paranthropus boisei: an example of evolutionary stasis?
TRANSCRIPT

AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 9 5 1 17-136 (1994)
Paranthropus boisei: An Example of Evolutionary Stasis? BERNARD WOOD, CHRISTOPHER WOOD, AND LYLE KONIGSBERG Hominid Palaeontology Research Group, Department of Human Anatomy and Cell Biology, The University of Liverpool, PO Box 147, Liverpool L69 3BX, UK (B. W., C. W.) and Department of Anthropology, University of Tennessee, Knoxville, Tennessee 37996-0720 (L.K.)
KEY WORDS analysis, Punctuated equilibrium, Phyletic gradualism
Paranthropus boisei, Masticatory evolution, Trend
ABSTRACT Of the presently recognised early hominid species, Paran- thropus boisei is one of the better known from the fossil record and arguably the most distinctive; the latter interpretation rests on the numbers of appar- ently derived characters it incorporates. The species as traditionally diag- nosed is distributed across approximately one million years and is presently confined to samples from East African sites. The hypodigm has been exam- ined for evidence of intraspecific phyletic evolution, with particular emphasis on the areas best represented in the fossil record, namely the teeth and mandible. The results of this examination of 55 mandibular and dental vari- ables, which uses the r test statistic for the detection of trend, and nonpara- metric spline regression (Loess regression) for investigating pattern and rate of temporal change, show that within Paranthropus boisei sensu stricto most evidence of temporally related morphological trends relates to the morphology of the P, crown. There is little or no evidence of any tendency to increase in overall size through time. Fossils from the Omo Shungura Formation and West Turkana which have been recovered from a time period earlier than the P. boisei sensu stricto hypodigm resemble the latter taxon in some features, but differ from it in aspects of cranial morphology. There is insufficient fossil evidence of the earlier taxon to tell whether it changes through time, but when trends of 47 mandibular and dental variables are assessed across the com- bined East African “robust” australopithecine sample, there is evidence €or a relatively abrupt change around 2.2-2.3 Myr in approximately 25% of the dental and mandibular remains. Some of these changes correspond with the temporal trends within P. boisei sensu stricto, but others, such as mandible height, do not. If the earlier material is ancestral to P. boisei sensu stricto, evidence from the teeth and jaws is consistent with a punctuated origin for the latter taxon. 0 1994 Wiley-Liss, Inc.
Hominid fossil evidence has been cited as providing support both for (e.g., Gould and Eldredge, 1977; Rightmire, 1981, 1990) and against (e.g., Cronin et al., 1981; Wolpoff, 1984) a punctuated equilibrium evolution- ary model. While some of these studies have considered change across most, if not all, of the hominid fossil record (Gould and El- dredge, 1977; Cronin et al., 19811, others have concentrated on just one species (Rightmire, 1981; Wolpoff, 1984). The latter
two authors have respectively cited evidence for, and against, there being statistically significant temporal trends in morphology within material which has been allocated to the species Homo erectus. However, the util-
Received November 23,1993; accepted April 20,1994. Address reprint requests to Bernard Wood, Hominid Palaeon-
tology Research Group, Department of Human Anatomy and Cell Biology, The University of Liverpool, PO Box 147, Liverpool L69 3BX, UK.
0 1994 WILEY-LISS. INC

B. WOOD ET AL. 118
ity of all such studies is greatly reduced if there is no consensus about alpha taxonomy with inevitable and crucial disagreements about what specimens should be included in species hypodigms.
Of all the early hominid species available for study, there is evidence that Paranthro- pus boisei offers a particularly suitable model for determining both the mode and rate of evolutionary change. Its suitability rests on its morphological distinctiveness (Wood, 19911, its relatively restricted geo- graphical range, the quantity and quality of the fossil evidence, and the quality of the stratigraphic and chronological control of much of the hypodigm (Feibel et al., 1989). Nonetheless, it would be surprising if any hominid species was free from taxonomic controversy and P. boisei is no exception. When P. boisei is defined according to the criteria set out by Wood (1991), that is, using a diagnosis which concentrates on cranial features which are seen consistently in both presumed male and female crania from Koobi Fora and elsewhere, the taxon can be recognised from remains which span the time period between 2.0 and 1.4 Myr (Fig. 1). Variations in cranial morphology have been demonstrated (Wood, 1991; Brown et al., 1993) but the basic design of the cranium is consistent within the hypodigm. However, Suwa (1988), using an admittedly narrow suite of mandibular premolar features he has identified in suitable teeth from the Shungura Formation, proposes that the spe- cies can be traced back to Member G times, that is to around 2.3 Myr (Fig. 1). In the present study the hypodigm recognised by Wood (1991) and Suwa (1988) is identified as P. boisei sensu stricto.
Remains dating from before 2.3 Myr which variously incorporate large, low, and wide mandibular corpora, large postcanine teeth and heavily crested crania with a more sagittally oriented petrous axis and a wide mid-face (Coppens, 1979; Walker et al., 1986; Leakey and Walker, 1988) are re- ferred to as P. aff. P. boisei. Arambourg and Coppens (1968) placed earlier discoveries from the Shungura Formation in this time range in a novel genus and species, Paraus- tralopithecus aethiopicus, the holotype of which is the edentulous mandibular corpus
Omo 18-18 from Member C. More recently Coppens (1979) listed this specimen as part of the P. boisei hypodigm. Walker et al. (1986) and Leakey and Walker (1988) have suggested that there may be grounds for re- viving P. aethiopicus on the basis of similar- ities between KNM-WT 16005 and Omo 18- 18. However, the precise composition of the hypodigm and the taxonomic status of this earlier subset of East African “robust” aus- tralopithecines remains uncertain, with some workers content to place KNM-WT 17000 in P. aethiopicus (Kimbel and White, 1988; Kimbel et al., 1988) while others con- sider it falls within an acceptable range of variation for a palaeontological species, and provisionally retain it in P. boisei (Walker and Leakey, 1988; Brown et al., 1993).
While many of the features of this earlier material are matched by the morphology of P. boisei, it is becoming increasingly evident that a substantial number of morphological characters, particularly in the cranium, find a different expression in KNM-WT 17000 (Delson, 1986; Chamberlain and Wood, 1987; Kimbel et al., 1988; Wood, 1991) (Ta- ble 1). These characters have been pursued both individually (Rak and Kimbel, 1991, 1993, but see Walker et al., 1993 for a con- trary view) and considered at a simpler level en masse (Skelton and McHenry, 1992). In- evitably perhaps, the conclusions differ, for while Walker et al. (1986) and Leakey and Walker (1988) see the morphological differ- ences as compatible with the type of evolu- tionary change that one might expect within a species which is coextensive with an evolv- ing lineage (sensu Bown and Rose, 1987), others have interpreted the differences as sufficient to warrant species recognition (Kimbel et al., 1988) and some have gone as far as to recommend placing the older subset of East African “robust” fossil hominids in a distinct clade, only distantly related to P. boisei sensu stricto (Skelton and McHenry, 1992).
The present paper addresses the question of mode and rate in hominid evolution by concentrating on what is widely recognised as one of the most easily diagnosable early hominid taxa. First, it reports the results of temporal trend analyses of the teeth and mandible of P. boisei sensu stricto, paying

PARANTHROPUS BOISEI EVOLUTION 119
1 .o 1.1
1.2
1.3
1.4
1.5
1.6
1.7
1.8
1.9
2.0
2.1
2.2
2.3
2.4
2.5
2.6
2.1
2.8
2.9
3.0
(OH 3, 38)
KNM-CH 1, 302, 304, P E ” J 1; OH 32; F203-1
KNM-ER 404, 725, 728, 729, 733, 805, 1467, 3887, 3889, 5877, 6080, 6082, 15950; KNM-WT 17396
KNM-ER 3230 KNM-ER 726, 818, 1468
KNM-ER 816; OH 5 KNM-ER 407, 1477, 1806, 3229, 3952, 6128
KNM-ER 405, 1469, 1803, 3731, 13750, 23000
KNM-ER 403, 406, 727, 732, 801. 802, 810. 812, 814, 1170, 1171. 1479, 1509. 1804, 1816, 1818, 1819. 1820, 2607. 3737, 3885, 3886, 3890, 3954, 5429,
KNM-WT 17400; OH 26,30,46,60
3729,
L726-11, Omo 47-46, Omo 47-1500. Omo 323-896
L7A-125, L74A-21, L427-7, M28-1, M28-2, L628-3, L6284, L628-5, Om0 75-14a,b, Om0 136-1,
F22-lq F22-lb. L10-21, L28-58, L28-126, L157-35, L209-17, L209-18, L238-35. L338~-34, L338~-35, L338~-39. L338x-40. L398-14. L398-120, L398-266. I Om0 136-2. Omo 136-3. Om0 141-2 L398-630; L398-847; L398-1223. L398-2608, I U2CL15. U65-111. L860-2. Omo 33-9. Orno 33-62.
1 Om0 3363,Omo 33-65. G o 33-506, Omo 33-507. Omo 33-508,01110 33-6172, Orno 57-41, Omo 57.4-42. Omo 76-37, Omo 76r-11
KNM-WT 16005; L26-59, L64-2, L338x-32, L338x-33, L338y-6, L704-2, Omo 44-1410, I Om0 44-2466, Omo 57.4-147, Omo 57.4-148
KNM-W 17000
L55-33, Om0 18-18, Omo 18-31, Omo 84-100
Fig. 1. Temporal distribution of East African ‘robust’ australopithecine craniodental specimens. Ver- tical bars represent the age ranges for large groups of specimens. Ages taken from Brown and Feibel (19881, Wood (19911, and White (19881.
particular attention to variables reflecting apparently distinctive features of the taxon. For example, does P. boisei sensu stricto be- come “more specialised through time, or are its distinctive features, once established, maintained relatively unchanged for more than one million years? Does this hominid
species follow Cope’s Law, and increase in size through time? Moreover, if there are changes through time, what is their time course?
Second, this paper compares the dental and mandibular evidence €or P. boisei sensu stricto with the temporally earlier, but ap-

