obtenciÓn de enzimas lignocelulolÍticas y … · departamento de ingeniería ... diseño y puesta...
TRANSCRIPT

1
OBTENCIÓN DE ENZIMAS LIGNOCELULOLÍTICAS Y
POLISACÁRIDOS A PARTIR DE RESIDUOS LIGNOCELULÓSICOS DEL
DEPARTAMENTO DE CALDAS EMPLEANDO MACROMICETOS DE
PUDRICIÓN BLANCA POR FERMENTACIÓN SUMERGIDA Y
FERMENTACIÓN EN ESTADO SÓLIDO
SANDRA MONTOYA BARRETO
Facultad de Ciencias Agropecuarias
Doctorado en ciencias Agrarias
Universidad de Caldas
Manizales, Colombia, 2012

OBTENCIÓN DE ENZIMAS LIGNOCELULOLÍTICAS Y POLISACÁRIDOS A
PARTIR DE RESIDUOS LIGNOCELULÓSICOS DEL DEPARTAMENTO DE CALDAS
EMPLEANDO MACROMICETOS DE PUDRICIÓN BLANCA POR FERMENTACIÓN
SUMERGIDA Y FERMENTACIÓN EN ESTADO SÓLIDO
Tesis para optar el título de
Doctora en Ciencias Agrarias de:
SANDRA MONTOYA BARRETO
Bajo la dirección del Doctor:
ÓSCAR JULIÁN SÁNCHEZ TORO
Facultad de Ciencias Agropecuarias
Doctroado en Ciencias Agrarias
Manizales, 2012

Agradecimientos
Institutciones y oficinas
Universidad de Caldas:
Vicerrectoría de Investigaciones y Postgrados, por la financiación de la Tesis Doctoral
Departamento de Ingeniería, por el aval para la comisión de estudios.
Instituto de Biotecnología Agropecuaria, por el uso de los laboratorios y sus instalaciones
para el desarrollo de la Tesis.
Grupo de alimentos y Agroindustria, por el apoyo permanente durante el desarrollo de la
investigación.
Universidad de Buenos Aires, por el apoyo con la pasantía realizada en el Departamento de
Biodiversidad y biología Experimental.
Universidade Federal Do Paraná, por el apoyo con la pasantía realizada en el Departamento de
Bioquímica y Biología Molecular.
Doctores e investigadores
Doctor Óscar Julián Sánchez Toro, Director de la tesis Doctoral, por su apoyo y
orientación permanentes en cada uno de los procesos académicos durante el desarrollo de
todo el trabajo.
Doctora Laura Levín, Codirectora de la Tesis Doctoral, por su apoyo permanente y
orientaciones pertinentes durante todo el proceso de formación doctoral.
Doctoras Lucía Atehortúa, profesora de la Universidad de Antioquia y Aida Rodriguez de
Stouvenel, porfesora de la Universidad del Valle, quienes conforman el Comité tutorial de
mi Tesis Doctoral, por su evaluación oportuna y participación en los eventos académicos
relacionados con la Tesis Doctoral.
Prof. Arkady P. Sinitsyn, Institute of Biochemistry, Russian Academy of Sciences,
Department of Chemical Enzymology, Moscow State University, Russia, por su apoyo y
orientación durante visita académica a la Universidad de Caldas en el año 2011. Estudiantes de Postgrados, Ingenieros y Compañeros
Profesor Óscar Darío Hernández, por su apoyo y colaboración en la construcción y
desarrollo de los modelos matemáticos desarrollados en este trabajo.
Profesional Diego Fernado Arias Monsalve, por su colaboración con el trabajo de campo
realizado para la Tesis.
Profesionales Patricia Salazar Villegas y Silvio Alexandro Gómez saldarriaga, por la
colaboración y apoyo con el trámite de la solcitud de Patente de Biorreactor de lecho fijo.
Agradecimientos personales
Especialmente a mi hijo por su compañía permanente, su amor y apoyo complice; a mi esposo por su paciencia y solaridad.

TABLA DE CONTENIDO
1. Introducción 4
1.1. Contexto general 4
1.2. Objetivos de la tesis 8
2. Estado del Arte y Marco Teórico 10
2.1. Los hongos 10
2.1.1 Generalidades de los hongos 10
2.1.2. Biología d elos hongos 12
2.1.3 Crecimeinto y desarrollo de los hongos 13
2.1.4. Investigaciones con hongos en Colombia 15
2.2. materiales lignocelulósicos 20
2.2.1. Celulosa 22
2.2.2. Hemicelulosa 24
2.2.3. Lignina 25
2.2.4. Otros polímeros presentes en los materiales lignocelulósicos 27
2.2.5. Pretratamiento de los materiales lignocelulóscios 28
2.3. Enzimas lignocelulolíticas 30
2.3.1. Enzimas celulolíticas 31
2.3.2. Enzimas xilanolíticas 37
2.3.3. Enzimas ligninolíticas 38
2.3.4. Otras enzimas producidas por hongos de pudrición blanca 43
2.3.5. Aplicaciones de las enzimas 44
2.4. Polisacáridos fúngicos 45
2.5. fermentación sumergida 48
2.6. Fermentación en estado sólido 50
2.7. Modelamiento de las cinéticas de fermentación de hongos de pudrición blanca 54
2.8. Evaluación de biomasa fúngica, enzimas lignocelulolíticas y polisacáridos 57
2.8.1. Detarminación de biomasa celular 57
2.8.2. Rastreo de cepas para la producción de biosustancias 58
2.8.3. determinación de actividades enzimáticas lignocelulolíticas 59
2.8.4. Determinación de polisacáridos 59
Referencias 60
3. Evaluación de Actividades Endoglucanasa, Exoglucanasa, Lacasa y Lignina Peroxidasa en Diez hongos de pudrición blanca 72

4. Producción de Polisacáridos por Cultivos Sumergidos y en Estado sólido a partir de diferentes basidiomicetos 86
5. Selección de la mejor Combinación Hongo – Formulación para la degradación de Sustratos Lignocelulósicos mediante Fermentación en Estado Sólido con Pleurotus ostreatus, Coriolus versicolor y Lentinus edodes 98
6. Selección de la mejor Combinación Hongo – Formulación para la degradación de Sustratos Lignocelulósicosproducción de polisacáridos mediante Fermentación en Estado Sólido con Pleurotus ostreatus, Coriolus versicolor y Lentinus edodes 120
7. Selección de la mejor Combinación Hongo – Formulación para la degradación de Sustratos Lignocelulósicos mediante Fermentación Sumergida con Pleurotus ostreatus, Coriolus versicolor y Lentinus edodes 139
8. Selección de la mejor Combinación Hongo – Formulación para la producción de exo-polisacáridos mediante Fermentación Sumergida con Pleurotus ostreatus, Coriolus versicolor y Lentinus edodes 163
9. Biorreactor para la Obtención de Ssutancias Bioactivas por Fermentación en Estado Sólido Empleando Hongos Macromcietos (Solicitud de Patente) 181
10. Conclusiones Generales 184
10.1. Contribuciones de la Tesis 184
10.2. Impactos Potenciales de la Tesis 186
10.3. Trabajos Futuros 187
Anexos 1. Documento Solictud de Patente
Anexo 2. Obtención de polisacáridos del cuerpo fructífero Grifola frondosa
Anexo 3. Aprobación de proyecto Colciencias

6
1. Introducción
1.1. Contexto general
1.1.1. Campo de aplicación y motivación
La producción de enzimas lignocelulolíticas y polisacáridos a partir de hongos de pudrición blanca
empleando residuos lignocelulósicos como sustratos ha sido estudiada en diversos centros de
investigación y universidades en el mundo con múltiples resultados a escala de laboratorio. Sin embargo,
no hay disponible información suficiente sobre los procesos industriales para la obtención de estos
productos, ni sobre su escalamiento. Tampoco existe información suficiente y disponible en torno a la
producción de polisacáridos fúngicos empleando residuos lignocelulósicos como sustratos, diferenciando
la fuente de los polisacáridos fúngicos como son el cuerpo fructífero de los hongos, el micelio vegetativo
o los medios de cultivo procedentes de las fermentaciones sumergidas. De igual forma, se ha detectado
que muchos de los problemas que se presentan en el escalamiento y montaje de los procesos industriales
para la producción de biosustancias empleando hongos macromicetos sobre matrices sólidas heterogéneas,
como son las diversas mezclas de materiales lignocelulósicos, se deben a la ausencia de modelos
matemáticos adercuados que describan y permitan la articulación entre el crecimiento y desarrollo de los
macromicetos de pudrición blanca, el consumo de los sustratos y la producción de las sustancias con valor
comercial.
Varios fueron los motivos que nos llevaron a encarar este trabajo en el marco del Doctroado en
Ciencias Agrarias de la Universidad de Caldas, entre los que se destacaron:
En Colombia son escasos los trabajos de investigación en torno a la producción de enzimas
lignocelulolíticas obtenidas a partir de hongos de pudrición blanca empleando residuos
lignocelulósicos como sustratos en procesos de fermentación sumergida y en estado sólido.
En la producción de polisacáridos de hongos de pudrición blanca se requiere información en torno a
la descripción de las clases de polisacáridos que pueden obtenerse dependiendo si éstos se encuentran
en el cuerpo fructífero, en el micelio o son excretados al medio de cultivo. Asimismo, es reducida la
información en torno a los esquemas de extracción de polisacáridos y a su identificación y
cuantificación.
No sehan establecido pleanemente los medios de cultivo basados en materiales lignocelulósicos
pretratados utilizados en fermentación sumergida para la producción de polisacáridos a partir de
hongos de pudrición blanca.
Existen escasas propuestas en referencia a modelos matemáticos que permitan la descripción del
crecimiento y desarrollo de los hongos de pudrición blanca y la producción de biosustancias como
metabolitos primarios y secundarios a fin de buscar el escalamiento de los procesos (nivel piloto)
para su producción industrial.
La producción de enzimas lignocelulolíticas y polisacáridos de hongos de pudrición blanca
empleando mezclas de residuos lignocelulósicos es bastante compleja por todas las interacciones que se
presentan entre la biomasa, el sustrato y las nuevas sustancias producto del metabolismo del hongo; estas
sustancias son a su vez necesarias para realizar la conversión de la matriz polimérica del sustrato,
constituida principalmente por celulosa, hemicelulosa y lignina, en sustancias pequeñas que le sirvan de
fuente de energía para su crecimiento y desarrollo. Esta tesis doctoral intenta describir los procesos de
obtención de enzimas lignocelulolíticas y polisacáridos, como productos del metabolismo de estos hongos.
Este trabajo fue realizado entre agosto de 2009 y diciembre de 2012, con el soporte económico de la
Vicerrectoría de Investigaciones y Postgrados, el Instituto de Biotecnología Agropecuaria y el
Departamento de Ingeniería de la Universidad de Caldas; adicionalmente, se contó con el apoyo parcial de
la Gobernación de Caldas con cargo a los recursos de la estampilla para proyectos de investigación de las

universidades públicas del Departamento de Caldas. La autora de esta tesis realizó una pasantía
internacional en dos instituciones de reconocida trayectoria en el campo de aplicación que aborda el
presente trabajo: Departamento de Biología y Bioquímica de la Universidade Federal do Paraná
(Curitiba, Brasil) y el Departamento de Biodiversidad y Biología Experimental de la Universidad de
Buenos Aires (Argentina).
Durante el desarrollo de esta investigación se realizaron dos trabajos paralelos que complementan
esta tesis y hacen parte de los productos obtenidos como aplicaciones de los conocimientos adquiridos, los
cuales son: i) Diseño y puesta en marcha de la Planta de Bioprocesos de la Universidad de Caldas, la
cual consta de seis unidades de proceso, compostaje, cultivo de hongos macromicetos, planta de
tratamiento de aguas residuales, lombricultivo, unidad de fermentaciones sumergidas con biorreactores
bajo condiciones controladas y unidad de fermentaciones en estado sólido con biorreactores bajo
condiciones controladas; la cual se encuentra operando desde diciembre de 2011; y ii) planteamiento y
desarrollo del proyecto Desarrollo y evaluación de los parámetros de cultivo a escala semi industrial del
hongo Grifola frondosa en climas tropicales y su transformación con fines comerciales como productos
nutraceúticos. Este proyecto se encuentra en la ejecución de su primera etapa que corresponde al estudio
de mercado del producto y a pruebas piloto de producción.
El espectro de aplicación de este trabajo se resume en la importancia de ampliar el conocimiento en
torno a la producción de enzimas lignocelulolíticas de los hongos de pudrición blanca Coriolus versicolor,
Lentinus edodes y Pleurotus ostreatus (seleccionados de entre 10 especies de hongos) sobre 12
formulaciones diferentes en fermentaciones sumergida y en estado sólido bajo condiciones controladas.
Esta importancia radica no solamente en la relevancia comercial que puede llegar a tener la producción de
enzimas de este tipo, sino porque el conocimiento acerca de cómo estos hongos producen estas enzimas,
permiten profundizar en el conocimiento del funcionamiento de los hongos de pudrición blanca, lo cual
posibilita incrementar la eficiencia en los cultivos de hongos para la obtención de sus cuerpos fructíferos,
y de otras sustancias como polisacáridos de micelio y exopolisacáridos de los medios de cultivo en
procesos de fermentación sumergida. De igual forma, se amplía el conocimiento en referencia al potencial
que presentan los polisacáridos de los hongos macromicetos y su importancia como potenciadores del
sistema inmunológico, para lo cual es relevante dilucidar los esquemas de extracción y la importancia de
la obtención de las diferentes fracciones y su acción sinérgica.
1.1.2. Estructura de la tesis
El documento de la tesis se desarrolló en 10 capítulos. En el capítulo dos se presenta el estado del arte
de la tesis, el cual contiene una descripción acerca de los hongos, los materiales lignocelulósicos, las
enzimas lignocelulolíticas, los procesos de fermentación sumergida y en estado sólido, la producción de
polisacáridos fúngicos, los modelos matemáticos, los métodos analíticos para determinar las actividades
enzimáticas y polisacáridos, y la realización de rastreos para la producción de enzimas y polisacáridos de
cepas fúngicas. En la revisión realizada sobre los hongos se contemplaron sus generalidades, su biología,
crecimiento y desarrollo y un compendio de trabajos realizados en Colombia empleando hongos. En la
sección de materiales lignocelulósicos, se encontrará información relacionada con la composición química
de estos materiales y su distribución dentro de la matriz lignocelulósica, que permita dilucidar cómo
pueden ser atacadas estas estructuras por las enzimas lignocelulolíticas producidas por los hongos de
pudrición blanca. De igual forma, en esta sección se encontrará un compendio sobre tipos de
pretratamientos que se les puede realizar a los materiales lignocelulósicos a fin de desordenar la matriz
lignocelulósica y permitir más fácilmente el acceso de los hongos de pudrición blanca a estos materiales
en procesos de fermentación sumergida. En la sección 2.3 se encontrará información general sobre las
enzimas lignocelulolíticas, los tipos de estas enzimas producidas por los macromicetos de pudrición
blanca, su modo de acción sobre los materiales lignocelulósicos y las aplicaciones de ellas en diferentes
sectores industriales. En la sección 2.4 se encontrará un compendio sobre polisacáridos fúngicos, las
clases de polisacáridos contenidos en la pared celular fúngica, cómo se producen, las funciones
fisiológicas y su utilidad en la industria farmacéutica. En las secciones 2.5 y 2.6 se tratan los procesos de

fermentaciones sumergidas y en estado sólido respectivamente con hongos de pudrición blanca, en las
cuales se describe la importancia de conocer el comportamiento de estos hongos en cada uno de los
procesos con sus ventajas y desventajas para producción a escala industrial. En la sección 2.7 se hace un
compendio sobre el modelamiento de las cinéticas de fermentación de hongos de pudrición blanca, en el
que se exponen las expresiones matemáticas más utilizadas para el ajuste de datos experimentales de
procesos de fermentación sumergida y en estado sólido, así como los problemas que representa la ausencia
de modelos para el escalamiento de estos procesos productivos a tamaño industrial. La última sección de
este capítulo trata sobre cómo se realiza la evaluación de biomasa fúngica, enzimas lignocelulolíticas y
polisacáridos; en ella se discute cómo se determina la biomasa en procesos de fermentación en fase sólida
por estimación indirecta, utilizando la medida de N-acetil-D-glucosamina (NAGA), el cual es precursor de
quitina, componente mayoritario de la pared celular de los hongos. En esta sección también se hace
referencia a los métodos para realizar el rastreo de cepas para la producción de biosustancias, en particular
de enzimas lignocelulolíticas y polisacáridos. En la última parte de esta sección se describen los métodos
para determinar actividades enzimáticas lignocelulolíticas y polisacáridos.
En el capítulo tres se encontrará el primer artículo producto de esta tesis doctoral: “Polysaccharide
production by submerged and solid state cultures from several basidiomicetes”, aceptado para su
publicación en la revista International Journal of Medicinal Mushrooms. Este artículo es el resultado del
rastreo de polisacáridos realizado a diez especies de hongos de pudrición blanca con el fin de seleccionar
la mejor cepa para la producción de exopolisacáridos y el cuerpo fructífero de mayor contenido de
intrapolisacáridos. A su vez, el capítulo cuatro contiene el artículo: “Evaluación de las actividades
endoglucanasa, exoglucanasa, lacasa y lignina peroxidasa en diez hongos de pudrición blanca”, como
resultado del rastreo de enzimas lignocelulolíticas realizado a las diez cepas de hongos de pudrición
blanca con el fin de seleccionar las mejores cepas productoras de celulasas y ligninasas.
En los capítulos cinco, seis, siete y ocho se encontrarán los artículos realizados como compendio de
los resultados obtenidos en el diseño experimental como elemento central de esta tesis. Los capítulos
cinco y seis corresponden a la producción de enzimas lignocelulolíticas y polisacáridos por fermentación
en estado sólido, respectivamente, y los capítulos siete y ocho están relacionados con la producción de
enzimas lignocelulolíticas y polisacáridos por fermentación sumergida.
En el capítulo nueve se encontrará el documento petitorio de la patente: “Biorreactor para la
obtención de sustancias bioactivas por fermentación en estado sólido empleando hongos macromicetos”,
en el cual se describe el diseño, construcción y puesta en marcha de un biorreactor de lecho fijo para
fermentación en fase sólida con hongos de pudrición blanca para la producción de biosustancias como
enzimas lignocelulolíticas y polisacáridos. Por último en el capítulo diez se encontrará la discusión general
de la tesis, en la que se condensarán los resultados generales obtenidos, los impactos y las perspectivas
futuras del trabajo.
1.2. Objetivos de la Tesis
1.2.1. Objetivo general
Obtener enzimas lignocelulolíticas y polisacáridos a partir de residuos lignocelulósicos disponibles en
el Departamento de Caldas empleando macromicetos de pudrición blanca tanto por fermentación
sumergida como por fermentación en estado sólido.
1.2.2. Objetivos específicos
Determinar la influencia de los medios de cultivo formulados con base en los residuos
lignocelulolíticos seleccionados disponibles en el Departamento de Caldas sobre la producción de
enzimas lignocelulolíticas y polisacáridos empleando macromicetos de pudrición blanca por
fermentación en medio líquido.

Determinar la influencia de los medios de cultivo formulados con base en los residuos
lignocelulolíticos seleccionados disponibles en el Departamento de Caldas sobre la producción de
enzimas lignocelulolíticas y polisacáridos empleando macromicetos de pudrición blanca por
fermentación en estado sólido.
Realizar y caracterizar cinéticamente los procesos de fermentación sumergida y en estado sólido a
nivel piloto para la producción de enzimas lignocelulolíticas y polisacáridos empleando macromicetos
de pudrición blanca.

10
2. Estado del Arte y Marco Teórico
2.1. Los hongos
2.1.1. Generalidades de los hongos
Los hongos tienen una importancia fundamental como organismos descomponedores en el ambiente
terrestre, ellos juegan un rol importante en los ciclos bio-geoquímicos de carbono, nitrógeno, fósforo y otros
(Wainwright, 1988; Gadd, 1999). Los hongos hacen parte de una comunidad natural con plantas, otros
microorganismos e invertebrados, esta combinación demuestra la condición mutualista de los hongos en el
planeta y los pone como integrantes importantes en los procesos que involucran los ciclos ambientales de la
tierra (Bosell et al., 2003). Adicionalmente, la importancia de los hongos se incrementa por su uso en la
producción industrial de alcoholes, antibióticos, ácidos orgánicos, prebióticos, diversas sustancias de uso
farmacológico, enzimas, además del gran potencial de uso en procesos de biorremediación, control
biológico, entre otros (Burgstaller y Schinner, 1993; Tobin et al., 1994; Gadd, 2001).
Los hongos son organismos celulares sin cloroplastos y por lo tanto heterótrofos que poseen paredes
celulares compuestas de quitina y células con especialización funcional. Actualmente se consideran como un
grupo heterogéneo, formado por organismos pertenecientes por lo menos a tres líneas evolutivas
independientes (Whittaker, 1969; Naeem, 2002). Los hongos son organismos eucarióticos uni o
pluricelulares que se desarrollan en sitios húmedos y con poca luz. Las células de los hongos pluricelulares
se agrupan en filamentos llamados hifas que en conjunto reciben el nombre de micelio (Hudson, 1986;
Chang y Miles, 2004). Los hongos descomponen la materia orgánica por medio de enzimas, absorbiendo las
sustancias nutritivas. La reproducción puede ser asexual o sexual, principalmente por esporas. La mayor
parte de los hongos son saprofitos; algunos se consideran parásitos; otros mutualistas o simbiontes basados
en asociaciones con algas, líquenes o con otro grupo en forma de micorrizas, en los que los hongos
acompañan a la mayor parte de las plantas, residiendo en sus raíces y ayudándolas a absorber nutrientes del
suelo. Se piensa que esa simbiosis fue esencial para la conquista del medio terrestre por las plantas y para la
existencia de los ecosistemas continentales (Hudson, 1986; Naeem, 2002).
Otras características de los hongos pueden resumirse diciendo que carecen de tejido vascular y que no
tienen movimiento. Mientras el componente principal de la pared celular de las plantas está formado por
celulosa, la de los hongos contiene mayoritariamente quitina (Carlile et al., 2001). Los hongos son
heterotróficos, igual que los animales, pero los animales primero ingieren y después digieren y los hongos
primero digieren y después ingieren por medio de la producción de moléculas más simples. La mayoría de
los hongos, como los animales, almacenan alimentos como glucógeno; mientras las plantas almacenan
almidones, celulosas y azúcares simples. Los saprófitos se alimentan de materia orgánica en descomposición.
La falta de clorofila afecta profundamente su forma de vida: no necesitan de la luz. Crecen en cualquier
dirección; invaden el sustrato con filamentos absorbentes. Junto con las bacterias, su función principal es
reciclar el carbono, el nitrógeno y los minerales esenciales para la nutrición. Como parásitos, usan la materia
orgánica de organismos vivos, causando daño a plantas, animales y humanos (por las toxinas que producen).
En simbiosis, pueden beneficiar a otros organismos, como las micorrizas en las raíces de las plantas y los
líquenes (asociación de un hongo y un alga). Son organismos muy útiles por su versatilidad genética y
fisiológica, producen enormes cantidades de esporas que permanecen viables hasta que las condiciones
climatológicas favorecen su multiplicación. Descomponen toda clase de productos manufacturados,
exceptuando los plásticos y algunos plaguicidas. Los hongos son muy útiles al ser humano, los hay que
producen antibióticos, hormonas y esteroides y muchas más sustancias. Son muy importantes en la
investigación, debido a que se reproducen rápida y fácilmente, ocupan poco espacio y su ciclo de vida puede
ser controlado a conveniencia de los procesos (Whittaker, 1969; Moore, 1998a; Carlile et al., 2001). Las
características moleculares (recién descubiertas) indican que los hongos están más relacionados con los
animales que con las plantas (Hudson, 1986; Naeem, 2002).
Existen diferencias entre los micromicetos y macromicetos. En las hifas de los micromicetos u hongos

