physiological testosterone levels enhance chondrogenic extracellular matrix synthesis by male...
TRANSCRIPT

The Spine Journal 14 (2014) 455–468
Basic Science
Physiological testosterone levels enhance chondrogenicextracellular matrix synthesis by male intervertebral disc cells in vitro,
but not by mesenchymal stem cells
Alessandro Bertolo, PhDa, Martin Baur, MDb, Niklaus Aebli, MD, PhDc,d,Stephen J. Ferguson, PhDe, Jivko Stoyanov, PhDa,f,*
aBiomedical Laboratories, Swiss Paraplegic Research, G.A. Zaech Strasse 4, 6207 Nottwil, SwitzerlandbCantonal Hospital of Lucerne, Spitalstrasse 16, 6000 Lucerne, Switzerland
cSwiss Paraplegic Centre, Zaechstrasse 1, 6207 Nottwil, SwitzerlanddSchool of Medicine, Griffith University, University Drive, 4131 Meadowbrook Qld, Brisbane, Queensland, Australia
eInstitute for Biomechanics, Schafmattstrasse 30, 8093 ETH Zurich, SwitzerlandfInstitute for Surgical Technology and Biomechanics, University of Bern, Stauffacherstrasse 78, 3014 Bern, Switzerland
Received 4 June 2013; revised 11 September 2013; accepted 17 October 2013
Abstract BACKGROUND CONTEXT: Testosterone (
FDA device/drug
Author disclosures
NA: Nothing to disc
disclose.
1529-9430/$ - see fro
http://dx.doi.org/10.10
T) is a hormone and regulator involved in the pro-cesses of development of the organism (ie, promoting development of bone and muscle mass).Although T effects on the mesenchyme-derived muscle, bone, and adipose tissues are well studied,T effects on intervertebral disc (IVD) have not been reported.PURPOSE: The aim was to test the following hypothesis: if a physiological concentration ofT (~30 nM) can improve in vitro chondrogenesis of human IVD cells and mesenchymal stem cells(MSCs).STUDY DESIGN/SETTING: Human IVD cells and MSCs were differentiated to chondrogeniclineage on gelatin scaffolds for 4 weeks, in the presence or absence of T.METHODS: Chondrogenesis was assessed by cell viability, by measuring gene expression withquantitative polymerase chain reaction and extracellular matrix (ECM) accumulation with immuno-blotting, immunohistochemical, and biochemical methods.RESULTS: Supplementation of T to chondrogenic culture did not affect viability. In male IVDcells, T had a beneficial impact on chondrogenesis, particularly in nucleus pulposus cells, demon-strated by an increased expression of aggrecan, collagen type I, and especially collagen type II.Conversely, T had no effects on chondrogenesis of female IVD cells or MSCs from both genders.A gene expression array of transforming growth factor b/bone morphogenetic protein signaling cas-cade showed that in male IVD cells, T promoted a stable general but nonsignificant increase in geneexpression. Furthermore, aromatase inhibitor anastrazole repressed the effect of T on ECM expres-sion by IVD cells. The results suggest that T increased ECM accumulation in male IVD cells incombination with its conversion to estradiol by the enzyme aromatase.CONCLUSIONS: We demonstrated that T effectively enhances in vitro chondrogenesis inmale IVD cells, rising the interest in the possible role of sex hormones in IVD degeneration. Never-theless, T does not affect chondrogenic differentiation of female IVD cells and MSCs from bothgenders. � 2014 Elsevier Inc. All rights reserved.
Keywords: Intervertebral disc cells; Mesenchymal stem cells; Chondrogenesis; Aromatase; Testosterone and three-
dimensional cultures; Sex hormones
status: Not applicable.
: AB: Nothing to disclose. MB: Nothing to disclose.
lose. SJF: Nothing to disclose. JS: Nothing to
* Corresponding author. Biomedical Laboratories, Swiss Paraplegic
Research, G.A. Z€ach Strasse 4, CH-6207 Nottwil, Switzerland. Tel.: (41)
41-939-6635; fax: (41) 41-939-6640.
E-mail address: [email protected] (J. Stoyanov)
nt matter � 2014 Elsevier Inc. All rights reserved.
16/j.spinee.2013.10.018

Table 1
Demographic details of intervertebral disc donors (average age541 years)
Sample Sex
Donor’s
age, y
Thompson
grading scale Type of operation
MD F 32 IV DDD, L5–S1
MD M 50 III Trauma, L2–L3
MD M 37 V DDD, L5–S1
AF M 25 IV DDD, L5–S1
AF F 57 II Trauma, Th10–Th11
AF M 45 III DDD, L5–S1
NP F 34 II Trauma, Th12–L1
NP F 39 II Trauma, C4–C5
NP M 47 IV DDD, C5–C6–C7
NP M 46 III DDD, L5–S1
AF, annulus fibrosis; DDD, disc degeneration disease; L, lumbar verte-
brae; MD, mixed disc (no distinction between AF and NP); NP, nucleus
pulposus; S, sacral vertebrae; Th, thoracic vertebrae.
456 A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
Introduction
Back pain (BP) is one of the most prevalent health prob-lems in industrialized countries, leading to productivitylosses for the economy and generating additional coststo support the treatment of affected individuals [1,2]. Trau-matic injuries, poor posture, smoking, and, last but notleast, genetics [3,4] are all variables linked with BP. Incertain cases, BP is caused by degeneration of the interver-tebral discs (IVDs), the largest avascular and anervous tis-sues of the body, which comprise three distinct tissues: aninner gel-like nucleus pulposus (NP) surrounded by thefibro-cartilaginous annulus fibrosus (AF) and cartilaginousend plates. The etiology of disc degeneration is complex,but the occurrence of IVD degeneration is strongly associ-ated with age: 97% of individuals 50 years or older havedegenerated discs [5]. During degeneration, the homeo-static mechanisms maintaining the biochemical composi-tion and biomechanical structure of the IVD are altered,leading eventually to mechanical failure of the extracellularmatrix (ECM). Such failure is facilitated by a misregulationof the balance between anabolic and catabolic processes,that is, the expression of the major structural moleculescomprised in the IVD, such as collagens and proteoglycans[6], and their turnover catalyzed by matrix metalloprotei-nases (MMPs) [7]. A general loss of collagens and proteo-glycans results in a loss of fixed negative charges from theNP, gradual dehydration of the IVD, and a consequent de-crease in disc height [8].
Aging is also well known to reduce testosterone (T) syn-thesis and its concentration in the circulatory system in bothmen and women [9]. Testosterone, as a steroid hormone,belongs to the group of androgens and binds to the andro-gen receptor (AR), which is a nuclear transcription factor,regulating transcriptional activation of several downstreampathways [10]. Androgen receptor has been localized in hu-man mesenchymal stem cells (MSCs) [11,12], althoughthere is no evidence if it is expressed also by IVD cells.Not only does T play a key role in the development of malereproductive tissues, it also promotes increased muscle andbone mass synthesis and accumulation [13]. The concentra-tion of T circulating in the blood is directly correlated withmuscular mass [14], whereas T supplementation decreasedfat mass in older men with low T levels [15]. Very few stud-ies report effects of T on cartilaginous tissues, for examplethe absence of T induced apoptosis and decreased prolifer-ation and the total number of chondrocytes was studied incastrated rabbits [16], but T stimulated the growth of chon-drocyte cell layers in organ culture of mice mandibular con-dyles, as well as the local production of insulinlike growthfactor (IGF-I) and IGF-I receptor [17]. In parallel, it hasbeen shown that human articular chondrocytes are also af-fected by the female sex hormone b-estradiol (E2), whichpromoted DNA synthesis, sulfate incorporation, and alka-line phosphatase activity in female-derived chondrocytes[18]. As articular chondrocytes have a similar morphology
and gene expression profile a IVD cells, our hypothesis wasthat IVD might experience similar effects of the sexhormones.
Therefore, we supplemented cell cultures with T at con-centrations equivalent to upper physiological levels foundin male blood, expecting that it may stimulate anabolicIVD activity, and may correlate with an improvement ofthe disc homeostasis by promoting the expression of de-sired ECM proteins, such as aggrecan and collagen type II.
