromatic compounds; the sugar is mostly D-glucose,but can be other sugars too, for example gentibiose, primeverose or other (seeTable 1).
2. Occurrence, structure, distribution
Whereas most plants produce a small amount of cyanide associated withethylene production, between 3±12000 plant species produce su�cient quantitiesof cyanogenic compounds (McMahon et al., 1995). The CGs are glycosides of a-hydroxynitriles, all known compounds are b-linked, mostly with D-glucose. Thestructure of some cyanogenic glycosides and some examples of their occurrence
JaÂnos Vetter / Toxicon 38 (2000) 11±3612

are given in Table 1. The enzymic hydrolysis produces the aglycone and the sugarmoiety. The CGs can be grouped according to chemical nature of substituents,namely aliphatic, aromatic groups and into the glycosides with a free a-hydroxynitrile. Some of these CGs are better known than the others because thecarrying plant species (group) has a greater practical importance, severaleconomically important plants are highly cyanogenetic (linamarin in Manihotesculenta, Linum usitatissimum, Trifolium repens, dhurrin in Sorghum species,amgydalin in rosaceous plants, lotaustralin in Lotus corniculatus, etc.).
The linamarin and lotaustralin have a relatively broad distribution in the plantkingdom, having been demonstrated in the following plant families: Compositae,Euphorbiaceae, Linaceae, Papaveraceae and Fabaceae (Leguminosae). A similarwide distribution has been observed for prunasin in six families (Polypodiaceae,Myrtaceae, Rosaceae, Saxifragaceae, Scrophulariaceae and Myoporaceae).Sambunigrin, vicianin, amygdalin, which are closely related to prunasin, havebeen demonstrated in three (Caprifoliaceae, Mimosaceae, Oleaceae), in two(Polypodiaceae, Fabaceae) and only in one (Rosaceae) families, respectively. Themore common distribution pattern is that a particular cyanogenic compound willoccur in one or two families. Conversely, it is generally true, that, with fewexceptions, only one or two characteristic glycoside will occur in a given plantfamily (Poaceae: dhurrin; Compositae: linamarin; Polypodiaceae: prunasin andvicianin, Rosaceae: amygdalin and prunasin). The cyanogenic compounds ofplants belong undoubtedly to secondary plant metabolites which have or can havea chemotaxonomical character. Let's see their taxonomical distribution from thispoint of view. The majority of these families belongs to the Angyospermatophyta,but there are some exceptions (Polypodiaceae/Pteridophyta/, Taxaceae/Gymnospermatophyta). Both the class Dicotyledonopsida andMonocotyledonopsida have plant families with cyanogenetic compounds, but themost families belong to the dicots. The families Saxifragaceae, Rosaceae,Mimosaceae, Fabaceae, Myrtaceae, Linaceae and Euphorbiaceae are in thesubclass Rosiidae, other cyanogenous families are in subclasses Ranunculidae(family Papaveraceae), in Lamiidae (families Caprifoliaceae, Sambucaceae,Oleaceae), Asteridae (family Compositae). The occurrence or missing of thecyanogenetic compounds has probable chemotaxonomical importance too, butthese relations are less, or not, documented or investigated.
CGs have been reported from many members of the three subfamilies ofFabaceae, but some of these reports should be recon®rmed (Seigler et al., 1989).Cyanogenic members of the Papilionoideae have been reported from 18 tribes (theability to produce HCN upon hydrolysis), but the compounds responsible havebeen isolated only from a few of these tribes. Linamarin and lotaustralin havebeen found in many species of the tribes Loteae and Trifoliae, probably occur inthe Coronilleae. Vicianin occurs in seeds of several Vicia species. At least onemember of the Crotalarieae appears to contain prunasin. Cyanogenic compoundsfrom the Galegeae have been reported, but their chemical nature is unclear.Species of the Indigofereae appear to contain prunasin or sambunigrin, whereasthose of the Phaseoleae contain linamarin and lotaustralin.
JaÂnos Vetter / Toxicon 38 (2000) 11±36 13

Table
1
Thechem
icalstructure
ofsomecyanogenic
glycosides
(accordingto
Tapper
andReay,1973)
Substituent
Glycoside
Sugar
Con®gurationatC1
Occurrence
(A)Glycosides
withaliphaticsubstitutes
R1R'1
CH
3-
linamarin
D-glucose
Linum
spp.
Trifolium
spp.
R1CH
3-,R'1
CH
3CH
2-
lotaustralin
D-glucose
Lotusspp.
Maniholtspp.
R1(C
H3) 2CH
aciapetalin
D-G
lucose
Acaciaspp.
R1HCO
2CH1CH(C
O2HCH
2)C
Triglochinin
D-glucose
Triglochin
spp.
R1H,R'1
Hdeidaclin
D-glucose
Deidamia
spp.
tetraphyllin
AD-glucose
Tetrapathaea
spp.
R1OH,R'1
Htetraphyllin
BD-G
lucose
Tetrapathaea
spp.
R1OH,R'1
HGynocardin
D-glucose
Gynocardia
spp.
Pangium
spp.
JaÂnos Vetter / Toxicon 38 (2000) 11±3614
Table
1(continued
)
Substituent
Glycoside
Sugar
Con®gurationatC1
Occurrence
(B)Glycosides
witharomaticsubstituents
phenyl
prunasin
D-glucose
DPrunusspp.
phenyl
amygdalin
gentiobiose
DPrunusspp.
phenyl
lucumin
primeverose
DLucumaspp.
phenyl
vicianin
vicianose
DVicia
spp.
phenyl
sambunigrin
D-glucose
LSambucusspp.
p-hydroxyphenyl
dhurrin
D-glucose
LSorghum
spp.
p-hydroxyphenyl
taxiphyllin
D-glucose
DTaxusspp.
p-H
ydroxyphenyl
Zierin
D-G
lucose
±Zieriaspp.
p-G
lucosyloxyphenyl
Proteacin
D-G
lucose
LMacadamia
spp.
C.Glycosides
withafree
a-hydroxynitrile
p-G
lucosyloxymandelonitrile
±Nandinaspp.
JaÂnos Vetter / Toxicon 38 (2000) 11±36 15

The new, more developed analytical methods isolate and identify newcyanogenic compounds, or new derivates of known compounds. The 6-trans-2butenoyl ester of prunasin was isolated as a new CG from Centaurea aspera var.subinermis (Cardona et al., 1992), the purshianin from the Purshia tridentata(Rosaceae: Nakanishi et al., 1994), the multi®din from the latex of Jatropamulti®da (Euphorbiaceae: Berg et al., 1995). An other new CG (esteri®ed with aniridoid glycoside) was isolated from Canthium schimperianum (Schwartz et al.,1996). In other cases CGs (or HCN) were identi®ed and determined from plantspecies or varieties: in seeds of Hevea brasiliensis (Mallika et al., 1991), in leavesand callus cultures of Schlecterina mitostemmatoides/Passi¯oraceae (Jager et al.,1995), in di�erent morphological parts of Moringa oleifera (Moringaceae: Makkarand Becker, 1997), in some legumes, grasses and other plants (Aikman et al.,1996).
3. Estimation assays
There are di�erent possibilities for the qualitative, semi-quantitative orquantitative determinations of the compounds. The ®rst group of the estimationscontains direct methods, the second one is based on the preliminary hydrolysisand quanti®cation of HCN. Because of the medical signi®cance of cyanide, it isnot surprising that most research data are reported in terms of potential cyanideyields rather than the glycoside content itself. Kawamura et al. (1993) suggested amethod for the direct estimation of linamarin in beans, bean paste products andcassava ¯our. A sample was extracted with a mixture of acetonitrile and water,cleaned up by using a solid-phase extraction cartridge and trimethylsilated, thenlinamarin was quantitated by FID-GC and con®rmed by GC/MS (detection limitsof linamarin were 0.1±1 mg/g plant material). Application of Sephadex LH-20chromatography was described by Amarowicz and Shahidi (1994). This methodcan be used as a ®rst step for the separation of CGs and hydrophilic phenoliccompounds from ¯axseed. The taxiphyllin content of bamboo shoots wasdetermined by HPLC and air steam distillation±alkaline titration (SD±AT)method (Chang and Hwang, 1993). A solvent extraction, either directly usingfresh shoots or using a freeze-dried sample, was a pretreatment for the HPLCmethod. The direct extraction with methanol was more convenient and less timeconsuming. The HPLC method determined taxiphyllin quantitatively andqualitatively, whereas the ASD±AT method measures taxiphyllin as the HCNreaction product.
The second main group of methods are based on the quantitation of HCNcontent liberated from the original plant material. To this group belong theclassical (conventional) simple and rapid semi-quantitative methods. The classicaltest of Guignard (1906) is based on the reaction of alkaline picrate paper withHCN liberated in a closed test tube by spontaneous enzymatic hydrolysis ofcrushed, digerated plant material. Cyanide can be detected at 30±50 mg/gconcentration. This test has been adapted for quantitation by Boyd et al. (1938),
JaÂnos Vetter / Toxicon 38 (2000) 11±3616