B. WOOD ET AL.
TABLE 1. Cranial, mandibular, and dental features of P. boisei sensu strict0 compared with those of P. aff. P. boisei'
120
P . boisei (SS) P. aff. P. boisei (R) Feature
Cranial vault Receding frontal squama with trigonum frontale 111 Supraorbitals in form of costa supraorbitalis [ l ] Temporoparietal overlap at asterion [ l ] Posteriodanterior temporalis ratio large [11* Extensive temporal squama pneumatisation Ill* Strongly flared parietal mastoid angle (?asterionic notch) [I]* Mastoid bulbous, inflated beyond supramastoid crest [ l ] Reduced medial inflection of mastoid process [ l ] Occipital marginal venous drainage system absent [21*
Flat cranial base Ill* Short cranial base [ l l Heart-shaped foramen magnum [ l ] Foramen magnum level witWposterior to bitympanic line [3]* Petrous inclined coronally [11 Tympanic vertically deep, with strong vaginal process [ l ] Vertically inclined tympanic plate inferosuperiorly concave [ l ]
Cranial base
Temporomandibular joint Temporomandibular joint flat, open anteriorly Ell* Postglenoid process anterior to tympanic plate [11* Large horizontal distance between molars and TMJ [11*
Dished midface 111 Strong upper facial prognathism Ill* Inferior orbital margin rounded laterally [21* Low position of infraorbital foramen [21 Single infraorbital foramen [21 Anterior pillars obscured by infilling [21 Upper canine jugae absent [21
Zygomatic process forward relative to palate length [ l l Zygomaticoalveolar crest weakly arched in facial view [ l l Very robust zygomatic arches
Flat, shallow palate [11*
Nasals widest superiorly [ l l Nasion coincident with high glabella [1]* Anterior vomer insertion coincident with anterior nasal spine [ l ] Guttered nasoalveolar clivus grades into nasal cavity floor [ l ] Nasoalveolar contour projects weekly anterior to bicanine line [I] Nasoalveolar clivus convex [21*
Narrow frontal portion [41 Low dorsal cerebral height [41 Laterally-flaring and posteriorly-protruding cerebellar lobes I41 Slight left occipital and right frontal petalia [41
Receding symphysis [21* Wide extramolar sulcus [21 High mandibular ramus origin on corpus [21*
Relatively enlarged postcanine toothrow [ l ] Incisors in bicanine line [ l ] Absolutely large anterior toothrow [11* Maxillarv dental arch convergent Dosteriorlv 111*
Face
Zygomatic region
Palate
Nasal region
Endocast
Mandible
Dentition
X X X
X X
X
X X X X
X X X
X
X X X
X
X
X X
X' X" X Xa X" X" X'
X
X"
X X" X' X"
Xb.c.d
Xb,c,d
Xb,c,d
axi ill& lateral incisor roots medial to naiai aperture margins [I] X xb.c
No mandibular diastema I21 x X' Oval p3 occlusal outline [2] x X
High P, talonid relative to protoconid [21 X X' Strong P, metaconid development [2] X X b . 4
Round P, occlusal outline [21 X Xb.c,d
'Features taken from: 111 Kimbel et al., 1988; 121 Skelton and McHenry, 1992; 131 Wood, 1991; 141 Holloway, 1988. Superscripts indicate that this feature is also shared with a, A. afaremis; b, A. ~ f r i ~ ~ n t ~ ; c, P. robustus; d, Homo spp. Features marked with an asterisk differ between P. boisel semu strict0 andP. aff. P. boisei.

PARANTHROPUS BOISEI EVOLUTION 121
parently cranially distinctive, subset to de- termine whether the cranial distinctions are matched by any general modifications to gnathic morphology in addition to the de- tailed changes that Suwa (1988) noted in the mandibular premolar crown morphology.
Third, if there are differences in mastica- tory variables between the early and late subsets, the study examines whether these variables continue to change within P. boisei sensu stricto.
MATERIALS AND METHODS Some 144 separate specimens comprise
the complete East African ‘robust’ australo- pithecine sample, 70 of which are allocated to P. boisei sensu stricto according to the cri- teria set out in Suwa (1988) and Wood (1991), and 62 of which are allocated to P. aff. P. boisei; a further 12 specimens are too fragmentary or eroded to be included in P. boisei sensu stricto although their allocation to this taxon is likely. Table 2 provides a list of these specimens, together with their taxo- nomic allocation, geological age, morpholog- ical category, and brief anatomical descrip- tion. The sample is heavily biased toward dental and mandibular remains (Table 31, and around 90% of specimens have been re- covered from Koobi Fora and the Omo Shun- gura Formation. Figure 1 shows that the majority of the Omo specimens come from Tuff G and below (c. 2.2-2.5 Myr) while the Koobi Fora material largely derives from de- posits which are a t least 300-400 Kyr years younger. The variables chosen to investigate for temporal changes are listed in Table 4 and their definitions are given in Wood (1991). The criterion for variable inclusion is that the sample size for P. boisei sensu stricto should comprise 5, or more, speci- mens; this constraint effectively confines the metrical and statistical analysis to man- dibular and dental measurements.
Two techniques were used to test for tem- porally related morphological trends within P. boisei sensu stricto and P. aff. P. boisei and for the combined sample. Both of these methods have been used recently by Leigh (1992) in his study of cranial capacity evolu- tion in Homo erectus: the theoretical basis and appropriateness of both of these meth- ods are discussed in Leigh (1992) and are
not rehearsed again here. The nonparamet- ric r (gamma) test statistic (Hubert et al., 1985; Konigsberg, 1990) was used to test for “the presence and significance of directional change” for a given variable (Leigh, 1992: 5). This test simply reveals whether or not vari- ation is distributed randomly over time. Gamma is the inner-product of two vectors, one for the chronological order of specimens and the other for the variable in question. There are N ! permutational gammas possi- ble for randomly rearranged orders: gamma is significant if 5% or less of the random values are equal to or more extreme than the original (correct) ordination. The number of permutations carried out in this study was set at 3000 for any variable where N > 7 and to the exact number for any variable where N was 6 or less.
A second technique, nonparametric Loess regression (Cleveland, 1979; Cleveland and Devlin, 1988; Cleveland et al., 1988; Efron and Tibshirani, 1991; Eubank, 1988) was then used to explore the pattern and rates of any trends made apparent by the former technique. Loess regression works by fitting a series of locally weighted regression lines for different values ofX (i.e., geological age). The weighting used here is the “tricube” (see Cleveland, 1979), so thqt X values further removed from the current point are down- weighted. A regression window of 75% was used for all analyses. In order to make the regressions more resistant to outliers, a bi- weight was used on the residuals (scaled against the median of the residuals) to itera- tively reweight the regressions. All of these procedures are described in detail in Cleve- land (1979). Ninety-five percent confidence limits were found by bootstrapping on the residuals from the observed Loess regression 1000 times, recalculating the Loess regres- sions, and then selecting the 25th and 975th values for each X point.
Prior to the use of these more sophisti- cated techniques the sample parameters of P. boisei sensu stricto and P. aff. P. boisei were computed and the two-tailed Student’s t test, modified by incorporating Satter- thwaite’s estimation oft’ (Snedecor and Co- chran, 1980) which allows for unequal vari- ances, was used to check for significant differences between the two subsets.

