parte uno tesis edali diciembre 2010
TRANSCRIPT

DEPARTAMENTO DE BIOTECNOLOGÍA AGRÍCOLA
ANÁLISIS PROTEÓMICO DE LA RESISTENCIA INDUCIDA POR MICORRIZACIÓN AL PATÓGENO FOLIAR Sclerotinia sclerotiorum EN FRIJOL
(Phaseolus vulgaris L.)
T E S I S
PARA OBTENER EL GRADO DE
MAESTRA EN
RECURSOS NATURALES Y MEDIO AMBIENTE
PRESENTA
I.BQ. EDALHÍ QUINTERO ZAMORA
GUASAVE SINALOA DICIEMBRE DEL 2010
INSTITUTO POLITÉCNICO NACIONAL
CENTRO INTERDISCIPLINARIO DE INVESTIGACIÓN PARA EL DESARROLLO INTEGRAL REGIONAL
UNIDAD SINALOA

INSTITUTO POLITÉCNICO NACIONALSECRETARÍA DE INVESTIGACIÓN Y POSGRADO
CARTA CESIÓN DE DERECHOS
En la Ciudad de Guasave Sinaloa el día .lQ del mes diciembre del año 2010, el (la) que
suscribe Edalhí Quintero Zamora alumno (a) del Programa de Maestría en Recursos Naturales
y Medio Ambiente con número de registro B081126, adscrito a CIIDIR-IPN Unidad Sinaloa,
manifiesta que es autor (a) intelectual del presente trabajo de Tesis bajo la dirección de el Dr.
Ignacio Eduardo Maldonado Mendoza y el Dr. Sergio Medina Godoy y cede los derechos del
trabajo intitulado "Análisis proteómico de la resistencia inducida por micorrización al patógeno foliar
Sc/erotinia sc/erotiorum en frijol (Phaseolus vulgaris L.)", al Instituto Politécnico Nacional para su
difusión, con fines académicos y de investigación.
Los usuarios de la información no deben reproducir el contenido textual, gráficas o datos del
trabajo sin el permiso expreso del autor y/o director del trabajo. Este puede ser obtenido
escribiendo a la siguiente dirección yo [email protected], [email protected],
[email protected]. Si el permiso se otorga, el usuario deberá dar el agradecimiento
correspondiente y citar la fuente del mismo.
Edot \h( Q..¡l~o -Z.Edalhí Quintero Zamora
Nombre y firma

SIP-13-BIS
INSTITUTO POLITÉCNICO NACIONALSECRETARIA DE INVESTIGACiÓN Y POSGRADO
ACTA DE REGISTRO DE TEMA DE TESISY DESIGNACIÓN DE DIRECTORES DE TESIS
Guasave, Sinaloa a ~ de Noviembre del 2010
El Colegio de Profesores de Estudios de Posgrado e Investigación de CIIDIR-Sinaloa en su sesiónExtraordinaria No. 19 celebrada el día 12 del mes de Noviembre conoció la solicitudpresentada por el (la) alumno(a):
Quintero Zamora
Con registro: ~~~--~~~--~~
Apellido paterno Apellido materno
Aspirante de:
1.- Se designa al aspirante el tema de tesis titulado:"Análisis proteómico de la resistencia inducida por micorrización al patógeno foliar Sclerotiniasc/erotiorum en frijol (Phaseolus vulgaris L.)".
De manera general el tema abarcará los siguientes aspectos:La identificación de proteínas diferenciales relacionadas a la resistencia sistémica inducida en frijolasociadas a la simbiosis micorrízica arbuscular con el hongo Glomus intraradices
2.- Se designan como Directores de Tesis a los Profesores:Dr. Ignacio Eduardo Maldonado Mendoza y Dr. Sergio Medina Godoy
3.- El trabajo de investigación base para el desarrollo de la tesina será elaborado por el alumno en:Las instalaciones del CIIDIR-Sinaloa
que cuenta con los recursos e infraestructura necesarios.
4.- El interesado deberá asistir a los seminarios desarrollados en el área de adscripción deltrabajo desde la fecha en que se suscribe la presente hasta la aceptación de la tesis porla Comisión Revisora correspondiente:
Directores de Tesis
Dr.Dr. Ignacio Eduardo Maldonado Mendoza
Aspirante Presidente del Colegio
I.BQ. Edalhí Quintero Zamora Dr. J«<~ntiel Montoya el 1DI R - 1PNUNIOAD SINALOA
OIRECCION
-------------------------------------------

SIP-14-BISINSTITUTO POLITÉCNICO NACIONALSECRETARíA DE INVESTIGACiÓN Y POSGRADO
ACTA DE REVISIÓN DE TESIS
En la Ciudad de Guasave, Sin. siendo las 17:00 horas del día 6 del mes de
diciembre del 2010 se reunieron los miembros de la Comisión Revisora de Tesis, designadapor el Colegio de Profesores de Estudios de Posgrado e Investigación de CIIDIR-SINALOA
para examinar la tesis titulada:"Análisis proteómico de la resistencia inducida por micorrización al patógeno foliar Sc/erotinia sc/erotiorum
en frijol (Phaseolus vulgaris L.)".
Presentada por el alumno:Quintero Zamora
Apellido paterno Apellido materno
aspirante de:MAESTRIA EN RECURSOS NATURALES Y MEDIO AMBIENTE
Después de intercambiar opiniones los miembros de la Comisión manifestaron APROBAR LATESIS, en virtud de que satisface los requisitos señalados por las disposiciones reglamentariasvigentes.
LA COMISiÓN REVISORA
Directores de tesis
Dr. Ignacio Eduardo Maldonado Mendoza
Adolfo Dagobe
PRESIDENTE DEL COLEGIO DE PROFESORES
------:D=-r.-;¡¡~f-org----7l:"~:-on--:7tl~L.,-1 :-:"Mo'O"":n-:-'toy=a=-<---.;.tn-·-----tCII O 1R - 1P NUNIOAU ~INALúA
DIRfCCION

El hombre encuentra a Dios detrás de cada puerta que la ciencia logra abrir…
Albert Einstein.

Dedicatoria Te dedico mi trabajo de tesis a ti mamá Librada, porque siempre te entusiasmaron mis
logros y sé que esta vez no es la excepción. Gracias por formar parte de tu esencia y porque
no habrá una persona más admirable para mí en la vida que tú. Yo sé que eres el angelote que
alguna vez alguien dijo que me seguía...solo espero encontrarte otra vez, en algún lugar…te
quiero mucho má…

El presente trabajo se llevó a cabo en el Departamento de Biotecnología Agrícola en
el Laboratorio de Alimentos Funcionales del Centro Interdisciplinario de Investigación para
el Desarrollo Integral Regional (CIIDIR) Unidad Sinaloa del Instituto Politécnico
Nacional (IPN) bajo la dirección del Dr. Sergio Medina Godoy y el Dr. Ignacio Eduardo
Maldonado Mendoza. Para este proyecto se recibió el financiamiento por parte del Consejo
Estatal de Ciencia y Tecnología del Estado de Sinaloa (CECYT, Convocatorias 2008 y
2009) y la Secretaria de Investigación y Posgrado del IPN (Proyectos: 20090022,
20100318). La autora agradece al CIIDIR Unidad Sinaloa por su apoyo en infraestructura
para la realización de la presente investigación así como a CONACYT, COFAA-IPN e IPN
por las becas otorgadas (Beca CONACYT y PIFI).

A G R A D E C I M I E N T O S
Primero que nada gracias Dios por estar siempre muy cerquitita de mí y nunca
abandonarme. Te agradezco el haber puesto en mi camino a todas aquellas personas que han
sido mi compañía y que me han ayudado en la realización de este trabajo. Quiero dejar
constancia de todas ellas y agradecerles con sinceridad su participación.
Muchísimas gracias Dr. Sergio Medina Godoy por haberme aceptado como su alumna
de maestría, por haber puesto todos los medios para que yo hiciera mi tesis bajo su dirección y
por su ayuda para que yo asistiera a tres congresos en los que presente mi trabajo. Doc. Sergio
también le agradezco sus enseñanzas, el tiempo invertido y la cordialidad con la que siempre
me apoyó. Y aún más, gracias por su amistad, sus consejos y sus palabras para levantar mi
ánimo. Un especial agradecimiento a usted Dr. Ignacio E. Maldonado Mendoza por su
colaboración como director de esta tesis. Le agradezco Dr. Nacho sus enseñanzas, sus
valiosas sugerencias y el tiempo invertido para la revisión de mi tesis. Gracias a mi comité
tutorial por la revisión de mi tesis: Dra. Melina López Meyer le agradezco su especial interés
en mi trabajo y todas las aportaciones brindadas. Dr. Wenceslao Valenzuela Quiñónez le
agradezco su colaboración para el desarrollo de esta tesis. Dr. Dagoberto Armenta
Bojórquez, gracias por sus valiosas clases. Gracias Dr. José Ángel López Valenzuela el haber
proporcionado el software PDQuest para llevar a cabo el análisis de imágenes de este trabajo.
También agradezco enormemente tu ayuda M.C. Luz Isela Peinado Guevara en el uso de
este software y en el análisis de los resultados, de igual manera agradezco tus enseñanzas y
sugerencias en las metodologías aplicadas en el análisis proteómico. Muchas gracias M.C.
Arlene Guadalupe Mora Romero por proporcionar las muestras de hojas de frijol de dos
líneas de frijol colonizadas y no colonizadas con G. intraradices indispensables en la
realización de este trabajo. Gracias también M. C. Hugo Galindo Flores por ayudarme a
resolver mis dudas y compartir sus conocimientos conmigo.
Este trabajo de tesis también es resultado de personas que participaron indirectamente,
las cuales me demostraron su aprecio y cariño. Un millón de gracias a todos ustedes familia y
amigos por los momentos llenos de risas, lágrimas, sueños, secretos, anhelos y hasta de enojo.
Blanca Elvira te agradezco enormemente tu amistad desde que fuimos niñas. Hemos
compartido muchas etapas de nuestra vida juntas, en la que me has contagiado de tu alegría y

de tu espíritu de lucha. Gracias también Victor y Glenda, porque juntos recorrimos
muchísimas horas de camino, las cuales pudieron haber sido muy aburridas y cansadas, pero
con ustedes fueron superdivertidas. A ti Odeta mi amiga a primera vista, un millón de gracias
por todos los momentos compartidos. Por las riso-terapias que me hicieron tanto bien en los
momentos de estrés y por tu contribución a mi trabajo de tesis al acompañarme a sacar mis
geles durante la noche y vivir esos momentos de miedo en el CIIDIR conmigo. Mil gracias
por las muchas molestias dadas y tu como si nada negra…también les agradezco a Eymer,
Cindy y Libna porque estuvieron presentes en esos momentos tan importantes de mi trabajo
y su apoyo fue incondicional, gracias por su amistad. Muchísimas gracias también a ti Lucky
por todas las cosas que has hecho por mí. Sobre todo gracias por nuestros arrebatos, porque
gracias a ellos nos divertimos mucho, espero que surjan más y a ti Raquel gracias por tu
apoyo, porque aunque tranquila siempre has sido parte del show y nuestra gran amiga. Paola
no podías faltar tu, aunque ha sido poco el tiempo que hemos compartido en esta vida, han
sido momentos llenos de buena vibra y de sabios consejos, gracias por brindarme tu amistad y
por considerarme tu hermanita. Gracias también a mis compañeros de generación: Karla,
Dámaris, Leonardo, Breydi, Magda, Carmen, Rocío y Nataly y del lab: Luis Daniel,
Damian, Alejandro, Claudia María y Claudia Moreno.
Gracias mamá y papá porque su ilusión en la vida ha sido convertir a mis hermanos y
a mí en personas de bien. Porque ustedes sin escatimar esfuerzo alguno, han sacrificado gran
parte de su vida en formarnos y educarnos. Porque ni con las riquezas más grandes del mundo
podré pagarles la familia que tengo. Gracias por todo su apoyo en esta meta de mi vida.
Por último quiero agradecer a mis grandísimos amores: a ti Moisés y a nuestro hijo
Luis Diego. Porque al mirar sus ojos crecen en mi ilusiones de vivir, de alcanzar sueños que
tenemos en común. Gracias por que sin el amor de ustedes no lo hubiera logrado. Perdonen
mis ausencias y mis momentos de frustración y enojo. Los amo, lo saben.
He llegado al final de este camino y en mi han quedado marcadas huellas profundas de
éste recorrido. Son profesores sus palabras y sabios consejos. Son amigos sus sonrisas. Son
madre tu mirada y tu aliento. Son padre tu trabajo y esfuerzo. Son esposo e hijo sus abrazos y
besos. Sin ustedes esto no sería porque, cada uno de ustedes en algún momento de la vida hizo
algo por mi e influyó en mi destino…gracias por participar :)

ÍNDICE RESUMEN. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I
ABSTRACT. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . II
ÍNDICE DE CUADROS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . III
ÍNDICE DE FIGURAS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . IV
GLOSARIO. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . VI
I. INTRODUCCIÓN. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
II. ANTECEDENTES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
2.1. El frijol. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
2.1.1. Azufrado Higuera. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
2.1.2. Azufrado Regional 87. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
2.2. Interacciones planta-microorganismo en la rizosfera. . . . . . . . . . . . . . 5
2.2.1. Micorrizas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
2.2.2. Bioprotección por hongos micorrízicos arbusculares. . . . . . . . . 9
2.2.2.1. Posibles mecanismos de bioprotección. . . . . . . . . . . . . 10
2.2.2.2. Bioprotección por hongos micorrízicos arbusculares en frijol. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
13 2.3. Glomus intraradices. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.4. Proteómica. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
2.4.1. Análisis de proteínas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
2.5. Resistencia sistémica. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
2.5.1. Resistencia sistémica adquirida (RSA). . . . . . . . . . . . . . . . . . . 20
2.5.2. Resistencia sistémica inducida (RSI). . . . . . . . . . . . . . . . . . . . . 21
2.5.3. Resistencia inducida por micorrización (RIM). . . . . . . . . . . . . . 23
2.6. Métodos de genómica y proteómica empleados en el estudio de la interacción planta-microorganismo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
24 III. JUSTIFICACIÓN. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
IV. HIPÓTESIS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
V. OBJETIVOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
5.1. Objetivo general. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
5.2. Objetivos específicos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33

VI. MATERIALES Y MÉTODOS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
6.1. Tejido Vegetal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
6.2. Estandarización del método de extracción de proteínas. . . . . . . . . . . 36
6.2.1. Método A de extracción con Fenol (Fenol-A). . . . . . . . . . . . . . 36
6.2.2. Método B de extracción con Fenol (Fenol-B). . . . . . . . . . . . . . 37
6.2.3. Método A de extracción con Ácido Tricloroacético/Acetona (TCA-A). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
37
6.2.4. Método B de extracción con Ácido Tricloroacético/Acetona (TCA-B). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
38
6.3. Cuantificación de proteínas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38
6.4. Protocolo de limpieza de proteínas para 2D-SDS-PAGE. . . . . . . . . . . . 39
6.5. Separación de proteínas por punto isoeléctrico [Isoelectroenfoque (IEF)]. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
39
6.6. Separación de proteínas por peso molecular (SDS-PAGE). . . . . . . . 40
6.7. Adquisición y análisis de imágenes. . . . . . . . . . . . . . . . . . . . . . . . . . . 41
6.8. Identificación de proteínas mediante espectrometría de masas. . . . . . 41
VII. RESULTADOS Y DISCUSIÓN. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
7.1. Estandarización del método de extracción de proteínas. . . . . . . . . . . . 43
7.2. Electroforesis bidimensional (2D-SDS-PAGE). . . . . . . . . . . . . . . . . . 46
7.3. Análisis proteómico de plantas colonizadas con G. intraradices vs. no colonizadas en dos líneas de frijol. . . . . . . . . . . . . . . . . . . . . . . . . . . . .
49
7.3.1. Extracción en hojas de frijol. . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
7.3.2. Perfil de proteínas inducido en hojas de frijol. . . . . . . . . . . . . . 50
7.4. Interpretación del papel funcional de las proteínas implicadas en la simbiosis micorrízica arbuscular. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
61
7.4.1. Análisis proteómico en hojas de plantas de frijol de la línea Az. Higuera susceptible y Az. Regional 87 tolerante a S. sclerotiorum en asociación con G. intraradices. . . . . . . . . . . . . . . . . .
63
VIII. CONCLUSIONES. . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 68
IX. RECOMENDACIONES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70
X. BIBLIOGRAFÍA. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71

Quintero-Zamora, 2010 I
RESUMEN
En los últimos años se han reportado diversos estudios sobre la resistencia inducida
por micorrización al ataque de patógenos, en éstos se han determinado los cambios de
expresión de genes y proteínas que ocurren en las raíces de plantas colonizadas por hongos
micorrízicos arbusculares. Recientemente, se han iniciado estudios para entender cómo
afecta esta interacción a la parte aérea de la planta. En nuestro grupo de investigación se
llevó a cabo el estudio del efecto de la micorrización con Glomus intraradices en la
tolerancia al moho blanco causado por Sclerotinia sclerotiorum en tres variedades de frijol.
Los resultados obtenidos muestran una resistencia inducida con G. intraradices a S.
sclerotiorum en dos de las tres variedades probadas. Para identificar proteínas diferenciales
en la parte aérea de variedades de frijol con capacidad contrastante para inducir la
tolerancia a patógenos por colonización micorrízica, en el presente trabajo primeramente se
evaluaron cuatro diferentes métodos de extracción de proteínas ya reportados para otros
tejidos vegetales. La comparación cuantitativa de extractos de proteínas reveló que el
método de extracción con ácido tricloroacético-acetona fue el mejor en cuanto a cantidad y
calidad de proteínas resueltas en el análisis 2D-SDS-PAGE. Posteriormente, se realizó un
análisis proteómico comparativo entre plantas de frijol de las variedades Azufrado Higuera
(susceptible a S. sclerotiorum, cuando es colonizada con G. intraradices) y Azufrado
Regional 87 (tolerante a S. sclerotiorum, cuando es colonizada con G. intraradices)
colonizadas y no colonizadas con el hongo G. intraradices. El análisis del perfil proteico de
dos variedades de frijol a estudiar en las diferentes condiciones, mostró un promedio de
200 proteínas en un rango de pH 4-7 y una masa molecular de 10 a 100 kDa. El análisis
proteómico (colonizada vs. no colonizada) de la variedad Az. Higuera, reveló cinco
proteínas diferenciales, las cuales disminuyeron su expresión en plantas colonizadas. En
cambio, la variedad Az. Regional 87 reveló una proteína que sólo se encuentra en la
condición colonizada, once aumentan su expresión relativa y dos la disminuyen en cuanto a
plantas colonizadas vs. no colonizadas. Los resultados de identificación mediante
espectrometría de masas, muestran once proteínas cuya función se conoce, una con función
desconocida y dos proteínas no reportadas en tejidos vegetales. Tripsinógeno y
transcetolasas fueron reguladas de manera contrastante entre las variedades. La mayoría de
las proteínas diferencialmente inducidas están relacionadas a fotosíntesis.

Quintero-Zamora, 2010 II
ABSTRACT
In recent years, there have been several studies on mycorrhiza-induced resistance to
pathogen attack, in which changes of expression of genes and proteins that occur in the
roots of arbuscular mycorrhizal fungi (AMF)-colonized plants have been determined.
Recent studies have begun to deal with the question of how this interaction affects plant
shoots. In our research group, the effect of mycorrhiza colonization with Glomus
intraradices on the induction of tolerance to white mold caused by Sclerotinia sclerotiorum
in three varieties of beans have been investigated. Results showed an induced tolerance to
S. sclerotiorum in colonized plants in two out of the three varieties tested. In order to
identify differential proteins between AMF-colonized and non colonized plants, first, four
protein extraction methods reported for other plant tissues were evaluated. Quantitative
comparison of protein extracts revealed that trichloroacetic acid-acetone extraction method
was the best in terms of quantity and quality of proteins resolved by 2D-SDS-PAGE
analysis. Subsequently, we conducted a comparative proteomic analysis of shoots of the
varieties Azufrado Higuera and Azufrado Regional 87. Azufrado Higuera showed
susceptibility to S. sclerotiorum, when colonized with G. intraradices, whereas Azufrado
Regional 87 was able to induce tolerance to this pathogen, when colonized with the AMF.
Protein profile analysis of these two bean varieties under colonized and non colonized
conditions showed an average of 200 proteins in the range of pH 4-7 and molecular mass of
10 to 100 kDa. Proteomic analysis on Az. Higuera, revealed five differential proteins that
showed decreasing expression in AMF-colonized plants. Instead, the variety Az. Regional
87 revealed a protein found only in colonized plants, whereas eleven increased their relative
expression, and two showed a decrease in relative expression in colonized vs. non-
colonized plants. Identification by mass spectrometry showed eleven proteins whose
function is already known, whereas putative function of one of them is currently unknown,
and two proteins have not been reported in plant tissues. Trypsinogen and transketolases
were regulated in a contrasting manner between the two varieties. Most of the induced
proteins are related to photosynthesis.

Quintero-Zamora, 2010 III
ÍNDICE DE CUADROS
Cuadro 1 Rendimientos de extracción de proteínas de hojas de plantas
mediante cuatro diferentes métodos. . . . . . . . . . . . . . . . . . . . . . .
44
Cuadro 2 Proteínas (spots) resueltas por cada uno de los diferentes
métodos de extracción. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
49
Cuadro 3 Valores obtenidos por el software PDQuest en pixeles, así
como el promedio y desviación estándar de estos para las
diferentes condiciones. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
54
Cuadro 4 Relación de densidad de proteínas expresadas diferencialmente
en hojas de plantas de frijol Az. Higuera. . . . . . . . . . . . . . . . . . .
57
Cuadro 5 Relación de densidad de proteínas expresadas diferencialmente
en hojas de plantas de frijol Az. Regional 87. . . . . . . . . . . . . . . .
57
Cuadro 6 Proteínas diferenciales como resultado de la colonización con
HMA en hojas de plantas de frijol variedad Az. Higuera. . . . . .
59
Cuadro 7 Proteínas diferenciales como resultado de la colonización con
HMA en hojas de plantas de frijol variedad Az. Regional 87. . .
60

Quintero-Zamora, 2010 IV
ÍNDICE DE FIGURAS
Figura 1 Esquema de los diferentes pasos involucrados en un análisis de
proteómica comparativa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
16
Figura 2 Plantas de frijol de la variedad Az. Regional 87, mantenidas en
estantes cubiertos con mallas antiáfidos bajo condiciones de
invernadero. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
34
Figura 3 Plantas de frijol de las variedades Az. Regional 87 y Az. Higuera,
cultivadas bajo condiciones controladas en cámaras de
crecimiento. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
35
Figura 4 Curva estándar para determinar la concentración de proteína. . . . . 44
Figura 5 Análisis por SDS-PAGE de proteínas extraídas por diferentes
métodos de extracción. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
45
Figura 6 Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el
método de extracción con Fenol-A. . . . . . . . . . . . . . . . . . . . . . . . .
47
Figura 7 Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el
método de extracción con Fenol-B. . . . . . . . . . . . . . . . . . . . . . . . .
47
Figura 8 Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el
método de extracción con TCA-A. . . . . . . . . . . . . . . . . . . . . . . . .
48
Figura 9 Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el
método de extracción con TCA-B. . . . . . . . . . . . . . . . . . . . . . . . . .
48
Figura 10 Análisis mediante electroforesis en gel desnaturalizante (SDS-
PAGE) de proteínas extraídas de hoja mediante el método de
extracción TCA-A. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
50
Figura 11 Geles preparativos bidimensionales (2D-SDS-PAGE) de plantas
de frijol (Phaseolus vulgaris) línea Az. Regional 87 colonizadas
con Glomus intraradices. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
51
Figura 12 Geles preparativos bidimensionales (2D-SDS-PAGE) de plantas
de frijol (Phaseolus vulgaris) línea Az. Regional 87 no
colonizadas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
51

Quintero-Zamora, 2010 V
Figura 13 Geles preparativos bidimensionales (2D-SDS-PAGE) de plantas
de frijol (Phaseolus vulgaris) línea Az. Higuera colonizadas con
Glomus intraradices. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
52
Figura 14 Geles preparativos bidimensionales (2D-SDS-PAGE) de plantas
de frijol (Phaseolus vulgaris) línea Az. Higuera no colonizadas. . .
52
Figura 15 Gel maestro comparativo entre las proteínas de la línea de frijol
Az. Higuera, colonizadas con G. intraradices vs. no colonizadas,
obtenido en el programa PDQuest . . . . . . . . . . . . . . . . . . . . . . . . . .
55
Figura 16 Gel maestro comparativo entre las proteínas de la línea de frijol
Az. Regional 87, colonizadas con G. intraradices vs. no
colonizadas, obtenido en el programa PDQuest. . . . . . . . . . . . . . . .
56

Quintero-Zamora, 2010 VI
GLOSARIO
Arbustivo tipo I. Las variedades arbustivo tipo uno se caracterizan por tener
floración y maduración uniformes, lo que en parte se debe a su hábito de
crecimiento compacto y en algunos casos a su respuesta neutral al fotoperíodo,
aunque su potencial de rendimiento es menor que otros tipos (II, III, IV).
Bioprotección. Protección conferida a las plantas, contra estrés biótico o abiótico,
por microorganismos benéficos como hongos micorrízicos arbusculares o
rizobacterias promotoras de crecimiento vegetal.
Elicitor. Son moléculas del patógeno o sustancias químicas, que actúan como
receptores de las plantas, activando en ellas respuestas de defensa.
Fotobionte. Todo organismo capaz de absorber la energía de luz y transformarla en
energía química.
Hábito de crecimiento. Las plantas se clasifican por su hábito de crecimiento en
erectas (plantas con tallos de crecimiento erecto), rastreras (plantas cuyos tallos
crecen tendidos sobre la superficie del suelo) y trepadoras (plantas que interfieren
con el cultivo, con tallo de crecimiento oblicuo). De acuerdo con el hábito de
crecimiento que presentan sus plantas, los cultivares de fríjol son agrupados en
cuatro tipos principales: Hábito de crecimiento determinado arbustivo (Tipo I),
hábito de crecimiento indeterminado arbustivo (Tipo II), hábito de crecimiento
indeterminado postrado (Tipo III) y hábito de crecimiento indeterminado trepador
(Tipo IV).
Hifa. Elementos filamentosos cilíndricos característicos que en conjunto forman el
micelio de la mayoría de los hongos.