later calibrated for colorimetry in the 5±50 mg/g extract range by Gilchrist et al.(1967). There are three distinct steps in the HCN assay: liberation of cyanide; itsextraction, isolation from the plant material and determination. Most of the earlymethods relied on acidic or spontaneous autolysis by the own enzymes in rupturedplant tissues. Later the hydrolysis was carried out by both the endogenous and theexogenous added enzymes, or was performed by b-glucosidase added exogenous.The duration and temperature of the hydrolysis are di�erent in di�erent methods.Cyanide recovery from the plant substrate is normally carried out by aspirationwith air, nitrogen or water vapour and trapping of cyanide in alkaline solution.Critical factors include timing, temperature, volume of substrate sample size andothers. Numerous quantitative methods, di�ering in sensitivity and detectionlimits, haven been employed in determining the isolated cyanide. Titration withacid or alkaline silver nitrate is a well-known classical standard analyticalprocedure (AOAC 1965). The colorimetry with picric acid detects cyanide in the5±50 mg range. A modi®ed colourimetric method and a simple equipment weredeveloped by the author and his coworker (Vetter and Haraszti, 1975: Fig. 1).This method is based on the enzymatic hydrolysis of homogenate of fresh plantmaterial (100±200 mg) The spontaneous hydrolysis was supplemented adding of 1mg of b-glucosidase enzyme (to 100 mL homogenate).The aspiration of theliberated HCN was performed by CO2 free air, the transported HCN was trappedin alkaline solution of picric acid. After the reaction time (30 min) at 528Ctemperature the colour was measured at 480 nm. The quantity of the liberated
Fig. 1. Equipment for the determination of HCN from plant tissue: (1) water bath; (2) air compressor;
(3) trapping of CO2; (4) tubes of air stream; (5) liberation of HCN; (6) plant homogenate; (7) vessel of
HCN trapping.
JaÂnos Vetter / Toxicon 38 (2000) 11±36 17

HCN can be measured by cyanide-sensing electrodes. An other variant of theestimation is the potentiometric measurement by cianide-sensitive electrodes. The®rst successful attempt was made by Gillingham et al. (1969), according to ourexperience, this type of determination requires relative more time although itsdetection's limit is better than the sensitivity of the colourimetric method. In thenew work of Prokofjeva et al. (1990) a good correlation was found between theresults obtained by cyanide-sensitive electrode and by the mercurometric method.This method is recommended for the rapid evaluation of cyanogenic potential ofplant seeds.
A modern variant of the picric acid methods was developed by Hin et al.(1996). Their system is based on the hydrolysis of linamarin by stabilised leaflinamarase with detection of the cyanide by an alkaline picrate reagent. A positivereaction was observed within 15 min at 378C and it could detect linamarin at aconcentration as low as 0.5±1 mM. These paper strips were used for theestimation of linamarin content semi-quantitatively. In the second variant of themethod of Hin et al. (1997) the colour from the picrate paper was eluted withwater and its absorbance was measured at 510 nm.
Some other new methods contain di�erent elements of older and newer ones.For example, some new chromatographic steps. Chadha et al. (1995) described anew method for the estimation of cyanide in ¯axseeds, it is carried out byhomogenising the sample with water, letting it stand, ®ltering it through amembrane and then injecting the ®ltrate into an HPLC system (consisting of ananion exchange column and an electrochemical detector). Kobaisy et al. (1996)used a crude enzyme extracted from the ¯axseed for the hydrolysis of cyanogenicglycosides. Two colourimetric methods using the crude enzyme extract and anestablished HPLC method were compared. The total cyanide values obtained byall three methods were not signi®cantly di�erent. Some new methods weredeveloped for the estimation of cyanogens of M. esculenta (cassava). Essers et al.(1993) improved the enzymatic method with a more acceptable colourdevelopment procedure and other minor changes. Isonicotinate-1,3-dimethylbarbiturate as the reagent is less poisonous and more stable than theearlier piridin/pyralozone colour reagent. Chassagne et al. (1996) extracted thecyan glycosides using Amberlite XAD from fruits of several edible Passi¯oraspecies.
4. Cyanogenesis: genetic background
The inheritance, the genetic control of cyanogenesis has not an uniquemechanism. The results regarding to di�erent cyanogenic plant taxa are di�erent.All the cyanogenic plant species which have been investigated in any detail showvariation in the amount of the produces HCN. This variation re¯ects variation inboth the production of CGs themselves and the enzyme which degrades them.Variation occurs within individual plants or genotypes, depending on di�erentphysiological or ecological factors.
JaÂnos Vetter / Toxicon 38 (2000) 11±3618

In white clover (Trifolium repens ) the discrete form of variation is controlled bytwo genes according to the classic result of Corkill (1942). The presence orabsence of the both the glycosides is regulated by alleles of a single gene,designated Ac, whereas the presence or absence of the enzyme linamarase isgoverned by alleles of another, independently inherited, gene (Li). Only plantswhich posses dominant functional alleles of both genes liberate HCN whendamaged. However, the presence of a functional allele of only one of these genesmay be shown by adding either a glycoside or enzyme extract to the test material.Both genes also e�ect vegetative and reproductive characters of the plants,especially seed and ¯ower production. Female reproductive ®tness of acac plantsis about double the of Acac plants (Kakes, 1997). It is suggested that di�erencesin male fertility between Acac and acac plants are either due to di�erences in¯ower attraction and/or a di�erence in fertilisation e�ciency between Ac and acbearing pollen. There are very interesting studies on the e�ect of di�erentecological factors on cyanogenetic character of white clover. The cyanogeneticfrequency in the US white clover germplasm collection was greater in accessioncollected at low altitudes and at sites with high winter temperature, lower summerprecipitation, greater spring cloudiness and less snow cover (Pederson et al., 1996).Caradus and Forde (1996) collected white clover populations from the Caucasusand from moist, lowland, highly fertile environment. There was a negativerelationship between the altitude of the origin of the population and theproportion of cyanogenic plants; populations from severely grazed sites were lessupright and less cyanogenic than populations from infrequently or nil defoliatedsites. The comparison of the cyanogenic and the acyanogenic morphs has aagricultural, practical importance too. Noitsakis and Jacquard (1992)demonstrated in experimental conditions that the acyanogenic plants have acompetitive advantage over the cyanogenic ones (the relative ®tness, i.e. thebiomass production and the number of ¯owers per plant are higher). Thebiological production and the cyanogenic or acyanogenic character of a whiteclover cultivar are complex questions, importance of which is demonstrated as aproblem in the adaptation of US-cultivars in New Zealand (Widdup et al., 1996).
M. esculenta: there are cultivar-dependent di�erences in root cyanogenicglycoside content (Cook, 1985; Wheatley et al., 1993). Most cassava cultivars haveroot CG contents less than 100 mg/kg fresh w., but there are many `bitter' ones,that contain up to 500 mg/kg. Signi®cantly no acyanogenic cultivars have beenidenti®ed. The genetic background of the inheritance in cassava is problematic,but the e�ect on di�erent factors on actual concentration of CG as well as thegenes of cyanogenic enzymes linamarase and hydroxynitrile liase is betterdemonstrated (McMahon et al., 1995).
The phenotypic distribution and genetic variation for cyanogenesis of Turneraulmifolia was studied (Shappert and Shore, 1995. Controlled crosses andgreenhouse studies showed that there is a genetic basis to the variation withbetween family variance accounting for more than 80% of the variation in onepopulation. The dhurrin containing Sorghum species and hybrids (S. bicolor, S.sudanense etc.) show a great variability in the HCN production. Authors di�er in
JaÂnos Vetter / Toxicon 38 (2000) 11±36 19

their conclusions about the number of genes involved and the dominancerelationships of these genes (Nass, 1972). Some of these concluded that in thesudangrass there is one pair of genes determining high versus low cyanide contentand that low HCN production is dominant over the high one. Others showed thatthe inheritance is multigenic, with high HCN production partially dominant tolow production. Earlier Carlson (1958) founded low HCN production associatedwith the following loci: green striped-1, golden-2, yellow leaf tip, single seededspikelets. Studies on the variation, changes of cyanogenic glycoside (and/or HCN)contents of ¯axseed (Linum usitatissimum ) estimated its dependency on cultivar inCanada (Oomah et al., 1992). No signi®cant di�erences were demonstrated inHCN content of di�erent cultivars of Lotus corniculatus in a study of the author'swork team (Borsos et al., 1976), but we measured sharp changes during thedevelopment of plants.
5. Cyanogenesis: biosynthesis and transport
The biosynthesis of cyanogenic glycosides. The relationship between aminoacids and CGs was earlier clearly demonstrated with the incorporation of 14C-labelled tyrosine into the hydrolysis products of dhurrin from Sorghum. Similarresults have been obtained with other glycosides in a range of plants (theprecursors are: L-valine (for linamarin); L-isoleucine (for lotaustralin); L-phenilalanine (for prunasin). The general pathway of biosynthesis of cyanogenicglycosides is given after Tapper and Reay (1973) (Fig. 2), i.e. the a-amino acidsare hydroxylated to form a N-hydroxylamino acid, which is then converted to analdoxime, this in turn to a nitrile. The nitrile is hydroxylated to form an a-hydroxynitrile, which is glucosylated to form the corresponding CG (McFarlane etal., 1975). 2-Hydroxyisobutyraldoxime, 2-hydroxiphenylacetaldoxime and theirglycosides should be considered to be part of a possible alternative pathway ofCG biosynthesis (see the dotted arrows of the ®gure). Stereochemical studiesindicated the involvement of a monooxygenase in the hydroxylation step.Researches on linamarin biosynthesis in cassava were and are coming intoprominence. The precursor of the linamarin synthesis is the valine. Recently,Moller's group has demonstrated that the conversion of valine to acetonecyanohydrin (non-glycosylated form of linamarin) is catalysed by NADPH-dependent cytochrome P-450 (Koch et al., 1992). Such a cytochrome P-450enzyme (hemethiolate enzyme) was isolated from microsomes prepared frometiolated Sorghum seedlings too (Sibbesen et al., 1994) which gave rise to thespeci®c formation of a type I substrate binding spectrum in the presence of L-tyrosine. The initial step is the N-hydroxylation of valine followed by theformation of 2-methyl-propanal oxime and its dehydration to yield 2-methylpropionitrile. The addition of oxygen forms acetone cyanohydrin, which isthen glycosylated (by a soluble UDPG-glucosyltransferase) to form linamarin(Koch et al., 1992; Bokanga et al., 1993). The initial reactions take place onmembrane fractions, more recently it has been suggested that acetone cyanohydrin
JaÂnos Vetter / Toxicon 38 (2000) 11±3620

