original articlesjpp.krakow.pl/journal/archive/02_13/pdf/11_02_13_article.pdfcells are connected by...
TRANSCRIPT

INTRODUCTION
Cells are connected by several types of junctions includingtight junctions (1), gap junctions (2), desmosomes and adherensjunctions (3). Among these, tight junctions are intimatelyadherent junction between the cells and apically located in thecells. Tight junctions are known to play numerous roles includingsecuring adjacent cells, forming barriers against extracellularfluids and materials, and acting as paracellular transporters (4).Tight junctions are mainly composed of three types of proteinssuch as transmembrane, cytoskeletal, and cytoplamic plaqueproteins (5). Transmembrane proteins penetrate into the cellularmembrane, restrict the passage of certain substances, and areresponsible for tight junction permeability and paracellulartransport (6). Occludin, claudins, and junction adhesionmolecules (JAM)s are typical transmembrane proteins (7).
Occludin is tetraspan transmembrane protein with two loopsthat protrude into the intercellular spaces, and interact with thePDZ domain of cytoplasmic plaque components such as the ZO-1/ZO-2 complex (8, 9). The expression of occludin has beenfound in various tissues including the digestive tract, kidney,urinary bladder, skin, liver, and blood vessels (10, 11). Occludin
is known as be involved in determining paracellular permeabilityby sealing the tight junctions. Toxins or pathogens includingcholera enterotoxin (12), enteropathogeinc E. coli (13), C.difficile toxin (14), and rotaviruses (15) are known to target tightjunction components such as occludin and cause diarrhea.Decreased expression of occludin has been observed in cancer ortumors, and it has been reported that oncogenes including SV40are involved in breaking down tight junctions (16, 17). Abnormalexpressions of occludin with other tight junction proteins are alsoassociated with allergies (18), and diabetes (19).
JAMs are also classified as transmembrane proteins.However, unlike occludin they have only one trans-membraneloop. JAMs are members of the immunoglobulin subfamily andare closely associated with the immune system. JAM-A is animmunoglobulin-like molecule found in tight junctions for thefirst time (20). Expression of JAM-A has been observed invarious types of blood cells such as monocytes, neutrophils, andlymphocytes (21, 22). Since JAM-A is related with the immuneresponse, there are many physiologic disorders and diseasesassociated with unbalanced JAM-A expression. These includethrombocytopenia (23), thrombosis (24), vascular diseases (25),and virus infection. In JAM-A null mice, carcinoma
JOURNAL OF PHYSIOLOGY AND PHARMACOLOGY 2013, 64, 1, 11-18www.jpp.krakow.pl
Original articles
I. HWANG1, B.S. AN1,2, H. YANG1, H.S. KANG1, E.M. JUNG1, E.B. JEUNG1
TISSUE-SPECIFIC EXPRESSION OF OCCLUDIN, ZONA OCCLUDENS-1, AND JUNCTIONADHESION MOLECULE A IN THE DUODENUM, IELUM, COLON, KIDNEY, LIVER, LUNG,
BRAIN, AND SKELETAL MUSCLE OF C57BL MICE
1Laboratory of Veterinary Biochemistry and Molecular Biology, College of Veterinary Medicine, Chungbuk National University,Cheongju, Chungbuk, 361-763 Republic of Korea; 2Department of Biomaterials Science, College of National Resources & Life
Science, Pusan National University, Republic of Korea
Tight junctions are the most apically positioned intercellular junction and play many roles such as securing adjacentcells, forming barriers from extracellular materials, and facilitating paracellular transport. Occludin and junctionadhesion molecule A (JAM-A) are classified as transmembrane proteins that are directly involved in paracellulartransport. Zona occludens-1 (ZO-1) is a protein that contains a PDZ domain which forms a binding site for other tightjunction proteins. In this study, we assessed the differential expression of these tight junction components in variousmouse organs including the intestine (duodenum, ileum, and colon), kidney, liver, lung, brain, and skeletal muscle. Real-time PCR and Western blot assays were performed to measure the gene and protein expression of occludin, JAM-A, andZO-1. Similar levels of occludin gene expression were detected in all tissues except for skeletal muscle in whichoccludin expression was not found. The JAM-A and ZO-1 genes were highly expressed in all the tested tissues.Localization of occludin, JAM-A, and ZO-1 was determined by immunohistochemistry. These proteins were detected inthe intercellular apical junctions in each tissue except for occludin (which was not observed in skeletal muscle). Theseimmunostaining data were consistent with the gene expression profiles we obtained. Our results suggest that occludin,JAM-A, and ZO-1 genes are normally expressed in the intestine, kidney, liver, lung, and brain indicating that thesefactors may be essential for maintaining appropriate physiological concentration of ions, solutes and water.
K e y w o r d s : junction adhesion molecule, occludin, zona occludens-1, tissue-specific expression, tight junction protein 1,duodenum, skeletal muscle

development is inhibited through increased cellular apoptosis(26). In addition, many other studies have revealed existence ofJAM-A in tissues including brain, trachea, lung, liver, pancreas,and kidney (20, 21).
