ofa 77-nucleotide-long rna methionine-acceptance. natl. acad. sci. usa vol. 85, pp. 5764-5768,...
TRANSCRIPT

Proc. Natl. Acad. Sci. USAVol. 85, pp. 5764-5768, August 1988Chemistry
Total chemical synthesis of a 77-nucleotide-long RNA sequencehaving methionine-acceptance activity
(solid-phase synthesis of tRNA/ribonucleoside phosphoramldites/amino-acylation)
K. K. OGILVIE*t, N. USMAN*4, K. NICOGHOSIAN§, AND R. J. CEDERGREN§*Department of Chemistry, McGill University, Montr6al, PQ, H3A 2K6; and §Department of Biochemistry, Universitd de Montr6al, Montreal, PQ,H3C3Jy,Canada
Communicated by Alexander Rich, April 11, 1988
ABSTRACT Chemical synthesis is described of a 77-nucleotide-long RNA molecule that has the sequence of anEscherichia coli Ado47-containing tRNAP41t species in whichthe modified nucleosides have been substituted by their un-modified parent nucleosides. The sequence was assembled on asolid-phase, controlled-pore glass support in a stepwise mannerwith an automated DNA synthesizer. The ribonucleotide build-ing blocks used were fully protected 5'-monomethoxytrityl-2'-silyl-3'-N,N-diisopropylaminophosphoramidites. p-Nitro-phenylethyl groups were used to protect the o' of guanineresidues. The fully deprotected tRNA analogue was character-ized by polyacrylamide gel electrophoresis (sizing), terminalnucleotide analysis, sequencing, and total enzyme degradation,all of which indicated that the sequence was correct andcontained only 3-5 linkages. The 77-mer was then assayed foramino acid acceptor activity by using E. coli methionyl-tRNAsynthetase. The results indicated that the synthetic product,lacking modified bases, is a substrate for the enzyme and hasan amino acid acceptance 11% of that of the major nativespecies, tRNA e't containing 7-methylguanosine at position 47.
Ribonucleic acids (RNAs) play a wide range of importantroles in living cells. These include the traditional roles intranscription and translation (mRNA, tRNA, and rRNA) ofgenetic information. Excitement in RNA chemistry hasgrown with the discovery of RNA sequences with catalyticactivity (1), the formation of "lariats" during the processingof pre-RNA (2), and the development of recombinant RNAtechnology (3, 4). RNA sequences of the tRNA type arerequired in the first step of chlorophyll biosynthesis (5).tRNAs are particularly attractive as synthetic targets becausetheir length, 70-85 nucleotides, is sufficient to demonstratea successful chemical procedure. In addition, they havespecific biological functions that can be demonstrated in thesynthetic product. They are also interesting molecules intheir own right, having been implicated in gene expression(6), in propagation of tumor viruses (7), and in cellulardifferentiation and development in cancer induction (8-10).The chemical synthesis of RNA molecules allows completeflexibility in the choice of sequence and the ability tointroduce modifications at specific sites. Thus, the ability tochemically synthesize molecules of this size should permit arapid increase in the rate of understanding of the fullbiological roles of RNA molecules paralleling that whichoccurred earlier when the chemical synthesis ofDNA oligo-mers became routine.Our laboratory has had as a major objective the develop-
ment ofmethods for the chemical synthesis ofRNA sequence(11, 12). These methods have now progressed to the pointwhere the synthesis of 70- to 80-unit molecules is a practicalpossibility. To demonstrate the chemical method, a sequence
was chosen that was identical to that of an Escherichia coliAdo-47 tRNAPet species (i.e., containing adenosine insteadof 7-methylguanosine at position 47), except that 4-thio-uridine at position 8, dihydrouridine at position 21, ribothy-midine at position 55, and pseudouridine at position 56 werereplaced by uridine, and 2'-O-methylcytidine (Cm) at posi-tion 33 was replaced by cytidine. Our beliefthat this syntheticRNA would retain some residual aminoacylation activity ofthe native tRNAPfet was founded on the observation ofOhtsuka et al. (13) that such an unmodified molecule con-taining guanosine in position 47 possessed some of the nativeactivity. The synthetic target sequence is shown in Fig. 1.To date, there have been two reports ofcombined chemical
and enzymatic syntheses of RNA sequences the length of atransfer RNA molecule. Ohtsuka et al. (13) reported thesynthesis of a 77-nucleotide-long sequence related to thetRNA14et from E. coli. The approach involved the enzymaticjoining of short, chemically synthesized oligomers (3-10nucleotides). The completed molecule possessed 4-6% oftheaminoacylation activity ofthe native tRNAv et molecule. Thesecond synthesis was described by Wang (14). This teamprepared the 76-nucleotide sequence of yeast alanine tRNAincluding the nine modified nucleosides. Short segments oftwo to eight units were prepared chemically and joinedenzymatically. The final product was reported to have 90% ofthe amino acid acceptor activity of the native tRNA. In thismanuscript we report the complete solid-phase chemicalsynthesis of the 77-nucleotide-long RNA molecule shown inFig. 1. This synthetic molecule was found to have 11% of theamino acid acceptance activity of the native 7-methylguano-sine47-containing tRNAfet (commercial sample).
