parasitoid es
DESCRIPTION
ecologiaTRANSCRIPT

Introducción
El parasitoidismo es una relación intermedia entre la depredación y el parasitismo. Los parasitoides (en su gran mayoría insectos ) como parte de su ciclo de vida depositan un huevo en o cerca de su hospedador o huésped (también por lo general un insecto), luego las larvas viven como ectoparásitos o endoparásitos , según la especie. De esta manera se desarrollan en su víctima durante su ciclo larval.
Los parasitoides tienes características distintivas como que al final de su ciclo larval el hospedador muere (característica que lo diferencia de los parásitos comunes). Al igual que cada parasitoide utiliza sólo un hospedador durante su ciclo de vida (diferencia respecto de los depredadores, que matan varias víctimas a lo largo de su vida).
Para que un parasitoide deje futuras generaciones tiene que tener un hospedador, pero este debe de cumplir con elementos nutritivos o protección del nuevo organismo en su mayoría el huésped son otros insectos, rara vez otros artrópodos.
Los insectos parasitoides se distinguen de los insectos parasitos por que están mayor mente restringidos a los órdenes Hymenoptera y Diptera. La relación de parasitismo se da solo en larvas aunque algunos adultos pueden alimentar al huésped, las larvas del parasitoide matan al huésped solo al completar su desarrollo.
La diferencia respecto a depredador y parasito radica en que el depredador se alimenta de un solo huésped, pero no le produce la muerte. Por tanto en muchos aspectos un parasitoide se encontraría entre un depredador y una parasito. Como depredador porque siempre le produce la muerte a su huésped y como parasito porque solamente se alimenta de un huésped. Los estados evolutivos de un parasitoide son: huevo, pupa y adulto.
Otra característica de los parasitoides son sus adaptaciones para ovipositar, lo que ha llevado a la evolución de sofisticados ovopositores especialmente en especies que parasitan larvas protegidas en lugares de difícil acceso como troncos de árboles y túneles en el suelo.

Los parasitoides se clasifican de muchas maneras, especialmente por el habito de su larvas. de acuerdo al estado del hospedero que atacan, hay parasitoidedes de huevos, del larvas, de pupas o de adultos, cuando sus hospederos son holometábolos. Cuando atacan hemimetábolos los parasitoides son de huevos o de ninfas. Como veremos más adelante, algunos parasitoides pueden pasar de un estado de desarrollo al siguiente, entonces hay parasitoides de huevo-larva o de larva-pupa. los parasitoides pueden ser endoparasitoides cuando se desarrollan dentro del cuerpo del hospedero o ectoparasitoides cuando se desarrollan externamente sobre el cuerpo de la víctima. Cuando un sólo individuo se desarrolla en un hospedero el parasitoide es solitario. Cuando más de uno se desarrolla en un solo hospedero el parasitoide es gregario, en este caso se pueden desarrollar desde 2 hasta varios miles de individuos dentro del mismo hospedero. esta terminología permite combinaciones tales como "endoparasitoide solitario”
Tipos de parasitoidismo
Superparasitismo
Cuando las hembras de la misma especie de parasitoide ponen más de un huevo en el mismo hospedero, se produce SUPERPARASITISMO. En estos parasitoides se presentan larvas de primer instar móviles y agresivas que tienen mandíbulas bien desarrolladas.
Multiparasitismo
Cuando una hembra de una segunda especie oviposita en un hospedero que ya estaba previamente parasitado por una primera especie, pueden suceder dos cosas: si la progenie de la segunda especie compite con las larvas residentes de la primera especie por los recursos que proporciona el hospedero, se produce MULTIPARASITISMO y eventualmente las dos especies se desarrollan dentro del mismo hospedero.
Hiperparasitismo
Si la hembra de la segunda especie va en busca de la larva residente de la primera especie, esto equivale a ser un parasitoide de otro parasitoide lo que se conoce como HIPERPARASITISMO. Los hiperparasitoides tambien se conocen como parasitoides secundarios. Hay casos de parasitoides terciarios. Los hiperparasitoides pueden atacar sus hospederos de dos formas.