is probably synthesised by enzymes localised on the tonoplast. McMahon andSayre (1993) demonstrated that linamarin biosynthesis is NADPH-dependent. Thenew results suggest that the initial biosynthetic steps are localised on the tonoplastand the enzyme of glucosyltransfer is presumably vacuolar (Gruhnert et al., 1994).A vacuolar site for CG storage has also been demonstrated in Sorghum speciesand Hevea brasiliensis (Saunders and Conn, 1978; Gruhnert et al., 1994).Regarding the morphological localisation of the linamarin synthesis, Du et al.(1995) established that at least part of CGs are synthesised in the roots of cassava.In young cassava roots the CGs are more evenly distributed across the tubers. In
Fig. 2. Pathways of cyanogenic glycoside biosynthesis (dotted arrows indicate alternative pathways)
(according to Tapper and Reay, 1973).
JaÂnos Vetter / Toxicon 38 (2000) 11±36 21

old roots (12 months) the highest concentration is found primarily in thephelloderm and with a gradient of decreasing concentration of CG from thephelloderm towards the inner core. In etiolated seedlings of cassava, CGs aresynthesised in the cotyledons and transported to roots. In the mature plant, atransport of CGs from the leaves to the roots has been postulated. Thus, thebiosynthesis of linamarin is located both in the young root and in the green leavesof mature plants.
The term `cyanogenesis' means not only the synthesis or presence of an CG, butthe enzymatic hydrolysis producing free HCN and other compounds. Since noHCN is released from intact cyanogenic plants, the substrates (the CGs) and theenzymes must be located in di�erent compartments of cell. The generation ofcyanide from linamarin is a two-step process involving the initial deglycosilationof linamarin and the cleavage of linamarin to acetone cyanohydrin to formacetone and cyanide (Fig. 3). These reactions are catalysed by a b-glucosidase(linamarase) and by a-hydroxynitrile lyase (HNL). Since acetone cyanohydrin mayenzymatically as well spontaneously decompose, it has been generally assumedthat the linamarase activity is the rate-limiting step. The di�erent biochemicalproperties of linamarase of di�erent origin are summarised by McMahon et al.,1995. The linamarase of cassava consists of three di�erent isoforms, which di�erin their tissue speci®c localisation, and in their speci®c activities too. Thelinamarases of some plants have such broad substrate speci®cities that they aremore designated generalised b-glucosidases rather than linamarase (for examplethe b-glucosidase of Hevea brasiliensis is the only such enzyme present in the plant
Fig. 3. Cyanogenesis from linamarin (according to McMahon et al., 1995).
JaÂnos Vetter / Toxicon 38 (2000) 11±3622

and will hydrolyse a variety of b-linked glycones and aglycons (Tull et al., 1991;Yeoh and Yeoh, 1994). Most plants, however, have b-glucosidases with narrowsubstrate speci®city. New and important step was achieved with the isolation andsequencing of a cassava linamarase cDNA clone (Hughes et al., 1992). This DNAencodes a protein of 532 residues, having a predicted molecular mass of 65 kDa.Sequence analysis has demonstrated that the protein of cassava origin has 43%amino acid sequence identity with white clover linamarase, but is most similar tothe b-glucosidase of Agrobacterium tumefaciens. Analysis of the deduces aminoacid sequence of the cyanogenetic b-glucosidases from T. repens and M. esculentaand a non-cyanogenetic b-glucosidase from T. repens revealed considerablehomology between the three genes and with several cloned prokaryotic b-glucosidase enzymes (Hughes and Esen, 1993). The T. repens cyanogenic enzymewas extracellular whereas the enzyme was intracellular in cassava. Using immuno-gold localisation techniques, Mkpong et al. (1990) demonstrated that cassavalinamarase was localised in the cell wall, on the other hand, Pancoro and Hughes(1992) described that mRNA of linamarase was localised in laticifer cells. Thelatex has elevated linamarase activity, but probably accounts for only a fraction ofthe total leaf linamarase. The distribution of linamarase activity and linamarinvaries between di�erent organs and tissues of the same plant, in addition there aretissue speci®c di�erences in linamarase levels within the same plant organ. Theouter layer of the root cortex may have a 15-fold higher linamarase activity thanthe inner cells of root parenchyma. The distribution of linamarin in the roots isnot uniform, the root peel has a higher (12-fold) linamarin concentration than theinner parenchymatic tissue (Bradbury et al., 1991).
The second, i.e. the ®nal step of the cyanogenesis is the breakdown of acetoneto cyanide and acetone. This can occur both spontaneously (at temperaturesgreater than 358C or at pH greater than 4.0) and enzymatically catalysed byhydroxynitrile-liase (HNL). These enzymes have two subgroups (classes), thosethat contain ¯avins (FAD) and those that do not. The ¯avin-containing enzymesoccur in the family Rosaceae and have a higher speci®c activities and lower KM
values than the others. Cassava-HNL in contrast to HNL enzymes from otherplants has non-Michaelis±Menten rate kinetics (Hughes et al., 1994). The HNLshave been classi®ed into non-FAD and FAD-containing proteins. FAD-containingenzymes have been isolated exclusively from the family Rosaceae, whereas theFAD-independent ones, which are more heterogeneous in structure, have beencharacterised from various plant families (Euphorbiaceae, Linaceae, Oleaceae,Filitaceae, Poaceae: Hickel et al., 1996). The tissue and subcellular localisation ofHNLs were determined in Sorghum and Linum usitatissimum (Wajant et al., 1994),the highest level of the enzyme was found in the cytoplasm with lower levels inorganelles, and was not detected in the cell wall or in the vacuoles. A suicidalrelease of HCN in Sorghum as well as in ¯ax is prevented by separatecompartmentation of enzyme and substrate on the subcellular level. The twospeci®c b-glucosidases (amygdalin hydrolase and prunasin hydrolase) of the plumseeds are restricted to protein bodies of speci®c procambial cells and are absentfrom the cotyledonary parenchyma, bundle sheet and endosperm cells (Poulton
JaÂnos Vetter / Toxicon 38 (2000) 11±36 23
and Li, 1994). The distribution of CGs is di�erent, and thus the tissue levelcompartmentation prevents large scale CG hydrolysis in intact Prunus seeds.
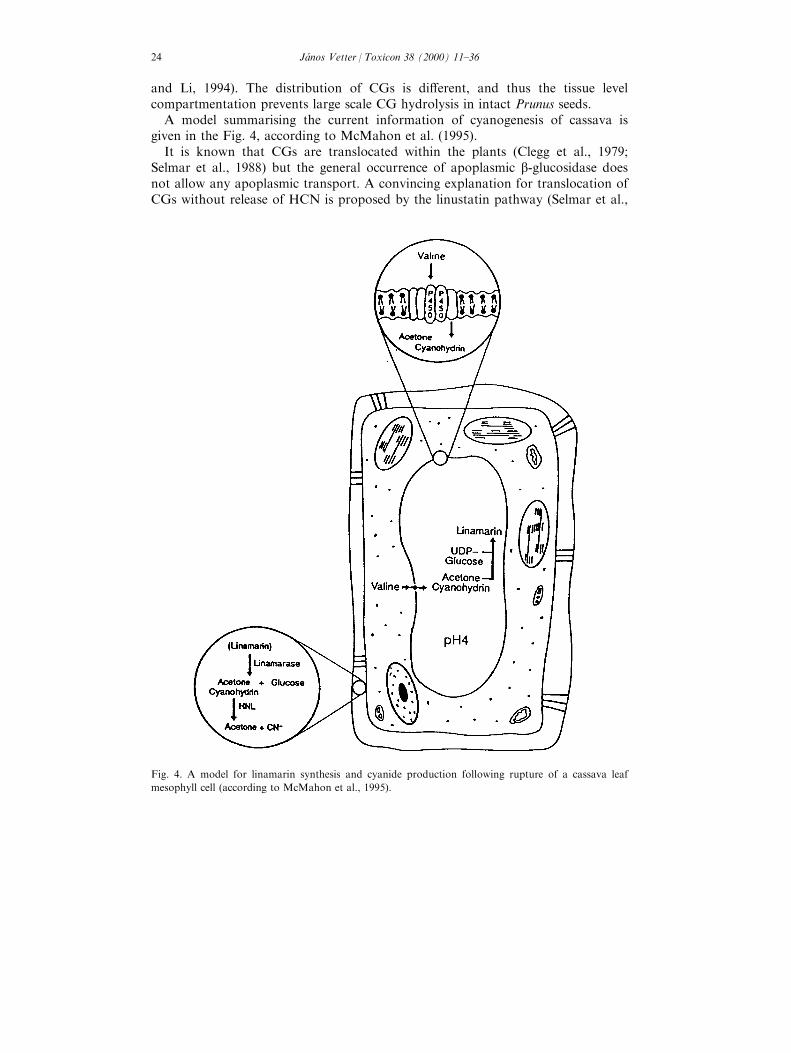
A model summarising the current information of cyanogenesis of cassava isgiven in the Fig. 4, according to McMahon et al. (1995).
It is known that CGs are translocated within the plants (Clegg et al., 1979;Selmar et al., 1988) but the general occurrence of apoplasmic b-glucosidase doesnot allow any apoplasmic transport. A convincing explanation for translocation ofCGs without release of HCN is proposed by the linustatin pathway (Selmar et al.,
Fig. 4. A model for linamarin synthesis and cyanide production following rupture of a cassava leaf
mesophyll cell (according to McMahon et al., 1995).
JaÂnos Vetter / Toxicon 38 (2000) 11±3624