MATERIALS AND METHODSPreparation of Chemical Reagents. The basic protocols for
the preparation of the ribonucleoside phosphoramidite re-agents were as described (12, 15). The ribonucleoside phos-phoramidite reagents used were: N6-benzoyl-5'-O-mono-methoxytrityl-2'-O-t-butyldimethylsilyladenosine-3'-N,N-diisopropylmethylphosphoramidite, N"-benzoyl-5 '-O-monomethoxytrityl-2'-O-t-butyldimethylsilylcytidine-3 '-N,N-diisopropylmethylphosphoramidite, N2-benzoyl-5'-O-monomethoxytrityl-06-p-nitrophenylethyl-2-O-triisopropyl-silylguanosine-3'-N,N-diisopropylmethylphosphoramidite,and 5'-O-monomethoxytrityl-2'-O-t-butyldimethylsilyluri-dine-3'-N,N-diisopropylmethylphosphoramidite and areshown schematically in Fig. 2. The adenosine-derivatized (25Amol/g) long-chain-alkylamine controlled-pore glass support
Abbreviation: Bu4NF, tetra-n-butylammonium fluoride.tTo whom reprint requests should be sent at the present address:Vice-President (Academic), Acadia University, Wolfville, NS, BOP1XO, Canada.tPresent address: Department of Biology, Massachusetts Institute ofTechnology, Cambridge, MA 02139.
5764
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 85 (1988) 5765
ACC
I AC AG - CC - GG - CG - C
U G0-C0. -C.
77
U A* % CGGCC As4U I IsII. A 58
C A G G UCGG CC CGAG C TY
18 U I I I IU WGG C U C G A* Gm U
D AGU-AC-GU G0-CG - CG - C
C *Cm AU ACAU
35
FIG. 1. Sequence of an E. coli Ado-47-containing tRNA1Iet.Arrows indicate points at which specific substitutions were made inthe synthetic 77-mer. The diamond indicates the difference betweenthe major (containing 7-methylguanosine at position 47) and minor(containing adenosine at position 47) tRNAf species. D, dihydro-uridine; S4U, 4-thiouridine; 4i, pseudouridine; Cm, 2'-O-methylcy-tidine.
was prepared as described (12) and is illustrated in Fig. 2.HPLC grade acetonitrile was first dried over activated (byheating at 600°C) 4-A molecular sieves (BDH Canada) anddistilled after continuously refluxing over CaH2. Dichloro-methane (CH2CI2), used for the detritylation solution, wasdistilled. CC13COOH (AnalaR grade; minimum assay, 99%o)was obtained from BDH Canada. All other reagents for theautomated synthesis were as supplied by Applied Biosystems(Foster City, CA).Automated Synthesis of the 77-mer E. coli tRNAPet Ana-
logue. An Applied Biosystems 381A automated DNA syn-thesizer was used for the synthesis of the 77-mer. The columnused to contain the solid support consisted of a Teflon bodywith Teflon end fittings crimped into place by using com-mercially available Hypo-Vial (Pierce) seals. A Pharmacia
Frac-100 fraction collector was attached to the instrument tocollect the released trityl cation. Since this instrument usedargon pressure to deliver the reagents, the flow rate was notidentical for the different solutions. The approximate flowrates were 1.6 ml/min for acetonitrile (CH3CN); 1.3 ml/minfor the capping solutions [0.5 M acetic anhydride/lutidine intetrahydrofuran and 0.5 M dimethylaminopyridine in tetrahy-drofuran (the two separate stock solutions were mixed in a1:1 (vol/vol) ratio by the instrument during delivery)] and forthe 5% CCl3COOH/CH2C12 detritylation solution; and 1ml/min for the ribonucleoside phosphoramidites, tetrazole,and oxidation solutions. The ribonucleoside phosphorami-dite/CH3CN and the tetrazole/CH3CN solutions were fil-tered anhydrously prior to use with a 13-mm Swinny stainlesssteel filtering unit (Millipore) fitted with a 0.5-gum filter(FHLP 013 00). Solid support (30-33 mg) bearing 25 ,umol/gof adenosine per g was used in each synthesis. The synthesiscycle shown in Table 1 was used to perform the syntheses andwas a modification of the pulsed-delivery protocol providedby the Applied Biosystems software. The coupling step of 300sec was simply a waiting step, during which time theactivated nucleoside phosphoramidite was allowed to remainin contact with the support.