122 B. WOOD ET AL.
TABLE 2. List of specimens included in the complete East African <robust' australopithecine (EAFROB) hypodigm'
Suecimen
KNM-ER 403 KNM-ER 404 KNM-ER 405
KNM-ER 406 KNM-ER 407
KNM-ER 725 KNM-ER 726
KNM-ER 728 KNM-ER 727 __ ~~
KNM-ER 729 KNM-ER 732 KNM-ER 733
KNM-ER 801
KNM-ER 802 KNM-ER 805
KNM-ER 810
KNM-ER 812
KNM-ER 814 KNM-ER 816 KNM-ER 818
KNM-ER 1170 KNM-ER 1171
KNM-ER 1467 KNM-ER 1468 KNM-ER 1469 KNM-ER 1477
KNM-ER 1479 KNM-ER 1509 KNM-ER 1803
KNM-ER 1806 KNM-ER 1816
KNM-ER 1818 KNM-ER 1819 KNM-ER 1820 KNM-ER 2607 Kh'M-ER 3229
KNM-ER 1804
KNM-ER 3230 KNM-ER 3729 KNM-ER 3731 KNM-ER 3737 KNM-ER 3885 KNM-ER 3886 KNM-ER 3887 KNM-ER 3889 KNM-ER 3890 KNM-ER 3952 KNM-ER 3954 KNM-ER 5429
KNM-ER 5679 KNM-ER 5877 KNM-ER 6080 KNM-ER 6082 KNM-ER 6128 KNM-ER 13750
TC
ss ss ss ss ss ss ss ss ss ss ss ss ss ss ss ss ss ss cf ss ss ss ss ss ss ss ss ss ss ss ss ss cf ss ss cf ss ss ss cf ss ss cf cf cf ss cf ss ss ss ss cf ss ss ss
- Age 1.70 1.53 1.90
1.70 1.85
1.53 1.60 1.70 1.53 1.53 1.70 1.52
1.70
1.70 1.53
1.78
1.78
1.78 1.80 1.60
1.70 1.70
1.54 1.60 1.90 1.88
1.75 1.70 1.90 1.78 1.85 1.70
1.70 1.77 1.70 1.75 1.85
1.65 1.90 1.90 1.70 1.78 1.78 1.54 1.53 1.70 1.88 1.77 1.77
1.77 1.53 1.54 1.54 1.85 1.90
MC2 Description
D D B
A A
D D D D C A B P
D
E D
D
D
B E D
B E
E D D C
E E D B D E
E E D E D
C D D E E E E D E E D D
E D E E E B
Right side mandibular corpus; weathered crowns P,-M,, roots I,-C Right side mandibular corpus; shattered crowns M,-M, Palate lacking teeth, base of zygomatic process of maxilla and part
conjoined sphenoid Cranium lacking teeth Partial calvaria-frontoparietal frag., base and sides of vault, left
petrous temporal Left side mandibular corpus; roots P,-P, Left side mandibular corpus; roots P,-M, Frag. right side mandibular corpus; roots MI-M, Right side mandibular corpus; roots MI-M, Mandible with most of right ramus; all teeth except right I, Demi-cranium-facial, occipital frags.; right P4, 14, and roots M1-M3 Right side mandibular corpus with M, and roots C-M,; left maxilla with M2 root and M' crown; cranial frags. Right side mandibular corpus; M,-M,, roots MI and associated left
M,-M, and right I, Right and left P,, right M,, left M,-M,, right M,, left upper C and P3 Frag. left side mandibular corpus; frag. right alveolar process and
corpus
left M,
left 11-1, and P,-P,
Left side mandibular corpus and small part of anterior right side;
Frag. left side juvenile mandibular corpus; roots di,-dm,, germs of
Cranial frags., including part of right frontal Left upper C and molar frags. Left side mandibular body; cracked crowns left P3-M3, roots right I,
and left I,-C Cranial vault frags. Right I,, left P4 germ, left and right M,-M, germs, left and right M2 germs, right upper C germ Isolated right M, Right side mandibular corpus; roots P,-M, Left side mandibular corpus; crowns M,-M,, roots P3-M3 Juvenile mandible lacking rami; left dc, M,, left and right dm,-dm,,
Frag. left and right molar crown frags. Left C, P,, M,-M,, and C frag. Frag. right side mandibular corpus; roots I,-P4 Frag. right maxilla; worn crowns P3-Mz and roots C Mandible; no teeth but roots present Frag. juvenile mandible; left dm,, P,, M,, left M, germ, right MI, left
and right P4 germs Isolated left I' Isolated left M, crown Frag. left side juvenile mandibular corpus; dm,, M, and germs Il-P3 Lower molar frag. Left and right mandibular corpus with tooth roots; isolated left and
Mandibular corpus with complete dentition Left side mandibular corpus; partial roots C-M, Left side mandibular corpus; partial roots Iz-M2 Frag. right M, and M, Isolated right P, Isolated left dm, (?M') Isolated left molar (?M3) Frag. mandibular symphysis; roots left I, through to right P, Isolated left M, Frag. left I,, left lower C, left P3, MI, and M3 Mandibular corpus; roots left I,-M, and right I,-M, Right mandibular frag. and symphysis; weathered right P, and M,
Partial left M, crown Right side mandibular corpus; M, and roots P,-M, Germ of right M, Germ of left P, Right P3 crown Partial cranium and two molar frags.
left I, crown, C and P, germs
right P,, and left I,
and tooth roots
(continued1

123 PARANTHROPUS BOISEI EVOLUTION
TABLE 2. Llst of specimens included in the complete East African hbust'australopithecine (EAPROBI hypodigm' (continued)
Specimen TC Age MC2 Description
KNM-ER 15930 KNM-ER 15940 KNM-ER 15950 KNM-ER 16841 KNM-ER 17760 KNM-ER 23000 KNM-WT 16005
KNM-WT 17000 KNM-WT 17396 KNM-WT 17400
KNM-CH 302 KNM-CH 304 PENINJ 1 OH 3 OH 5 OH 26 OH 30 OH 32 OH 38 OH 46 OH 60 F22-la F22-lb F203-1 L7A-125 L10-21 L26-59 L28-58 L28-126 L55-33 L64-2 L74A-21 L157-35
KNM-CH 1
L209-17 L209-18 L238-35 L338x-32
L338x-34 L338x-35 L338x-39
L338y-6
L398-120
L338~-33
L338~-40
L398-14
L398-266 L398-630
L398-1223 L398-2608
L398-847
L420-15 L427-7
L628-1 L628-2
L465-111
L628-3 L628-4 L628-5 L704-2 L726-11 L860-2 Omo 18-18 Omo 18-31 Omo 33-9
ss ss ss ss ss ss R
R ss ss ss ss ss ss ss ss ss ss cf ss cf cf R R ss ss R R R R R R ss R R R R R R R R R R R R R R R R R R R R R R R R R R R ss R R R R
1.78 ?1.78 ?1.40 ?1.78 ?1.78 1.90 2.40
2.50 ?1.50 1.78 1.40 1.40 1.40 1.40 1.20 1.80
?1.70 1.70
?1.40 1.20 1.70 1.70 2.34 2.34 1.42 2.20 2.38 2.40 2.34 2.30
?2.60 ?2.40 2.20 2.34 2.35 2.35 2.30 2.40 2.40 2.38 2.38 2.38 2.38 2.40 2.35 2.35 2.35 2.35 2.35 2.35 2.35 2.34 2.20 2.34 2.25 2.25 2.25 2.25 2.25
?2.40 2.10 2.34 2.60
?2.60 2.35
D E E C E A C
A E A B E B C E A E B E E E E E E E C E E E E D E D E E E E E E E E E E B E E E E E E E E D E E E E E E E E D D E E
Left side mandibular corpus; M1-M3 crowns and roots C-P, Isolated left and right M, or M, g e m s Isolated left M, Edentulous mandibular corpus Isolated lefi I2 Calvaria with mesial root and mesial crown frag. M, Frag. mandibular corpus; crowns left P,-M, and right P3-MI, and roots of anterior teeth except left I, Cranium with isolated right P3 or left P4 and tooth roots Isolated left M, Anterior cranium with left and right I*-M' and left M3 perm Right facial skeleton with right palate and C-M3 Frag. upper molars Frag. cranium in five parts Mandible with complete dentition Upper dm2 and lower dc Cranium and full maxillary dentition Isolated right M, Cranial frags.; dm', dm', C, P3-M1, Il-C, M, Deciduous tooth frags. (? dml) Left and right I,, and right M, Isolated right P" Isolated M3 Isolated right M, crown Isolated right M, crown Isolated right M, Mandibular corpus with all teeth except three incisors Incomplete right M, Incomplete left M2 or M3 Worn right M2 Partial crown of lower premolar Left mandibular frag., I, and P, Isolated left dm, Right hemi-mandible; C-P, Isolated left M, Incomplete right MI or M, Lower molar frag. Isolated right M2 Incomplete left dm2 Incomplete right P4 Left M1 or M2 Isolated right P3 Isolated left M, crown Incomplete left P, with root Parietooccipital portion of cranium Incomplete right dm, (or dm, or M,) Isolated right P, Incomplete right M, Incomplete right M, Distal portion of left M, Incomplete right P, Isolated right M3 Left P, crown Right side mandibular corpus; P,-PI, M, and roots dm, and M, Incomplete left P, Isolated left P4 Incomplete left M3 Isolated left M, Isolated left P, Isolated left p4 Isolated left dm, Incomplete p3 crown Partial mandibular corpus; right P,, M, and left P,-P, Mandibular corpus and symphysis with tooth roots Isolated P, Isolated right M,
(continued)

B. WOOD ET AL.
TABLE2. List ofsmx%nens included in the complete East African ‘robust’australopithecine (EAFROB) hypodigm’ (continudl
124
Specimen TC Age MC2 Description
Orno 33-62 R Om0 33-63 R Orno 33-65 R Om0 33-506 R Orno 33-507 R Orno 33-508 R Orno 33-6172 R Orno 44-1410 R Orno 44-2466 R Orno 47-46 ss Orno 47-1500 ss Orno 57-41 R Orno 57.4-42 R Orno 57.4-147 R Orno 57.4-148 R Orno 75-14a,b R
Orno 76-37 R Orno 76r-11 R Om0 84-100 R Orno 136-1 R Orno 136-2 R Orno 136-3 R Orno 141-2 R Orno 323-896 ss
2.30 2.30 2.35 2.30 2.30 2.35 2.30 2.40 2.40 2.10 2.10 2.38
?2.38 2.40 2.40 2.20
2.30 2.30
?2.60 2.20 2.20 2.20 2.25
E E E E E E E E D E E D E E E D
E E E E E
Incomplete lower premolar or right M3 Isolated right M’ Left M, germ Isolated right P3 Right P, or left M3 Isolated left P, Isolated right M, Incomplete right P4 Frag. left side mandibular corpus; roots M, Worn right M, Isolated right M, Right side mandibular corpus and symphysis; incomplete P, and MI Isolated MI Left P, frag. or left M3 frag. Incomplete right lower PM Associated right maxilla and mandibular frags. with P4-M3 and
Buccal half left M, Incomplete right M, Isolated right M, Isolated left M, Isolated left M,
P3-M3
E Tooth crown frag. E M’ or M2, distal portion germ left M3
2.10 B Partial cranium; P3-M1
‘Specimens marked SS in the taxonomic category column (TC) are allocated top. boisei sensu stricto according t o criteria set out in Wood (1991) and Suwa (1988). Some ‘robust‘ specimens are too fragmentary or eroded to assign at the species level though allocation to P. boisei is likely (cn; better preserved material which either lacksP. boiser autapomorphies or which antedate the time range ofP. botsei is referred to asp. aff. P. bnrser (R). This table provides morphological categories of specimens (MC) and brief anatomical descriptions. Ages taken from Brown and Feihel(1988), Wood (19911, and White (1988). ‘Key to morphological categories: A, well-preserved crania; B, fragmentary crania; C, well-preserved mandibles; D, fragmentary mandibles; E, isolated teeth.
RESULTS The hypodigm of P. boisei sensu stricto
provides sufficient data to test 55 linear and areal variables and seven computed indices for evidence of directional change across its approximately one million years time range. Only six variables, or <lo%, were identified as showing evidence of non-random change (Table 5a). One, the breadth of the mandibu- lar canine socket, involves the anterior den- tition, the remainder being concentrated on the size of the P, crown and the length of the postcanine tooth row, There is redundancy in the six variables. Variables 30,32, and 22 are unlikely to change independently of one another. Likewise a change in variable 40 will lead inevitably to a change in variable 23. Thus, apart from the mandibular canine, only P, size and M, mesiodistal length show
evidence of directional change in P. boisei sensu stricto. The data were too meager to investigate whether there was evidence of nonrandom trends in P. aff. P. boisei.
The visible representations in the Loess regression plots (Fig. 2A-C) help to deter- mine whether the few dental and mandibu- lar variables that change nonrandomly in P. boisei sensu stricto do so in ways that em- phasise its distinctiveness. For example, there is sound evidence to suggest that one of the distinctive features of P. boisei sensu stricto is the small absolute and relative size of the canine tooth crowns (Wood and Stack, 1980; Wood and Chamberlain, 1987; Wood, 19911, but the increase through much of the time span of P. boisei sensu stricto in the maximum labiolingual diameter of the man- dibular canine crown (Fig. 2A) not only fails