1988) the cyanogenic monoglucoside (e.g. linamarin) is translocated as thecorresponding diglucoside which is produced by a glycosylation of themonoglucoside. In the seedlings of Hevea brasiliensis the diglucoside linustatinfunctions as a transport metabolite, as a transport form. From di�erent literaturedata arises the question of where the aglycone is glucosilated (in last step ofbiosynthesis) and how the synthesised CGs are accumulated in the vacuole. Whenglucosylation takes place in the cytosol, the produced glycosides must be activelytransported into the vacuole. The long-distance transport of linamarin couldproceed via linustatin to the sink. At the sink site, linustatin is deglycosylatedeither by a `simultaneous' diglycosidase (producing gentiobiose and acetonecyanohidrin), or it is sequentially deglycosylated to linamarin and then acetonecyanohydrin. In the simultaneous diglycosidase pathway, acetone cyanhidrin iseither converted to cyanide and acetone or reglycosylated by a UDP-glycosyltransferase to form linamarin. In the sequential deglycosilation pathway,linamarin may be stored at the sink or hydrolysed to acetone cyanohidrin. Thesimultaneous linustatin diglycoside pathway plays a role in nitrogen metabolism ofHevea seeds during germination. The linamarin of Hevea endosperm may functionas a mobilisable form of nitrogen for the growing seedlings, contrary in thecassava seedlings the asparagine and not linamarin is the major transportableform of N. There are con¯icting reports on the presence or absence of linamarinand linustatine in phloem of cassava according to Pereira and Splittstoesser (1987)was not found linamarin, according to Calatayud et al. (1994) was foundlinamarin. The later result suggests that linamarin can be transportedsymplastically.
6. Regulation of CG level in plants
The actual level of CGs of a genetically cyanogen plant is in¯uenced,determined by various factors both developmental (endogenous) and ecological(exogenous) ones. The whole development cycle of the cyanogenic plants showcharacteristic changes in their CG (and HCN) contents. The seeds of di�erentplant taxa contain di�erent, generally low concentration of CG. According to ourearlier work (Vetter and Haraszti, 1975) the seeds of Fabaceae (Trifolium species,Lotus corniculatus, Vicia species, Lathyrus sativus ) plants and the ¯ax haverelative higher (16±63 mg/kg fresh weight), the sudangrass and Sorghum cultivarslower (0±33 mg/kg fresh weight) concentration. During the ®rst day ofgermination (that is the phase of intensive water uptake, the imbibition) the CGlevel of fabaceous increases slightly, whereas the Sorghum group does not showchanges. The CG level of 4-day-old seedlings (i.e. in a very early phase of seedlingdevelopment) however shows di�erences. A relative unchanged level in the groupof fabaceous plants can be measured, but the plants of the Sorghum-group showeda sharp increase. The HCN content of the seedlings were 8±10-fold higher than inthe previous phase (in the 1-day `seedling'). These data indicate that importantchanges of the metabolism (biosynthesis) of CGs must take place during the ®rst
JaÂnos Vetter / Toxicon 38 (2000) 11±36 25
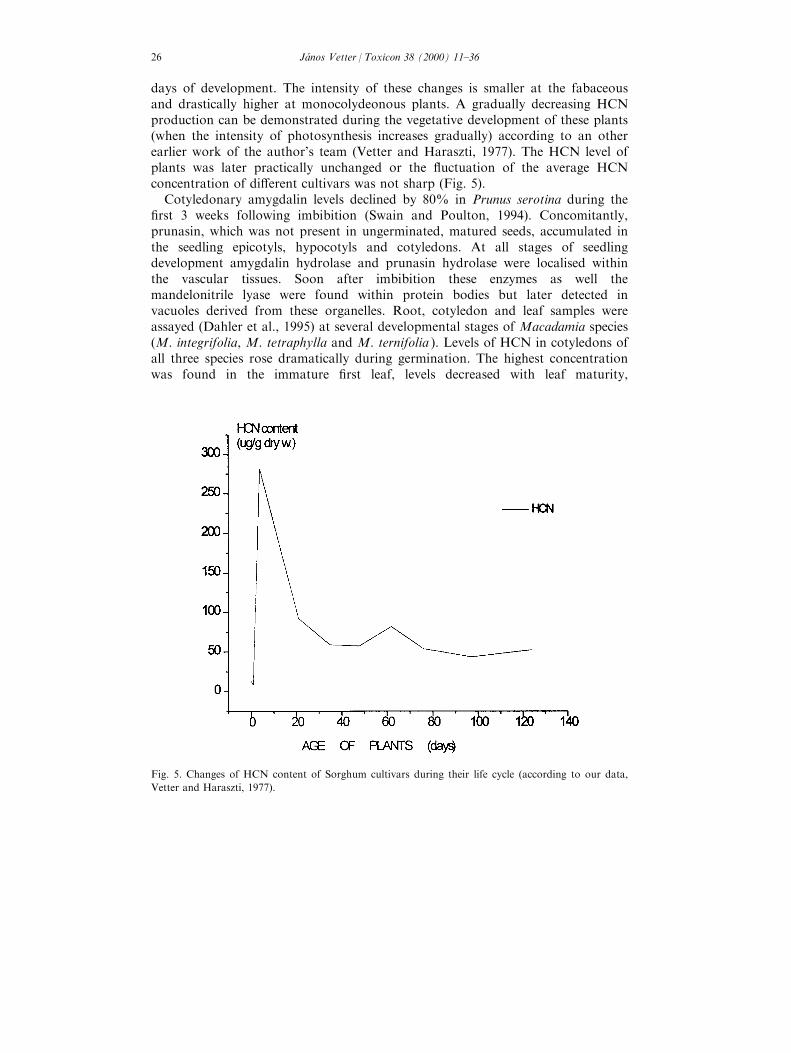
days of development. The intensity of these changes is smaller at the fabaceousand drastically higher at monocolydeonous plants. A gradually decreasing HCNproduction can be demonstrated during the vegetative development of these plants(when the intensity of photosynthesis increases gradually) according to an otherearlier work of the author's team (Vetter and Haraszti, 1977). The HCN level ofplants was later practically unchanged or the ¯uctuation of the average HCNconcentration of di�erent cultivars was not sharp (Fig. 5).
Cotyledonary amygdalin levels declined by 80% in Prunus serotina during the®rst 3 weeks following imbibition (Swain and Poulton, 1994). Concomitantly,prunasin, which was not present in ungerminated, matured seeds, accumulated inthe seedling epicotyls, hypocotyls and cotyledons. At all stages of seedlingdevelopment amygdalin hydrolase and prunasin hydrolase were localised withinthe vascular tissues. Soon after imbibition these enzymes as well themandelonitrile lyase were found within protein bodies but later detected invacuoles derived from these organelles. Root, cotyledon and leaf samples wereassayed (Dahler et al., 1995) at several developmental stages of Macadamia species(M. integrifolia, M. tetraphylla and M. ternifolia ). Levels of HCN in cotyledons ofall three species rose dramatically during germination. The highest concentrationwas found in the immature ®rst leaf, levels decreased with leaf maturity,
Fig. 5. Changes of HCN content of Sorghum cultivars during their life cycle (according to our data,
Vetter and Haraszti, 1977).
JaÂnos Vetter / Toxicon 38 (2000) 11±3626