In the present study, we also investigated the effects of Tonthe regulation of in vitro ECM accumulation during chondro-genic differentiation of IVD cells in comparisonwithMSCs, amain candidate for IVD regeneration therapies, derived fromdonors of both sexes. Cells were differentiated in three-dimensional cultures, and cell viability and chondrogenesiswere assessed by quantification of gene and protein expres-sion, including glycosaminoglycan accumulation.
Materials and methods
IVD cell isolation and cell expansion
IVD cells were isolated from degenerated human disc tis-sue (n510, Table 1) obtained during full or partial discec-tomy, with patient consent and approval by the ethicscommittee of Canton Lucerne. IVD cells were divided intogroups depending on the origin of tissue: AF, NP, or mixeddisc (MD), when disc degeneration did not allow precise dis-crimination between tissues. Disc fragments were digestedin 0.05% collagenase (Sigma-Aldrich, St. Louis, MO,USA), 5% fetal bovine serum (FBS), penicillin (100 units/mL)/streptomycin (100 mg/mL) in Dulbecco’s modified Ea-gle’s medium (DMEM)/F12þGlutaMAX (all Gibco) for 6hours at 37�C. After incubation, cell suspensions were fil-tered through a 100-mm cell strainer (TPP, Faust Lab,Schaffhausen, Switzerland), centrifuged at 1 g, and the pel-lets were washed with 1� phosphate-buffered saline (PBS).IVD cells were expanded in culture for 20 days (two to threepassages) as a monolayer in DMEM/F12þGlutaMAX,

457A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
supplemented with 10% FBS, penicillin (100 units/mL)/streptomycin (100 mg/mL) (all Gibco; Grand Island, NY,USA), 2.5 mg/mL amphotericin B (Applichem, Darmstadt,Germany), and 5 ng/mL recombinant basic fibroblast growthfactor (bFGF) (Peprotech, Rocky Hill, NJ, USA) at 37�C in ahumid atmosphere containing 5% CO2, with media changedthree times a week. Cells used in this study were expandedfor no more than three passages in culture.
MSC isolation and culture
Fresh bonemarrow (BM) samples were obtained from theiliac crest of the donors during surgery, with informed con-sent and approval by the ethics committee of cantonLucerne. MSCs were isolated from BM of 10 patients(5 men and 5 women; average age: 3469 years). The BM as-pirates were diluted twofold in 3.8% sodium citrate and PBS,and filtered through a 100-mm cell strainer (Falcon; BD Bio-science, San Jose, CA, USA). Mononuclear cells were sep-arated by Ficoll gradient centrifugation (density 1.077 g/mL; GE Healthcare, Madison, WI, USA) in a Leucosep tube(Greiner, Monroe, NC, USA) at 800g for 20minutes, washedwith PBS, centrifuged again at 210g for 10 minutes, resus-pended in PBS and counted using trypan blue dye in asingle-use Neubauer chamber (C-Chip Typ Neubauer; Zeiss,Jena, Germany). Cells were placed in a tissue culture flask(TPP) in nonhematopoietic stem cell media (Miltenyi, Ber-gisch Gladbach, Germany) at 37�C in a humid atmospherecontaining 5% CO2. After 2 days, nonadherent cells werediscarded, whereas adherent cells were cultured inDMEM/F12þGlutaMAX, supplemented with 10% FBS,penicillin (100 units/mL)/streptomycin (100 mg/mL), 2.5mg/mL amphotericin B, and 5 ng/ml bFGF with mediumchanged three times a week. Cells used in this project wereexpanded for no more than four passages in culture.
Construct preparation and chondrogenic culture
The scaffold used in this project was a sponge-shapedmedical device made of gelatin–partially hydrolyzed colla-gen (Spongostan; Ferrosan/Pfizer, St Louis, MO, USA)[19]. From this material, cubes with 3-mm side length werecut and used as a support for cellular growth. At 90% conflu-ence, cells were harvested by dissociation with 0.025% tryp-sin, and resuspended in PBS at 4�106 cells/mL density; 18mL cell suspension (equivalent to ~72,000 cells) was pipet-ted on the construct and completely absorbed by it. IVD andMSC cell constructs were kept at room temperature for 30minutes to allow cells to anchor to the matrix, before thecareful addition of media to the 6-well plate. The resultingcell matrix constructs were maintained for 28 days in con-trol, and chondrogenic and chondrogenic media supple-mented with 10 ng/mL (34.7 nM) testosterone (CT10;Sigma, St. Louis, MO, USA), 30 nM b-Estradiol (E2; Sig-ma), or 30 nM dihydrotestosterone (DHT; Sigma). Differentconcentrations of Twere also tested (10 ng/mL, 100 ng/mL,and 1000 ng/mL) and results are presented in
Supplementary Fig. 1. Chondrogenic medium consisted ofAdvanced DMEM/F12þGlutaMAX (Gibco), supplementedwith 2.5% FBS, 40 ng/mL dexamethasone (Applichem),50 mg/mL ascorbate-2-phosphate (Sigma), 100 U/mL peni-cillin/100 mg/mL streptomycin, 2.5 mg/mL amphotericinB, 1� insulin (10 mg/mL)–Transferrin (5.5 mg/mL)–Sele-nium (0.67 ng/mL)–X Supplement (ITS-X; Gibco), and10 ng/mL transforming growth factor-b1 (TGF-b1; Pepro-tech). Aromatase inhibitor anastrazole (100 nM; Sigma)was added to chondrogenic medium with and without T.Negative controls were maintained in Advanced DMEM/F12þGlutaMAX, supplemented with penicillin/streptomy-cin, amphotericin B, FBS, ascorbate-2-phosphate, and ITS.The media was replaced three times per week.
Cell viability assay
To investigate the proportion of live and dead cells after7, 14, and 28 days in different culture conditions, constructscontaining cells were incubated for 2 hours with 0.05% col-lagenase, 0.3% pronase E (Sigma-Aldrich), 5% FBS, andpenicillin (100 units/mL)/streptomycin (100 mg/mL) inDMEM/F12þGlutaMAX at 37�C. After digestion of theconstructs, cells were centrifuged at 250 g for 10 minutes,resuspended in PBS, counted, and the ratio of live and deadcells was determined using trypan blue dye in a Neubauerchamber (C-Chip Typ Neubauer; Zeiss).
Detection of AR by immunofluorescence
The localization of AR was carried out in MSCs and IVDcells cultured for 24 hours on eight-chamber slides (NalgeNunc International) with or without testosterone (10 ng/mL). Cells were fixed for 20 minutes in PBSþ4% parafor-maldehyde (Applichem), washed with PBS, followed byblocking with PBS containing 1 mg/mL bovine serum albu-min (BSA), 10% FBS, and 0.1% Triton (Applichem) for 1hour. After blocking, cells were incubated with polyclonalrabbit anti-AR antibody (N-20; Santa Cruz Biotechnology,Santa Cruz, CA, USA) in a blocking mix overnight at 4�C.Fluorescence labeling was performed with anti-rabbitsecondary antibody linked to TRITC (sc-3841; Santa CruzBiotechnology). The slides were counterstained with 40,6-diamidino-2-phenylindole (DAPI) and were examined undera fluorescence microscope (Olympus, Tokyo, Japan).
RNA isolation, cDNA synthesis, and real-time PCR
RNA was isolated from IVD cell and MSC constructs atdays 7, 14, and 28 and then stored at –80�C as follows: cellconstructs were previously disaggregated and homogenizedin cell lysis buffer using a Dispomix homogenizer (Axonlab,Baden, Germany) and Aurum Total Mini Kit (Bio-Rad, Her-cules, CA, USA), following the manufacturers’ instructionswith modification of adding 2 mL polyacryl carrier (Lucerna-Chem, Lucerne, Switzerland) to the lysis buffer. Total RNA(500 ng/sample) was used for synthesis of cDNA (VILO

458 A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
cDNA Synthesis Kit; Bio-Rad). Complementary DNA(cDNA) was diluted 1:10 with PCR-grade water and the re-sulting cDNA template (5 mL) was mixed with the PCR reac-tion solution (IQ SYBR Green Supermix; Bio-Rad)containing 0.25-mM specific primers as described inTable 2. Specific products were amplified in triplicate in a fi-nal volume of 25 mL in 96-well plates (Bio-Rad) using quan-titative PCR (qPCR) (CFX96 Real Time System; Bio-Rad).Real-time PCR was carried out with the following settings:denaturation 95�C, 3 minutes (one cycle); 95�C, 15 seconds;64�C, 20 seconds; and 72�C, 20 seconds (40 amplificationcycles), followed by melting curve analysis. Gene expressiondifferences were determined using the 2�DDCt method and theresults were normalized to the expression of 60S gene.