Quintero-Zamora, 2010 VII
Micorriza. Asociación simbiótica que se establece entre raíces de una planta y un
hongo micorrízico.
Preacondicionamiento o “priming”. Estado fisiológico causado en plantas por
algunos compuestos o microorganismos benéficos, en el cual la planta se potencía
contra diferentes tipos de estrés como el ataque de un patógeno.
Resistencia sistémica. Estado fisiológico de aumento de la capacidad defensiva en
plantas, dado por un elicitor de manera local y la señal es transportada a zonas
distales de la planta.
Simbiosis. Asociación de dos o más organismos de distintas especies, en la que
todos salen beneficiados.
Susceptible. Que no tiene capacidad para resistir enfermedades ocasionadas por
organismos patógenos.
Tolerante. Que tiene capacidad para resistir enfermedades ocasionadas por
organismos patógenos.
Virosis. Son todas aquellas patologías provocadas por virus en plantas.

Quintero-Zamora, 2010 1
I. INTRODUCCIÓN
El frijol común (Phaseolus vulgaris L.) es originario de América, es una especie
dicotiledónea anual perteneciente a la familia de las fabáceas, antiguamente conocida como
familia de las papilionáceas. El frijol es la leguminosa de grano más consumida y estudiada
en el mundo por ser una de las fuentes principales de proteínas, así como formar parte
importante de los hábitos alimentarios de la población. Su mayor área de producción se
encuentra concentrada en Asia donde se localiza cerca del 50% de la producción mundial y
en el Continente Americano el cual participa aproximadamente con un 25%. La producción
de frijol en el mundo se concentra en 129 países de los cinco continentes. Entre los países
productores de la leguminosa destacan la India con un 26%, Brasil 23%, Myanmar 17%,
Estados Unidos con 8%, y México en quinto lugar con un 8%. Estas naciones, junto con
China, contribuyeron con el 80% producido, según la FAO (2008). En México, los
principales estados productores de frijol son Zacatecas, Durango y Chihuahua, en el ciclo
primavera-verano; y Nayarit y Sinaloa en el ciclo otoño-invierno. En Sinaloa en el ciclo
agrícola otoño-invierno 2009 se sembraron 94,650.07 hectáreas (ha), obteniendo un total de
162,344.21 toneladas (t) de grano, lo cual representa la mayor superficie sembrada del país
en ese ciclo (SAGARPA, 2009). Por lo que el cultivo de frijol ocupa un lugar importante en
la economía del estado.
La producción de frijol común es afectada por factores bióticos y abióticos. Los
factores bióticos incluyen: plagas, enfermedades y malezas. Los factores abióticos
incluyen: calor, sequía, suelos ácidos, baja fertilidad del suelo y salinidad (Rodríguez,
1996). Dentro de los factores bióticos también existen microorganismos en el suelo que
actúan principalmente en la rizosfera. Estos microorganismos interaccionan de diferente
manera con las plantas; en algunos casos actúan como patógenos y en otros establecen
interacciones benéficas con las mismas (Azcón-Aguilar y Barea, 1996; Peterson et al.,
2004). Dentro de las interacciones benéficas microorganismo-planta más importantes están
la simbiosis con bacterias fijadoras de nitrógeno y la simbiosis con hongos micorrízicos
arbusculares (HMA). Ambos tipos de interacción se pueden llevar a cabo con plantas
leguminosas y ambas corresponden a asociaciones complejas lo cual requiere de una
participación coordinada entre los organismos que interactúan (Gamalero et al., 2004).

Quintero-Zamora, 2010 2
La interacción de la micorriza arbuscular (MA) es una asociación mutualista que se
establece entre un grupo de hongos pertenecientes al filo Glomeromycota y la gran mayoría
de las plantas (Schuessler et al., 2001). Aproximadamente el 80% de todas las especies de
plantas terrestres pueden formar este tipo de asociación. En esta simbiosis, el hongo provee
fósforo, nitrógeno y otros nutrimentos poco móviles a la planta y la planta a cambio le
brinda carbohidratos al hongo (citado en Hause et al., 2002). Además de las ventajas
nutricionales que la asociación brinda a las plantas, se ha observado que la colonización
micorrízica confiere tolerancia a patógenos tanto de raíz (Bodker et al., 2002; Colditz et al.,
2005; Elsen et al., 2008) como de parte aérea (Liu et al., 2007; Galindo-Flores, 2008;
Mora-Romero, 2008).
En base a la resistencia inducida por micorrización (RIM) al ataque de patógenos,
en los últimos años se han reportado diversos estudios donde se han determinado los
cambios de expresión de genes y proteínas que ocurren en las raíces de plantas colonizadas
por HMA (Salzer et al., 2000; Liu et al., 2003; Whipps, 2004; Colditz 2004, 2005; Liu et
al., 2007). Sin embargo, pocos son los estudios encaminados a entender cómo afecta esta
interacción la parte aérea de la planta (Liu et al., 2007; Fiorilli et al., 2009). Se ha sugerido
que la micorrización promueve la resistencia a patógenos mediante la acumulación de
proteínas en estado inactivo, de tal manera que una vez enfrentada la planta colonizada con
HMA con el patógeno, éstas proteínas son activadas y la respuesta de defensa es mucho
más rápida e intensa. Se ha planteado que en el establecimiento de la asociación MA deben
participar señales moleculares que se intercambien entre la planta y el hongo (Salzer y
Boller, 2000). Se ha encontrado que en plantas colonizadas con HMA se produce una
respuesta transitoria, similar a la producida por patógenos, de acumulación de proteínas
relacionadas a patogenicidad, la cual es atenuada a medida que avanza la colonización
(Dumas-Gaudot et al., 2000).
En el Laboratorio de Interacción Microorganismo-Planta del Departamento de
Biotecnología Agrícola del CIIDIR-Sinaloa, se llevó a cabo el estudio del efecto de la
micorrización con Glomus intraradices en la tolerancia al moho blanco causado por
Sclerotinia sclerotiorum en tres variedades de frijol por Mora-Romero (2008). Los
resultados obtenidos muestran una tolerancia inducida por micorrización con G.
intraradices a S. sclerotiorum en dos de las tres variedades probadas.

Quintero-Zamora, 2010 3
En el presente trabajo de tesis de maestría se planteó realizar un análisis proteómico
comparativo entre plantas de frijol colonizadas y no colonizadas con el hongo G.
intraradices. Este estudio permitirá sentar las bases para el entendimiento del mecanismo
molecular de la comunicación sistémica que se desencadena en una planta de frijol cuando
es colonizada con un hongo micorrízico y con ésto entender el mecanismo de tolerancia
inducida por micorrización en respuesta al ataque por patógenos.

Quintero-Zamora, 2010 4
II. ANTECEDENTES
2.1. El frijol
El frijol común (Phaseolus vulgaris L.) es una planta anual, herbácea, intensamente
cultivada desde zonas tropicales hasta templadas. Se le conoce con diferentes nombres:
poroto, haricot, caraota, judía, aluvia, habichuela y otros. El frijol es una de las especies de
la familia de las leguminosas más producidas y consumidas a nivel mundial, el grano
presenta alto contenido de proteínas, vitaminas y minerales para la dieta del humano
(Broughton et al., 2003). En México, en el año agrícola 2009, se sembraron
aproximadamente un millón 676 mil ha del cultivo de frijol, ocupando el tercer lugar en
superficie sembrada, después del sorgo y del maíz (SAGARPA, 2009). La producción de
frijol en nuestro país empezó en el año de 1943, con la primera colecta de germoplasma
nativo, mismo que ha sido recombinado con germoplasma exótico, razón por la cual existen
cultivares muy diversos, tanto por características agroecológicas como por demanda de
consumo (Rosales-Serna et al., 2005). De 1943 al 2000, el Instituto Nacional de
Investigación Forestal, Agrícola y Pecuaria (INIFAP) liberó 120 variedades mejoradas
(Rosales-Serna et al., 2003, 2004).
En cuanto a las variedades utilizadas para el presente trabajo, existe también
variabilidad entre ellas.
2.1.1. Azufrado Higuera
Azufrado Higuera (CCZT 10163) es producto de la cruza entre Canario DIVEX
8130 y Royal Red, realizada en 1981, en el Centro Internacional de Agricultura Tropical
(CIAT), a través del programa de intercambio de germoplasma y cruzas entre INIFAP y el
CIAT; este material fue introducido en 1982 al Campo Agrícola Experimental Valle del
Fuerte (Salinas-Pérez et al., 1995). Bajo las condiciones del Valle del Fuerte se hicieron
selecciones en forma individual y masiva hasta la sexta generación filial (F6), siendo su
nomenclatura CCZT 10163-4-2-CB-(10)-M-M. Azufrado Higuera adquirió de Canario-
Divex 8130 (originario del Perú), su ciclo vegetativo intermedio, adaptación y color de
grano amarillo intenso; mientras que de Royal Red (originario de EUA) su hábito de
crecimiento determinado arbustivo tipo I, tolerancia a virosis, tamaño y uniformidad de

Quintero-Zamora, 2010 5
color de grano, con un peso de cien semillas que varía de 43.5 a 55.3 gramos, propiedad por
la cual se distingue de las demás variedades dentro de su clase comercial (Salinas-Pérez et
al., 1998). En cuanto a su capacidad de rendimiento, esta variedad ha rendido desde 2,500 a
3,300 kg ha-1. Azufrado Higuera manifiesta sus más altos rendimientos en el Norte de
Sinaloa, con una alta calidad de grano, propiedades por las cuales se distingue dentro de las
demás variedades dentro de su calidad comercial e inclusive ha penetrado en el mercado de
exportación generando una alta demanda tanto en el mercado interno como exterior. Esta
variedad es resistente a royas, tolerante a virus, pero susceptible al moho blanco causado
por S. sclerotiorum, sin embargo puede escapar a la enfermedad por su hábito de
crecimiento el cual no es rastrero (Salinas-Pérez y Rodríguez-Cota, 2008).
2.1.2. Azufrado Regional 87
Azufrado Regional 87 nomenclatura II 8 Fr Mo-5-3-M-M y registro FRI-043-
150228, es un cultivar logrado por la cruza entre Canario 101 y Azufrado 200, realizada en
el Campo Experimental del Valle del Fuerte (INIFAP-CEVAF) durante el ciclo otoño –
invierno 1979-80, seleccionándose hasta F6, adquiriendo de canario 101 su ciclo vegetativo
precoz, amplia adaptación y potencial de rendimiento, mientras que de Azufrado 200 su
hábito de crecimiento arbustivo tipo I, tolerante a virosis y tipo de grano amarillo claro
(azufrado). Su peso promedio es de 34.2 g por 100 semillas, con un rendimiento promedio
observado de 2,400 kg ha-1. Azufrado Regional 87 fue liberado en 1988, es resistente a
roya, tolerante a virosis, susceptible a la antracnosis, tizón común y al moho blanco aunque
puede escapar a esta enfermedad debido a su hábito de crecimiento determinado semierecto
(Salinas-Pérez et al., 1989).
2.2. Interacciones planta-microorganismo en la rizosfera
La rizosfera es el volumen de suelo que está influenciado directamente por las raíces
de las plantas, las cuales liberan gran cantidad de exudados y producen una estimulación
muy significativa en la actividad y densidad microbiana (citado en Gamalero et al., 2004).
Los microorganismos interaccionan de diferente manera con las plantas; en algunos casos
actuando como patógenos, y en otros estableciendo interacciones mutualistas, es decir
beneficiosas para ambos. Dentro de las interacciones planta-microorganismo benéficas se

Quintero-Zamora, 2010 6
encuentran la simbiosis con bacterias fijadoras de nitrógeno y la simbiosis MA (Deguchi et
al., 2007). Los principales nutrientes que aportan estos microorganismos simbióticos a la
planta huésped son los macronutrientes fósforo (P) y en menor medida nitrógeno (N). Esta
simbiosis tiene una influencia importante en la fisiología de la planta e incluso en la
función del ecosistema.
2.2.1. Micorrizas
La palabra micorriza proviene del griego “myco”, hongo y “rhiza”, raíz; y
literalmente puede traducirse como “raíz fungosa”. El término fue utilizado por primera vez
en 1985 para describir la simbiosis entre raíces de las plantas y los hongos del suelo
(Munyanziza et al., 1997). Existen dos tipos de micorrizas, las ectomicorrizas, en las que el
tejido fúngico micelial coloniza la raíz de la planta de manera extracelular y las
endomicorrizas o micorrizas arbusculares, en donde las hifas que invaden el tejido radical
penetran las células de la raíz para establecer una simbiosis intracelular (Bonfante, 2003;
Peterson et al., 2004).
La micorriza arbuscular (MA) es una simbiosis que se presenta entre un grupo de
hongos y las raíces de las plantas. Estos hongos pertenecen a un filo nuevo, descrito de
manera independiente: el Glomeromycota (Schuessler et al., 2001). Hasta ahora se
encuentran clasificados en cuatro subórdenes: Archaeosporales, Paraglomerales,
Diversisporales y Glomerales. Ocho géneros de HMA han sido reconocidos basados
principalmente en características morfológicas de esporas asexuales. Estos géneros,
Glomus, Paraglomus, Sclerocystis, Scutellospora, Gigaspora, Acaulospora, Archaeospora
y Entrophospora incluyen aproximadamente 150 especies (Peterson et al., 2004). Sólo
algunas familias de las plantas superiores no son capaces de formar este tipo de asociación,
tales como miembros de la familia Brassicaceae, Caryophyllaceae, Urticaceae y
Amaranthaceae (Bouwmeester et al., 2007). Trabajos realizados en el Laboratorio de
Ecología Molecular del CIIDIR-Sinaloa indican que ciertas especies de la familia
Amaranthaceae se han encontrado colonizadas con HMA (Maldonado-Mendoza,
comunicación personal).
Los registros fósiles más antiguos indican que dicha simbiosis tiene más de 460
millones de años, lo cual los ubica en el período Devónico. Esto ha llevado a considerar la

Quintero-Zamora, 2010 7
compleja coevolución de las plantas y estos hongos asociados como un mecanismo para
que las plantas acuáticas hayan podido colonizar el ambiente terrestre. Esta asociación tan
antigua se manifiesta en la amplia distribución de la simbiosis pues se estima que hasta el
80% de las plantas terrestres pueden asociarse con HMA.
Las micorrizas arbusculares constituyen una asociación que se forma con la mayoría
de las especies de plantas de interés agrícola. Durante la simbiosis, la planta hospedera
recibe nutrientes minerales del suelo tomados por el hongo (principalmente fósforo),
mientras que éste obtiene compuestos de carbono derivados de la fotosíntesis (Brundrett et
al., 1996). Esta simbiosis se considera de tipo mutualista, en la cual el hongo es un
simbionte de tipo obligado, es decir que no puede completar su ciclo de vida sin la planta y
requiere siempre asociarse a ella para completar su ciclo de vida. Esto implica que no puede
ser cultivado in vitro de manera pura, siempre debe estar asociado a algún hospedero
vegetal.
El proceso de formación de la simbiosis MA comienza cuando la raíz secreta
algunas sustancias que promueven la germinación de las esporas del hongo en el suelo. Las
hifas que forman el micelio crecen hasta encontrar el área de la rizosfera de una planta
hospedera. Cuando la hifa alcanza la epidermis de la raíz se forma una estructura de anclaje
denominada apresorio. A partir de éste, la hifa empieza a crecer penetrando las células de la
epidermis de la raíz hasta alcanzar las células corticales internas. En estas células, el
extremo distal de las hifas es invaginado dentro de la célula vegetal cortical sin causar la
ruptura de la membrana vegetal. Posteriormente se ramifica abundantemente para formar
unas estructuras denominadas arbúsculos, que son las estructuras donde se realiza el
intercambio de nutrientes entre el hongo y la planta. Los arbúsculos están envueltos por
membranas de origen vegetal y fúngica denominada por el espacio peri-arbuscular. La
transferencia de nutrientes entre los simbiontes se lleva a cabo en los arbúsculos y
particularmente a través de la membrana peri-arbuscular vegetal (Peterson et al., 2004;
Harrison, 2005). Dentro de la raíz colonizada, en el micelio interno, algunos hongos
micorrízicos forman vesículas, las cuales son estructuras que contienen reservas lipídicas,
que le sirven al hongo para responder a la falta de nutrientes si la planta es de ciclo anual o
al final de su ciclo de vida usando toda esa energía acumulada para la producción de
esporas. Estos hongos además forman una red micelial externa en donde se encuentran

Quintero-Zamora, 2010 8
hifas especializadas en la absorción de nutrientes en el suelo. Es en el micelio externo
donde la mayoría de las especies forman esporas asexuales las cuales son estructuras de
reproducción y de resistencia. A la fecha, no se ha demostrado la existencia de fases
sexuales en estos organismos.
Actualmente son bien conocidos los efectos beneficiosos de las MA, las
investigaciones muestran que los HMA dirigen la diversidad y productividad de sus plantas
hospederas en diferentes ecosistemas. Uno de los principales beneficios comprende la
mayor absorción de elementos poco móviles en el suelo como el fósforo, cobre y zinc por
parte de las plantas colonizadas por HMA en comparación con las no colonizadas (Smith y
Read, 1997). A cambio de recibir algunos nutrientes minerales, la planta le brinda al hongo
una fuente de carbono en forma de glucosa y otros carbohidratos de tipo hexosa, para que
este pueda desarrollarse y completar su ciclo de vida. El fósforo es el nutrimento mejor
estudiado en esta asociación, ya que es el más importante para que esta asociación se lleve
a cabo. Junto con el nitrógeno, es de los nutrientes que la planta requiere en mayores
cantidades. Concentraciones bajas de fósforo favorecen la asociación micorrízica
arbuscular, en cambio concentraciones elevadas del mismo en el suelo pueden inhibir la
formación de la asociación. El hongo capta fósforo en forma de fosfato, que es la manera
asimilable del fósforo para la planta en el suelo, lo acumula en su micelio, lo transporta y
transloca a la planta. Recientemente también se encontró que los HMA también podrían
participar mejorando el metabolismo de nitrógeno en la planta (Leigh et al., 2009). Gracias
al uso más eficiente que hacen las plantas colonizadas con HMA de los nutrientes del suelo,
permiten disminuir el uso de fertilizantes químicos y reducir por consiguiente los
problemas de contaminación que el uso excesivo de fertilizantes conlleva. Por otra parte,
las plantas colonizadas con HMA son capaces de hacer un mejor uso de los fertilizantes
orgánicos, bien sea debido a la producción de fosfatasas por parte de los mismos hongos, lo
cual hace biodisponible el fósforo presente en el suelo (Dodd et al., 1987; Joner y Johansen,
2000), o bien gracias a la asociación existente entre las hifas de los HMA y los
microorganismos que participan en la mineralización de la materia orgánica (Azcón-
Aguilar y Barea, 1992). También son conocidos los efectos de las MA en la formación de la
estructura del suelo, a través de su papel en la constitución de agregados de suelo estables
al agua, en los que el micelio externo de los HMA tiene una notable participación (Miller y

Quintero-Zamora, 2010 9
Jastrow, 2000). Esto se logra a través de la producción de una glicoproteína llamada
glomalina, la cual por sus características químicas favorece la agregación de las partículas
de suelo (Rillig, 2004).
A pesar de que existe una gran información sobre esta simbiosis, aún se desconocen
detalles sobre los mecanismos de adaptación de estos hongos, así como su relación con
otros microorganismos y aspectos fundamentales de su biología.
2.2.2. Bioprotección por hongos micorrízicos arbusculares
Los HMA tienen un impacto significativo en la salud y nutrición mineral del
fotobionte y aunque los mecanismos moleculares sean en gran parte desconocidos, ha sido
demostrado que los HMA pueden translocar el fosfato del suelo a la raíz de la planta (citado
en Maldonado-Mendoza et al., 2001). Las micorrizas constituyen una asociación
multifuncional con las plantas, cuyos beneficios van más allá de los aspectos nutricionales.
Además de las ventajas nutricionales que un hongo micorrízico arbuscular confiere a la
planta, existe evidencia en la literatura que indica que esta asociación aumenta la habilidad
del fotobionte para tolerar diferentes tipos de estrés tanto abióticos como bióticos (Pozo y
Azcón-Aguilar, 2007).
Los HMA protegen a las plantas del déficit hídrico (Ruiz-Lozano y Azcón, 1995) y
también existen diversos estudios sobre la bioprotección conferida por HMA a las plantas
ante el ataque de patógenos. El control biológico de patógenos agronómicamente
importantes ha sido consistentemente demostrado en muchos casos. Estos incluyen especies
de hongos patógenos del suelo como: Aphanomyces, Fusarium, Phytophthora, Pythium,
Rhizoctonia, Sclerotinia, Sclerotinium, Verticillium entre otras. Incluso el impacto de los
HMA como agentes de biocontrol ha sido demostrado también para especies de nemátodos
como Heterodera, Meloidogyne, Pratylenchus, y Radopholus. Sin embargo, la bioproteción
conferida por HMA no es efectiva para todos los hongos patógenos de plantas, y el nivel de
control biológico conferido por la colonización de HMA es específico de la especie de
planta y aislados específicos de HMA. La simbiosis MA puede mejorar la resistencia o
tolerancia del huésped para patógenos de raíz, aunque el grado de mejoramiento varía
(citado en Harrier y Watson, 2004).