Deprotection of the 77-mer E. coli tRNA""e Analogue. Allmanipulations were carried out under sterile conditions.Tetra-n-butylammonium fluoride (Bu4NF, 1 M) in tetrahy-drofuran was obtained from Aldrich. The first two steps ofthe deprotection, the thiophenoxide treatment to remove themethyl phosphate protecting group and the 3:1 (vol/vol)NH40H/95% EtOH treatment at room temperature to cleavethe sequence from the support, were performed on thecolumn itself. A 2-ml sterile polypropylene syringe (Aldrich)was attached to the column, and 5 ml of the thiophenoxidereagent, 1:2:2 (vol/vol) thiophenol/triethylamine/dioxane,was drawn into the column through a sterile needle fitted tothe other end of the column. The reaction was allowed toproceed for 1 hr, and the solution was expelled. The supportwas washed six times with 2 ml of 95% EtOH, and then 1 mlof 3:1 NH40H/95% EtOH was drawn into the column. Thesolution was allowed to stand for 30 min, and then theammoniacal solution was collected in a 5-ml sterile test tube.This was repeated twice more, and the total 3 ml ofammonia/EtOH solution was sealed in its test tube and treated at 55°Cfor 18 hr. The ammonia solution was evaporated to drynessin a Speed-Vac apparatus (Savant), and the resulting residuewas extracted first with 95% EtOH and then with 1:1(vol/vol) 95% EtOH/H20. The UV spectrum of the 95%EtOH fraction revealed the presence of 255 A270 units,whereas only 6.7 units were found in the EtOH/H20 extract.
ABZMMTO <
SiO 0
1. 5% TCA/DCM2. WASH
B3. MMT O
0 OSI
P' RRO" ~NR1R2
4. Ac20/DMAP
5. 0.1 M 12/H206. WASH
BMMTO a
0 OSiO=P-OR ABZ0Si
SiO
FIG. 2. Steps involved in each chain-extension cycle in the automated synthesis of the 77-mer: 1, removal of monomethoxytrityl (MMT)group with 5% CC13COOH (TCA) in dichloromethane (DCM) in 150-180 sec; 2, a wash with acetonitrile for 180 sec; 3, pulsed addition cyclewith 0.055 M ribonucleoside phosphoramidite 2 (R = CH3, R1 = R2 + isopropyl; B = N6-benzoyladenine, N4-benzoylcytosine,N2-benzoyl-06-p-nitrophenylethylguanine, and uracil; Si = t-butyldimethylsilyl for A, C, and U, triisopropylsilyl for G) and a 9-fold excess oftetrazole in acetonitrile for 321 sec; 4, capping step with 0.25 M (each) acetic anhydride (Ac2O), dimethylaminopyridine (DMAP), and lutidinein tetrahydrofuran for 90 sec; 5, oxidation of phosphate linkages to phosphates with 0.1 M iodine in tetrahydrofuran/pyridine/H20, 7:2:1(vol/vol) for 30 sec; 6, a wash with acetonitrile for 105 sec.
Chemistry: Ogilvie et al.

Proc. Natl. Acad. Sci. USA 85 (1988)
Table 1. Synthesis cycle for the synthesis of RNA on theApplied Biosystems 381A synthesizer
Step1234567891011121314Total
Reagent
CH3CN0.5 M tetrazole0.11 M amidite/0.5 M tetrazole, 1:10.5 M tetrazole0.11 M amidite/0.5 M tetrazole, 1:10.5 M tetrazole0.11 M amidite/0.5 M tetrazole, 1:10.5 M tetrazoleCouple0.25 M Ac20/DMAP/lutidine in H4furan0.1 M 12 in H4furan/pyridine/H20, 7:2:1CH3CN5% CC13COOH/CH2CI2CH3CN
Time, sec
6025
25
25
23009030105180120878
H4furan, tetrahydrofuran; DMAP, dimethylaminopyridine.