Ovipositar antes que llegue el parasitoide primario y "esperar" a que este sea parasitado para atacar su hospedero parasitoide. En este caso se denominan HIPERPARASITOIDES FACULTATIVOS. Cuando los hiperparasitoides sólo se pueden desarrollar como parasitoides de parasitoides, y deben encontrar un hospedero que ya esté parasitado se denominan HIPERPARASITOIDES OBLIGATORIOS.
Cleptoparasitismo
Un cleptoparasitoide obligatoriamente requiere la presencia de otra especie de parasitoide, pero no se alimenta de este sino que utiliza recursos del primero en su propio beneficio. Por ejemplo: los parasitoides de laravas barrenadoras de madera que carecen de ovipositor largo y fuerte para perforar el tronco, esperan que otro parasitoide, que sí tiene la adaptación del ovipositor, perfore y se "aprovechan" de este comportamiento para "robar" la perforación del anterior y ovipositar en su hospedero dentro del tunel en el tronco.
Koinobiontes
Aquellos parasitoides que permiten que el hospedero continue su crecimiento despues de haber sido parasitado. Esto ocurre con frecuencia en parasitoides que permiten el paso de un estadío del hospedante al otro, como aquellos que son parasitoides de huevo-larva y larva-pupa. En muchos casos los koinobiontes no paralizan el hospedero durante la oviposición. Muchos koinobiontes pueden suspender el desarrollo durante su estado larval o pueden no alimentarse de organos vitales del hospedero para lograr completar su desarrollo.
Idiobiontes
Parasitoides que solo disponen de los recursos del hospedero al momento de la oviposicion, para compleatar su desarrollo. Estos no tienen la oportunidad de que el hospedero continue su crecimiento despues del parasitismo: son típicamente parasitoides de huevos, de pupas o de adultos y no pasan de un estado al siguiente. Estos parasitoides en muchos casos paralizan la presa durante la oviposición.
Los parasitoides en su mayoría como ya se mencionó son de los órdenes Huymenoptera y Diptera acontinucaion de muestra una tabla desde orden asta familia

Orden Suborden Superfamilia FamiliaHymenoptera Symphyta Orussoidea Orussidae
Apocrita Trygonalyoidea Trigonalyidae
Evanoidea EvaniidaeAulacidaeGasteruptiidae
Cynipoidea IbaliidaeCharipidaeFigitidaeEucolidae
Chalcidoidea LeucospidaeChalcididaeEurytomidaeTorymidaeAgaoinidaePerilampidaePteromalidaeSigniphoridaeEncyrtidaeAphelinidaeTrichogrammatidaeMymaridaeEulophidaeEucharitidae
Proctotrupoidea ProctotrupidaeDiapriidaeScelionidaePlatygastridaeRoproniidae
Ceraphronoidea MegaspilidaeCeraphronidae
Ichneumonoidea IchneumonidaeBraconidae
Aculeata Chrysidoidea DryinidaeBethylidaeChrysididae
Vespoidea TiphiidaePompilidae
Diptera Nematocera CecyidomiidaeBrachycera Acroceridae
BombylidaeNemestrinidae
Cyclorrhapha PhoridaePipunculidaeConopidaeSarcophagidaeTachinidae
Tabla 1° ordenes representativos de parasitoides

Fig 1 principales parasitoides y sus hospederos
Los parasitoides están adaptados a reaccionar a una serie de estímulos y a interpretar ciertas claves provenientes del hospedero o de su hábitat para encontrar un hospedero adecuado para dejar su descendencia. Una vez hallado el hospedero el parasitoide, especialmente la hembra debe decidir si este ofrece las condiciones óptimas para maximizar su tasa de transmisión de su genoma a la siguiente generación. La optimización que los parasitoides han hecho evolutivamente de estos aspectos de la localización del hospedero y de la maximización de sus tasas reproductivas han hecho de ellos organismos de mucha importancia en la regulación de población de insectos plaga. En esta parte vamos a describir en más detalle los mecanismos de los parasitoides para la localización del hospedero, a partir de la localización de su hábitat y el hospedero, las decisiones que debe tomar la hembra para optimizar su tasa reproductiva cuando se enfrenta a un foco de hospederos y una de las teorías de optimización del manejo de esos focos, el teorema del valor marginal.