inferiores, el citoplasma es generalmente continuo, no está separado por septos, posee numerosos núcleos
minúsculos, que se desplazan libremente por una corriente citoplasmática que permite el transporte de
nutrientes al interior del micelio. Los hongos superiores o macromicetos tiene septos con perforaciones
centrales que permiten el paso del citoplasma (Carle - Urioste et al., 1997).
Las setas o champiñones son macrohongos que tienen cuerpos fructíferos definidos, que pueden ser
epigeos (encima de la tierra o aéreos) o hipogeos (bajo la tierra), son observados por el ojo humano y
recolectados. Conforme con esta definición, champiñón no es un basidiomiceto, aéreo, suave y comestible
exclusivamente; champiñón, también puede ser un ascomiceto no aéreo (Carlile et al., 2001; Chang y Miles,
2004). El tipo más común de champiñones es el que posee píleo (sombrero o cabeza) en forma de sombrilla
y estípite (pie) como el Lentinus edodes; sin embargo existen macrohongos que tiene anillos, como Agaricus
bisporus, o volva, como el Volvariella volvacea, o ambos como el Amanita phalloides. Adicionalmente,
existen los que tienen forma de taza plegable, palo de golf, en forma de coral, de globo, además de diversos
colores, lo que muestra que los cuerpos fructíferos de los hongos llamados champiñones son ricos en
diversidad de formas y colores y que son un cúmulo de micelios en su estado de madurez reproductiva
(Hudson, 1986; Chang y Miles, 2004). Principalmente en hongos superiores (Ascomycota y Basidiomycota)
la parte recolectada del hongo no es más que el órgano de reproducción del hongo, llamado carpóforo (Gow,
1995a). El verdadero cuerpo del hongo, o cuerpo vegetativo, está escondido formado por una red de
filamentos microscópicos inmersa en el substrato, llamada micelio (Carlile et al., 2001).
Los hongos tanto por su capacidad hidrolítica como por su distribución, son los organismos
lignocelulolíticos por excelencia. Entre ellos existen algunos con mayor capacidad degradativa de lignina:
los que producen la llamada “pudrición blanca” que podrían utilizarse en el proceso de bioconversión de
materiales lignocelulósicos. Esta categoría definida por el tipo de pudrición que causan en la madera
contiene cientos de especies de Basidiomycetes. Todos son capaces de degradar la lignina, la celulosa y la
hemicelulosa de la madera, pero la velocidad y extensión de la degradación de cada componente de la pared
celular varía considerablemente (Joselau y Ruel, 1994; Carlile et al., 2001).
Los hongos de pudrición castaña o café, clasificados dentro de los basidiomycetes, degradan
preferentemente celulosa, la alta concentración de lignina que queda en la madera deteriorada evidencia la
limitada capacidad de estos hongos para degradar la lignina. La lignina residual sin embargo, está
químicamente alterada y diversas investigaciones revelan que ocurre cierto grado de degradación de lignina,
principalmente a través de reacciones de demetoxilación. Atacan las capas de la pared a distancia, desde el
lumen celular (no son capaces de degradar extensivamente a la lignina; lo que les facilitaría el acceso a los
carbohidratos), por lo tanto los agentes del deterioro deben ser moléculas muy pequeñas para pasar por los
poros de las paredes. No tienen exoglucanasa y en cambio llevarían a cabo la solubilización de la celulosa
nativa por un mecanismo que involucra a la endoglucanasa y a factores no proteicos, entre ellos H2O2 y
Fe(II) (reactivo de Fenton: que produce radicales libres -OH muy reactivos que oxidan las cadenas de
celulosa). Producen extracelularmente H2O2 y la madera contiene suficiente Fe(III). Recientemente se
describió en Gloeophyllum trabeum un ciclo hidroquinona-quinona redox extracelular que reduce Fe(III) y
produce H2O2. Junto con la producción de enzimas, Trichoderma también excreta sustancias que generan
hinchamiento e hidratación sobre las moléculas que va a degradar, presumiblemente como coadyuvantes de
la degradación (Carlile et al., 2001). En estadíos tempranos del decaimiento hay reducción de la fuerza
mecánica de la madera pero no pérdida de peso seco, mientras que en la pudrición blanca son dos procesos
que progresan en paralelo. Esto se debe a que los hongos de pudrición castaña clivan completamente a través
de las regiones amorfas de las microfibrillas antes de utilizar la celulosa; mientras que los hongos de
pudrición blanca erosionan progresivamente la superficie de las microfibrillas y consumen la celulosa a
medida que esta es degradada.
El desperdicio de enzimas secretadas al medio es probablemente minimizado por la existencia de una
cubierta o vaina de exopolisacáridos que se desarrolla alrededor de la hifa durante la colonización del hongo
sobre la madera. Las moléculas de enzimas son retenidas en estas vainas y no se dispersan muy lejos de la
zona del sustrato a ser degradado. Es sorprendente como esta acción permite que el hongo pueda digerir
enzimáticamente el micelio viejo como fuente de nitrógeno orgánico y reutilizar su misma biomasa para
desarrollar nuevas hifas (Hudson, 1986; Gow, 1995a; Carlile et al., 2001).

Los hongos de pudrición blanca son los principales causantes de la degradación de la madera. Son más
numerosos que los hongos de pudrición café, están representados por basidiomicetos y ascomicetos. Son
capaces de degradar completamente la lignina hasta su mineralización, dejando la madera de color blanco en
las zonas atacadas, por la celulosa remanente. Estos hongos producen grandes cantidades de oxido-
reductasas, las cuales son las responsables de la mineralización de la lignina, además de tener la capacidad de
producir y transformar sustancias no fenólicas en fenólicas para su fácil degradación y la obtención de
sustancias aromáticas de menor tamaño que la lignina y de menor toxicidad. Este concepto lo describe Wong
(2009), cuando expone el mecanismo de acción de la enzima lignina peroxidada (LiP) sobre la estructura de
la lignina. La estructura de la lignina no es única, es decir depende de cómo se unen sus precursores, los
alcoholes sinapílico, coniferílico y p-cumarílico, basados en substituciones metoxi sobre el anillo aromático.
Estos precursores se unen por medio de diversos tipos de enlaces, los cuales conforman una matriz de
unidades estructurales por enlaces C-C y éter. Estos enlaces incluyen el acoplamiento de β-O-4, β-5, β-β, 5-
5, 4-O-5, y 1-β. Este tipo de agrupamientos a través de tan diversos enlaces demuestran que la lignina se
sintetiza como una estructura fenólica y no-fenólica. Teniendo en cuenta que en la estructura de la lignina
están presentes sólo alrededor del 10-20% de subunidades fenólicas. Sin embargo, reacciones de
demetilación y ruptura de los enlaces éter catalizados por la enzima LiP durante la degradación pueden
generar productos fenólicos, que a su vez pueden servir de sustrato para la subdivisión de la lignina. En
general, la ruptura de los restos no-fenólicos, componen el 80-90% de la estructura de lignina y se convierten
en el indicador principal de la degradación enzimática. Asimismo, dentro del grupo de hongos de pudrición
blanca existen los hongos que degradan selectivamente la lignina, los cuales degradan primero la mayor
parte de la lignina presente en la vecindad del material que se encuentra rodeando la hifa antes de iniciar la
degradación de la celulosa y los que atacan simultáneamente celulosa y lignina en la madera. De todos
modos los hongos de pudrición blanca y café comparten varias características y ambos pueden ser utilizados
con diversos fines en la industria agrícola (Hudson, 1986; Carlile et al., 2001).
2.1.2. Biología de los hongos
Los principales responsables del reciclaje de las plantas muertas son los hongos saprófitos. Esta actividad
de reciclaje es esencial para la continuación de la vida en el planeta. El ciclo del carbono involucra la fijación
de dióxido de carbono atmosférico dentro de moléculas orgánicas por fotosíntesis; los hongos juegan un
papel importante en la degradación de estas moléculas reponiendo nuevamente el dióxido de carbono a la
atmósfera. Además de madera aprovechable, una explotación forestal produce anualmente dos toneladas por
hectárea de residuos forestales húmedos, los cuales pueden ser degradados exclusivamente por hongos
especializados (Moore, 1998a; Chang y Miles, 2004). Además de la importancia que tiene la degradación de
estos residuos forestales en el ciclo de otros elementos como el nitrógeno, fósforo y potasio, los cuales deben
ser incorporados como componentes insolubles de las células vegetales (Moore, 1998a).
Los hongos por ser organismos eucariotas típicos, poseen un núcleo que contiene varios cromosomas
(siete en Candida albicans, ocho en Aspergillus nidulans y 16 en Saccharomyces cerevisiae) delimitado por
una membrana nuclear, con nucléolo rico en ARN y orgánulos citoplásmicos como mitocondrias, vacuolas,
retículo endoplásmico, aparato de Golgi y ribosomas 80S. El citoplasma se encuentra limitado por la
membrana citoplásmica, que es una doble capa de lípidos que contiene proteínas y esteroles y que controla la
permeabilidad celular y participa en la síntesis de la pared celular. La estructura de las células de los hongos
es muy diferente de la de las bacterias que son organismos procariotas (Whittaker, 1969). Aunque comparten
muchas estructuras, las células de los hongos se diferencian de las de las plantas en la composición de la
pared celular y en que carecen de cloroplastos y clorofila, y de las humanas en que tienen pared celular y en
la presencia de ergosterol en la membrana citoplásmica. Por el exterior de la membrana citoplásmica,
presentan una pared celular que está compuesta fundamentalmente por polisacáridos y por diversas
proteínas. Los polisacáridos más importantes son la quitina (polímero de N-acetil glucosamina), el manano
(polímero de manosa) y el glucano (polímero de glucosa) (Carlile et al., 2001), siendo la quitina el
componente mayoritario de la pared celular de los hongos, el cual es un polímero de la N-acetil-D-
glucosamina; esta sustancia puede ser vista como un derivado de la celulosa, en la cual los grupos hidroxilos

del segundo carbono de cada unidad de glucosa son reemplazados con grupos acetamido (NH(C=O)CH3).
El desarrollo fúngico se concentra en el ápice de la hifa, con numerosas vesículas liberando enzimas
líticas y biosintéticas de la membrana plasmática a una alta velocidad. Se han desarrollado amplios estudios
sobre la biosíntesis de la quitina y de otros componentes de la pared celular (Carlile et al., 2001; Campbell y
Farrell, 2004). Pero aún resta por estudiar en profundidad las rutas metabólicas de la biosíntesis de las
enzimas que involucran la conversión de glucosa-6-fosfato en el precursor de quitina.
2.1.3. Crecimiento y desarrollo de los hongos
El crecimiento de los hongos es un fenómeno complejo que no tiene una definición sencilla. Este
crecimiento es un incremento ordenado de los componentes celulares que involucran un aumento de
biomasa. Las velocidades de crecimiento celular pueden variar por diferentes factores, tales como las
características genéticas y el número de generaciones (multiplicaciones) de la cepa. Intervienen factores
externos como las condiciones ambientales, las características físico-químicas y el estado (sólido o líquido)
de los sustratos en los que crecen, las condiciones de asepsia de las plantas de producción donde se
desarrollan los cultivos, entre otros. Estos factores también afectan de forma directa o indirecta la producción
de metabolitos de los hongos durante las fases de crecimiento y desarrollo (Chang y Miles, 2004).
El desarrollo de los hongos en cultivos líquidos induce el micelio a la formación de pellets, generalmente
de forma esférica. La hifa en el interior del pellet está expuesta a condiciones limitadas de oxígeno, lo que
no sucede con las hifas que se encuentran en la superficie de éste. Los micelios que se desarrollan en cultivos
líquidos no están limitados espacialmente, como sí es el caso de los que crecen en medios sólidos (Stames,
1993; Chang y Miles, 2004).
La fase vegetativa de los hongos filamentosos es especializada y está dividida en dos estructuras
micelianas: rizomorfas, que corresponde a los ramales de hifas bien formados, y esclerotia, que son los
ramales de hifas entrecruzadas o trenzadas. Estas estructuras probablemente cumplen con la función primaria
de sobrevivencia de los hongos en condiciones ambientales adversas (Chang y Miles, 2004).
En la fase reproductiva de los hongos intervienen múltiples estructuras especializadas. En referencia a la
reproducción sexual de los ascomicetos, ésta involucra la producción de meiosporas dentro de una estructura
en forma de bolsas llamadas ascas. Las meiosporas haploides son denominadas ascosporas. Para estudios
genéticos los ascomicetos más trabajados han sido los géneros Saccharomyces, Neurospora y Sordaria
(Carlile et al., 2001; Chang y Miles, 2004).
Los basidiomicetos producen sus meiosporas (basidiosporas) sobre una estructura llamada basidio. Para
estudios genéticos los géneros de basidiomicetos más empleados son Ustilago, Schizophyllum y Coprinus, y
de los géneros comestibles se reconocen Agaricus, Lentinus, Volvariella, Flamulina y Pleurotus (Hudson,
1986; Chang y Miles, 2004).
En referencia a los requerimientos nutricionales de los hongos, es necesario considerar tanto los
requerimientos de oxígeno, como las concentraciones del ión hidrógeno. Si bien existen hongos, como las
levaduras que pueden vivir en condiciones anaeróbicas, la mayoría de los hongos son considerados aeróbicos
y las bajas concentraciones de oxígeno en los medios donde los hongos se desarrollan pueden generar un
efecto negativo por tensión (déficit) de oxígeno. Del mismo modo, el ión hidrógeno (pH) puede provocar
efectos positivos y negativos en el desarrollo morfológico del hongo, ya que este factor es determinante para
la absorción y asimilación de nutrientes. En general, la mayoría de los hongos se desarrollan adecuadamente
en intervalos de pH de 4 a 8, aunque hay excepciones y, por lo tanto, estas especies deben ser manejadas por
separado. Los factores tensión de oxígeno y pH influyen de forma directa en los procesos metabólicos y
consecuentemente en la habilidad de las especies fúngicas para utilizar las sustancias del medio como
nutrientes en poco tiempo (Hudson, 1986; Carlile et al., 2001; Chang y Miles, 2004). De igual forma, en la
utilización de las fuentes de carbono; éste provee los requerimientos energéticos y estructurales a las células
fúngicas. El carbono es tomado por los hongos de diversas fuentes, como de polisacáridos, monosacáridos,
ácidos orgánicos, aminoácidos, alcoholes, compuestos policíclicos, celulosas y hemicelulosas, además de
tener la habilidad de degradar sustancias como la lignina. El nitrógeno es esencial en la síntesis de proteínas,
purinas y pirimidinas. La quitina, un polisacárido de ocurrencia común en las paredes celulares de muchos

hongos, también contiene nitrógeno. Los hongos utilizan una variedad de fuentes para obtener el nitrógeno
necesario para la síntesis de estos compuestos esenciales. Se ha reportado por Burnett (1976) que existen
organismos eucarióticos que tienen la habilidad de usar el nitrógeno atmosférico para la síntesis de estas
sustancias; otras fuentes de nitrógeno utilizadas son los nitratos, el ión amonio y las fuentes de nitrógeno
orgánico.
Los minerales y las vitaminas también son muy importantes para el crecimiento y desarrollo de los
hongos, además de intervenir directamente en la producción de sus metabolitos primarios y secundarios. En
la Tabla 1 se muestran las proporciones de los minerales y vitaminas denominados esenciales para el
adecuado crecimiento y desarrollo de los hongos. Cuando se pretende producir alguna sustancia en particular
o se busca la fructificación de una especie específica, debe indagarse respecto a las necesidades puntuales del
hongo en cuanto a inductores o inhibidores de las sustancias deseadas a fin de obtener el medio con las
mejores características para un fin específico.
En conjunto con los requerimientos nutricionales que demandan los hongos para su crecimiento y
desarrollo, éstos también, necesitan unas condiciones físicas mínimas en los sustratos para su adecuado
desarrollo. Los factores físicos más importantes son la temperatura, la luz, la humedad, la aireación y la
gravedad. Otros factores que pueden incidir en el crecimiento de algunos hongos son la presión hidrostática,
viscosidad, y radiación mutagénica. Comúnmente, se hace referencia a condiciones mínimas necesarias para
el adecuado desarrollo de los hongos. Sin embargo, cada especie tiene unos requerimientos propios que
deben ser suplidos cuando se desea optimizar los procesos de producción de alguna especie en particular
(Chang y Miles, 2004).
Tabla 1. Requerimientos de minerales y vitaminas esenciales para el desarrollo fúngico
Compuesto Tipo/forma Función Cantidad
media
Minerales
Azufre (S): como sulfato de magnesio
o suministrado por aminoácidos que
contienen S.
Para la síntesis de diversos metabolitos secundarios
como las penicilinas y mercaptanos y para la síntesis
de aminoácidos que contienen S.
10-4
M
Fósforo (P): como fosfatos de
potasio o por el mismo material,
cuando éste contiene P.
El P se requiere para la síntesis de adenosin trifosfato
(ATP), ácidos nucleicos y los fosfolípidos de la
membrana celular.
10-3
M
Potasio (K): como fosfatos de potasio El K cumple el rol como cofactor en algunos sistemas
enzimáticos, está involucrado en el metabolismo de
los carbohidratos y es importante en el
mantenimiento del balance iónico de los hongos.
10-3
M
Magnesio (Mg): como sulfato de
magnesio.
Es esencial para todos los hongos, muchas enzimas
son activadas por Mg y es importante en el
metabolismo del ATP
10-3
M
Elementos menores o microelementos
(traza): Hierro (Fe)
Activador de enzimas oxido-reductasas. 10-6
M
Elementos menores o microelementos
(traza): Zinc (Zn)
Activador de enzimas, alcohol deshidrogenasa,
contiene cuatro átomos de Zn/molécula.
10-8
M
Elementos menores o microelementos
(traza): Manganeso (Mn)
Activador de muchas enzimas (oxido-reductasas), se
requiere en el ciclo ATC y en la síntesis de ácidos
nucleicos.
10-6
- 10-7
M
Elementos menores o microelementos
(traza): Cobre (Cu)
Requerido para el desarrollo normal de los hongos,
intervine como activador de varias enzimas, en
cantidades muy altas resulta tóxico, ya que tiene
actividad fungicida.
10-7
M
Elementos menores o microelementos
(traza): Molibdeno (Mo)
Requerido esencialmente si la fuente de nitrógeno
tiene nitratos, ya que es un elemento constituyente de
la flavoproteína nitrato reductasa, la cual reduce los
nitratos hasta el ión amonio.
10-6
M
(muy
variado)
Elementos menores o microelementos Requerido para la formación de carpóforos de varias 10-6
M

(traza): Calcio (Ca) especies de macromicetos.
Vitaminas
Tiamina (vitamina B1): adicionando
pirimidinas, tiazoles o algunas
especies requieren la molécula de
tiamina completa.
Actúa como coenzima de la carboxilasa en la
regulación del metabolismo de carbohidratos por
conversión del ácido pirúvico en acetaldehído y
dióxido de carbono. En especies de macromicetos,
puede requerirse en la fase vegetativa, para fructificar
o en la maduración de los carpóforos.
100 µg/L
Biotina (vitamina B7 o H): por
materiales que la contengan o
adicionándola directamente.
Como coenzima para la carboxilación, es coenzima
de la piruvato carboxilasa, actúa como donador o
aceptor de dióxido de carbono, en reacciones de
carboxilación de ácidos grasos, tales como el ácido
aspártico.
5 µg/L
Ácido nicotínico (B3), Ácido
pantoténico (B5) y ácido para-amino-
benzoico: suplidas por materiales
naturales, que son heterogéneos, si
los medios son sintéticos, deben
adicionarse.
Como coenzimas de diversos grupos de enzimas. Muy
pequeñas
cantidades
(<1µg/L)
Fuente: Chang y Miles (2004)
2.1.4. Investigaciones con hongos en Colombia
En la Tabla 2 se hace una recopilación de trabajos de investigación y revisiones publicados sobre hongos
en general, hechos en Colombia con énfasis en hongos de pudrición blanca. Los trabajos representan el
interés de las instituciones y de los académicos en la profundización, investigación y desarrollo en procesos y
productos utilizando hongos y el conocimiento de estos organismos enfocados a las necesidades que
demanda un país con vocación agrícola como Colombia. En la búsqueda realizada se encontró una mayor
cantidad de trabajos referidos al conocimiento de hongos entomopatógenos, causantes de infecciones
agrícolas y micromicetos de uso industrial. En la Tabla 2 se ubicaron algunas publicaciones seleccionadas
que se refieren a temas como modificaciones genéticas, modo de acción y control biológico de hongos en
general. De acuerdo al contexto de este trabajo en particular, se realizó con detalle la búsqueda de trabajos
con hongos macromicetos, en especial con hongos de pudrición blanca; obteniéndose como resultado una
cantidad de publicaciones muy importantes sobre diferentes tópicos como son el reconocimiento de las
especies nativas de los bosques colombianos, su taxonomía, caracterización morfológica, y diversas
aplicaciones, entre las que se destacan procesos de biorremediación a nivel de laboratorio, evaluación de
técnicas de cultivo de varios hongos con atributos comestibles y medicinales a escala artesanal o de
laboratorio, y caracterizaciones químicas de diversos grupos funcionales de interés de los cuerpos fructíferos
de estos hongos; así como, trabajos donde se desarrollaron procesos de fermentación en fase líquida y sólida
con estos organismos. Sin embargo, no se encontraron trabajos que planteen el escalamiento de procesos de
fermentación en estado sólido y líquido para la producción de enzimas lignocelulolíticas y polisacáridos con
hongos de pudrición blanca silvestres o cepas adaptadas. Asimismo, no se encontraron referencias sobre
investigación y desarrollo de exo e intrapolisacáridos a una escala de producción diferente a la de
laboratorio.
Tabla 2. Trabajos de investigación y revisiones sobre hongos en general hechos en Colombia
Título del trabajo Descripción Referencia
Utilización de residuos de plátano
para la producción de metabolitos
secundarios por fermentación en
estado sólido con el hongo Lentinus
crinitus
Residuos del cultivo de plátano empleados como sustrato en un proceso de
fermentación en estado sólido con el hongo de la podredumbre de la madera
Lentinus crinitus. La producción de metabolitos fue determinada durante un
periodo de 21 días de fermentación conformado por 7 combinaciones de
sustrato. Evaluaron la presencia de los compuestos aromáticos ácido ferúlico,
vainilla, ácido vainillínico y eugenol en los días 11, 16 y 21.
Granada et al.
(2005)

Estudio del efecto de dos
inductores y un protector
enzimático sobre la actividad de las
enzimas MnP y lacasa producidas
por Trametes versicolor y su acción
en la decoloración
Evaluación del uso de Tween 80 como protector enzimático y variación de
concentraciones de sales de cobre y manganeso como activadoras de las
enzimas MnP y lacasa evaluadas sobre los materiales coloreados. Gómez-
Dorado et al.
(2005)
Establecimiento de un medio de
cultivo para una cepa nativa de un
hongo poliporal
Producción de fenoles totales, actividad antioxidante, crecimiento micelial y
consumo de sustrato de una cepa nativa de un hongo poliporal en cultivo
sumergido.
Peña-Serna et
al. (2006)
Inmovilización de hongos
ligninolíticos para la remoción del
colorante negro reactivo 5
Evaluaron el efecto de los hongos Trametes versicolor, Pleurotus ostreatus y
Phanerochaete chrysosporium sobre la decoloración de un agua que contiene
colorante negro reactivo 5 (NR5), usado en la industria textil. El hongo con
mayor capacidad para la decoloración fue el Trametes versicolor.
Fernández et
al. (2009)
Macromicetes (Ascomycota,
basidiomycota) de la región del
medio Caquetá, departamentos de
Caquetá y Amazonas (Colombia)
Listado taxonómico de macromicetos encontrados en los departamentos de
Caquetá y Amazonas (Colombia). Vasco-
Palacios et al.
(2005)
Nuevos registros de
Aphyllophorales (Basidiomycota)
en bosque montano húmedo
y de niebla de Colombia
Los hongos Aphyllophorales (Basidiomycota) contribuyen de gran manera al
mantenimiento de la estructura y la función de los ecosistemas forestales a
través de su actividad saprofítica como descomponedores de madera. Los
estudios sobre estos organismos han sido escasos y esporádicos en Colombia;
de acuerdo con la literatura se han inventariado 262 especies para el país.
Ruíz y Varela
(2006)
Actividad enzimática, degradación
de residuos sólidos orgánicos y
generación de biomasa útil por el
macromiceto Grifola frondosa
Se determinaron las actividades enzimáticas de las enzimas celulolíticas:
endoglucanasa, exoglucanasa y β-glucosidasa; ligninolíticas: lacasa, lignin
peroxidasa (LiP) y manganeso-peroxidasa (MnP) y xilanolíticas: endoxilanasa
del hongo en dos formulaciones diferentes basadas en aserrín de roble y borra
de café
Montoya
(2008)
Effect of culture parameters on the
production of the edible
mushroom Grifola frondosa
(maitake) in tropical weathers
Desarrollo de las condiciones de cultivo del hongo Grifola frondosa en climas
tropicales. En Manizales Colombia se adaptó y desarrolló una cepa del hongo
en cultivos a nivel de laboratorio, con una eficiencia biológica de 34%
comparable con las eficiencias obtenidas por Shen y Royse (2002)
Montoya et al.
(2008)
Producción de biomasa y
exopolisacáridos de Grifola
frondosa bajo cultivo sumergido
utilizando fuentes de carbono no
convencionales
Evaluaron la incidencia de diferentes fuentes de carbono (FC) en la producción
de biomasa y exopolisacáridos (EPS), bajo condiciones de cultivo sumergido.
La máxima producción de biomasa micelial (21,10 ± 0,82) g/l y la máxima
producción de EPS (6,53 ± 0,14) g/l se logró utilizando el medio MB
suplementado con FC
Zapata et al.
(2007)
Producción de enzimas
ligninolíticas por Basidiomycetes
mediante la técnica de
fermentación en fase sólida
Evaluación del crecimiento de los hongos de pudrición blanca: Trametes
versicolor, Phanerochaete chrysosporium y Pleurotus florida sobre residuos
sólidos agroindustriales para la producción de las enzimas ligninolíticas: LiP,
MnP y lacasa, utilizando matraces
García y
Torres (2003)
Dehidroergosterol: un artefacto
generado durante el proceso de
extracción de esteroles en el hongo
Pleurotus sajor-caju
Estudiaron la fracción esterólica del hongo Pleurotus sajor-caju. Establecieron
que el dehidroergosterol (ergosta-5, 7, 9 (11), 22-tetraen-3-ol) presente en el
extracto obtenido del hongo no es un producto natural sino un artefacto
producido durante el proceso de extracción de los esteroles con cloroformo. El
valor nutricional de los hongos cultivados en desechos agroindustriales del café
puede verse afectado por la presencia de polifenoles.
Rivera et al.
(2005b)
Determinación de ácidos grasos y
compuestos triterpenoides del
cuerpo fructífero de Suillus luteus
Del cuerpo fructífero de Suillus luteus extrajeron e identificaron, con base en el
análisis de sus espectros de masas, dieciséis compuestos, los cuales
corresponden al ácido palmítico, oléico, linolénico y linoléico, octadecanoato
de etilo y otros a fin de buscar nuevas fuentes de sustancias con diversos usos
industriales.
Nieto y Ávila
(2008)
Degradación de colorantes
industriales con hongos
ligninolíticos
Evaluaron siete cepas de hongos ligninolíticos en función de su capacidad para
degradar el colorante Orange II y los colorantes industriales Rojo Cibacrón®,
Rojo Erionyl®, Azul Terasil® y Turquesa Erionyl® en medio semisólido y
líquido. Phanerochaete chrysosporium y Phanerochaete sordida mostraron
alta capacidad de decoloración, alcanzando 98% en medio líquido para Orange
II y entre 82 y 86% para los colorantes industriales después de ocho días de
tratamiento. En medio semisólido todos los colorantes se eliminaron
completamente.
Cardona et al.
(2009)