PARANTHROPUS BOISEI EVOLUTION 125 TABLE 3. Specimens included in the complete East African ‘robust’ australopithecine (EAFROB) hypodigm listed
accordinp to morDholopica1 cateporv and site’
A Well-preserved crania KOOBI FORA-KNM-ER 406; 407; 732; 23000
OLDUVAI4H 5 WEST TURKANA-KNM-WT 17000; 17400
B Fragmentary crania
CHESOWANJA-KNM-CH 1; 304 KOOBI FORA-KNM-ER 405; 733; 814; 1170; 1804; 13750
OLDYVAI-OH 30 OMO-L338y-6; Omo 323-896 C Well-preserved mandibles KOOBI FORA-KNM-ER 729; 1477; 3230; 16841 WEST TURKANA-KNM-WT 16005 WEST NATRON-PENINJ 1 OMO-L7A-125
N = 7 (4.8%)
N = 11 (7.6%)
N = 7 (4.8%)
D Fragmentary mandibles N = 33 (22.8%) KOOBI FORA-KNM-ER 403; 404; 725; 726; 727; 728; 733; 801; 805; 810; 812; 818; 1468; 1469; 1803; 1806; 1820;
3229; 3729; 3731; 3889; 3954; 5429; 5877; 15930 OMO-L55-33; L74A-21; IA27-7; L860-2; Orno 18-18; Orno 44-2466; Om0 57-41; Orno 75-14a,b E Isolated teeth N = 87 (60.0%) KOOBI FORA--KNM-ER 802; 816; 1171; 1467; 1479; 1509; 1816; 1818; 1819; 2607; 3737; 3885; 3886; 3887; 3890;
3952; 5679; 6080; 6082; 6128; 15940; 15950; 17760 WEST TURKANA-KNhf-WT 17396 CHESOWANJA-KNM-CH 302 OLDUVAI-OH 3; 26; 32; 38; 46; 60 OMO-F22-la; F22-lb; F203-1; L10-21; L26-59; L28-58; L28-126; L64-2; L157-35; L209-17; L209-18; L238-35;
L338~-32; L338~-33; L338~-34; L338~-35; L338~-39; L338~-40; L398-14; L398-120; L398-266; L398-630; L398-847; L398-1223; L398-2608; L420-15; L465-111; L628-1; L628-2; L628-3; L628-4; L628-5; L704-2; L726-11; Orno 18-31; Om0 33-9; Orno 33-62; Orno 33-63; Orno 33-65; Om0 33-506; Orno 33-507; Orno 33-508; Orno 33-6172; Orno 44-1410; Orno 47-46; Orno 47-1500; Orno 57.4-42; Orno 57.4-147; Orno 57.4-148; Orno 76-37; Orno 76r-11; Om0 84-100; Orno 136-1: Orno 136-2: Orno 136-3: Orno 141-2
‘This table provides the numbers of specimens within each category and the percentage each category contributes to the whole sample. Note that KNM-ER 733 is included in categories B and D.
to emphasise but actually deemphasises this dental distinction. Otherwise, the in- creased size of the P4 crown and the increase in the length of one of the components of the mandibular molar tooth row (Fig. 2B and C) are trends which tend to increase the dis- tinctiveness of P. boisei sensu stricto (Wood, 1991; Suwa, 1988). It is noteworthy that the characteristically large cross-sectional area and the substantial robusticity of the man- dibular corpora of P. boisei sensu stricto ap- parently do not change, other than ran- domly, across the one million years, or so, of its fossil history (Fig. 2D and E).
Forty-seven mandibular and dental vari- ables were available to test for any signifi- cant differences between the early (P. aff. P. boisei) and later (P. boisei sensu stricto) sub- sets of East African “robust” australopithe- cines (EAFROB). Of these nine, or approxi- mately 20%, were significantly different (Table 6). It should be borne in mind, how- ever, that some of the sample sizes are small and asymmetric. Consequently the power to
detect differences is low in some cases, the Satterthwaite approximation t’ notwith- standing. A further important consideration is that these are dependent univariate tests, so that the percentage of significant differ- ences may actually fluctuate around the fig- ure given above. Differences include the height of the mandible, as measured at the symphysis (Var. No. 1) (Fig. 3A) and at MI War. No. 91, the size and shape of the P, War. Nos. 30-33, 35) (Fig. 3B) and the length of the mandibular molar crowns (Var. Nos. 37 and 45). When the gamma test sta- tistic was applied to the combined EAFROB sample (Table 5b) seven of the variables mentioned above were identified as showing significant directional change across the whole sample. In addition, non-random change across the larger sample is sug- gested for the width of the mandibular cor- pus at M, War. No. 131, the area of the P4 talonid (Var. No. 36) and the buccolingual diameter of the M, talonid (Var. No. 48). Thus, apart from the increase in mandible

B. WOOD ET AL. 126
TABLE 4. List of variables used in the present study'
Variable Description
1. (141) 2. (142) 3. (143) 4. (144) 5. (145) 6. (147) 7. (148) 8. (149) 9. (150)
10. (151) 11. (152) 12. (154) 13. (155) 14. (156) 15. (157) 16. (158) 17. (159) 18. (161) 19. (162) 20. (164) 21. (165) 22. (167) 23. (168) 24. (193) 25. (194) 26. (195) 27. (257) 28. (258) 29. (259) 30. (271) 31. (272) 32. (273) 33. (274) 34. (275) 35. (276) 36. (277) 37. (285) 38. (286) 39. (289) 40. (313) 41. (314) 42. (317) 43. (318) 44. (324)
45. (341) 46. (342) 47. (343)
48. (344)
49. (345) 50. (346) 51. (347) 52. (349)
53. (350) 54. (352)
55. (354)
56. (lvi) 57. (lix) 58. (Ixiv) 59. (lxv) 60. (lxvi) 61. (lxvii)
Symphyseal height (69) Symphyseal depth (Max) Symphyseal depth (Sl"l') Symphyseal depth (IT") Symphyseal height (GF) Corpus height a t P, Corpus width at P, Corpus area at P, (Co) Corpus height at MI Corpus width at MI Corpus area at MI (Co) Corpus height a t M, Corpus width at M, Corpus area at M, (Co) Corpus height a t M3 Corpus width a t M, Corpus area a t M, (Co) Height of mental foramen (B) Height of mental foramen (Al) Canine socket (LL) Canine socket (MD) P3-P, alveloar length MI-M, alveolar length P3: Mesiodistal crown diameter (Co) P: Buccolingual crown diameter P3: Crown area (Co) P,: Mesiodistal crown diameter P,: Buccolingual crown diameter P,: Crown area (Co) P,: Mesiodistal crown diameter (Co) P,: Buccolingual crown diameter P,: Crown area (Co) P,: Crown area (M) P,: Buccal cusp area P,: Lingual cusp area P,: Talonid area MI: Mesiodistal crown diameter (Co) MI: Buccolingual crown diameter (Max) MI: Crown area (Co) M,: Mesiodistal crown diameter (Co) M,: Buccolingual crown diameter (Max) M,: Crown area (Co) M,: Crown area (M) M,: Reconstructed entoconid area
M,: Mesiodistal crown diameter (Co) M,: Buccolingual crown diameter (Max) M,: Buccolingual crown diameter
M,: Buccolingual crown diameter
M,: Crown area (Co) M,: Crown area (M) M,: Protoconid cusp area M,: Reconstructed metaconid area
M,: Hypoconid cusp area M,: Reconstructed entoconid area
M, Reconstructed hypoconulid area
MI: Corpus Robusticity Index (151/150.100) P3: Crown Shape Index (BL'MD.100) P3: Crown Shape Index (BL'MD. 100) P,: Crown Shape Index (BUMD.100) M,: Crown Shape Index (BL'MD.lOO) M.,: Crown Shape Index (BL'MD.lOO)
(Entoconid 11)
(Protoconid)
(Hypoconid)
(Metaconid 11)
(Entoconid 11)
(Hypoconulid 11)
62. (lxviii) M;: Crown Shape Index (BWMD.100)
'Variahlrs chosen whew N : 5 tiur theP. botier d e ~ u slrrrlu sample. Thr numhcrs in parenrhrses refer 111 thr variahle nuniber in Wuod 119911. The same publication should be culuul1t.d fur vanable drtinitiuns
height and width and the increase in width of the distal M, crown, the majority of the other significant morphological differences between the early and late subsets continue the same trend in P. boisei sensu stricto.
DISCUSSION There have been a considerable number of
discussions surrounding temporal trends in hominid evolution (e.g., Cronin et al., 1981; Rightmire, 1981,1990; Wolpoff, 1984,1988), but the appropriate statistical treatment of the available data is still in its infancy. Con- nor (1986) gives an excellent discussion of methods for assessing trend and periodicity in fossil records, while Manly (1991) dis- cusses general issues in time series analysis that have important implications for the analysis of fossil material. The reliance of many investigators on ordinary regression models is unfortunate, for as Connor (1986: 125) points out, "the application of least- squares regression to autocorrelated data requires that estimates of the regression model account for autocorrelation." Simply stated, the use of ordinary linear regression as a descriptor of temporal trend may be appropriate, but the inferential basis is sus- pect if there is strong serial correlation. Fur- ther, there is every reason to believe that there is correlation between fossils sampled relatively closely together on the geological time scale.
Connor (1986; 125) indicates that Daniels' and the Mann-Kendall tests are "the most powerful and robust tests" for trend analysis on a time series. The Mann-Kendall test has been generalised by Hubert et al. (1985) to account for the magnitude of changes in the series of measurements, and this test has been applied both by Konigsberg (1990) and by Leigh (1992) to human cranial material. Other tests described in Manly (1991) (e.g., the sign test and runs above and below the median test) are not as powerful as the gen- eralisation of the Mann-Kendall test, though they require fewer assumptions about the structure of the data and the pro- cesses that generate the data.
The primary purpose of the present study was to examine the dental and mandibular fossil evidence for P. boisei sensu stricto to determine whether its distinctive morphol- ogy remained static or underwent gradual modification during the million years or so