ZO-1 (also known as tight junction protein 1) is acytoplasmic plaque protein. This protein interacts withtransmembrane proteins through a PDZ domain (27). Severalstudies have demonstrated that down-regulation of ZO-1 iscorrelated with cancer and tumor development (28). A previousknock-down study showed that decreased expression of ZO-1induces delayed tight junction recruitment (29), and this proteinis essential for embryonic development (30). Several studies alsodemonstrated that down-regulation of ZO-1 is correlated withcancer and tumor development (28). Expression of ZO-1 hasbeen reported in numerous organs of human and mice; kidney,intestine, liver, lung, and brain (31-33).
The regulation and expression of occludin, JAM-A, and ZO-1, have been examined in many studies under various conditions.Although these proteins are expressed and regulated in a tissuespecific manner, comparison of expressions of these genes in thetissues under identical condition has not been studied.
Since the tight junction proteins are closely correlated andfunction concurrently, in this study, we profiled the tissuedistribution of occludin, JAM-A, and ZO-1 in the duodenum,ileum, colon, kidney, liver, lung, brain, and skeletal muscle inmice at the transcriptional and translational levels. Furthermore,localization of these genes in the tissues was also verified byimmunohistochemistry.
MATERIALS AND METHODS
Animals and tissues
Male C57BL/6 mice (9~10 weeks) were obtained fromKOATECH (Pyeongtaek-si, Gyeonggi-do, Republic of Korea).All mice were euthanized by ether and tissue samples from theduodenum, ileum, colon, kidney, liver, lung, brain, and skeletalmuscle of the hind leg were obtained. These samples were usedfor real-time PCR, Western blotting, and immunohistochemistryanalysis.
RNA extraction and quantitative real-time PCR
Total RNA was extracted with Trizol reagent (Ambion,Austin, TX, USA) according to the manufacturer's instructions.The total RNA concentration was measured at 260 nm ofabsorbance by EPOCH (BioTeK, Vermont, USA). First strandcomplement DNA (cDNA) was synthesized from 1 µg of totalRNA by reverse transcription using Moloney murine leukemiavirus (MMLV) reverse transcriptase (1:1000, Invitrogen Co.,Carlsbad, CA, USA), and random primers (9-mers; TaKaRa BioInc., Otsu, Shiga, Japan). Reverse transcription (RT) PCR wasperformed with a 7300 real-time PCR system (AppliedBiosystems, Foster City, CA, USA) according to themanufacturer's instructions. β-actin was used for normalizationand the relative gene expression levels were quantified using RQsoftware (Applied Biosystems). The primer sequences foroccludin were: 5'-ACT GGG TCA GGG AAT ATC CA-3' (sense)and 5'-TCA GCA GCA GCC ATG TAC TC-3' (antisense). Theprimer sequences for JAM-A were: 5'-CTG ATC TTT GAC CCCGTG AC-3' (sense) and 5'-ACC AGA CGC CAAAAA TCAAG-3' (antisense). The primer sequences for ZO-1 were: 5'-ACT CCCACT TCC CCAAAAAC-3' (sense) and 5'-CCA CAG CTG AAGGAC TCA CA-3' (antisense). The primer sequences for β-actinwere: 5'-ACA GGC ATT GTG ATG GAC TC-3' (sense), and 5'-ATT TCC CTC TCA GCT GTG GT-3' (antisense).
Quantitative real-time PCR reactions contained 1 µL cDNAtemplate with 10 µL of 2 × SYBR Premix Ex Taq (TaKaRa BioInc.) and 10 pmol of the specific primers. Each reaction wascarried out for 40 cycles of denaturation at 95°C for 15 s,annealing at 62°C for 15 s, and extension at 72°C for 30 s. Atthe end of the extension phase of each cycle, fluorescenceintensity was measured. Threshold values for the fluorescenceintensity of the samples were manually set. The cycle at whichsample fluorescence reached the threshold, was defined as thethreshold cycle (CT). β-actin was used as the control fornormalization.
Western blot analysis
Proteins were extracted with RIPA buffer [(50 mM Tris-HCl (AmrescoInc, Solon, OH, USA), 1% NP-40 (Calbiochem,Germany), 150 mM NaCl (DuchefaBiochemie, Haarlem, TheNetherlands), 1 mM EDTA (Boehringer Mannheim Germany),and 2 protease inhibitor cocktail tablets (Roche Diagnostics,Germany) for 50 ml of total RIPA solution] and homogenized.After centrifugation at 14,000 rpm, each protein (20 µg perlane) were separated in 7.5~10% SDS-PAGE gels andtransferred to nitrocellulose membranes (Millipore, Bedford,MA, USA). The membranes were blocked with 5% skim milkthat is resolved in Tris-Buffered Salined Tween-20 (TBS-T) for2 hours at room temperature and then incubated overnight withthe following primary antibodies at 4°C: rabbit anti-occludin(1:1000, Invitrogen Co.), rabbit anti-ZO-1 (1:1000, InvitrogenCo.), rabbit anti-JAM-A (1:1000, Santa Cruz, California,USA), or rabbit anti-β-actin (1:1000, Santa Cruz) for O/N at4°C. Next, the membranes were washed with TBS-T for 1 hourat room temperature, and incubated with anti-rabbithorseradish peroxidase-conjugated secondary antibodies(1:3000, Santa Cruz) for 2 hours at room temperature. Afterwashing with TBS-T again, antibody binding was detectedwith enhanced chemiluminescence reagent (AmershamBiosciences, Little Chalfont, UK) and exposed to Biomaxtm
Light film (Kodak Express, London, United Kingdom) for 1 to20 min. To ensure signal specificity, the membranes werelabeled with secondary antibody in the absence of primaryantibody.