Estudio químico de la fracción
insaponificable del hongo
macromiceto Lentinula edodes
(Shiitake)
Obtuvieron la fracción insaponificable del Lentinula edodes alcanzando 5
fracciones denominadas: LE1, LE2, LE3, LE4 y LE5. Del estudio de los
espectros de masas de las diferentes fracciones por comparación con patrones y
análisis de fragmentación, identificaron 8 esteroles tipo ergostano.
Benavidez-
Calvache
(2004)
Aislamiento y evaluación de la
actividad enzimática de hongos
descomponedores de madera en la
reserva natural La montaña del
ocaso, Quimbaya-Quindío
Evaluaron hongos seleccionados recolectados de la reserva natural La montaña
del ocaso, Quimbaya-Quindío, les determinaron actividad ligninolítica
semicuantitativa inoculando los hongos sobre agar y realizando inducción de
las ligninasas con ABTS.
Chaparro y
Rosas (2006)
Evaluación de la síntesis de
proteína a partir del crecimiento
vegetativo de Pleurotus spp. sobre
residuos de algarrobo y uva pasa
Estudio sobre la capacidad de síntesis de proteína de los hongos Pleurotus
ostreatus, P. sajor-caju y P. pulmonarius sobre uno de los residuos de la
Industria Licorera de Caldas, uva pasa y algarrobo, en su fase vegetativa.
Montoya y
Restrepo
(2006)
Validación de deshidratación
convencional para la conservación
del hongo comestible Pleurotus
sajor-caju
Deshidratación convencional del hongo, le realizaron evaluación
microbiológica al producto obtenido y el análisis estadístico. La deshidratación
de esta seta la realizaron a cuatro temperaturas (45, 50, 55 y 60°C).
Castro-Ríos
(2006)
Incorporación de cafeína en el
hongo Pleurotus sajor-caju
cultivado sobre
pulpa de café
Estudiaron la composición química de los metabolitos secundarios de
Pleurotus sajor-caju cultivado en pulpa de café, encontraron que el hongo
tiene la capacidad de incorporar en su fructificación la cafeína, componente del
sustrato (cerca del 1,3% en base seca), sin modificar la estructura del alcaloide.
Nieto-
Ramírez et al.
(2007)
Macrocybe titans (Bigelow y
Kimbr.) Pegler, Lodge y Nakasone,
un registro nuevo para Colombia
Descripción macro y microscópica a partir de colecciones realizadas durante
los años 2001-2004 del Macrocybe titans (Bigelow y Kimbr.) Pegler Lodge, y
Nakasone en los departamentos Colombianos de Antioquia y Santander. Este
género y especie se registran por primera vez para Colombia.
Corrales y
López (2005)
Nuevos aspectos sobre la
clasificación de los hongos y su
posible aplicación médica
Revisión sobre la importancia de los hongos para el hombre y los aspectos
indeterminados sobre la clasificación y ubicación como reino, evolución:
conceptos filogenéticos, biológicos y morfológicos
Montes et al.
(2003)
Producción de enzimas
ligninolíticas con hongos
basidiomicetos cultivados sobre
materiales lignocelulósicos
Evaluación de la propagación de los hongos Bjerkandera adusta y
Phanerochaete chrysosporium sobre suelos contaminados mezclados con
materiales lignocelulósicos que les suministran la fuente de carbono necesaria
para sostener su crecimiento e inducir la producción del complejo enzimático
ligninolítico y la biodegradación de contaminantes.
Quintero et al.
(2006)
Screening of white rot fungal
species for their capacity to
degrade lindane and other isomers
of hexachlorocyclohexane (HCH)
Evaluación de la capacidad de degradación de las especies Bjerkandera adusta,
Irpex lacteus, Lentinus tigrinus, Phanerochaete chrysosporium, Phanerochaete
sordida, Phlebia radiata, Pleurotus eryngii, Polyporus cialatus, y Stereum
hirsutum actuando sobre varias concentraciones de sustancias tóxicas como α-,
β-, γ- y δ isómeros de hexaclorociclohexano (HCH)
Quintero et al.
(2008)
Aislamiento e identificación de
hongos filamentosos de muestras
de suelo de los páramos Guasca y
Cruz verde
Aislaron e identificaron hongos filamentosos en los Páramos de guasca y cruz
verde específicamente en frailejones. Los principales géneros encontrados
fueron Aspergillus, Penicillium, Cladosporium, Curvilaria, Mucor,
Paicelomyces, Trichoderma, entre otros.
Arias y
Piñeros
(2008)
Cultivo de hongos comestibles del
género Pleurotus sobre residuos
agrícolas de la zona cafetera
Desarrollo de cultivos de hongos comestibles del género Pleurotus sobre
mezclas de residuos de café a nivel de planta piloto con el método de
anaerobiosis.
Rodríguez y
Jaramillo
(2005)
Producción comercial del
Champiñón/Pleurotus ostreatus/ en
pulpa de café
Producción de champiñón Pleurotus ostreatus sobre pulpa de café como una
alternativa ambiental y nutricional de la zona cafetera Lozano
(1989)
Producción de hongos comestibles
en residuos de caña de azúcar. VI
congreso Colombiano de la
Asociación de Técnicos de la Caña
de Azúcar
Evaluación de la producción artesanal de los hongos comestibles Pleurotus
ostreatus; Pleurotus djamur; Pleurotus sajor-caju; Volvariella esculenta;
Volvariella volvacea; Lentinus edodes sobre residuos de caña de azúcar Guzmán et al.
(2003)
Hongos comestibles: otro producto
de las plantaciones de coníferas no
explotado en Colombia
(Agaricales, Aphyllophorales,
Lycoperdales, Pezizales)
Evaluación de las posibilidades de producción de hongos comestibles de los
órdenes Agaricales; Aphyllophorales; Lycoperdales; Pezizales sobre residuos
de coníferas en Colombia. Uribe-López
(1983)
El cultivo de hongos comestibles
sobre desechos agroindustriales
Eficiencias biológicas (kg de hongos frescos/kg sustrato, base seca) X 100 de
hongos del género Pleurotus medidas sobre sustratos a base de pulpa de café
proveniente de despulpado sin agua.
Rodríguez
(1993)

Aprovechamiento de los residuos
sólidos generados en el cultivo e
industrialización del café para la
producción de hongos comestibles
y medicinales
Se determinó la factibilidad técnica y económica de cultivar el hongo
comestible Pleurotus sajor caju, el hongo comestible y medicinal Lentinula
edodes y el hongo medicinal Ganoderma lucidum, sobre sustratos preparados
con los subproductos generados durante el proceso de cultivo e
industrialización del café. Con intervalo de relación C/N de 40 a 60.
Rodríguez
(2003)
Utilización de los desechos del
banano para el cultivo del hongo
comestible Pleurotus ostreatus
Evaluación de los residuos del cultivo de banano para el cultivo artesanal de
hongos comestibles Pleurotus ostreatus. Arteaga-
Hoyos (2002)
Evaluación de algunos residuos
orgánicos como sustrato para el
cultivo de hongos comestibles
Evaluación de residuos de jardín, como sustrato para la producción de dos
especies de hongos, Pleurotus ostreatus y Pleurotus pulmonarius y
estandarización de la producción de semilla.
Garcés-
Molina et al.
(2006)
Fortificación de hongos
comestibles (Pleurotus ostreatus)
con calcio, selenio y vitamina C
Fortificación del hongo comestible Pleurotus ostreatus con calcio y vitamina C
como tratamiento posterior a la cosecha y determinaron vida de anaquel del
hongo.
Cortés et al.
(2007)
Registro de Gimnopilus rugulosus
(Agaricales, Cortinariaceae) de
Colombia
Se registra por primera vez en Colombia Gimnopilus rugulosus (Agaricales,
Cortinariaceae), prospera en zonas húmedas subtropicales der Colombia, entre
los 2.500 y 3.200 m s.n.m. Se ha reportado en Colombia, Costa rica y México.
Cardona et al.
(2005)
Ácidos grasos, ésteres y esteroles
del cuerpo fructífero del hongo
Laccaria laccata
Del extracto en acetato de etilo del hongo comestible Laccaria laccata se
aislaron 3 ácidos grasos, 6 esteres etílicos, 5 esteroles y un triterpeno
ergostánico. Los compuestos se identificaron como ácido palmítico, ácido
linoléico y ácido oléico, hexadecanoato de etilo, 8-octadecenoato de etilo, 9-
octadecenoato de etilo, 9,12-octadecadienoato de etilo, estearato de etilo,
eicosanoato de etilo, ergosta- 2,5,7,9(11), 22-pentaeno, ergosta-5,7,22-trien-3_-
ol (ergosterol), ergosta-7,22-dien-3_-ol, ergosta-7-en-3_-ol, ergosta-
5,7,9(11),22-tetraen-3_-ol y estigmast-5-en-3-ol.
Nieto y
Cucaita
(2007)
Actividad enzimática de hongos y
su patogenicidad
sobre Hypothenemus hampei
Evaluaron seis aislamientos de Beauveria bassiana y Metarhizium anisopliae
para cuantificar la actividad catalítica de cinco enzimas y su relación con la
patogenicidad sobre la broca del café (Hypothenemus hampei).
Delgado et al.
(2001)
Aislamiento y evaluación de la
actividad enzimática de hongos
descomponedores de madera
(Quindío, Colombia)
De hongos colectados de troncos caídos con diferentes estados de
descomposición en la reserva natural La Montaña del Ocaso, se evaluó su
actividad ligninolítica y la celobiosa deshidrogenasa (CDH). Seleccionaron los
hongos con mayor actividad enzimática de troncos con diferente grado de
descomposición: Cookeina sulcipes, Corticiaceae, Xylaria polymorpha y
Earliella sp.
Chaparro et
al. (2009)
Micorrizas Arbusculares del Sur de
la Amazonia Colombiana y su
Relación con Algunos Factores
Fisicoquímicos y Biológicos del
Suelo
Evaluaron la presencia natural de hongos micorrícicos de tipo arbuscular
(HMA) en suelos ácidos de textura franco-arcillosa a arcillosa del sur de la
Amazonia colombiana bajo bosque, rastrojo joven, y praderas establecidas, a
dos profundidades diferentes.
Peña-Venegas
et al. (2007)
Avances en conocimiento y
mejoramiento del hongo
entomopatógenos Beauveria
bassiana para el control de la
broca del café, Hypothenemus
hampei
En Cenicafé se están usando dos aproximaciones para producir un hongo
mejorado para el control de la broca: 1). Estudio de la genómica de Beauveria
bassiana. conocimiento de genes y modificación genética y 2) Exploración y
uso de la diversidad genética de Beauveria bassiana
Góngora
(2009)
Caracterización de una cepa nativa
de Aspergillus niger y evaluación
de la producción de ácido cítrico
Evaluación de la cepa nativa de Aspergillus niger en la producción de ácido
cítrico sobre un sustrato con sacarosa y otro con almidón. Los rendimientos
fueron bajos comparados con los que se reportan en la literatura.
Sáez et al.
(2002)
Contribución al estudio de
microhongos filamentosos en los
ecosistemas páramo de guasca y el
tablazo
Realizaron el aislamiento e identificación de hongos filamentosos originarios
de los páramos de Guasca y El Tablazo, presentes en muestras del suelo y en
las hojas de las plantas Espeletia barclayana y Espeletia killipii.
Chitiva -
Jaramillo et
al. (2009)
Diversidad taxonómica y ecológica
de la entomofauna micófaga en un
bosque alto-andino de la cordillera
oriental de Colombia
Determinaron la estructura y composición de la comunidad de insectos
micófagos encontrados en robledales (Quercus spp.) de la región de Iguaque
(Villa de Leyva-Boyacá). Colectaron 1778 insectos en estado adulto; en
laboratorio criaron 3409 para un total de 5187 individuos distribuidos en 48
morfoespecies, éstas fueron infectadas por los hongos entomopatógenos.
Amat-García
et al. (2001)

Evaluación de la patogenicidad de
los hongos Bauveria basiana y
Metarhizium anisopliae en el
control de la garrapata del ganado
Rhipicephalus micraplus en su fase
parasítica en su estado larval y
ninfal
Evaluación del efecto de los hongos Bauveria basiana y Metarhizium
anisopliae sobre los estados parásitos de la garrapata del ganado Rhipicephalus
micraplus con diferentes concentraciones de esporas de cada una de las
especies de los hongos.
Valbuena y
Alzate (2007)
Problemas fitopatológicos en
especies de la familia Solanáceas
causados por los géneros
Phytophthora, Alternaria y
Ralstonia en Colombia. Una
revisión
Se expone la recopilación de los estudios ubicados realizados en Colombia,
referentes a las enfermedades causadas por tres de los patógenos más
importantes que afectan la familia Solanáceas: Phytophthora, Alternaria y
Ralstonia.
Carreño et al.
(2007)
Evaluación de la susceptibilidad de
hongos endófitos aislados de rosa
(Rosa hybrida) a insecticidas
comerciales
Evaluación de la capacidad de hongos endófitos aislados de rosa como
controladores biológicos de las plagas preferentes en rosa y su resistencia a los
insecticidas.
Corredor et al.
(2007)
El género Pseudocyphellaria
VAIN. (Lobariaceae - ascomycetes
liquenizados) en
Colombia
Descripciones morfoanatómicas, complementadas con datos de pruebas
químicas con K, P, C, KCl y cromatografía en capa fina de las especies
encontradas, comentarios de datos ecológicos y de distribución geográfica.
Moncada y
Forero (2006)
Aislamiento de hongos
solubilizadores de fosfatos de la
rizosfera de arazá (Eugenia
stipitata, Myrtaceae)
Encontraron que los principales solubilizadores del fosfato de calcio fueron
Trichoderma aureoviride, Aspergillus aculeatus, Trichoderma cepa 1 y
Trichoderma cepa 2 y para el fosfato de hierro: Aspergillus oryzae,
Paecilomyce cepa 3, Gongronella butleri y Fusarium oxysporum.
Vera et al.
(2002)
Macromicetos observados en
bosques del Departamento de
Caldas: su influencia en el
equilibrio y la conservación de la
biodiversidad
Se analizaron algunos patrones de diversidad y distribución de macrohongos en
relación con el paisaje antropogénico en varios tramos de bosques del
departamento de Caldas. Se hace la relación de los géneros encontrados, como
indicadores para el monitoreo biológico de la Eco-región. Los géneros
encontrados corresponden al orden Agaricales con 12 familias y 35 géneros;
seguido del orden Polyporales con cinco familias y 11 géneros.
Montoya et al.
(2010)
Influencia del sustrato utilizado
para el crecimiento de hongos
comestibles sobre sus
características nutraceúticas
El objetivo del presente estudio fue el de evaluar el efecto del sustrato sobre las
propiedades nutricionales o nutriceúticas de hongos del género Pleurotus.
Como resultado se determinó que efectivamente la composición de los hongos
en cuanto al contenido de proteínas netas, fibra, humedad, cenizas,
carbohidratos y grasas totales varía con el sustrato empleado.
Nieto y
Chegwin,
(2010)
Dehidroergosterol: un artefacto
generado durante el proceso de
extracción de esteroles en el hongo
Pleurotus sajor-caju
Durante el estudio químico de la fracción esterólica del hongo Pleurotus sajor-
caju se estableció que el dehidroergosterol (ergosta-5,7,9 (11),22-tetraen-3-ol)
presente en el extracto.
Rivera et al.
(2005a)
Growth, fruiting and
lignocellulolytic enzyme
production by the edible mushroom
Grifola frondosa (maitake)
Efecto de las actividades enzimáticas sobre los diferentes estados de
producción del hongo G. frondosa en diversas mezclas de materiales
lignocelulósicos.
Montoya et al.
(2011a)
Evaluación in vitro de celulasas
producidas por cepas nativas de
Trichoderma reesei, Cladosporium
herbarum y Aspergillus niger
La celulosa es el polisacárido más abundante en la naturaleza, los organismos
capaces de degradarlo son principalmente hongos y bacterias, ellos son
reconocidos por la habilidad de producir enzimas no sólo celulasas sino
también amilasas, proteasas y peptidasas entre otras; que les permite el
reciclado de material orgánico de nuevo al suelo. La hidrólisis de ésta se
realiza mediante un complejo enzimático llamado celulasas, constituido
básicamente por tres enzimas: Endoglucanasa, exoglucanasa y β-glucosidasas.
Gutiérrez
Ramírez et al.
(2012)
Macromicetos en Zona Rural de
Villavicencio
Mediante colecciones puntuales se recolectaron 30 especímenes en la zona
rural de la ciudad de Villavicencio, que posee un paisaje de Piedemonte. En el
muestreo predominaron los basidiomicetos. El orden Polyporales y la familia
Polyporaceae fueron los mejor representados y los géneros más frecuentes
Trametes y Auricularia. Se espera que este estudio contribuya al conocimiento
de la diversidad micológica en los Llanos Orientales de Colombia.
Ortiz-Moreno
(2010)
Modeling Grifola frondosa fungal
growth during solid-state
fermentation
Aplicación de modelos matemáticos a fin de describir el crecimiento y
desarrollo de la fase vegetativa del hongo G. frondosa sobre un sustrato sólido
con materiales lignocelulósicos.
Montoya et al.
(2011b)

Production of Biomass,
Polysaccharides, and Ganoderic
Acid using Non-conventional
Carbon
Production of Biomass, Polysaccharides, and Ganoderic Acid using Non-
conventional Carbon Sources under Submerged Culture of the Lingzhi or
Reishi Medicinal Mushroom, Ganoderma lucidum (W.Curt.:Fr.)P. Karst.
(Higher Basidiomycetes)
Zapata et al.
(2012)
2. 2. Materiales lignocelulósicos
El desarrollo de los hongos de pudrición blanca requiere de sustratos compuestos de materiales que le
proporcionen las características físicas y la composición química adecuada para su crecimiento y desarrollo
en condiciones controladas. Los materiales lignocelulósicos cumplen con las condiciones generales
requeridas por estos hongos. Estos materiales son los principales constituyentes de los vegetales,
representados por lignina, celulosa, hemicelulosa y pectina en sus paredes celulares y el almidón utilizado
como reserva energética de la célula. Estos materiales son los que descomponen los hongos de pudrición
blanca. La celulosa, componente mayoritario de la pared celular de todas las plantas, es el material natural
más abundante en el planeta. Aunque su estructura química es simple, por estar compuesta de unidades de
glucosa y ser una cadena lineal tiene diversas propiedades químicas que han sido de interés científico y
técnico. Asimismo, la lignina que es otro componente importante de los materiales lignocelulósicos se
localiza entre los carbohidratos que componen las maderas formando una red a través de enlaces covalentes
cross-liking (ver Figura 2). Por ejemplo, la lignina en las maderas blandas los enlaces covalentes se dan entre
la lignina y ambas hemicelulosas (glucomanano y Xilano) y en maderas duras el contenido de glucomanano
es bajo, por lo que el enlace dominante se da entre la lignina y el Xilano (Ek et al., 2009). Además de
considerar que, estos materiales lignocelulósicos se constituyen como uno de los materiales más acumulables
del planeta, debido a que en este grupo se encuentran los residuos agroindustriales, forestales y residuos
sólidos urbanos, entre otros (Papinutti et al., 2003; Sánchez y Cardona, 2007).
Los carbohidratos como componentes importantes de los materiales lignocelulósicos, son muy versátiles,
ya que hacen parte de una gran variedad de productos en la industria, como la alimentaria, farmacéutica,
textil, papelera y en la producción de empaques biodegradables, entre otros. De la misma manera, los
carbohidratos juegan un rol muy importante en el establecimiento y evolución de la vida en la tierra por la
creación directa de una fuente de energía química a partir de la energía solar. Los carbohidratos son
producidos durante el proceso de fotosíntesis. Los carbohidratos están distribuidos en plantas y animales y
cumplen diversas funciones: i) reserva de energía como el almidón, fructanos y glucógeno, ii) materiales
estructurales como la celulosa, quitina, xilanos y mananos, iii) sustancias protectoras, algunas plantas
producen carbohidratos repelentes de insectos o inhibidores de formación de hielo como el arabinoxilano en
la supervivencia de los cereales en climas templados, iv) fracciones de polisacáridos, los cuales pueden ser
oligosacáridos conjugados con proteínas (glicoproteínas) o con lípidos (glicolípidos) como componentes
importantes de la membrana celular que pueden servir como sondas con las cuales la célula interactúa con su
entorno, v) como agentes de transferencia de información, como los ácidos nucleicos (Cui, 2005).
Los materiales lignocelulósicos se clasifican de acuerdo a su origen. En la Tabla 3 se muestra una
clasificación general de los principales materiales con alto contenido de biomasa lignocelulósica,
componentes básicos para el desarrollo fúngico. Los residuos agrícolas compuestos por pajas, rastrojos,
capachos y otros residuos como las pulpas de café, desechos de cultivos de hortalizas (de corta duración),
leguminosas y oleaginosas, etc. y en general, los más representativos de la región cafetera y el país se
mencionan en la Tabla 5. Los materiales provenientes de maderas, se componen de dos subgrupos, los que
pertenecen a las maderas duras, provenientes principalmente de angiospermas, como el álamo, el eucalipto,
el aspen, el roble, el arce, entre otros. Las maderas blandas, en cambio corresponden a maderas de coníferas,
ubicándose los árboles de gimnospermas como el pino, el abeto, la pícea, el alerce, entre otros, haciendo la
consideración que las maderas blandas contienen más lignina que las maderas duras. La biomasa herbácea
proviene de las plantas que no generan madera, sus tallos son verdes, en general se consideran los follajes,
pastos, etc. Los residuos sólidos urbanos son muy heterogéneos y en general contienen un alto contenido de
material lignocelulósico, como residuos de papel, cartón, cáscaras de frutas y vegetales en general, residuos
de jardinería, entre otros (Sánchez y Cardona, 2007).