correlating with leaf toughening. Other investigations were carried out at theLotus corniculatus cultivars (Borsos et al., 1976). The average changes of HCNconcentration showed that the characteristic level of the phase of vegetativegrowth decrease till the beginning of ¯owering, than increase till the whole¯owering, and it is especially high in the second growth (in the growing plantparts after the cutting). These fresh plants, plant parts show signi®cant or extremehigh values, the highest ones of the whole life cycle of Lotus corniculatus. It wasan important practical conclusion that some cyanogenic forage plants can achievethe highest level (which is or can be poisonous for the consuming animals) in thephase of the intensive growth (whereas the cells are in the enlargement phase andhave very intensive metabolic activities). Other investigations revealed the phase ofseed- and fruit development. Ohtsubo and Ikeda (1994) found that the total CGcontent increased throughout seed development in apricot. In the early stages, theovule contained relative high amount of prunasin which disappeared at the onsetof stage II of fruit development. Concurrently, amygdalin attained a maximum inthe embryo maturity, whereas the prunasin concentration in the endosperm islower. The ®rst slow growth period of the fruit coincided with the decrease inendosperm prunasin content and the rapid increase in embryo amygdalin content.
The regulating role of di�erent ecological (exogenous) factors on the CG(HCN) level of plants has been known since many years. Some such results werepublished in the `classical' physiological works of the sixties±seventies. Nutritionalfactors (®rst of all the N), temperature (or sudden change of temperature) havethe most important e�ects on the actual level of CG (or HCN). According to a®eld experiment of El-Essawi et al. (1995) the increasing NP fertiliser levels led tosigni®cant increase in HCN content of Sorghum (cv. Pioneer 988).
7. Biological e�ects of CGs
7.1. Role in plant physiological processes
Production of HCN by both rice and cocklebur (Xanthium pennsylvanicum )increased during a pregermination period and preceded ethylene production(Hasegawa et al., 1994). Its concentration declined abruptly when the radiclesemerged before the peak in ethylene production. That HCN production duringinitial imbibition may be derived from cyanogenic reserves and controlled by bothpreexisting and subsequently developing beta-cyanoalanine synthase. Applicationof ethylene stimulates the activity of this enzyme and reduced their cyanogencontent. In other experiments (Hasegawa et al., 1995) the addition of cysteingreatly reduced the CG content of seeds, but increased the HCN evolution. Esashiet al. (1996) established that some nitrogenous inhibitors of respiration (namelyKCN, NaN3, etc.), which promoted the germination of Xanthium pennsylvanicumseeds, enhanced the accumulation of cyanogen compounds. Part of theexogenously applied KCN was converted to CG and it is suggested that
JaÂnos Vetter / Toxicon 38 (2000) 11±36 27

endogenous cyanogens might be involved in the germination of Xanthium seeds.The promoting e�ect of KCN on germination was con®rmed by Maruyama et al.(1996).
7.2. Role in plant defence mechanism
One of the most probable explanations of the biological role of CGs in someplants is the participation in defence mechanisms against di�erent phytopatogens,despite that both plants and animals posses the ability to detoxify cyanide. So thepresence of CGs in plants is not necessarily inimical to herbivory (larvae ofSpodoptera eridania prefer to graze on CG-containing plants and grow betterwhen cyanide is present in their diet: Brattsen et al., 1983). Studies have shownthat CGs can act as either feeding deterrents or phagostimulants, depending onthe insect species. Berenbaum and Zangerl (1991) quote research that both doesand does not implicate the presence or absence of cyanogenesis with feedingpreference and behaviour in molluscs. Malagon and Garrido (1990) concludedthat bitter almond plants are resistant to larvae of buprestid Capnodis tenebrionis,owing to the high concentration of CGs. The larvae of Hypera postica(Coleoptera, Curculionidae) preferred the acyanogenic lea¯ets of Trifolium repenscv. `Louisiana S-1' as well cv. `Regal' (Ellsbury et al., 1992) suggesting that thelevels of CGs could be selected in T. repens to improve resistance to insect pests.Fourteen Sorghum varieties were investigated for their susceptibility to ¯y speciesAtherigona soccata and Chilo partellus (Alborn et al., 1992). The CG dhurrin wasin greater quantities in susceptible cultivars CSH-1, Swarna and IS10795, and itwas suggested that dhurrin acts as an oviposition activator for the pests. Clearexamples of the protective e�ect of cyanogenesis do exist, even against apparentlyspecialist pests. Thus the CG of cassava tubers improved its resistance to thecassava root borer (Bellotti and Arias, 1993).
7.3. E�ects on animals
All cyanogenic glycosides have a potential danger because of the production ofHCN via hydrolysis (either by spontaneous or by enzimatically regulatedreactions). A lot of investigations, practical data or experiments were published indi�erent journals of veterinary sciences, of toxicology or others. Summarised, theconclusions can establish the general toxicity of cyanide and the cyanide (CGs)containing plants, but there are di�erences in the sensitivity of di�erent animalspecies, in the poisonous (and in lethal) doses of plants (in the actualconcentrations of CGs or cyanide), in the rate of production of HCN from CGsand in other facts.
Di�erent parts (the lower stems, leaves and racemes) of Prunus padus (birdcherry) was eaten by three aged cows (Sargison et al., 1996). The animals showedsign of anorexia, weakness, depression, stupor, circling, bruxism, excessivesalivation and tenesmus. Clinical examination established ruminal stasis, brightred mucous membranes, tachycardia with cardiac dysrhytmia, tachypnoe and
JaÂnos Vetter / Toxicon 38 (2000) 11±3628

scant tarry faeces. All animals became recumbent with generalised fasciculationand died within 10 h of onset of clinical sings. PM ®nding were consistent with adiagnosis of cyanide poisoning, acute hepatocyte degeneration may have been dueto P. padus, the whole blood cyanide concentration was 0.53 mg/L. Anexperimental poisoning was induced by the fresh leaves of Prunus sellowii (Gava etal., 1992). The symptoms of these animals were tachycardia, rumen atony,muscular tremors, instability, di�culty in breathing, sternal or lateral decubitus,and in some cases, death. The plant Mimosa invisa var. inermis St. Adalbertcaused a total lethal poisoning of 22 swamp bu�aloes in Thailand(Tungtrakanpoung and Rhienpanish, 1992). The animals were dead 18±36 h aftereating, the symptoms included salivation, sti�ness, lack of mastication, musculartremor, dyspnoe and recumbency, the lesions revealed congestion and petechialhaemorrhage of heart, liver, kidneys, lungs, rumen and intestines. Jackson (1995)demonstrated the sudden death of two donkeys in New York after intake of thewild cherry, Prunus virginiana, although no signi®cant gross or histopathologicallesions were found in either donkeys.
The `sudden death' syndrome of sheep induced by Phalaris aquatica was studiedby Bourke and Carrigan (1992). Potentially toxic levels of HCN (20±36 mg/100 g)were measured in the three toxic Phalaris pastures tested. It was suggested thatPhalaris `sudden death' syndrome could have as many as four di�erent underlyingmechanisms and that these might re¯ect the presence of cyanogenic and of othercompounds (a cardio-respiratory toxin, a thiaminase and amine cosubstrate andnitrate compounds). Gajendragad et al. (1992) examined the pathology of brain inacute HCN poisoning in sheep. Shortly after eating pods of an Acacia sp. tree 50sheep showed signs of acute cyanide poisoning (dyspnoe, salivation andrecumbency) and some died. PM examination showed severe congestion of all thevisceral organs. The blood was unclotted and dark red in colour. The neuroneswere darkly stained, their nucleus marginated with various degree of degeneration,the Nissl substance had degraded and central cromatolysis was found. Nucleardegenerations occurred in glial cells. The microscopic changes observed in thecerebellum varied from congestion, nuclear degeneration in Purkinje cells andGolgi cells, and central chromatolysis in the motor neurones, the nerve ®bres wereswollen. The histological lesions seen were similar to anoxic type of lesion Theselenium status and the thyroid function of sheep was studied by Gutzwiller(1993) under the e�ect of cyanogenic glycosides containing diet. The animals hadeaten a white clover/grass silage with high and with low content of CGs duringthe last 69±97 days of pregnancy. Results show that the intake of CGs a�ects Sestatus of sheep and increases the susceptibility of lambs to nutritional myopathyif the Se content of the diet is marginal. Intake of CG presumably had noor at most only a slight e�ect on thyroid function, presumably because bodyiodine reserves of ewes were su�cient to o�set the goitrogenic e�ect ofthyocyanate.
The intake of leaves of Manihot glaziovii (cearra rubber) increased signi®cantlythe white blood cell count of goats in Sri Lanka (Jayasekara and Horadagoda,1992).
JaÂnos Vetter / Toxicon 38 (2000) 11±36 29

The possibility of HCN (or CG) poisoning in horses is a slightly contradictoryquestion. Hintz (1993) concluded that the danger of cyanide poisoning of horsesgiven seed of Linum usitatissmium is slight. The intake of Sorghum vulgare can bemore problematical. After being fed for over 2 months on Sorghum bicolor (S.vulgare ) three of 11 horses showed incordination of the hind legs, 7±10 daysdeveloped frequent urination, urinary incontinence and haematuria, followed by aserious nasal discharge, increased body temperature, depression and reducedappetite (Varshney et al., 1996). Landrace X Large White wenare pigs were givendiets containing HCN 0, 239 and 419 mg/kg to study the e�ects of low and highcyanide content in cassava on animals (Iyayi, 1994). Weight gain, feed intake feedconversion e�ciency and protein e�ciency were not a�ected. Serum albumin andtotal protein were not di�erent, but serum urea increased signi®cantly withincreasing cyanide intake.
Panigraki et al. (1992) examined the e�ect of fast- and slow-dried meal ofcassava on broiler chicks. Growth rate of animals fed on slow-dried meal weresigni®cantly higher than those of chicks fed on fast-dried meal. Thus, slower ratesof drying cassava roots produce meals with lower cyanogen concentration thatare, consequently, less toxic to broiler chicks. The level of cassava meal over 10±15% in the diet of chicks will reduce gains, but adult hens seems to be quitetolerant and maintain egg production, even when fed tapioca leaves with a highHCN content.
Nutritional experiments were carried out with a cassava-containing diet for dogs(Ibebunjo et al., 1992; Kamalu, 1993). The gari diet increased the plasmathiocyanate concentration, raised 24 h thiocyanate excretion and increased urinaryprotein excretion, lowered serum albumin. The e�ect of linamarin ondiabetogenicity of dog was studied (Ragoobirsingh et al., 1993). In the linamarintreated undernourished dogs plasma insulin and insulin binding to erythrocytesand mononuclear leukocytes were signi®cantly lower than in control dogs and thisresulted in abnormally high glucose values.
7.4. E�ects on human organism
The investigations on the biological e�ect of CGs on human organism were andare focused on some species, ®rst of all on M. esculenta (cassava). The cassavaroot is an important dietary staple in a lot of tropical countries. In both Africaand Latin America, cassava is the second or the third most important dietaryenergy source for more than 500 million people (Boccas, 1987). The CGs, i.e.linamarin and lotaustratalin, are presented in all tissue of the plant, they canundergo hydrolysis producing the poisonous HCN. The cassava root with a highcyanogenic content is processed to remove the glycosides and to convert it into astorable form such as gari. The areas in which the kwashiorkor is commoncoincide with areas of intensive human su�ering of M. esculenta; furthermore, thedisease may be found in countries other than those in which cassava is eaten, forexample in North America (Rossouw, 1989) or in di�erent countries of Europe.To the observed pathological features belongs the cellular oedeme. Kamalu (1993)
JaÂnos Vetter / Toxicon 38 (2000) 11±3630