Quintero-Zamora, 2010 10
La reducción de la enfermedad entre las plantas hospederas colonizadas por HMA
es el resultado de un complejo mecanismo de interacciones entre planta, patógeno y hongo
micorrízico. Por lo que distintos mecanismos alternativos han sido propuestos.
2.2.2.1. Posibles mecanismos de bioprotección
Se han propuesto algunos posibles mecanismos para explicar la bioprotección
conferida por HMA. Uno de ellos sugiere que la bioprotección se debe a una mejora del
estatus en los nutrientes minerales de la planta. El incremento en crecimiento de la planta,
seguido de la colonización de la raíz por HMA, es normalmente debido a una mejora del
estado de nutrición mineral de la planta. Dependiendo del aislado de hongo micorrízico y
de la planta huésped, la colonización del sistema radicular por HMA puede incrementar la
nutrición por fósforo y otros nutrientes minerales (Clark y Zeto, 2000). La susceptibilidad
de la planta huésped a infecciones por patógenos puede también ser influenciada por el
estado nutricional del huésped y el estado de fertilidad del suelo (Bødker et al., 1998). Sin
embargo, hay trabajos en donde la mejora de nutrientes en plantas colonizadas con HMA,
en particular fósforo no tiene un efecto en la bioprotección contra especies de patógenos
como S. sclerotiorum y X. campestris (Galindo-Flores, 2008; Mora-Romero, 2008). Por lo
tanto, la mejora por nutrición mineral en simbiosis micorrízica no aclara la bioprotección
conferida por HMA a sus plantas hospederas (Harrier y Watson, 2004). La alteración en la
morfología del sistema radicular de las plantas por la colonización de HMA es otro
mecanismo propuesto. Sin embargo, Norman et al. (1996) demostró que la alteración
inducida con un HMA a la estructura de la raíz en plantas de fresa no se encontró
directamente involucrada en la bioprotección contra Phytophthora fragariae Hickman. La
relación entre la colonización con el HMA y el tiempo de vida de la raíz es dependiente de
la especie, y pocos resultados han sido publicados. Hodge et al. (2000) trabajando en
Plantago lanceolata L. encontraron que el tiempo de vida de la raíz no es afectada por la
inoculación con HMA. Black (1997) observó que el tiempo de vida de la raíz de trébol
(Trifolium repens L.) se incrementa cuando las raíces fueron colonizadas con un HMA.
Cualquier cambio en la longevidad de la raíz resultante de la colonización por HMA puede
estar vinculado a los cambios en la morfología de la raíz como se describe anteriormente.
Cuando los HMA colonizan el sistema radicular de la planta se observan cambios

Quintero-Zamora, 2010 11
morfológicos, así como también un aumento en la lignificación de pared celular de la planta
huésped como resultado de la elevación del metabolismo fenólico, traduciéndose en un
incremento en la bioprotección contra patógenos. Otro mecanismo comprende la
competencia por sitios de colonización e infección por los HMA en relación a los hongos
patógenos del suelo y nemátodos parásitos de plantas que ocupan los mismos sitios en la
raíz y por lo tanto compiten directamente desde el punto de vista espacial cuando la
colonización está ocurriendo al mismo tiempo (Smith, 1988). Aunque los HMA y
patógenos colonicen al mismo tejido huésped, comúnmente se desarrollan dentro de
diferentes células de la raíz, indicando la competencia por espacio (Azcón-Aguilar y Barea,
1996). El número de sitios de infección en un sistema de raíces particular determinará el
alcance del ingreso del patógeno. Vigo et al. (2000) reportaron que el número de sitios de
infección se reduce en los sistemas de raíces colonizadas por HMA y que la colonización
por el HMA no tiene efecto perceptible sobre la propagación de la necrosis. Esto sugiere
que el número de sitios de colonización del HMA es importante para infecciones
subsecuentes por patógenos. Un mecanismo adicional podría involucrar modificaciones de
las poblaciones microbianas del suelo. Las plantas colonizadas por HMA difieren de las no
colonizadas en la composición de la comunidad microbiana de la rizosfera. El número de
bacterias anaerobias facultativas como Pseudomonas fluorescens y diferentes especies de
Streptomyces varían dependiendo de la interacción entre la planta hospedera y el HMA
específico (Marschner et al., 1997). El establecimiento de la comunidad de la rizosfera
altamente específica puede tener un impacto importante en el subsecuente establecimiento
de organismos patogénos de plantas.
Aunque la colonización micorrízica se considera normalmente como benéfica para
la planta, la reacción de la planta puede ser positiva o negativa, dependiendo de parámetros
genéticos y ambientales (Taylor y Harrier, 2001).
Uno de los mecanismos más importantes que se ha propuesto es el de la activación
de respuestas de defensa de la planta por HMA. Varios genes y productos de las enzimas
correspondientes, tales como metabolitos involucrados en respuestas de defensa en plantas
han sido ampliamente estudiados en la simbiosis MA y han mostrado ser expresados parcial
y temporalmente. Esto incluye fitoalexinas, deposición de callosa, glicoproteínas ricas en
hidroxiprolina, fenoles, peroxidasas, quitinasas, β-1-3 glucanasas y proteínas PR

Quintero-Zamora, 2010 12
relacionadas a patogénesis (citado en Harrier y Watson, 2004). La bioprotección conferida
por HMA en sistemas de raíces de plantas hospederas, es tanto localizada como sistémica.
El establecimiento de la simbiosis MA puede predisponer a la planta a responder más
rápidamente al ataque de patógenos. Cordier et al. (1998) y Pozo et al. (2002) demostraron
que la resistencia inducida contra el patógeno Phytophthora parasitica en raíces
colonizadas con HMA de tomate se debió a respuestas de defensa localizadas en células
conteniendo arbúsculos y respuestas de defensa sistémica en raíz en sitios tanto colonizados
como sitios no colonizados.
Existe evidencia experimental que la asociación MA confiere bioprotección contra
fitopatógenos de raíz. Los HMA tienen la habilidad de inducir resistencia sistémica contra
nemátodos en raíz. La presencia de G. intraradices ejerce un efecto protector contra
Radopholus similis y Prathylenchus coffea en plantas colonizadas de plátano (Musa sp.) y
en raíces transformadas de zanahoria afectando negativamente la reproducción de P.
coffeae (Elsen et al., 2003; 2008).
También, se ha reportado que esta asociación simbiótica induce resistencia contra
patógenos de parte aérea. Existen genes que se expresan de manera diferencial en la parte
aérea de plantas de Medicago truncatula colonizadas con el HMA G. intraradices y no
colonizadas. El análisis de los perfiles de transcripción indican la inducción sistémica de
genes involucrados con mecanismos de respuesta a estrés o patógenos en planta, además de
la disminución significativa de unidades formadoras de colonias de plantas de M.
trúncatula colonizadas con G. intraradices y retadas con el patógeno foliar Xanthomonas
campestris; lo cual sugiere que las plantas micorrizadas pueden mejorar la resistencia a
enfermedades (Liu et al., 2007). En un estudio posterior de la interacción entre el HMA G.
intraradices y el patógeno foliar X. campestris en plantas de tomate, se obtuvieron
resultados que muestran una inducción de tolerancia por micorrización contra mancha
bacteriana, esto comparado con plantas no micorrizadas (Galindo-Flores, 2008).
La bioprotección de plantas colonizadas con HMA es el resultado de complejas
interacciones entre la planta, el patógeno y el HMA. Hay numerosos mecanismos
propuestos para conferir esta bioprotección, pero se ha sugerido que la bioprotección
efectiva es el resultado acumulativo de todos estos mecanismos trabajando separadamente,
y/o juntos (Harrier y Watson, 2004). El desafío para desarrollar más sistemas de producción

Quintero-Zamora, 2010 13
sustentables en el futuro incluye obtener un mejor entendimiento de los mecanismos
involucrados en las interacciones planta hongo y patógeno y los factores del ambiente que
en conjunto dictan la escala y tiempo de su expresión. Más aún, el entendimiento de estas
interacciones permitirá un mejor empleo como biocontrol de los HMA.
2.2.2.2. Bioprotección por hongos micorrízicos arbusculares en frijol
Mora-Romero (2008) reportó un estudio del efecto de la micorrización con G.
intraradices en la bioprotección de las variedades de frijol (P. vulgaris) A-55, Az. Regional
87 y Az. Higuera contra S. sclerotiorum. Ensayos en hojas desprendidas y en planta
completa demostraron que la respuesta de bioprotección por la colonización de frijol con
HMA depende de la variedad de frijol estudiada. Las variedades de frijol Az. Regional 87 y
A-55 mostraron mayor tolerancia al patógeno en plantas colonizadas con G. intraradices
que en plantas no colonizadas. Sin embargo, no se observó la misma respuesta en la
variedad Az. Higuera.
La tolerancia de las plantas de frijol de las tres variedades se evaluó primero
comparando el comportamiento de los tejidos de las plantas de frijol ante S. sclerotiorum
bajo condiciones de no micorrización, donde no se observaron diferencias significativas en
respuesta al patógeno. Las tres variedades de frijol probadas resultaron susceptibles a S.
sclerotiorum bajo esta condición. A la fecha, no existen reportes de variedades de frijol
resistentes a S. sclerotiorum, lo cual podría deberse al hecho de que los programas de
mejoramiento genético de variedades no consideran las condiciones de micorrización.
Al comparar la respuesta de una variedad colonizada con G. intraradices y no
colonizada se observó que las lesiones ocasionadas por S. sclerotiorum fueron menores en
las variedades Az. Regional 87 y A-55 sin embargo la variedad Az. Higuera no mostró
inducción de tolerancia. Es posible que el programa de mejoramiento para la obtención de
esta variedad, se enfocara en aspectos de rendimiento y alta calidad de grano y tal vez la
inducción de tolerancia de esta variedad por micorrización se haya perdido durante el
proceso de mejoramiento. La no inducción de tolerancia de Az. Higuera también podría
relacionarse al bajo porcentaje de colonización obtenido para esta variedad en planta
completa (23%). Puede ser posible que exista un nivel mínimo requerido de colonización
para que se presente la tolerancia y que dependa de la variedad. Sin embargo, no existen

Quintero-Zamora, 2010 14
reportes que sustenten si la bioprotección sistémica contra S. sclerotiorum depende del
grado de colonización con HMA o si es suficiente un porcentaje mínimo de colonización
para encender el mecanismo de bioprotección (Mora-Romero, 2008).
Es por esto, que experimentos posteriores resultan necesarios para entender el
mecanismo de bioprotección que se induce por micorrización. La expresión diferencial de
proteínas en las variedades Az. Regional 87 (tolerante) y Az. Higuera (susceptible); las
cuales responden de diferente manera al ataque de S. sclerotiorum cuando están
colonizadas con HMA, nos permitirá visualizar proteínas posiblemente involucradas con
el mecanismo de bioprotección por HMA. Si aprendemos en que consiste el mecanismo,
podremos entender mejor sus implicaciones en el mejoramiento genético para buscar la
resistencia a S. sclerotiorum en un cultivo tan importante en nuestro país, como lo es el
frijol.
2.3. Glomus intraradices
El HMA G. intraradices es una especie cosmopolita que se ha utilizado en muchos
estudios de micorrizas arbusculares. G. intraradices forma esporas redondeadas (entre 40 y
190 µm de diámetro) en el interior de las raíces de la planta hospedadora. La pared es de
tipo amorfo y reacciona con el reactivo de Melzer, presentando una capa externa
evanescente y una o dos capas internas laminadas de color más oscuro. El grosor de la
pared varía entre 3 y 15 µm y se extiende hacia el pedúnculo de la espora en forma de tubo.
El color de la espora puede variar desde amarillo hasta marrón claro. A pesar de que las
esporas de G. intraradices se forman en el interior de la raíz, pueden encontrarse también
grupos de esporas en el suelo, provenientes de la disgregación de una raíz (Schenck y
Smith, 1982) o formadas en el micelio externo.
2.4. Proteómica
La mayoría de las funciones biológicas que ocurren en los organismos vivos son
mediadas por proteínas. El proteoma es modificado en función de factores bióticos y
abióticos, entre algunos se pueden mencionar estados de enfermedades o de desarrollo,
daños por insectos, tipos de células y tejidos, estrés ambiental y condiciones de suelo,
etcétera (Chinnasamy, 2005). De aquí la necesidad de un área enfocada en el estudio

Quintero-Zamora, 2010 15
funcional de las proteínas como lo es la proteómica, término sugerido por Marc Wilkins, en
1994. La proteómica es definida como el análisis sistemático y la documentación de todas
las especies de proteínas y sus modificaciones post-traduccionales en un organismo, tejido,
célula u organelo a un tiempo o condición específica de desarrollo o crecimiento dado
(Rose et al., 2004; Chinnasamy, 2005; Chen y Harmon, 2006).
El desarrollo de las ciencias denominadas “omicas” como es el caso de la
proteómica ha permitido su aplicación al entendimiento de distintas problemáticas actuales,
incluyendo las concernientes a la agricultura, entre las que se encuentra la interacción
planta-microorganismo.
El objetivo de la proteómica comparativa no es el de identificar el conjunto entero
de proteínas en una muestra en particular, sino de caracterizar diferencias entre poblaciones
de proteínas diferentes (Figura 1) (Rose et al., 2004). Tradicionalmente, la proteómica se
basa en la combinación de electroforesis bidimensional (2D-SDS-PAGE) permitiendo la
separación de proteínas desnaturalizadas de acuerdo a su punto isoeléctrico (primera
dimensión) y peso molecular (segunda dimensión), y de métodos de identificación basados
en espectrometría de masas (Bestel-Corre et al., 2004).
2.4.1. Análisis de proteínas.
El análisis proteómico de los tejidos vegetales, son típicamente mas problemáticos
que con otros organismos ya que implica un número de desafíos prácticos. Además de
tener concentraciones bajas de proteínas, los tejidos vegetales son a menudo ricos en
proteasas y en materiales que interfieren seriamente con la separación y análisis
multifactorial de la proteína, incluyendo pared celular y almacenamiento de polisacáridos,
lípidos, compuestos fenólicos y una amplia variedad de metabolitos secundarios (Granier,
1988; Tsugita y Kamo, 1999). Tales contaminantes son un problema particular para la
electroforesis bidimensional (2D-SDS-PAGE), dando por resultado en los geles después de
la electroforesis rayas horizontales (que las proteínas no se logren separar en base a su
punto isoelétrico) y verticales (que las proteínas no se logren separar en base a su peso
molecular), manchas y una reducción en el número de puntos resueltos de la proteína.

Quintero-Zamora, 2010 16
Extracción de proteínas
Separación en2D SDS-PAGE
Generación de péptidos
Hidrólisis con
Tripsina
Espectrometría de masas
Huella peptídica(Peptide mass fingerprinting)
Etiqueta peptídica(Secuencia de aminoácidos)
Relación masa/carga (m/z)
Inte
nsid
ad re
lativ
a %
Figura 1. Esquema de los diferentes pasos involucrados en un análisis de
proteómica comparativa.

Quintero-Zamora, 2010 17
La electroforesis bidimensional en geles de poliacrilamida (2D-SDS-PAGE) es una
herramienta muy utilizada en proteómica para la separación y cuantificación de proteínas.
La identificación de proteínas de manera rápida y masiva es factible debido a los avances
en espectrometría de masas. MALDI–TOF–MS (por sus siglas en inglés, Matrix-Assisted
Laser Desorption/Ionization Time of Flight Mass Spectrometry) y LC–MS/MS (por sus
siglas en inglés, Liquid Chromatography Mass Spectrometry and Electrospray) son
métodos sensibles y precisos para caracterizar e identificar perfiles de proteínas. Junto con
la técnica de 2D–SDS-PAGE, estas herramientas se pueden utilizar para visualizar y
comparar mezclas complejas de proteínas, además de obtener información acerca de las
proteínas individuales implicadas en respuestas biológicas específicas (Adessi et al., 1997;
Finnie et al., 2002).
Recientemente, las metodologías en electroforesis 2D-SDS-PAGE han tenido
avances; mediante el desarrollo de protocolos para el procesamiento de muestras y del uso
de tiras de gel de gradiente de pH inmovilizado para llevar a cabo el isoelectroenfoque
(IEF, separación de proteínas por su punto isoeléctrico) de las proteínas. Estos avances han
permitido obtener proteínas separadas en cantidades micropreparativas para su
caracterización e identificación mediante espectrometría de masas y el análisis cuantitativo
de proteínas expresadas diferencialmente. Sin embargo, el aislamiento de proteínas sigue
siendo un reto respecto a la resolución en el análisis exacto de proteínas. Lo anterior debido
a la presencia de contaminantes que afectan el rendimiento en la 2D-SDS-PAGE. La
extracción de proteínas de tejidos vegetales puede ser particularmente un desafío debido a
la alta relación carbohidratos/proteínas en la mayoría de los tejidos vegetales (Thelen,
2007). Aunque varios métodos de análisis por 2D-SDS-PAGE de plantas y semillas han
sido reportados para una variedad de cultivos, no existen reportes en tejidos de plantas de
frijol. La mayoría de los estudios de proteómica en tejidos vegetales, utilizan una
estrategia básica de la precipitación de la proteína con ácido tricloroacético (TCA) y
acetona seguidos por la resolubilización en un amortiguador específico para el
isoelectroenfoque (IEF) que contiene agentes caotrópicos (urea, tiourea) y detergentes [3-
(cloroamido propil)-dimetilamonio-1-propanosulfato (CHAPS), Tritón X-100] (Santoni y
Bellini, 1994). Bajo la anterior estrategia se aumenta la concentración de la proteína y se
ayuda a remover los contaminantes, aunque algunos contaminantes poliméricos son

Quintero-Zamora, 2010 18
coextraídos a menudo. Esto es un problema particular con los tejidos que son ricos en
compuestos tales como polisacáridos y polifenoles solubles de la pared celular
(Saravanan y Rose, 2004).
Se ha desarrollado un protocolo alternativo con el cual las proteínas se solubilizan
en solución saturada de fenol, con o sin el detergente aniónico dodecil sulfato de sodio
(SDS); y se precipitan posteriormente con acetato de amonio y metanol, seguida por la
resolubilización en el amortiguador para IEF (Hurkman y Tanaka, 1986; Meyer et al.,
1988). Este método ha mostrado su eficacia para la obtención de proteínas con la
contaminación mínima, se ha utilizado para extraer las proteínas de las hojas verdes de
plantas de olivo, que contienen grandes cantidades de polifenoles, dando por resultado un
aumento substancial en la calidad de las proteínas resueltas mediante 2D-SDS-PAGE
(Wang et al., 2003). Estos dos protocolos se han utilizado en estudios proteómicos de un
número de diversas especies y tejidos de planta. Sin embargo, la compatibilidad de éstos
con la 2D-SDS-PAGE depende de la muestra analizada.
Debido a que la mayoría de los estudios de proteómica publicados de tejidos
vegetales, utilizan un procedimiento en el cual las proteínas se precipitan con TCA y
acetona, o bien un método basado en la solubilización de proteínas con soluciones saturadas
de fenol, Saravanan y Rose (2004) reportaron los resultados de poner en contraste estas
estrategias de una manera sistemática con métodos alternativos contra distintos tejidos
vegetales. Para tratar esto, el protocolo basado en método de fenol propuesto por Hurkman
y Tanaka (1986) fue combinado con un protocolo basado en precipitación con TCA
propuesto por Santoni y Bellini (1994) para proteínas de tejidos de tomate. Sus resultados
muestran que el método de extracción con fenol dio un mayor rendimiento y una mayor
intensidad y resolución de proteínas (spots en el gel) en 2D-SDS-PAGE, particularmente
con los extractos de los tejidos que contenían niveles de polisacáridos solubles. Los
diferentes métodos también generaron diferencias notables en los patrones de spots de
proteínas en geles de electroforesis bidimensionales. La estrategia de espectrometría de
huella de masa péptidica (peptide mass fingerprinting) fue utilizada para identificar los
polipéptidos que eran comunes a los múltiples métodos de extracción evaluados o que
estaban presentes únicamente en un método. Los autores sugieren que el protocolo de
solubilización con fenol es altamente eficaz con tejidos más recalcitrantes y que una

Quintero-Zamora, 2010 19
combinación de métodos de TCA y de fenol es mejor en 2D-SDS-PAGE en base al análisis
proteómico de la mayoría de los tejidos vegetales evaluados (raíces, tallos, hojas y flores de
tomate; tejido de aguacate, banana y naranja).
Sarma et al. (2008) publicaron un método de extracción con fenol modificado, este
método no solo mejoró la resolución y reproducibilidad de 2D-SDS-PAGE sino también
acortó el tiempo de análisis. La aplicabilidad del método fue demostrada usando varios
tejidos de soya. La precipitación se llevó a cabo por fenol alcalino y acetato de
amonio/metanol durante 2 a 3 horas a -80°C, se usó etanol para el lavado final de la
proteína precipitada. El reactivo ditiotreitol (DTT) fue usado en todas las soluciones desde
el principio, mejorando considerablemente la solubilidad de las proteínas precipitadas. La
solubilización fue mejorada más a fondo mediante el uso de una mezcla de detergentes y
desnaturalizantes en altas concentraciones junto con grandes cantidades de DTT en el
amortiguador IEF.
En el caso de los geles bidimensionales (2D-SDS-PAGE), la identificación de las
proteínas o manchas conduce a la creación de mapas de referencia, que definen las
proteínas expresadas por un organismo o tejidos en condiciones determinadas. Las
proteínas pueden ser identificadas por diversos procedimientos, entre los que se incluyen
la secuenciación del extremo N-terminal, detección con anticuerpos específicos,
composición de aminoácidos, o co-migración con proteínas conocidas. Debido a su
rapidez y elevada sensibilidad, la espectrometría de masas se ha convertido en el método
de detección para la identificación de proteínas a gran escala y el primer paso para el
estudio del proteoma de distintos organismos. También permite la caracterización de
modificaciones post-traduccionales que presentan relevancia fisiológica, tales como
glicosilación y fosforilación. El análisis de las proteínas mediante espectrometría de
masas ha sido posible gracias al desarrollo de varios métodos de ionización suave para
convertir biomoléculas grandes, polares y no volátiles en iones en fase gaseosa. Los
espectrómetros de masa están formados al menos por una fuente de iones, un analizador
de masas y un detector que mide la relación masa/carga (m/z) de los iones en fase
gaseosa.
Los espectrómetros de masas tienen en común que miden la relación m/z de iones
gaseosos haciendo uso de campos eléctricos y/o magnéticos. La medida exacta de este valor

Quintero-Zamora, 2010 20
para las moléculas gaseosas ionizadas del analito (ion molecular) dará información sobre su
masa molar (peso molecular). Si además se induce la fragmentación del ión molecular, cada
fragmento iónico será detectado con su propio valor de m/z (McLafferty et al., 1993). El
conjunto de fragmentos formados es característico de cada analito y depende de su
estructura química (“huella dactilar”). Por lo tanto, del análisis de los fragmentos obtenidos
se obtiene información sobre la estructura química del analito en cuestión, en nuestro caso
de interés, la identificación de proteínas.
2.5. Resistencia sistémica
Las plantas son expuestas a muchas formas de estrés, como el ataque de herbívoros
y patógenos, o condiciones adversas a luz, agua, temperatura, nutrientes o sales. La
percepción de señales de estrés seguido resulta en la biosíntesis de una o más moléculas de
señalización secundaria como jasmonatos, etileno y acido salicílico. La producción de estas
hormonas genera una transducción de señales que lleva a una cascada de eventos para la
adaptación fisiológica de la planta a condiciones de estrés (citado en Memelink, 2009).
La resistencia sistémica es un “estado fisiológico de aumento de la capacidad
defensiva“, inducido por estímulos ambientales específicos, donde las defensas
constitutivas de la planta son potenciadas contra subsecuentes desafíos bióticos. Este
aumento del estado de resistencia es efectivo contra un amplio rango de patógenos y
parásitos, incluyendo hongos, bacterias, virus, nemátodos, y hasta insectos herbívoros
(Vallad y Goodman, 2004; Chinnasamy, 2005; Walters y Heil, 2007).
Las dos formas más claras y definidas de resistencia sistémica son resistencia
sistémica adquirida (RSA) y resistencia sistémica inducida (RSI), las cuales pueden ser
diferenciadas con base en la naturaleza del inductor de la resistencia y la vía de regulación
involucrada.
2.5.1. Resistencia sistémica adquirida (RSA)
Es producida en plantas después de la expresión de la respuesta hipersensible, es
decir, después de que el tejido vegetal al ser atacado de manera local por un patógeno
desata una serie de mecanismos locales, incluyendo la producción de especies reactivas de
oxígeno, que causan inclusive la muerte celular programada, para contener el ataque de un