Tabla 3. Clasificación de los principales materiales con alto contenido de biomasa lignocelulósica.
Tipo de Biomasa Ejemplos
Residuos Agrícolas Pajas: Trigo, arroz, cebada, algodón, sorgo, millo, cogollos de caña, etc.
Otros residuos: rastrojo de maíz, capacho de coco, algodón de desecho, zoca de café, pulpa de
café, vainas de leguminosas. Residuos
Agroindustriales Bagazos: caña, sorgo dulce.
Otros residuos: cascarilla de arroz, huesos de aceituna, borra, cascarilla y película plateada de
café, cascarilla de cacao y de cereales como arroz, sorgo, maíz; tortas de oleaginosas (algodón,
soya y palma) Madera y Residuos
Forestales Madera dura: álamo, eucalipto, aspen.
Madera blanda: pino, picea.
Aserrín, ramas: de roble, mezclas de industrialización de madera en construcción de muebles. Residuos Lignocelulósicos Papel: periódico, de oficina utilizado, lodo de papel reciclado Biomasa Herbácea Pastos: pasto de pradera, alpiste rosado, pasto Bermuda de la variedad costera, fleo de los prados,
Heno de alfalfa. Residuos Sólidos Urbanos Fracción celulósica: papel, cartón, madera, residuos de jardinería, cáscaras de frutas y verduras.
Modificado de Sánchez y Cardona (2007)
En la Tabla 4 se relaciona la composición en base seca de diferentes materiales lignocelulósicos,
incluyendo algunos residuos de la industria pecuaria. Se observa que son muy representativos los contenidos
de los polímeros celulosa y hemicelulosa en las maderas duras y blandas, bagazo de caña, residuos de papel,
pajas de cereales (en este caso de trigo), tusa de maíz y pastos (Sánchez y Cardona, 2007). Estos altos
contenidos de celulosa en todos estos materiales, los convierten en relevantes para la obtención de productos
de valor agregado por fermentación de los azúcares derivados de la hidrólisis de este polisacárido. De igual
forma, todos estos materiales contienen lignina, la cual puede ser removida o tratada con hongos de
pudrición blanca a través de sus enzimas oxido-reductasas (Kirk y Farrel, 1987).
Tabla 4. Composición porcentual (en base seca) de varios materiales lignocelulósicos
Materiales Celulosa
(%)
Hemicelulosa (%) Lignina (%)
Bagazo de caña 50 25 25
Madera dura 40-55 24-40 18-25
Madera blanda 45-50 25-35 25-35
Cubierta de la nuez 25-30 25-30 30-40
Tusa de maíz 45 35 15
Pastos 25-40 35-50 10-30
Papel 85-99 0 0-15
Paja de trigo 30 50 15
Basura surtida 60 20 20
Hojas 15-20 80-85 0
Hebras de semilla de algodón 80-95 5-20 0
Papel periódico 40-55 25-40 18-30
Papeles de desecho de pulpas químicas 60-70 10-20 5-10
Sólidos primarios de aguas residuales 8-15 - 24-29
Desechos de cerdo 6 28 -
Estiércol bovino sólido 1,6-4,7 1,4-3,3 2,7-5,7
Pasto bermuda de la variedad costera 25 35,7 6,4
Pasto de pradera 45 31,4 12
Fuente: Sánchez y Cardona (2007)
En la Figura 1 se muestra un esquema simple del complejo lignocelulósico de los tejidos vegetales donde
se puede observar el arreglo de las fibras de celulosa, hemicelulosa y la matriz lignina-hemicelulosa
haciendo parte de la pared celular primaria y secundaria. Asimismo, en la Figura 2 se muestra un esquema de
la distribución molecular de la lignina y los polisacáridos en los materiales lignocelulósicos, donde se
evidencia la distribución de las diferentes concentraciones de lignina en la estructura. La lignina se deposita
en las zonas precedidas de una deposición de carbohidratos. El primer depósito de lignina inicia en las
esquinas de la pared celular y continua en la laminilla media. El segundo estado de lignificación ocurre

después de la deposición de celulosa y hemicelulosa en la capa S2. La principal lignificación se da en la
etapa S3, aunque la mayor concentración de lignina se presenta en la capa S2 (media) tanto para maderas
duras como blandas y su asociación en red parece más claramente con el xilano que con mananos y celulosa
(Ek et al., 2009).
Figura 1. a) Estructura simplificada de la pared celular que muestra la laminilla media (ML), pared primaria (P),
capas secundarias de la pared celular (S1, S2, S3) y lignina en forma de capas verrugosas sobre la pared celular (W).
Las flechas indican la orientación de las microfibrillas de celulosa en las capas individuales de la pared celular
secundaria. La microfotografía de la derecha, muestra la localización de la lignina en la laminilla media intercelular
(ML) y la laminilla de la esquina entre las traqueidas de pino, b) y las fibras de abedul, c). Las capas S1 y S3 no
aparecen en las microfotografías. Fuente: Ek et al. (2009)
Las materias primas de mayor utilización en los procesos de fermentación son los subproductos
agropecuarios y agroindustriales, por su gran disponibilidad y por su bajo precio. En Colombia, por tener una
alta vocación agrícola y por ser un país tropical, existe una gran cantidad de residuos que pueden ser
utilizados como materias primas para los medios de cultivo, en la Tabla 5 se muestra la distribución por
grupos de materiales orgánicos naturales disponibles en Colombia. En el Departamento de Caldas, los
materiales lignocelulósicos más representativos son: i) residuos del cultivo e industrialización del café, como
la pulpa, soca, cascarilla, borra de café; ii) residuos agroindustriales, como el capacho de coco y otros
residuos de frutas, iii) residuos de la agroindustria de la caña de azúcar y iv) residuos de la agroindustria de
la madera, especialmente del beneficio del roble. Algunos de estos residuos están siendo utilizados como
combustible en los mismos procesos productivos, como materias primas de procesos de compostaje o en
otros usos secundarios en avícolas, caballerizas, entre otros.
2.2.1. Celulosa
La celulosa, que es un biopolímero de D-glucosa, es el material orgánico más abundante sobre la corteza
terrestre. Las plantas sintetizan la celulosa como material estructural para soportar su peso. Las estructuras
longitudinales de celulosa, llamadas micro fibrillas, forman haces por los puentes de hidrógeno que se crean
entre los numerosos grupos OH de los anillos de glucosa. En la celulosa las unidades de D-glucosa están
unidas por enlaces glucosídicos β(1,4), que le confieren una disposición bastante rígida y muy estable. En la
Figura 3 se muestra la estructura parcial de la celulosa.

Figura 2. Distribución de la lignina y polisacáridos a nivel molecular en los materiales lignocelulósicos. Fuente: Ek
et al. (2009)
Tabla 5. Principales materiales orgánicos naturales disponibles en Colombia
Materiales (según cultivo) Ubicación Lignocelulósico proteico
Caña de azúcar (bagazos de
ingenio y de trapiche, melaza,
desechos de cultivo)
Valle del Cauca, Caldas,
Santander, Antioquia X
Café (borra, cascarilla, pulpa,
mucílago, zoca)
Caldas, Risaralda, Quindío,
Antioquia, Tolima, Nariño,
Huila
X
Cereales (cascarilla, jarabes,
residuos de cultivo)
Tolima, Antioquia, Valle,
Llanos Orientales X
Leguminosas (residuos de
cultivo y de cosecha)
Tolima, Valle, Eje cafetero,
Santander, Cundinamarca,
Antioquia
X X
Oleaginosas (residuos de
cosecha y tortas)
Tolima, Antioquia, Costa
Atlántica, Valle del Cauca X X
Maderas (aserrín, cascarilla,
viruta)
Caldas, Antioquia, Chocó,
Cundinamarca X
Frutales (cáscaras, semillas y
residuos de cosecha) Todo el país X
Hortalizas (residuos de cosecha,
hojas, tallos)
Caldas, Cundinamarca,
Antioquia X
Centrales de sacrificio de
animales (sangre, plumas,
cascos, cuernos)
Todo el país X
Cacao (cascarilla y residuos de
cultivo)
Caldas, Antioquia,
Cundinamarca X X
Figura 3. Estructura de la celulosa mostrando los enlaces glucosídicos β(1,4). Fuente: Sánchez y Cardona (2007)

La longitud del polímero es altamente variable y depende del organismo del cual la celulosa provenga,
así como de la edad y estado metabólico al momento de su extracción. Un grado de polimerización promedio
de alrededor de 10.000 se determinó para la celulosa de la madera (Zabel y Morrel, 1992b). Cada monómero
de glucosa presenta una rotación de 180º respecto de los residuos contiguos, estabilizada por la formación de
puentes de hidrógeno intramoleculares. Las microfibrillas están conformadas por zonas cristalinas donde las
moléculas de celulosa se hallan ordenadas y regiones para cristalinas donde se hallan desorganizadas. La
estructura cristalina de la celulosa hace que la pared de la célula sea anisótropa (que presentan más de un
índice de refracción de la luz en función de la dirección de vibración o polarización de las ondas luminosas)
y por consiguiente con doble refracción (birrefringente) cuando se mira con luz polarizada (Meier, 1955).
Se consideran dos tipos de celulosa: la nativa o cristalina, caracterizada por un alto grado de cristalinidad
u ordenamiento y de polimerización, resultando así insoluble (ej.: Avicel, fibras de algodón, papel de filtro,
etc.); y la celulosa modificada, la cual resulta soluble como la celulosa amorfa, carboximetilcelulosa, y los
celooligosacáridos, en los cuales el grado de cristalinidad y el grado de polimerización es menor (Levin,
1998; Sánchez y Cardona, 2007).
En resumen las fibras de celulosa están ordenadas en cristales de tres dimensiones. La primera dimensión
está dada por enlaces covalentes, impuestos por puentes de hidrógenos a lo largo de la cadena de celulosa,
que le da la longitud final de la fibrilla, la cual depende de su grado de polimerización. La segunda
dimensión está constituida por los puentes de hidrógeno, la estructura se pliega en hojas. La formación de
puentes entre las hojas de celulosa y las fibrillas provocadas por las interacciones-X y las fuerzas de Van der
Waals generan la tercera dimensión de la celulosa (Ek et al., 2009). Hecho que hace que la celulosa sea una
sustancia altamente estable frente a procesos químicos como la hidrólisis, donde se requieren altas
concentraciones de ácidos o bases para obtener éxito en la ruptura de la estructura. Los microorganismos
juegan un rol importante en el reciclado del carbono celulósico, ya que hay un gran número de
microorganismos, que incluyen bacterias y hongos, los cuales son capaces de romper las unidades de
celulosa hasta monosacáridos bajo condiciones aeróbicas o anaeróbicas. Las bacterias anaeróbicas incluyen
Bacteroids cellulosolvens, Bacillus sp., Clostridium (C. celluloliticum, C. cellulovorans, C. papyrosolvens,
C. Thermocellum), Fibrobacter succinogenes, Cellvibrio gilvus, Ruminococcus flavefaciens, etc., levaduras
como Candida (C. wickerhamii, C. lucitance), hongos del rumen como Neocallimastix frontalis (Leschine,
1995; Joshi y Pandey, 1999b), y todos los hongos aeróbicos filamentosos micro y macromicetos (Carlile et
al., 2001). Todos estos organismos emplean exoenzimas para la degradación del polímero de celulosa, entre
las que se destacan la endoglucansa, exocelobiohidrolasas y β-glucosidasa de las cuales se hace referencia
más adelante, y la celobiosa quinasa y fosfo- β-glucosidasa que también pueden estar presentes en muchos de
estos organismos mencionados.
2.2.2. Hemicelulosa
La hemicelulosa junto con la celulosa son tanto carbohidratos estructurales de las plantas como los
componentes mayoritarios de la pared celular. Las hemicelulosas son encontradas en la matriz entre las
fibras de celulosa y la pared celular, ubicándose entre la celulosa y la lignina. Los tipos y cantidades de
hemicelulosas son diversas en la naturaleza, dependiendo de los materiales, el tipo de tejido, estado de
desarrollo, condiciones de desarrollo, almacenamiento y métodos de extracción. Las hemicelulosas aparecen
como heteropolisacáridos de diversas hexosas y pentosas, principalmente D-xilosa, L-arabinosa, D-manosa,
D-glucosa, D-galactosa, y ácido D-glucururónico, que aparecen en las paredes celulares en forma amorfa y
poseen una especie de columna vertebral formada por una cadena plana de azúcares unidos casi siempre por
enlaces β (1,4) de menor longitud que la de celulosa con grado de polimerización 100-200 (ver Figura 4) de
la que pueden salir ramificaciones muy cortas, generalmente de un solo azúcar de longitud. Las
hemicelulosas se clasifican usualmente de acuerdo a los azúcares residuales presentes (Ek et al., 2009). Los
tipos y cantidades de las hemicelulosas contenidas en las paredes celulares de las maderas de angiospermas y
coníferas son diferentes. Estas últimas contienen menos hemicelulosas constituidas principalmente por
glucomananos y galactoglucomananos (Moore, 1998b). En los glucomananos la cadena principal está
formada por residuos alternos de β-D-glucopiranósido y β-D-manopiranósido; en los galactoglucomananos

sustituyentes de D-galactopiranosil terminales están asociados al O-6 de manosa o glucosa (Zabel y Morrel,
1992a; Joselau y Ruel, 1994; Goodenough, 1996; Sánchez y Cardona, 2007; 2008).
Los xilanos son las principales hemicelulosas en la madera de angiospermas (constituyen del 10 al 35%
del peso seco) (Zabel y Morrel, 1992a; Joselau y Ruel, 1994). Consisten de una cadena principal de unidades
β-D-xilopiranósido con uniones β(1,4) y, dependiendo de su origen, esta cadena puede tener diferentes
sustituyentes y cadenas laterales. Se consideran tres familias principales de xilanos (Coughlan y Hazlewood,
1993; Goodenough, 1996): Xilano de la madera de angiospermas, de la madera de coníferas y el xilano de
los pastos.
Las hemicelulosas cubren y unen a las microfibrillas de celulosa en una matriz común. La corta
extensión de las cadenas, que incrementa la solubilidad de las hemicelulosas y la posición expuesta de las
mismas en la superficie de las microfibrillas, explicaría por qué este polímero está entre los primeros
componentes de la pared celular atacados por los hongos causantes de pudrición (Zabel y Morrel, 1992b;
Goodenough, 1996).
(A)
(B)
Figura 4. (A) Estructura de xilano en maderas blandas, con una cadena adicional de L-arabinofuranosa; (B)
Estructura de xilano en maderas duras, con 4-O-metil-D-glucuronxilano. Fuente: Ek et al. (2009)
2.2.3. Lignina
La lignina es el material aromático renovable más abundante del planeta, es un biopolímero aromático,
no polisacárido (Kirk y Farrel, 1987). Es un compuesto amorfo e insoluble en agua, de alto peso molecular y
tridimensional. La lignina se encuentra principalmente en la pared celular de las plantas formando parte de
una matriz ordenada con las microfibrillas de la celulosa. Los precursores de la lignina son denominados
monolignoles: alcoholes sinapílico (3,5-dimetoxi-4-hidroxicinamil), p-cumarílico (p-hidroxicinamilico) y
coniferílico (4-hidroxi-3-metoxicinamil alcohol), cada uno aportando unidades de siringil, p-hidroxifenil y
guaiacil respectivamente en el biopolímero (Adler, 1977; Buswell y Odier, 1987; Reid, 1995) (ver Figura 5).
La copolimerización de los radicales libres de estos alcoholes produce polímeros heterogéneos
ópticamente inactivos, entrelazados y altamente polidispersos. Las unidades de guaiacil se encuentran en
mayor proporción conformando las ligninas de las gimnospermas, mientras que las ligninas de las

angiospermas están conformadas por cantidades aproximadamente iguales de guaiacil y siringil. Ambos tipos
de ligninas contienen pequeñas cantidades de unidades de p-hidroxifenil (Adler, 1977; Kirk y Farrel, 1987).
El proceso de polimerización de los monómeros se inicia con una oxidación que da semiquinonas, luego
estos radicales libres reaccionan unos con otros y forman enlaces covalentes entre C-O o C-C, siendo el
predominante el enlace β-O-4, el cual usualmente con adición de agua da una forma más estable de enlace
éter β-O-4. Alrededor del 40-60% de todas las uniones que conforman la lignina corresponden a enlaces tipo
éter. En la Figura 6 se observa un acercamiento de la estructura de la lignina de maderas blandas y el
segmento que se repite en las diferentes estructuras de lignina presentes en los otros productos. Tres tipos de
lignina pueden ser reconocidos según el contenido de monolignoles: i) lignina de maderas blandas
(gimnospermas, eudicotiledonas, etc.) o guayacil lignina que está compuesta mayoritariamente de alcohol
coniferílico y pequeñas cantidades de alcohol p-cumaríclico y sólo trazas de alcohol sinapílico. ii) lignina de
maderas duras o lignina siringil-guayacil contienen los alcoholes coniferílicos y sinapílicos en proporciones
iguales con pequeñas cantidades de alcohol p-cumarílico. Este tipo de lignina está presente en angiospermas
eudicotiledoneas y genotipos del grupo de gimnospermas que estén evolucionando a angiospermas y iii)
ligninas de hierbas y pastos o lignina hidroxi-fenol, guayacil y siringil, contienen todos los monolignoles y
contienen un alto contenido de p-cumarílico. Este tipo de lignina está presente en árboles de palma,
musáceas, helechos, etc.
Figura 5. Monómeros precursores de la molécula de lignina. Fuente: Bommarius y Riebel (2004a).
La lignina imparte resistencia a la degradación microbiana de la madera. La íntima asociación espacial
existente entre los polisacáridos y la lignina dentro de las paredes celulares de la madera provee una barrera
protectora que impide la degradación de la celulosa. Una vez que la lignina que rodea a las fibrillas de
celulosa es removida o modificada, la celulosa resulta más accesible a las enzimas microbianas y puede ser
degradada eficientemente. Así, la lignificación en muchas plantas puede ser un mecanismo de resistencia a la
enfermedad y se produce como un mecanismo de defensa hacia los hongos patógenos o en respuesta a
heridas (Blanchette, 1995). Hasta el momento, los únicos organismos capaces de mineralizar eficientemente
la lignina hasta CO2 y agua como productos finales son los hongos causantes de pudrición blanca
(Leonowicz et al., 1999). Estos hongos en su mayoría pertenecen a los ordenes Aphyllophorales y Agaricales
(Pelaez et al., 1995) y poseen un complejo de enzimas oxidasas y peroxidasas que catalizan las primeras
reacciones que rompen uniones dentro de la compleja molécula de lignina, generando moléculas más
pequeñas. Los productos de degradación se incorporan luego a los ciclos metabólicos del organismo
originando como producto final CO2 (Kirk y Farrel, 1987). Se ha demostrado que la lignina como sustrato
único no es una fuente aprovechable para el crecimiento de los hongos, sino que es necesaria la presencia de
un co-sustrato (Kirk et al., 1976).
Alcohol p-cumarílico Alcohol coniferílico Alcohol sinapílico
Notación de la estructura
general simplificada

(A)
(B)
Figura 6. (A) Estructura sugerida de lignina de madera blanda. Las ligninas de maderas duras y monocotiledoneas
difieren mucho en el contenido de grupos metoxilo (-OCH3). (B) Estructura que se repite en las diferentes ligninas
encontradas en los materiales lignocelulósicos. Fuente: (Ek et al.(2009)
2.2.4. Otros polímeros presentes en los materiales lignocelulósicos
El almidón y las pectinas pueden estar presentes en el complejo lignocelulósico, dependiendo de la
procedencia. El almidón es un homopolisacárido de unidades de D-glucosa unidos con enlaces glicosídicos
α(1,4). El almidón tiene dos componentes, amilosa y amilopectina. La amilosa es una molécula de peso
molecular cercano a 5000 y naturalmente está conformada por un grupo terminal hemiacetal reducido y otro
no reducido en cada cadena. La amilopectina es un polisacárido con enlaces α(1-4) y α(1-6) cada 25-30
unidades de glucosa. Su peso molecular es diferente para distintas fuentes de almidón.
El almidón se encuentra ampliamente distribuido en microalgas, algas y plantas superiores, se
constituye en el mejor material de almacenamiento. Para los organismos heterótrofos es la fuente más común
de energía. Una gran cantidad de microorganismos tienen la capacidad de utilizar este polisacárido (Joshi y
Pandey, 1999b). La hidrólisis del almidón en glucosa en los sistemas biológicos es llevada a cabo por
múltiples enzimas, como α-amilasa, β-amilasa, glucoamilasa, maltosa hidrolasa (β-amilasa), y otras enzimas
como la 1-6, glucosidasa, la dextrinasa límite y las pulanasas (Takasaki, 1976; Hyun y Zeikus, 1985).
Las pectinas son otro componente importante de la pared celular de las plantas, lo que lo constituye en
un polisacárido que hace parte del complejo lignocelulósico y es de los principales constituyentes de la fibra
soluble. Las sustancias pécticas se localizan principalmente en la laminilla media y la pared primaria de las
células vegetales. En los árboles la laminilla media de los elementos xilemáticos jóvenes está constituida

mayoritariamente por pectina, pero al envejecer éstos, se torna altamente lignificada. La pectina está
constituida principalmente por ácido D-galacturónico, un derivado de la galactosa donde el grupo hidroxilo
del carbono 6 ha sido oxidado para formar un grupo carboxílico y, en menor medida está compuesto de
ramnosa, arabinosa y galactosa. La importancia comercial de las pectinas se centra en su uso como
gelificante en el procesamiento de diversos alimentos, entre los que se encuentran los yogurts, conservas,
jaleas y mermeladas (Rodríguez-Palenzuela et al., 2000; Campbell y Farrell, 2004). Las pectinas tienen un
papel esencial en la estabilidad de la pared celular al actuar como material aglutinador de las fibras de
celulosa (Pasquel, 2001).
2.2.5. Pretratamientos de los materiales lignocelulósicos
Los residuos lignocelulósicos son potencialmente utilizables como materias primas para la obtención de
diversos productos. Actualmente, para la producción de etanol de biomasa lignocelulósica es necesario
realizar una hidrólisis previa de la celulosa y hemicelulosa para la obtención de azúcares fermentables y
separación de la lignina residual. El fin del pretratamiento es romper la estructura de la lignina y
desestabilizar la celulosa cristalina con el propósito de incrementar la accesibilidad de las enzimas a la
estructura de la celulosa durante la hidrólisis (Mosier et al., 2005a; Shi et al., 2009). Los materiales
lignocelulósicos tienen diferentes características físico-químicas, lo que hace necesario adoptar diferentes
tecnologías de pretratamiento según las características de cada material crudo. Los aspectos generales a tener
en cuenta para un efectivo pretratamiento del material lignocelulósico a fin de conseguir material rico en
monosacáridos fermentables son: la composición química del material lignocelulósico, la edad del cultivo, el
tiempo de cosecha, los requerimientos de digestibilidad final de los materiales, la degradación de los
azúcares obtenidos, la cantidad de compuestos tóxicos, el tamaño de partícula, residuos sólidos generados del
pretratamiento, contenido de humedad, compatibilidad del material pretratado con el proceso de
fermentación posterior, recuperación de la lignina y mínimos requerimientos de calor y energía. Para lo cual
se puede someter el material, por ejemplo a pretratamientos alcalinos con cal, explosión de la fibra con
amonio (AFEX) y percolación con reciclado del amonio (ARP), reducen el contenido de lignina de residuos
agrícolas, aunque no son tan apropiados para el pretratamiento de maderas blandas. Pretratamientos con
ácido-base han presentado mejores rendimientos para materiales lignocelulósicos, aunque más costosos
(Mosier et al., 2005b; Chandra R.P. et al., 2007). Las tecnologías de pretratamiento de la biomasa
lignocelulósica se presentan en la Tabla 6.
Tabla 6. Tecnologías de pretratamiento a biomasa lignocelulolósica
Pretratamiento Descripción
Materiales
pretratados
(ejemplos)
Referencia
Biológico
Empleando hongos de pudrición blanca, parda y blanda selectivos por la lignina a menor costo de degradación para la obtención de azúcares fermentables, menor
impacto ambiental y menor producción de sustancias tóxicas. Sin embargo, aún es
necesario mejorar la capacidad de los pretratamientos biológicos con hongos basidiomicetos a fin de mejorar su habilidad para delignificar los materiales
lignocelulósicos en menor tiempo y más eficientemente.
Maderas, pajas,
bagazos, semillas
Kumar y
Wyman (2009)
Físicos
Los materiales son sometidos a:
Conminución mecánica: reducción del tamaño de partícula y cristalización del
material a fin de incrementar la superficie específica y reducir el grado de polimerización. Se utilizan molinos de diferentes tipos según los requerimientos
finales, son costosos por el alto consumo de energía e infraestructura.
Extrusión: los materiales son sometidos a calentamiento, mezclado y agitación, obteniéndose una modificación física y química del material cuando es pasado
por el extrusor, por desfibrilación, fibrilación y corte de las fibras del material,
incrementando al final la accesibilidad de los carbohidratos al ataque enzimático. Esta es una tecnología promisoria, nueva y costosa.
Maderas duras, bagazo de caña,
pajas
Karunanithy et al. (2008);
Alvira et al.
(2009); Sánchez y Cardona
(2007)