Los cinco aspectos más importantes de la localización del hospedero por parte de un parasitoide listo para ovipositar implican:
1. Localización del hábitat al que está asociado el hospedero.
2. Búsqueda y descubrimiento del hospedero mismo.
3. Localización y reconocimiento.
4. Evaluación y aceptación.
5. Uso de focos del hospedero ("Patch use")
1 Localización del hábitat al que está asociado el hospedero. El parasitoide emerge como adulto, en muchos casos, directamente en el hábitat que ocupa su hospedero. Sin embargo, si no ha copulado, o si no esta maduro reproductivamente, muy probablemente tendrá que abandonar ese hábitat para ir en busca de pareja y copular. A continuación tendrá que localizar de nuevo su hospedero. En otras ocasiones el adulto emerge lejos del hospedero, por ejemplo, porque este ya no está en el área debido a que pasó su ataque, o a que su población fue diezmada por factores ambientales, por parasitismo o depredación. En este caso como en el anterior, el parasitoide adulto tiene que localizar el hospedero, muy probablemente a grandes distancias de donde se halla. Estas han sido algunas de las razones por las cuales los parasitoides han evolucionado mecanismos muy eficientes para localizar el hábitat del hospedero, especialmente a largas distancias, como primer paso para la localización de su hospedero.
Orientación a larga distancia: A grandes distancias el parasitoide tiene la opción de localiza su hospedero directamente (difícil) o localizar el hábitat del hospedero donde aumenta la probabilidad de encontrarlo. Al orientarse bien sea hacia el hábitat o directamente hacia el hospedero, el parasitoide se enfrenta con el problema de "confiabilidad/detectabilidad" descrito por Vet y Dicke (1992). Las señales que obtiene del hospedero son generalmente muy confiables pero son poco detectables, mientras que las producidas por el hábitat son altamente detectables pero poco confiables en términos de que allí se halle el hospedero adecuado. Las señales más comúnmente usadas por los parasitoides para localizar el hábitat de sus hospederos son olfativas, visuales o auditivas.

Señales olfativas: Son de dos tipos principales: feromonas, que actúan sobre individuos de una misma especie (p.ej. feromonas sexuales) y las alelo químicas que se usan para comunicación entre dos o más especies. Las señales olfativas se clasifican de acuerdo al efecto que tienen sobre el organismo emisor y el receptor
Tipo Efecto sobre Emisor
Efecto sobre Receptor
Ejemplo
Alomonas Positivo Negativo VenenosKairomonas
Negativo Positivo Volátiles en las heces
Sinomonas Positivo Positivo Volátiles de plantas
Tabla 2. Efecto de alelo químicos sobre el emisor y el receptor.
Señales Visuales. El parasitoide puede usar señales visuales, como color y forma, para localizar el hábitat del hospedero.
Señales Auditivas: Orientación a corta distancia
Gustativas:
Mecano receptivas: Evidencia localización hábitat, Hospedero polífago atacado solo en una planta: Helicoverpa armigera Microbracon brevicornis - Antirrhinum sp.
2 Búsqueda y Descubrimiento del Hospedero mismo: Las claves provienen de:
Plantas bajo ataque: Plantas atacadas por un herbívoro producen mayores volúmenes y diferentes tipos de volátiles que plantas sanas.

Heces del herbívoro u otros componentes asociación insecto/planta: Respuesta es mayor hacia plantas que tienen el hospedero, el daño u otro material relacionado.
Otros organismos asociados: Hongos y bacterias asociados a la descomposición del hospedante producen compuestos que usa el parasitoide
El mismo hospedero: Claves auditivas y visuales, especialmente importante en grupos que usan comunicación auditiva
3 Localización y Reconocimiento del hospedero:
Claves del micro hábitat y la planta
Claves indirectas del hospedero: volátiles plantas atacadas, heces
Claves directas del hospedero: acústicas (sonidos, vibraciones) visuales, táctiles, olfativas.
4 Evaluación y Aceptación.
¿Se puede usar el hospedero para ovi-posición?
¿Se puede usar como alimento?
¿Esta previamente parasitado?
¿Se ignora o se parasita?
Experiencia previa, información obtenida durante la búsqueda, Aprendizaje
5 Uso del hospedero.
Teoría de la optimización del forrajeo. Entre ellas la más importante en este caso es la que tiene que ver con el Teorema del Valor Marginal, propuesto por Charnov (1976). Este teorema predice que el tiempo óptimo que tiene un parasitoide para abandonar el foco del hospedero es cuando su tasa instantánea de ganancia reproductiva llega al promedio máximo que se puede obtener en ese foco. Esto se explica mejor en una gráfica:

Figura 2. Teorema de Charnov sobre la optimización de la utilización de un foco de hospederos por una hembra parasitoide.
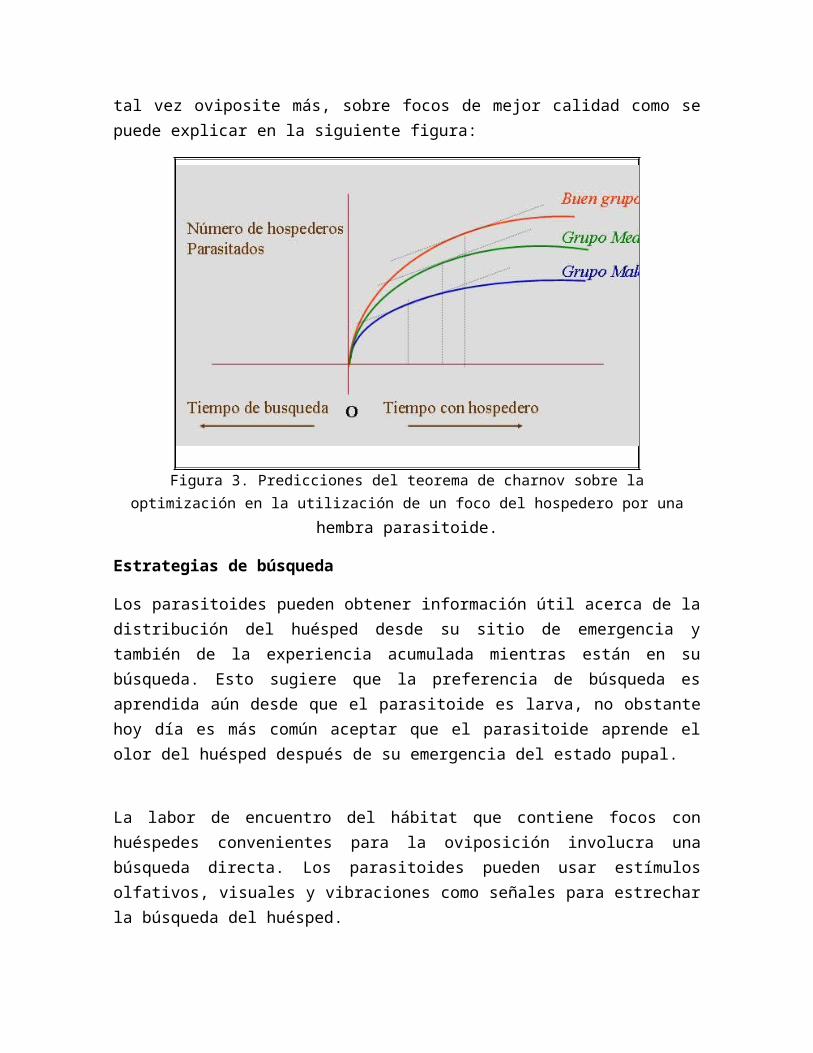
No todos los focos del hospedero que el parasitoide encuentran son de la misma calidad. La hembra debe evaluar la calidad del foco para tomar la decisión y optimizar su postura. Charnov predice que la hembra pasaría más tiempo, y tal vez oviposite más, sobre focos de mejor calidad como se puede explicar en la siguiente figura:
Figura 3. Predicciones del teorema de charnov sobre la optimización en la utilización de
un foco del hospedero por una hembra parasitoide.

Estrategias de búsqueda
Los parasitoides pueden obtener información útil acerca de la distribución del huésped desde su sitio de emergencia y también de la experiencia acumulada mientras están en su búsqueda. Esto sugiere que la preferencia de búsqueda es aprendida aún desde que el parasitoide es larva, no obstante hoy día es más común aceptar que el parasitoide aprende el olor del huésped después de su emergencia del estado pupal.
La labor de encuentro del hábitat que contiene focos con huéspedes convenientes para la oviposición involucra una búsqueda directa. Los parasitoides pueden usar estímulos olfativos, visuales y vibraciones como señales para estrechar la búsqueda del huésped.
Una o mas modalidades de acuerdo a la ecología del huésped, pueden ser usadas para localizarlo. Un concepto muy empleado es el de "sistema tritrófico", que relaciona tres niveles de movimiento de la energía a lo largo del sistema de plantas, insectos fitófagos y sus enemigos naturales. Información importante para el parasitoide esta disponible desde el primer y segundo nivel trófico. Por "espionaje" de la comunicación química entre la planta y el insecto fitófago, el parasitoide puede detectar la presencia de potenciales huéspedes. No obstante, señales químicas derivadas del huésped pueden ser poco detectables a largas distancias. Aun así, el parasitoide puede emplear información menos confiable como semioquímicos producidos por la planta, señales visuales como forma y color y señales vibratorias (Vet 1991).
Los parasitoides han involucrado al menos tres estrategias para explotar el "sistema tritrófico".
La primera estrategia es llamada el "desvío infoquímico". Esta involucra el uso de la información del estado mas detectable en el ciclo de vida del huésped. Por ejemplo, Opius lectus un parasitoide de huevos y larvas de Rhagoletis pomonella aprovecha la feromona que disuade la oviposición en el adulto de R.. pomonella (Prokopy and Webster 1978).
La segunda estrategia es llamada "sinomonas que inducen al herbívoro" y se refieren a la capacidad del parasitoide para responder a la emisión de volátiles causados por la planta cuando es dañada por el insecto. Por ejemplo, plantas de maíz dañadas por larvas de Spodoptera liberan volátiles que atraen al parasitoide.