Human TGFb/bone morphogenetic protein (BMP)signaling-specific RT2 profiler PCR array (PAHS-035; SABioscience Co. Qiagen, Valencia, CA, USA) was used toquantify 84 genes related to the TGFb/BMP-mediated sig-nal transduction pathway, including members of the TGFb
Table 2
Human genes used in quantitative reverse transcriptase polymerase chain reactio
Gene Primer nucleotide seq
Housekeeping gene
60S F - GAGGCCCCTAC
R - CCTCGCTTGGT
Chondrogenic differentiation markers
Aggrecan F - AGGCTATGAGC
R - GCACGCCATAG
Collagen type I F - CCTCCTGGCTCT
R - AGGGAGACCGT
Collagen type II F - GAAGTGCTGGT
R - GGCCTCTCCTT
MMPs
MMP1 F - ACGAATTTGCCG
R - GTCCTTGGGGT
MMP13 F - CCAGTCTCCGA
R - AAAAACAGCTC
TGFb/BMP signaling pathway
LTBP4 F - TGTTCCCCCACG
R - AGAACGGGGCT
C-Fos F - ACTACCACTCAC
R - CCAGGTCCGTG
JUN F - CCAAAGGATAG
R - CTGTCCCTCTCC
Chordin F - CCAGCCAGGAG
R - GTGCCCACGTT
GDF5 F - GCTTTATTGACA
R - GGGCACTAATG
Nucleus pulposus markers
Keratin 19 F - GCCACTACTACA
R - CAAACTTGGTT
PAX1 F - GCAATGACCTTC
R - GGCAGTCCGTG
Androgen-related genes
Androgen F - GCCTTGCTCTCT
Receptor R - GGTCGTCCACG
Aromatase F - GAATATTGGAAG
R - GGGTAAAGATC
bp, base pair; GDF5, growth differentiation factor 5; F, forward; MMP, mat
beta/bone morphogenetic protein.
superfamily of cytokines and their receptors crucial forchondrogenic differentiation. At day 28, RNA was isolatedfrom IVD cells undergoing chondrogenic differentiation inpresence or absence of T, 500 ng/sample RNA was used tosynthesize cDNA and subjected to PCR array analysis. Rawdata were analyzed using PCR Array Data Analysis soft-ware (Qiagen), and fold changes in relative gene expressionwere presented after normalization to five housekeepinggenes (GAPDH, B2M, Actin, RPL13A, and HPRT1).
Histological and immunohistological analysis
Constructs were harvested following 28 days of culture inchondrogenic and CT10media, embedded in OCT compound(Sysmex Digitana, Kobe, Japan) for 30 minutes, and then fro-zen at�80�C. Constructs were subsequently cryosectioned at20-mm thickness using a cryostat (CM 1850; Leica).
Histological detection of sulfated glycosaminoglycan(GAG) accumulation was carried out by alcian blue staining.
n
uence, 50 / 30 Amplicon, bp
CACTTCC 82
TTTGTGG
AGTGTGAACG 125
GTCCTGA
CCTGGT 106
TGAGTCCAT
GCTCGTG 125
GCTCACC
ACAGAGAT 95
ATCCGTGTA
GGAGAAACA 88
CGCATCAAC
AACTTCT 69
AGCAGGT
CCGCAGAC 75
CAGAAGT
TGCGATGTTT 62
ACTGCAAC
GACACAC 99
CAGGAAG
AAGGGCAAGA 75
TCAAACACG
CGACCATCC 126
CGGAAGTCAT
AAGCATCC 91
TAAGCTACTG
AGCCTCAA 103
TGTAAGTTG
GATGCACAGACT 293
ATTTCCAGCATGT
rix metalloproteinase; R, reverse; TGFb/BMP, transforming growth factor

Fig. 1. (Top) Images of the intervertebral disc (IVD) cell constructs (top
row) and mesenchymal stem cell (MSC) constructs (bottom row) in nega-
tive control, chondrogenic, and CT10 differentiation cultures at day 28
(entire scale bar is 2 mm). (Middle) Cell constructs from each type of cul-
ture were enzymatically digested and the number of cells per construct
(IVD cells, upper graph; MSCs, lower graph) was determined over a period
of 28 days (n510, the number of cells per construct is represented as
mean6standard deviation [SD]). (Bottom) DNA quantification of IVD cell
and MSC constructs at day 28 (n510, DNA content is represented as
mean6SD).
459A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
Sections were stained overnight with 0.4% alcian blue (Fluka;Sigma) dissolved in 0.01% H2SO4 (Applichem) and 0.5 Mguanidine hydrochloride (Fluka). Next, sections were washedfor 30 minutes in 40% dimethyl sulfoxide and 0.05 MMgCl2(both from Applichem). Finally, sections were mounted with70% glycerol and examined by light microscopy.
Immunohistochemical analysis was used to detect aggre-can and collagen type I and type II accumulation. Endoge-nous peroxidase was quenched by 3% H2O2 in PBS atroom temperature for 10 minutes, and washed with PBS. Be-fore incubation, sections for immunodetection with antiag-grecan antibody were predigested with chondroitinaseABC (0.25 U/mL, Sigma-Aldrich) in 0.1 M Tris-HCl and0.03 M acetate buffer, pH 6.5, for 3 hours at 37�C. Nonspe-cific background was blocked with PBS containing 1mg/mLBSA, 10% FBS, and 0.1% Triton (Applichem) for 30minutes. Sections were incubated overnight at 4�C withmonoclonal mouse antiaggrecan antibody (969D4D11;Biosource), mouse anticollagen type I antibody (M-38;Development Studies Hybridoma Bank, University of Iowa,Iowa City, IA, USA), and monoclonal mouse anticollagentype II antibody (II-II6B3; Development Studies HybridomaBank) in blocking solution. After washing with PBS, allsections were incubated with a secondary biotinylated goatanti-mouse antibody (B0529; Sigma), and then withstreptavidin-horseradish peroxidase (HRP) (S2438; Sigma)for 45 minutes at room temperature. Aggrecan and collagentype I and type II were visualized by reaction with 0.075%solution of 3-amino-9-ethylcarbazole (Applichem) in0.01% H2O2. Sections were mounted with 70% glycerol(Applichem) and examined by light microscopy.
Glycosaminoglycan accumulation and DNA assays
Glycosaminoglycan accumulation was quantified withalcian blue binding assay after 6 hours of digestion of threeconstructs per sample at 60�C with 125 mg/mL papain(Sigma-Aldrich) in 5 mM L-cysteine-HCl (Fluka), 5 mMNa-citrate, 150 mM NaCl, and 5 mM EDTA (all Appli-Chem). Glycosaminoglycan accumulation was determinedby binding to alcian blue, and absorption was measuredat 595 nm and quantified against chondroitin sulfate(Sigma-Aldrich) reference standards [20].
Total double-stranded DNAwas measured for each sam-ple after papain digestion, as previously described. Theamount of DNAwas determined using SYBR green (LuBio-Science, Toepferstrasse, Switzerland) fluorescent assay (ab-sorption measured at 535 nm), quantified by referring to calfthymus DNA (Sigma-Aldrich) standards.