Immunohistochemistry
Tissue-specific localization of tight junction proteins wasexamined using immunohistochemistry. Samples of duodenum,ileum, colon, kidney, liver, lung, brain, and muscle wereembedded in paraffin, cut into 5-µm sections, mounted on slidesglasses (Matunami, Ishikawa, Japan) which are coated withamino silane (APS), deparaffinized using xylene, and they werehydrated in descending graded ethanol solutions. Endogenousperoxidase activity was blocked with 3% hydrogen peroxidasein PBS for 30 min at room temperature. To remove non-specificreaction, the sections were incubated with 10% goat serum(Vecter Laboratories, Burlingame, CA, USA) in PBS for 1 hourat room temperature. After washing using TBS-T, the sectionswere incubated with primary antibodies which were used inWestern blotting (rabbit anti-occludin, rabbit anti-JAM-A andrabbit anti-ZO-1, 1:250) for O/N at room temperature. After that,the sections were washed with TBS-T again. The sections werereacted with biotinylated secondary antibodies (1:500, RabbitIgG; Vector Laboratories, Inc.) for 1 hour at 37°C followed byincubation with ABC-Elite (Vector Laboratories, Inc.) for 30min at 37°C. Diaminobezidine (Sigma, St Louis, MO, USA) wasused as a chromogen. The sections were then counterstainedwith hematoxylin and mounted in Cytoseal*60 (Richard-AllanScientific Co., Kalamazoo, MI, USA).
12
RESULTS
Tissue distribution and location of occludin
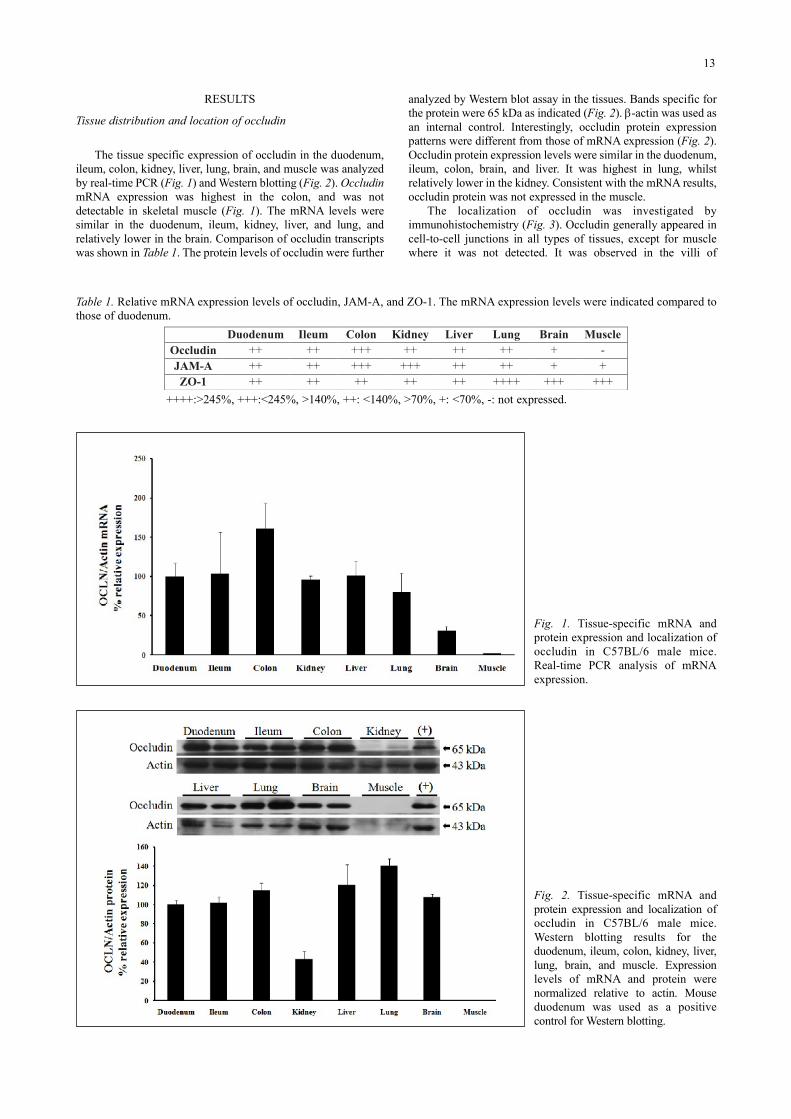
The tissue specific expression of occludin in the duodenum,ileum, colon, kidney, liver, lung, brain, and muscle was analyzedby real-time PCR (Fig. 1) and Western blotting (Fig. 2). OccludinmRNA expression was highest in the colon, and was notdetectable in skeletal muscle (Fig. 1). The mRNA levels weresimilar in the duodenum, ileum, kidney, liver, and lung, andrelatively lower in the brain. Comparison of occludin transcriptswas shown in Table 1. The protein levels of occludin were further
analyzed by Western blot assay in the tissues. Bands specific forthe protein were 65 kDa as indicated (Fig. 2). β-actin was used asan internal control. Interestingly, occludin protein expressionpatterns were different from those of mRNA expression (Fig. 2).Occludin protein expression levels were similar in the duodenum,ileum, colon, brain, and liver. It was highest in lung, whilstrelatively lower in the kidney. Consistent with the mRNA results,occludin protein was not expressed in the muscle.