La tercera estrategia es llamada aprendizaje asociativo. Los parasitoides son muy buenos relacionando diversas señales. El beneficio de¡ aprendizaje es que pueden asociar fácilmente señales detectables y poco confiables, como los volátiles de la planta, con señales mas confiables pero difícilmente detectables, como los olores específicos del huésped (Vet 1991).
Además, estudios han indicado que las hembras parasitoides pueden aprender a responder a nuevos olores. Una de las implicaciones del aprendizaje asociativo es que crías de parasitoides pueden ser "entrenadas" en laboratorio para mejorar el éxito en el forrajeo inicial.
Mecanismo general de selección del huésped
En algún momento una hembra parasitoide desarrolla un afán por buscar huéspedes, el cual, presumiblemente esta inducido por un conjunto de condiciones fisiológicas. Algunas especies pueden tener un período de preoviposición y de igual manera no sentir el afán de búsqueda luego de que este periodo ha pasado; así mismo, otras especies pueden empezar a buscar el huésped inmediatamente han emergido. El afán de búsqueda se asocia con una serie de factores internos tales como hambre, necesidad de limpieza, edad, carga de huevos y experiencia y factores externos como presencia de predadores o competidores, disponibilidad de refugio, condiciones ambientales, enfermedades y disponibilidad de alimento.La experiencia previa de una hembra es muy importante para la localización del huésped. Involucra una cadena de sucesos que son específicos como el comportamiento de oviposición, si se trata de un endo- o ectoparasitoide, etc. Esta experiencia puede ser de tres tipos, así:
La primera, denominada innata o inexperiencia, se refiere a aquellas hembras que no han tenido contacto con huéspedes o plantas asociadas a ellos. En estas hembras los genes ("fundamento genético") determinan la primera señal a la cual responden, el tipo de señales a las cuales responden, el comportamiento que generan dichas señales, el rango de especies que puede atacar la hembra y la intensidad del ataque. El estimulo puede variar hasta un umbral luego del cual se puede generar una repuesta. Este comportamiento puede estar o no realzado por una combinación de estímulos. En general la detección y respuesta innata a señales estaría sujeta a selección natural.

El segundo factor que afecta la respuesta de selección del huesped es la experiencia. Experiencia con oviposición o localización puede mejorar la destreza o realzar la respuesta para selección del huesped por la hembra. Las hembras con experiencia responden mas rápido y vuelan mas a prisa hacia el complejo huésped / planta con respecto a hembras inexpertas Tal experiencia puede extenderse a todos los niveles del proceso de selección del huésped; por ejemplo, el tiempo necesario para reconocer y parasitar el huésped disminuye con el aumento en la sucesiva manipulación de huéspedes. Otro tipo de aprendizaje es el condicionado, el cual es definido como la respuesta a una nueva señal resultado de la asociación con una sería innata o una señal para la cual la hembra esta programada genéticamente para responder durante una experiencia que involucre a ambas. Vinson se refiere al, aprendizaje condicionado como experiencia y al condicionado como aprendizaje asociativo.
El tercer componente de una experiencia previa es el aprendizaje asociativo en el cual una hembra asocia una "nueva" señal con señales" innatas". Esto resulta en hembras que tienen la capacidad de responder apropiadamente a señales que ellas pueden detectar pero para las cuales ellas no están programadas genéticamente para responder. El aprendizaje asociativo redirecciona y amplia la respuesta de las hembras. El resultado es un sistema flexible que permite a las hembras responder a cambios en el ambiente (Lewis et al., 1990; Vet et al., 1990). El aprendizaje asociativo también permite que las hembras respondan a nuevos huéspedes o a huéspedes en nuevas situaciones sin la influencia genética de la población del parasitoide, dado que la selección puede actuar en la capacidad para aprender asociativamente y no en la capacidad para detectar o responder a una señal particular.