Collagen type II immunoblot analyses
Total protein content was isolated from both IVD cellsand MSCs (control, chondrogenic, and CT10 groups) duringRNA extraction by collecting the lysis and washing flow-through using a Total RNAMini Kit (Bio-Rad). After precip-itation overnight at �80�C, the samples were centrifuged at
14,000 g for 15 minutes, the protein pellets were washedtwice in 70% ethanol, and then resuspended in CelLytic MCell Lysis Reagent (Sigma-Aldrich). Protein extracts werefractionated by Mini-Protean TGX 4% to 15% polyacryla-mide gels (4561083, Bio-Rad), and electrophoreticallytransferred to a nitrocellulose membrane (Bio-Rad). The ni-trocellulose membranes were incubated with mouse mono-clonal antibodies against collagen type II (1:400, II-II6B3;Development Studies Hybridoma Bank), and housekeeping

Fig. 2. Androgen receptor (AR) presence (red fluorescence) in intervertebral disc (IVD) cells (Top) and mesenchymal stem cells (MSCs) (Bottom) cultured
without (top row) or with 10 ng/mL testosterone (bottom row) was determined by immunocytochemistry. Nuclear translocation of the cytoplasmic AR occurs
in the presence of testosterone in IVD cells and MSCs cultured for 24 hours in monolayer, and 40,6-diamidino-2-phenylindole (DAPI) counterstain was used
to localize the nuclei. (Entire scale bar is 120 mm).
460 A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
gene actin (1:100, JLA20-s; Development Studies Hybrido-ma Bank). Blocked membranes were probed with primaryantibodies diluted in 5% milk (Applichem) for 2 hours,
Fig. 3. Gene expression according to gender of collagen type I (COL1), collage
(Top) and mesenchymal stem cell (MSC) constructs (Bottom) after 28 days of cul
to 60S and compared with expression in chondrogenic medium, data represented
MSCs: 5 male and 5 female).
followed by HRP-conjugated mouse secondary antibody(1:1000; Bethyl) diluted in PBS, for 1 hour at room temper-ature. Membranes were developed with EZ-ECL-Kit
n type II (COL2), and aggrecan (ACAN) by intervertebral disc (IVD) cell
ture in negative control and CT10 media. (Gene expression was normalized
as a mean6standard deviation, *p!.05; IVD cells: 6 male and 4 female,

461A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
(LucernaChem). The results were normalized to the relativeamount of actin.
Statistical analysis
Data were expressed as the mean6standard deviation.The nonparametric Mann-Whitney–Wilcoxon U test for de-pendent variables was used to compare gene expression,DNA quantification, and GAG accumulation, because anal-ysis of variance would assume normal distribution of thedata, which cannot be guaranteed in this data set. For alltests, p!.05 was considered significant. Data analysis wasperformed with SPSS 14.0 for Windows (SPSS Inc., Chica-go, IL, USA).
Fig. 4. Gene expression of collagen type II (COL2) (Top), collagen type I
(COL1) (Middle), and aggrecan (ACAN) (Bottom) by male intervertebral
disc cell constructs after 28 days of culture in control and CT10 media, ac-
cording to tissue source: mixed disc (MD), nucleus pulposus (NP), and an-
nulus fibrosus (AF). (Gene expression was normalized to 60S and
compared with expression in chondrogenic medium, data represented as
a mean6standard deviation, *p!.05; MD n52, NP n52, AF n52).
Results
Cell morphology, cell number, and viability of IVD cellsand MSC constructs
After 28 days in chondrogenic, chondrogenic plus T(CT10), and negative control media, IVD cells and MSCsseeded in gelatin scaffolds had a similar size and shape(Fig. 1, Top). Decreased translucency and increased reflec-tion of light suggested a more compact and dense consis-tency in the constructs cultured in chondrogenic andCT10 media.
The number of cells per construct was time dependentand strongly influenced by the type of culture (Fig. 1, Mid-dle). At day 7, the range of IVD cell numbers per constructwas between approximately 50,000 cells (control medium),approximately 75,000 cells (CT10 medium), and appro-ximately 100,000 cells (chondrogenic medium). Within4 weeks, cell number in the negative control culture waspractically unchanged, whereas cells in both chondrogenicand CT10 cultures more than doubled their numbers byday 28. MSC constructs in chondrogenic and CT10 mediahad initially a similar number of cells retained, and onlyslightly lower in control medium. After 4 weeks, the numberof cells retained by MSC constructs in control medium de-creased further to 50,000 cells, whereas in chondrogenicand CT10 cell media, the number remained constant. Viabil-ity for both IVD cells and MSCs was analyzed and the per-centage of viable cells (~95%), was similar for each type ofculture over the 4-week time course (data not shown).
Similarly, DNA quantification of IVD cell and MSCconstructs at day 28 confirmed previous observations(Fig. 1, Bottom): at the end of the differentiation period,IVD cells were largely more than MSCs and the presenceof T did not influence the number of cells retained.
Testosterone induces nuclear translocation of AR
Immunofluorescence microscopy was used to detect theexpression of AR protein in IVD cells and MSCs (Fig. 2).In the control expansion culture, AR receptor localizationwas mostly restricted in the cytoplasm. Conversely, after
24 hours of incubation with culture medium supplementedwith 10 ng/mL T, a large fraction of AR was translocatedto the nucleus of the cells. Cells were counterstained withDAPI to reveal the respective position of the nuclei. Interest-ingly, both IVD cells and MSCs had heterogeneous levels ofAR protein in the cytoplasm, which translocated fully to thenuclei of IVD cells or partially to the nuclei in of MSCs.
After 28 days in chondrogenic and CT10 cultures, geneexpression of AR was analyzed in IVD cells and MSC con-structs. AR expression was not affected by T, and did notdiffer according to gender (Supplementary Fig. 2A) andIVD tissue type (Supplementary Fig. 2B).
Gene expression analysis of IVD cells and MSCscultured in presence of T, depending on gender
After 28 days in culture, quantitative real-time PCR wasused to assess expression of the most important ECM mol-ecules of the IVD, namely aggrecan (ACAN) and collagentype I (COL1) and type II (COL2) by IVD cells (Fig. 3,

Fig. 5. T effects on extracellular matrix (ECM) accumulation was determined by immunohistochemistry, Western blot, and glycosaminoglycan (GAG)/DNA
ratio. Immunohistochemistry microphotographs (Top) of sections of male mesenchymal stem cell (MSC) and intervertebral disc (IVD) cell (mixed disc [MD]
cells) constructs cultured in chondrogenic and CT10 media for 28 days (on the left as negative control, sections of MSC constructs in control media). Col-
lagen type II, type I, and aggrecan accumulation was depicted by positive red-colored signal, and on the bottom of the panel, deposition of proteoglycan-rich
ECM was proved by alcian blue staining (entire scale bar is 160 mm). Collagen type II Western blot (Middle) was performed on proteins isolated from male
IVD cells (MD cells) and MSC construct after 28 days in culture, and normalized to actin. At days 7, 14, and 28, the accumulation of GAG (Bottom) accord-
ing to gender by MSC and IVD cell constructs in chondrogenic and CT10 media was measured and normalized to the content of DNA (values represent the
mean6standard deviation; n510).