Quintero-Zamora, 2010 21
patógeno. Como respuesta a esta infección local, la planta responde con una cascada de
señales dependientes de ácido salicílico, el cual se acumula inicialmente en el sitio mismo
de la infección. Posteriormente, un compuesto de origen desconocido se mueve en el
floema y transporta la señal a lugares distales de la planta. Entonces, el ácido salicílico se
acumula en la región distal y es convertido a metil salicilato el cual puede servir como una
señal a plantas vecinas. Proteínas con actividades antimicrobianas tales como β-1,3-
glucanasa, quitinasas, proteínas ricas en cisteína y proteínas PR (proteínas relacionadas a
patogénesis) aparecen en la porción distal de la planta y seguramente participan en la
resistencia sistémica ante ataques de patógenos posteriores (Vallad y Goodman, 2004).
La RSA se asocia con un aumento en los niveles de acido salicílico (AS), y se
caracteriza por la activación coordinada de un grupo de proteínas específico relacionados a
patogénesis (PR), muchas de las proteínas del grupo codifican para proteínas PR con
actividad antimicrobiana (Van Loon et al., 2006). Estudios con plantas mutantes y
transgénicas que son perjudicadas en la percepción del AS demostraron el papel central de
esta fitohormona en la RSA (Loake y Grant, 2007; Vlot et al., 2008). La proteína NPR1
(non-expressor of PR genes-1) es un importante co-activador transcripcional de expresión
de genes PR, una vez que ésta es activada por la acción de ácido salicílico (Dong, 2004).
Además de AS, la hormona gaseosa etileno se encuentra implicada en RSA.
Verberne et al. (2003) demostraron que la percepción de etileno se requiere para el inicio
de RSA, el cual es activado una vez infectado tabaco con el virus del mosaico de tabaco.
Además Truman et al. (2007) demostraron que el ácido jasmónico (AJ) juega un papel
importante en RSA en mutantes en la señalización de ácido jasmónico, al retar estas plantas
mutantes con una cepa virulenta de Pseudomonas syringae. Sin embargo, otras mutantes en
señalización de AJ mostraron una activación normal en los niveles de RSA (Attaran et al.,
2009; Cui et al., 2005; Pieterse et al., 1998; Ton et al., 2002). Por lo que el papel exacto
del AJ en RSA necesita aún ser definido en estudios posteriores (Citado en Van der Ent et
al., 2009).
2.5.2. Resistencia sistémica inducida (RSI)
La RSI se ha definido como un estado fisiológico identificado por el mejoramiento
de la capacidad de defensa a patógenos en respuesta a la interacción de un rango de

Quintero-Zamora, 2010 22
organismos no patogénicos y agentes de biocontrol. Las defensas innatas de la planta se
potencian contra el subsecuente ataque de patógenos y parásitos, incluyendo bacterias,
virus, hongos, nemátodos y hasta insectos herbívoros. La RSI es fenotípicamente similar a
la RSA, la cual es activada por un patógeno virulento, no virulento, algún microorganismo
no patogénico o de manera artificial por algún químico (Vallad y Goodman, 2004). A
diferencia de la RSA, la RSI no involucra la acumulación de proteínas relacionadas con
patógenos o AS (Pietersen et al., 1996) y en cambio, es regulada por AJ y etileno dos
grupos de fitoreguladores de naturaleza volátil.
La RSI es disparada por microorganismos benéficos y es efectiva contra diferentes
tipos de ataques, confiriendo una resistencia de amplio espectro. La fitohormona AJ y sus
derivados llamados jasmonatos son los reguladores de esta respuesta sistémica (Van der Ent
et al., 2009).
La RSI se produce por la interacción de la planta con algunas bacterias que
colonizan las raíces pero que no causan enfermedad. A estas bacterias se les denomina
bacterias promotoras de crecimiento o PGPR (por sus siglas en inglés, plant growth
promoting rhizobacteria). A diferencia de la RSA, la RSI no involucra la acumulación de
proteínas relacionadas a patogénesis (PR) o de AS. La interacción con PGPR induce la
síntesis de manera transitoria de AJ y etileno, los cuales, de alguna manera transportan una
señal a las zonas distales de la planta para encender un estado de protección (Vallad y
Goodman, 2004).
La activación de RSI en plantas micorrizadas pareciera involucrar una señal móvil
inducida por la micorriza. Sin embargo, poco se conoce sobre la señal concerniente a la
inducción de RSI posterior al establecimiento de la simbiosis (Hause y Fester, 2005). Sin
embargo, el AJ, el cual se ha encontrado que juega un papel importante para el
establecimiento de la simbiosis, podría ser un candidato apropiado, considerando su
participación en el movimiento a larga distancia en otros fenómenos, como en la respuesta
a daño mecánico (Noval et al., 2007).
La proteína NPR1 reguladora de defensa en RSA, también ha sido implicada en la
RSI (Dong, 2004; Pieterse y Van Loon, 2004). Mutantes en plantas del género Arabidopsis
mostraron bloqueada su habilidad para expresar RSI una vez colonizadas las raíces con
PGPR (Hossain et al., 2008; Stein et al., 2008; Segarra et al., 2009). Sin embargo, el papel

Quintero-Zamora, 2010 23
de la proteína NPR1 en RSI podría ser diferente que en RSA. Sin embargo, los mecanismos
moleculares por los que NPR1 ejerce su función no han sido delucidados (Van der Ent et
al., 2009).
2.5.3. Resistencia inducida por micorrización (RIM)
Se conoce muy poco de los mecanismos moleculares responsables de la RIM. Sin
embargo, se ha propuesto que comparte mecanismos con la resistencia sistémica inducida
por rizobacterias promotoras de crecimiento (Verhagen et al., 2004), la hipótesis
establecida dice que la simbiosis micorrízica dispara un estado de pre-acondicionamiento o
“priming” que actúa de manera sistémica a lo largo de toda la planta, no únicamente en los
tejidos en donde se lleva a cabo la interacción. Este pre-acondicionamiento enciende un
mecanismo de alerta que le permite a la planta tolerar de mejor manera un ataque por
patógenos foliares (Pozo y Azcón Aguilar, 2007).
Cuando una planta pre-acondicionada es atacada por herbívoros, patógenos, o bien,
sometida a estrés abiótico, ésta responde activando genes de defensa celular de una manera
más rápida e intensa que las plantas no preacondicionadas (Beckers y Conrath, 2007;
Goellner y Conrath, 2008).
A partir de estudios con rizobacterias promotoras de crecimiento, se ha hipotetizado
que el pre-acondicionamiento podría estar asociado con la acumulación de proteínas de
señalización en estado inactivo. Una vez que se lleva a cabo una exposición subsecuente al
ataque por un patógeno, un segundo evento de señalización podría activar las proteínas en
cuestión, amplificando de esta manera la señal y llevando a una activación del sistema de
defensa de una manera mucho más rápida e intensa (Beckers y Conrath, 2007). La
posibilidad de responder de manera más expedita a un ataque por patógenos sin la
necesidad de encender los mecanismos de defensa antes de enfrentarse al patógeno,
representaría una ventaja para la planta. Es posible que los mecanismos de pre-
acondicionamiento inducidos por rizobacterias y los inducidos por la micorrización
arbuscular compartan algunos de sus elementos.

Quintero-Zamora, 2010 24
2.6. Métodos de genómica y proteómica empleados en el estudio de la interacción
planta-microorganismo
En estudios recientes del proteoma de la raíz de M. truncatula después de la
infección con el patógeno Oomycete Aphanomyces euteiches, se identificaron proteínas que
se regulan después de la infección con dicho patógeno (Colditz et al., 2004). Dieciséis
proteínas fueron expresadas diferencialmente con respecto a la condición infectada con el
patógeno. La mayoría de las proteínas inducidas pertenecen a la familia de la clase 10 de
proteínas relacionadas a patogénesis (PR10). Tres de estas proteínas también fueron
inducidas por aplicación exógena de ácido abscisíco (ABA). Otras tres de estas
proteínas inducidas por el patógeno disminuyen su expresión en estrés por sequía. Esto
sugiere que las proteínas tipo PR10 desempeñan un papel fundamental durante las
adaptaciones de la raíz a diversas condiciones de estrés.
Un segundo estudio acerca del proteoma de la raíz de M. truncatula se llevó a cabo.
Como primer estrategia se analizaron mediante proteómica comparativa dos líneas de M.
truncatula las cuales muestran un aumento (línea F83.005-9) o disminución (línea F83.005-
5) a la susceptibilidad al patógeno A. euteiches comparadas con la línea A-17 (línea
susceptible) analizada anteriormente por Colditz et al. (2004). Varias proteínas fueron
identificadas como diferencialmente inducidas después de retadas con el patógeno
mostrando un aumento en su acumulación en la línea más resistente. Entre estas
proteínas, se identificaron dos subunidades del α-proteosoma, las cuales pudieran estar
implicadas en el mecanismo que lleva a un aumento de resistencia a la enfermedad.
Como segunda estrategia, la mejora de resistencia de la planta hacia A. euteiches fue
estudiado en la línea A-17 (línea susceptible) la cual mostró un efecto bioprotector de
simbiosis micorrízica preestablecida. Los análisis del proteoma revelaron la inducción
de patrones de proteínas similares a las encontradas en las accesiones de M. truncatula
con un nivel de resistencia alto a A. euteiches. Además, se analizaron plantas de M.
truncatula susceptibles al patógeno después de la aplicación exógena (ABA) antes de la
infección de la raíz. Los resultados mostraron niveles de expresión elevados del grupo
de proteínas relacionadas a patógenesis (PR10) las cuales fueron previamente
identificadas como reguladas después de la infección con A. euteiches. Estos resultados

Quintero-Zamora, 2010 25
sugieren un aumento de susceptibilidad a la infección causado por la aplicación de ABA
(Colditz et al., 2005).
Castillejo et al. (2004) en el estudio del proteoma de raíces de una línea tolerante y
una susceptible de plantas de chícharo (Pisum sativum) en la interacción con el parásito
jopo de las habas (Orobanche crenata) demostraron cambios en el perfil proteómico. En
respuesta a la inoculación, en la línea susceptible se obtuvieron 34 proteínas suprimidas en
su expresión, una con un aumento en su expresión y tres proteínas diferenciales de manera
cualitativa fueron inducidas. De este grupo de proteínas los autores lograron identificar
siete y correspondieron al metabolismo de los carbohidratos y a la ruta de transporte
electrónico mitocondrial. Estos incluyen una fructokinasa, una fructosa bifosfato aldolasa y
una ferredoxin-NADP reductasa, una enoyl-CoA hidratasa putativa y un precursor
mitocondrial de la oxidasa-2 alternativa. Para la línea resistente pocos cambios fueron
observados en respuesta a O. crenata. Quince proteínas aumentaron su expresión y tres
disminuyeron, una proteína cualitativa fue encontrada en el control sin infectar. Ocho
proteínas fueron identificadas. Estas incluyen dos glutamino sintetasas de raíz, la isovaleril-
CoA deshidrogenasa, tres precursores de peroxidasas 43 y dos precursores de glucan endo-
1,3-β-glucosidasa.
En el curso temporal de la interacción simbiótica en raíces de plantas de frijol
colonizadas con G. mosseae, cubriendo todos los estados de desarrollo de la simbiosis, se
detectaron cambios moderados en la expresión de genes relacionados con respuestas de
defensa en plantas, tales como quitinasa, β-1,3-glucanasa y fenilalanina amonio-liasa (PAL)
en comparación con plantas no colonizadas como control (Morh et al., 1998).
Algunos genes de respuesta y de la biosíntesis del AJ son expresados en células que
contienen arbúsculos. Además, las raíces colonizadas con HMA presentan un incremento
en el nivel de AJ endógeno (Hause et al., 2002; Hause et al., 2007), por lo que es posible
especular que el AJ pudiera funcionar como una señal endógena en la resistencia inducida
por micorrización. Otras hormonas como el ABA muestran la inducción o represión de
algunos genes en micorrizas arbusculares y pueden también afectar la resistencia de la
planta a elicitores (Hause et al., 2007).
Estudios a nivel de análisis transcripcional se han enfocado en describir genes que
se expresan de manera diferencial en plantas colonizadas con HMA frente a patógenos de

Quintero-Zamora, 2010 26
raíz con el objeto de generar información que permita elucidar los mecanismos de la
resistencia observada por micorrización (Salzer et al., 2000; Liu et al., 2003; Whipps, 2004;
Liu et al., 2007). Aunque en menor abundancia, se han realizado también algunos trabajos
en raíz utilizando proteómica comparativa para identificar proteínas relacionadas con el
mecanismo de resistencia (Colditz et al., 2004; 2005).
Aunque la expresión global de genes a nivel transcripcional (transcriptómica)
proporciona información importante respecto a los estados tempranos de transcripción del
genoma, los niveles de mRNA no siempre son consistentes con la abundancia de proteínas
(citado en Rose et al., 2004). En comparación con el nivel de mRNA, el nivel de proteína
integra procesamientos post-transcripcionales y post-traduccionales que modulan la
cantidad, localización y eficiencia del producto final en la célula. La conjunción de estudios
de proteómica con transcriptómica permite un mejor conocimiento del papel que estos
genes juegan en la simbiosis y en el mecanismo de inducción sistémica (Rose et al., 2004;
Chen y Harmon, 2006).
Fontana et al. (2008) llevaron a cabo un estudio mediante la técnica de hibridación
sustractiva, la cual aisla secuencias tags expresadas, representando genes que son
expresados diferencialmente en diferentes poblaciones de RNAm. El estudio se llevó a
cabo en plantas de Platanus acerifolia tratadas con cerato-platanina. La cerato-platanina es
una proteína fitotóxica secretada por el ascomiceto Ceratocystis platani, el cual es un
patógeno virulento en plantas de plátano. Ellos demostraron que cuando las hojas de
plátano son tratadas con la proteína fitotóxica, Ceratocystis ya no puede crecer en ella y
hay sobreexpresión de la proteína transcetolasa. Además, sus resultados muestran un
incremento en el metabolismo del carbono. Algunas proteínas involucradas en la
fotosíntesis tales como la ferredoxina A, la ribulosa 1-5,bifosfato carboxilasa y algunas
enzimas cloroplásticas del fotosistema I y II se encontraron expresadas diferencialmente en
este estudio.
Shoresh y Harman (2008) analizaron el perfil proteómico de hojas de plántulas de
maíz colonizadas con el hongo Trichoderma harzianum el cual es empleado como agente
de biocontrol contra varios patógenos de suelo. T. harzianum induce cambios en el
proteoma de hojas de plántulas de maíz (Zea mays). Se identificaron 91 proteínas, de 114
inducidas y 30 proteínas, de 50 cuya acumulación disminuyó en las hojas. Una gran

Quintero-Zamora, 2010 27
cantidad de proteínas inducidas en esta asociación estan involucradas en el metabolismo de
aminoácidos, metabolismo de carbono, fotosíntesis (fructoquinasa, fructosa bifosfato
aldolasa, gliceraldehido-3-fosfato deshidrogenasa, malato deshidrogenasa, oxalato oxidasa,
β-glucosidasa), entre otras.
El perfil de expresión de genes en hojas y raíces de plantas de tomate (Solanum
lycopersicum) colonizadas con el HMA G. mosseae ha sido reportado previamente (Fiorilli
et al., 2009). Mediante el empleo de microarreglos, los autores reportan 362 genes
sobreexpresados y 293 genes cuya expresión fue reprimida en raíces, también se detectó
una modulación importante de genes de parte aérea; 85 genes sobreexpresados y 337 genes
reprimidos. La mayoría de los genes de respuesta en ambos tejidos estuvieron relacionados
con metabolismo primario y secundario, defensa, respuesta a estímulo, organización
celular, modificación de proteínas, y regulación transcripcional.
El análisis del perfil de expresión de células corticales conteniendo arbúsculos
empleando una metodología de microdisección laser permitió la identificación de seis
genes expresados diferencialmente. Los resultados sugieren que el metabolismo de auxinas
y ABA está involucrado en la formación y/o funcionamiento de arbúsculos (Fiorilli et al.
2009).
El análisis por electroforesis bidimensional ha demostrado ser muy útil para revelar
modificaciones de proteínas en raíces relacionados con la micorriza arbuscular (Samra et
al., 1997; Bestel-Corre et al., 2002; Chinnasamy, 2005; Colditz et al., 2005; Valot et al.,
2005), o bien del hongo en sistemas monoxénicos de raíces transformadas de zanahoria
colonizados con G. intraradices (Simoneau et al., 1994; Dumas-Gaudot et al., 2004).
Samra et al. (1997) detectaron por 2D-SDS-PAGE, proteínas relacionadas con la
simbiosis durante estadíos tempranos del establecimiento de la micorriza arbuscular entre
G. mosseae y raíces de Pisum sativum, sin embargo en este estudio no se identificó su
identidad por análisis de espectrometría de masas. Benabdellah et al. (2000) evaluaron
usando esta misma estrategia cambios inducidos por la formación de micorrizas
arbusculares (G. mosseae) en el patrón de polipéptidos de la membrana plasmática en raíz
de tomate. Ellos encontraron una reducción en la regulación de algunos polipéptidos
constitutivos y la síntesis de nuevos polipéptidos o endomicorrizinas. Sin embargo, solo se
identificó una proteína con regulación negativa, la subunidad tipo V de la H+-ATPasa

Quintero-Zamora, 2010 28
vacuolar, la cual podría estar relacionada con una disminución en el nivel de síntesis de la
membrana vacuolar y a su vez con el papel de la vacuola en el mantenimiento de la
homeostasis iónica en el citosol. El mismo trabajo muestra en una comparación de los
cambios inducidos por dos diferentes niveles de colonización por HMA encontrando que 16
polipéptidos fueron desplegados diferencialmente en ambos estados de colonización, sin
embargo éstos no fueron identificados.
Bestel-Corre et al. (2002) estudiaron perfiles de proteínas de raíz realizando un
análisis de curso temporal en plantas de M. truncatula inoculadas con el HMA G. mosseae.
En respuesta a la colonización por HMA se encontraron dos polipéptidos que
disminuyeron, 12 polipéptidos que aumentaron su acumulación y 41 fueron polipéptidos
inducidos solamente en la condición de plantas colonizadas con respecto al control. Dentro
de estos polipéptidos se encontraron proteínas involucradas en respuesta a defensa contra
patógenos, como peroxidasa y proteína inhibidora de poligalacturonasa; proteínas
involucradas en la fisiología de la raíz, como una proteína tipo miosina cadena pesada (la
cual podría participar en la reorganización del citoesqueleto en células colonizadas) y
proteínas implicadas en vía respiratoria mitocondrial, como una serina hidroximetil-
transferasa y citocromo-c-oxidasa.
Valot et al. (2005) identificaron mediante proteómica comparativa subcelular 25
proteínas de membrana cuya expresión fue significativamente alterada por la simbiosis MA
en raíces de M. truncatula inoculadas con el hongo G. intraradices. Valot et al. (2006)
reportaron dos estrategias complementarias por espectrometría de masas, basada en un
sistema automatizado de cromatografía líquida masas/masas (LC-MS/MS) con un fuerte
intercambio catiónico y; SDS-PAGE y cromatografía de fase reversa combinado con un
análisis sistemático LC-MS/MS. Esto permitió la identificación de proteínas relacionadas
con MA en fracciones de membrana plasmática de raíz de M. truncatula inoculada con el
hongo G. intraradices.
La estrategia del empleo de sistemas monoxénicos ha permitido la identificación de
nuevos polipéptidos en raíces transformadas de tomate durante el desarrollo de micorrizas
arbusculares (Simoneau et al., 1994) y la identificación de proteínas extraradicales de
hongo se ha llevado a cabo en un sistema de raíces transformadas de zanahoria colonizadas
con G. intraradices (Dumas-Gaudot et al., 2004). Este trabajo permitió la identificación de

Quintero-Zamora, 2010 29
proteínas secretadas por HMA, entre las cuales se incluyen proteínas del metabolismo
celular central como: una óxido reductasa y un precursor de la ATP sintasa-β mitocondrial;
además de una proteína tipo NmrA (por sus siglas en inglés: nitrogen metabolite repression
in Aspergillus nidulans), el cual es un represor de la transcripción involucrado en la
regulación del metabolismo de nitrógeno en Aspergillus nidulans y una proteína de choque
térmico, cuya función consiste en proteger a la célula a estrés térmico y también podría
interactuar con otras proteínas en el plegado y translocación de polipétidos en los diferentes
compartimentos celulares.
Pozo et al. (2002) estudiaron la capacidad de dos HMA (G. mosseae y G.
intraradices) para inducir la resistencia sistémica y localizada al hongo fitopatógeno
Phytophthora parasitica en raíces de tomate utilizando un sistema experimental de raíces
divididas, realizando análisis bioquímicos de algunas enzimas relacionadas a defensa a
partir de extractos de proteínas de las raíces. Las plantas de tomate respondieron de manera
diferencial a la inoculación con los dos hongos al ser infectados con el patógeno; los
autores describieron nuevas isoformas de proteínas relacionados a defensa como quitinasa,
quitosanasa, β-1,3-glucanasa y superóxido dismutasa inducidas localmente en la parte
colonizada con el hongo micorrízico y observaron ciertos efectos sistémicos como los
resultados obtenidos en la actividad lítica contra la pared celular del patógeno, concluyendo
en la existencia de una RSI mediada por la micorrización.
A pesar de que ya existe evidencia experimental de la expresión diferencial de genes
en hojas de plantas colonizadas con HMA con respecto a las no colonizadas (Taylor y
Harrier, 2003; Liu et al., 2007), y a la fecha se han publicado trabajos de expresión
diferencial de proteínas en hojas infectadas y no infectadas con patógenos foliares o de
parte aérea (Bestel-Corre et al., 2004; Colditz et al., 2004; Whipps, 2004; Chinnasamy,
2005), aún no se ha estudiado la expresión diferencial de proteínas de hojas de plantas de
frijol colonizadas con HMA y no colonizadas.
Entre los trabajos relacionados con la expresión diferencial de genes en hojas de
plantas colonizadas con HMA con respecto a las no colonizadas se encuentra el trabajo de
Taylor y Harrier (2003). Estos autores emplearon la estrategia de despliegue diferencial de
ARNm en tejido de hojas de plantas de tomate colonizadas y no colonizadas con el HMA
G. mosseae, encontrando cinco ADNc regulados diferencialmente. En años recientes, se ha

Quintero-Zamora, 2010 30
documentado que la micorrización reduce los síntomas causados por fitoplasmas (Lingua et
al., 2002) y por Alternaria solani (Fritz et al., 2006; de la Noval et al., 2007) en tomate. Liu
et al. (2007) estudiaron el impacto de la asociación micorrízica arbuscular sobre la
interacción de la planta con bacterias patógenas foliares. Empleando plantas de M.
truncatula infectadas con la bacteria foliar X. campestris pv. alfalfae se confirmó que las
plantas colonizadas con HMA presentaron un aumento en la tolerancia al patógeno aéreo.
Esta respuesta inducida por la micorrización correlacionó a su vez, con la alteración
diferencial de la expresión de algunos genes. En este trabajo, empleando microarreglos de
ADN se identificaron 468 genes cuya expresión aumentó y 131 genes cuya expresión
disminuyó en hojas de plantas colonizadas con HMA con respecto a hojas de plantas no
colonizadas. Aproximadamente el 25% de los genes inducidos en hojas de plantas
colonizadas con HMA pudieron asociarse in silico con proteínas involucradas en
mecanismos de defensa descritos en otros sistemas, sugiriendo que la micorrización podría
estar induciendo un mecanismo de defensa en la parte aérea de las plantas.
Actualmente en nuestro grupo se han llevado a cabo estudios a nivel transcripcional
en hojas de plantas de tomate colonizadas con HMA como resultado de la infección por el
agente patógeno X. campestris, donde se ha encontrado un efecto bioprotector en plantas de
tomate colonizadas con G. intraradices independientemente de su estado nutricional
fosforado (Galindo-Flores, 2008). También, se han conducido estudios en plantas crecidas
bajo condiciones controladas (16 hr luz/25 °C y 8 hr oscuridad/20 °C), donde se han
identificado mediante análisis bidimensional de proteínas y espectrometría de masas,
proteínas diferencialmente expresadas en hojas de plantas de tomate colonizadas con G.
intraradices comparadas con plantas no colonizadas. En ese trabajo, se detectaron 27
proteínas expresadas diferencialmente de la comparación de las plantas colonizadas y no
colonizadas, de las cuales 22 fueron identificadas por espectrometría de masas. La mayoría
de las proteínas identificadas disminuyeron su expresión en la condición colonizada y
fueron proteínas relacionadas a la fotosíntesis y al estado celular de óxido-reducción,
mientras que dos proteínas aumentaron su expresión y estuvieron relacionadas una con la
síntesis de auxina y la otra es una germina (Peinado-Guevara, 2010 en revisión). Las
auxinas también se han reportado en raíces de plantas colonizadas con HMA (Hause et al.,
2007; Ludwig-Muller y Guther, 2007), sin embargo el papel funcional de estas proteínas se

Quintero-Zamora, 2010 31
desconoce. Las proteínas tipo germina participan en muchos procesos que son importantes
para el desarrollo y defensa de la planta. Éstas han sido implicadas en el fortalecimiento de
la pared celular y resistencia al ataque de patógenos y estrés abiótico (Bernier y Berna,
2001; Godfrey et al., 2007; Gucciardo et al., 2007; Park et al., 2004). En otro estudio, se
llevó a cabo la comparación de las plantas colonizadas con G. intraradices y no
colonizadas y además ambas infectadas con el patógeno foliar X. campestris pv vesicatoria.
Se detectaron 27 proteínas expresadas diferencialmente, 23 de ellas aumentaron en la
condición colonizada con HMA e infectada con el patógeno con respecto al control. De las
proteínas inducidas trece se lograron identificar por espetrometría de masas. Siete proteínas
son proteínas relacionadas a fotosíntesis, las cuales aumentan su expresión en la condición
colonizada con HMA e infectada con patógeno. También se lograron identificar dos
chaperonas y las restantes estuvieron reacionadas a otras funciones moleculares (Peinado-
Guevara, 2010 en preparación). Finalmente, en experimentos realizados por Mora-Romero
(2008) se ha observado un fenómeno de bioprotección sistémico contra el patógeno S.
sclerotiorum en variedades de frijol al momento de estar colonizadas con el hongo G.
intraradices. Se ha demostrado que esta capacidad de bioprotección es específica de la
variedad de frijol empleada. Los resultados de dos variedades contrastantes: Az. Higuera
(susceptible en condiciones de micorrización) y Az. Regional 87 (tolerante en condiciones
de micorrización) nos impulsan a emplear la proteómica comparativa, para la identificación
de proteínas que pudieran estar involucradas en el mecanismo de defensa inducido por
micorrización y tratar de explicar más a fondo este fenómeno.