Only the 95% EtOH fraction was used for subsequent manip-ulations. Three 33-A270-unit aliquots were subjected to Bu4-NF treatments as follows: (0) 100 /l of 1 M Bu4NF/tetra-hydrofuran for S hr; (ii) 100 ,ul of 1M Bu4NF/tetrahydrofuranfor S hr, followed by the addition of 10 /l of H20 and 10 ILof pyridine for an additional 3 hr; (iii) 100 /l of 1 M Bu4NF/tetrahydrofuran with 10 Al each ofH20 and pyridine for 6 hr.All three samples were quenched with an equal volume of0.05 M NH4OAc and applied to a 30 x 1 cm Sephadex G-25Fcolumn. The material eluted in the void volume was collectedand quantitated. The eluants containing product were evap-orated to dryness. Preliminary analysis of the crude productswas by polyacrylamide gel electrophoresis visualized by UVshadowing over a fluorescent TLC plate. A total tRNAextract, obtained from Boehringer Mannheim, was used as areference marker. Only samples i and ii were used for furthermanipulations.
Purification. Phenol, CHC13, and gel electrophoresis re-agents were prepared as described by Maniatis et al. (16).Samples of up to 200 ;kg of crude i and ii eluted from theSephadex G-25F column were removed, dried down, andresuspended in 250 ul of sterile H20. These solutions wereextracted sequentially with 250 Al of phenol, 250 /l of 1:1(vol/vol) phenol/CHC13, 250 /l of CHCl3, and 250 Al ofH20-saturated Et2O. To the aqueous solution was added 150/l of 5 M NH4OAc, and the RNA was precipitated with 1000Al of EtOH at - 20'C overnight. The RNA was pelleted at40C, and the resulting pellet was washed with 500 ,ul of 70oEtOH to yield approximately 4 A260 units of material. Thesynthetic RNA (up to 200 gg) was taken up in 20 /l of a 50mM Tris borate/1 mM EDTA buffer and applied to a 0.8-mm-thick 20% polyacrylamide slab gel containing 7M urea inthe above buffer. Electrophoresis was carried out at 1200 Vfor 12-16 hr. Detection of RNA was accomplished by UVshadowing of the resulting gel. RNA was extracted from thegel by rinsing the appropriate excised gel bands twice withH20. The aqueous solution was then desalted on a SephadexG-50 column equilibrated in H20. The excluded volume wasIyophilized to dryness.
Sequencing. RNA samples of 1 ,ug were labeled at the 5'ends with [32P]ATP and polynucleotide kinase (17) or at the3' end with [32P]pCp and RNA ligase (18). Labeled materialwas purified by using 15% PAGE with 7 M urea, extractedwith H20, and precipitated by the addition of 10 Ag of carrierRNA per ul, 0.1 vol of 2 M NaOAc (pH 4.5), and 2.5 vol of95% ethanol. Sequencing was carried out by the enzymatichydrolysis of 5'-labeled RNA RNase T1 (guanosine specific,0.0254 unit/pg of RNA), RNase U2 (adenosine specific, 2units/gg), RNase of Bacillus cereus (C and U specific, 0.1
unit/,ug) all at 50°C for 5 min, and RNase Phy M (A and Uspecific, 0.5 unit/,g) at 55°C for 5 min. All enzymes werepurchased from Pharmacia, except for RNase U2, which wasfrom GIBCO-BRL. A sequence "ladder" was obtained byincubation of the end-labeled RNA for 30 min at 100°C indeionized formamide containing 1 mM MgC12. The 3' labeledRNA was subjected to the chemical degradation method (17)under the following reaction conditions: guanosine, 30 sec at90°C; adenosine, 7.5 min at 90°C; cytidine and uridine, 5 minat 0°C; and the aniline cleavage reaction, 3 min at 60°C in thedark.Terminal Nucleotide Analysis. The 5'-labeled RNA pre-
pared as described above was treated with 1 unit of nucleaseP1 (Pharmacia) per ,g ofRNA for 3 hr at 37°C and subjectedto two-dimensional thin-layer chromatography on a cellulosesheet in the following system (19): first-dimension eluant, 5:3(vol/vol) isobutyric acid/0.5 M ammonium hydroxide; sec-ond dimension eluant, 70:15:15 (vol/vol) 2-propanol/concen-trated HCI/H20. Autoradiography was used to detect theterminal nucleotide. The 3'-labeled RNA was treated with0.05 unit ofRNase T2 per ,g ofRNA for 14-16 hr at 37°C andsubjected to two-dimensional thin-layer chromatography asabove.