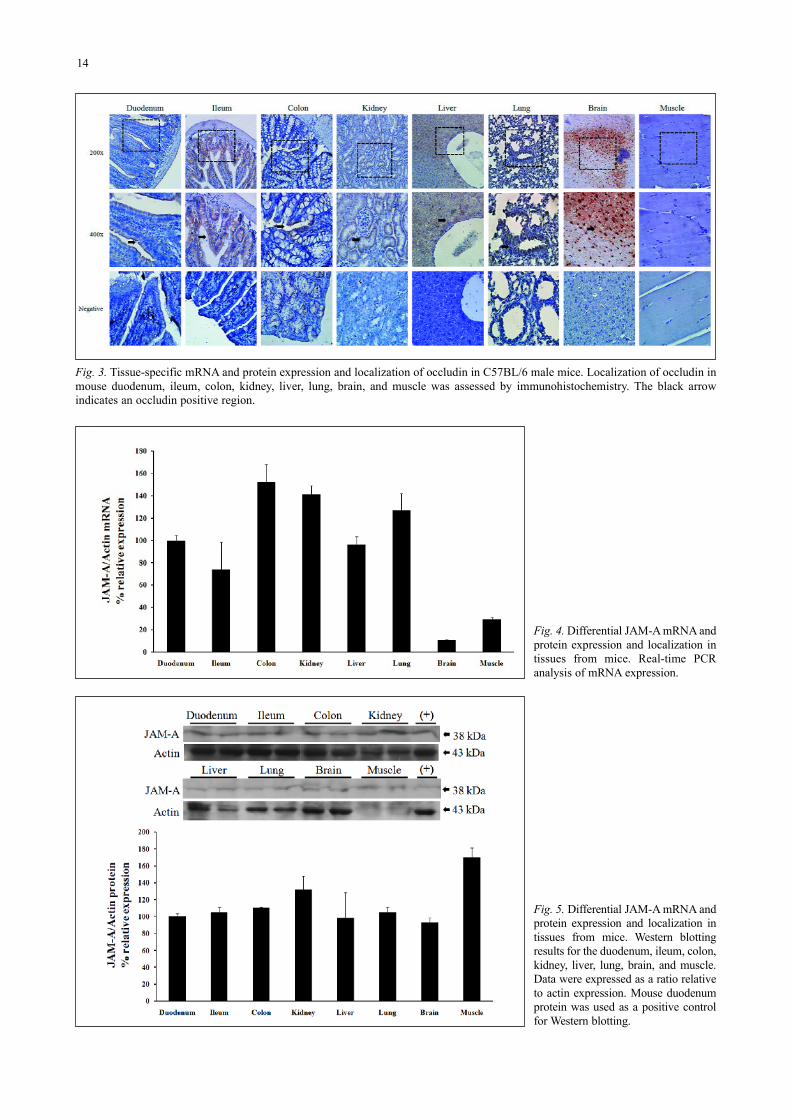
The localization of occludin was investigated byimmunohistochemistry (Fig. 3). Occludin generally appeared incell-to-cell junctions in all types of tissues, except for musclewhere it was not detected. It was observed in the villi of
13
Duodenum Ileum Colon Kidney Liver Lung Brain Muscle Occludin ++ ++ +++ ++ ++ ++ + - JAM-A ++ ++ +++ +++ ++ ++ + + ZO-1 ++ ++ ++ ++ ++ ++++ +++ +++
++++:>245%, +++:<245%, >140%, ++: <140%, >70%, +: <70%, -: not expressed.
Table 1. Relative mRNA expression levels of occludin, JAM-A, and ZO-1. The mRNA expression levels were indicated compared tothose of duodenum.
Fig. 1. Tissue-specific mRNA andprotein expression and localization ofoccludin in C57BL/6 male mice.Real-time PCR analysis of mRNAexpression.
Fig. 2. Tissue-specific mRNA andprotein expression and localization ofoccludin in C57BL/6 male mice.Western blotting results for theduodenum, ileum, colon, kidney, liver,lung, brain, and muscle. Expressionlevels of mRNA and protein werenormalized relative to actin. Mouseduodenum was used as a positivecontrol for Western blotting.
14
Fig. 3. Tissue-specific mRNA and protein expression and localization of occludin in C57BL/6 male mice. Localization of occludin inmouse duodenum, ileum, colon, kidney, liver, lung, brain, and muscle was assessed by immunohistochemistry. The black arrowindicates an occludin positive region.