Químicos
Pretratamiento con álcali: su eficiencia depende del contenido de lignina en el material. Este pretratamiento incrementa la digestibilidad de la celulosa y es más
efectivo para solubilizar la lignina, aunque genera menor solubilidad de la
celulosa y hemicelulosa. Este pretratamiento puede ser más efectivo para residuos agrícolas, que para residuos de maderas. Se emplean hidróxidos de
sodio, potasio, calcio y amonio. La selección de la base depende de la
composición del material. El hidróxido de sodio incrementa la digestibilidad de las maderas duras y reduce los contenidos de lignina, disminuyendo el grado de
polimerización y cristalinidad. Con hidróxido de calcio se remueven sustancias
amorfas como la lignina, la cual incrementa el índice de cristalinidad. También, remueve grupos acetilo de la hemicelulosa e incrementa la digestibilidad de la
celulosa. Los pretratamientos con cal son menos costosos comparados con los
realizados con NaOH y KOH.
Pertratamiento con ácido: El objetivo de este pretratamiento es la
solubilización de la fracción de la hemicelulosa y hacer la celulosa más accequible al ataque enzimático. El tratamiento con ácido diluido aparece como
el método más favorable a escala industrial. Las ventajas de este pretratamiento
se centran en la disponibilidad de reactores para hacerlo, a altas temperaturas y cortos períodos de tiempo; presenta ventajas en cuanto a la alta solubilización de
la hemicelulosa, principalmente el xilano, convirtiendo la hemicelulosa
solubilizada en azúcares fermentables. Debe tenerse en cuenta la temperatura a fin de disminuir la degradación de algunos azúcares hasta furfural y la
degradación de los compúestos aromáticos de la lignina, los cuales pueden
afectar el proceso de fermentación. El ácido más utilizado es el sulfúrico, aunque se utilizan el ácido clorhídrico, nítrico, fosfórico y ácidos orgánicos como
fumárico y maleico. La ventaja comparativa del uso de los ácidos orgánicos
frente al sulfúrico es la menor formación de furfural durante la hidrólisis.
Ozonólisis: el ozono es un potente oxidante y presenta una alta eficiencia en la
deslignificación. La deslignificación ocurre generalmente a temperatura de salón y presión normal y no se dirige hacia la formación de compuestos inhibitorios
que afecten la subsecuente hidrólisis y fermentación. Ha sido utilizado en el
pretratamiento de pajas de cereales y otros residuos agrícolas. Sin embargo, se requieren grandes cantidades de ozono, lo que hasta al momento lo hace inviable
económicamente.
Organosolvatación: Es una estrategia promisoria. Mezclas de solventes
orgánicos acuosas o no, pueden ser utilizadas, entre las cuales se incluyen
metanol, etanol, acetona, etilen glicol y tetrathidrofurfuril alcohol, solubilizan la lignina y viabilizan la celulosa para la posterior hidrólisis enzimática. Un
aspectos importante de este pretratamiento es la recuperación de la lignina pura
como un subproducto. Este pretratameinto ha sido combinado con tratamiento catalítico con ácidos clorhídrico, sulfúrico, oxálico y salicílico a fin de
solubilizar la hemicelulosa. Los altos costos de este proceso son la principal
restricción de este pretratamiento, es necesario recuperar los solventes para bajar costos de operación y evitar riesgos de inhibición en los procesos de
fermentación posteriores.
Pretratameinto con líquidos iónicos: es un pretratamiento que se encuentra aún en fase de laboratorio. Se ha realizado con 1-etil-3-metil imidazolim dietil
fosfato, presenta excelentes propiedades de estabilidad química y térmica, trabaja
a bajas temperaturas. Los carbohidratos y la lignina pueden ser disueltos simultáneamente, generando un desorden efectivo sin formación de sustancias
tóxicas. No ha sido aplicado industrialmente y hasta el momento presenta altos
costos de operación, ya que se presentan dificultades en la recuperación de la hemicelulosa y la lignina de la solución, después del pretratamiento.
Maderas duras,
residuos agrícolas, rastrojo de maíz,
pajas,
Mosier et al. (2005b); Kim y
Holtzapple
(2006); Kootstra et al. (2009);
Sun y Cheng
(2002); Li et al. (2009)

Fisico-químicos
Explosión con vapor: SO2-explosión con vapor. La explosión con vapor es uno de los pretratamientos más eficientes para la biomasa lignocelulósica. Este
proceso combina fuerzas mecánicas con efectos químicos por el incremento y la
caída de presión. Ocurre una autohidrólisis de los grupos acetil presentes en la hemicelulosa formando ácido a altas temperaturas; mientras que, los efectos
mecánicos reducen las fibras generando una separación de la misma durante la
descompresión. En combinación con la hidrólisis parcial y solubilización de la hemicelulosa, la lignina es redistribuida y removida del material, quedando la
celulosa expuesta a fin de incrementar la accesibilidad de las fibras. Los factores
más importantes a tener en cuenta para el pretratamiento con explosión a vapor son el tamaño de partícula, temperatura, tiempo de residencia y el efecto
combinado temperatura-tiempo (Ro) [Ro = t*e(τ-100/14.75)], el cual describe la
condición óptima para la producción máxima de azúcares fermentables: el factor debe estar entre 3 y 4,5. Altas temperaturas incrementan la remoción de la
hemicelulosa del sólido y la digestibilidad de la celulosa. Las ventajas, incluyen
menor impacto ambiental, mayor eficiencia energética, menores costos de operación, recuperación completa de los azúcares, entre otros.
Pretratamiento con agua caliente: Se aplica agua al material a temperaturas de 160-240°C para provocar alteración en la estructura de la lignocelulosa. El
objetivo del líquido es solubilizar la hemicelulosa y hacer más accequible la
celulosa, asimismo evitar la formación de inhibidores. Hay menor formación de inhibidores y mayor recuperación de pentosas. Sin embargo, consumen grandes
cantidades de energía y no se utiliza a escala industrial.
Explosión de fibra con amonio (AFEX): Los materiales son tratados con líquidos amoniacales a temperaturas entre 60 y 100°C en tiempos variables. Solo
produce pretratamiento al material sólido, es decir que se solubiliza poca cantidad de material sólido; disminuye la cristalinidad de la celulosa y rompe los
enlaces carbohidratos-lignina; pequeñas cantidades de hemicelulosa y lignina
son removidas, baja formación de sustancias tóxicas e inhibidoras de los postratamientos enzimáticos. La recuperación del amonio es costoso. Hay un
pretratamiento de explosión con amonio acuoso, denominado percolación de
amonio reciclado (ARP), en el cual se utiliza solución de amonio acuoso (5-15% p/p) y temperatura entre 140 y 210°C, con tiempos de reacción de 90 min y
velocidades de percolación de 5mL/min. Este procedimiento solubiliza
hemicelulosa, pero mantiene intacta la celulosa. Una ventaja de este método es la
disminución en el consumo de energía.
Oxidación húmeda: Se emplea aire u oxígeno como catalizador a temperaturas de 170 a 200°C y presiones de 10 a 12 bar O2. Ese proceso produce reacciones
de formación de ácidos del proceso hidrolítico y reacciones oxidativas. Se
solubiliza la hemicelulosa y la lignina y se incrementa la digestibilidad de la celulosa. Adicionando Na2CO3 se disminuye la formación de compuestos
inhibitorios y se mantiene el pH neutro-alcalino. Asimismo, se logra la
recuperación del 96% de la celulosa (65% convertida a glucosa). La principal desventaja es el costo del oxígeno.
Pretratamiento con microondas: El material es sometido entre 5 y 20 min, se
hace comparable con los tratamientos con álcali. Se realizan a escala de laboratorio.
Pretratamiento ultrasonido: se utiliza en diferentes tiempos, obteniendo los mejores resultados a 50°C. Abre los poros del material por impacto mecánico,
haciendo que las macromoléculas se desordenen y se incremente el rendimiento
de la hidrólisis enzimática. No hay formación de sustancias tóxicas.
Explosión con CO2: El CO2 es utilizado como fluido supercrítico a temperaturas
por encima del punto crítico, removiendo efectivamente la lignina e
incrementando la digestibilidad de la celulosa.
Residuos agrícolas, maderas
blandas, duras,
bagazo de caña, rastrojo de maíz.
Zhu et al.
(2006);
Yachmenev et al. (2009);
Palonen et al. (2004); Kim et
al. (2008);
Wyman et al. (2005); Mosier
et al. (2005b);
Hamelinck et al. (2005)
2.3. Enzimas lignocelulolíticas
De todas las funciones de las proteínas, la catálisis es tal vez la más importante, la cual es realizada por
las enzimas. Las enzimas son producidas en el interior de la célula, de las cuales se conocen enzimas
extracelulares que actúan fuera de la célula y se hará referencia más abajo, y las enzimas intracelulares que
son responsables de todas las reacciones metabólicas que mantienen en equilibrio los diferentes sistemas
biológicos y que actualmente son aisladas y utilizadas con diversos fines industriales. Estas enzimas
intracelulares pueden ser obtenidas por métodos de extracción químicos o físicos de las diferentes partes de
los organismos que las contienen, según se requiera. Asimismo, ya hay desarrollos de técnicas para la
separación y purificación de las fracciones enzimáticas obtenidas de los procesos de extracción, como

procesos combinados de centrifugación, ultrafiltración y cromatografía (Blanch y Clark, 1997; Bommarius y
Riebel, 2004b).
Las enzimas extracelulares actúan fuera de la célula. Su función principal es disminuir el tamaño de las
moléculas complejas que se encuentran en el medio cultivo, las cuales posteriormente son absorbidas como
fuentes de nutrientes y energía por los organismos que las excretan (Bommarius y Riebel, 2004b). El sistema
de secreción enzimática en los hongos filamentosos tiene como propósito principal el crecimiento y
transporte de nutrientes hacia el ápice hifal, siendo ésta el área activa de secreción enzimática. No se descarta
que exista una cierta actividad secretora a lo largo del resto del micelio con el fin de mantener la actividad
metabólica (Papinutti et al., 2003).
Las enzimas para la degradación de polímeros (no necesariamente polisacáridos) pueden emplear dos
estrategias de ataque. Pueden actuar sobre el polímero aleatoriamente fragmentando efectivamente la
molécula polimérica en oligómeros, siendo responsables de esta forma de ataque las endoenzimas.
Alternativamente en el caso de los polisacáridos, pueden actuar a partir de un extremo (exoenzimas),
generalmente el extremo no reductor de la molécula, liberando monómeros o dímeros de modo secuencial.
Los sistemas se completan con enzimas capaces de actuar sobre oligómeros o dímeros, llevando a la
degradación completa del polímero. Estos complejos de enzimas actúan de modo sinérgico (Moore, 1998b).
En general las enzimas degradadoras de polisacáridos están sujetas a mecanismos de regulación de su
síntesis. Es decir no se producen de modo constante, sino que su síntesis es inducida por el sustrato adecuado
y es reprimida por azúcares fácilmente utilizables, en particular glucosa. El inductor más eficiente es el
polímero-sustrato de las enzimas que serán sintetizadas. Sin embargo, debido a su alto peso molecular, no
son compuestos que puedan penetrar en las células y ejercer su efecto (Béguin y Aubert, 1994). Algunas de
las enzimas extracelulares que producen los hongos se encuentran dispersos en la película de líquido que
rodea la hifa, pero otras están fijas en el espacio sujetas a la pared de la hifa, a la matriz extracelular o al
propio sustrato (Hudson, 1986).
Cabe destacar que en medios de cultivo ligeramente diferentes se sintetizan diversas formas de una
misma enzima, que a pesar de poseer características catalíticas similares, difieren notablemente en sus
características fisicoquímicas, por lo que se deduce que hay formas múltiples que coexisten en determinado
momento pero también puede variar la composición de isoenzimas a lo largo del desarrollo del hongo
(Carlile et al., 2001). Las enzimas extracelulares del complejo celulolítico y ligninolítico existen en múltiples
formas. La síntesis de isoenzimas puede deberse a productos de genes diferentes, aunque también pueden
deberse a cambios postraduccionales como proteólisis, glicosilación, o agregados cuaternarios. Se pueden
sintetizar como consecuencia de estados fisiológicos diferentes (composición del medio de cultivo, pH,
temperatura, etc.) a partir de genes regulados diferencialmente. En la naturaleza, el hongo se encuentra con
distintos sustratos, distintas condiciones ambientales, y pueden entonces producirse distintas formas
enzimáticas, con propiedades más adecuadas a las nuevas circunstancias que les permitan degradar al
polímero de la manera más eficiente. Sin embargo, también el producto de un gen puede ser modificado
diferencialmente mediante glicosilaciones o digestiones proteolíticas luego de la secreción (Joselau y Ruel,
1994; Blanchette, 1995; Barr et al., 1996; Carlile et al., 2001).
2.3.1. Enzimas celulolíticas
El complejo de enzimas celulolíticas de los hongos de pudrición blanca consiste de tres enzimas
hidrolíticas: endoglucanasa, exoglucanasa y glucosidasas, las cuales trabajan sinérgicamente (Lemaire, 1996)
y se detallan a continuación.
i. Endoglucanasas
La engolucanasa como 1,4-β-D-glucan 4-glucanohidrolasas (EC 3.2.1.4) produce una disminución en el
grado de polimerización de la celulosa, por cortes al azar de la cadena glucosídica, produciendo glucosa,
celobiosa (disacáridos) y celotriosa (trisacáridos). Los sustratos sobre los que actúan las endoglucanasas son
carboximetilcelulosa, celulosa amorfa, celooligosacáridos; sin atacar en forma significativa a la celulosa

cristalina, como las fibras de algodón.
ii. Exoglucanasas
Causan principalmente rupturas secuenciales en la molécula de celulosa a partir de un extremo no
reductor liberado y están representadas por dos grupos de enzimas: exocelobiohidrolasas o celobiohidrolasas
(CBH) (1,4-β-D-glucan celobiohidrolasas) (EC 3.2.1.91) que liberan unidades de celobiosa a partir del
extremo no reductor y exoglucohidrolasas (1,4-β-D-glucan glucohidrolasas) (EC 3.2.1.74) que liberan
unidades de glucosa a partir del extremo no reductor liberado e incluyen también actividad de las
celobiohidrolasas produciendo celobiosa por ataque en el extremo no reductor del polímero. Las
celobiohidrolasas son las que se encuentran con mayor frecuencia (Ângelo, 2004).
iii. β-glucosidasas
La β-glucosidasa o β-D-glucósido glucohidrolasas (EC 3.2.1.21) hidrolizan celobiosa y otros β-1,4-
oligoglucósidos de cadena corta, para formar glucosa, siendo este monosacárido el resultado final de la
acción degradativa del complejo celulolítico sobre el polímero de celulosa (Moore, 1998b).
La celulosa es atacada inicialmente por la endoglucanasa (ver Figura 7), la cual se liga al azar sobre las
microfibrillas de celulosa en regiones amorfas, produciendo hidrólisis de los enlaces β-1,4 glucosídicos,
generando así múltiples sitios de ataque para las exo-1,4-β-D-glucanasas, las cuales actúan sobre los
extremos no reductores de la cadena, liberando una unidad de celobiosa. Existe una cooperación continua
entre la acción de ambas enzimas y la de la β-glucosidasa, la cual actúa sobre la celobiosa o también sobre
pequeños oligómeros produciendo moléculas de glucosa. El resultado de la acción de estas tres actividades
enzimáticas es la degradación sinérgica de la celulosa como se muestra en la Figura 7.
Existen actividades complementarias entre los tres tipos de enzimas y se consideran los responsables del
sinergismo, dado que la acción de dos o más enzimas combinadas es mayor que la suma de las actividades
individuales en la degradación (Papinutti et al., 2003). Puede ocurrir sinergismo tanto entre, como dentro de
las diferentes clases de enzimas celulolíticas (Barr et al., 1996; Boisset et al., 2000). Aunque el sinergismo
entre los distintos componentes del complejo celulasa aún no ha sido completamente elucidado, está claro
que depende de diferentes factores como la naturaleza del sustrato, la afinidad del componente celulasa por
el sustrato, la estereo-especificidad de dicho componente, la concentración de la enzima y la proporción entre
los componentes enzimáticos (Barr et al., 1996; Mansfield et al., 1999).
En adición a los tres grupos principales de enzimas, se han encontrado en distintas especies fúngicas,
especialmente en hongos de pudrición blanca, un número de actividades enzimáticas oxidativas auxiliares,
las cuales no son esenciales para la hidrólisis de la celulosa pero pueden asistir a la descomposición del
polímero llevando a cabo conversiones muy útiles, por ejemplo oxidaciones o la remoción de productos
finales que pueden provocar represión catabólica o inhibición de la actividad de los grupos principales de
enzimas celulolíticas (Coughlan y Ljungdahl, 1988; Markham, 1998).
Cuando un hongo de pudrición blanca se desarrolla sobre la celulosa se producen dos celobiosas-oxido-
reductasas: celobiosa-quinona oxido-reductasa (CBQ) y celobiosa-oxidasa (CBO). Esta serie convierte la
celobiosa en δ-lactona que puede a su vez convertirse en ácido celobiónico y entonces obtenerse glucosa y
ácido glucónico. Las enzimas celobiosa-oxido-reductasas son capaces de utilizar una gran variedad de
aceptores de electrones y son de gran significancia en la regulación de los niveles de celobiosa y glucosa,
cuya acumulación puede inhibir la acción de la endoglucanasa. La CBQ degrada las uniones entre celulosa y
lignina (Moore, 1998b). La celobiosa-oxidasa reduce el Fe+3
conjuntamente con el peróxido de hidrógeno,
generando radicales hidroxilo. Estos radicales pueden degradar tanto la celulosa como la lignina,
posiblemente indicando que la celobiosa oxidasa tiene un rol central en degradación de maderas por hongos
degradadores de madera (Ander, 1994). Actualmente aún se desconocen muchas de las formas de ataque
bioquímico que utilizan los hongos filamentosos para la degradación de los materiales celulósicos
heterogéneos, como los residuos lignocelulósicos.
La síntesis de celulasas está sujeta a inducción por el sustrato adecuado (celulosa cristalina) y represión

por una fuente hidrocarbonada fácilmente utilizable, como la glucosa. El inductor más efectivo es el sustrato
natural: la celulosa cristalina. Sin embargo este material, es un polímero de gran tamaño, que no puede
penetrar en la célula e inducir la síntesis de celulasas a nivel transcripcional. No obstante, existen niveles
basales de celulasas constitutivas que a partir de un bajo nivel de hidrólisis de la celulosa, producirían algún
producto soluble capaz de penetrar en la célula induciendo la síntesis enzimática (Carle - Urioste et al., 1997;
Forchiassin y Levin, en imprenta).
Los hongos de pudrición castaña utilizan un sistema celulolítico bien diferenciado del sistema
hidrolítico de los hongos de pudrición blanca. Éstos pueden despolimerizar celulosa rápidamente a través de
un mecanismo que involucra a la endoglucanasa y a factores no proteicos, entre ellos H2O2 y Fe+2
(Coughlan
y Ljungdahl, 1988; Wood y García, 1994), ya que producen endoglucanasas y β-glucosidasas, pero
aparentemente no producen exoglucanasas (Gow, 1995b).
Figura 7. Representación del modo de acción de las celulasas:
Glucosa extremo reductor ; Glucosa extremo no reductor
(a) Estructura cristalina y amorfa de la celulosa, (b) Acción del complejo celulolítico sobre la estructura de la
celulosa, endoglucanasas sobre la región interna amorfa, exoglucanasas sobre los extremos no reductores de la región
cristalina y β-glucosidasa sobre los dímeros formados.
Las celulasas fúngicas son producidas por diversos hongos de importancia económica, incluyendo
especies de Trichoderma spp., Aspergillus spp. y Phanerochaete chrysosporium y un gran número de
especies más (Lockington et al., 2002). La secreción de celulasas por los hongos, así como las demás
enzimas que producen estos organismos, está regulada por factores relacionados con la composición química
del sustrato y el metabolismo celular. Los hongos pueden disminuir la producción de celulasas por represión
catabólica de carbono (mediada por ruta metabólica vía CreA ) (Ilmen et al., 1996). Los hongos
filamentosos, en general, son potentes productores de celulasas. Sin embargo, no es posible asegurar un
Región cristalina Región amorfa
Endoglucanasa β-glucosidasa Exoglucanasa
(a)
(b)

mayor productor de estas enzimas bajo una sola premisa, ya que la experiencia ha demostrado que se deben
priorizar los factores asociados a la maximización de la producción de este grupo de enzimas, y aún en el
caso en que se requiera la selección de una de las celulasas en particular (Dekker, 1985; Coughlan y
Ljungdahl, 1988; Jennings, 1995; Ilmen et al., 1996; Lia et al., 2000; Lockington et al., 2002; Sohail et al.,
2009).
Los sistemas de celulasas están compuestos de múltiples proteínas que poseen funciones
complementarias que interactúan con la hidrólisis de la celulosa en la formación de complejos
multiproteínicos. Sin embargo, estas interacciones exhiben diferentes comportamientos cuando los
componentes del sistema de celulasas se presentan combinados, o cuando ellas están presentes de forma
aislada. Los sustratos celulósicos se presentan en diversas estructuras macroscópicas insolubles con una
distribución de partícula, una forma y en algunos casos composición variada, lo que genera una
heterogeneidad en el modo de ataque de los sistemas celulolíticos al material celulósico. Asimismo, influyen
también, la procedencia del sistema celulolítico, la dotación y la presencia de activadores de éstos en los
sustratos. Por ejemplo, Trichoderma reesei ha sido investigado desde hace 50 años. T. reesei, el cual produce
como mínimo dos exoglucanasas (CBHI y CBHII), cinco endoglucanasas (EGI, EGII, EGIII, EGIV y EGV)
y dos β-glucosidasas (βGI y βGII), con muchos esfuerzos realizados por años, se han modificado
genéticamente las cepas de este hongo y se ha logrado obtener hasta 0.33 g de proteína/g de carbohidrato
utilizado y un reconocimiento de su modo de acción sobre diversos materiales (Lynd et al., 2002).
En la Tabla 7 se enumeran actividades enzimáticas celulolíticas de varios hongos de pudrición blanca,
como Phanerochaete chrysosporium, Pleurotus ostreatus, Lentinus edodes, en contraste con diferentes
especies de Trichoderma spp. sobre diferentes tipos de sustratos compuestos en su mayoría, por materiales
lignocelulósicos heterogéneos como los residuos agrícolas. Los sustratos se presentan tanto en estado sólido,
como en medio líquido. Uno de los principales aspectos relevantes que se observan en la Tabla 7 es la
variación directa en la expresión de las celulasas, cualquiera que sea la que haya sido determinada, con el
sustrato sobre el cual se desarrolla el organismo. Es visible la influencia del estado de la fermentación en las
actividades de las enzimas celulolíticas; Elisashvili et al. (2001a) por ejemplo, muestra que las actividades de
endoglucanasa como CMasa determinadas en fermentaciones sumergidas corresponden a por lo menos tres
veces más que las actividades de la CMasa sobre medio sólido para Lentinus edodes IBB363 y L. edodes
IBB1233. En la determinación de la actividad de la enzima endoglucanasa (EG), se observa que ésta fue
determinada por varios investigadores sobre carboximetilcelulosa (CMC) y por otros como CMCasa. La
actividad FPasa es equivalente a la determinación de la exoglucanasa, debido a que se mide la actividad de la
enzima (o el organismo) sobre papel de filtro, el cual se compone de aproximadamente 99% de celulosa
cristalina; sin embargo, es considerada comúnmente como la actividad celulolítica total o global de un
organismo productor de estas enzimas (Dobozi et al., 1992; Rodríguez y Piñeros, 2007). Todo lo
anteriormente expuesto acerca del comportamiento de los organismos que producen celulasas hace pensar
que aún falta mayor exploración sobre la producción de enzimas celulolíticas tanto en medios sólidos, como
líquidos utilizando como sustratos diferentes mezclas de residuos lignocelulósicos y rastrear un mayor
número de especies fúngicas que pueden expresar enzimas celulolíticas si las condiciones medioambientales
y las características de los sustratos promueven su producción.
Tabla 7. Comparación de las actividades enzimáticas celulolíticas de hongos sobre diversos sustratos
Hongo
Material sobre el
que se cultivó el
hongo
Actividad enzimática
Tipo de enzima Sustrato empleado Actividad Referencia
Trichoderma
reesei
- CBHI y CBHII
purificadas
Avicel+AR, 50°C,
3h*
0,014-0,027 U/mg Tomme et al.
(1988)
- CBHI y CBHII
purificadas
Avicel+AR, 40°C,
3h*
0,26-0,48 U/mg El Gogary et al.
(1989)
- Proteína cruda Avicel, 37°C, 21 h* 0,000241 U/mg Johnson et al.
(1982)
Trichoderma
viride T12
Residuos de
palma (RP) sin
pretratamiento
EXG
EG
Papel filtro
CMC
0,004±0,008 U/mL
0,086±0,022 U/mL Rodríguez y
Piñeros (2007)