Aminoacylation. The tRNA acceptor activity of the 77-merRNA species was assayed in 25 ,u of a buffer containing 50mM sodium cacodylate (pH 7.5), 12.5 mM MgCl2, 2.5 mMATP, 2.5 mM CTP, 25 mM KCI, 10 ug of bovine serumalbumin, and 0.1 ,uCi (1 Ci = 37 GBq) of L-[14C]methionine(Amersham, 60.4 mCi/mmol). Incubation was for 15 min at37°C with 280 units of aminoacyl-tRNA synthetases [Sigma;1 unit = 1 pmol of labeled amino acid (arginine) incorporatedin 10 min at 37°C and pH 7.6]. The RNA was precipitated onfilter paper and assayed for 5-min periods in a scintillationcounter. All assays were done in duplicate.
RESULTS AND DISCUSSIONChemical Synthesis. The general procedure for assembling
an RNA sequence by the addition of nucleotide units one ata time in a continuous chemical process in the 3'-5' directionis outlined in Table 1 and Fig. 2. The synthesis was conductedon the surface of long-chain alkylamine controlled-pore glassto which an adenosine unit (25 ,mol/g) had been attached(Fig. 2, step 1). Ribonucleoside phosphoramidite 2 (see Fig.2 legend) was activated with tetrazole and then allowed toreact with the exposed 5'-hydroxyl group on the polymersurface, thus adding one more unit to the chain. A cappingreagent (acetic anhydride) was then added, followed by aniodine solution to oxidize the phosphite linkage to the desiredphosphate internucleotide linkage. After washing steps, themonomethoxytrityl group at the 5' position was removedwith 5% CC13COOH. The released methoxytrityl cation wascollected and quantitated by its absorption at 478 nm. Anadditional washing step prepared the polymer for anotheraddition cycle. Each complete cycle required 14.63 min and76 complete cycles were required. The average coupling yieldwas 98%, giving an overall yield of 24%. The steps in thissynthesis should all be compatible with any of the commer-cial synthesizers designed for phosphoramidite reagents inDNA synthesis.We have shown (15) that the use of guanine o6 protection
is not strictly necessary to prevent modification of the o6position during the synthesis of DNA or RNA. However, itdoes offer the advantage of conferring increased solubility inthe medium used for deprotection to the nucleoside oroligonucleotide as compared with the 06-unprotected nucle-oside or oligonucleotide (20, 21). Furthermore, the p-nitrophenylethyl group is removed concomitantly with thealkylsilyl groups by using the Bu4NF/tetrahydrofuran re-agent.
5766 Chemistry: Ogilvie et al.

Proc. Natl. Acad. Sci. USA 85 (1988) 5767
Deprotection of the 77-mer E. coli tRNA;'t Analogue. Uponcompletion ofthe synthesis, the product was deprotected andremoved from the polymer. The methyl groups were removedfrom the phosphate triester intemucleotide linkages by usingthiophenol in triethylamine and dioxane, 1:2:2 (vol/vol), for1 hr. The controlled-pore glass was washed with 95% ethanoland allowed to stand for 30 min in 1 ml of 3:1 (vol/vol)NH40H/ethanol at room temperature (to cleave the nucle-otide chain from the controlled-pore glass support). Thisprocess was repeated twice more, and the combined wash-ings were heated at 550C for 18 hr (this treatment removed thebenzoyl protecting group from the adenine, cytosine, andguanine bases).
This product was then treated with 1 M Bu4NF/tetra-hydrofuran'(35-40 equivalents per silyl group) alone or in thepresence of pyridine and H20 for 5 hr for simultaneousremoval of all of the silyl (t-butyldimethylsilyl for adenosine,cytidine, and uridine and triisopropylsilyl for guanosine)protecting groups and the 06-p-nitrophenylethyl groups fromthe guanine bases (22). An equal volume of 0.05 M NH4OAcwas added, and the mixture was applied directly to aSephadex G-25F column. The column was eluted with 0.05MNH4OAc and the nucleotide product was recovered in thevoid volume. Where only the Bu4NF/tetrahydrofuran solu-tion was used or where it was followed by the addition ofpyridine/H20, 23 A260 units of material with a Amk = 260 nm(indicative of complete deprotection) were recovered. In thecase where pyridine/H20 was included at the beginning ofthe fluoride treatment, 27 A270 units (i.e., Am., = 270 nm) ofmaterial, indicative of the presence of uncleaved 06-p-nitrophenylethyl protecting groups, were recovered. Sam-ples of the first two preparations were identical when run ona 16% polyacrylamide/8 M urea gel.