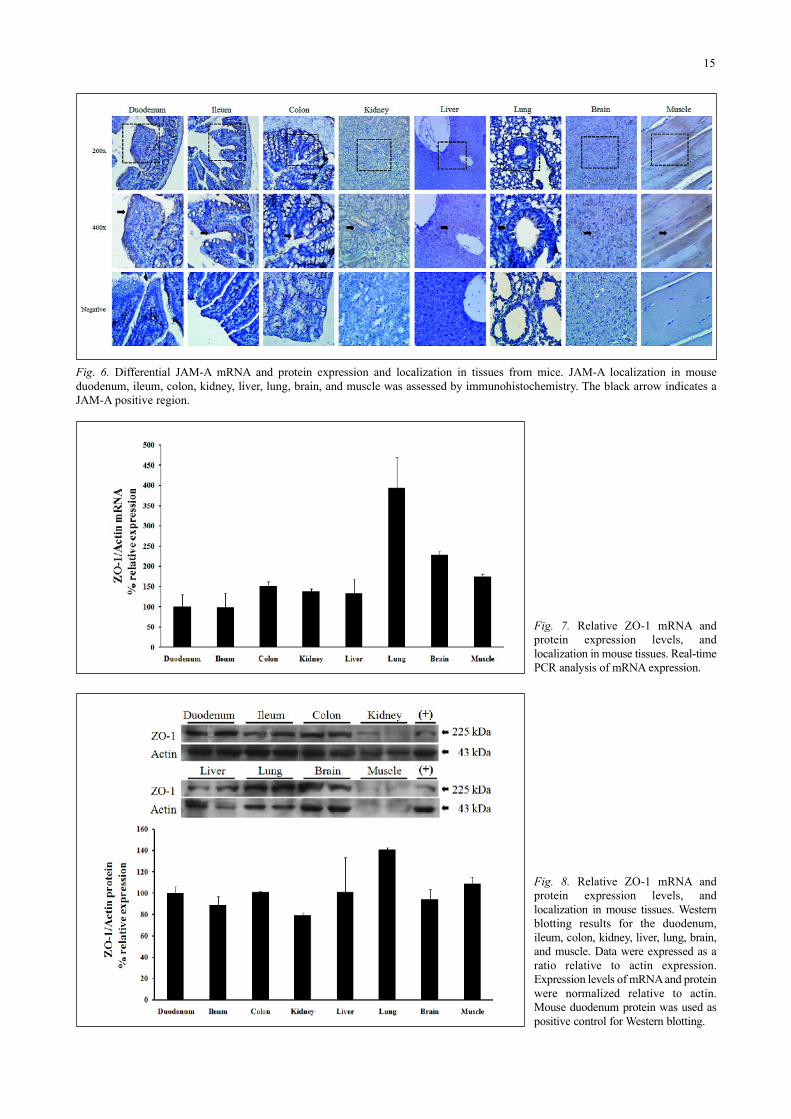
Fig. 4. Differential JAM-A mRNA andprotein expression and localization intissues from mice. Real-time PCRanalysis of mRNA expression.
Fig. 5. Differential JAM-A mRNA andprotein expression and localization intissues from mice. Western blottingresults for the duodenum, ileum, colon,kidney, liver, lung, brain, and muscle.Data were expressed as a ratio relativeto actin expression. Mouse duodenumprotein was used as a positive controlfor Western blotting.
15
Fig. 6. Differential JAM-A mRNA and protein expression and localization in tissues from mice. JAM-A localization in mouseduodenum, ileum, colon, kidney, liver, lung, brain, and muscle was assessed by immunohistochemistry. The black arrow indicates aJAM-A positive region.
Fig. 7. Relative ZO-1 mRNA andprotein expression levels, andlocalization in mouse tissues. Real-timePCR analysis of mRNA expression.
Fig. 8. Relative ZO-1 mRNA andprotein expression levels, andlocalization in mouse tissues. Westernblotting results for the duodenum,ileum, colon, kidney, liver, lung, brain,and muscle. Data were expressed as aratio relative to actin expression.Expression levels of mRNA and proteinwere normalized relative to actin.Mouse duodenum protein was used aspositive control for Western blotting.

enterocytes in the duodenum and ileum. In the colon and kidney,this protein was highly expressed in colonic mucosa, and renaltubules. Occludin was also highly expressed in hepatocytes,apical cell borders of the bronchial epithelium, endothelium ofthe lung, and epithelial and endothelial cells of the brain.
Tissue distribution and location of JAM-A
JAM-A gene expression was also examined by real-timePCR and Western blotting in the same tissues (Figs. 4-6). ThemRNA levels of JAM-A were higher in the colon, kidney thanthe other tissues (Fig. 4). JAM-A was moderately expressed inthe duodenum, ileum, liver, and lung while lower levels wereobserved in the brain and muscle. The relative mRNA levels ineach tissue were presented in Table 1. Expression levels of JAM-A protein were different those of mRNA (Fig. 5). The relativeexpression levels of JAM-A in the muscle were highest while itwas similar in other tissues.
JAM-A localization was analyzed by immunohistochemistry(Fig. 6). Immunochemical signals corresponding to this proteinwere observed in all the tissues. In general, JAM-A existed in theepithelial cells of the tested tissues. Expression of this proteinwas detected in the same region as occludin (Fig. 3).