462 A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
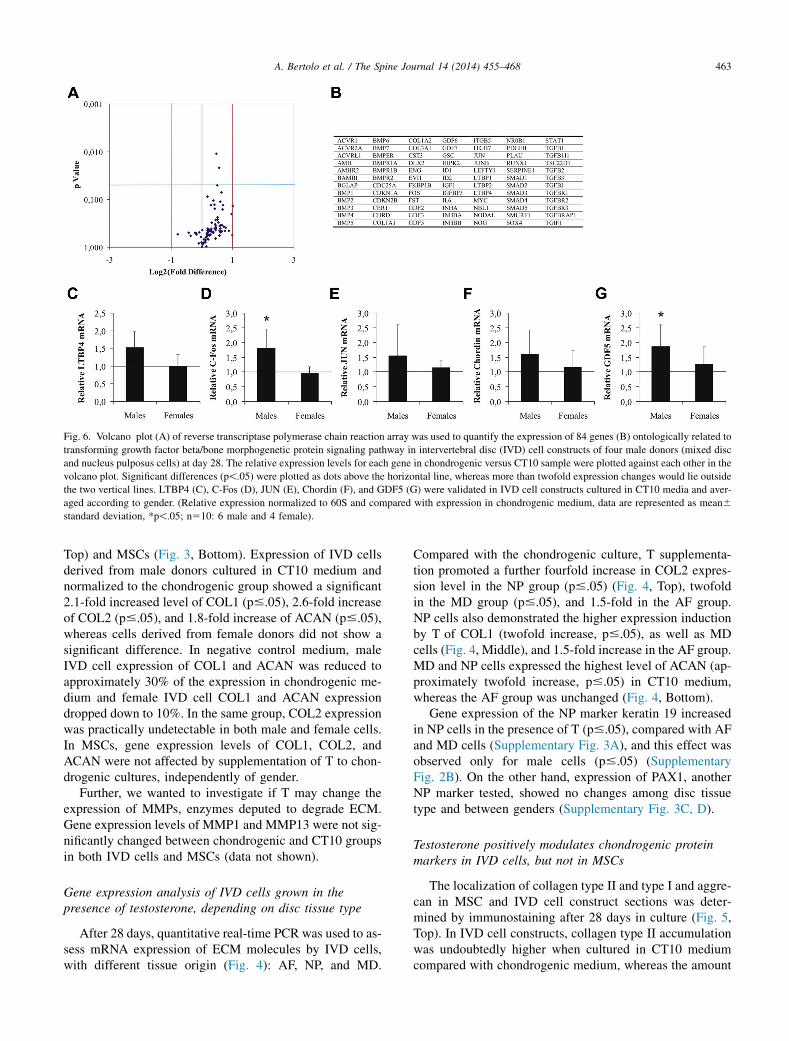
Fig. 6. Volcano plot (A) of reverse transcriptase polymerase chain reaction array was used to quantify the expression of 84 genes (B) ontologically related to
transforming growth factor beta/bone morphogenetic protein signaling pathway in intervertebral disc (IVD) cell constructs of four male donors (mixed disc
and nucleus pulposus cells) at day 28. The relative expression levels for each gene in chondrogenic versus CT10 sample were plotted against each other in the
volcano plot. Significant differences (p!.05) were plotted as dots above the horizontal line, whereas more than twofold expression changes would lie outside
the two vertical lines. LTBP4 (C), C-Fos (D), JUN (E), Chordin (F), and GDF5 (G) were validated in IVD cell constructs cultured in CT10 media and aver-
aged according to gender. (Relative expression normalized to 60S and compared with expression in chondrogenic medium, data are represented as mean6
standard deviation, *p!.05; n510: 6 male and 4 female).
463A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
Top) and MSCs (Fig. 3, Bottom). Expression of IVD cellsderived from male donors cultured in CT10 medium andnormalized to the chondrogenic group showed a significant2.1-fold increased level of COL1 (p#.05), 2.6-fold increaseof COL2 (p#.05), and 1.8-fold increase of ACAN (p#.05),whereas cells derived from female donors did not show asignificant difference. In negative control medium, maleIVD cell expression of COL1 and ACAN was reduced toapproximately 30% of the expression in chondrogenic me-dium and female IVD cell COL1 and ACAN expressiondropped down to 10%. In the same group, COL2 expressionwas practically undetectable in both male and female cells.In MSCs, gene expression levels of COL1, COL2, andACAN were not affected by supplementation of T to chon-drogenic cultures, independently of gender.
Further, we wanted to investigate if T may change theexpression of MMPs, enzymes deputed to degrade ECM.Gene expression levels of MMP1 and MMP13 were not sig-nificantly changed between chondrogenic and CT10 groupsin both IVD cells and MSCs (data not shown).
Gene expression analysis of IVD cells grown in thepresence of testosterone, depending on disc tissue type
After 28 days, quantitative real-time PCR was used to as-sess mRNA expression of ECM molecules by IVD cells,with different tissue origin (Fig. 4): AF, NP, and MD.
Compared with the chondrogenic culture, T supplementa-tion promoted a further fourfold increase in COL2 expres-sion level in the NP group (p#.05) (Fig. 4, Top), twofoldin the MD group (p#.05), and 1.5-fold in the AF group.NP cells also demonstrated the higher expression inductionby T of COL1 (twofold increase, p#.05), as well as MDcells (Fig. 4, Middle), and 1.5-fold increase in the AF group.MD and NP cells expressed the highest level of ACAN (ap-proximately twofold increase, p#.05) in CT10 medium,whereas the AF group was unchanged (Fig. 4, Bottom).
Gene expression of the NP marker keratin 19 increasedin NP cells in the presence of T (p#.05), compared with AFand MD cells (Supplementary Fig. 3A), and this effect wasobserved only for male cells (p#.05) (SupplementaryFig. 2B). On the other hand, expression of PAX1, anotherNP marker tested, showed no changes among disc tissuetype and between genders (Supplementary Fig. 3C, D).
Testosterone positively modulates chondrogenic proteinmarkers in IVD cells, but not in MSCs
The localization of collagen type II and type I and aggre-can in MSC and IVD cell construct sections was deter-mined by immunostaining after 28 days in culture (Fig. 5,Top). In IVD cell constructs, collagen type II accumulationwas undoubtedly higher when cultured in CT10 mediumcompared with chondrogenic medium, whereas the amount
464 A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
of collagen type I and aggrecan staining showed similar in-tensity between the groups. Interestingly, collagen type IIand type I and aggrecan accumulation was visibly higherin the inner part of MSC constructs compared with IVD cellconstructs, but there were no differences in staining be-tween chondrogenic and CT10 cultures. In negative controlmedium, MSC constructs showed, as expected, no accumu-lation of collagen type II, but only collagen type I and, to alesser extent, aggrecan. Western blot on protein extracts ofIVD cell and MSC constructs confirmed the higher accu-mulation of COL2 in the presence of T in IVD cells, andno changes in MSCs (Fig. 5, Middle).
Proteoglycan (GAG) accumulation was evaluated byhistological staining of construct sections and quantifiedby alcian blue precipitation reaction (Fig. 5, Bottom).MSC constructs accumulated more proteoglycan comparedwith IVD cell constructs. Histologically, chondrogenic andCT10 groups appeared to have similar proteoglycan stain-ing in both MSC and IVD cell construct sections. The de-tailed quantitative GAG precipitation analysis of IVD cellconstructs showed that cells in CT10 accumulated almost25% more GAG than cells in chondrogenic cultures in maleand 35% more than female cells; however, both did notreach significance, whereas no differences were present atday 7 or day 14.
Testosterone influences expression of TGFb/BMPpathway genes
Because expression and accumulation of the most extrac-ellular molecules of the disc are under the control of theTGFb/BMP pathway, we studied how this pathway was af-fected by T, using a specific qPCR array containing 84genes, the expression of which is related to the TGFb/BMP pathway (Fig. 6, Top, and a list of genes in Fig. 6, Mid-dle). The relative volcano plot showed a modest but stableupregulation of gene expression levels in CT10 culturescompared with the chondrogenic group, demonstrated by aright shift of the data points. The response of five genes spe-cifically upregulated only in male-derived IVD cells wasfurther validated by reverse transcriptase PCR (RT-PCR):latent TGFb binding protein 4 (LTBP4) showed 1.5-fold in-duction (p#.05), C-Fos showed 1.8-fold induction (p#.05),C-Jun showed 1.5-fold induction, Chordin showed 1.5-foldinduction, and growth differentiation factor-5 (GDF5)showed 1.8-fold induction (p#.05).