RP tratamiento
ácido 0,5%
EXG
EG
Papel filtro
CMC
0,012±0,004 U/mL
0,000±0,000 U/mL
RP tratamiento
ácido 1%
EXG
EG
Papel filtro
CMC
0,052±0,012 U/mL
0,048±0,022 U/mL
RP tratamiento
biológico (P.
ostreatus) 10 días
EXG
EG
Papel filtro
CMC
0,146±0,088 U/mL
0,614±0,078 U/mL
RP tratamiento
biológico (P.
ostreatus) 20 días
EXG
EG
Papel filtro
CMC
0,150±0,038 U/mL
0,558±0,091 U/mL
Trichoderma
reesei
Plantas
aromáticas
medicinales:
citronela, artemisa
y desechos de
jardín
EXG
EG
βG
Papel filtro
CMC
p-nitrofenil-β-D-
glucopiranósido
13,01 U/g
94,13 U/g
16,85 U/g
Chandra et al.
(2009)
Trichoderma
viride
Plantas
aromáticas
medicinales:
citronela, artemisa
y desechos de
jardín
EXG
EG
βG
Papel filtro
CMC
p-nitrofenil-β-D-
glucopiranósido
-
-
45 U/g
Trichoderma
citrinoviride
Plantas
aromáticas
medicinales:
citronela, artemisa
y desechos de
jardín
EXG
EG
βG
Papel filtro
CMC
p-nitrofenil-β-D-
glucopiranósido
10,0 U/g
97,60 U/g
94,77 IU/g
Trichoderma
harzianum
Plantas
aromáticas
medicinales:
citronela, artemisa
y desechos de
jardín
EXG
EG
βG
Papel filtro
CMC
p-nitrofenil-β-D-
glucopiranósido
10,0 U/g
28,66 U/g
90,0 U/g
Trichoderma
fasciculatum
Plantas
aromáticas
medicinales:
citronela, artemisa
y desechos de
jardín
EXG
EG
βG
Papel filtro
CMC
p-nitrofenil-β-D-
glucopiranósido
5,0 U/g
72,80 U/g
16,34 U/g
Trichoderma
koningii
Plantas
aromáticas
medicinales:
citronela, artemisa
y desechos de
jardín
EXG
EG
βG
Papel filtro
CMC
p-nitrofenil-β-D-
glucopiranósido
3,0 U/g
60,0 U/g
63,09 U/g
Trichoderma
viride
Residuo de
algodón molido.
EG CMC 30 U/mL
Mandels y
Reese (1956)
Rastrojo de
algodón. EG CMC
21 U/mL
Residuo de
algodón
parcialmente
degradado.
EG CMC
8 U/mL
Lodos de madera EG CMC 29 U/mL
Celulosa
Chromista
(Saprolegnia)
EG CMC
4 U/mL
Celulosa bacterial
(Acetobacter) EG CMC
5 U/mL
Celulosa animal EG CMC 7 U/mL
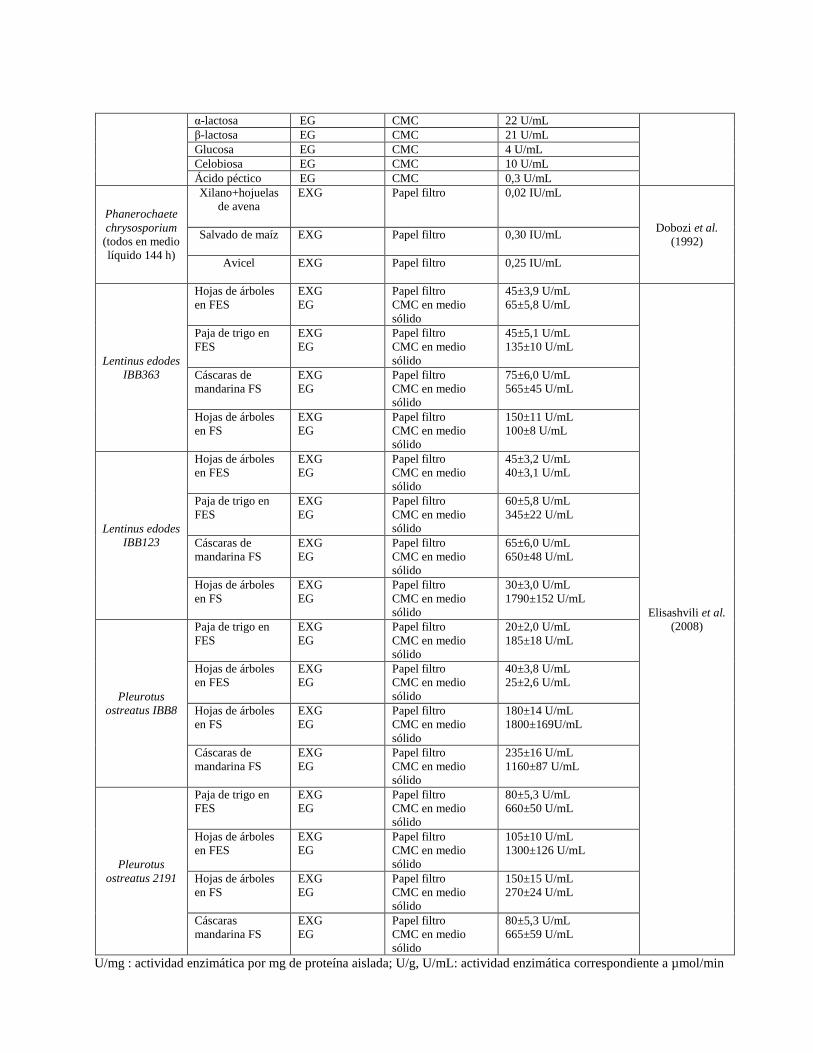
α-lactosa EG CMC 22 U/mL
β-lactosa EG CMC 21 U/mL
Glucosa EG CMC 4 U/mL
Celobiosa EG CMC 10 U/mL
Ácido péctico EG CMC 0,3 U/mL
Phanerochaete
chrysosporium
(todos en medio
líquido 144 h)
Xilano+hojuelas
de avena
EXG Papel filtro 0,02 IU/mL
Dobozi et al.
(1992) Salvado de maíz
EXG Papel filtro 0,30 IU/mL
Avicel
EXG Papel filtro 0,25 IU/mL
Lentinus edodes
IBB363
Hojas de árboles
en FES
EXG
EG
Papel filtro
CMC en medio
sólido
45±3,9 U/mL
65±5,8 U/mL
Elisashvili et al.
(2008)
Paja de trigo en
FES
EXG
EG
Papel filtro
CMC en medio
sólido
45±5,1 U/mL
135±10 U/mL
Cáscaras de
mandarina FS
EXG
EG
Papel filtro
CMC en medio
sólido
75±6,0 U/mL
565±45 U/mL
Hojas de árboles
en FS
EXG
EG
Papel filtro
CMC en medio
sólido
150±11 U/mL
100±8 U/mL
Lentinus edodes
IBB123
Hojas de árboles
en FES
EXG
EG
Papel filtro
CMC en medio
sólido
45±3,2 U/mL
40±3,1 U/mL
Paja de trigo en
FES
EXG
EG
Papel filtro
CMC en medio
sólido
60±5,8 U/mL
345±22 U/mL
Cáscaras de
mandarina FS
EXG
EG
Papel filtro
CMC en medio
sólido
65±6,0 U/mL
650±48 U/mL
Hojas de árboles
en FS
EXG
EG
Papel filtro
CMC en medio
sólido
30±3,0 U/mL
1790±152 U/mL
Pleurotus
ostreatus IBB8
Paja de trigo en
FES
EXG
EG
Papel filtro
CMC en medio
sólido
20±2,0 U/mL
185±18 U/mL
Hojas de árboles
en FES
EXG
EG
Papel filtro
CMC en medio
sólido
40±3,8 U/mL
25±2,6 U/mL
Hojas de árboles
en FS
EXG
EG
Papel filtro
CMC en medio
sólido
180±14 U/mL
1800±169U/mL
Cáscaras de
mandarina FS
EXG
EG
Papel filtro
CMC en medio
sólido
235±16 U/mL
1160±87 U/mL
Pleurotus
ostreatus 2191
Paja de trigo en
FES
EXG
EG
Papel filtro
CMC en medio
sólido
80±5,3 U/mL
660±50 U/mL
Hojas de árboles
en FES
EXG
EG
Papel filtro
CMC en medio
sólido
105±10 U/mL
1300±126 U/mL
Hojas de árboles
en FS
EXG
EG
Papel filtro
CMC en medio
sólido
150±15 U/mL
270±24 U/mL
Cáscaras
mandarina FS
EXG
EG
Papel filtro
CMC en medio
sólido
80±5,3 U/mL
665±59 U/mL
U/mg : actividad enzimática por mg de proteína aislada; U/g, U/mL: actividad enzimática correspondiente a µmol/min

formados del producto esperado ( azúcares reductores) por g o por mL de sustrato seco o volumen de extracto crudo del
cual proviene; AR: azúcares reductores, *tiempo de reacción enzimática, el número en paréntesis indica el día de
máxima actividad, FES: fermentación en estado sólido, FS: fermentación sumergida, EG: endoglucanasa, CMC:
carboximetilcelulosa, EXG: exoglucanasa, Cel: celulosa cristalina, βG: β-glucosidasa, MD: extracto de malta, SM:
salvado de maíz, BC: borra de café.
En la Tabla 8 se muestran las actividades enzimáticas máximas de los hongos Grifola frondosa y
Trametes trogii en procesos de fermentación en estado sólido para G. frondosa y en estado líquido para T.
trogii. Hay que resaltar que se muestran los tiempos de fermentación en los que se logran las máximas
actividades enzimáticas, hecho importante, ya que esto refleja que cada una de las enzimas manifiesta su
actividad a diferentes tiempos de fermentación. Asimismo, como se ve afectada esta actividad por la
composición de los medios de cultivo.
Tabla 8. Máximas actividades enzimáticas celulolíticas de los hongos T. trogii y G. frondosa
sobre sustratos sintéticos para dos procesos de fermentación en estado sólido.
Hongo
Material sobre el
que se cultivó el
hongo
Actividad enzimática
Tipo de
enzima Sustrato empleado Actividad Referencia
Trametes trogii
Madera
EG CMC(29) 167,25 U/g
Levin et al.
(2008)
βG p-nitrofenil-β-D-
glucopiranósido (43)
0,35 U/g
Madera+MD
EG CMC(36) 253,49 U/g
βG p-nitrofenil-β-D-
glucopiranósido (43)
0,66 U/g
Madera+MD+pep
tona
EG CMC(7) 503,81 U/g
βG p-nitrofenil-β-D-
glucopiranósido (14)
0,89 U/g
Madera+MD+pep
tona+Cu
EG CMC(17) 512,21 U/g
βG p-nitrofenil-β-D-
glucopiranósido (14)
0,87 U/g
Grifola
frondosa
PSUMCC 922
Aserrín+SM en
FES
EG
EXG
βG
CMC(45)
Cel(30)
p-nitrofenil-β-D-
glucopiranósido(60)
11,45±0,81U/g
15,63±0,55 U/g
2,20±0,013 U/g
Montoya (2008) Aserrín+SM+BC
en FES
EG
EXG
βG
CMC(30)
Cel(30)
p-nitrofenil-β-D-
glucopiranósido(75)
8,84±0,804 U/g
10,8±0,61 U/g
1,90±0,075 U/g
U/g, U/mL: actividad enzimática correspondiente a µmol/min formados del producto esperado ( azúcares reductores)
por g o por mL de sustrato seco o volumen de extracto crudo del cual proviene; el número en paréntesis indica el día de
máxima actividad, FES: fermentación en estado sólido, EG: endoglucanasa, CMC: carboximetilcelulosa, EXG:
exoglucanasas, Cel: celulosa cristalina, βG: β-glucosidasa, MD: extracto de malta, SM: salvado de maíz, BC: borra de
café.
2.3.2. Enzimas xilanolíticas
Las enzimas que degradan hemicelulosa se nombran según el sustrato sobre el que actúan; por ejemplo,
mananasas degradan mananos, xilanasas degradan xilanos, etc. (Moore, 1998b). Dentro de las hemicelulosas,
el más abundante es el xilano, por ello las xilanasas constituyen el grupo enzimático más estudiado entre las
hemicelulasas. Las D-xilanasas, similares a las celulasas, se componen básicamente de dos grupos de
enzimas: endo-1,4--D-xilanasas (EC 3.2.1.8), y 1,4--D-xilosidasas (EC 3.2.1.37), las cuales en muchos
casos están presentes en forma de distintas isoenzimas (Goodenough, 1996; Lia et al., 2000).
La hidrólisis de estas moléculas complejas requiere de la interacción de numerosas enzimas que corten
la cadena principal y también las laterales. La endo-1,4--D-xilanasa actúa sobre la cadena de xilano al azar,
disminuyendo el grado de polimerización con liberación de xilooligosacáridos, xilobiosa y xilosa. La 1,4--
D-xilosidasa interviene luego de la endoxilanasa, actuando sobre los xilooligosacáridos (a partir de los

extremos no reductores) o sobre la xilobiosa, dando como producto xilosa (Bajpai, 1997). Sin embargo, un
sistema xilanolítico completo requiere de las enzimas que actúan sobre sus ramificaciones tales como las α-
arabinofuranosidasas que remueven cadenas laterales de L-arabinosa, las α-glucuronosidasas que liberan
ácidos glucurónicos de las cadenas laterales y las acetilxilano-esterasas que remueven los grupos
sustituyentes acetílicos de la xilosa (Forchiassin y Levin, en imprenta), como se ilustra en la Figura 8.
Las estructuras de varios xilanos y xilanasas han sido estudiadas por diversos autores (Beily et al., 1981;
Visser et al., 1991; Tolan et al., 1996; Lia et al., 2000) utilizando técnicas electrónicas como difracción de
rayos X, espectroscopía de barrido electrónico y otras. El modo de acción de varias endoxilanasas sobre
sustratos conocidos ha sido estudiado (Dekker y Richards, 1975; Comtat y Joseleau, 1981; Meagher et al.,
1988). Sin embargo, aún falta por estudiar y comparar los modos de acción de varias xilanasas sobre
sustratos heterogéneos, como mezclas de residuos lignocelulósicos. Las investigaciones que se realicen en
torno a este tema adquieren importancia por las diversas aplicaciones industriales que poseen estas enzimas.
2.3.3. Enzimas ligninolíticas
En la Tabla 9 se presenta una descripción sobre el complejo ligninolítico, y en la Figura 9 se muestra un
esquema de acción del mismo sistema producido por hongos de pudrición blanca. El sistema ligninolítico
funciona en sinergismo con los demás complejos enzimáticos. En especial en la Figura 9 se ilustra como la
hifa de los hongos de pudrición blanca a través de su complejo de celulasas degradan la celulosa hasta
glucosa. La glucosa en el medio, activa la formación de la enzima glucosa oxidasa (GOX), que en presencia
de oxígeno produce gluconolactona (producto intermedio) y peróxido de hidrógeno. Posteriormente por
reacciones químicas diferentes a las enzimáticas la gluconolactona se convierte en ácido glucónico.
Asimismo, la hifa produce otro metabolito llamado glioxal sobre el cual actúa la glioxal oxidasa (Gliox),
activada por el mismo metabolito a través de la cual también se obtiene peróxido de hidrógeno. Estas
enzimas productoras de peróxido de hidrógeno son necesarias para el funcionamiento de las peroxidasas, ya
que sin ellas no habría presencia de H2O2 en el medio. Este mecanismo corresponde a parte de la
interpretación que probablemente puede dársele al funcionamiento de los hongos de pudrición blanca para
lograr descomponer sustancias tan complejas como la lignina, ya que la presencia de peróxido de hidrógeno
en el medio, el manganeso y el alcohol veratrílico (metabolito secundario producido por la hifa) son
necesarios para la activación y acción de las enzimas oxido-redutasas lacasa, manganeso peroxidasa (MnP) y
lignin peroxidasa (LiP), degradadoras de los compuestos fenólicos y no fenólicos en todas las direcciones,
para su conversión en radicales y productos de bajo peso molecular. La acción de las enzimas oxido-
reductasas resulta en una despolimerización parcial y las roturas ocurren sin estereoespecificidad. Estos
mecanismos oxidativos que producen radicales libres, pueden reaccionar entre sí volviendo a polimerizarse.
Sin embargo, existe un equilibrio entre polimerización-despolimerización y pueden iniciarse reacciones no
específicas que después pueden seguir cualquier camino sin subsiguiente control por parte de las enzimas, lo
que se ha llamado “combustión enzimática” (Kirk y Farrel, 1987).

Figura 8. Estructura de xilano y enzimas involucradas en su degradación. Xilosa , ácido acético
arabinofuranosa , ácido glucurónico o 4-O-metilglucurónico , 1-acetilxilanoestearasa, 2-arabinofuranosidasa, 3-
endoxilanasa, 4-β-xilosidasa, 5-α-glucuronosidasa. Fuente: Forchiasin y Levin, (en imprenta)
La LiP, MnP y peroxidasa versatil (PV) son peroxidasas con un grupo hemo como cofactor, que
emplean peróxido de hidrógeno como primer aceptor de electrones. La LiP puede oxidar directamente
sustratos aromáticos fenólicos y no fenólicos siendo el más estudiado el alcohol veratrílico (3,4 dimetoxi-
fenol). La MnP oxida Mn+2
a Mn+3
y este último es quelado por ácidos orgánicos sintetizados por el hongo
(Pérez y Jeffries, 1992). Es decir que la MnP puede oxidar sustratos del tipo fenólico a través del Mn+2
(Tuor et al., 1992), pero también puede hacerlo con sustratos no fenólicos a través de los productos de
peroxidación de ácidos grasos insaturados (Bao et al., 1994). La PV oxida Mn+2
a Mn+3
, como lo realiza la
MnP y también oxida compuestos no fenólicos, como la LiP lo que significa que comparte propiedades
catalíticas con ambas peroxidasas fúngicas (Martinez et al., 1996; Martinez, 2002). La lacasa es una oxidasa
con cuatro átomos de cobre como cofactor y cuyo aceptor de electrones es el oxígeno dando agua como
producto (Thurston, 1994). Las lacasas oxidan sustratos fenólicos y no fenólicos formando radicales
catiónicos, quinonas o radicales fenoxi. Esta enzima en presencia de cooxidantes como el 1-
hidroxibenzotriazol (HBT), el 2,2’-azinobis (ácido 3-etilbenzotiazoline-6-sulfónico) (ABTS) o el ácido
violúrico; capacita a la enzima para atacar sustratos más difícilmente oxidables como los de tipo no fenólico
(Soares et al., 2001). La lacasa es la enzima ligninolítica presente más frecuentemente en los hongos de
pudrición blanca.
3 3
1 5 2
2 5 3
1 2
4
4

Figura 9. Esquema de acción del sistema ligninolítico. Cel: celulosa; Lig.: lignina; [Glox]: glioxal oxidasa; [GOX]:
glucosal oxidasa; [MnP]: manganeso peroxidasa; [LiP]: lignin peroxidasa; [Lc]: lacasa VA: alcohol veratrílico.
Generalmente las materias primas que componen los sustratos que se utilizan para el desarrollo y
crecimiento de hongos de pudrición blanca son materiales heterogéneos, los cuales poseen además de las
fuentes de carbono una cantidad de minerales asociados que pueden servir como inductores dependiendo de
las concentraciones en las que se encuentren presentes, de otro modo puede complementarse adicionando
sales minerales que mejoren las reacciones de las oxido-reductasas producidas por los hongos de pudrición
blanca.
Los títulos de las enzimas óxido-reductasas, como lacasa, MnP, glioxal-oxidasa, se ven afectadas por la
concentración de metales como el cobre en el medio de cultivo; así no sea un hecho común que la MnP se
induzca con otro metal que no sea Mn+2
ya se ha mostrado por Mouso et al.(2003) y Niku-Paavola et al.
(1990) que en Stereum hirsutum el Cu+2
y el Cd+2
inducen la síntesis de esta enzima y por Levin et al. (2002)
en Trametes trogii la producción de lacasa y MnP son incrementadas con el agregado de CuSO4 al medio. La
concentración de los metales que son utilizados como inductores debe ser regulada de modo que no superen
la concentración de inhibición del crecimiento del hongo. Por ejemplo, Levin et al. (2002) realizaron
pruebas de crecimiento del hongo Trametes trogii variando la concentración de sulfato de cobre en el medio
como inductor de lacasa y MnP, obteniendo como resultado los mayores títulos de lacasa, MnP y glioxal-
oxidasa con 1mM de sulfato de cobre en el medio, mientras que con 1,5 mM de la misma sal se presentó
inhibición del crecimiento. Este hecho resulta ser muy importante para utilizar un metal como el cobre para
inducir la formación de enzimas ligninolíticas, aunque aún no se conozca la razón por la cual la mayor parte
de las ligninasas se ven favorablemente afectadas en su producción cuando al medio de cultivo se le adiciona
una sal de cobre. Asimismo, estos trabajos pueden convertirse en puntos de partida interesantes en
aplicaciones de estas enzimas, como en procesos de biorremediación, dado que en un mismo medio se
produce la máxima cantidad de enzima aprovechándose la acción sinérgica de todo el sistema enzimático de
los hongos de pudrición blanca en los diferentes procesos de degradación.
La degradación de sustancias polifenólicas se puede llevar a cabo mediante la reacción Fenton, la cual
se define como una reacción de oxido-reducción en presencia de iones metálicos, especialmente el Fe+2
o
Fe+3
y peróxido de hidrógeno (Gold y Alic, 1993; Rodríguez et al., 2003; Hammel et al., 2002). Varios
trabajos dan indicios de que los hongos de pudrición blanca además de la producción de H2O2, sintetizan
[Glox]
Glioxal [GOX]
O2
H2O2 H2O2
[Celulasas] Glc
O2
Cel.
Lig.
Gluconolactona ácido glucónico
Cu+2
[Lc] [MnP]
Mn+2
[LiP] Radicales: productos
de despolimerización
de la lignina
VA
Hifa

ciertos metabolitos aromáticos como productos intermedios, inclusive que tienen la capacidad de convertir
los compuestos no fenólicos de la lignina en compuestos fenólicos cuando en el medio ocurre una reacción
de oxidación de Fe+3
a Fe+2
en presencia de H2O2, reacción que generalmente es llevada a cabo por medio de
la acción de la LiP. La macromolécula de lignina está compuesta de 10 a 20% de estructura fenólica y entre
el 80-90% de estructura no fenólica, lo que hace que estos hongos deban contener mínimo un mecanismo
con alto potencial redox por el cual las unidades de lignina son degradadas o convertidas a unidades
fenólicas a fin de fomentar la oxidación por acción de la MnP y lacasa (Arantes y Milagres, 2007b).
Asimismo, a los hongos de pudrición blanca no se les conoce un único mecanismo por el cual degraden la
lignina, sino que esta biodegradación es atribuida probablemente a la contribución que hace cada enzima en
dependencia de las condiciones ambientales y la composición y estructura del sustrato. Sin embargo, la
evaluación completa del sistema oxidativo de los hongos de pudrición blanca no se ha totalizado aún. Ha
sido reportada la secreción de muchas sustancias de bajo peso molecular por los hongos de pudrición blanca,
como ácidos orgánicos, ácidos grasos y compuestos de Fe+3
con capacidad reductora y quelante, tales como
catecol-quelante y glucopéptidos. Los compuestos reducidos de Fe+3
juegan un rol importante en la
degradación de la madera por reducción a Fe+2
que en presencia de H2O2 generan radicales •OH, el cual es
uno de los más potentes oxidantes conocidos, de amplio espectro sobre compuestos orgánicos naturales
diversos, estos radicales •OH reaccionan de diferentes formas con la lignina y algunas de estas reacciones
convierten sustancias no fenólicas en fenólicas, como por ejemplo cuando ocurre la adición de un radical OH
en una posición insustituible de un anillo y el radical favorece la oxidación y libera un protón para ser
introducido a un nuevo grupo fenólico hidroxílico. En este punto MnP o lacasa pueden fomentar una nueva
forma fenólica de lignina; lo que genera en el proceso global de degradación de lignina una explicación
mínima de la degradación completa de esta estructura en ausencia de LiP (Buswell y Odier, 1987; Kirk y
Farrel, 1987; Blanchette, 1995; Blanchette et al., 1997; Arantes y Milagres, 2007b).
En diferentes estudios previos se reporta la degradación de la lignina fenólica y no fenólica utilizando el
reactivo de Fenton (Fe+3
/H2O2) sintético reductor. Un sistema referido a una reacción mediadora de Fenton
como quelante. Según estos resultados, probablemente en mayor o menor magnitud la reacción de Fenton
puede ser involucrada en el proceso global de la biodegradación de la lignina por hongos de pudrición
blanca, también reportada para hongos de pudrición café, ya que ellos poseen una forma de generar la
solubilización del Fe+2
, el cual en presencia de H2O2 una especie de oxígeno reactivo (ROS) es formado vía
reacción de Fenton (Gold y Alic, 1993; Blanchette et al., 1997; Rodríguez et al., 2003; Hammel et al., 2002).
Existen evidencias de la degradación del compuesto no fenólico azure B (indicador de LiP) han sido
llevados a cabo utilizando el posible sinergismo dado entre el reactivo de Fenton (Fe+3
/H2O2) con hongos de
pudrición blanca que producen solamente MnP y lacasa, evidenciando que la degradación del azure B está
sujeta a la modificación de su estructura, promovida por Fe+3
/H2O2 a fin de que el sustrato sea oxidado por el
sistema lacasa/MnP de los hongos de pudrición blanca (Arantes y Milagres, 2007a; Arantes y Milagres,
2007b).
Estas investigaciones se constituyen en un excelente avance para el desarrollo de procesos, como el de
biorremediación, ya que hay evidencias de sustratos previamente tratados con Fenton (Fe+3
/H2O2) y
posteriormente inoculados con el hongo de pudrición blanca no productor de LiP o adicionando un extracto
rico en MnP/lacasa fueron ampliamente degradados, demostrándose la importancia de la LiP para la
degradación de compuestos recalcitrantes, asimismo la asociación positiva que puede generarse entre la
reacción Fenton y los hongos de pudrición blanca, teniendo en cuenta que Fenton no sólamente corresponde
a reacciones en presencia de hierro férrico o ferroso, sino que también puede generarse en presencia de otros
metales como el cobre (Arantes y Milagres, 2007b; Arantes y Milagres, 2007a; Henry, 2003).
Tabla 9. Principales enzimas ligninolíticas de hongos de pudrición blanca
Enzima
ligninolítica Sustratos Propiedades Ocurrencia natural Referencias