Quintero-Zamora, 2010 32
III. JUSTIFICACIÓN
Si bien existen diversos estudios para determinar los cambios de expresión de genes
y proteínas que ocurren en las raíces de las plantas colonizadas con HMA, pocos son los
estudios que se han realizado para entender los mecanismos moleculares involucrados en la
MA y cómo ésta afecta la expresión génica en la parte aérea de la planta.
La identificación de las proteínas expresadas específica o diferencialmente en
plantas colonizadas por HMA permitirá entender la complejidad de la interacción planta-
hongo. Dado que aún cuando la interacción se manifiesta de manera local dentro de los
tejidos de la raíz, el efecto es sistémico alterando de manera global la fisiología de la
planta.
Para esto se propone identificar aquellas proteínas relacionadas a la simbiosis MA
relacionadas a la inducción de tolerancia en el tejido foliar de dos variedades de frijol
mediante proteómica comparativa. Para esto, se emplearán las variedades Az. Regional 87
y Az. Higuera, las cuales responden diferencialmente al ataque de S. sclerotiorum cuando
están colonizadas con G. intraradices.
Por lo anterior, este trabajo tiene como finalidad aumentar el conocimiento respecto
a la interacción frijol-HMA. En parte por la importancia de este cultivo en la región, y por
otra parte desde un punto de vista básico para conocer específicamente las proteínas que
puedan estar funcionando como mensajeros, o como parte del mecanismo de la inducción
de la defensa sistémica por micorrización en plantas. El conocimiento de los mecanismos
inducidos por el establecimiento de la asociación MA en la parte aérea de frijol, permitirá
aprovechar los beneficios de dichos mecanismos en el futuro, empleando la inducción
controlada de resistencia a patógenos de importancia económica.

Quintero-Zamora, 2010 33
IV. HIPÓTESIS
• La colonización radical por HMA induce la resistencia a patógenos en plantas de
frijol dependiendo de la variedad y se encuentra mediada por la expresión
diferencial de proteínas en parte aérea.
• El comportamiento diferencial entre las variedades Azufrado Regional 87 y
Azufrado Higuera colonizadas con G. intraradices con respecto a resistencia al
patógeno S. sclerotiorum, permitirá establecer los mecanismos moleculares
relacionados a la resistencia inducida por la simbiosis.
V. OBJETIVOS
5.1. Objetivo general
• Identificar proteínas diferenciales en hoja relacionadas a la resistencia que se induce
en frijol en respuesta a la simbiosis con el hongo Glomus intraradices.
5.2. Objetivos específicos
• Estandarizar la técnica de extracción de proteínas en hojas de plantas de frijol.
• Identificar las proteínas diferenciales presentes en la parte aérea de plantas de frijol
de las variedades Azufrado Regional 87 y Azufrado Higuera en respuesta a la
colonización con Glomus intraradices, mediante electroforesis bidimensional y
espectrometría de masas.
• Determinar las rutas metabólicas o mecanismos moleculares que están siendo
inducidas en la parte aérea de la planta por micorrización en frijol.

Quintero-Zamora, 2010 34
VI. MATERIALES Y MÉTODOS
6.1. Tejido vegetal
Se utilizó material criopreservado a -70 °C de hojas de plantas colonizadas con G.
intraradices y no colonizadas de dos variedades de frijol (Az. Regional 87 y Az. Higuera),
todo el material es proveniente de experimentos realizados por Mora-Romero (2008). La
estandarización del método de extracción se llevó a cabo con hojas de plantas de frijol de la
variedad Az. Regional 87 sin colonizar. Las plantas se tomaron del experimento fisiológico
preliminar en hoja desprendida realizado por Mora-Romero (2008). El material se preparó
de la siguiente manera: las semillas fueron desinfectadas en etanol al 70% durante 2 min,
seguido por 2 min de inmersión en hipoclorito de sodio al 0.5% y enjuagadas de tres a
cuatro veces con agua destilada estéril, sembradas en macetas de plástico, utilizando como
sustrato Peat Moss. Ocho días posteriores a la germinación las plantas fueron trasplantadas
a macetas individuales. Las plantas fueron fertilizadas semanalmente con 50 ml de solución
nutritiva de Hoagland (Ca(NO3)2·4H2O 2.5 mM, KNO3 2.5 mM, MgSO4·7 H2O 1 mM ,
Na-FeEDTA 50 μM, KH2PO4 20 μM, H3BO3 10 μM, Na2MoO4·2H2O 0.2 μM,
ZnSO4·7H2O 1.0 μM, MnCl2·4H2O 2.0 μM, CuSO4·5H2O 0.5 μM, CoCl2·6 H2O 0.2 μM,
HCl 3N 25.0 μM, MES Amortiguador 0.5 mM) (Millner y Kitt, 1992), regadas con agua
corriente según se requería y mantenidas bajo condiciones de invernadero, cubiertas con
malla antiáfidos durante diez semanas. Las plantas fueron crecidas de enero a marzo de
2007 (Figura 2).
Figura 2. Plantas de frijol de la variedad Az. Regional 87, mantenidas en estantes cubiertos con mallas antiáfidos bajo condiciones de invernadero (tomado de Mora-Romero, 2008).

Quintero-Zamora, 2010 35
Para llevar a cabo el análisis mediante proteómica comparativa se utilizaron
muestras de hojas de plantas de dos variedades de frijol (Az. Regional 87 e Higuera)
colonizadas con G. intraradices y no colonizadas. Las muestras provienen del crecimiento
y colonización de plantas de frijol para el experimento en planta completa, donde las
semillas de dos variedades de frijol (Az. Regional 87 y Az. Higuera) fueron desinfectadas
bajo el mismo procedimiento anterior, sembradas en macetas de plástico, utilizando como
sustrato vermiculita-arena (1:1; v/v), esterilizado a 121 °C, en autoclave, durante 1 hora a
15 lb de presión. Ocho días después de la germinación cada planta fue inoculada con
aproximadamente 500 esporas de G. intraradices contenidas en un ml de agua, y
trasplantadas a macetas individuales. Las plantas fueron fertilizadas semanalmente con 50
ml de solución nutritiva de Hoagland (2x) (composición completa de Hoagland con 20 μM
de fosfato de potasio) (Millner y Kilt, 1992), y regadas con agua corriente según se
requería. Las plantas fueron crecidas bajo condiciones controladas en cámaras de
crecimiento BINDER (Fotoperíodo: 18/8 hr luz/oscuridad; temperatura: 25/18 °C) (Figura
3) durante el período de abril a mayo. A las ocho semanas después de la adición de esporas
se midió el porcentaje de colonización con G. intraradices, obteniendo para la variedad Az.
Regional 87 un 23% y para Az. Higuera un 20% ((± 10 desviación estándar: DS).
Figura 3. Plantas de frijol de las variedades Az. Regional 87 y Az. Higuera, cultivadas
bajo condiciones controladas en cámaras de crecimiento (tomado de Mora-Romero, 2008).

Quintero-Zamora, 2010 36
6.2. Estandarización del método de extracción de proteínas
Se compararon cuatro diferentes métodos de extracción de proteínas en hojas de
plantas de frijol de la variedad Az. Regional 87. Se analizaron tres muestras biológicas por
duplicado. El método que resultó más eficiente con respecto a la solubilización de proteínas
y su compatibilidad con el análisis 2D-SDS-PAGE para la subsecuente identificación por
LC-MS/MS, es el que se tomó para llevar a cabo la extracción de hojas en plantas de frijol
colonizadas con G. intraradices y no colonizadas de las variedades Az. Regional 87 y Az.
Higuera. Una vez extraídas las muestras anteriormente mencionadas, se llevó a cabo
mediante proteómica comparativa la identificación de proteínas diferenciales presentes en
la parte aérea de la planta en respuesta a la micorrización con G. intraradices, mediante
electroforesis bidimensional para posteriormente identificar las proteínas diferenciales
mediante espectrometría de masas. Finalmente, determinamos las rutas metabólicas o
mecanismos moleculares que están siendo inducidas en la parte aérea de la planta por
micorrización en frijol.
Los métodos de extracción de proteínas son descritos en el apartado siguiente:
6.2.1. Método A de extracción con Fenol (Fenol-A)
Se pulverizó en presencia de nitrógeno líquido aproximadamente 250 mg de tejido
de hoja de planta de frijol congelado, empleando un molino (Retsh MM 301, Haan
Alemania) a una velocidad de 30 oscilaciones/segundo por 5 min. El tejido pulverizado se
resuspendió en 500 µl de amortiguador buffer de extracción (polivinilpolipirrolidona
(PVPP) 1% (p/v), sacarosa 0.7 M, KCl 0.1 M, Tris-HCl pH 8.0 al 0.5 M, EDTA 50 mM, 2-
mercaptoetanol 0.4% (v/v) y coctel de inhibidor de proteasas 1 mM). La mezcla fue
homogenizada en frío durante 15 min usando como homogenizador el equipo Disperser
T10 Basic (IKA, Staufen, Suiza). Se adicionó un volumen igual de fenol saturado con Tris-
HCl 0.1 M a pH 7.5 y se volvió a homogenizar por 15 min en frío. Se centrifugó a 10,000 x
g por 30 min a 4 °C. Se removió la fase superior de fenol y se volvió a extraer con
amortiguador de extracción. Las proteínas precipitaron en la fase final de fenol con cinco
volúmenes de acetato de amonio 0.1 M en metanol toda la noche a -20 ºC y se obtuvo la

Quintero-Zamora, 2010 37
pastilla por centrifugación a 10,000 x g por 30 min. Finalmente se resuspendió en 100 µl de
urea-tiourea (7M-2M) (Hurkman y Tanaka, 1986).
6.2.2. Método B de extracción con Fenol (Fenol-B)
Aproximadamente 250 mg de tejido pulverizado como se describió anteriormente se
resuspendió en 500 µl de amortiguador de extracción consistiendo de Tris-HCl (pH 8.8) 0.1
M, conteniendo DTT 50 mM, EDTA 10 mM, coctel de inhibidor de proteasas 1 mM. Se
homogenizó durante 15 min usando el Disperser T10 Basic (IKA). Después se centrifugó a
10,000 x g durante 10 min a 4 °C, el sobrenadante clarificado se combinó en una relación
1:1 con fenol Tris-saturado (pH 7.5) y se mezcló con un vortex a velocidad media, a
temperatura ambiente de 20 a 30 min, y se centrifugó a 10,000 x g por 10 min a 4 °C. La
fracción acuosa superior extraída anteriormente se combinó otra vez en una relación 1:1
con fenol Tris-saturado (pH 7.5), se mezcló con vortex a velocidad media, a temperatura
ambiente de 20 a 30 min, y se centrifugó a 10,000 x g por 10 min a 4 °C. Las dos fases de
fenol fueron combinadas y re-extraídas con Tris-HCl (pH7.5) 100 mM con DTT 10 mM.
Después, 4 volúmenes de metanol 100% conteniendo acetato de amonio 100 mM y DTT 10
mM (refrigerado a -70 °C) se agregó a las fracciones fenólicas agrupadas y precipitadas por
2 a 3 h a -70 °C y entonces centrifugadas a 10,000 x g por 10 min a 4 °C. Las proteínas se
lavaron 2 ó 3 veces con metanol 100% conteniendo acetato de amonio 100 mM y DTT 10
mM con centrifugación a 10,000 x g por 10 min y después con varios volúmenes de etanol
frío al 90% (v/v) con DTT 10 mM y centrifugado a 10,000 x g por 10 min. La pastilla fue
disuelta en urea 7 M, tiourea 2 M, CHAPS 5% (p/v), triton X-100 2% (v/v), DTT 100 mM,
y anfolitos pH 3-10 al 1% (v/v) y agitada con vórtex a velocidad media por una hora a
temperatura ambiente. Las soluciones de las proteínas fueron clarificadas por
centrifugación a máxima velocidad (Sarma et al., 2008).
6.2.3. Método A de extracción con Ácido Tricloroacético/Acetona (TCA-A)
Aproximadamente 250 mg de hoja de tejido pulverizado como se describió
anteriormente se resuspendió en 500 µl de amortiguador de extracción
(Polivinilpolipirrolidona (PVPP) 1% (p/v), KCl 0.1 M, Tris-HCl pH 8.0 al 0.5 M, EDTA 50
mM, 2-mercaptoetanol 0.4% (v/v) y coctel de inhibidor de proteasas 1 mM). La mezcla se

Quintero-Zamora, 2010 38
homogenizó usando el Disperser T10 Basic (IKA) durante 15 min y se centrifugó a 10,000
x g por 30 min a 4 °C. Se recuperó el sobrenadante y las proteínas se precipitaron
agregándole cuatro volúmenes de acetona fría conteniendo TCA 10% (p/v) y 2-
mercaptoetanol 2% (v/v). La mezcla fue homogenizada y las proteínas fueron precipitadas
en congelación a -20 °C durante toda la noche. Se recuperó la pastilla por centrifugación a
10,000 x g por 30 min. Finalmente, se resuspendió en urea-tiourea (7 M-2 M) (Saravanan y
Rose, 2004).
6.2.4. Método B de extracción con Ácido Tricloroacético/Acetona (TCA-B)
Aproximadamente 250 mg de tejido pulverizado como se describió anteriormente se
homogenizó con 500 µl de solución que contenía TCA 10% (p/v) en acetona con 2-
mercaptoetanol 0.07% (v/v), coctel de inhibidor de proteasas 0.001 M usando el Disperser
T10 Basic (IKA) durante 15 min. Las proteínas totales fueron precipitadas durante toda la
noche a -20 °C. La pastilla se recuperó por centrifugación a máxima velocidad durante 30
min a 4 °C. Se resuspendió en 1 ml de amortiguador lisis (urea 7 M, tiourea 2 M, CHAPS
2% (p/v), Tritón 2% (v/v), anfolitos 0.2% (v/v) [pH 3–10] y TBP 4 mM). El material
insoluble se removió por centrifugación a máxima velocidad por 20 min a 4 °C, y se
almacenó el sobrenadante a -70 °C hasta su uso en el análisis 2D–SDS-PAGE (Cascardo et
al., 2001).
6.3. Cuantificación de proteínas
Una vez resuspendida la pastilla se cuantificó empleando el reactivo de Bradford
(Sigma, San Louis, MO, EUA). Para esta cuantificación se realizó una curva estándar con
tres repeticiones. Se obtuvo la absorbancia de la mezcla del reactivo con la proteína a 595
nm en un espectrofotómetro (Thermospectronic, Genesis Spectrophotometer Spectronic).
Se preparó una curva estándar con concentraciones de proteínas conocidas (rango de 0.25 a
1.40 mg/ml) con albúmina de suero bovino (Sigma, San Louis, MO, EUA), corridas en
paralelo con soluciones de proteínas desconocidas para analizar cuantitativamente las
proteínas, al mismo tiempo se corrió un blanco como control de la reacción.
Una vez obtenida la concentración de la muestra se calculó el rendimiento (mg/250
mg de tejido) promedio para cada método de extracción. A los resultados se les realizó un

Quintero-Zamora, 2010 39
análisis de varianza con una P<0.05, para observar si había diferencias significativas entre
los métodos en cuanto a rendimiento de proteína.
6.4. Protocolo de limpieza de proteínas para 2D-SDS-PAGE
Antes de realizar la electroforesis bidimensional todas las muestras fueron tratadas
con el kit de limpieza 2-D Clean Up Kit (Amersham Biosciences, no. de catálogo 80-6484-
51) el cual consistió en el siguiente procedimiento:
Se transfirieron 80 µg de muestra de proteína en un tubo Eppendorf de 1.5 ml. Se le
agregó 300 µl de solución precipitante (Amersham Biosciences, no. de catálogo 80-6484-
51) se mezcló bien por vórtex. Se incubó en hielo durante 15 min. Posteriormente, se
agregaron 300 µl de solución co-precipitante (Amersham Biosciences, no. de catálogo 80-
6484-51) a la mezcla anterior (mezcla de proteínas y precipitante) y se mezcló con vórtex a
baja velocidad. Se centrifugó a 4 °C, a 12,000 x g por 5 min. Se observó la formación de
una pequeña pastilla en este paso. Se removió con mucho cuidado el sobrenadante por
decantación o pipeteo. Después se agregaron 40 µl de solución de co-precipitado sobre la
pastilla y se colocaron los tubos en hielo durante 5 min. Se volvió a centrifugar a 4 °C, y
12,000 x g durante 5 min. Se agregaron 25 µl de agua destilada desionizada estéril (a 4 °C)
sobre la pastilla. Se agitó en vórtex cada tubo durante 5-10 seg. Posteriormente, se agregó 1
ml de amortiguador de lavado (wash buffer, Amersham Biosciences, no. de catálogo 80-
6484-51) (almacenado a –20 °C) y 5 µl de aditivo de lavado (wash aditive, Amersham
Biosciences, no. de catálogo 80-6484-51). Se agitó con vórtex hasta que la pastilla estuvo
completamente dispersa. Se incubó a -20 °C durante 30 min (se agitó la suspensión
mediante vortex de 20 a 30 seg cada 10 min). Se centrifugó a una velocidad de 12,000 x g
durante 5 min. Cuidadosamente se removió y descartó el sobrenadante. Se secó la pastilla
por no más de 5 min. Finalmente se resuspendió la pastilla en 130 µl de amortiguador de
IEF (De-streak Rehydration Solution no. de catálogo 17-6003-19 de GE Healthcare) para
continuar con la rehidratación e hidratación de las tiras.
6.5. Separación de proteínas por punto isoeléctrico [Isoelectroenfoque (IEF)]
En esta etapa de estandarización se realizó la separación de proteínas en escala
analítica. Se tomaron 80 µg de proteínas en 125 µl de amortiguador IEF (De-streak

Quintero-Zamora, 2010 40
Rehydration Solution) adicionando anfolitos de rango de pH 3-10 al 1.0% (p/v) y se
homogenizó. Se empleó esta muestra para rehidratar tiras de gel de poliacrilamida en
gradiente inmovilizado pH 3-10 (Ready Strip IPG Strip, 7 cm, BioRad). La rehidratación
consistió en 14 h a 50 V/20 ºC. Después de la etapa de rehidratación, las proteínas se
enfocaron en el sistema Protean IEF Cell (BioRad) aplicando un total de 22,000 V/h. Las
tiras tratadas se almacenaron a -70 °C hasta su aplicación en la segunda dimensión. En una
primera etapa se realizó en formato analítico lo que nos permitió identificar las zonas de pH
y peso molecular donde se observaron diferencias, posteriormente se emplearon tiras de
IPG Strip de 17 cm con el objetivo de tener suficiente proteína para identificación. Se
utilizó un marcador de peso molecular (2D-SDS-PAGE Standards, BioRad, Cat. no. 161-
0320), el cual se separó en paralelo a las muestras.
6.6. Separación de proteínas por peso molecular (SDS-PAGE)
El paso de la primera a la segunda dimensión involucró dos etapas: primero, la
reducción, y segundo la alquilación de los grupos sulfhidrilos. Las tiras de IPG fueron
equilibradas adicionando 2.0 ml de amortiguador de equilibrio (urea 6 M, SDS 2% (p/v),
Tris/HCl 0.05 M pH 8.8, glicerol 20% (v/v), DTT 2% (p/v) para reducir los grupos
sulfhidrilos se agitó durante 15 min. Posteriormente, se decantó y se adicionó el
amortiguador de equilibrio II (urea 6 M, SDS 2% (p/v), Tris/HCl 0.05 M, pH 8.8, glicerol
20% (v/v), iodo-acetamida 2.5% (p/v)) con agitación durante 15 min, para alquilar los
grupos sulfhidrilos. Se agitaron las tiras con el amortiguador I y II para correr en una
segunda dimensión.
Las proteínas resueltas y equilibradas, se separaron en base a su peso molecular en
geles desnaturalizantes en condiciones reductoras (SDS-PAGE). Se utilizó un marcador de
peso molecular (BenchMark 10 a 220 kDa (Invitrogen, Carlsbad CA, EUA)). La corrida
electroforética se realizó en una cámara vertical (Mini Protean Tetra Electrophoresis
System, Bio-Rad) a temperatura ambiente y 70 V, por 4 horas y en buffer de electroforesis
1X (Tris base, 25mM, glicina 192 mM, SDS 0.1% (p/v)). Para visualizar las manchas de
proteínas, los geles fueron teñidos con azul de Coomasie R-250.

Quintero-Zamora, 2010 41
6.7. Adquisición y análisis de imágenes
Después de realizar la corrida de SDS-PAGE, los geles se tiñeron en solución de
tinción (azul de Coomassie R-250 0.1% (p/v), metanol:ácido acético glacial (45:45, (v/v)),
con agitación suave durante toda la noche, después se destiñó en solución de destinción
(ácido acético glacial:metanol (10:10, (v/v)) por al menos 3 horas en agitación.
Una vez estandarizado el método, se tomó el método más eficiente y más fiable para
llevar a cabo el análisis proteómico comparativo entre las variedades Az. Regional 87 y Az.
Higuera de plantas no colonizadas y colonizadas con G. intraradices. Una vez teñidas las
proteínas, se documentaron en el sistema Chemi Doc (BioRad). Las imágenes de los geles
bidimensionales se analizaron empleando el programa PDQuest (BioRad), analizándose al
menos tres muestras biológicas para cada condición.
6.8. Identificación de proteínas mediante espectrometría de masas
Una vez realizada la comparación, se seleccionaron proteínas para ser identificadas
mediante espectrometría de masas. Las manchas de las proteínas de interés fueron cortadas
manualmente de los geles teñidos con azul de Coomassie. Dado que existen bases de datos
de secuencias de transcritos de frijol y de otras leguminosas, fue factible emplear
cromatografía líquida masas/masas (LC-MS/MS). Este análisis se realizó en la Unidad de
Proteómica del Instituto de Biotecnología de la UNAM en Cuernavaca, Morelos. Las
muestras fueron digeridas “en el gel” con tripsina y los péptidos
resultantes fueron aplicados en un sistema LC-MS constituido de un cromatógrafo líquido
de micro-flujo Accela (Thermo-Ficher Co. San Jose, CA) con “spliter” (1/20) y un
espectrómetro de masas LTQ-Orbitrap XL (Thermo-Ficher Co.) con sistema de ionización
tipo nano-electrospray (ESI). Para la fragmentación de los péptidos
se utilizó los métodos de CID (Collision-Induced Dissociation) y HCD (High-energy
Collision Dissociation). Solamente iones con carga 2+ y 3+ fueron seleccionados para los
eventos de fragmentación. No fueron considerados los iones con cargas 1+, 4+ y de cargas
indefinidas. Todos los espectros fueron adquiridos en modo de detección positivo. Durante
la captura automática de los datos fue utilizada la exclusión dinámica de iones, con un
tiempo de exclusión de 60 seg. Las secuencias de los péptidos resultantes se analizaron en
la base de datos del NCBI, mediante el empleo del programa BlastP

Quintero-Zamora, 2010 42
(http://www.ncbi.nlm.nih.gov/BLAST). El peso molecular y el punto isoeléctrico de los
polipéptidos fueron evaluados en EXPASy (http://www.expasy.org/tools/pi_tool) para la
confirmación final de acuerdo a sus posiciones en el mapa del gel bidimensional.