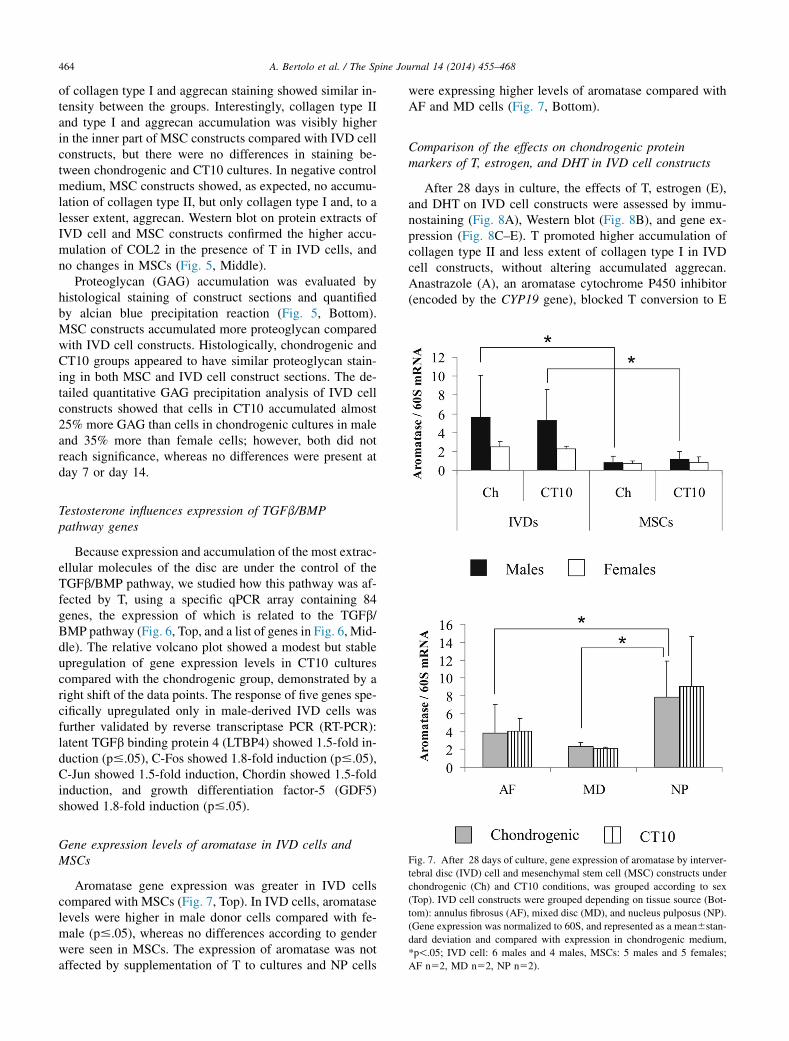
Fig. 7. After 28 days of culture, gene expression of aromatase by interver-
tebral disc (IVD) cell and mesenchymal stem cell (MSC) constructs under
chondrogenic (Ch) and CT10 conditions, was grouped according to sex
(Top). IVD cell constructs were grouped depending on tissue source (Bot-
tom): annulus fibrosus (AF), mixed disc (MD), and nucleus pulposus (NP).
(Gene expression was normalized to 60S, and represented as a mean6stan-
dard deviation and compared with expression in chondrogenic medium,
*p!.05; IVD cell: 6 males and 4 males, MSCs: 5 males and 5 females;
AF n52, MD n52, NP n52).
Gene expression levels of aromatase in IVD cells andMSCs
Aromatase gene expression was greater in IVD cellscompared with MSCs (Fig. 7, Top). In IVD cells, aromataselevels were higher in male donor cells compared with fe-male (p#.05), whereas no differences according to genderwere seen in MSCs. The expression of aromatase was notaffected by supplementation of T to cultures and NP cells
were expressing higher levels of aromatase compared withAF and MD cells (Fig. 7, Bottom).
Comparison of the effects on chondrogenic proteinmarkers of T, estrogen, and DHT in IVD cell constructs
After 28 days in culture, the effects of T, estrogen (E),and DHT on IVD cell constructs were assessed by immu-nostaining (Fig. 8A), Western blot (Fig. 8B), and gene ex-pression (Fig. 8C–E). T promoted higher accumulation ofcollagen type II and less extent of collagen type I in IVDcell constructs, without altering accumulated aggrecan.Anastrazole (A), an aromatase cytochrome P450 inhibitor(encoded by the CYP19 gene), blocked T conversion to E

Fig. 8. At day 28, immunohistochemical staining (A) and gene expression analysis were used to compare the effects of chondrogenic cultures supplemented
with testosterone (CT10), testosterone and aromatase inhibitor anastrazole (TA), b-estradiol (E), and dihydrotestosterone (DHT) on male intervertebral disc
(IVD) cell constructs. Accumulation of collagen type II and type I, and aggrecan is depicted in red on sections of IVD cell constructs (mixed disc cells)
cultured in different media (entire scale bar is 160 mm). Collagen type II Western blot (B) was performed on proteins isolated from male IVD cell constructs
after 28 days in culture, and normalized to actin. The protein extracts used were negative control (1), chondrogenic (2), chondrogenicþA (3),
chondrogenicþT (4), and chondrogenicþTA (5). Gene expression of collagen type II (C), collagen type I (D), and aggrecan (E) by male IVD cell constructs
was evaluated in control and chondrogenic cultures supplemented with either anastrazole (ChþA), T (CT10), T and anastrazole (ChþTA), b-estradiol
(ChþE), or dihydrotestosterone (ChþDHT). (Relative expression normalized to 60S and compared with chondrogenic medium, data are represented as a
mean6standard deviation, *p!.05; male IVD cell n56).
465A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
in IVD cells and canceled the beneficial effects observedwith T alone by reducing collagen type II and type I accu-mulation to levels observed in standard chondrogenic cul-ture. On the other hand, E reduced the accumulation ofcollagen type I and aggrecan, in comparison with chondro-genic cultures, without visible effects on collagen type II.DHT induced higher accumulation of collagen type I inIVD cell constructs, and promoted a moderate increase incollagen type II and aggrecan accumulation.
Quantitative real-time PCR showed that the expressionof COL2, COL1, and ACAN by IVD cell constructs cul-tured with E and DHT was similar to the chondrogenicgroup, and only T induced significant increases in the geneexpression (p#.05). These data, as well as collagen type IIWestern blot, confirmed the already observed inhibition of
the positive effects of T by A. As a control, chondrogeniccultures of IVD cell constructs supplemented with onlyA did not show notable differences in gene expression.
Discussion
In this study, we tested if there would be a place for Tsupplementation in association with future tissue engineer-ing approaches for IVD regeneration, based on the specula-tion that T deficiency might be a factor involved in IVDdegeneration. For this purpose, we used three-dimensionalcultures of human IVD cells derived from degenerated tis-sue and BM-derived MSCs as an in vitro model system toinvestigate the effects of 10 ng/mL T on chondrogenesis.

Fig. 9. Proposed mechanism of testosterone (T) on male-derived interver-
tebral disc cells. T is metabolized by cells in three ways: (1) it might be
converted to dihydrotestosterone (DHT) by the enzyme SRD5A1, (2) inter-
nalized in its original form, or (3) converted to b-estradiol (E) by aroma-
tase. In the cytoplasm, T and DHT bind to androgen receptor (AR) and E
binds to estrogen receptor (ESR1). We propose that the binding of T to cy-
toplasmic AR promotes its translocation to the nucleus, and in association
with conversion of T to E, enhances extracellular matrix (ECM) production
via the transcription-inducing complex c-FOS/c-JUN, which is also part of
the SMAD independent pathway of transforming growth factor beta
(TGFb) signaling. BMP, bone morphogenetic protein; TGF-b1R, TGF-
b1 receptor.
466 A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
This T concentration was chosen as an upper physiologicalvalue in young men, normal range 2.5 to 11.0 ng/mL(the normal range in women being an order of magnitudelower). We demonstrated by immunohistochemistry thatT had an immediate biological effect on the IVD cells iso-lated from male donors and induced nuclear translocationof the AR. In our study, confirmed at transcriptional (quan-titative RT-PCR) and translational (immunohistochemistryand Western blot) levels, T promoted chondrogenesis byupregulating the mRNA and protein expression in IVDcells of key ECM components of the disc, namely aggrecanand collagen type I and type II, and did not regulate expres-sion of the common MMP1 and MMP13. There were nosignificant differences in cell response to T based onthe grade of degeneration of the origin disc tissue, compar-ing IVD cells between mild (Thompson grades II–III) andsevere (Thompson grades IV–V) degeneration grades(Supplementary Fig. 4). Also for MSCs, we demonstratedthat T neither stimulates nor inhibits chondrogenic differ-entiation. Gender-specific effects on chondrogenesis ofT have been reported before with controversial results: inrat chondrocyte culture, T increased the percentage of col-lagen production in male cells only [21], but in human pro-genitor chondrocytes, T increased collagen type I and typeII gene expression only in female-derived cells [22].