Lignin-
peroxidasa
(E.C.1.11.1.14)
(LiP)
Grupos fenólicos y
no fenólicos Azure
B, alcohol
veratrílico, Poly R-
478 y oxida el
ABTS.
Hemoproteína glicosilada,
extracelular, dependiente de H2O2.
Es capaz de oxidar alcohol veratrílico,
metabolito producido durante el
crecimiento secundario junto con la
enzima. Es importante porque por su
pequeño tamaño puede difundir y
actuar a cierta distancia de la enzima.
Cumple un rol protector de la enzima
ya que previene su oxidación.
Hongos de pudrición blanca:
Phanerochaete crysosporium,
Grifola frondosa, Oxyporus
latemarginatus CTM 10133 y
CTM 10136, Trametes trogii
CTM 10154 y CTM 10156,
Polyporus sp., Coriolus
antarcticus, Pycnoporus
sanguineus, Trametes extenuata,
Pleurotus lindquistii entre otros.
Glenn y Gold
(1985); Koduri y
Tien, (1994);
Leonowicz et al.,
(1999); Levin et
al. (2004); Dhouib
et al. (2005);
Kersten y Cullen,
(2007).
Manganeso
Peroxidasa
(E.C.1.11.1.13)
(MnP)
Actúa sobre sustratos
fenólicos, rojo fenol
y sales de Mn.
Hemoproteina glicosilada con fuerte
preferencia por el Mn+2 como
sustrato. Lo oxida a Mn+3, el cual
forma complejos con ácidos
dicarboxílicos o con α-hidroxiácidos,
estos complejos pueden difundir a
cierta distancia de la enzima y oxidar
a su vez a la lignina. La regulación de
su producción está dada por la
limitación de nitrógeno y por la
presencia de Mn.
Phlebia sp., Oxyporus
latemarginatus, CTM 10133 y
10136 Trametes trogii,
Polyporus sp. Grifola frondosa,
Ganoderma applanatum,
Ganoderma lucidum, Lentinus
edodes, Pleurotus sp., Fomes
sclerodermeus, Daedaleopsis
confragosa, Marasmius allaceus,
entre otros.
Glenn y Gold
(1985);
Leonowicz et al.
(1999); Levin et
al. (2004);
Buswell et al.
(1995);
Maganhotto de
Souza-Silva et al.
(2005);
Rodriguez-Couto
y Sanromán,
(2005); Kersten y
Cullen, (2007);
Levin et al.
(2008).
Lacasa
(E.C.1.10.3.2)
Modelos de lignina
que contiene grupos
fenólicos libres
formando radicales
fenoxi y mediante la
acción de
mediadores del
proceso actúa sobre
grupos no fenólicos.,
ABTS.
Es una polifenoloxidasa que contiene
cobre en su molécula. Son
extraordinariamente no-específicas
respecto al sustrato. Presentan una
amplia gama de aplicaciones:
blanqueo de pulpas, decoloración de
textiles, tratamiento de aguas
residuales, biorremediación, etc.
Hongos de pudrición blanca,
algunas bacterias, insectos y
plantas superiores: Phlebia sp.,
Oxyporus latemarginatus, CTM
10133 y 10136 Trametes trogii,
Polyporus sp. Grifola frondosa,
Ganoderma applanatum,
Ganoderma lucidum, Lentinus
edodes, Pleurotus sp., Fomes
sclerodermeus, Daedaleopsis
confragosa, Marasmius allaceus,
entre otros.
Levin et al.
(2004); Dhouib et
al. (2005); Levin
et al. (2008)
Thurston, (1994);
Eggert et al.
(1996); Mayer y
Staples, (2002);
Rodriguez-Couto
y Toca-Herrera,
(2007).
Celobiosa-
quinona-oxido-
reductasa
(CBQ)
Celobiosa, quinonas
y metabolitos
producidos por la
acción de enzimas
ligninolíticas.
Oxida celobiosa a celobionolactona y
reduce quinonas y otros radicales
producidos por LiP, MnP, lacasa y
protege así de la repolimerización.
Por ser enzimas que intervienen
siempre en las reacciones de
transición en las reacciones de
las enzimas MnP, LiP, lacasa y
celulasas. Aún no se determinan
especies particulares de
producción de éstas. En general
todos los hongos que producen
enzimas ligninolíticas, también
producen CBQ
Evans et al.
(1994); Zhao y
Janse, (1996);
Carlile et al.
(2001).
Glucosa-oxidasa
(GOX: glucosa
1-oxidasa)
El requerimiento
necesario es la
presencia de glucosa
y oxígeno. Polímeros
de celulosas,
almidones, etc.
Su síntesis acompaña la síntesis de
peroxidasas. Actúa específicamente
sobre la glucosa mediante la
incorporación de O2, para producir
gluconolactona y H2O2,
posteriormente, a partir de la
glucolactona se obtiene ácido
glucónico.
En general, por todos los hongos
que utilizan la glucosa como
fuente de energía.
Evans et al.
(1994); Zhao y
Janse, (1996);
Vroemen (2002)

Glioxal-oxidasa
(Glox)
Oxida varios
sustratos: glioxal,
metilglioxal.
Es extracelular. El glioxal es un
metabolito excretado por los hongos
de pudrición blanca. Los azúcares
necesarios para la glioxal-oxidasa
pueden ser producidos por la acción
de celulasas o hemicelulasas sobre los
polisacáridos de la madera.
Por ser enzimas que intervienen
siempre en las reacciones de
transición en las reacciones de
las enzimas MnP, LiP, lacasa y
celulasas. En general todos los
hongos de pudrición blanca que
producen enzimas ligninolíticas,
también producen glioxal-
oxidasa
Evans et al.
(1994); Zhao y
Janse, (1996);
Carlile et al.
(2001).
2.3.4. Otras enzimas producidas por hongos de pudrición blanca
i. Proteasas
Hidrolizan los enlaces peptídicos de las proteínas y los péptidos; rompen las moléculas de sustrato en
fragmentos más pequeños (Moore, 1998a). Las enzimas proteolíticas son producidas intra y extra
celularmente, juegan un rol importante en los procesos de regulación de la célula y la pared celular,
contribuyendo con la nutrición de los hongos a través de la degradación de las proteínas presentes en los
nutrientes. Las proteasas extracelulares son producidas por muchas especies de hongos, aunque las más
conocidas son las producidas por Aspergillus sp. y Neurospora crassa. Los basidiomicetos Agaricus,
Coprinus y Volvariella son organismos que se pueden destacar porque además de utilizar la glucosa como
fuente de energía, pueden también usar proteínas como fuente de nitrógeno y azufre (Kalisz et al., 1987;
Kalisz, 1988; Kalisz et al., 1989).
ii. Pectinasas
Las enzimas pectinolíticas constituyen un grupo heterogéneo de enzimas que catalizan la degradación
de las pectinas. Las plantas y los microorganismos son los principales productores de estas enzimas. Las
pectinasas se clasifican en tres tipos principales: pectinas esterificantes o pectinesterasas, enzimas
despolimerizantes, las hidrolasas y liasas y protopectinasas. Las pectinestearasas catalizan la
desesterificación de los grupos metilo de la pectina dando lugar a ácido poligalacturónico o pectato (Burns,
1991; Alkorta et al., 1998). Su actividad se puede determinar midiendo la liberación de metanol. Las
despolimerasas se clasifican atendiendo a tres aspectos, que se refieren al modo de ataque al esqueleto de
ácido poligalacturónico. Estos aspectos siguen los siguientes criterios: a) el mecanismo químico de ruptura
de los enlaces: hidrólisis o transeliminación; b) la naturaleza del sustrato preferido: pectina o ácido
poligalacturónico; c) el modo de corte del polímero: endo o exo (Chang et al., 1994). Las enzimas
despolimerizantes pueden romper los enlaces α(1-4) glucosídicos entre los monómeros de ácido
galacturónico de las pectinas por hidrólisis (hidrolasas) o por transeliminación (liasas). Las hidrolasas
rompen los enlaces entre los monómeros del ácido galacturónico por adición de agua. Son activas a pH ácido
y pueden ser inhibidas por el ión calcio (Alkorta et al., 1998). Generalmente las exopoligalacturonasas de
hongos y plantas liberan ácido galacturónico, mientras que las bacterianas liberan ácido digalacturónico. Las
polimetilgalacturonasas degradan el polímero metilado, la pectina. Todas las pectinasas descritas hasta el
momento, actúan de forma endo (Baker y Wicker, 1996; Alkorta et al., 1998). Son sintetizadas
mayoritariamente por hongos, aunque también se han identificado en algunas bacterias (Chang et al., 1994).
Las pectinasas se usan ampliamente en la industria alimentaria en la clarificación de jugos y vinos, en la
extracción de aceites, en la remoción de cáscaras de frutas cítricas y para incrementar la firmeza de varias
frutas (Rodríguez-Couto y Sanromán, 2005).
iii. Lipasas y esterasas
Catalizan la hidrólisis de ésteres formando alcoholes y ácidos orgánicos, generalmente presentan baja
especificidad y podría decirse que casi cualquier lipasa puede hidrolizar cualquier éster orgánico variando

solamente la velocidad de reacción. Los principales factores que influyen en la especificidad son las
longitudes y formas de las cadenas hidrocarbonadas que se encuentran a lado y lado del grupo éster. El
término esterasa es generalmente aplicado a enzimas que prefieren cadenas de carbono cortas en el grupo
acilo. Las lipasas apropiadas tienden a favorecer las cadenas largas de carbono en el grupo acilo, como las
grasas. Los componentes lipídicos de las lipoproteínas y el enlace éster son fosfolípidos. La producción de
lipasas extracelulares han sido detectadas en Agaricus bisporus durante la degradación bacteriana (Fermor y
Wood, 1981); en cultivos de fermentación han sido producidas en fase estacionaria como metabolitos
secundarios por Rhizopus sp., teniéndose que controlar las concentraciones de carbono, nitrógeno y oxígeno
para obtener buenos rendimientos de lipasas (Mukhamedzhanova y Bezborodov, 1982; Guiseppin, 1984).
Las fosfatasas son también estearasas que actúan sobre ésteres de alcoholes con ácido fosfórico. Éstas
son enzimas de baja especificidad y se dividen en grupos dependiendo de su actividad, como
fosfomonoestearasas, fosfodiestearasas y polifosfatasas. Se distinguen por su acción bajo condiciones
alcalinas o ácidas. Por ejemplo las fosfatasas producidas por Agaricus bisporus durante su crecimiento sobre
compost, se convierten en un factor de nutrición importante para el hongo; ya que éstas pueden hacer una
recuperación importante de sulfatos o de polisacáridos sulfatados, tales como los que se encuentran en el
suelo (Moore, 1998a).
2.3.5. Aplicaciones de las enzimas lignocelulolíticas
En el contexto general, es notable el incremento de la integración de la catálisis biológica en una gran
variedad de procesos industriales de artículos básicos de consumo y en la industria química, tales como acril-
amida, fructosa, ácido málico y aspártico, como intermediarios de complejos farmacéuticos intermedios,
insumos para la producción de drogas, entre otros (Schulze y Wubbolts, 1999; Zaks, 2001). Las hidrolasas se
consideran un grupo importante de enzimas para ser utilizadas en la producción de químicos finos como
enantiómeros de moléculas quirales, productos químicos con diferentes grados de pureza, ya que por la
selectividad de este grupo de enzimas, se pueden obtener mayores combinaciones de estos productos a
menores costos de producción (Stead et al., 1996; Ladner y Ditrich, 1999).
Las enzimas oxido-reductasas son utilizadas para la conversión de compuestos proquirales en moléculas
quirales con alta selectividad. En la industria se encuentran aplicaciones de reacciones de bio-oxidación, que
hacen uso de la regioselectividad de la biocatálisis. Por ejemplo, se han utilizado procesos de fermentación
para la producción de ácido 6-hidroxinicotínico, para la fabricación de agroquímicos, a partir de ácido
nicotínico a gran escala con Achromobacter xilososidans y para la conversión de ácido (R)-2-
fenoxipropanoico a (R)-2-4-hidroxifenoxi-ácido propanoico (utilizado para síntesis de muchos
agroquímicos) con Bauveria bassiana (utilzada por la BASF) (Meyer et al., 1997; Schulze y Wubbolts,
1999).
Las enzimas ligninolíticas de hongos de pudrición blanca tienen una amplia aplicación en procesos de
transformación y mineralización de órgano-contaminantes con estructuras similares a las de la lignina. Estas
enzimas en la actualidad están siendo involucradas en los procesos de depuración de aguas residuales,
pesticidas bifenil policlorados, hidrocarburos policíclicos aromáticos, efluentes de plantas de blanqueo,
residuos sintéticos, polímeros sintéticos, entre otros (Pointing, 2001). Las enzimas ligninolíticas por su baja
especificidad son activas sobre una amplia gama de sustratos; su modo de acción se basa en la generación de
radicales libres. Estas enzimas son capaces de realizar transformación o mineralización in vitro de productos
altamente recalcitrantes como fenantrenos, benzopirenos, pentaclorofenol, 2,2’,4,4’-tetraclorofenil y muchos
otros compuestos aromáticos policíclicos (Reddy, 1995; Pointing, 2001; Ali, 2010). Esto es de particular
importancia para el tratamiento de residuos con xenobióticos que están generando un problema de polución
ambiental. Referente al tema de biorremediación, en particular, se requiere mayor investigación sobre el
modo de acción de las enzimas ligninolíticas sobre diversos compuestos aromáticos. Falta información
disponible acerca de la determinación de condiciones ambientales para realizar las transformaciones y
mineralizaciones de estas sustancias denominadas recalcitrantes in situ. Asimismo, es importante realizar las
comparaciones de las eficiencias de procesos de fermentación con procesos enzimáticos, utilizando los
extractos crudos de enzimas o las enzimas purificadas.

Las ligninasas son utilizadas para la deslignificación de los materiales lignocelulósicos dispuestos para
alimentación de rumiantes. Por ejemplo, la industria de producción de carne bovina en el mundo presenta
dificultades para mantener estable el suministro de alimentos para los animales. Uno de los principales
inconvenientes es la variación del clima, lo que ha generado que este segmento de la industria alimentaria
busque nuevas alternativas para el mantenimiento de la alimentación animal (Koutinas et al., 2004). En el
planeta se desechan grandes cantidades de materiales lignocelulósicos que generan contaminación y que
pueden ser utilizados para este fin utilizando complejos de enzimas ligninolíticas para la degradación de la
lignina y posteriormente realizar los ensilajes o las transformaciones y suplementaciones requeridas por los
animales (Weinberg, 2000; Koutinas et al., 2004).
Las enzimas celulolíticas son consideradas de gran importancia industrial. Por ejemplo, ellas han sido
ampliamente utilizadas para reemplazar la hidrólisis ácida como método de conversión de celulosa en
azúcares fermentables, especialmente glucosa para la producción de etanol y otros alcoholes de importancia
industrial, asimismo, la glucosa obtenida es utilizada como materia prima para la manufactura de diferentes
alimentos para animales, humanos y productos químicos. Las celulasas también son utilizadas en diversos
procesos de fermentación para la producción de sustancias de uso industrial como el ácido láctico, en
procesos de fermentación aceto-butílica; en el mejoramiento de la textura del papel (liso), biodesentintado
para reciclado del papel utilizando una mezcla de celulasas, pectinasas, xilanasas y lipasas; en el biopulido
de telas para el tratamiento superficial de las pelusas, en la elaboración de paños super absorbentes, en la
extracción de jugos, aromas, aceites y pigmentos con agregados enzimáticos de celulasas, pectinasas y
xilanasas, éstas rompen paredes celulares y hace más eficiente la extracción; en la elaboración de aditivos no
calóricos; ampliamente utilizada en la formulación de detergentes para mejoramiento del liso de las telas y
de la evacuación de la formación de fibras indeseables en telas de algodón (Joshi y Pandey, 1999a; Carlile et
al., 2001; Bommarius y Riebel, 2004b; Sánchez y Cardona, 2007).
Las enzimas lignocelulolíticas juegan un rol importante en la conversión del complejo lignocelulósico
en azúcares fermentables para la producción de alcohol carburante (Sánchez y Cardona, 2008). Los
materiales lignocelulósicos generalmente son sometidos a pretratamientos físicos y químicos como
adecuación de los materiales a fin de mejorar las condiciones de procesos y de trabajo en los procesos de
fermentación para la producción de etanol. A pesar de que existen alternativas de desarrollo para realizar los
pretratamientos de estos materiales lignocelulósicos utilizando enzimas comerciales. Falta más investigación
para el desarrollo de estas tecnologías de transformación de los materiales lignocelulósicos, en azúcares
fermentables con extractos crudos de enzimas lignocelulolíticas para el proceso de producción de etanol a
escala industrial.
2.4. Polisacáridos fúngicos
Los polisacáridos de los hongos macromicetos han sido reconocidos como excelentes
inmunoestimuladores debido al potencial de sus propiedades terapéuticas, a que no son tóxicos y son escasos
sus efectos secundarios en comparación con otros inmunoestimulantes. Estos polisacáridos bioactivos son
producidos en cantidades relevantes a partir de los cuerpos fructíferos de los hongos, su micelio y además,
de los filtrados obtenidos de sus cultivos en fermentación sumergida. En mayor medida las estructuras de los
polisacáridos procedentes de los macromicetos corresponden a homoglicanos, heteroglicanos o complejos de
gluco-proteínas. En general, la actividad inmunomoduladora de los polisacáridos de los macromicetos está
relacionada con las estructuras β-(1→3), β-(1→6) glucanos, α-(1→3) glucanos y complejos de polisacáridos
de galactomananos y glucuromananos unidos a proteínas presentes en la pared celular de los macromicetos
(Enshasy, 2010). El β-D-glucano es un polisacárido que produce exclusivamente D-glucosa luego de la
hidrólisis ácida (Mizuno, 1999). A los biopolímeros fúngicos quitina y quitosano no se les ha reportado
actividad antitumoral.
Los polisacáridos pueden ser obtenidos por medio de fermentación sumergida con una selección
apropiada del medio de cultivo, procedimiento que en ocasiones puede ser más útil que esperar que las
diferentes especies de macromicetos fructifiquen para utilizar estos carpóforos como materia prima de los
polisacáridos (Komura et al., 2010).

Los β-glucanos forman moléculas largas y cilíndricas que pueden contener hasta 250.000 unidades de
glucosa. Los β-glucanos también se encuentran en las paredes de las células del endosperma de granos como
la cebada y la avena, y ayudan a reducir las enfermedades del corazón bajando el nivel de colesterol y
reduciendo la reacción glicémica de los carbohidratos. Se usan comercialmente para sustituir grasas y para
modificar la textura de los productos alimenticios (Mohammad-Fata et al., 2007). En efecto, de los 700
hongos comestibles, solo unos 50 tienen valor medicinal y entre ellos Grifola frondosa es uno de los más
notables e investigado, especialmente como estimulador del sistema inmune y por sus propiedades en la
lucha contra el cáncer. Sin embargo, según Chang y Miles (2004) se ha considerado que existen unas 1800
especies de macrohongos con potencial medicinal en el planeta. Se han aislado muchas sustancias bioactivas
con efecto inmunoestimulador a partir de los macromicetos entre los que se incluyen polisacáridos de alto y
bajo peso molecular, glicoproteínas, triterpenoides y proteínas fúngicas que estimulan el sistema
inmunológico. Estas sustancias bioactivas pueden ser obtenidas de diferentes estructuras del macromiceto
como de extractos del cuerpo fructífero en diferentes solventes, desde agua caliente hasta distintas mezclas
de solventes orgánicos; también se pueden extraer del micelio obtenido de los cultivos líquidos y de los
caldos de cultivo de las fermentaciones sumergidas. En mayor proporción estas sustancias se obtienen del
cuerpo fructífero sometido a proceso de extracción con un promedio del 77, 3%, seguida del micelio
conseguido del procesos en medio líquido con un 20,8% y finalmente un 2% derivadas de los caldos de
cultivo de las fermentaciones sumergidas. Sin embargo, cada especie de hongo produce distintos tipos de
polisacáridos con diferentes propiedades y actividad biológica, lo cual, a pesar de sus aplicaciones, no se ha
discutido sistemáticamente en la literatura más que a nivel de laboratorio. De igual forma, la estructura de la
cual se obtiene el polisacáridos bioactivo, del carpóforo, del micelio o de los caldos de cultivo afectan las
estructuras moleculares, la unidad estructural que se repite (el monómero), los pesos moleculares, entre otros
factores, lo cual influye directamente en su modo de acción y su actividad biológica (Tsukagoshi et al., 1984;
Ishikawa et al., 1992; Hamlyn y Schmidt, 1994; Kobayashi et al., 1995; Zhuang y Mizuno, 1999; Chang y
Miles, 2004; Mohammad-Fata et al., 2007).
Los exopolisacáridos de basidiomicetos de pudrición blanca juegan un rol importante en los procesos de
crecimiento y desarrollo de estos organismos. Estos exopolisacáridos pueden inmovilizar las exoenzimas que
ellos mismo excretan (Tang y Zhong, 2002). Según Catley (1992) el gel formado por estos biopolímeros
previene la deshidratación de la hifa, permitiendo la adherencia de otras células a las superficies de los
sustratos, posibilitando la selección de las moléculas del ambiente que los rodea (Maziero et al., 1999).
Adicionalmente, estos biopolímeros tienen una amplia expectativa de aplicaciones industriales. Un ejemplo
es el exopolisacárido conocido como schizophyllan producido por el basidiomiceto Schizophyllum commune.
Este polímero es el β-(1-3), (1-6)-glucano soluble en agua, que forma una solución viscosa con alta
estabilidad térmica y ha sido utilizado con buenos resultados en pacientes con cáncer cervical (ver Tabla 10)
(Jong y Birgmingham, 1992; Hamlyn y Schmidt, 1994). En la Tabla 10 se hace referencia a algunos
polisacáridos obtenidos de hongos de pudrición blanca con su actividad antitumoral e inmunomoduladora y
sus estructuras generales. Asimismo, a estos compuestos se les ha encontrado otras funciones biológicas, en
las que se incluye actividad antiviral, propiedades inmunoreguladoras (Kumari y Sahoo, 2006; Tao et al.,
2006), en la industria cosmética (Mansel, 1994; Zanchetta et al., 2003). Además, de haberse reportado usos
de estos biopolímeros en la industria alimentaria (Moon et al., 2006), y en la remoción de iones metálicos de
aguas residuales (Chu y Kim, 2006). Otros investigadores han reportado actividad antitumoral de los
complejos de polisacáridos extraídos del micelio o del cuerpo fructífero de hongos macromicetos, como el
complejo proteína-polisacárido PSK del hongo Coriolus versicolor (Ooi y Liu, 2000), el complejo
polisacárido-proteína del cultivo de Tricholoma lobayense (Liu et al., 1996), exobiopolímeros de varias
especies de Cordyceps a las cuales se les ha demostrado actividad antitumoral (Chen et al., 1997),
inmunopotenciador (Nakamura et al., 1999), actividad hipoglicémica (Kiho et al., 1999) y efecto
hipocolesterolémico (Koh et al., 2003).
Los polisacáridos de los macromicetos y sus complejos polisacárido-proteina estimulan el sistema
inmune no específico y por lo tanto ejercen actividades antitumorales a través de la estimulación de los
propios mecanismos de defensa del cuerpo (Wasser y Weis, 1999). Los polisacáridos de las setas pueden
actuar como efectores y antiproliferativos en la inducción de la apoptosis (muerte celular programada) en las