found periportal vacuolation of the liver, swelling, vacuolation and rupture of theepithelial cells of the proximal convoluted tubules of the kidney in the gari-fedexperimental dogs. The swelling and vacuolation (cellular oedema) observed was adirect consequence of linamarin acting as a glycoside. Proteinuria occurred asearly as the end of week 1 of the experiment, thus the kidney was alreadydamaged and nephrosis was presented. Kamalu (1993) observed congestion andhaemorrhage in the experimental animals, degeneration and swelling of themyocardial ®bres resulted the generalised chronic passive congestion. Impairedglucose tolerance and reduced or absent insulin response to glucose have beendemonstrated by various authors in children with kwashiorkor (Crace et al.,1990). Experiments with dogs demonstrated (Kamalu, 1991) that the states ofpotassium depletion and hypocalcemia did play a role in the reduces insulin level.Biochemical examination (Kamalu, 1991) revealed an abnormal elevation inplasma activity of pancreatic lipase and haemorrhage, necrosis, ®brosis andatrophy of pancreas. According to the new hypothesis of Kamalu (1993),linamarin absorbed from cassava diets, causes inhibition of Na±K±ATPase, givingrise to electrolyte imbalance with potassium depletion. This depletion causescellular swelling, vacuolation and rupture of the epithelial cells of the proximaltubules, which results in proteinuria and causes low serum albumin concentration.
Several cases of acute cassava poisoning have been reported in Nigeria since1989 (Akintonwa et al., 1994). Eight patients died shortly after eating a meal ofgari, a grated or fried cassava product. They vomited and complained ofabdominal pain immediately after the meal, they became comatose with acuterenal failure and died of cardio-pulmonary arrest.
In Nigerians, a syndrome referred to as tropical ataxic neuropathy compriseslesions of skin, mucous membranes, optic and auditory nerves, spinal cord andperipheral nerves. The disease a�ected all age groups, patients subsisted mainly ona cassava diet, the epidemiological studies showed correlation of prevalence of thedisease with intensity of cassava cultivation and consumption. Theneuropathology of the disease would be compatible with the e�ects of chroniccyanide intoxication (Osuntokun, 1973).
Cassava contains a potential goitrogen agent that may aggravate iodinede®ciency disorders (Delange et al., 1994), that have goitre and cretinism. Themain part of the dietary cyanide will be converted to thyocyanate, its e�ectmimics entirely that of iodine de®ciency. The goitrogenic e�ect of cassava can becorrected by an increasing intake of iodine. Experiments on thyocianate values inhuman serum and urine samples indicate that conversion of cyanide tothyocianate is a signi®cant pathway in the metabolism of HCN (Eminedoki et al.,1994). Research in Liberia and other parts of humid tropical Africa (Jackson,1994) suggests that a diet containing cassava may in¯uence the biological ®tnessand distribution of certain haemoglobin genes, alter basic metabolic processessuch as gluconeogenesis, enzyme function, the activity of thyroid hormone anddecelerates the usual patterns of human growth and development. The highphenotypic and genotypic variability observed among Africans may re¯ect thepotentially dramatic in¯uence of chronic dietary cassava use.
JaÂnos Vetter / Toxicon 38 (2000) 11±36 31

References
Aikman, K., Bergman, D., Ebinger, J., Seigler, D., 1996. Variation of cyanogenesis in some plant
species of the Midwestern United States. Biochemical Systematics and Ecology 24, 637±645.
Alborn, H., Stenhagen, G., Leuschner, K., Rizvi, S.J.H., 1992. Biochemical selection of sorghum crop
varieties resistant to sorghum shoot ¯y (Atherina soccata ) and stem borer (Chilo partellus ): role of
allelochemicals. In: Rizvi, V. (Ed.), Allelopathy: Basic and Applied Aspects. Chapman and Hall
Ltd, London, pp. 101±117.
Akintonwa, A., Tunwashe, O., Onifade, A., 1994. Fatal and non-fatal acute poisoning attributed to
cassava based meal. Acta Holticulturae 375, 285±288.
Amarowicz, R., Shahidi, F., 1994. Application of Sephadex LH-20 chromatography for the separation
of cyanogenic glycosides and hydrophylic phenolic fraction from ¯axseed. Journal of Liquid
Chromatography 17, 1291±1299.
Association of O�cial Agricultural Chemists, 1965. In: AOAC o�cial methods of analysis, 10th ed., p.
341.
Belloti, A.C., Arias, B., 1993. The possible role of HCN on the biology and feeding behaviours of the
cassava burrowing bug (Cyrtomenus bergi, Froeschner). In: Roca, W.M., Thro, A.M. (Eds.),
Proceedings of the 1st International Scienti®c Meeting of the Cassava Biotechnology Network. CIA,
Coli USA, pp. 406±409.
Berg van den, A.J.J., Horsten, S.F.A.J., Kettenes van den Bogsch, J.J., Kroes, B.H., Labadie, R.P.,
1995. Multi®din a cyanoglucoside in the latex of Jatropha multi®da. Phytochemistry 40, 597±598.
Berenbaum, M.R., Zangerl, A.R., 1991. Genetics of secondary metabolism and herbivore resistance in
plants. In: Rosenthal, G.A., Berenbaum, M.R. (Eds.), Herbivores, their Interactions with Secondary
Plant Metabolits. Academic Press, San Diego, pp. 415±438.
Borsos, O., Haraszti, E., Vetter, J., 1976. Change of cyanogenetic glycoside content of some cvars. of
Lotus corniculatus during vegetation period. Botanikai KoÈ zleme nyek 63, 63±71.
Boccas, B., 1987. Cassava staple food crop of prime importance in the tropics. Courier 101, 72±73.
Bokanga, M., Halkier, B., Moller, B., 1993. Studies on biosynthesis of cyanogenic glucosides in cassava.
In: Roca, W.M., Thro, A.M. (Eds.), Proceedings of the 1st International Scienti®c Meeting of the
Cassava Biotechnology Network. Coli, USA, pp. 418±423.
Bourke, C.A., Carrigan, M.J., 1992. Mechanism underlaying Phalaris aquatica `sudden death' syndrome
in sheep. Australian Veterinary Journal 69, 165±167.
Boyd, F.T., Aamodt, O.S., Bohestedt, G., Troug, E., 1938. Sudangrass managements for control of cya-
nide poisoning. Journal American Society Agronomy 30, 569±582.
Bradbury, J.H., Egan, S.V., Lynch, M.J., 1991. Analysis of cyanide in cassava using acid hydrolysis of
cyanogenic glycosides. Journal of Science and Food Agriculture 55, 277±290.
Brattsen, L.B., Samuelian, J.H., Long, K.Y., Kincaid, S.A., Evans, K., 1983. Cyanide as a feeding
stimulant for the southern armyworm, Spodoptera eridania. Ecological Entomology 8, 125±132.
Calatayud, P.A., Rakbe, Y., Delobel, B., Khuong, H.F., Tertuliano, M., Le Ru, B., 1994. In¯uence of
secondary compounds in the phloem sap of cassava on expression on antibiosis towards the mealy-
bug Phenacoccus manihoti. Entomology Experimental Application 72, 47±57.
Caradus, J.R., Forde, M.B., 1996. Characterization of white clover populations collected from the
Caucasus and high altitude regions of eastern Turkey. Genetic Resources and Crop Evolution 43,
143±155.
Cardona, L., Fernandez, I., Pedro, J.R., Vidal, R., 1992. Polyoxygenated terpenes and cyanogenic glu-
cosides from Centaurea alpera var. subinermis. Phytochemistry 31, 3507±3509.
Carlson, I.T., 1958. Inheritance of hydrocianic acid production in crosses between sorghum and sudan-
grass. Agronomy Journal 50, 302±306.
Chadha, R.K., Lawrence, J.F., Ratnayake, W.M.N., 1995. Ion chromatographic determination of cya-
nide released from ¯axseed under autohydrolisation. Food Additives and Contaminants 12, 527±
533.
Chang, J.Y.C., Hwang, L.S., 1993. Study on quantitative determination of taxiphyllin in bamboo
shoots. Journal of the Chinese Agricultural Chemical Society 31, 506±517.
JaÂnos Vetter / Toxicon 38 (2000) 11±3632