Quintero-Zamora, 2010 43
VII. RESULTADOS Y DISCUSIÓN
7.1. Estandarización del método de extracción de proteínas
Este estudio fue imprescindible para evaluar, estandarizar y seleccionar el método
más eficiente para el análisis de las proteínas en hojas de plantas de frijol, ya que no
existían protocolos previamente reportados para este cultivo en la literatura.
Se evaluaron cuatro diferentes métodos de extracción de proteínas de hojas de
plantas de frijol. Esto con la finalidad de encontrar un protocolo que permitiera la
solubilización de la mayor cantidad y resolución de proteínas en el análisis por 2D-SDS-
PAGE (separación tanto por punto isoeléctrico, como por peso molecular) y la subsecuente
identificación por espectrometría de masas.
Primeramente, las muestras proteicas obtenidas de los distintos métodos se
cuantificaron empleando el reactivo de Bradford, ya que este método es compatible con
agentes reductores y bajas concentraciones de detergentes. Para esta cuantificación se
obtuvo una curva estándar (Figura 4), con el promedio de tres réplicas a concentraciónes
de 0.25, 0.50, 0.75, 1.0, 1.4 mg/ml de proteína albúmina de suero de bovino, con una
longitud de onda de 595 nm. El criterio de elección tomado en consideración fue un
coeficiente de determinación (R2) mayor a 0.95. Los resultados obtenidos muestran un (R2)
de 0.9991. Esto indica que el modelo ajustado es capaz de explicar la respuesta
(absorbancia) a partir de los valores de concentración. De manera que el intervalo de
concentración comprendido, satisface las condiciones de linealidad del método analítico.
Con la ecuación obtenida y = 0.4299x-0.0112, se calculó la concentración desconocida (x)
que corresponde a cada una de las muestras de concentrado de proteinas.

Quintero-Zamora, 2010 44
Figura 4. Curva estándar para determinar la concentración de proteína. Se
cuantificó con el reactivo de Bradford con albúmina de suero de bovino (BSA).
Los rendimientos de los métodos evaluados se muestran en el Cuadro 1. El
promedio de los rendimientos fue aproximadamente de 0.42 ± 0.1 mg de proteína/250 mg
de tejido vegetal fresco, siendo el mayor el del método TCA A (0.513 mg/250 mg de
tejido).
Cuadro 1. Rendimientos de extracción de proteínas de hojas de plantas mediante cuatro diferentes métodos
Método Concentración Rendimiento de extracción (mg/ml)1 mg/250mg de tejido2 1 Fenol A 3.02 ± 0.1 0.302 2 Fenol B 3.96 ± 0.1 0.396 3 TCA A 5.13 ± 0.1 0.513 4 TCA B 3.09 ± 0.1 0.464
1. Dos extracciones por método y cuantificación por duplicado. 2. Tejido congelado de hojas de plantas de frijol Az. Reg. 87.
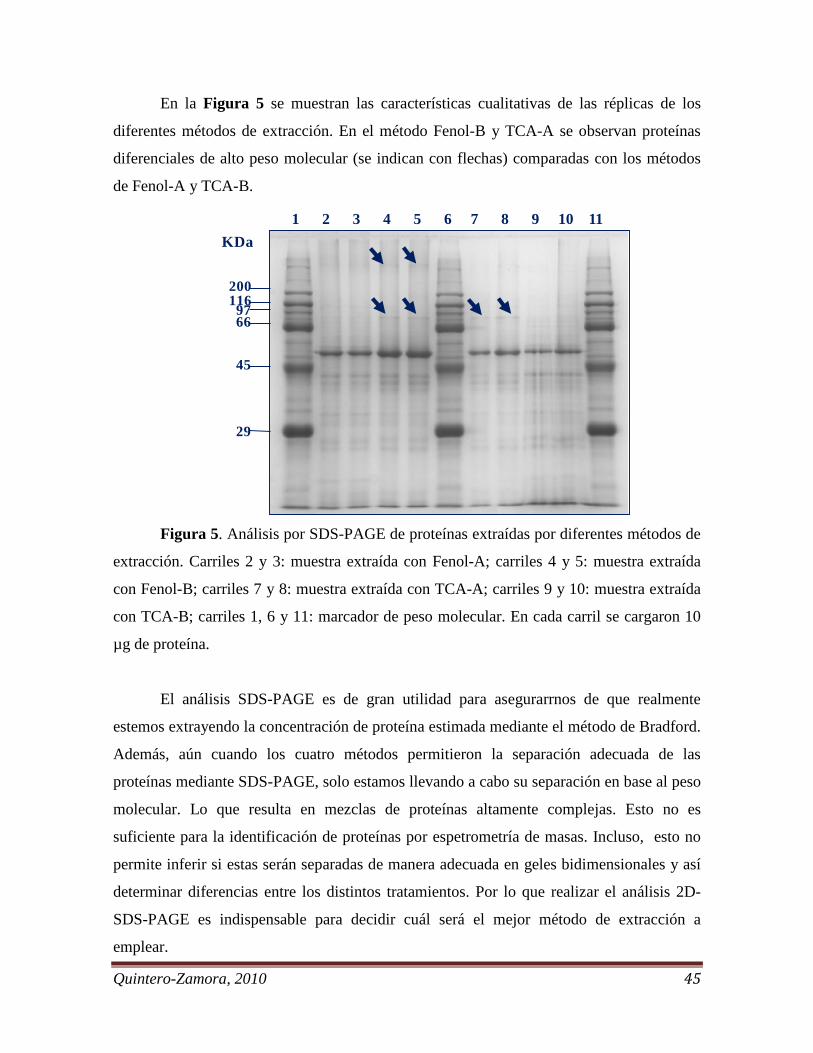
Quintero-Zamora, 2010 45
En la Figura 5 se muestran las características cualitativas de las réplicas de los
diferentes métodos de extracción. En el método Fenol-B y TCA-A se observan proteínas
diferenciales de alto peso molecular (se indican con flechas) comparadas con los métodos
de Fenol-A y TCA-B.
1 2 3 4 5 6 7 8 9 10 11
29
2001169766
45
KDa
Figura 5. Análisis por SDS-PAGE de proteínas extraídas por diferentes métodos de
extracción. Carriles 2 y 3: muestra extraída con Fenol-A; carriles 4 y 5: muestra extraída
con Fenol-B; carriles 7 y 8: muestra extraída con TCA-A; carriles 9 y 10: muestra extraída
con TCA-B; carriles 1, 6 y 11: marcador de peso molecular. En cada carril se cargaron 10
µg de proteína.
El análisis SDS-PAGE es de gran utilidad para asegurarrnos de que realmente
estemos extrayendo la concentración de proteína estimada mediante el método de Bradford.
Además, aún cuando los cuatro métodos permitieron la separación adecuada de las
proteínas mediante SDS-PAGE, solo estamos llevando a cabo su separación en base al peso
molecular. Lo que resulta en mezclas de proteínas altamente complejas. Esto no es
suficiente para la identificación de proteínas por espetrometría de masas. Incluso, esto no
permite inferir si estas serán separadas de manera adecuada en geles bidimensionales y así
determinar diferencias entre los distintos tratamientos. Por lo que realizar el análisis 2D-
SDS-PAGE es indispensable para decidir cuál será el mejor método de extracción a
emplear.

Quintero-Zamora, 2010 46
7.2. Electroforesis bidimensional (2D-SDS-PAGE)
Para una mejor separación de mezclas de proteínas, se llevó a cabo la 2D-SDS-
PAGE. Los cuatro métodos evaluados (Figura 6, 7, 8 y 9) han sido reportados previamente
para otros tejidos vegetales. Las imágenes son representativas del análisis por duplicado. La
comparación cuantitativa de extractos de proteínas usando los cuatro protocolos de
extracción reveló que el método de extracción con TCA-A fue el mejor en cuanto a
cantidad (20 spots) y calidad de proteínas resueltas en el análisis 2D-SDS-PAGE (Figura
8). Aún cuando en el análisis en primera dimensión (Figura 5) no evidenció muchas
diferencias en cuanto al patrón de proteínas obtenido por los diferentes métodos; en el
análisis 2D-SDS-PAGE hubo menor cantidad de spots resueltos para los métodos de Fenol-
A (10 spots, Figura 6), Fenol-B (8 spots, Figura 7) y TCA-B (4 spots, Figura 9). Es difícil
determinar los factores por los cuales se obtuvieron mejores resultados con el método de
TCA-A. Esto debido al análisis multifactorial de la muestra.
Estos resultados coinciden con los de Saravanan y Rose (2004) donde el método de
extracción con TCA proporciona mejoras en 2D-SDS-PAGE en base al análisis proteómico
de la mayoría de los tejidos vegetales. Pridmore et al. (1999) y Natarajan et al. (2005)
utilizaron un método de precipitación con TCA el cual aumentó la resolución de manchas
de proteínas individuales debido en parte a la inhibición de la actividad proteolítica.
Cascardo et al. (2001), Damerval et al. (1986) y Santoni et al. (1994) demostraron una alta
resolución de proteínas mediante 2D-SDS-PAGE usando el método de precipitación con
TCA en Arabidopsis thaliana y diferentes líneas de semillas de trigo.

Quintero-Zamora, 2010 47
3 10pI
M
1510
20
3025
506070100
KDa
Figura 6. Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el método de
extracción con Fenol-A. M. Indica marcador de peso molecular. 80 µg de proteínas fueron
enfocados en tiras IPG (pH 3-10, 7cm).
KDa3 10
pIM
29
45
6697
116
Figura 7. Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el método de
extracción con Fenol-B. M. Indica marcador de peso molecular. 80 µg de proteínas fueron
enfocados en tiras IPG (pH 3-10, 7cm).

Quintero-Zamora, 2010 48
15
2530
4050
7060
20
100KDa
3 10pI
M
Figura 8. Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el método de
extracción con TCA-A. M. Indica marcador de peso molecular. 80 µg de proteínas fueron
enfocados en tiras IPG (pH 3-10, 7cm).
3 10pI
M
25
30
40
50
7060
20
100KDa
80
Figura 9. Análisis 2D-SDS-PAGE de proteínas de hojas de frijol con el método de
extracción con TCA-B. M. Indica marcador de peso molecular. 80 µg de proteínas fueron
enfocados en tiras IPG (pH 3-10, 7cm).

Quintero-Zamora, 2010 49
En el Cuadro 2 se presenta la cantidad de proteínas y el rango de pH en el que se
encuentran para cada método. Cabe mencionar que el número de proteínas encontradas en
el mejor método (TCA-A) son sólo 20 spots. Este número se verá incrementado en los geles
preparativos, donde se analizarán aproximadamente 500 µg por gel, es decir, cinco veces
más a lo empleado en estos ensayos analíticos.
Cuadro 2. Proteínas (spots) resueltas por cada uno de los diferentes métodos de
extracción.
Método de extracción No. de Spots Rango de pH
Fenol-A 10 5 a 7
Fenol-B 8 5 a 7
TCA-A 20 5 a 8
TCA-B 4 5 a 7
7.3. Análisis proteómico de plantas colonizadas con G. intraradices vs. no colonizadas
en dos variedades de frijol
7.3.1 Extracción en hojas de frijol
Para la identificación de patrones de proteínas inducidos en el proteoma de hoja de
frijol Az. Regional 87 e Higuera por la colonización con G. intraradices, el total de
proteínas de hojas de plantas de frijol inoculadas y no inoculadas fueron extraídas.
La extracción de las proteínas para evaluar el efecto de la micorrización en el tejido
foliar de frijol se llevó a cabo con el método modificado TCA-A (Saravanan y Rose, 2004)
descrito en la sección 6.2.3. Las extracciones de las diferentes condiciones (variedades Az.
Higuera y Az. Regional 87 ambas colonizadas y no colonizadas) se observaron
homogéneas al analizarse mediante SDS-PAGE (Figura 10), aún cuando se comparan las
proteínas de las dos distintas variedades de frijol, lo que muestra la limitante de la
comparación de proteínas separadas sólo en base a su peso molecular.

Quintero-Zamora, 2010 50
Figura 10. Análisis mediante electroforesis en gel desnaturalizante (SDS-PAGE) de
proteínas extraídas de hoja mediante el método de extracción TCA-A. Carril 1: marcador de
peso molecular; carril 2: frijol Az. Regional 87 colonizada con G. intraradices; carril 3:
frijol Az. Regional 87 no colonizado; carril 4: frijol Az. Higuera colonizado con G.
intraradices; carril 5: frijol Az. Higuera no colonizado. En cada carril se cargaron 10 µg de
proteína.
7.3.2. Perfil de proteínas inducido en hojas de frijol
En las Figuras 11, 12, 13 y 14 se muestran el patrón electroforético bidimensional
(2D-SDS-PAGE) de cada una de las plantas analizadas, en cada figura se exponen tres
geles bidimensionales que corresponden a las réplicas biológicas de los experimentos
realizados por Mora-Romero (2008). Las imágenes de los geles corresponden al formato
preparativo en tiras de gel de poliacrilamida en gradiente inmovilizado de 17 cm, pH 4-7,
de proteínas extraídas en hoja de plantas de frijol de dos variedades Az. Regional 87 y Az.
Higuera colonizadas con G. intraradices y no colonizadas.

Quintero-Zamora, 2010 51

Quintero-Zamora, 2010 52

Quintero-Zamora, 2010 53
Las imágenes de los geles preparativos bidimensionales se obtuvieron con la
finalidad de ser analizadas mediante el software PDQuest. Este software lleva a cabo el
análisis cualitativo o cuantitativo de proteínas mediante los estadísticos t-student y/o
mínimos cuadrados, promediando la intensidad por área (densidad del spot en pixeles) de
los spots (Cuadro 3) en base a tres réplicas (imágenes de geles bidimensionales) por
condición.
El análisis del perfil proteico de la variedad Az. Higuera colonizada con G.
intraradices comparado con la condición no colonizada de la misma variedad se observa
mediante un gel maestro (master gel) con el software PDQuest (Figura 15). Se observaron
un promedio de 200 proteínas (spots) distribuidas en un rango de pH de 4 a 7 y con un peso
molecular de 10 a 100 kDa. El análisis comparativo de proteínas reveló 5 proteínas
diferenciales las cuales disminuyeron su expresión de manera significativa en plantas
colonizadas comparadas con las no colonizadas (Cuadro 4) de acuerdo a los estadísticos t-
student y/o mínimos cuadrados (algunas proteínas resultaron diferenciales mediante los dos
estadísticos, spots en rojo y amarillo, Figura 15). Cabe mencionar que de acuerdo a Mora-
Romero (2008) la variedad Az. Higuera al encontrarse colonizada por G. intraradices no
mostró diferencias a la no colonizada respecto a la tolerancia contra el patógeno moho
blanco, como sí se observó en la variedad Az. Regional 87. Las pocas diferencias
observadas en los patrones de proteínas de Az. Higuera, podrían coincidir con los pocos
cambios a nivel fisiológico reportados previamente.

Quintero-Zamora, 2010 54

Quintero-Zamora, 2010 55
Figura 15. Gel maestro comparativo entre las proteínas de la variedad de frijol Az.
Higuera, colonizadas con G. intraradices vs. no colonizadas, obtenido en el
programa PDQuest.
El perfil proteico de la variedad Az. Regional 87 colonizada con G. intraradices al
ser comparado con la condición no colonizada de la misma variedad muestra un promedio
de 200 proteínas; las cuales tienen un intervalo en un rango de pH 4-7 y una masa
molecular de 20 a 100 kDa (Figura 16). El intervalo de las proteínas fue similar a lo
encontrado para las dos condiciones de la variedad Az. Higuera. El análisis comparativo del
patrón de proteínas reveló 14 proteínas diferenciales: una proteína cualitativa (que se
encuentra en las plantas colonizadas y no en las plantas sin colonizar, indicada en azul en
Figura 16) y 13 proteínas cuantitativas donde 11 aumentan su expresión y dos la
disminuyen en cuanto a plantas micorrizadas comparadas con las plantas no micorrizadas
(Cuadro 5) de acuerdo a los estadísticos t-student y/o mínimos cuadrados (algunas
proteínas resultaron diferenciales mediante los dos estadísticos, spots en rojo y amarillo,
Figura 16).
De acuerdo a Mora-Romero (2008), la variedad Az. Regional 87 mostró un
incremento a la tolerancia al moho blanco al encontrarse bajo la condición micorrízica, esto
en tejido foliar, por lo que es de gran interés la interpretación de las 14 proteínas
diferenciales, donde 13 de estas aumentaron su expresión en la condición micorrízica.

Quintero-Zamora, 2010 56
Figura 16. Gel maestro comparativo entre las proteínas de la variedad de
frijol Az. Regional 87, colonizadas con G. intraradices vs. no colonizadas, obtenido
en el programa PDQuest.
Para darnos una idea de qué tanto aumentaron o disminuyeron los spots inducidos,
se llevó a cabo una relación de densidad de spots. Se obtuvo el promedio de densidad del
spot (pixeles) de tres réplicas por condición (Cuadro 4 y 5). La relación de densidad se
obtuvo en base a plantas colonizadas con G. intraradices con respecto a no colonizadas de
cada variedad. La flecha indica un aumento (↑) o una disminución (↓) en la expresión de
proteínas de plantas colonizadas con G. intraradices con respecto a no colonizadas, la
ausencia de flecha indica una proteína cualitativa (Cuadro 4 y 5).

Quintero-Zamora, 2010 57
Cuadro 4. Relación de densidad de proteínas expresadas diferencialmente en hojas
de plantas de frijol Az. Higuera
Código del Spot Expresión Relación de Densidad5203 ↓ 0.315803 ↓ 0.335804 ↓ 0.366302 ↓ 0.537001 ↓ 0.41
Cuadro 5. Relación de densidad de proteínas expresadas diferencialmente en hojas
de plantas de frijol Az. Regional 87.
Código del Spot Expresión Relación de Densidad37011205 ↑ 1.982302 ↑ 3.472802 ↑ 2.663201 ↑ 1.983801 ↑ 2.525205 ↑ 4.495503 ↑ 2.266201 ↑ 3.346301 ↑ 1.496404 ↑ 1.837801 ↑ 2.76301 ↓ 0.80
2401 ↓ 0.50
Cualitativa

Quintero-Zamora, 2010 58
Un total de 19 spots fueron inducidos (aumento o disminución en intensidad*area
del spot, en base a los estadísticos mencionados). Un aumento de expresión de proteínas de
plantas colonizadas vs. no colonizadas de las dos variedades, se presentó en una relación de
densidad que va de 1.49 a 4.49 y una disminución de expresión en un rango menor de 0.8.
Los spots presentados en el Cuadro 4 y 5 se enviaron a identificar mediante LC-MS/MS a
la Unidad de Proteómica del Instituto de Biotecnología de la UNAM en Cuernavaca,
Morelos. En la variedad Az. Higuera de 5 péptidos enviados se obtuvo la secuencia de 3
(Cuadro 6) y en la variedad Az. Regional 87 de 14 enviados se obtuvo la secuencia de 11
péptidos (Cuadro 7). Las secuencias fueron analizadas en la base de datos del NCBI
(Cuadros 8 y 9), los valores teóricos de punto isoeléctrico y peso molecular de las
proteínas de las secuencias obtenidas se analizaron con el programa EXPASY para la
confirmación final de acuerdo a sus posiciones en el mapa del gel 2D-SDS-PAGE.

Quintero-Zamora, 2010 59

Quintero-Zamora, 2010 60

Quintero-Zamora, 2010 61
7.4. Interpretación del papel funcional de proteínas implicadas en la simbiosis
micorrízica arbuscular
La descripción de cambios en transcritos o proteínas en parte aérea de la planta
como respuesta a la simbiosis micorrízica se plantea de interés, debido a las observaciones
de que la simbiosis incrementa la tolerancia a distintos tipos de estrés. En el caso de frijol,
se tienen evidencias en nuestro grupo de trabajo de la inducción de tolerancia por
micorrización a S. sclerotiorum, un hongo de importancia agronómica. Existen reportes
adicionales de la inducción de tolerancia por micorrización en otros sistemas planta-HMA-
patógeno foliar (Lingua et al., 2002; Fritz et al., 2006; Liu et al., 2007; Galindo-Flores,
2008), sin embargo los reportes son escasos. En el caso de la descripción de cambios
transcripcionales como respuesta a la simbiosis micorrízica, sólo existen dos reportes
basados en análisis de microarreglos en parte aérea de plantas colonizadas con HMA: Liu
et al. (2007) en M. truncatula, una planta leguminosa, y Fiorelli et al. (2009) en tomate,
una solanácea. El objetivo de este estudio fue la identificación de proteínas inducidas
diferencialmente en hojas de dos variedades de frijol, una que respondió a la colonización
induciendo tolerancia a S. sclerotiorum (Az Regional 87) y otra que no mostró tal
inducción por la colonización micorrízica con G. intraradices. Estas proteínas podrían estar
potencialmente relacionadas a la tolerancia observada contra el patógeno foliar. Cambios
significativos en el perfil de proteínas fueron detectados en las hojas de P. vulgaris en
asociación simbiótica con G. intraradices. Considerando las dos variedades de frijol, se
obtuvo la identificación de 14 proteínas expresadas diferencialmente en hojas en respuesta
a la simbiosis MA, tres para Az Higuera, y 11 para Az Regional 87. Dentro de estas
proteínas, algunas de ellas fueron detectadas como diferenciales en ambas variedades. Una
de las proteínas encontradas en ambas variedades no ha sido reportada previamente en
tejidos vegetales (spots 7001 y 1205), una identificada como una proteína de función
desconocida (spot 2302) y once proteínas cuya función ya ha sido descrita con
anterioridad (spots, 5803, 5804, 3701, 2802, 3801, 5503, 6201, 6301, 6404, 7801 y 2401)
(Cuadro 6 y 7).
Liu et al. (2007) identificaron 468 genes cuya expresión aumentó y 131 genes cuya
expresión disminuyó en hojas de plantas de M. truncatula colonizadas con G. intraradices
con respecto a hojas de plantas no colonizadas. Aproximadamente el 25% de los genes

Quintero-Zamora, 2010 62
inducidos en hojas de plantas colonizadas por HMA pudieron asociarse in silico con
proteínas involucradas en mecanismos de defensa descritos en otros sistemas, sugiriendo
que la micorrización podría estar induciendo un mecanismo de defensa en la parte aérea de
las plantas.
Peinado-Guevara (2010, en revisión) encontró 27 proteínas expresadas
diferencialmente en hojas de plantas de tomate colonizadas con G. intraradices comparadas
con plantas no colonizadas. La mayoría de las proteínas identificadas disminuyeron su
expresión en la condición colonizada y fueron proteínas relacionadas a la fotosíntesis y al
estado celular de óxido-reducción. Una proteína reportada en raíces de plantas colonizadas
cuyo papel funcional se desconoce y una proteína importante en defensa de plantas fueron
encontradas inducidas.
Por otro lado, Mora-Romero (2008) reportó que plantas de frijol de la variedad Az.
Regional 87 en la condición colonizada con G. intraradices muestran tolerancia al moho
blanco (S. sclerotiorum), ésto, a diferencia de la variedad Az. Higuera también en la misma
condición, la cual fue susceptible al patógeno bajo las mismas condiciones de cultivo. En
base a esto, se sugiere que las proteínas diferenciales pueden estar relacionadas a tal
tolerancia observada. En plantas colonizadas por HMA de la variedad capaz de inducir
tolerancia (Az. Regional 87), la mayoría de las proteínas aumentaron su expresión con
respecto a la otra variedad (Az. Higuera). Ya que Az. Higuera fue incapaz de inducir
tolerancia a S. sclerotiorum, y en esta variedad se presentó un número menor de proteínas
diferenciales (tres proteínas diferenciales), mientras que en Az Regional 87 se identificaron
11, de las cuales 10 aumentaron su expresión en la condición colonizada, es posible sugerir
que al menos algunas de las proteínas diferenciales identificadas en Az Regional 87,
pudieran estar relacionadas con un mecanismo de inducción de tolerancia por
micorrización, o preacondicionamiento, tal como el observado en respuesta inducida por
rizobacterias promotoras de crecimiento (Van Loon et al., 1998).