PARAiiTHROPUS BOISE1 EVOLUTION 127 TABLE 5. Significant results of the I‘ test for trends in both the P. boisei sensu stricto and the complete
EAFROB hypodigm’
Variable
20 22 23 30 32 40
1 13 30 31 32 33 ~~
35 36 37 48
Description N
(a) P. boisei sensu stricto Canine socket (LL) 13 P,-P, alveolar length 13
12
M,: Mesiodistal crown diameter (Co) 14
Symphyseal height 16 Corpus width at M, 28
21
M,-M, alveolar length 19 P,: Mesiodistal crown diameter (Co) P,: Crown area (Co) 12
(b) Complete EAFROB sample
P,: Mesiodistal crown diameter (Co) P,: Buccolingual crown diameter 19 P,: Crown area (Co) 20 P,: Crown area (M) 12 P,: Lingual cusp area 11 P4: Talonid area 11
M,: Buccolingual crown diameter 11 M,: Mesiodistal crown diameter KO) 17
(Hypoconid)
Permutations
3000 3000 3000 3000 3000 3000
3000 3000 3000 3000 3000 3000 3000 3000 3000 3000
Probability
0.024 0.038 0.049 0.022 0.038 0.036
0.019 0.044
<0.001 0.022
<0.001 0.010 0.020 0.004 0.030 0.040
‘Significance level set a t P rr 0.05. Tests in section (a) were carried out on all variables listed in Table 4; missing data in the P. aff. P. boisei subset reduced the numbers of variables examined to 47, comprising 40 linear or area measurements and seven indices.
of its existence. Others have studied subsets of early hominid tooth crown dimensions, but the hypotheses they tested differed from those in the present analysis and thus the design of the studies also differed. Hunt and Vitzhum (1986) also studied premolar and molar crown size, but their P. boisei sample (EAR) apparently comprised both the early and late subsets which are treated sepa- rately in our own analysis. Wolpoff (1988) made a temporal distinction between his P. boisei subsamples of M1, P4, P4, and M, com- puted occlusal areas, with the time bands being 2.6-1.9 Myr and 1.9-1.4 Myr for the “middle” and “late” spans, respectively. His demonstration that the mean values for P4 and M, occlusal area in the “late” span sub- set exceed those in the middle span sample is not directly comparable to our own find- ings which are based on samples subdivided at 2.3 and not 1.9 Myr.
The results from the present study sug- gest that stasis is the predominant signal coming from the masticatory morphometric data which form the data base for this study. However, although P. boisei sensu stricto is a relatively well-demarcated hominid species it does not follow that the hypodigm is neces- sarily free from problems of sample bias which may have been introduced by tapho- nomic processes. For example, the levels of sexual dimorphism which are inferred for P. boisei sensu stricto are substantial and are
comparable in degree with those observed in the modern gorilla (Wood, 1985; Kimbel and White, 19881, thus the differential preserva- tion of one or other of the sexes could mimic, or even obscure, evidence of directional change. Likewise any trend to either in- crease or decrease body size through time could obscure evidence of directional change. There is, however, no evidence that P. boisei sensu stricto does change in size through time. Its cranial capacity shows no such trend (Brown et al., 1993), nor do the dimen- sions of the mandibular corpus (Wood, 1991; Aiello and Wood, 1994) or those of the major- ity of the postcanine teeth.
Taphonomic processes normally favour the preservation of larger taxa and individu- als and any change in climate or habitat preference could have influenced the way in which the fossils are modified post mortem. One of us has suggested that at least some of the high variability in P. boisei sensu stricto mandible size-a degree of variability, more- over, which has led some workers to question whether this is acceptable within a single species (e.g., Dean, 1988)-may in fact be due to the enlarging effects of matrix infil- tration and the size-reducing effect of abra- sion. Such apparently excessive variation at 2.2,1.9, and 1.7 Myr is seen in Figure 2D, but closer analysis reveals that the four “large” specimens (i.e., L7A-125, KNM-ER 1469, 1468, and 818, reading from left to right)
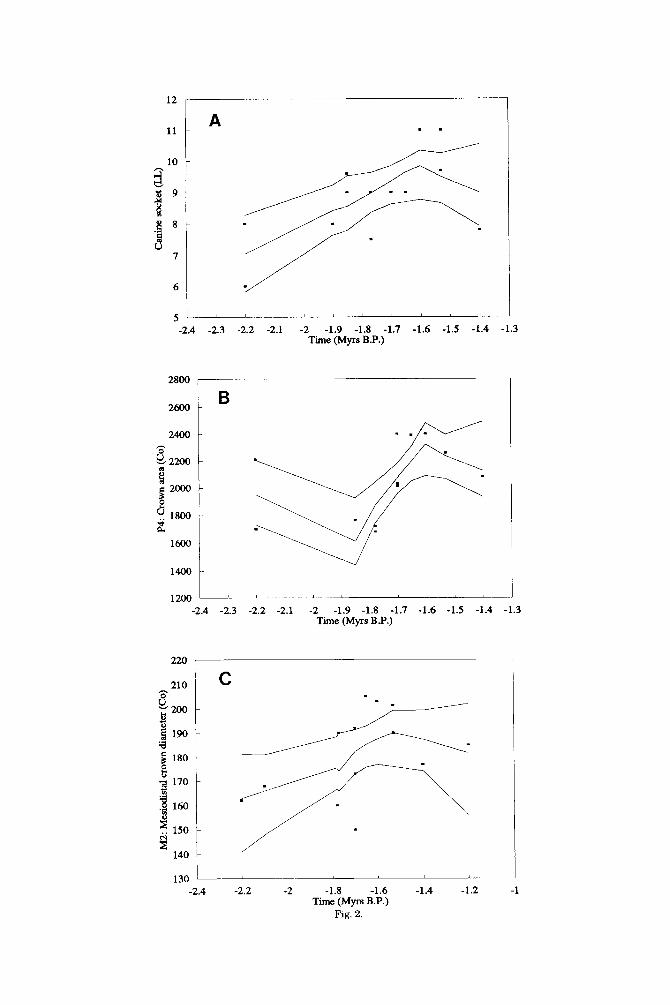
12
11
10
- 9 - s
-2.4 -2.3 -2.2 -2.1 -2 -1.9 -1.8 -1.7 -1.6 -1.5 -1.4 -1.3 Time (Myrs B.P.)
. . A -
-
220 I
C
u 7 -
6 -
-2.4 -2.2 -2 -1.8 -1.6 -1.4 -1.2 -1 Time (Myrs B.P.)
Fig. 2.
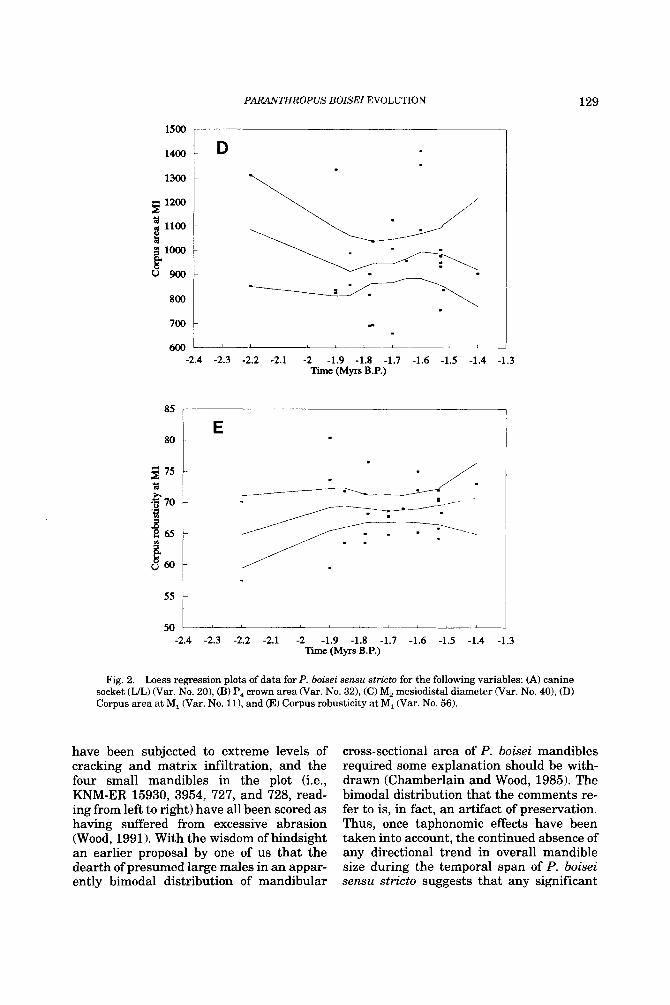
PARANTHROPUS BOISEI EVOLUTION 129
1500
1400 D
Q 800
700
600 -2.4
85
80
g 75 Y
i? ‘G 70
265 f
55
50
L
L I
-2.3 -2.2 -2.1 -2 -1.9 -1.8 -1.7 -1.6 -1.5 -1.4 Time (Myrs B.P.)
E
2
-1.3
-2.4 -2.3 -2.2 -2.1 -2 -1.9 -1.8 -1.7 -1.6 -1.5 -1.4 -1.3 Time (Myrs B.P.)
Fig. 2. Loess regression plots of data for P. boisei sensu stricto for the following variables: (A) canine socket (U) War. No. 20), (B) P, crown area War. No. 321, (C) M, mesiodistal diameter (Var. No. 40), (D) Corpus area at M, (Var. No. 111, and (E) Corpus robusticity at M, War. No. 56).
have been subjected to extreme levels of cracking and matrix infiltration, and the four small mandibles in the plot (i.e., KNM-ER 15930, 3954, 727, and 728, read- ing from left to right) have all been scored as having suffered from excessive abrasion (Wood, 1991). With the wisdom of hindsight an earlier proposal by one of us that the dearth of presumed large males in an appar- ently bimodal distribution of mandibular
cross-sectional area of P. boisei mandibles required some explanation should be with- drawn (Chamberlain and Wood, 1985). The bimodal distribution that the comments re- fer to is, in fact, an artifact of preservation. Thus, once taphonomic effects have been taken into account, the continued absence of any directional trend in overall mandible size during the temporal span of P. boisei sensu stricto suggests that any significant