Tissue distribution and location of ZO-1
ZO-1 mRNA and protein expression was investigated byreal-time PCR and Western blotting (Figs. 7-9). The ZO-1 geneexpression was detected in all the tested organs. The mRNAlevels were similar in all organs except for the lung, brain, andmuscle (Fig. 7). Expression of ZO-1 was highest in the lung, andrelatively higher in the brain, and muscle compared to the othertissues. The expression pattern of ZO-1 protein in the differenttissues was similar to that of mRNA (Figs. 7 and 8). The relativeZO-1 mRNA expression levels are presented in Table 1.
ZO-1 localization determined by immunohistochemistry wassimilar to that of occludin and JAM-A (Fig. 9).
DISCUSSION
Tight junctions have been studied in various fields due totheir involvement in many physiological functions including
immune reactions, barrier formation, and paracellular passivetransport. Therefore, investigating the tissue-specificdistribution of tight junction gene expression is valuable forelucidating the functions of these proteins in various organs. Inthe present study, we selected eight organs in mice (theduodenum, ileum, colon, kidney, liver, lung, brain, and skeletalmuscle from the hind leg), and examined the expression andlocalization of two transmembrane proteins (occludin, and JAM-A) and one cytoplasmic plaque protein (ZO-1).
We found that, occludin expression levels were relativelysimilar among all the tested tissues except for skeletal muscle. Aprevious study also determined that occludin is not expressed inskeletal muscle (34). Except the kidney, the expression of mRNAand protein was consistent in the organs. In the kidney, the proteinexpression levels were lower compare to those of mRNA. It ispossible that occludin protein is modulated via post-transcriptonalor post-translational regulations. In other studies, occludin wasknown to have various post-transcriptional and post-translationalmodifications in diverse circumstances by proteolyticdegradation, phosphorylation and ubiquitation (35). The occludinexpression patterns we observed were concurred with the findingsof previous studies. For example, Claude and Goodenoughidentified the occludin expression in the distal tubules of kidney(36). In our study, the occludin was expressed in all tubules of thekidney. In addition, we confirmed occludin expression inepithelial cells and endothelium of the brain. Occludin is acomponent of tight junction in the brain endothelium and theblood-brain barrier (BBB), and may control the selective transferof ions to the brain (11, 34). Intestinal occludin regulatesmacromolecule flux by controlling the intestinal epithelial tightjunction barrier (37). We observed expression of occludin inenterocytes of the small and large intestine in the present study.The expression of occludin was also found in the liver and lung.Occludin is involved in hepatic regeneration, and function as anair-blood barrier in the lung (38-40).
JAM-A is a single-pass transmembrane protein and alsoknown as F11r. Similar to other transmembrane proteins, JAM-Aalso seals the intercellular space, and acts as a physical barrierthat restricts free paracellular movement of solutes and water.This protein also acts as a receptor in immune reactions againstretroviral infection (41) or in platelets (42), and also regulatesmonocyte transmigration (20). JAM-A is responsible forregulating tight junction assembly in the epithelium, which may
16
Fig. 9. Relative ZO-1 mRNA and protein expression levels, and localization in mouse tissues. Localization of ZO-1in mouseduodenum, ileum, colon, kidney, liver, lung, brain, and muscle was assessed by immunohistochemistry. The black arrow indicates aZO-1 positive region.

be accomplished by direct or indirect interaction with occludin(22). In our experiment, the mRNA expression levels of JAM-Ain the intestine, kidney, liver, and lung were similar, while theexpression was lower in the brain and skeletal muscle. Proteinexpression levels were concurred with those of mRNA except thebrain and muscle. In the brain and muscle, the mRNA levels ofJAM-A were relatively lower than other organs, while the proteinexpression showed similar levels with other organs. Post-translational modification of JAM-A may be linked with thisdiscrepancy of mRNA and protein expressions. Although thefunctions of JAM-A such as involvement in immune reactions ortight junction regeneration were not investigated in the currentstudy, our results indicate that JAM-A is present in the intestine,kidney, liver, lung, brain, and skeletal muscle. It means that JAM-A is generally expressed in the body and may function essentialroles including immune reaction and intercellular sealing.
ZO-1 is a cytoplasmic plaque protein and alternatively calledtight junction protein 1 (TJP1). ZO-1 is a ubiquitous protein,suggesting that it is always expressed in tight junction. In our study,the expression levels of ZO-1 were similar in the intestine, kidney,liver, and muscle while the levels were higher in the lung and brain.The mRNA and protein expression patterns of ZO-1 wereconsistent in all tested tissues. ZO-1 belongs to the membrane-associated guanylate kinase (MAGUK) family (43). As a feature ofMAGUK family members, ZO-1 has PDZ domains that formbinding sites for transmembrane proteins, and connections with thecytoskeleton (44, 45). Furthermore, many studies have reportedthat ZO-1is involved in intracellular signaling and the regulation ofgene expression (46). Other studies have reported that the tightnessof tight junction is related with the expression level of ZO-1.
In conclusions, the expression of tight junctional proteins,occludin, JAM-A, and ZO-1 were examined in mice organs.Since most tight junction proteins interact with each other toperform their physiological functions, the relative levels andbalances of these tight junction is important. Three tested tightjunction proteins were expressed in the intestines, kidney, liver,lung, brain, and skeletal muscle, except occludin in the muscle.The expressions were not static, but rather varied in the testedorgans, suggesting that occludin, JAM-A and ZO-1 may havedifferent functions depending on the tissues. These findings mayhelp to design further studies related to the regulation andfunctions of occludin, JAM-A, and ZO-1 in the tissues.