Quintero-Zamora, 2010 63
7.4.1. Análisis proteómico en hojas de plantas de frijol de la variedad Az. Higuera
susceptible y Az. Regional 87 tolerante a S. sclerotiorum en asociación con G.
intraradices
El análisis proteómico de planta colonizada con G. intraradices vs. no colonizada de
de la variedad Az. Higuera, la cual no mostró inducción de tolerancia a S. sclerotiorum
reveló cinco proteínas diferenciales, las cuales disminuyeron su expresión en plantas
colonizadas. De estas cinco proteínas, se lograron identificar tres, una de ellas fue
identificada como una proenzima llamada tripsinógeno (Cuadro 6: spot 7001) y las otras
dos corresponden a transcetolasas (Cuadro 6: spot 5803 y 5804).
El spot 7001 identificado como una proenzima llamada tripsinógeno y reprimida en
Az. Higuera, fue encontrado sobreexpresado en Az. Regional 87 (spot 1205). Este péptido
ha sido reportado en jabalí (Sus acrofa) y en otros tejidos animales. Muchas de las tripsinas
son sintetizadas como zimógenos o proenzimas los cuales son precursores inactivos de las
enzimas. Las proenzimas se rompen durante la proteólisis para generar sus formas activas.
Las enzimas digestivas, algunos factores de la coagulación y otras proteinas son
sintetizadas como zimógenos. Sin embargo, esta proteína no ha sido reportada previamente
en tejidos vegetales. Por lo que se sugiere llevar a cabo estudios mas detallados en base a la
funcionalidad de esta enzima en plantas colonizadas.
Los spots 5803 y 5804 (cuadro 6) los cuales en la variedad Az. Higuera disminuyen
su expresión en hojas plantas colonizadas con respecto a no colonizadas, fueron
identificados como transcetolasas. Una transcetolasa también se encontró inducida en hojas
de la variedad Az. Regional 87 (Cuadro 7: spot 6301), la cual corresponde al mismo
péptido encontrado en Az. Higuera (spot 5804). Sin embargo en Az. Regional 87 se obtuvo
un efecto contrario, en esta variedad esta proteína se encontró inducida en la condición
colonizada. Transcritos correspondientes a estos péptidos (Spots 5804 y 6301) se
encontraron expresados diferencialmente en hojas de plantas de Platanus acerifolia tratadas
con la proteína fitotóxica cerato-platanina (Fontana et al., 2008) la cual inhibe el
crecimiento del patógeno Ceratocystis. Sus resultados también muestran diferentes
proteínas involucradas en la fotosíntesis tal como una proteína ferredoxina A, la ribulosa
1,5-bifosfato carboxilasa oxigenasa y enzimas cloroplásticas del fotosistema I y II. En
nuestros resultados, además de transcetolasas también encontramos otras proteínas

Quintero-Zamora, 2010 64
inducidas relacionadas a fotosíntesis (Cuadro 7: spots 2802, 3801, 5503, 6201, 6301, 6404
y 7801). En la variedad Az. Regional 87 (la cual muestra tolerancia a S. sclerotiorum en
condiciones de micorrización) se encontró un mayor perfil de proteínas inducidas en el
análisis proteómico (colonizadas vs. no colonizadas), la mayoría involucradas en el proceso
de fotosíntesis. Estas enzimas son: fructosa bifosfato aldolasa isoenzima citoplásmica (spot
5503), triosa fosfato isomerasa (spot 6201), ribulosa bifosfato carboxilasa subunidad grande
(spot 7801), ferredoxina-NADP reductasa isoenzima de hoja cloroplástica (spot 6404), ATP
sintasa CF1 subunidad alfa (spot 2802) y ATP sintasa subunidad B unida a membrana (spot
3801).
Nuestros resultados se podrían correlacionar con varios reportes que demuestran
que la fotosíntesis se incrementa en plantas en diferentes asociaciones simbióticas: Glycine
max colonizado con G. mosseae (Brown y Bethlenfalvay, 1988), tomate colonizado con G.
fasciculatum (Sanchez-Rocha et al., 2005), Citrus aurantium asociado con G. intraradices
(Johnson, 1984) y C. tangerine colonizado con G. versiforme (Wu and Xia, 2006).
A nivel proteómico en parte aérea, nuestros resultados coinciden con los obtenidos
por Shoresh y Harman (2008). Ellos encontraron una gran cantidad de proteínas inducidas
involucradas en el metabolismo de carbono en la asociación simbiótica de plántulas de
maíz colonizadas con el hongo T. harzianum. Algunas de las proteínas reguladas coinciden
con nuestros resultados, estas incluyen: fructosa bifosfato aldolasa (spot 5503) y la
subunidad grande de la ribulosa 1,5-bifosfato carboxilasa/oxigenasa (spot 7801).
Trichoderma harzianum al igual que G. intraradices es un agente de biocontrol y en ambos
sistemas su efecto en parte aérea de plantas, ocasiona cambios en el metabolismo de
carbono, lo cual probablemente prepara a la planta para la producción de subsecuentes
elicitores.
A nivel de genes en parte aérea, nuestros resultados también coinciden con los de
Liu et al. (2007). El patrón de expresión de transcriptos en hojas de M. truncatula
colonizadas con G. intraradices, mostró la sobreexpresión de genes que codifican para
proteínas relacionadas a fotosíntesis tales como ATP sintasa (ATPasa subunit IV), y la
subunidad pequeña de la ribulosa bifosfato carboxilasa. Estos resultados son similares en
plantas de Az. Regional 87 (Spots: 2802, 3801 y 7801).

Quintero-Zamora, 2010 65
El ciclo de calvin es un ciclo llevado a cabo por organismos eucarioticos
fotosintéticos con la finalidad de reducir el dióxido de carbono a carbohidratos.
Transcetolasas participan en varias reacciones del ciclo de calvin. En base a esto se podría
sugerir que la fotosíntesis aumenta en condiciones de estrés/defensa de la planta a través de
la sobreexpresión de transcetolasas. Esto podría jugar un papel en la inducción de la
tolerancia a patógenos foliares por micorrización. Por lo que la sobreexpresión de enzimas
involucradas en fotosíntesis en la planta como transcetolasas, puede ser interpretada como
una respuesta de defensa de la misma al ataque por un patógeno.
Un efecto contrario se presenta en otros reportes como el de Fiorilli et al. (2009)
llevado a cabo en hojas de tomate colonizadas por el HMA G. mosseae y el trabajo de
Peinado-Guevara (2010, en revisión) llevado a cabo en hojas de tomate colonizadas con G.
intraradices, donde la mayoría de los genes y proteínas relacionadas a fotosíntesis
disminuyen su expresión. Galindo-Flores (2008), reporta una inducción de tolerancia en
hojas de plantas de tomate colonizadas con G. intraradices y retadas con el patógeno X.
campestris. Se ha demostrado en muchas interacciones planta-microorganismo que la
respuesta a una condición es dependiente del sistema involucrado. Aquí ambos sistemas
consisten en la asociación de un HMA con plantas de tomate y el estudio se llevó a cabo en
parte aérea. Probablemente esta asociación, requiera de la disminución de fotosíntesis como
un mecanismo para preparar a la planta contra diferentes tipos de estrés.
Un transcrito correspondiente a la enzima biosintética de tiamina fue reportada
disminuida en su expresión para M. truncatula por Liu et al. (2007). De manera interesante
se observó que en frijol Az. Regional 87 una proteína correspondiente a una enzima
biosintética de tiamina también disminuye su expresión (spot 2401). La tiamina funciona
como precursor de la tiamina difosfato la cual sirve como coenzima de enzimas
involucradas en varias rutas metabólicas como el ciclo del ácido tricarboxilico, ciclo de
Calvin, y en la biosíntesis de pigmentos en plantas (Friedrich, 1987). Además,
recientemente se ha reportado que tiene funciones como cofactor en respuesta a estrés
biótico y abiótico en plantas.
Rapala-Kozik et al. (2008), reportaron un aumento en el contenido de tiamina, en
hojas de plántulas de maíz bajo condiciones de estrés abiótico (sequía, salinidad y estrés
oxidativo). Además de que en este estudio también reportan un incremento en

Quintero-Zamora, 2010 66
transcetolasas (proteína ya discutida anteriormente) bajo condiciones de salinidad y estrés
oxidativo.
Tunc-Ozdemir et al. (2009), demostraron la acumulación de tiamina y pirofosfato
de tiamina en hojas de plantas de A. thaliana bajo condiciones de estrés oxidativo como
salinidad, frío, elevada intensidad de luz y tratamientos oxidativos. La acumulación de
tiamina y pirofosfato de tiamina fue acompañado con la expresión de transcriptos que
codifican para enzimas biosintéticas de tiamina. Las investigaciones muestran un aumento
en actividad de la enzima de la biosíntesis de tiamina en plantas bajo la condición de estrés
abiótico. Sin embargo, en nuestro trabajo y en el de Liu et al. (2007) se observa un efecto
contrario. La proteína enzima biosintética de tiamina disminuye su expresión en plantas
colonizadas con HMA. Análisis proteómicos en parte aérea de plantas de Az. Regional 87
colonizada con el hongo y retada con el patógeno podrían llevarse a cabo, para observar el
efecto del patógeno en la expresión de estas proteinas.
El trabajo de Liu et al. (2007) es el único trabajo de expresión diferencial de genes
llevado a cabo en parte aérea de plantas leguminosas colonizadas con un HMA. Los autores
reportaron un incremento a la tolerancia al patógeno foliar X. campestris en parte aérea de
plantas micorrizadas con G. intraradices, fenómeno similar a lo observado por Mora-
Romero (2008) en parte aérea de la variedad Az. Regional 87. En el trabajo de Liu et al.
(2007) se identificaron un número elevado genes diferenciales, aunque el número de
proteínas diferenciales encontradas en este trabajo corresponden a un número mucho
menor. Esto refleja las diferencias técnicas de las dos metodologías. Además, la proteómica
de plantas cuando está enfocada a evaluar tejido completo, presenta la dificultad que la
proteína Rubisco es la mayoritaria, haciendo difícil la detección de proteínas menos
abundantes. Estos resultados también abren la necesidad de llevar a cabo esfuerzos para
realizar subproteomas, buscando estrategias para la optimización del método de extracción
y el análisis bidimensional, con lo cual se podrían analizar distintos grupos de proteínas.
Se encontró sobreexpresada en la variedad Az. Regional 87 en condiciones de
micorrización una proteína de función desconocida (spot 2302). Esta proteína no se
encontró diferencialmente regulada en la variedad Az. Higuera. Experimentos posteriores,
resultan necesarios para establecer la función de esta proteína.

Quintero-Zamora, 2010 67
La inducción de una proteína identificada como tipo alfa faseolina (Cuadro 7: spot
3701), detectada solamente en la parte aérea de la variedad tolerante en la condición
micorrizada, podría estar directamente relacionada con la simbiosis de la planta con G.
intraradices y podría jugar un papel importante en la inducción de tolerancia por
micorrización. Es interesante la presencia de la alfa faseolina, ya que ésta pertenece a la
familia de las globulinas 7S, las cuales son proteínas de almacenamiento en grano, lo que
nos indica las repercusiones que pueden estarse presentando en las plantas colonizadas por
HMA a nivel de expresión génica. Por esto, trabajos posteriores deberán realizarse para
conocer la función que desempeña esta proteína en parte aérea en plantas micorrizadas.
En el presente estudio se encontró un perfil de proteínas diferenciales contrastantes
en hojas de plantas colonizadas con G. intraradices de la variedad Az. Regional 87, la cual
mostró inducción de tolerancia al patógeno S. sclerotiorum y Az. Higuera, la cual fue
incapaz de inducir tal tolerancia. Esto podría sugerir que al menos algunas de las proteínas
encontradas como reguladas de manera diferencial están participando en un mecanismo de
bioprotección que enciende la MA en la variedad Az. Regional 87 y que la preparan para
enfrentar a condiciones de estrés posteriores. Lo que implica que el mecanismo involucrado
es dependiente de la variedad, ya que en Az. Higuera se presenta un efecto contrario en el
patrón de expresión de proteínas.

Quintero-Zamora, 2010 68
VIII. CONCLUSIONES
El método de extracción de proteínas más adecuado, en hojas de frijol para
electroforesis bidimensional, involucra la precipitación con TCA/Acetona.
La proteómica comparativa es una herramienta que nos permitió la identificación de
proteínas que pudieran estar involucradas en la resistencia que se induce por
micorrización.
Las proteinas diferenciales transcetolasa y tripsinógeno, se encontraron reguladas de
manera distinta en hojas de plantas colonizadas con G. intraradices. En la variedad
Az. Regional 87 (la cual mostró inducción de tolerancia al patógeno S.
sclerotiorum) aumentaron su expresión y Az. Higuera (la cual fue incapaz de
inducir tal tolerancia) disminuyeron su expresión. Además, la proteína alfa faseolina
fue encontrada solo en la variedad Az. Regional 87 colonizada con G. intraradices.
Lo cual sugiere que estas proteínas podrían jugar un papel importante en la
inducción de la tolerancia a patógenos foliares por micorrización.
La mayoría de las proteínas inducidas en Az. Regional 87 corresponden a enzimas
implicadas en fotosíntesis: transcetolasa, fructosa bifosfato aldolasa isoenzima
citoplásmica, triosa fosfato isomerasa, ribulosa bifosfato carboxilasa subunidad
grande, ferredoxina-NADP reductasa isoenzima de hoja cloroplástica, ATP sintasa
CF1 subunidad alfa y ATP sintasa subunidad B unida a membrana. Por lo que se
sugiere que la fotosíntesis está siendo inducida por la MA en condiciones de
estrés/defensa de la variedad Az. Regional 87 colonizada con G. intraradices. Lo
cual puede ser interpretado como una respuesta de defensa de la variedad Az.
Regional 87 colonizada con G. intraradices al ataque por un patógeno.

Quintero-Zamora, 2010 69
Al menos algunas de las proteínas encontradas como reguladas de manera
diferencial están participando en un mecanismo de bioprotección que enciende la
MA en la variedad Az. Regional 87 y que la preparan para enfrentar a condiciones
de estrés posteriores. Lo que implica que el mecanismo involucrado es dependiente
de la variedad, ya que en Az. Higuera se presenta un efecto contrario en los patrones
de acumulación de algunas proteínas analizadas.
Este es el primer trabajo realizado con proteómica en la parte aérea de plantas de
frijol en donde se hace el estudio comparativo de proteínas que se inducen por
micorrización.

Quintero-Zamora, 2010 70
IX. RECOMENDACIONES
• Primeramente se sugiere fortalecer la infraestructura en cuanto al equipo y material
utilizado en en análisis proteómico.
• Perfeccionar aún más el método de extracción de proteínas mediante TCA-A, para
lograr un patrón más alto de proteínas. Ya que el análisis del proteoma es el
resultado de una combinación una serie de técnicas y posiblemente estemos
perdiendo una gran cantidad de proteínas.
• Generar oligonucleótidos para el gen que codifica para la proteína cualitativa alfa
faseolina y llevar a cabo experimentos en plantas colonizadas y no colonizadas y
además retadas con el patógeno y asi determinar si el gen es específico de
resistencia inducida por micorrización e interpretar su papel en la inducción de
tolerancia.
• Se recomienda realizar experimentos llevando a cabo el silenciamiento o
sobreexpresión de genes que codifican para las proteínas diferenciales encontradas.
Especialmente de las proteínas encontradas de manera constrastante en las dos
variedades, tripsinógeno y transcetolasa, así como la proteína cualitativa, alfa
faseolina. Esto mediante la tecnología de transgénicos y retando con el patógeno
para comprobar su función en la defensa sistémica de la planta.

Quintero-Zamora, 2010 71
X. BIBLIOGRAFÍA
• Adessi C., Miege C., Albrieux C. y Rabilloud T. (1997). Two-dimensional
electrophoresis of membrane proteins: a current challenge for immobilized
pH gradients. Electrophoresis 18:127-135.
• Annamraju D. Sarma, Nathan W. Oehrle y David W. Emerich (2008). Plant protein
isolation and stabilization for enhanced resolution of two-dimensional
polyacrylamide gel electrophoresis. Analytical Biochemistry 379: 192-195.
• Attaran E., Zeier T. E., Griebel T. y Zeier J. (2009). Systemic acquired resistance
inArabidopsis is independent of methyl salicylate production and jasmonate
signaling. Plant Cell 21: 954-971.
• Azcón-Aguilar C. y Barea J. M. (1992). Interactions between mycorrhizal fungi and
other rhizosphere microorganisms. En Allen MF (Ed.) Mycorrhizal
Functioning. An Integrative Plant Fungal Process. Chapman & Hall. New
York, EEUU. Pp 163-198.
• Azcón-Aguilar C. y Barea J. M. (1996). Arbuscular mycorrrhizas and biological
control of soil-borne plant pathogens-an overview of the mechanisms
involved. Mycorrhiza 6:457-464.
• Beckers, G. J. M. y U. Conrath. (2007). Priming for stress resistance: from the lab
to the field. Plant Biology 10:425-431.
• Benabdellah, K., C. Azcón-Aguilar y N. Ferrol. (2000). Alterations in the plasma
membrane polypeptide pattern of tomato roots (Lycopersicum esculentum)
during the development of arbuscular mycorrhiza. Journal of Experimental
Botany 51:747-754.
• Bernier, F. y Berna, A. (2001). Germins and germin-like proteins: plant do-all
proteins. But what do they do exactly? Plant Physiolology and Biochemistry.
39:545-554.
• Bestel-Corre G., Dumas-Gaudot E. y Gianinazzi S. (2004). Proteomics as a tool to
monitor plant-microbe endosymbioses in the rhizosphere. Mycorrhiza 14:1-
10.

Quintero-Zamora, 2010 72
• Bestel-Corre G., Dumas-Gaudot E., Poinsot V., Dieu M., Dierick J.F., Van Tuinen
D., Remacle J., Gianinazzi-Pearson V. y Gianinazzi S. (2002). Proteome
analysis and identification of symbiosis-related proteins from Medicago
truncatula Gaertn by two-dimensional electrophoresis and mass
spectrometry. Electrophoresis 23:122-137.
• Black K. E. (1997). Root longevity as affected by biotic and abiotic factors, PhD
Thesis, University of Aberdeen.
• Bødker L., Kjoller R. Kristensen K. y Rosendahl S. (2002). Interactions between
indigenous arbuscular mycorrhizal fungi and Aphanomyces euteiches in
field-grown pea. Mycorrhiza 12:7-12.
• Bødker L., Kjoller R. y Rosendahl S. (1998). Effect of phosphate and the arbuscular
mycorrhizal fungus Glomus intraradices on disease severity of root rot of
peas (Pisum sativum) caused by Aphanomyces euteiches. Mycorrhiza 8:169–
174.
• Bonfig K. B., Schreiber U., Gabler A., Roitsch T. y Berger S. (2006). Infection with
virulent and avirulent P. syringae strains differentially affects photosynthesis
and sink metabolism in Arabidopsis leaves. Planta 225: 1-12.
• Bonfante P. (2003). Plants, Mycorrhyzal Fungi and Endobacteria: a Dialog Among
Cells and genomes. Biology Bulletin 204:215-220.
• Bouwmeester H.J., Roux C., Lopez-Raez J.A. y Becard G. (2007). Rhizosphere
communication of plants, parasitic plants and AM fungi. Trends in Plant
Science 12:224-230.
• Broughton W. J., Hernández G., Blair M., Beebe S., Gepts P. y Vanderleyden J.
(2003). Beans (Phaseolus spp.) – model food legumes. Plant and Soil 252:
55-128.
• Brown M. S. y Bethlenfalvay G. J. (1988). The Glycine-Glomus- Rhizobium
symbiosis. VII. Photosynthetic nutrient use efficiency in nodulated,
mycorrhizal soybeans. Plant Physiology. 86:1292-1297.
• Brundrett M., Bougher N., Dell B., Grove T. y Malajczuc N. (1996). Working with
Micorrizas in Forestry and Agriculture. ACIAR Monograph. Camberra.

Quintero-Zamora, 2010 73
• Cascardo J. C., Buzeli R. A., Almeida R. S., Otoni W. C. y Fontes E.P. (2001).
Differential expression of the soybean BiP gene family. Plant Science
160:273-281.
• Castillejo M. A., Amiour N., Dumas-Gaudot E., Rubiales D. y Jorrín J.V. (2004). A
proteomic approach to studying plant response tocreanate broomrape
(Orobanche crenata) in pea (Pisum sativum). Phytochemistry 65:1817–
1828.
• Chen S. y Harmon A.C. (2006). Advances in plant proteomics. Proteomics 6:1-13.
• Chinnasamy G. (2005). A proteomics perspective on biocontrol and plant defense
mechanism. PGPR: Biocontrol and Biofertilization. Pp 233-255.
• CIAT (1993). Trends in CIAT commodities. Working Doc. No 128. Cali, Colombia
• Clark R. B. y Zeto S.K. (2000). Mineral acquisition by arbuscular mycorrhizal
plants. Journal of Plant Nutrition 23:867-902.
• Colditz F., Braun H.P., Jacquet Ch., Niehaus K. y Krajinski F. (2005). Proteomic
profiling unravels insights into the molecular background underlying
increased Aphanomyces euteiches – tolerances of Medicago truncatula.
Plant Molecular Biology 59:387-406.
• Colditz F., Nyamsuren O., Niehaus K., Eubel H., Braun H.P. y Krajinski, F. (2004).
Proteomic approach: Identification of Medicago truncatula proteins induced
in roots after infection with the pathogenic oomycete Aphanomyces
euteiches. Plant Molecular Biology 55:109-120.
• Cordier C., Pozo M. J., Gianinazzi S. y Gianinazzi-Pearson V. (1998). Cell defence
responses associated with localised and systemic resistance to Phytophthora
parasitica induced in tomato by an arbuscular mycorrhizal fungus.
Molecular Plant-Microbe Interactions 11:1017-1028.
• Cui J., Bahrami A. K., Pringle E. G., Hernandez-Guzman G., Bender C. L. Pierce,
N. E. y Ausubel, F. M. (2005). Pseudomonas syringae manipulates systemic
plant defenses against pathogens and herbivores. Proceedings of the National
Academy of Sciences of the United States of America. 102, 1791-1796.
• Damerval C., De Vienne D., Zivy M. y Thiellement H. (1986). The technical
improvements in two dimensional electrophoresis increase the level of

Quintero-Zamora, 2010 74
genetic variation detected in wheat seedling proteins. Electrophoresis 7:52–
54.
• Deguchi Y., Banba M., Shimoda Y., Chechetka S. A., Suzuri R., Okusako Y., Ooki
Y., Toyokura K., Suzuki A., Uchiumi T., Higashi S., Abe M., Kouchi H.,
Izui K. y Hata S. (2007). Transcriptome profiling of Lotus japonicas Roots
During Arbuscular Mycorrhiza Development and Comparison with that of
Nodulation. DNA Research 14:117-133.
• De la Noval B., Pérez E., Martínez B., León O., Martínez-Gallardo N. y Délano-
Frier J. (2007). Exogenous systemin has a contrasting effect on disease
resistance in mycorrhizal tomato (Solanum lycopersicum) plants infected
with necrotrophic or hemibiotrophic pathogens. Mycorrhiza 17:449-460.
• Dodd J. C., Burton C. C., Burns R. G. y Jeffries P. (1987). Phosphatase activity
associated with the roots and the rhizosphere of plants infected with
vesicular-arbuscular mycorrhizal fungi. New Phytologist 107:163-172.
• Dong X. (2004). NPR1, all things considered. Current Opinión in Plant Biology 7:
547-552.
• Dumas-Gaudot E., Gollotte A., Cordier C., Gianinazzi S. y Gianinazzi-Pearson V.
(2000). Modulation of host defence systems. Arbuscular Mycorrhizas:
Physiology and Function. (Y. Kapulnik & D.D. Douds Jr. eds.). Kluwer
Academic Publishers. Pp.173-200.
• Dumas-Gaudot E., Valot B., Bestel-Corre G., Recorbet G., St-Arnaud M., Fontaine
B., Dieu M., Raes M., Saravanan R.S. y Gianinazzi S. (2004). Proteomics as
a way to identify extra-radicular fangal proteins from Glomus intraradices –
RiT-DNA carrot root mycorrhizas. FEMS Microbiology Ecology 48:401-
411.
• Elsen A., Declerck S. y De Waele D. (2003). Use of root organ cultures to
investigate the interactions between Glomus intraradices and Pratylenchus
coffeae. Applied Enviromental Microbiology 69(7):4308-4311.
• Elsen A., Gervacio D., Swennen R. y De Waele D. (2008). AMF-induced biocontrol
against plant parasitic nematodes in Musa sp.: a systemic effect. Mycorrhiza
18:251-256.