130 B. WOOD ET AL.
TABLE 6. Nine variables which uary significantly between P. boisei sensu stricto and P. afT. P. boisei using Satterthwaite's modified t'l
~ ~~~
Var
1
9
30
31
32
33
35
37
45
____
TC
ss R ss R ss R ss R ss R ss R ss R ss R ss R
N
13 3
27 2
12 9
12 7
12 8 9 3 8 3
15 2
17 13
Mean SE
49.5 1.3 37.0 1.2 41.7 0.9 35.0 0.3
136.2 3.6 115.0 2.6 150.5 4.6 129.0 4.0
2053.7 81.8 1434.1 82.5 1626.9 83.9 1244.0 108.0 467.9 35.6 350.3 26.5 162.9 2.8 156.5 0.5 193.6 3.7 180.4 4.2
SD
4.6 2.0 4.8 0.4
12.4 7.8
15.9 10.6
283.3 233.3 251.6 187.0 100.6 46.0 10.8 0.7
15.0 15.3
Min
40.0 35.0 30.0 35.0
117.0 103.0 127.0 115.0
1680.0 1058.0 1230.0 1130.0 285.0 301.0 146.0 156.0 172.0 151.0
Max
57.0 39.0 50.0 35.0
157.0 129.0 189.0 150.0
2400.0 1800.0 1910.0 1460.0 560.0 392.0 185.0 157.0 222.0 202.0
cv 9.3 5.4
11.5 1.1 9.1 6.8
10.6 8.2
13.8 16.3 15.5 15.0 21.5 13.1 6.6 0.4 7.7 8.5
t' d f
7.22*** 7
6.73*** 26
4.77*** 18
3.51** 16
5.33*** 17
2.80* 4
2.65* 8
2.26* 14
2.37* 25
'See Table 2 for key to abbreviations used in taxonomic categories (TC) and Table 4 for definitions of variables War). SE, standard error of the mean; SD, standard deviation; CV, coefficient of variation. *P < 0.05; **P < 0.01; ***P < 0.001.
change in overall body size can be dis- counted. This is not to say that the lack of detectable change in corpus cross-sectional area necessarily means that mandibular function was consistent through this time period. Daegling and Grine (1991), for exam- ple, have shown how similarities in mandib- ular cross-sectional area can conceal differ- ences in shape and in biomechanical properties.
These findings of a consistent mandibular morphology across the time range of P. boi- sei sensu stricto are echoed in the cranial evidence which, while it shows evidence of polymorphism (Leakey and Walker, 1988; Wood, 1991; Brown et al., 1993) and sexual dimorphism (Wood, 1991), nonetheless con- forms to a basic pattern. Polymorphism is particularly marked in the arrangement of the ectocranial crests, but is also evident in the face. Previous reports had suggested that P. boisei sensu stricto crania could be recognised because of the apparent empha- sis on the anteriormost fibres of the tempo- ralis, as demonstrated by the size of the sag- ittal crest anteriorly (Kimbel et al., 1984; Walker et al., 1986). However, in several cranial specimens which are otherwise typi- cal of P. boisei sensu stricto (e.g., KNM-ER 406) the crests are highest posteriorly. Simi- lar contrasts are seen in the form of the mar- gins of the nasal aperture which are rounded in OH 5, but sharp in KNM-ER 406. The addition of recently discovered speci-
mens like KNM-ER 13750 (Leakey and Walker, 1988) and KNM-ER 23000 (Brown et al., 1993) t o the P. boisei sensu stricto hypodigm also suggests that yet other fea- tures that were once thought to be relatively invariant in this taxon, like the morphology of the zygomatic arches and supraorbital re- gions (e.g., Rak, 1983), also show a range of expressions (Brown et al., 1993).
Although the dominant signal in P. boisei sensu stricto is stasis, some variables do un- dergo directional change between 2.0 and 1.4 Myr. Apart from those variables involv- ing P,, which reflect the changes already referred to by Suwa (19881, the other changes which occur during this time span are a moderate increase in the dimension of the canine and in the length of the M, crown. However, these two variables, after showing an increase in size between 1.9 and 1.7 Myr, thereafter appear to decrease in size be- tween 1.6 and 1.4 Myr. Whereas taphonomic processes such as abrasion and matrix-infil- tration can result in quite major changes in mandibular size, the crowns of the teeth are less affected by such influences. All the oc- clusal areas in the present study have been corrected for wear. However, the tooth crown area measures which do show change are computed areas; these are both less ac- curate and precise than measured crown ar- eas, but are available for much larger num- bers of teeth. The evidence from the P, and M, of an enlargement of the postcanine

PAR.ANTHR0PUS BOISEI EVOLUTION 131
A S
_- s s >a- s
I 1 I , I I I
-2.8 -2.6 -2.4 -2.2 -2 -1.8 -1.6 -1.4 -1.2 Time (Myrs B.P.)
3000
2500
n 0 2 E 2000 i3 8 1500 4 &
lo00
500
B
S
-2.8 -2.6 -2.4 -2.2 -2 -1.8 -1.6 -1.4 -1.2 Time (Myrs B.P.)
Fig. 3. Loess regression plot of data for the combined EAFROB sample for (A) symphyseal height (Var. No. 1) and for (B) P, crown area (Co) War. No. 32). s = P. boisei sensu stricto; r = P. aff. P. boisei.

132 B. WOOD ET AL.
tooth row coincides with, but of course can- not be causally linked with, evidence of cli- mate change in the Eastern Rift Valley be- tween 1.9 and 1.6 Myr (Hay, 1976; Brown and Feibel, 1988).
East African “robust” australopithecines apparently display two cranial morpholo- gies (Table 1). The early form is poorly rep- resented with just one adult and one juve- nile specimen, the former being KNM-WT 17000, dating from around 2.5 Myr, and the latter L338y-6 from the Shungura Forma- tion, dated to around 2.4 Myr. The oldest evidence for the second morphology dates from around 2.0 Myr and consists of the cal- varia, KNM-ER 13750 with, a little later in time, the palate KNM-ER 405 and the calva- ria KNM-ER 23000 (Brown et al., 1993) around 1.9 Myr, and later still the calvarial fragment KNM-ER 407 at 1.8 Myr. No one who has compared, or reported upon, the two morphologies has failed to record mor- phological, if not functional (e.g., Rak, 1983, 1988), differences between them, but there is no unanimity about the significance of those differences.
Most of the debate about the taxonomic status of these two cranial morphologies has centered on the 40 characters which Skelton et al. (1986) extracted from the studies of White et al. (1981) and Kimbel et al. (1984). Of these, very few are shared exclusively between the early and late cranial morphs (see Table 1) and a considerable number of the character state expressions seen in KNM-WT 17000 are shared with Australo- pithecus afarensis. It is the scale of these primitive retentions, of which an unflexed cranial base and broad anterior tooth crowns are but two examples, which has led many commentators to the conclusion that the early and late morphs are sufficiently different to be treated as distinct species. Unfortunately, the nearly half a million years gap in the cranial fossil record does not allow researchers to use cranial evi- dence to determine whether the distinctive features of the younger of the two cranial morphs accumulated in a gradual, anage- netic, fashion or whether they appeared over a much shorter period of time and thus at a rate compatible with either a cladogenetic origin for P. boisei sensu stricto or even with
the more extreme hypotheses which suggest that the early and late forms may represent separate clades (Skelton and McHenry, 1992; Ramirez-Rozzi, 1993).
In a painstaking study of mandibular pre- molar crown morphology, Suwa (1988) dem- onstrated that the substantial P, talonid of P. boisei sensu stricto makes a relatively abrupt appearance in the fossil record in lower Member G of the Shungura Forma- tion, that is just prior to 2.2 Myr. Suwa (1988: 219) speculated that it would be of interest to “document what other anatomi- cal modification occurred synchronously with premolar specialization.” This study examined 47 mandibular and dental vari- ables and demonstrated that “other anatom- ical modifications” include an increase in the height and breadth of the mandible as well as providing evidence that post 2.2 Myr East African “robust” hominids also have an elongated molar tooth row relative t o the earlier subset. Compared with the scale of the differences between the cranial anatomy of the early and late subsets, the changes in the mandible and teeth, while significant, are much less impressive. It seems that once the hypermasticatory trend is established, by at least 2.5 Myr, subsequent changes to it are focused on the posterior premolars, on lengthening the molar tooth row and on a modest enlargement of the canine.
The results of this analysis of East African “robust” australopithecine mandibular and dental data suggest that stasis is the pre- dominant signal detected within the known time span of the lineage. Small sample sizes prevent a similar investigation into other re- gions of the cranium, but 18 features (marked with an asterisk in Table 1) differ between the early and late subsets, many of which have been cited by workers as evi- dence for retaining the specific distinction between the two subsets. Features such as the anteriorlposterior temporalis propor- tions, degree of squamous temporal pneu- matisation, presence or absence of an asteri- onic notch, basicranial flexion, morphology of the temporomandibular joint, degree of upper facial prognathism, curvature of the nasoalveolar clivus, and the relative propor- tions of the anterior dentition may, when taken together, suggest that there were func-