It is noteworthy that the sample still containing p-nitrophenylethyl groups not only had a slower mobility ondenaturing PAGE but also was a much poorer substrate forpolynucleotide kinase during labeling with [ry32P]ATP whencompared with labeling of the fully deprotected 77-mers orauthentic tRNAP'et (unpublished data). These findings inaddition to the change in Amx from 270 to 260 nm confirmedthe complete removal ofp-nitrophenylethyl groups from thefully deprotected samples. The fully deprotected 77-merswere subsequently purified, sequenced, and assayed for theiraminoacylation capability.
Purification, Labeling, Sequencing, and Terminal Nucleo-tide Analysis. After the desalting of the desilylation reactionmixture, the crude 77-mers were extracted with phenol,phenol/CHCI3, and CHC13. The samples were then precip-itated in EtOH, and the resulting pellets were washed with70o EtOH. After this, the products were purified by 10%polyacrylamide gel electrophoresis. Some samples had beenpurified by BD-cellulose as well, but in all cases furthercharacterization was preceded by thorough desalting of thesample.The synthetic RNA was characterized in several ways.
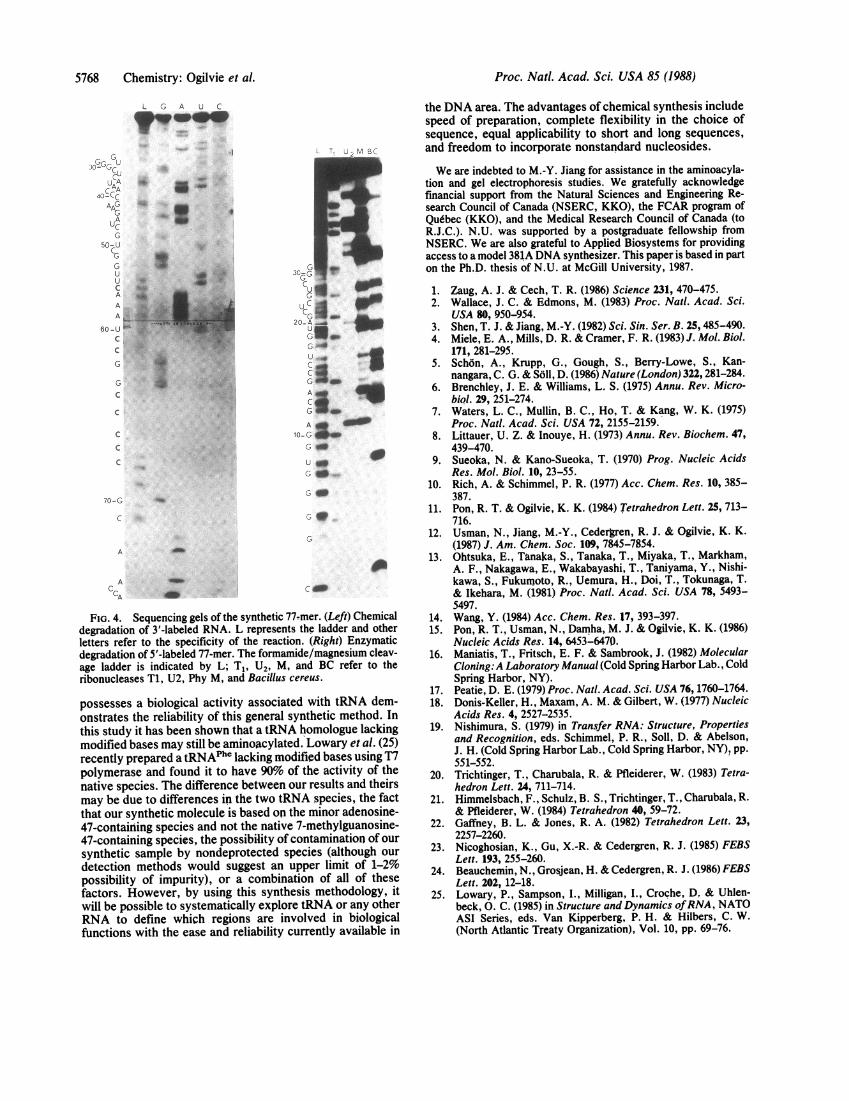
First, the crude sample was labeled with 32p at its 5' terminusby using polynucleotide kinase (23). Polyacrylamide gelelectrophoresis in 7 M urea showed that the labeled purifiedsample has a mobility similar to a labeled sample ofthe majortRNAP"'t isoaccepter from E. coli (Fig. 3). Subsequently, the5'-terminal nucleotide, pC, was determined by complete P1nuclease digestion and thin-layer chromatographic compar-ison of the labeled nucleotide with authentic samples. In likemanner, the 3'-terminal nucleoside was identified as adeno-sine after the synthetic RNA was labeled by using [32P]pCpand RNA ligase, followed by enzymatic hydrolysis to 3'nucleotides by using T2 RNase (24). Finally, rapid gelsequencing techniques were applied to 5'- and 3'-labeledsamples of the RNA (Fig. 4) (24). The 5'-labeled RNA wassequenced by the enzymatic degradation procedure, whereas
t RNAMet 77-merfOrigin
xc
BPB
FIG. 3. A 15% polyacrylamide/7- M urea gel electrophoresiscomparison oftRNASCe and synthetic 77-mer. Both tRNAf et and thepurified synthetic 77-mer were labeled with 32P by using polynucleo-tide kinase and [-32P]ATP. Authentic tRNA is in the left lane and the77-mer is in the right lane. The origin and the position ofthe two dyes,bromophenol blue (BPB) and xylenecyanol (XC), are indicated.