Acknowledgements: This work was supported by theNational Research Foundation of Korea (NRF) grant funded bythe Korea government (MEST) (No. 2010-0011433).
Conflict of interests: None declared.
REFERENCES
1. Schneeberger EE, Lynch RD. Structure, function, andregulation of cellular tight junctions. Am J Physiol 1992;262: L647-L661.
2. Bennett MV, Barrio LC, Bargiello TA, Spray DC, HertzbergE, Saez JC. Gap junctions: new tools, new answers, newquestions. Neuron 1991; 6: 305-320.
3. Steinert PM, Roop DR. Molecular and cellular biology ofintermediate filaments. Annu Rev Biochem 1988; 57: 593-625.
4. Farquhar MG, Palade GE. Junctional complexes in variousepithelia. J Cell Biol 1963; 17: 375-412.
5. Gumbiner BM. Cell adhesion: the molecular basis of tissuearchitecture and morphogenesis. Cell 1996; 84: 345-357.
6. Fanning AS, Mitic LL, Anderson JM. Transmembraneproteins in the tight junction barrier. J Am Soc Nephrol 1999;10: 1337-1345.
7. Tsukita S, Furuse M, Itoh M. Multifunctional strands in tightjunctions. Nat Rev Mol Cell Biol 2001; 2: 285-293.
8. Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S,Tsukita S. Occludin: a novel integral membrane proteinlocalizing at tight junctions. J Cell Biol 1993; 123: 1777-1788.
9. Fanning AS, Jameson BJ, Jesaitis LA, Anderson JM. Thetight junction protein ZO-1 establishes a link between thetransmembrane protein occludin and the actin cytoskeleton.J Biol Chem 1998; 273: 29745-29753.
10. Kimura Y, Shiozaki H, Hirao M, et al. Expression ofoccludin, tight-junction-associated protein, in humandigestive tract. Am J Pathol 1997; 151: 45-54.
11. Saitou M, Ando-Akatsuka Y, Itoh M, et al. Mammalianoccludin in epithelial cells: its expression and subcellulardistribution. Eur J Cell Biol 1997; 73: 222-231.
12. Fasano A, Baudry B, Pumplin DW, et al. Vibrio choleraeproduces a second enterotoxin, which affects intestinal tightjunctions. Proc Natl Acad Sci USA 1991; 88: 5242-5246.
13. Shifflett DE, Clayburgh DR, Koutsouris A, Turner JR, HechtGA. Enteropathogenic E. coli disrupts tight junction barrierfunction and structure in vivo. Lab Invest 2005; 85: 1308-1324.
14. Nusrat A, von Eichel-Streiber C, Turner JR, Verkade P,Madara JL, Parkos CA. Clostridium difficile toxins disruptepithelial barrier function by altering membranemicrodomain localization of tight junction proteins. InfectImmun 2001; 69: 1329-1336.
15. Obert G, Peiffer I, Servin AL. Rotavirus-induced structural andfunctional alterations in tight junctions of polarized intestinalCaco-2 cell monolayers. J Virol 2000; 74: 4645-4651.
16. Nunbhakdi-Craig V, Craig L, Machleidt T, Sontag E. Simianvirus 40 small tumor antigen induces deregulation of theactin cytoskeleton and tight junctions in kidney epithelialcells. J Virol 2003; 77: 2807-2818.
17. Li D, Mrsny RJ. Oncogenic Raf-1 disrupts epithelial tightjunctions via downregulation of occludin. J Cell Biol 2000;148: 791-800.
18. Robinson C, Baker SF, Garrod DR. Peptidase allergens,occludin and claudins. Do their interactions facilitate thedevelopment of hypersensitivity reactions at mucosalsurfaces? Clin Exp Allergy 2001; 31: 186-192.
19. Moolenaar WH. Lysophosphatidic acid, a multifunctionalphospholipid messenger. J Biol Chem 1995; 270: 12949-12952.
20. Martin-Padura I, Lostaglio S, Schneemann M, et al.Junctional adhesion molecule, a novel member of theimmunoglobulin superfamily that distributes at intercellularjunctions and modulates monocyte transmigration. J CellBiol 1998; 142: 117-127.
21. Williams LA, Martin-Padura I, Dejana E, Hogg N, SimmonsDL. Identification and characterisation of human junctionaladhesion molecule (JAM). Mol Immunol 1999; 36: 1175-1188.
22. Liu Y, Nusrat A, Schnell FJ, et al. Human junction adhesionmolecule regulates tight junction resealing in epithelia. J Cell Sci 2000; 113: 2363-2374.
23. Naik UP, Ehrlich YH, Kornecki E. Mechanisms of plateletactivation by a stimulatory antibody: cross-linking of a novelplatelet receptor for monoclonal antibody F11 with the Fcgamma RII receptor. Biochem J 1995; 310: 155-162.
24. Babinska A, Kedees MH, Athar H, et al. F11-receptor(F11R/JAM) mediates platelet adhesion to endothelial cells:role in inflammatory thrombosis. Thromb Haemost 2002;88: 843-850.