There are various possible underlying mechanisms thatmay lead to these effects. Sex hormones, including T, inthe body are extensively studied and have effects far beyondthe sexual function. On the biochemical, cellular, and organ-ism levels, T plays a complex role and, apart from being im-portant for sexual function, it is involved in the regulation ofbone density, adipose tissue distribution, mood, energy, andpsychological well-being [23]. Furthermore, T modulatesthe immune response and its immunomodulatory potentialhas been described for the treatment of atherosclerosis[24]. Testosterone replacement therapy reduces the proin-flammatory cytokines tumor necrosis factor a and interleu-kin-1b while increasing the anti-inflammatory cytokineinterleukin-10 [25]. The same cytokines have been detectedin the degenerated IVD [26], implying that in vivo treatmentof degenerate IVDs could act not only on improved ECMdeposition, but also onmodulation of the inflammatory state.
This beneficial influence of T on IVD cells, which weobserved, would make biological sense in vivo in termsof relation between IVD degeneration and sex hormonallevels. As known, IVD degeneration is strongly age related[5], but some cases of IVD degeneration in men have beenreported already in the second decade of life, and the num-ber of IVD degeneration cases of young aged-matched in-dividuals was slightly but significantly more frequentin men compared with women [5,27]. Consequently, youngmen are more inclined to IVD degeneration rather thanage-matched women, but it is unknown if the degenerationdifference is because of environmental factors, suchas increased mechanical stress and physical injuries or met-abolic reasons, for example, because these men had lower
levels of T. Remarkably, the demographics of IVD degener-ation is reversed in the elderly; in a radiographic study of55-year-old subjects and older, IVD space narrowing wasmore prevalent in postmenopausal women than in men[28] and male propensity for more severe IVD degenerationwas no longer evident by the age of 60 years [5]. This dif-ference reported between ages in disc degeneration suscept-ibility has been associated with alteration in the hormonalbalance. Sex hormones regulate the different stages ofaging, with a faster progressive loss in estrogens in women(menopause) and slower loss of T in men [9]. Indeed, lowerestrogen release in the blood, which occurs with meno-pause, has also been connected to IVD degeneration [29].Total lumbar IVD height was significantly lower in un-treated menopausal women compared with both premeno-pausal and hormone-treated postmenopausal women [30].Gruber et al. [31] showed that human AF cells express es-trogen receptor-b and estrogen (17-b-estradiol) stimulatedin vitro cell proliferation of AF cell cultures. No such studyhas been done for T but the estradiol link may be indicativeof some explanation of our results. Testosterone is con-verted into 17-b-estradiol by the aromatase CYP19, which

467A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
could link our results with the sex hormone changes de-scribed previously, but in this study we did not measurethe rate of conversion of T to 17-b-estradiol. Older studiesin cartilage hint against such a conclusion: chondrocytesfrom women were stimulated by 17-b-estradiol treatment[18] but cartilage volume in the knee showed no associationwhen T levels increased [32], suggesting that, if chondro-cytes have aromatase activity at all, possibly not enoughT was converted to 17-b-estradiol; however, until now nostudies of T and IVD have been published. In our study,by blocking aromatase activity with anastrazole, we wereable to show a significant decrease of anabolic ECM pro-ducing activity in male-derived IVD cells, suggesting stim-ulation by 17-b-estradiol. However, by adding synthetic17-b-estradiol, in the absence of T, the activity observedwith T was not achieved. Because anastrazole by itself (inabsence of T) did not inhibit chondrogenesis, we may con-clude that the observed stimulation by T is achieved only bya T/17-b-estradiol balance, a specific ratio that needs to bestudied further.
We hypothesized also which molecular mechanismmight underlay the beneficial effects of T on IVD ECM ac-cumulation (Fig. 9); by (1) inducing directly AR or (2) in-directly via SRD5A1 reductase (converting T to DHT)followed by their nuclear localization, or (3) via estrogenreceptor (aromatase convert T to 17-b-estradiol). In ourstudy, we did not observe a repression of the TGFb, onthe contrary there was a low but persistent level of upregu-lation of approximately 70 of 84 genes related to the TGFb/BMP pathway. The opposite has been reported for humanprostate cells, where the TGFb signaling pathway was re-pressed by AR signaling thorough interaction with SMAD3[33], suggesting that in different cells, AR may work ondifferent promoters. The TGFb signaling pathway is centralfor the increased ECM deposition in IVD cells and TGFb1is a growth factor commonly used in chondrogenic differ-entiation of cells in vitro, so considering our data we haveto accept that it is either the general low-level stimulationof this pathway that is responsible for the enhanced chon-drogenesis or that AR may promote its biological functionsin IVD cells through a different pathway. In addition, nei-ther AR nor SRD5A1 were expressed specifically in thecell groups responding more efficiently to T (ie, male-derived IVD cells, especially NP cells). An alternative hy-pothesis we tested was that T is converted by aromatase to17-b-estradiol, which binds to estrogen receptor-1. In ourstudy, aromatase was specifically expressed to a higher ex-tent in male-derived IVD cells and NP cells compared withMSCs and AF cells. We know that 17-b-estradiol mightpromote overexpression of Jun and Fos, which form thetranscription factor activator protein-1 (AP-1) [34], a heter-odimeric transcription factor already linked to the TGFbsignaling pathway [35]. Indeed, our results confirmed upre-gulation of Jun and Fos in the presence of T, proposingAP-1 to be responsible of the mechanism elicited by T toimprove ECM synthesis of IVD cells in vitro.
On the other hand, the same positive effects on chondro-genesis were not achieved with BM-derived MSCs, whichwe included in our study because these cells have a highproliferative capacity [36] and the ability to differentiate in-to several cell types [37], including chondrocytes [38] andpossibly IVD cells, and because of their potential for cell-based therapies of degenerated IVD [19]. The absence ofsignificant changes might be because of the diverse gradeof commitment of cells, the fully differentiated interverte-bral cells responding in a different way compared withthe MSCs, or because MSCs have higher (compared withIVD cells) expression of ECM and might have reached theirin vitro optimum, whereas the IVD cells have further poten-tial that can be stimulated by adding T. It has been shownthat T inhibits adipogenic differentiation in vitro [39] andstimulates myogenic differentiation [40] and muscle cellgrowth [41]. Together, these findings lead to the conclusionthat T has other possible effects on MSC differentiation,such as driving them toward the myogenic lineage, a possi-bility not investigated in our study.
In conclusion, we demonstrated that normal-to-highT levels stimulated the accumulation of ECM duringin vitro chondrogenic differentiation of three-dimensionalcultures of male-derived IVD cells, but not in MSCs. Thesefindings are in line with the relevance of decreasing andro-gen levels in aging individuals, and support the hypothesisthat not only estrogens but also androgens, might be linkedto the incidence of IVD degeneration. Our data also doesnot exclude the involvement of an additional pathway acti-vated by T [42]. Therefore, the study opens the way for fur-ther research of a new therapeutically attractive option: theuse of the well-studied and readily available T molecule toenhance ECM synthesis by human disc cells for preventiveand regenerative treatment of degenerated IVDs.
Acknowledgments
This workwas supported by the Swiss Paraplegic Founda-tion and Swiss National Foundation Grant CR3I3_140717/1.
Appendix
Supplementary data
Supplementary data related to this article can be found athttp://dx.doi.org/10.1016/j.spinee.2013.10.018.
References
[1] Wenig CM, Schmidt CO, Kohlmann T, Schweikert B. Costs of back
pain in Germany. Eur J Pain 2009;13:280–6.
[2] Wieser S, Horisberger B, Schmidhauser S, et al. Cost of low back
pain in Switzerland in 2005. Eur J Health Econ 2011;12:455–67.
[3] Pye SR, Reid DM, Adams JE, et al. Influence of weight, body mass
index and lifestyle factors on radiographic features of lumbar disc de-
generation. Ann Rheum Dis 2007;66:426–7.

468 A. Bertolo et al. / The Spine Journal 14 (2014) 455–468
[4] VidemanT, Saarela J, Kaprio J, et al. Associations of 25 structural, deg-
radative, and inflammatory candidate genes with lumbar disc desicca-
tion, bulging, and height narrowing. Arthritis Rheum 2009;60:470–81.