PAR.ANTHROPUS BOISEI EVOLUTION 133
tional differences in the masticatory appara- tus between P. boisei sensu stricto and P. aff. P. boisei.
Recently, Rak (1988) has noted that there is ‘(tremendous variation’’ with East African “robust” australopithecines as regards the achievement, but not the degree, of the level of mechanical advantage of the masseter on the postcanine dentition, reflected in differ- ences in facial topography among individual specimens. While Rak did not report the val- ues of these indices for the prognathic indi- vidual KNM-WT 17000, measurements taken on a cast suggest that, although the three values which measure the degree of palatal protrusion are similar to those for the two ape taxa and to A. ufricunus, the values for the indices measuring the degree of overlap of the masseter on the palate fall within the range of other (‘robust” australo- pithecines (Wood, 1990). This is interesting when one notes that a similar emphasis on the anterior placement of the temporalis muscle is not seen in KNM-WT 17000, where a clear posterior temporalis predomi- nance is indicated both by the morphology of the sagittal crest (e.g., the minuscule occipi- tal trigon and well-developed compound temporonuchal crest) and by an analysis of its biomechanics. Indeed, with regard to the orientation of the temporalis, KNM-WT 17000 is more similar to prognathic modern taxa which make use of the anterior denti- tion for initial preparation of foodstuffs in the mouth (see Jolly, 1970a,b). Further, the palate of this individual is particularly broad anteriorly (Leakey and Walker, 1988) and may have housed a relatively larger an- terior dentition than that in other “robust” australopithecines (e.g., Suwa, 1989).
The characteristic P. boisei sensu stricto infraorbital morphology was established by 2.5 Myr, while the coopting of the temporalis into this complex lagged behind; this lack of synchronicity in the alteration of mastica- tory muscle biomechanics is an example of mosaic evolution. The differences in tempo- ralis morphology between early and late subsets may be functionally related to the lack of the asterionic notch in the later sub- set and also explain differences noted in the extent and position of the squamous tempo- ral suture between KNM-WT 17000 and P.
boisei sensu stricto. This has been noted by Rak and Kimbel(1991,1993; but see Walker et al., 1993) who point out the similarities between the suture pattern in KNM-WT 17000 and in prognathic modern apes that show posterior temporalis predominance.
It is unfortunate that the mandible of KNM-WT 17000 is not preserved, since the relatively large distance of the temporoman- dibular joint behind the tooth row and its lack of height above the occlusal plane (see Osborn, 1987) are suggestive of a different biomechanical arrangement from that of later specimens and are indicative of an an- gulated mandibular ascending ramus. Leakey and Walker’s (1988) suggestion that KNM-WT 17000 ‘(would have had. . . one of the largest mandibles . . . ever found (p. 23) is in agreement with findings of the present study since both large and small mandibles are present throughout the duration of the lineage (Fig. 2D), but, on the basis of TMJ height and location, one would not predict a substantial depth to the ascending ramus. The two reconstructions by Stewart and Clarke of the OH 5 mandible, housed in the National Museums of Kenya, Nairobi, show how the biomechanics of mastication are af- fected by choice of ramal depth (Wood, in preparation). Height of the TMJ above the occlusal plane affects how the teeth come into occlusion, as does the slope of the artic- ular eminence. This slope is less marked in the earlier subset (Table 1) and indicates serial occlusion (Osborn, 1987) in contrast with the simultaneous approximation of the upper and lower postcanine teeth in the P. boisei sensu stricto sample. The accentua- tion of the Curve of Spee such as is seen in the temporally late Chesowanja individual, KNM-CH 1, may be a further refinement of a trend toward a more simultaneous bite, while an increase in the height of the as- cending ramus serves to equalise bite force along the tooth row (De Wolff-Exalto, 1951a,b; Ward and Molnar, 1980).
While there is now widespread recogni- tion that early hominids are relatively speci- ose, there is little, or no, consensus about the processes which result in new hominid spe- cies. Vrba (1985) reviewed the initiating causes of speciation and came down deci- sively in favour of environmental change as

134 B. WOOD ET AL.
the “motor” of speciation. Competition as an agent for speciation received little support in Vrba’s analysis, nor did it in a more recent discussion of the relationship between com- petition and macroevolution (Masters and Rayner, 1993). The latter paper quoted Coope’s (1979) eminently pragmatic analy- sis of the options which are open to species when faced with climate-induced changes in environments. In those circumstances, he says “species may adapt to the new condi- tions, they may become extinct, or they may move to an area where conditions are still acceptable” (Coope, 1979, p. 262). That pro- cess of adaptation is either manifest as mod- est morphological change within a species group, or more dramatic morphological change the nature andlor degree of which is more sensibly recognised by the allocation of a new species name.
However, Godfrey and Marks (1991) pro- vided a forceful reminder that “biological” speciation and “morphological” speciation do not always go hand-in-hand. Thus, in the eyes of some, even a relatively complete fos- sil record may fail to document all the epi- sodes of “biospeciation.” Tattersall (1986, 1991) has consistently stressed that the numbers of palaeontologically diagnosable primate morphospecies underestimate spe- cies numbers, but the counter argument, that palaeospecies overestimate species numbers, has been equally forcefully put (Templeton, 1989). Godfrey and Marks (1991, p. 60) admit that “the discrimination of biospecies from the fossil record is an in- tractable problem,” but advise that “the goal of fossil species diagnosis should be to ap- proximate neontological morphospecies.” They provide no reliable criteria for recogn- ising a primate or hominid speciation event, but the evident discontinuity in what is oth- erwise a relatively continuous distribution of morphology (Fig. 3) is circumstantial evi- dence for an episode of early hominid specia- tion around 2.3 Myr.
The hypothesis advanced here, that the observed changes in early hominid dental morphology are part of a more widespread sequence of morphological change around 2.2 Myr, results in the prediction that any East African “robust” australopithecine cra- nium discovered in strata older than 2.2 Myr
would resemble KNM-WT 17000 more closely than P. boisei sensu stricto. Likewise we would predict that crania found between 2.2 and 2.0 Myr would, within the limits of the polymorphisms which have been re- ferred to above, possess the autapomorphies which make P. boisei sensu stricto such a distinctive taxon.
CONCLUSION There are differences in the mandibular
corpus and mandibular molar variables be- tween P. aff. P. boisei and P. boisei sensu stricto, but the dental differences are con- centrated in variables that reflect the fea- tures which Suwa (1988) has already identi- fied. Thus, while this study has furnished fresh evidence for a morphological hiatus be- tween P. aff. P. boisei and P. boisei sensu stricto, if the latter species was the product of a cladogenetic event then, at least with respect to the mandible and dentition, that speciation event was mostly an acceleration of a general process of morphological change which continued thereafter. Any further res- olution of this interesting period in the evo- lution of the “robust” australopithecines must await the discovery of fossil evidence from the crucial period between 2.5 and 2.0 Myr so that the tempo and mode of the changes in cranial anatomy can be identi- fied. As yet we do not know whether the differences in cranial anatomy between P. aff. P. boisei and P. boisei sensu stricto (Ta- ble 1) occurred gradually between 2.5 and 2.0 Myr, or whether they were associated with the relatively abrupt changes in man- dibular and dental morphology at around 2.3-2.2 M y . Our findings lead us to suggest that the latter was the more likely course of events. Advances in our understanding of the relationships between early hominid taxa may also come from making novel obser- vations on the evidence that already exists. Recent studies suggest that observations about enamel microstructure (Ramirez- Rozzi, 1993) may help to clarify the relation- ships between P. aff. P. boisei and P. boisei sensu stricto.
ACKNOWLEDGMENTS Research reported in this manuscript was
supported by the Science-Based Archaeol-