Clegg, D.O., Conn, E.E., Janzen, D.H., 1979. Developmental fate of the cyanogenic glucoside linamarin
in Costa Rica wild Lima bean seeds. Nature 278, 343±344.
Cook, J.H., 1985. Cassava: New Potential for a Neglected Crop. West®eld Press, London 191 pp.
Corkill, L., 1942. Cyanogenesis in white clover. 5. The inheritance of cyanogenesis. New Zealand
Journal Science Technology (Ser. B) 23, 178±193.
Chassagne, D., Crouzet, J.C., Bayonove, C.L., Baumes, R.L., 1996. Identi®cation and quanti®cation of
passion fruit cyanogenic glucosides. Journal of Agriculture and Food Chemistry 44, 3817±3820.
Crace, C.J., Swenne, I., Kohn, P.G., Strain, A.J., Milner, R.D.G., 1990. Protein-energy malnutrition
induces changes in insulin sensitivity. Diabete et Metabolisme 16, 484±491.
Dahler, J.M., McConchie, C.A., Turnbull, C.G.N., 1995. Quanti®cation of cyanogenic glucosides in
seedling of three Macadamia (Proteaceae) species. Australian Journal of Botany 43, 619±628.
Delange, F., Ekpechi, L.O., Rosling, H., 1994. Cassava cyanogenesis and iodine de®ciency disorders.
Acta Horticulturae 375, 289±293.
Du, L.C., Bokanga, M., Moller, B.L., Halkier, B.A., 1995. The biosynthesis of cyanogenic glucosides in
roots of cassava. Phytochemistry 39, 323±326.
El-Essawi, T.M., Mashali, S.A., Kanany, R., 1995. E�ect of balanced manuring on Sorghum growth
and increasing utilization of nutrients. Egyptian Journal of Soil Science 35, 253±264.
Ellsbury, M.M., Pederson, G.A., Fairbrother, T.E., 1992. Resistance to foliar feeding hipergine weerils
(Coleoptera, Curculionidae) in cyanogenic white clover. Journal of Economic Entomology 85, 2467±
2472.
Eminedoki, D.G., Monann, M.O., Anosike, E.O., 1994. Thyocyanate levels of mainly dietary origin in
serum and urine from a human population sample in Port Harcourt, Nigeria. Plant Foods for
Human Nutrition 46, 277±285.
Esashi, Y., Maruyama, A., Sasaki, S., Tani, A., Yoshiyama, M., 1996. Involvement of cyanogens in the
promotion of germination of cocklebur seeds in response to various nitrogenous compounds, inhibi-
tors of respiratory (respiration) and ethylene. Plant and Cell Physiology 37, 545±549.
Essers, S.A.J.A., Bosveld, M., Grift van der, R.M., Voragen, S.A.G.J., 1993. Studies on quanti®cation
of speci®c cyanogenesis in cassava products and introduction of a new chromogen. Journal of the
Science of Food and Agriculture 63, 287±296.
Gajendragad, M.R., Gopalakvishna, S., Ravikumar, S.B., 1992. Pathology of the brain in acute hydro-
cyanic acid poisoning in sheep. Indian Veterinary Journal 69, 206±210.
Gava, A., Stolf, L., Neves, D.S., Stolf, P., Varaschim, M.S., Fereira, E.M.M., 1992. Experimental poi-
soning of cattle by Prunus sellowii (Rosaceae). Pesquisa Veterinaria Brasileira 12, 1±4.
Gillingham, J.F., Shiver, M.M., Page, N.R., 1969. Evaluation of the Orion cyanide electrode for esti-
mating the cyanide content of forage samples. Agronomy Journal 61, 717±718.
Gilchrist, D.G., Lueschen, W.E., Hittle, C.N., 1967. Revised method for the preparation of standards
in the sodium picrate assay of HCN. Crop Science 7, 267±268.
Gruhnert, C.H., Biehl, B., Selmar, D., 1994. Compartmentization of cyanogenic glucosides and their
degrading enzymes. Planta 195, 36±42.
Guignard, M.L., 1906. Le haricot a acide cianhydrique, Phaseolus lunatus L., Paris. Compt. Rend.
Paris 142, 545±553.
Gutzwiller, A., 1993. The e�ect of a diet containing cyanogenic glycosides on the selenium status and
the thyroid function of sheep. Animal Production 57, 415±419.
Hasegawa, R., Maruyama, A., Sasaki, H., Tada, T., Eashi, Y., 1995. Possible involvement of ethylene-
activated beta-cyanoalanine synthase in the regulation of cocklebur seed germination. Journal of
Experimental Botany 46, 551±556.
Hasegawa, R., Tada, T., Torii, Y., Eashi, Y., 1994. Presence of beta-cyanoalanine synthase in unim-
bibed dry seeds and its activation by ethylene during pregermination. Physiologia Plantarum 91,
141±146.
Hickel, A., Hasslacher, M., Griengl, H., 1996. Hydroxynitrile lyases: functions and properties.
Phisiologia Plantarum 98, 891±898.
Hin, Y.H., Egan, S.V., Yeoh, H.H., 1997. An enzyme-based dip-stick for the estimation of cyanogenic
potential of cassava ¯our. Food Chemistry 60, 119±122.
JaÂnos Vetter / Toxicon 38 (2000) 11±36 33

Hin, Y.H., Lim, L.S., Woo, H.C., Yeoh, H.H., 1996. An enzyme-bound linamarin indicator paper strip
for the semi-quantitative estimation of linamarin. Biotechnology Techniques 10, 319±322.
Hintz, H.F., 1993. Flaxseed: friend or foe. Equine Practice 15, 6±9.
Hughes, M.A., Brown, K., Pancoro, A., Murray, B.S., Hughes, J., 1992. A molecular and biochemical
analysis of the structure of the cyanogenic-glucosidase (linamarase) from cassava (Manihot esculenta
Crantz). Archives of Biochemistry and Biophysics 295, 273±279.
Hughes, J., Carvalho, F.J.P., De, C., Hughes, M.A., 1994. Puri®cation characterization and cloning of
a-hydroxynitrile lyase from cassava (Manihot esculenta Crantz). Archives of Biochemistry and
Biophysics 311, 496±502.
Hughes, M.A., Esen, A., 1993. Molecular genetics of plant cyanogenic beta-glucosidases. Beta-glucosi-
dases: Biochemistry and Molecular Biology. In: Symposium Sponsored by the Division of
Agricultural and Food Chemistry at the 204th National Meeting of the American Chemical Society,
Washington DC, August 23±28, 1992. ACS symposium series, vol. 533, pp. 153±169.
Ibebunjo, C., Kamalu, B.P., Ihemelandu, E.C., 1992. Comparison of the e�ects of cassava (Manihot
esculenta Crantz) organic cyanide and inorganic cyanide on muscle and bone development in a
Nigerian breed of dog. British Journal of Nutrition 68, 483±491.
Iyayi, E.A., 1994. Supplement e�ects of low- and high cyanide cassava peels on the performance, nutri-
ent digestibility and serum metabolites of growing pigs. Tropenlandwirt 95, 197±205.
Jackson, F.L., 1994. The bioantropological impact of chronic exposure to sublethal cyanogens in
Africa. Acta Horticulturae 375, 295±309.
Jackson, T., 1995. Cyanide poisoning in two donkeys. Veterinary and Human Toxicology 37, 567±568.
Jager, A.K., McAlister, B.C., Staden van, J., 1995. Cyanogenic glycosides in leaves and callus cultures
of Schlechterina mitostemmatoides. South African Journal of Botany 61, 274±275.
Jayasekara, S., Horadagoda, N.U., 1992. Hydrocianic acid content in Manihot glaziovii (cearra rubber)
leaves and toxicologic e�ects in goats. Sri Lankan Journal of Agricultural Sciences. 29, 9±15.
Kakes, P., 1997. Di�erence between the male and female components of ®tness associated with the
gene Ac in Trifolium repens. Acta Botanica Nederlandica 46, 219±223.
Kamalu, B.P., 1991. The e�ect of a nutritinally balanced cassava (Manihot esculenta Crantz) diet on en-
docrine function using the dog as a model. 1. Pancreas. British Journal of Nutrition 65, 365±372.
Kamalu, B.P., 1993. Pathological changes in growing dogs fed on a balanced cassava (Manihot escu-
lenta Crantz) diet. British Journal of Nutrition 69, 921±934.
Kawamura, Y., Hikidi, S., Maruyama, K., Uchiyama, S., Saito, Y., 1993. Improvement of the direct
determination method of linamarin in beans and bean paste products. Journal of the Food Hygienic
Society of Japan 34, 74±79.
Kobaisy, M., Oomah, B.D., Mazza, G., 1996. Determinations of cyanogenic glycosides in ¯axseed by
barbituric acid pyridine, pyridine-pyrazolone and high performance liquid chromatography methods.
Journal of Agricultural and Food Chemistry 44, 3178±3181.
Koch, B., Nielsen, V.S., Olsen, C.E., Moller, B.L., 1992. The biosynthesis of cyanogenic glucosides in
seedlings of cassava (Manihot esculenta Crantz). Archives of Biochemistry and Biophysics 292, 141±
150.
Makkar, H.P.S., Becker, K., 1997. Nutrients and antiquality factors in di�erent morphological parts of
the Moringa oleifera tree. Journal of Agricultural Science 128, 311±322.
Malagon, J., Garrido, A., 1990. Relation between cyanogenic glycosides content and the resistance to
Capnodis tenebrionis (L.) in stone fruits. Boletin de Sanidad Vegetal 16, 499±503.
Mallika, G.V., Jausz, E.R., Pieris, N.M., Abeysekara, A.M., 1991. The cyanogenic glycosides and glu-
cosidases of rubber seed kernel. Journal of the National Science Council of Sri Lanka 19, 99±106.
Maruyama, A., Yohiyama, M., Adachi, Y., Tani, Hasegawa, R., Esashi, Y., 1996. Promotion of cockle-
bur seed germination by allyl, sulfur and cyanogenic compounds. Plant and Cell Physiology 37,
1054±1058.
McFarlane, I.J., Lees, E.M., Conn, E.E., 1975. The in vitro biosynthesis of dhurrin, the cyanogenic glu-
coside of Sorghum bicolor. Journal Biological Chemistry 250, 4708±4713.
McMahon, J.M., Sayre, R.T., 1993. Di�erential biosynthesis and transport of linamarin in high- and
low-cyanide cultivars of cassava (Manihot esculenta Crantz). In: Roca, W.M., Thro, A.M. (Eds.),
JaÂnos Vetter / Toxicon 38 (2000) 11±3634