the 3'-labeled material was analyzed by the chemical degra-dation procedure. In both cases and in complementaryfashion, the synthetic RNA corresponded to the sequenceshown in Fig. 1.Aminoacylation of the 77-Nucleotide-Long RNA. The ami-
noacyl acceptance activity was obtained after a seconddesalting ofthe crude sample and a renaturation cycle of55TCfor 5 min, 37TC for 10 min, and 20TC for 10 min. Anappropriate amount of synthetic RNA was incubated for 15min with ["4C]methionine and an aminoacyl tRNA synthetasefraction from E. coli (24). The extent of aminoacylation wasascertained by counting the radioactivity of filter-precipi-tated samples. The activity was determined to be 60 ± 5 pmolof methionine per A260 unit (11% of theoretical activity atsaturation level) ofRNA based on duplicate analysis of threedifferent samples. Under identical conditions, purified com-mercial tRNA;1et charged 546 + 24 pmol of methionine perA260 unit (theoretical, 1600 pmol per A 60 unit). Aminoacy-lation assays have been performed on samples prepared byother purification techniques such as BD-cellulose and com-binations thereof. In these cases and in assays done in Hepesbuffer at pH 7.5, lower (5-6% of the authentic sample) butconsistent activities have been found.The synthesis of a molecule of ribonucleic acid consisting
of 77 nucleotides that may be fully characterized and that
Chemistry: Ogilvie et al.
Proc. Natl. Acad. Sci. USA 85 (1988)
L G A U CQW9_-c
A GAGA
UCG50- UCG
UC
UU
AAAL
60-U '-C,
C
G
C
Cc
i30
20
10.C
C
C
70-G
A _
A _
CCA -
FIG. 4. Sequencing gels of the synthetic 77-1degradation of 3-labeled RNA. L represents tletters refer to the specificity of the reactiondegradation of 5'-labeled 77-mer. The formamidage ladder is indicated by L; T1, U2, M, a'ribonucleases Ti, U2, Phy M, and Bacillus cer
possesses a biological activity associatedonstrates the reliability of this general syrthis study it has been shown that a tRNA I
modified bases may still be aminoacylated.recently prepared a tRNAPhC lacking modiipolymerase and found it to have 90% ofnative species. The difference between ou
may be due to differences in the two tRN.that our synthetic molecule is based on the47-containing species and not the native 747-containing species, the possibility of cosynthetic sample by nondeprotected speidetection methods would suggest an uppossibility of impurity), or a combinati(factors. However, by using this synthesiwill be possible to systematically exploreIRNA to define which regions are invofunctions with the ease and reliability cui
I
the DNA area. The advantages of chemical synthesis includespeed of preparation, complete flexibility in the choice ofsequence, equal applicability to short and long sequences,and freedom to incorporate nonstandard nucleosides.
We are indebted to M.-Y. Jiang for assistance in the aminoacyla-tion and gel electrophoresis studies. We gratefully acknowledgefinancial support from the Natural Sciences and Engineering Re-search Council of Canada (NSERC, KKO), the FCAR program ofQuebec (KKO), and the Medical Research Council of Canada (toR.J.C.). N.U. was supported by a postgraduate fellowship fromNSERC. We are also grateful to Applied Biosystems for providingaccess to a model 381A DNA synthesizer. This paper is based in parton the Ph.D. thesis of N.U. at McGill University, 1987.