25. Weber C, Fraemohs L, Dejana E. The role of junctionaladhesion molecules in vascular inflammation. Nat RevImmunol 2007; 7: 467-477.
26. Murakami M, Giampietro C, Giannotta M, et al. Abrogationof junctional adhesion molecule-A expression induces cell
17

apoptosis and reduces breast cancer progression. PLoS One6: e21242.
27. Itoh M, Furuse M, Morita K, Kubota K, Saitou M, Tsukita S.Direct binding of three tight junction-associated MAGUKs,ZO-1, ZO-2, and ZO-3, with the COOH termini of claudins.J Cell Biol 1999; 147: 1351-1363.
28. Nemeth Z, Szasz AM, Somoracz A, et al. Zonula occludens-1, occludin, and E-cadherin protein expression in biliarytract cancers. Pathol Oncol Res 2009; 15: 533-539.
29. Ikenouchi J, Umeda K, Tsukita S, Furuse M. Requirement ofZO-1 for the formation of belt-like adherens junctionsduring epithelial cell polarization. J Cell Biol 2007; 176:779-786.
30. Katsuno T, Umeda K, Matsui T, et al. Deficiency of zonulaoccludens-1 causes embryonic lethal phenotype associatedwith defected yolk sac angiogenesis and apoptosis ofembryonic cells. Mol Biol Cell 2008; 19: 2465-2475.
31. Anderson JM, Stevenson BR, Jesaitis LA, Goodenough DA,Mooseker MS. Characterization of ZO-1, a proteincomponent of the tight junction from mouse liver andMadin-Darby canine kidney cells. J Cell Biol 1988; 106:1141-1149.
32. Stevenson BR, Siliciano JD, Mooseker MS, GoodenoughDA. Identification of ZO-1: a high molecular weightpolypeptide associated with the tight junction (zonulaoccludens) in a variety of epithelia. J Cell Biol 1986; 103:755-766.
33. Anderson JM, Van Itallie CM, Peterson MD, Stevenson BR,Carew EA, Mooseker MS. ZO-1 mRNA and proteinexpression during tight junction assembly in Caco-2 cells. J Cell Biol 1989; 109: 1047-1056.
34. Vorbrodt AW, Dobrogowska DH. Molecular anatomy ofintercellular junctions in brain endothelial and epithelialbarriers: electron microscopist's view. Brain Res Brain ResRev 2003; 42: 221-242.
35. Cummins PM. Occludin: one protein, many forms. Mol CellBiol 2012; 32: 242-250.
36. Claude P, Goodenough DA. Fracture faces of zonulaeoccludentes from "tight" and "leaky" epithelia. J Cell Biol1973; 58: 390-400.
37. Al-Sadi R, Khatib K, Guo S, Ye D, Youssef M, Ma T.Occludin regulates macromolecule flux across the intestinal
epithelial tight junction barrier. Am J Physiol GastrointestLiver Physiol 2011; 300: G1054-1064.
38. Takaki Y, Hirai S, Manabe N, et al. Dynamic changes inprotein components of the tight junction during liverregeneration. Cell Tissue Res 2001; 305: 399-409.
39. Tobioka H, Tokunaga Y, Isomura H, Kokai Y, Yamaguchi J,Sawada N. Expression of occludin, a tight-junction-associated protein, in human lung carcinomas. VirchowsArch 2004; 445: 472-476.
40. Evans SM, Blyth DI, Wong T, Sanjar S, West MR.Decreased distribution of lung epithelial junction proteinsafter intratracheal antigen or lipopolysaccharide challenge:correlation with neutrophil influx and levels of BALF sE-cadherin. Am J Respir Cell Mol Biol 2002; 27: 446-454.
41. Barton ES, Forrest JC, Connolly JL, et al. Junction adhesionmolecule is a receptor for reovirus. Cell 2001; 104: 441-451.
42. Kornecki E, Walkowiak B, Naik UP, Ehrlich YH. Activationof human platelets by a stimulatory monoclonal antibody. J Biol Chem 1990; 265: 10042-10048.
43. Willott E, Balda MS, Fanning AS, Jameson B, Van Itallie C,Anderson JM. The tight junction protein ZO-1 ishomologous to the Drosophila discs-large tumor suppressorprotein of septate junctions. Proc Natl Acad Sci USA 1993;90: 7834-7838.
44. Sheng M, Sala C. PDZ domains and the organization ofsupramolecular complexes. Annu Rev Neurosci 2001; 24: 1-29.
45. Ranganathan R, Ross EM. PDZ domain proteins: scaffoldsfor signaling complexes. Curr Biol 1997; 7: R770-773.
46. Polette M, Gilles C, Nawrocki-Raby B, et al. Membrane-type 1 matrix metalloproteinase expression is regulated byzonula occludens-1 in human breast cancer cells. Cancer Res2005; 65: 7691-7698.
R e c e i v e d : October 19, 2012A c c e p t e d : February 20, 2013
Author's address: Prof. Eui-Bae Jeung, Laboratory ofVeterinary Biochemistry and Molecular Biology, College ofVeterinary Medicine, Chungbuk National University, Cheongju,Chungbuk, 361-763, Republic of Korea.E-mail: [email protected]
18