[5] Miller JA, Schmatz C, Schultz AB. Lumbar disc degeneration: corre-
lation with age, sex, and spine level in 600 autopsy specimens. Spine
1988;13:173–8.
[6] Antoniou J, Steffen T, Nelson F, et al. The human lumbar interverte-
bral disc: evidence for changes in the biosynthesis and denaturation
of the extracellular matrix with growth, maturation, ageing, and de-
generation. J Clin Invest 1996;98:996–1003.
[7] Crean JK, Roberts S, Jaffray DC, et al. Matrix metalloproteinases in
the human intervertebral disc: role in disc degeneration and scoliosis.
Spine 1997;22:2877–84.
[8] Dabbs VM, Dabbs LG. Correlation between disc height narrowing
and low-back pain. Spine 1990;15:1366–9.
[9] Liu PY, Pincus SM, Takahashi PY, et al. Aging attenuates both
the regularity and joint synchrony of LH and testosterone secretion
in normal men: analyses via a model of graded GnRH receptor block-
ade. Am J Physiol Endocrinol Metab 2006;290:E34–41.
[10] Mangelsdorf DJ, Thummel C, Beato M, et al. The nuclear receptor
superfamily: the second decade. Cell 1995;83:835–9.
[11] Mantalaris A, Panoskaltsis N, Sakai Y, et al. Localization of androgen
receptor expression in human bone marrow. J Pathol 2001;193:
361–6.
[12] Gupta V, Bhasin S, Guo W, et al. Effects of dihydrotestosterone on
differentiation and proliferation of human mesenchymal stem cells
and preadipocytes. Mol Cell Endocrinol 2008;296:32–40.
[13] Mooradian AD, Morley JE, Korenman SG. Biological actions of an-
drogens. Endocr Rev 1987;8:1–28.
[14] Sinha-Hikim I, Artaza J, Woodhouse L, et al. Testosterone-induced
increase in muscle size in healthy young men is associated with
muscle fiber hypertrophy. Am J Physiol Endocrinol Metab
2002;283:E154–64.
[15] Snyder PJ, Peachey H, Hannoush P, et al. Effect of testosterone treat-
ment on body composition and muscle strength in men over 65 years
of age. J Clin Endocrinol Metab 1999;84:2647–53.
[16] Irie T, Aizawa T, Kokubun S. The role of sex hormones in the kinetics
of chondrocytes in the growth plate. A study in the rabbit. J Bone
Joint Surg Br 2005;87:1278–84.
[17] Maor G, Segev Y, Phillip M. Testosterone stimulates insulin-like
growth factor-I and insulin-like growth factor-I-receptor gene expres-
sion in the mandibular condyle—a model of endochondral ossifica-
tion. Endocrinology 1999;140:1901–10.
[18] Kinney RC, Schwartz Z, Week K, et al. Human articular chondro-
cytes exhibit sexual dimorphism in their responses to 17beta-estra-
diol. Osteoarthritis Cartilage 2005;13:330–7.
[19] Bertolo A, Mehr M, Aebli N, et al. Influence of different commercial
scaffolds on the in vitro differentiation of human mesenchymal stem
cells to nucleus pulposus-like cells. Eur Spine J 2012;21:S826–38.
[20] Bjornsson S. Simultaneous preparation and quantitation of proteogly-
cans by precipitation with alcian blue. Anal Biochem 1993;210:
282–91.
[21] Schwartz Z, Nasatzky E, Ornoy A, et al. Gender-specific, maturation-
dependent effects of testosterone on chondrocytes in culture. Endo-
crinology 1994;134:1640–7.
[22] Koelling S, Miosge N. Sex differences of chondrogenic progenitor
cells in late stages of osteoarthritis. Arthritis Rheum 2010;62:
1077–87.
[23] Burger HG. Androgen production in women. Fertil Steril 2002;77:
S3–5.
[24] Malkin CJ, Pugh PJ, Jones RD, et al. Testosterone as a protective
factor against atherosclerosis—immunomodulation and influence
upon plaque development and stability. J Endocrinol 2003;178:
373–80.
[25] Malkin CJ, Pugh PJ, Jones RD, et al. The effect of testosterone re-
placement on endogenous inflammatory cytokines and lipid profiles
in hypogonadal men. J Clin Endocrinol Metab 2004;89:3313–8.
[26] Ahn SH, Cho YW, Ahn MW, et al. mRNA expression of cytokines
and chemokines in herniated lumbar intervertebral discs. Spine
2002;27:911–7.
[27] Takatalo J, Karppinen J, Niinimaki J, et al. Prevalence of degenera-
tive imaging findings in lumbar magnetic resonance imaging among
young adults. Spine 2009;34:1716–21.
[28] de Schepper EI, Damen J, vanMeurs JB, et al. The association between
lumbar disc degeneration and low back pain: the influence of age, gen-
der, and individual radiographic features. Spine 2010;35:531–6.
[29] Wang YX, Griffith JF. Effect of menopause on lumbar disk degener-
ation: potential etiology. Radiology 2010;257:318–20.
[30] Baron YM, Brincat MP, Calleja-Agius J, Calleja N. Intervertebral
disc height correlates with vertebral body T-scores in premenopausal
and postmenopausal women. Menopause Int 2009;15:58–62.
[31] Gruber HE, Yamaguchi D, Ingram J, et al. Expression and localiza-
tion of estrogen receptor-beta in annulus cells of the human interver-
tebral disc and the mitogenic effect of 17-beta-estradiol in vitro.
BMC Musculoskelet Disord 2002;3:4.
[32] Hanna FS, Bell RJ, Cicuttini FM, et al. The relationship between en-
dogenous testosterone, preandrogens, and sex hormone binding glob-
ulin and knee joint structure in women at midlife. Semin Arthritis
Rheum 2007;37:56–62.
[33] Chipuk JE, Cornelius SC, Pultz NJ, et al. The androgen receptor re-
presses transforming growth factor-beta signaling through interaction
with Smad3. J Biol Chem 2002;277:1240–8.
[34] Silbiger S, Lei J, Neugarten J. Estradiol suppresses type I collagen
synthesis in mesangial cells via activation of activator protein-1. Kid-
ney Int 1999;55:1268–76.
[35] Avouac J, Palumbo K, Tomcik M, et al. Inhibition of activator protein
1 signaling abrogates transforming growth factor beta-mediated acti-
vation of fibroblasts and prevents experimental fibrosis. Arthritis
Rheum 2012;64:1642–52.
[36] Colter DC, Sekiya I, Prockop DJ. Identification of a subpopulation of
rapidly self-renewing and multipotential adult stem cells in colonies
of human marrow stromal cells. Proc Natl Acad Sci U S A 2001;98:
7841–5.
[37] Jiang Y, Jahagirdar BN, Reinhardt RL, et al. Pluripotency of mesen-
chymal stem cells derived from adult marrow. Nature 2002;418:41–9.
[38] Yoo JU, Barthel TS, Nishimura K, et al. The chondrogenic potential
of human bone-marrow-derived mesenchymal progenitor cells.
J Bone Joint Surg Am 1998;80:1745–57.
[39] Singh R, Artaza JN, Taylor WE, et al. Testosterone inhibits adipogen-
ic differentiation in 3T3-L1 cells: nuclear translocation of androgen
receptor complex with beta-catenin and T-cell factor 4 may bypass
canonical Wnt signaling to down-regulate adipogenic transcription
factors. Endocrinology 2006;147:141–54.
[40] Singh R, Bhasin S, Braga M, et al. Regulation of myogenic differen-
tiation by androgens: cross talk between androgen receptor/beta-
catenin and follistatin/transforming growth factor-beta signaling
pathways. Endocrinology 2009;150:1259–68.
[41] Sinha-Hikim I, Taylor WE, Gonzalez-Cadavid NF, et al. Androgen
receptor in human skeletal muscle and cultured muscle satellite cells:
up-regulation by androgen treatment. J Clin Endocrinol Metab
2004;89:5245–55.
[42] Heinlein CA, Chang C. The roles of androgen receptors and
androgen-binding proteins in nongenomic androgen actions. Mol En-
docrinol 2002;16:2181–7.