Proceedings of the 1st International Scienti®c Meeting of the Cassava Biotechnology Network.
Centro International de Agricultura Tropical, Cali, Columbia, pp. 376±378.
Mcmahon, J.M., White, W.L.B., Sayre, R.T., 1995. Cyanogenesis in cassava (Manihot esculenta
Crantz). Journal of Experimental Botany 46, 731±741.
Mkpong, O.E., Yan, H., Chism, G., Sayre, R.T., 1990. Puri®cation, characterization and localization
of linamarase in cassava. Plant Physiology 93, 176±181.
Nakanishi, T., Nishi, M., Somekawa, M., Murata, H., Mzuno, M., Iinuma, M., Tanaka, T., Murata,
J., Lang, F.A., Inada, A., 1994. Structures of new and known cyanoglucosides from a North
American plant, Purshia tridentata DC. Chemical and Pharmaceutical Bulletin 42, 2251±2255.
Nass, H.G., 1972. Cyanogenesis: its inheritance in Sorghum bicolor, Sorghum sudanense, Lotus and
Trifolium repens: a review. Crop Science 12, 503±506.
Noitsakis, B., Jacquard, P., 1992. Competition between cyanogenic and acyanogenic morphs in
Trifolium repens. Theoretical and Applied Genetics 83, 443±450.
Ohtsubo, T., Ikeda, F., 1994. Seasonal changes of cyanogenic glycosides in mume (Prunus mume Sieb.
Et Zucc.). Journal of Japanese Society for Horticultural Science 62, 695±700.
Oomah, B.D., Mazza, G., Kenaschuk, E.O., 1992. Cyanogenic compounds in ¯axseed. Journal of
Agricultural and Food Chemistry 40, 1346±1348.
Osuntokun, B.O., 1973. Ataxic neuropathy associated with high cassava diets in West Africa. In:
Nestel, B., MacIntyre, R. (Eds.), Chronic Cassava Toxicity. International Development Reseach
Centre, Ottawa, Canada, pp. 127±138.
Pancoro, A., Hughes, M.A., 1992. In situ localization of cyanogenic-glucosidase (linamarase) gene ex-
pression in leaves of cassava (Manihot esculenta Crantz) using non-isotopic riboprobes. The Plant
Journal 2, 821±827.
Panigraki, S., Rickard, J., O'Brien, G.M., Gay, G., 1992. E�ects of di�erent rates of drying cassava
root on its toxicity to broiler chicks. British Poultry Science 33, 1025±1041.
Pederson, G.A., Faitbrother, T.E., Greene, S.L., 1996. Cyanogenesis and climatic relationships in US
white clover germplasm collection and core subset. Crop Science 36, 427±433.
Pereira, J.F., Splittstoesser, W.E., 1987. Exudate from cassava leaves. Agricultural Ecosystems and
Environment 18, 191±194.
Poulton, J.E., Li, C.P., 1994. Tissue level compartmentation of (R )-amygdalin and amygdalin hydrolase
prevents large-scale cyanogenesis in undamaged Prunus seeds. Plant Physiology 104, 29±35.
Prokofjeva, I.V., Benken, I.I., Gutman, F.N., 1990. Methodology of determining the content of cyano-
genic glycosides in the seeds of Vicia sativa. Nauchno-Tekhnicheski-Bulleten, Instituta
Rasteniovodstva imeni N.I. Vavilova 202, 31±34.
Ragoobirsingh, D., Robinson, H.M., Morrison, E.Y., St, A., 1993. E�ects of cassava cyanoglucoside
linamarin on blood sugar levels in the dog. Journal of Nutritional Biochemistry 11, 625±629.
Rossouw, J.E., 1989. Kwashiorkor in North America. American Journal of Clinical Nutrition 49, 588±
592.
Saunders, J.A., Conn, E.E., 1978. Presence of the cyanogenic glycoside dhurrin in isolated vacuoles
from Sorghum. Plant Physiology 61, 154±157.
Sargison, N.D., Williamson, D.S., Duncan, J.R., McCance, R.W., 1996. Prunus padus (bird cherry) poi-
soning in cattle. Veterinary Record 138, 188.
Shappert, P.J., Shore, J.S., 1995. Cyanogenesis in Turnera ulmifolia L. (Turneraceae). I. Phenotypic dis-
tribution and genetic variation for cyanogenesis on Jamaica. Heredity 74, 392±404.
Schwartz, B., Wray, V., Proksch, P., 1996. A cyanogenic glycoside from Canthium schimperianum.
Phytochemistry 42, 633±636.
Seigler, D.S., Maslin, B.R., Conn, E.E., 1989. Cyanogenesis in the Leguminosae. Monographs in
Systematic Botany from the Missouri Botanical Garden 29, 645±672.
Selmar, D., Lieberei, R., Biehl, B., 1988. Mobilization and utilization of cyanogenic glycosides: the
linustatin pathway. Plant Physiology 86, 711±716.
Sibbesen, O., Koch, B., Halkier, B.A., Moller, B.L., 1994. Isolation of the haeme-thiolate enzyme cyto-
chrome P-450TYR which catalizes the committed step in the biosynthesis of the cyanogenic glyco-
side dhurrin in Sorghum bicolor (L.) Moench. Proceedings of the National Academy of Sciences of
the USA 91 (21), 9740±9744.
JaÂnos Vetter / Toxicon 38 (2000) 11±36 35

Swain, E., Poulton, J.E., 1994. Utilization of amygdalin during seedling development of Prunus sero-
tina. Plant Physiology 106, 437±445.
Tapper, B.A., Reay, P.F., 1973. Cyanogenic glycosides and glucosinolates. In: Butler, C.W., Bailey,
R.W. (Eds.), Chemistry and Biochemistry of Herbage, 1. Academic Press, London, New York, pp.
447±476.
Tull, D., Withers, S.G., Gilkes, N.R., Kilburn, D.G., Warren, R.A.J., Aebersold, R., 1991. Glutamic
acid 274 is the nucleophile in the active site of a retaining exoglucanase from Cellulomonas ®mi.
Journal of the Biological Chemistry 266, 15621±15625.
Tungtrakanpoung, N., Rhienpanish, K., 1992. The toxicity of Mimosa invisa var. inermis Adalbert to
bu�aloes. Bu�alo Bulletin 11, 230±231.
Varshney, J.P., Gupta, A.K., Yadav, M.P., 1996. Occurrence of ataxia-cystitis syndrome in horses fed
on Sorghum vulgare in India. Indian Veterinary Journal 73, 985±986.
Vetter, J., Haraszti, E., 1975. Determination of cyanoglycosides of plant tissue by the modi®ed picric
acid technique. Agrokemia es Talajtan 24, 413±422.
Vetter, J., Haraszti, E., 1977. Changes in the hydrogen cyanide content of Sudan grass (Sorghum suda-
nense ) and broomcorn (Sorghum bicolor var. technicum ) during the growing season. Acta
Agronomica Sci. Hung. 26, 15±22.
Wajant, H., Riedel, D., Benz, S., Mundry, K.W., 1994. Immunocytological localization of hydroxyni-
trile lyases from Sorghum bicolor L. and Linum usitatissimum L. Plant Science 103, 145±154.
Wheatley, C.C., Orrego, J.I., Sanchez, T., Granados, E., 1992, 25±28 Augus± 1993. Quality evaluation
of the cassava core collection at CIAT. In: Roca, W.M., Thro, A.M. (Eds.), Proceedings of the ®rst
international scienti®c meeting of the Cassava Biotechnology Network, Cartegena, Colombia. CIA,
Cali, Columbia, pp. 255±264.
Widdup, K.H., Caradus, J.R., Green, J., Mueller, J.P., Pennell, C.P., 1996. White clover: New
Zealand's competition edge. In: Joint Symposium, 1995, Lincoln University, 21±22 November, 1995,
pp. 149±153.
Yeoh, H.H., Yeoh, Y.C., 1994. Some properties of b-glucosidases from tropical plant species.
Phytochemistry 35, 1391±1393.
JaÂnos Vetter / Toxicon 38 (2000) 11±3636