C 10 ,g,1. Zaug, A. J. & Cech, T. R. (1986) Science 231, 470-475.C: i -2. Wallace, J. C. & Edmons, M. (1983) Proc. Natl. Acad. Sci.-G _ USA 80, 950-954.U-A 3. Shen, T. J. & Jiang, M.-Y. (1982) Sci. Sin. Ser. B. 25, 485-490.5U - 4. Miele, E. A., Mills, D. R. & Cramer, F. R. (1983) J. Mol. Biol.G _ 171, 281-295.U. 5<< 5. Sch6n, A., Krupp, G., Gough, S., Berry-Lowe, S., Kan-c 2_ nangara, C. G. & Soll, D. (1986) Nature (London) 322, 281-284.
A _ 6. Brenchley, J. E. & Williams, L. S. (1975) Annu. Rev. Micro-ci biol. 29, 251-274.G-_ 7. Waters, L. C., Mullin, B. C., Ho, T. & Kang, W. K. (1975)A do-- Proc. Nat!. Acad. Sci. USA 72, 2155-2159.-G 4_ 8. Littauer, U. Z. & Inouye, H. (1973) Annu. Rev. Biochem. 47,G _0 439-470.U _ 9. Sueoka, N. & Kano-Sueoka, T. (1970) Prog. Nucleic AcidsG a. Res. Mol. Biol. 10, 23-55.
10. Rich, A. & Schimmel, P. R. (1977) Acc. Chem. Res. 10, 385-387.
11. Pon, R. T. & Ogilvie, K. K. (1984) Tetrahedron Lett. 25, 713-716.
12. Usman, N., Jiang, M.-Y., Cedergren, R. J. & Ogilvie, K. K.(1987) J. Am. Chem. Soc. 109, 7845-7854.
13. Ohtsuka, E., Tanaka, S., Tanaka, T., Miyaka, T., Markham,do A. F., Nakagawa, E., Wakabayashi, T., Taniyama, Y., Nishi-
kawa, S., Fukunmoto, R., Uemura, H., Doi, T., Tokunaga, T.c_-& Ikehara, M. (1981) Proc. Natl. Acad. Sci. USA 78, 5493-
5497.mer. (Left) Chemical 14. Wang, Y. (1984) Acc. Chem. Res. 17, 393-397.the ladder and other 15. Pon, R. T., Usman, N., Damha, M. J. & Ogilvie, K. K. (1986)(Right) Enzymatic Nucleic Acids Res. 14, 6453-6470.
le/magnesium cleav- 16. Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982) Molecularnd BC refer to the Cloning:A Laboratory Manual (Cold Spring Harbor Lab., Coldreus. Spring Harbor, NY).
17. Peatie, D. E. (1979) Proc. Nat!. Acad. Sci. USA 76, 1760-1764.I with tRNA dem- 18. Donis-Keller, H., Maxam, A. M. & Gilbert, W. (1977) Nucleicnthetic method. In Acids Res. 4, 2527-2535.iomologue lacking 19. Nishimura, S. (1979) in Transfer RNA: Structure, PropertiesLowary eta!. (25) and Recognition, eds. Schimmel, P. R., Soll, D. & Abelson,
J. H. (Cold Spring Harbor Lab., Cold Spring Harbor, NY)? pp.Fied bases using T7 551-552.the activity of the 20. Trichtinger, T., Charubala, R. & Pfleiderer, W. (1983) Tetra-r results and theirs hedron Lett. 24, 711-714.A species, the fact 21. Himmelsbach, F., Schulz, B. S., Trichtinger, T., Charubala, R.e minor adenosine- & Pfleiderer, W. (1984) Tetrahedron 40, 59-72.-methylguanosine- 22. Gaffney, B. L. & Jones, R. A. (1982) Tetrahedron Lett. 23,)ntamination of our 2257-2260.cies (although our 23. Nicoghosian, K., Gu, X.-R. & Cedergren, R. J. (1985) FEBS
limitof1-2% Lett. 193, 255-260.oper limi of 1hes 24. Beauchemin, N., Grosjean, H. & Cedergren, R. J. (1986) FEBSon of all of these Lett. 202, 12-18.is methodology, it 25. Lowary, P., Sampson, I., Milligan, I., Croche, D. & Uhlen-tRNA or any other beck, 0. C. (1985) in Structure and Dynamics ofRNA, NATOAlved in biological ASI Series, eds. Van Kipperberg, P. H. & Hilbers, C. W.rrently available in (North Atlantic Treaty Organization), Vol. 10, pp. 69-76.
5768 Chemistry: Ogilvie et A