Quintero-Zamora, 2010 75
• FAO (2008). Food and Agriculture Organization. http:/www.fao.org. Base de datos
estadísticos.
• Finnie C., Melchior S., Roepstorff P. y Svensson B. (2002). Proteome analysis of
grain Wlling and seed maturation in barley. Plant Physiology 129:1308-
1319.
• Fiorilli V., Catoni M., Miozzi L., Novero M., Accotto G. P. y Lanfranco L. (2009).
Global and cell-type gene expression profiles in tomato plants colonized by
an arbuscular mycorrhizal fungus. New phytologist 184: 975-987.
• Fontana F., Santini A., Salvini M., Pazzagli L., Cappugi G., Scala A., Durante M. y
Bernardi R. (2008). Cerato-platanin treated plane leaves restrict Ceratocystis
platani growth and overexpress defence-related genes. Journal of Plant
Pathology 90 (2): 295-306.
• Friedrich W. (1987). Thiamin (Vitamin B1, aneurin). In: Handbuch der Vitamine.
Munich: Urban and Schwartzenberg, 240-258.
• Fritz, M., I. Jakobsen, M.F. Lyngkjaer, H. Thordal-Christensen y J. Pons-
Kuhnemann (2006). Arbuscular mycorrhiza reduces susceptibility of tomato
to Alternaria solani. Mycorrhiza 16:413-419.
• Galindo-Flores H. (2008). Resistencia inducida por micorrización en tomate
(Lycopersicon esculentum Mill) ante Xanthomonas campestris pv.
Vesicatoria. Tesis de maestría en Recursos Naturales y Medio Ambiente.
CIIDIR-IPN U. Sinaloa.
• Gamalero E., Trotta A., Massa N., Copetta A., Martinotti M.G. y Berta G. (2004).
Impact of two fluorescent pseudomonads and an arbuscular mycorrhizal
fungus on tomato plant growth, root architecture and P acquisition.
Mycorrhiza 14:185-192.
• Gao L., Knogge O., Delp G., Smith F. A. y Smith S. E. (2004). Expression Patterns
of Defense-Related Genesin Different Typesof Arbuscular Mycorrhizal
Developmenin Wild-Type and Mycorrhiza-Defective Mutant Tomato.
Molecular Plant-Microbe Interactions 17:1103-1113.
• Godfrey D., Able A. J. y Dry I. B. (2007). Induction of a grapevine germin-like
protein (VvGLP3) gene is closely linked to the site of Erysiphe necator

Quintero-Zamora, 2010 76
infection: a possible role in defense? Molecular Plant-Microbe Interactions.
20(9):1112-1125.
• Goellner K. y Conrath U. (2008) Priming: it’s all the world to induced disease
resistance. European Journal of Plant Pathology 121: 233–242.
• Granier F. (1988). Extraction of plant proteins for two-dimensional electrophoresis.
Electrophoresis 9:712-718.
• Gucciardo S., Wisniewski J-P., Brewin N. J. y Bornemann S. (2007). A germin-like
protein with superoxide dismutase activity in pea nodules with high protein
sequence identity to a putative rhicadhesin receptor. Journal of Experimental
Botany 58(5): 1161-1171.
• Harrier L. A. y Watson C. A. (2004). The potential role of arbuscular mycorrhizal
(AM) fungi in the bioprotection of plants against soil-borne pathogens in
organic and/or other sustainable farming systems. Pest Manager Science
60:149-157
• Harrison M. J. (2005). Signaling in the arbuscular mycorrhizal symbiosis. Annual
Review of Microbiology 59:19-42.
• Hause, B., C. Mrosk, S. Isayenkov y D. Strack. (2007). Jasmonates in arbuscular
mycorrhizal interactions. Phytochemistry 68:101-110.
• Hause B. y Faster T. (2005). Molecular and cell biology of arbuscular mycorrhizal
symbiosis. Planta 221:184-196.
• Hause, B., W. Maier, O. Miersch, R. Kramelland y D. Strack. (2002). Induction of
jasmonate biosynthesis in arbuscular mycorrhizal barley roots. Plant
Physiology 130: 1213-1220.
• Hause B. y Faster T. (2005). Molecular and cell biology of arbuscular mycorrhizal
symbiosis. Planta 221:184-196.
• Hodge A., Robinson D. y Fitter A.H. (2000). An arbuscular mycorrhizal inoculum
enhances root proliferation in, but not nitrogen capture from, nutrient-rich
patches in soil. New Phytology 145:575-584.
• Hossain M. M., Sultana F., Kubota M. y Hyakumachi M. (2008). Differential
inducible defense mechanisms against bacterial speck pathogen in

Quintero-Zamora, 2010 77
Arabidopsis thaliana by plant-growth-promoting-fungus Penicillium sp.
GP16-2 and its cell free filtrate. Plant Soil 304: 227-239.
• Hurkman W. J. y Tanaka C. K. (1986). Solubilization of plant membrane proteins
for analysis by two-dimensional gel electroforesis. Plant Physiology 81:802-
806.
• Joner E.J. y Johansen A. (2000). Phosphatase activity of external hyphae of two
arbuscular mycorrhizal fungi. Mycological Research 104:81-86.
• Leigh J. Hodge A. y Fitter A. H. (2009). Arbuscular mycorrhizal fungi can transfer
substancial amounts of nitrogen to their host plant from organic material.
New phytology 181(1): 199-207.
• Lingua, G., G. D´Agostino, N. Massa, M. Antosiano y G. Berta. (2002).
Mycorrhiza-induced differential response to a yellows disease in tomato.
Mycorrhiza 12:191-198.
• Liu J., Maldonado-Mendoza I., López-Meyer M., Cheung F., Town C.D. y Harrison
M.J. (2007). Arbuscular mycorrhizal symbiosis is accompanied by local and
systemic alterations in gene expression and an increase in disease resistance
in the shoots. The Plant Journal 50:529-544.
• Liu, J., L. A. Blaylock, G. Endre, J. Cho, C. D. Town, K. A. VandenBosch, y M.
J. Harrison. (2003). Transcript profiling coupled with spatial expression
analyses symbiosis. Plant cell 15: 2106-2123.
• Loake G. y Grant M. (2007). Salicylic acid in plant defence-the players and
protagonists. Current Opinion in Plant Biology 10: 466-472.
• Ludwig-Mülle, J. y Güther M. (2007). Auxins as Signals in Arbuscular Mycorrhiza
Formation. Plant Signal & Behavior 2(3):194-196.
• Maldonado-Mendoza I. E., Dewbre G. R. y Harrison M. J. (2001). A Phosphate
Transporter Gene from the Extra-Radical Mycelium of an Arbuscular
Mycorrhizal Fungus Glomus intraradices Is Regulated in Response to
Phosphate in the Environment. Molecular Plant-Microbe Interactions 14 (10)
pp. 1140-1148.
• Marschner P., Crowley D. E. y Higashi R. M. (1997). Root exudation an
physiological status of a root colonising fluorescent Pseudomonad in

Quintero-Zamora, 2010 78
mycorrhizal and non-mycorrhizal pepper (Capsicum annuum L). Plant Soil
189:11-20.
• McLafferty (1993). Interpretation of mass spectra. USB Mill Valley. 37:118-120.
• Memelink J. (2009). Regulation of gene expression by jasmonate hormones.
Phytochemistry 70: 1560-1570.
• Meyer Y., Grosser J., Chartier Y. y Cleyet-Maurel J. C. C. (1998). Electrophoresis
9: 704-712.
• Miklas N. P., Kelly J. D., Beebe S. E. y Blair M.W. (2006). Common bean
breeding for resistance against biotic and abiotic stresses: From classical to
MAS breeding. Euphytica 147: 105-131.
• Miller R.M. y Jastrow J.D. (2000). Mycorrhizal fungi influence soil structure.
Arbuscular Mycorrhizas: Physiology and Function. Eds: Y Kapulnick and
DD Douds Jr. Kluwer Academic Press. Pp. 3-18.
• Mohr U., Lange J., Boller T., Wiemken y Vögeli-Lange R. (1998). Plant defence
genes are induced in the pathogenic interaction between bean roots and
Fusarium sonali, but not in the symbiotic interaction with the arbuscular
mycorrhiza fungus Glomus mosseae. New Phytology 138:589-598.
• Mora-Romero G. A., (2008). Efecto de la micorrización con Glomus intraradices
en la tolerancia al moho blanco causado por Sclerotinia sclerotiorum en
tres líneas de frijol. Tesis de maestría en Recursos Naturales y Medio
Ambiente. CIIDIR-IPN U. Sinaloa.
• Munyanziza E., Kehri H. K. y Bagyaraj D. J. (1997). Agricultural intensification,
soil biodiversity and agro-ecosystem function in the tropics: the role of
mycorrhiza in crops and trees. Applied Soil Ecology 6:77-85.
• Natarajan S., Xu C., Caperna T.J. y Garrett W.M. (2005). Comparison of protein
solubilization methods suitable for proteomic analysis of soybean seed
proteins. Analytical Biochemistry 342:214–220.
• Newsham K. K., Fitter A. H. y Watkinson A. R. (1995). Arbuscular mycorrhiza
protects an annual grass from root pathogenic fungi in the field. Journal of
Ecology 83:991-1000.

Quintero-Zamora, 2010 79
• Norman J. R., Atkinson D. y Hooker J. E. (1996). Arbuscular mycorrhizal fungal-
induced alteration to root architecture in strawberry and induced resistance
to the root pathogen Phytophthora fragariae. Plant Soil 185:191–198.
• Pachico D. (1989). Tendencias en la producción mundial de frijol común. In:
Pastor-Corrales, M.A. & Schwartz, H.F. (Eds.). Problemas de producción del
frijol en los trópicos. CIAT. Cali, Colombia. pp.1-10.
• Park C. J., An J. M., Shin Y. C., Kim K. J., Lee B. J. y Pack K. H. (2004).
Molecular characterization of pepper germin-like protein as the novel PR-16
family of pathogenesis-related proteins isolated during the resistance
response to viral and bacterial infection. Planta 219:797-806.
• Pearson J. N. y Jakobsen I. (1993). The relative contribution of hyphae and roots to
phosphorus uptake by arbuscular mycorrizal plants, measured by dual
labeling with 32P and 33P. New Phytology 124:489-494.
• Peterson R. L., Massicotte H. B. y Melville L.H. (2004). Mycorrhizas: Anatomy
and Cell Biology. NRC Research Press. Ottawa Ont.
• Pieterse C. M. J. y Van Loon L. C. (2004). NPR1: the spider in the web of induced
resistance signaling pathways. Current Opinión in Plant Biology 7: 456–464.
• Pieterse C. M. J., Van Wees S. C. M., Van Pelt J. A., Knoester M., Laan R., Gerrits
H., Weisbeek P.J. y Van Loon L. C. (1998). A novel signaling pathway
controllinginduced systemic resistance in Arabidopsis. Plant Cell 10: 1571-
1580.
• Pieterse C. M., Van Wees S. C., Hoffland E. y Van Loon L.C. (1996). Systemic
resistance in Arabidopsis induced by biocontrol bacteria independentof
salicylic acid accumulations and pathogenesis-related gene expression. Plant
Cell 8:1225-1237.
• Pozo M. J. y Azcón–Aguilar C. (2007). Unraveling mycorrhiza-induced resistance.
Current Opinión in Plant Biology 10:393-398.
• Pozo, M. J., C. Cordier, E. Dumas-Gaudot, S. Gianinazzi, J. M. Barea, y C. Azcón-
Aguilar. (2002). Localized versus systemic effect of arbuscular mycorrhiizal
fungi on defense responses to Phytophthora infection in tomato plants.
Journal of Experimental Botany 53(368): 525-534.

Quintero-Zamora, 2010 80
• Pridmore A. M., Devine D. A., Bonass W. A. y Silley P. (1999). Influence of
sample preparation technique on two dimensional gel electrophoresis of
proteins from Porphyromonas ingivalis. Lett in Applied Microbiology
28:245-249.
• Quintana H., Gómez C., Díaz J. y Camarena F. (2000). Evaluación de la calidad de
la Proteína de 4 Variedades de Frijol. Rev. Cubana Alimentaria Nutricional
14:22-27.
• Rapala-Kozik M., Kowalska E. y Ostrowska K. (2008). Modulation of thiamine
meabolism in Zea mays seedlings under conditions of abiotic stress. Journal
of Experimental Botany 59 (15): 4133-4143.
• Rillig M. (2004). Arbuscular mycorrhizae, glomalin, and soil aggregation.
Canadian Journal of Soil Sciencie 84: 355-363.
• Rodríguez R. (1996). Problemas, logros y posibilidades para el mejoramiento del
frijol negro. In: Taller de mejoramiento de frijol para el siglo XXI: Bases
para una estrategia para América Latina. Ed. por Singh, S.P. y Voysest, O.
Cali, Colombia, CIAT. Pp. 294-306.
• Rosales-Serna R., Acosta-Gallegos J. A., Durán-Durán R. P., Guillén-Andrade H.,
Pérez-Herrera P., Esquivel-Esquivel J., y Muruaga-Martíınez J. S. (2003).
Diversidad genética del germoplasma mejorado de frijol (Phaseolus vulgaris
L.) en México. Agric. Tec. Mexico 29:11-24.
• Rosales-Serna R., Acosta-Gallegos J.A., Muruaga-Martínez J.S., Hernández-
Casillas J.M., Esquivel-Esquivel G. y Pérez-Herrera P. (2004). Variedades
mejoradas de frijol del Instituto Nacional de Investigaciones Forestales,
Agrícolas y Pecuarias. (INIFAP). Libro Técnico No. 6. INIFAP-SAGARPA,
México.
• Rosales-Serna R., Hernández-Delgado S., González-Paz M., Acosta-Gallegos J.A.,
y Mayek-Pérez N. (2005). Genetic relationships and diversity revealed by
aflp markers in Mexican common bean bred cultivars. Crop Science
45:1951-1957.

Quintero-Zamora, 2010 81
• Rose J.K.C., Bashir S., Giovannoni, J.J., Jahn M.M. y Saravanan S. (2004).
Tackling the plant proteome: practical approaches, hardles and experimental
tool. The Plant Journal 39:715-733.
• Ruiz-Lozano J.M. y Azcón R. (1995). Hyphal cotribution to water uptake in
mycorrhizal plants as affected by the fungal species and water status.
Physiologia Plantarum 95:472-478.
• SAGARPA (2009). Secretaría de Agricultura Ganadería, Desarrollo Rural, Pesca y
Alimentación. http://www.sagarpa.gob.mx/
• Salinas-Pérez R.A., Hernández-Fernández A., Navarro-Sandoval F.J. y Rodríguez-
Cota, F.G. (1989). Azufrado peruano y azufrado regional 87 nuevas
variedades de frijol para el estado de Sinaloa. SARH. Folleto técnico No 5.
Octubre de de 1989.
• Salinas-Pérez R.A., Navarro-Sandoval F.J. y Rodríguez-Cota F.G. (1995).
Azufrado Noroeste y Azufrado Higuera Nuevas variedades de frijol para
Sinaloa. INIFAP-CIRNO-CEVAF. Folleto Técnico No. 11. Noviembre de
1995.
• Salinas-Pérez R.A. y Rodríguez-Cota F.G. (2008). Impacto del mejoramiento
genético del frijol en Sinaloa: variedad “azufrado higuera”. INIFAP-
CIRNO-CEVAF. (Trabajo que se presentó para concurso del premio tecno
agro 2008).
• Salzer P. y Boller T. (2000). Elicitor induced reactions in mycorrhizae and their
supresión. En. Arbuscular Mycorrhizas: Physiology and Function. (Y. Kap
ulnik y D.D. Douds Jr. eds.). Kluwer Academic Publishers. Pp : 1-10.
• Samra A., Dumas-Gaudot E. y Gianinazzi S. (1997). Detection of symbiosis-related
polypeptides during the early stages of the establishment of arbuscular
mycorrhiza between Glomus mosseae and Pisum sativum roots. New
Phytologist 135:711-722.
• Sanchez-Rocha S., Vargas-Luna M., Gutiérrez-Juárez G., Huerta Franco, R.,
Madueño L. y Olalde-Portugal V. (2005). Oxygen evolution from tomato
(C3) plants with and without mycorrhiza: Open photoacoustic cell

Quintero-Zamora, 2010 82
measurement and statistical analysis. Journal Physiology IV France.
125:803-806.
• Santoni, V., Bellini C. y Caboche M. (1994). Use of two-dimensional protein
pattern analysis for the characterization of Arabidopsis thaliana mutants.
Planta 192:557–566.
• Saravanan R. S. y Rose J. K. C. (2004). A critical evaluation of sample extraction
techniques for enhanced proteomic analysis of recalcitrant plant tissues.
Proteomics 4:2522–2532.
• Sarma A. D., Oehrle N. W. y Emerich D. W. (2008). Plant protein isolation and
stabilization for enhanced resolution of two-dimensional polyacrylamide gel
electrophoresis. Analytical Biochemistry 397: 192-195.
• Schenck N.C., y G.S. Smith, (1982). Additional new and unreported species of
micorrhizal fungi (Endogonaceae) from Florida. Mycologia 74:77-92.
• Scheublin T.R., Ridway K.P., Young J.P.W. y Van der Heiden M.G.A. (2004).
Nonegumes, Legumes, and Root Nodules Harbor Diferrent Arbuscular
Mycorrhizal Fungal Communities. Applied and Environmental
Microbiology 70:6240-6246.
• Schoonhoven A. y Van Voysest O. (1989). Common beans in Latin America and
their constraints. In: CIAT. Bean Production Problems in the Tropics. Cali,
Colombia. Pp.33-58.
• Schuessler A., Schwarzott D. y Walker C. (2001). A new fungal phylum, the
Glomeromycota: phylogeny and evolution. Mycological Research 105 (12):
1413-1421.
• Segarra G., Van der Ent S., Trillas I. y Pieterse C. M. J. (2009). MYB72, a node
ofconvergence in induced systemic resistance triggered by a fungal and
abacterial beneficial microbe. Plant Biology 11: 90-96.
• Shoresh M. y Harman G. E. (2008). The molecular basis of shoot responses ofmaize
seedlings to Trichoderma harzianum T22 Inoculaction of the root: a
proteomic approach. Plant Physiology 147: 2147-2163.

Quintero-Zamora, 2010 83
• Sieverding E. (1983). Manual de métodos de investigaciones en micorrizasvesículo-
arbusculares en el laboratorio. Centro Internacional de Agricultura Tropical
(CIAT). Palmira (Colombia).
• Simón L., Bousquet R., Levesque C. y Lalonde M. (1993). Origin and
Diversification of Endomycorrhizal Fungi and Coincidence with Vascular
Land Plants. Nature 363:67-69
• Simoneau P., Louisy-Louis N., Plenchette C. y Strullu D.G. (1994). Accumulation
of New Polypeptides in Ri T-DNA-Transformed roots of tomato
(Lycopersicon esculentum) during the development of vesicular-arbuscular
mycorrhizae. Applied and Environmental Microbiology 60:1810-1813.
• Singh S.P., Gutiérrez J.A. y Terán H. (2003). Registration of indeterminate tall
upright small black-seeded common bean germplasm A 55. Crop Science
43:1887-1888.
• Smith G. S. (1988). The role of phosphorous nutrition in interactions of vesicular
arbuscular mycorrhizal fungi with soilborne nematodes and fungi.
Phytopathology 78:371–374.
• Smith S.L. y Read D. (1997). Mycorrhizal symbiosis. 2ª edición. Capítulo 1.
Academic Press.Stein E., Molitor A., Kogel K. H. y Waller F. (2008).
Systemic resistance in Arabidopsisconferred by the mycorrhizal fungus
Piriformospora indica requires jasmonic acid signaling and the cytoplasmic
function of NPR1. Plant Cell Physiology 49:1747-1751.
• Swarbrick P.J., Schulze-Lefert P. y Scholes J.D. (2006). Metabolic consequences of
susceptibility and resistance in barley leaves challenged with powdery
mildew. Plant Cell and Environment 29: 1061-1076.
• Taylor J. y Harrier L.A. (2003). Expresión studies of plant genes differentially
expresed in leaf and root tisúes of tomato colonised by the arbuscular
mcorrhizal fungus Glomus moseae. Plant molecular Biology 51:619-629.
• Taylor J. y Harrier L. A. (2001). Comparison of development and mineral nutrition
of micropropagated Fragaria x ananassa cv Elvira (Strawberry) when
colonised by nine species of arbuscular mycorrhizal fungi. Applied Soil
Ecology18:205-215.

Quintero-Zamora, 2010 84
• Thelen J. J. (2007). Introduction to proteomics: A brief historical perspective on
contemporary approaches, in: Samaj J., Thelen J. J. (Eds.). Plant Proteomics
Springer, Berlin. Pp. 1–13.
• Ton J., De Vos M., Robben C., Buchala A., Métraux J. P., Van Loon L.C. y Pieterse
C.M.J. (2002). Characterization of Arabidopsis enhanced disease
susceptibility mutants that are affected in systemically induced resistance.
Plant Journal 29: 11-21.
• Trappe J.M. (1987). Phylogenetic and ecological aspects of mycotrophy in the
angiosperms from an evolutionary standpoint. En Safir DG (Ed.)
Ecophysiology of VA mycorrhizal plants. CRC. Boca Raton, FL, EEUU. pp
5-25.
• Truman W., Bennett M. H., Kubigsteltig I., Turnbull C. y Grant M. (2007).
Arabidopsis systemic immunity uses conserved defense signaling pathways
and is mediated by jasmonates. Proceedings of the National Academy of
Sciences of the United States of America 104: 1075-1080.
• Tsugita A. y Kamo M. (1999). 2D Electrophoresis of plant proteins. Methods
Molecular Biology 112:95-97.
• Tunc-Ozdemir M., Miller G., Song L., Kim J., Sodek A., Koussevitzky S., Narayan
Misra A., Mittler R. y Shintani D. (2009). Thiamin confers enhanced
tolerance to oxidative stress in Arabidopsis. Plant Physiology 151: 421-432.
• Vallad G.E. y Goodman R.M. (2004). Systemic Acquired Resistance and Induced
Systemic Resistance in Convetional Agricultura. Crop Science 44:1920-
1934.
• Valot B., L. Negroni, M. Zivy, S. Gianinazzi y E. Dumas-Gaudot. (2006). A mass
spectrometric approach to identify arbuscular mycorrhiza-related proteins in
root plasma membrane fractions. Proteomics 6:145-155.
• Valot B., Dieu M., Recobert G., Raes M. y Gianinazzi, S., (2005). Identification of
membrane-associated proteins regulated by arbuscular mycorrhizal
symbiosis. Plant Molecular Biology 59:565-580.

Quintero-Zamora, 2010 85
• Van der Ent S., Van Wees S.C.M y Pieterse C. M. J. (2009). Jasmonate signaling in
plant interactions with resistance-inducing beneficial microbes.
Phytochemistry 70: 1581-1588.
• Van Loon L. C. y Bakker P. A. H. M. (2006). Root-associated bacteria inducing
systemic resistance. In: Gnanamanickam, S.S. (Ed.), Plant-Associated
Bacteria. Springer Dordrecht, pp. 269–316.
• Van Loon L. C., Bakker P. A. H. M. y Pieterse C. M. J. (1998). Systemic resistance
induced by rhizosphere bacteria. Annual Review. Phytopathology. 36:453-
483.
• Verberne M. C., Hoekstra J., Bol J. F. y Linthorst H. J. M. (2003). Signaling of
systemic acquired resistance in tobacco depends on ethylene perception.
Plant Journal 35: 27-32
• Verhagen, B. W. M., Glazebrook J., Zhu T., Chang H. S, Van Loon L. C. y
Pieterse C.M.J. (2004). The transcriptome of rhizobacteria-induced systemic
resistance in Arabidopsis. Molecular Plant-Microbe Interactions 17(8): 895-
908.
• Vigo C., Norman J. R. y Hooker J.E. (2000). Biocontrol of the pathogen
Phytophthora parasitica by arbuscular mycorrhizal fungi is a consequence
of effects on infection loci. Plant Pathology 49:509–514.
• Vlot, A.C., Klessig, D.F., Park y S.-W. (2008). Systemic acquired resistance: the
elusive signal(s). Current Opinión in Plant Biology 11: 436-442.
• Walters D. y Heil M. (2007). Costs and trade-offs associated with induced
resistance. Physiological and Molecular Plant Pathology 71:3-17.
• Wang W., Scali M., Vignani R. y Spadafora A. (2003). Eletrophoresis 24: 2369-
2375.
• Whipps J. M. (2004). Prospect and limitations for mycorrhizas in biocontrol of root
pathogens. Canadian Journal of Botany 82:1198-1227.