PARANTHROPUS BOISE1 EVOLUTION 135
ogy Committee of the NERC and The Lever- hulme Trust. Particular thanks are ex- tended to Richard Leakey and the Trustees of the National Museums of Kenya, and the Government of Kenya, for allowing BAW to make a detailed study of the Koobi Fora re- mains. We thank Gabriele Macho and anon- ymous referees for comments upon the manuscript.
LITERATURE CITED Aiello LC, and Wood BA (1994) Cranial variables as
predictors of hominid body mass. 95:(in press). Arambourg C, and Coppens Y (1968) Decouverte d u n
Australopithecien nouveau dans le gisement de YOmo (Ethiopie) en 1967. S. Af. J. Sci. 64:58-59.
Bown TM, and Rose KD (1987) Patterns of dental evolu- tion in early Eocene Anaptomorphine primates (Omo- myidae) from the Bighorn Basin, Wyoming. J . Paleon-
Brown B, Walker A, Ward CV, and Leakey RE (1993) New Australopithecus boisei calvaria from East Lake Turkana, Kenya. Am. J . Phys. Anthropol. 91t137-159.
Brown FH, and Feibel CS (1988) “Robust” hominids and Plio-Pleistocene paleogeography of the Turkana Ba- sin, Kenya. In FE Grine (ed.): Evolutionary History of the “Robust” Australopithecines. New York: Aldine de Gruyter, pp. 325-341.
Chamberlain AT, and Wood BA (1985) A reappraisal of variation in hominid mandibular corpus dimensions. Am. J . Phys. Anthropol. 66r399-405.
Chamberlain AT, and Wood BA (1987) Early hominid phylogeny. J . Hum. Evol. 16:119-133.
Cleveland WS (1979) Robust locally weighted regression and smoothing scatterplots. J . Am. Stat. Assoc. 74: 829-836.
Cleveland WS, and Devlin SJ (1988) Locally weighted regression: An approach t o regression analysis by lo- cal fitting. J . Am. Stat. Assoc. 83:59&610.
Cleveland WS, Devlin SJ, and Grosse (1988) Regression by local fitting: Methods, properties, and computa- tional algorithms. J . Economet. 37r87-114.
Connor EF (1986) Time series analysis of the fossil record. In DM Raup and D Jablonski (eds.): Patterns and Processes in the History of Life. Berlin: Springer- Verlag, pp. 119-147.
Coope GR (1979) Late Cenozoic Coleoptera: Evolution, biogeography, and ecology. Annu. Rev. Ecol. Syst. 10: 247-267.
Coppens Y (1979) Les hominides du Pliocene et du Pleis- tocene de la Rift Valley. Bull. SOC. Geol. Fr. 21:313- 320.
Cronin JE, Boaz NT, Springer CB, and Rak Y (1981) Tempo and mode in hominid evolution. Nature (Lon- don) 292t113-122.
Daegling DJ, and Grine FE (1991) Compact bone distri- bution and biomechanics of early hominid mandibles. Am. J. Phys. Anthropol. 86:321-339.
Dean MC (1988) Growth of teeth and development of the dentition in Paranthropus. In FE Grine (ed.): Evolu- tionary History of the “Robust” Australopithecines. New York: Aldine de Gruyter, pp. 43-53.
tol. 61:1-162.
Delson E (1986) Human phylogeny revised again. Na- ture (London) 322: 49-97,
De Wolff-Exalto EA (1951a) On the differences in the lower jaw of animalivorous and herbivorous mam- mals. I. Proc. Kon. Ned. Akad. Weten. (C) 54237-246.
De Wolff-Exalto EA (1951b) On the differences in the lower jaw of animalivorous and herbivorous mam- mals. 11. Proc. Kon. Ned. Akad. Weten. (C) 54r405- 410.
Efron B, and Tibshirani R 11991) Statistical data analy- sis in the computer age. Science 253:390-395.
Eubank RL (1988) Spline Smoothing and Nonparame- tric Regression. New York: Marcel Dekker.
Feibel CS, Brown FH, and McDougall I (1989) Strati- graphic context of fossil hominids from the Omo Group deposits: Northern Turkana Basin, Kenya and Ethiopia. Am. J . Phys. Anthropol. 78595-622.
Godfrey L, and Marks J (1991) The nature and origins of primate species. Yb. Phys. Anthropol. 34t39-68.
Gould SJ, and Eldredge N (1977) Punctuated equilibria: the tempo and mode of evolution considered. Paleobi- ology 3:115-151.
Hay RL (1976) Geology of the Olduvai Gorge. A Study of Sedimentation in a Semiarid Basin. Los Angeles: Uni- versity of California Press.
Holloway RL (1988) “Robust” australopithecine brain endocasts: some preliminary observations. In FE Grine (ed.): Evolutionary History of the “Robust” Aus- tralopithecines. New York: Aldine de Gruyter, pp. 97- 105.
Hubert W, Golledge RG, Constanzo CM, and Gale N (1985) Tests of randomness: Unidimensional and mul- tidimensional. Environ. Planning 17r373-385.
Hunt K, and Vitzhum VJ (1986) Dental metric assess- ment of the Omo fossils: Implications for the phyloge- netic position of Australopithecus africanus. Am. J. Phys. Anthropol. 71:141-155.
Jolly CJ (1970a) The large African monkeys as an adap- tive array. In JR Napier and PH Napier (eds.): Old World Monkeys. New York: Academic Press, pp. 141- 174.
Jolly CJ (1970b) The seed-eaters: A new model of homi- nid differentiation based on a baboon analogy. Man 5:5-28.
Kimbel WH, and White TD (1988) Variation, sexual di- morphism and the taxonomy of Australopithecus. In FE Grine (ed.): Evolutionary History of the “Robust” Australopithecines. New York Aldine de Gruyter, pp. 175-192.
Kimbel WH, White TD, and Johanson DC (1984) Cra- nial morphology ofAustralopithecus afarensis: A com- parative study based on a composite reconstruction of the adult skull. Am. J . Phys. Anthropol. 64r337-388.
Kimbel WH, White TD, and Johanson DC (1988) Impli- cations of KNM-WT 17000 for the evolution of “ro- bust” Australopithecus. In FE Grine (ed.): Evolution- ary History of the “Robust” Australopithecines. New York: Aldine de Gruyter, pp. 259-268.
Konigsberg LW (1990) Temporal aspects of biological distance: serial correlation and trend in a prehistoric skeletal lineage. Am. J . Phys. Anthropol. 82t45-52.
Leakey REF, and Walker AC (1988) New Australopithe- cus boisei specimens from East and West Lake Tur- kana, Kenya. Am J. Phys. Anthropol. 76:l-24.

136 B. WOOD ET AL.
Leigh SR (1992) Cranial capacity evolution in Homo erectus and early Homo supiens. Am. J . Phys. Anthro-
Manly BFJ (1991) Randomization and Monte Carlo Methods in Biology. London: Chapman and Hall.
Masters JC, and Rayner FLJ (1993) Competition and macroevolution: The ghost of competition yet to come? Biol. J . Linn. SOC. 49:87-98.
Osborn JW (1987) Relationship between the mandibular condyle and the occlusal plane during hominid evolu- tion: Some of its effects on jaw mechanics. Am. J . Phys. Anthropol. 73:193-207.
Rak Y (1983) The Australopithecine Face. New York Academic Press.
Rak Y (1988) On variation in the masticatory system of Australopithecus boisei. In FE Grine (ed.): Evolution- ary History of the “Robust” Australopithecines. New York Aldine de Gruyter, pp. 193-198.
Rak Y, and Kimbel WH (1991) On the squamosal suture of KNM-WT 17000. Am. J. Phys. Anthropol. 81:l-6.
Rak Y, and Kimbel WH (1993) Reply to Drs. Walker, Brown, and Ward. Am. J . Phys. Anthropol. 90:50€- 507.
Ramirez-Rozzi FV (1993) Tooth development in East African Purunthropus. J . Hum. Evol. 24:429-454.
Rightmire GP (1981) Patterns in the evolution ofHomo erectus. Paleobiology 7t241-246.
Rightmire GP (1990) The Evolution of Homo erectus. Comparative Anatomical Studies of an Extinct Hu- man Species. Cambridge: Cambridge University Press.
Skelton RR, and McHenry HM (1992) Evolutionary rela- tionships among early hominids. J . Hum. Evol. 23: 309-439.
Skelton RR, McHenry HM, and Drawhorn GM (1986) Phylogenetic analysis of early hominids. Cum. An- thropol. 27:2143.
Snedecor GW, and Cochran, WG (1980) Statistical Methods, 7th ed. Ames: Iowa State University Press.
Suwa G (1988) Evolution of the “robust” australopithe- cines in the Omo succession: Evidence from mandibu- lar premolar morphology. In FE Grine (ed.): Evolu- tionary History of the “Robust” Australopithecines. New York: Aldine de Gruyter, pp. 199-222.
Suwa G (1989) The premolar of KNM-WT 17000 and relative anterior to posterior dental size. J . Hum.
Tattersall I (1986) Species recognition in human palae- ontology. J . Hum. Evol. 15:165-175.
Tattersall I(1991) What was the human revolution? J. Hum. Evol. 20:77-83.
Templeton AR (1989) The meaning of species and speci- ation: a genetic perspective. In D Otte and JA Endler
pol. 87:1-13.
Evol. 18t795-799.
(eds.): Speciation and Its Consequences. Sunderland, MA: Sinauer Associates, pp. 3-27.
Vrba ES (1985) Environment and evolution: Alternative causes of the temporal distribution of evolutionary events. S. Afr. J. Sci. 81:229-236.
Walker AC, Brown B, and Ward SC (1993) Squamosal suture of cranium KNM-WT 17000. Am. J . Phys. An- thropol. 90:501-505.
Walker AC, and Leakey REF (1988) The evolution of Austrulopithecus boisei. In FE Grine (ed.): Evolution- ary History of the “Robust” Australopithecines. New York Aldine de Gruyter, pp. 247-258.
Walker AC, Leakey REF, Harris JM, and Brown FH (1986) 2.5 Myr Austrulopithecus boisei from west of Lake Turkana, Kenya. Nature (London) 322517- 522.
Ward SC, and Molnar S (1980) Experimental stress analysis of topographic diversity in early hominid gnathic morphology. Am. J . Phys. Anthropol. 53:383- 393.
White TD (1988) The comparative biology of ‘robust’ Australopithecus: Clues from context. In FE Grine (ed.): Evolutionary History of the “Robust” Australop- ithecines. New York: Aldine de Gruyter, pp. 44M83.
White TD, Johanson DC, and Kimbel WH (1981) Aus- trulopithecus ufricunus: Its phylogenetic position re- considered. S. Afr. J . Sci. 77:445-470.
Wolpoff MH (1984) Evolution in Homo erectus: A ques- tion of stasis. Paleobiology 10t389-406.
Wolpoff MH (1988) Divergence between early hominid lineages: the roles of competition and culture. In FE Grine (ed.): Evolutionary History of the “Robust” Aus- tralopithecines. New York: Aldine de Gruyter, pp. 485-497.
Wood BA (1985) Sexual dimorphism in the hominid fos- sil record. In J Ghesquiere, RD Martin, and F New- combe (eds.): Human Sexual Dimorphism. London: Taylor and Francis, pp. 105-123.
Wood BA (1991) Koobi Fora Research Project. Volume 4. Hominid Cranial Remains. Oxford: Clarendon Press.
Wood BA, and Chamberlain AT (1987) The nature and affinities of the ‘robust’ australopithecines: a review. J. Hum. Evol. 16t625-641.
Wood BA, and Stack CG (1980) Does allometry explain the differences between ‘gracile’ and ‘robust’ australo- pithecines? Am. J . Phys. Anthropol. 52:55-62.
Wood CG (1990) The masticatory morphology of the ‘ro- bust’ australopithecines: an investigation into adap- tation using Paradigm Analysis. Unpublished Ph.D. thesis, The University of Liverpool.
Wood CG (in preparation) Masticatory functional mor- phology in ‘robust’ australopithecine specimens OH 5 and KNM-ER 406. J. Hum. Evol. (1994).