células antitumorales, activando los efectores de las células como macrófagos y los linfocitos para secretar
citoquinas, las cuales son capaces de promover la citoxicidad de los macrófagos; de modo que la producción
de las citocinas (citoquinas) por las células inmunes es considerada como un acontecimiento de la iniciación
y regulación de la respuesta inmune del cuerpo que consume estos polisacáridos bioactivos (Lull et al.,
2005).
Los polisacáridos bioactivos de los hongos han demostrado reducir el tamaño de los tumores hasta en un
50% en ratones prolongando la supervivencia del portador del tumor (Wasser, 2002). A través de los años se
ha comprobado que los extractos de los hongos macromicetos tienen actividad inmunomoduladora, aunque
el enfoque estándar haya sido aislar y caracterizar los compuestos bioactivos puros a fin de facilitar el
suministro vía intravenosa a las personas que lo requieran. De forma que surgen interrogantes de cómo los
polisacáridos de los macromicetos pueden introducirse en la célula, si éstos por su tamaño no pueden
penetrar la pared celular. Sin embargo, estos polisacáridos pueden actuar de forma sinérgica con otros
componentes, ya que las respuestas a diferentes polisacáridos son mediadas por el reconocimiento de
diferentes receptores que se encuentran en la superficie celular y su adecuada combinación ofrece una mejor
respuesta, dando mayores efectos terapéuticos al polisacárido o su combinación (Borchers et al., 2004; Lull
et al., 2005; Leung et al., 2006).
Los pesos moleculares de los polisacáridos o los compuestos asociados de los hongos juegan un rol
importante al momento de medir su actividad biológica o bioactividad, ya que todas las fracciones de estas
estructuras no exhiben una actividad biológica detectable. Por ejemplo, en extractos obtenidos de G.
frondosa se obtuvieron fracciones de diferentes tamaños, detectando la mayor actividad inmunomoduladora
en la fracción de masa molecular de 800 kDa (Adachi et al., 1990). De igual forma, cuando se realizaron las
separaciones de las fracciones del PSK (Krestin) de C. versicolor por ultrafiltraciones sucesivas, en cuatro
fracciones: F1, <50 kDa; F2, 50-100 kDa; F3, 100-200 kDa; F4, >200 kDa la fracción que mayor actividad
inmunomoduladora arrojó fue la de mayor masa molecular (Kim et al., 1990). Estos ejemplos son una
muestra de la potencialidad que tienen los hongos como fuente de sustancias bioactivas para la obtención de
potenciadores del sistema inmunológico, teniendo en cuenta que de las 140.000 especies de macromicetos
que se estima existen en el planeta, solamente se conocen cerca del 10%, de las cuales sólo han sido
examinadas desde su potencial inmunomodulador el 5% (Lindequist et al., 2005).
En la formación de exo e intra-polisacáridos están involucradas varias enzimas que conducen a su
formación y pueden estar divididas en cuatro grupos: enzimas responsables del metabolismo inicial de los
carbohidratos; enzimas que involucran síntesis de nucleótidos y azúcares y su interconversión;
glicosiltransferasas que frecuentemente sintetizan unidades glucosídicos de lípidos y; translocasas y
polimerasas que sintetizan los polímeros. La fosfoglucosaisomerasa (PGI) conduce a la formación de lactato
y la α-fosfoglucomutamasa (α-PGM) conduce a la formación de los polisacáridos, que son los puntos
ramificados entre la vía Embden–Meyerhof–Parnas (EMP) y la biosíntesis de polisacáridos. La
suplementación de los sustratos es una estrategia para incrementar la producción de biomasa celular y una
forma de mantener constante la producción de polisacáridos (Tang y Zhong, 2002).
Tabla 10. Polisacáridos bioactivos obtenidos de hongos de pudrición blanca con actividad antitumoral e
inmunomoduladora.
Especie
Tumor/cáncer
tratado/Actividad
inmunomoduladora
Origen del
polisacárido Estructura del polisacárido Referencia
Coriolus versicolor Sistema digestivo /
inmunomodulador Cultivo micelial
Glucoproteínas, (1-4), (1-3)-β-
D-glucan, Polisacaropéctido
Ishikawa et al.
(1992); Tsukagoshi et
al. (1984); Cui y
Chisti (2003)
Lentinus edodes
Cáncer de
estómago/actividad
inmunomoduladora
Cuerpo fructífero,
medio de cultivo
(1-3)-β-D-glucan,
manoglucanos, glucanos,
lentinan, complejo poliscáridos-
proteina
Chihara et al. (1969);
Kosaka y Yamashita
(1993); Hobbs (2000)
Cordyceps sinensis Inmunomodulador Cuerpo fructífero,
micelio, caldo de
Glucano, heteroglucanos,
cordiglucanos
Russell y Paterson
(2008); Yalin et al.
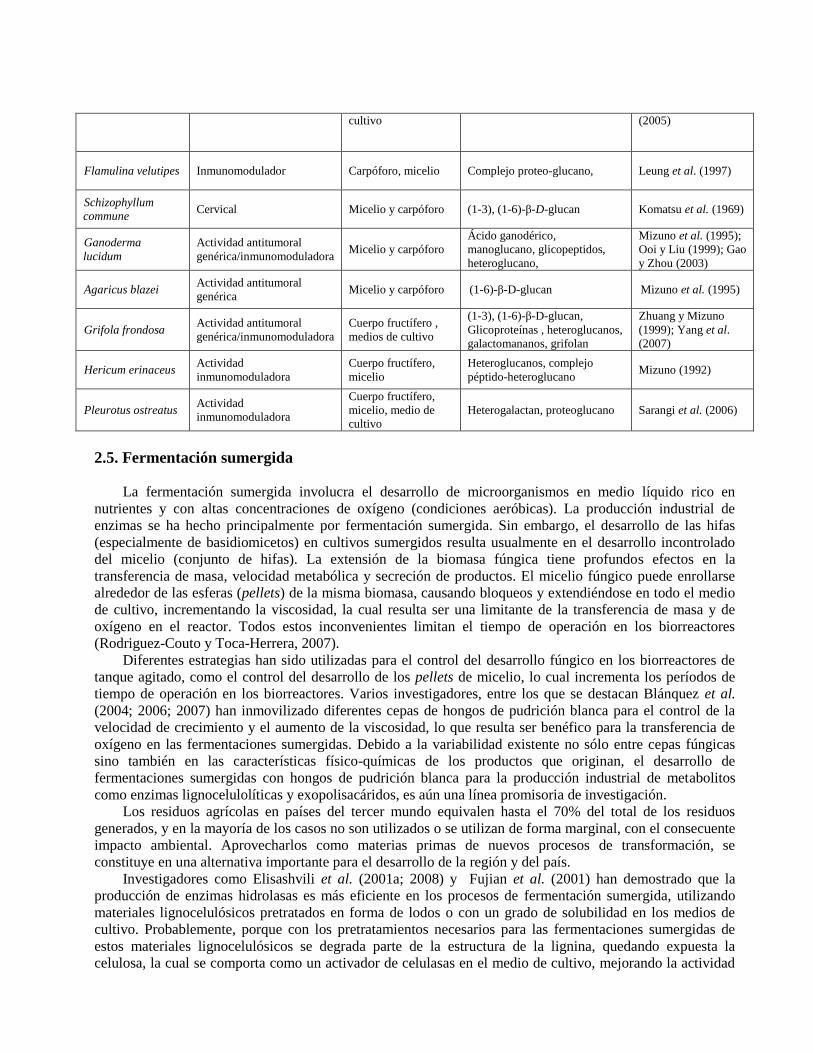
cultivo (2005)
Flamulina velutipes Inmunomodulador Carpóforo, micelio Complejo proteo-glucano, Leung et al. (1997)
Schizophyllum
commune Cervical Micelio y carpóforo (1-3), (1-6)-β-D-glucan Komatsu et al. (1969)
Ganoderma
lucidum
Actividad antitumoral
genérica/inmunomoduladora Micelio y carpóforo
Ácido ganodérico,
manoglucano, glicopeptidos,
heteroglucano,
Mizuno et al. (1995);
Ooi y Liu (1999); Gao
y Zhou (2003)
Agaricus blazei Actividad antitumoral
genérica Micelio y carpóforo (1-6)-β-D-glucan Mizuno et al. (1995)
Grifola frondosa Actividad antitumoral
genérica/inmunomoduladora
Cuerpo fructífero ,
medios de cultivo
(1-3), (1-6)-β-D-glucan,
Glicoproteínas , heteroglucanos,
galactomananos, grifolan
Zhuang y Mizuno
(1999); Yang et al.
(2007)
Hericum erinaceus Actividad
inmunomoduladora
Cuerpo fructífero,
micelio
Heteroglucanos, complejo
péptido-heteroglucano Mizuno (1992)
Pleurotus ostreatus Actividad
inmunomoduladora
Cuerpo fructífero,
micelio, medio de
cultivo
Heterogalactan, proteoglucano Sarangi et al. (2006)
2.5. Fermentación sumergida
La fermentación sumergida involucra el desarrollo de microorganismos en medio líquido rico en
nutrientes y con altas concentraciones de oxígeno (condiciones aeróbicas). La producción industrial de
enzimas se ha hecho principalmente por fermentación sumergida. Sin embargo, el desarrollo de las hifas
(especialmente de basidiomicetos) en cultivos sumergidos resulta usualmente en el desarrollo incontrolado
del micelio (conjunto de hifas). La extensión de la biomasa fúngica tiene profundos efectos en la
transferencia de masa, velocidad metabólica y secreción de productos. El micelio fúngico puede enrollarse
alrededor de las esferas (pellets) de la misma biomasa, causando bloqueos y extendiéndose en todo el medio
de cultivo, incrementando la viscosidad, la cual resulta ser una limitante de la transferencia de masa y de
oxígeno en el reactor. Todos estos inconvenientes limitan el tiempo de operación en los biorreactores
(Rodriguez-Couto y Toca-Herrera, 2007).
Diferentes estrategias han sido utilizadas para el control del desarrollo fúngico en los biorreactores de
tanque agitado, como el control del desarrollo de los pellets de micelio, lo cual incrementa los períodos de
tiempo de operación en los biorreactores. Varios investigadores, entre los que se destacan Blánquez et al.
(2004; 2006; 2007) han inmovilizado diferentes cepas de hongos de pudrición blanca para el control de la
velocidad de crecimiento y el aumento de la viscosidad, lo que resulta ser benéfico para la transferencia de
oxígeno en las fermentaciones sumergidas. Debido a la variabilidad existente no sólo entre cepas fúngicas
sino también en las características físico-químicas de los productos que originan, el desarrollo de
fermentaciones sumergidas con hongos de pudrición blanca para la producción industrial de metabolitos
como enzimas lignocelulolíticas y exopolisacáridos, es aún una línea promisoria de investigación.
Los residuos agrícolas en países del tercer mundo equivalen hasta el 70% del total de los residuos
generados, y en la mayoría de los casos no son utilizados o se utilizan de forma marginal, con el consecuente
impacto ambiental. Aprovecharlos como materias primas de nuevos procesos de transformación, se
constituye en una alternativa importante para el desarrollo de la región y del país.
Investigadores como Elisashvili et al. (2001a; 2008) y Fujian et al. (2001) han demostrado que la
producción de enzimas hidrolasas es más eficiente en los procesos de fermentación sumergida, utilizando
materiales lignocelulósicos pretratados en forma de lodos o con un grado de solubilidad en los medios de
cultivo. Probablemente, porque con los pretratamientos necesarios para las fermentaciones sumergidas de
estos materiales lignocelulósicos se degrada parte de la estructura de la lignina, quedando expuesta la
celulosa, la cual se comporta como un activador de celulasas en el medio de cultivo, mejorando la actividad

enzimática de endoglucanasas, exocelobiohidrolasas y β-glucosidasas en medios líquidos, en comparación
con las actividades hidrolíticas en sustratos sólidos para basidiomicetos. Cuando se trabaja en procesos de
fermentación en estado sólido generalmente no se realizan pretratamientos a los materiales lignocelulósicos y
los organismos que trabajan sobre ellos, probablemente excretan solo las hidrolasas que se requieren para la
obtención de la energía necesaria para su desarrollo.
La producción de polisacáridos extracelulares con hongos macromicetos por fermentación sumergida
resulta influida por las condiciones de fermentación, tales como el tiempo de proceso, temperatura,
composición del medio de cultivo, velocidad de agitación, pH inicial, tamaño del inóculo, etc. Realizar este
seguimiento y control de todas estas variables demanda tiempo, esfuerzo y dinero para obtener los datos
experimentales requeridos a fin de obtener la evaluación exhaustiva de la interacción de estas variables con
la producción directa de los polisacáridos. Actualmente se encuentran reportes de investigaciones realizadas
en la obtención de intra y exopolisacáridos a partir de macromicetos como Coprinus comatus, Agaricus
nevoi, Auricularia auricula-judae, y Flammulina velutipes extraídos con etanol y mostrando una actividad
antioxidante de más del 80% medida con 1,1 difenil-2-picril hidrazil (DPPH) y una producción de
exopolisacáridos de 0,6 a 4,2 g/L en cultivos sumergidos (Asatiani et al., 2008).
En referencia al medio de cultivo para la producción de polisacáridos a partir de hongos, diferentes
trabajos publicados en la última década evidencian la importancia de la presencia de aceites vegetales ricos
en ácido oleico como promotores de crecimiento de la biomasa fúngica y por ende de sus polisacáridos
constituyentes (Yang et al., 2000; Hsieh et al., 2008). Al momento de seleccionar el tipo de aceite vegetal o
ácido grasos a utilizar como inductor de polisacáridos es necesario considerar la composición de ácidos
grasos del aceite; ya que ha sido reportado por varios investigadores que cantidades importantes de ácido
linoleico suprimen la producción de biomasa y polisacáridos, mientras que la presencia de ácido oleico
promueve su producción (Park et al., 2002; Hsieh et al., 2006; Hsieh et al., 2008). Los aceites de soya y de
oliva han sido utilizados como promotores de crecimiento celular y de polisacáridos en cultivos de
macromicetos, tanto líquidos como sólidos. Al respecto, el aceite de oliva contiene 84% de ácido oleico y
entre 30 y 60% en aceite de soya con bajos contenidos de ácido linoleico (Wade, 1993; Hsieh et al., 2008).
Aún se requiere mayor investigación en torno al control de los procesos de fermentación sumergida con
hongos macromicetos. Por ejemplo, Wu et al. (2006) encontraron que para incrementar y mantener constante
la producción de biomasa celular y exopolisacáridos producidos por el macromiceto Auricularia auricula en
fermentación sumergida fue necesario controlar variables tales como composición de sustrato, pH del medio,
temperatura y condiciones ambientales, entre otras. Sin embargo, estudios previos a éste han demostrado que
el control del pH cumple un rol muy importante en el desarrollo de la biomasa miceliana y la producción de
polisacáridos (Fang y Zhong, 2002). Los estudios cinéticos de crecimiento fúngico y producción de
exopolisacáridos por macromicetos en fermentaciones sumergidas son de carácter prioritario para la
optimización de estos procesos.
Las materias primas que se requieren como fuente del complejo lignocelulósico para obtener extractos
crudos de enzimas lignocelulolíticas, celulasas y polisacáridos (exo y endopolisacáridos) utilizando hongos
de pudrición blanca, deben ser tomados de los residuos de la región donde se realicen los procesos. En este
caso en particular, se deben tener en cuenta los residuos de las industrias cafetera (pulpa, borra, cascarilla,
zoca), maderera (aserrín de roble y de pino y los residuos forestales de cosecha), de transformación de cacao
(cascarilla que contiene un alto contenido de proteína y puede ser utilizada como suplemento nutricional de
los sustratos). De la misma forma, la producción agrícola de frutas y hortalizas son el suministro de cáscaras
(plátano, cítricos, aguacate, etc.), los residuos provenientes de la industria de transformación frutícola, como
cáscaras y semillas de mango, mora, lulo, naranja, maracuyá, coco, entre otros. El Departamento de Caldas y
sus alrededores también, produce caña de azúcar y la transforma, obteniéndose subproductos como el
bagazo, la melaza y la cachaza, con valor industrial importante. Por ejemplo, el bagazo de caña posee unas
cantidades importantes de celulosa y hemicelulosa, representan el 75% en base seca de todo el material
(Sánchez y Cardona, 2008). De igual forma, se registra una creciente producción de maíz y arroz en Caldas,
de los cuales pueden utilizarse la tusa, las cáscaras, cascarillas, el tamo (hojas) y la planta después de la
cosecha.
Los hongos, como los demás organismos, utilizados en fermentación sumergida obedecen a una

selección cuando van a ser utilizados en procesos industriales, teniendo en cuenta aspectos básicos de
selección como son la estabilidad genética, la velocidad de crecimiento, la facilidad de adaptación a los
medios de cultivo, entre otros. La selección de los organismos en la mayoría de los casos, se realiza
probando medios de cultivo preferenciales, en la sección 2.7 se amplía la información sobre los métodos de
selección y rastreo de cepas fúngicas según los productos que se esperan obtener.
Las desventajas que limitan los tiempos de operación de los biorreactores de trabajo en fase líquida con
organismos filamentosos como hongos y actinomicetos, hacen que se prefieran los procesos de fermentación
discontinua; sin embargo, estos procesos han sufrido limitaciones tales como altos costos de proceso, control
de tiempos de frecuencias de inicio y terminado de la fermentación, el requisito del inóculo fresco para cada
fermentación y la recuperación de los productos en cantidades menores que cuando el proceso es continuo.
Actualmente se están desarrollando nuevos diseños de biorreactores para dar solución operacional a estos
problemas relacionados con los hongos filamentosos (Moreira et al., 2003).
La dispersión del micelio, comúnmente en fermentaciones líquidas industriales implica homogenización
y agitación de las hifas ramificadas. La dispersión del micelio produce un caldo con comportamiento no-
newtoniano y su viscosidad aparente se incrementa con la velocidad de agitación, reduciendo el transporte de
nutrientes, oxígeno y calor y por ende incrementando los costos de operación (Prosser y Tough, 1991). Este
crecimiento del micelio, normalmente forma pellets. Cuando estas esferas exceden al denominado radio
crítico, puede llevar a una disminución en el crecimiento de la masa celular. Esta situación aumenta las
dificultades, prácticas y técnicas del cultivo de hongos filamentosos en medio líquido y consecuentemente el
control y regulación de la extensión hifal y el tamaño del pellets se convierten en un factor de control crítico
en los procesos continuos (Allen y Robinson, 1989; Moreira et al., 2003). De hecho, los hongos de pudrición
blanca cada vez son más utilizados en procesos industriales para producir diferentes sustancias como
enzimas y polisacáridos y en procesos de biorremediación; por lo que las investigaciones ligadas a dichos
temas contribuirán a incrementar y optimizar sus aplicaciones industriales.
2.6. Fermentación en estado sólido
La fermentación en estado sólido involucra el desarrollo de organismos sobre sustratos sólidos en
ausencia de agua libre. En la última década se han incrementado sus aplicaciones en torno a la producción de
enzimas, antibióticos, surfactantes, biofertilizantes, bioinsecticidas, etc. así como también, en la adición de
valor a muchos residuos sólidos, ya que estos subproductos son utilizados como materias primas de estos
procesos.
Las ventajas de los usos biotecnológicos que presenta la fermentación en estado sólido sobre la
fermentación sumergida o fermentación clásica incluyen aspectos históricos, biológicos, ecológicos e
ingenieriles. Como aspectos históricos relevantes se puede mencionar que los procesos de fermentación en
estado sólido han sido convencionalmente utilizados por los asiáticos en la producción de koji ricos en
enzimas a partir de cepas de Aspergillus sp., bebidas alcohólicas como el sake, utilizando materiales
sacarificados de arroz, producción de colorantes, como el angkak (Monascus purpurea) y otros metabolitos y
sustancias de utilidad con la tecnología de fermentación en estado sólido a gran escala, realizando la
transferencia de tecnología entre las familias como secretos industriales. En occidente la tecnología de
fermentación en fase sólida debe competir con la fermentación sumergida, ya que a partir de la segunda
guerra mundial todos los desarrollos tecnológicos fueron orientados en torno a la fermentación sumergida.
Sin embargo, en la actualidad por la globalización, las tendencias han cambiado y ya se encuentran avances
en los procesos tecnológicos de las fermentaciones en estado sólido para la obtención de diversas sustancias,
así como para la transformación de una gran cantidad de materiales lignocelulósicos (Hölker y Lenz, 2005).
En referencia a los aspectos biológicos, la situación de mayor importancia probablemente es la condición de
vida natural de los hongos filamentosos, ascomicetos, basidiomicetos y deuteromicetos, los cuales tienen sus
hábitats silvestres sobre sustratos húmedos. Por esta condición la producción de enzimas, esporas o
metabolitos de los hongos se encuentran bien ajustados al desarrollo sobre sustratos sólidos. Como ejemplo,
la producción de esporas fúngicas por fermentación en estado sólido muestran una alta estabilidad, son más
resistentes a las épocas de sequía y exhiben mayores velocidades de de germinación por largos períodos de

tiempo, este hecho, probablemente se le atribuye a que las conidiosporas producidas en fermentación en
estado sólido tienen una alta hidrofobicidad o causado también por la formación de un enlace hidrofóbico en
las proteínas, una pared celular fuerte y un volumen pequeño (Munoz et al., 1995; Hölker y Lenz, 2005).
Utilizando fermentación en estado sólido con hongos filamentosos se ha logrado obtener mejores
rendimientos en la producción de enzimas, menores problemas de inhibición de crecimiento por sustratos y
mayor estabilidad de los organismos a cambios de temperatura y pH que cuando estos mismos organismos se
cultivan en medios líquidos, menor ocurrencia de degradación de enzimas por la presencia de proteasas
indeseables (Hölker et al., 2004). Interesantes estudios en este campo exploran las características fisiológicas
que se presentan durante el desarrollo de las células fúngicas en los procesos de FES. Ocurre una aparente
acumulación de glicerol, eritritol y arabinitol que generan una inducción de genes de glucoamilasas, hecho
que aparentemente está sujeto a la temperatura y actividad agua bajo estas condiciones de proceso. La
formación de metabolitos secundarios de numerosos hongos filamentosos (especialmente micromicetos) en
FES está asociada con la formación de las hifas aéreas y de esporas al inicio de la fase del metabolismo
secundario y únicamente descritas en este tipo de procesos y no en FS (Hölker y Lenz, 2005). Esta es otra
razón por la cual es necesario el desarrollo tecnológico en torno a FES, ya que existe mucho por explorar en
cuanto a la obtención de nuevas sustancias utilizando hongos filamentos en FES.
Una de las razones más serias que detienen los adelantos de los procesos FES son los problemas de
ingeniería. Los problemas de estandarización y escalamiento de los procesos limitan la reproducibilidad de
los resultados. El escalamiento en particular, genera un cuello de botella importante, debido al manejo de los
gradientes de temperatura, humedad, concentración de sustrato, los cuales pueden variar, incrementándose
durante el proceso, generando condiciones adversas en los procesos de cama sólida estática y en los reactores
de tambor giratorio u otros equipos de agitación intermitente. La interrelación entre las condiciones
ambientales, tales como concentración de oxígeno, niveles de humedad y temperatura contribuyen a la
dificultad de regular estos mismos parámetros. El desarrollo fúngico bajo condiciones aeróbicas en el
biorreactor resulta en un considerable incremento en la producción de calor causado por un aumento rápido
de la temperatura. Este efecto es deseable en procesos de compostaje, pero es totalmente adverso en procesos
biotecnológicos en biorreactores, ya que una gran parte de las enzimas producidas durante la FES pueden
desnaturalizarse al final del proceso. En ausencia de la fase acuosa libre el calor producido es muy difícil de
remover. Generalmente se utiliza para la remoción del calor metabólico, aireaciones inducidas que deben ser
cuidadosamente planeadas a fin de evitar la pérdida excesiva de agua del material sólido o el incremento de
la actividad agua del mismo, provocando un desequilibrio indeseable en el balance metabólico del proceso
estático, el cual puede contaminarse con otros organismos oportunistas. Toda esta situación descrita provoca
un desafío para nuevos desarrollos en cuanto al diseño y control de los procesos de FES, entendiendo mejor
los procesos biológicos, físicos y químicos que ocurren durante el desarrollo de la hifa en medios sólidos
(ver Figura 10) para así despejar las rutas de proceso más adecuadas para la industrialización de los procesos
FES (Muller dos Santos et al., 2004; Hölker y Lenz, 2005; Lenz et al., 2004). Aspectos tales como la
transferencia de masa y de calor, el diseño, control y escalamiento de los biorreactores en estado sólido
constituyen líneas de estudio de relevancia.
Los basidiomicetos de pudrición blanca se destacan por la producción de enzimas celulasas, xilanasas,
lacasas y manganeso peroxidasas en fermentaciones en estado sólido utilizando sustratos lignocelulósicos.
Perfiles de comparación sobre la producción de enzimas lignocelulolíticas hechos para los ciclos de vida de
cepas comerciales de Grifola frondosa, Pleurotus ostreatus y Lentinus edodes en fermentaciones sólidas
mostraron que las actividades de las lacasas y MnP fueron altas al final de la fase de colonización y
declinaron rápidamente en las fases reproductivas. En contraste con las actividades enzimáticas de celulasas
y xilanasas, las cuales expresaron actividades enzimáticas medianamente estables durante la colonización y
se incrementaron durante la fase reproductiva (Asatiani et al., 2008; Montoya et al., 2011a).
La selección de las materias primas para los sustratos de las fermentaciones en estado sólido depende de
los productos a obtener. En la Tabla 4 se muestran los materiales que mayoritariamente son utilizados como
materias primas para los procesos de fermentación en estado sólido, siempre obedeciendo a formulaciones
preestablecidas de acuerdo con las necesidades de cada proceso en particular. En la Tabla 5 se muestran
algunos materiales sólidos orgánicos naturales colombianos, los cuales hacen parte de los materiales

lignocelulósicos y que probablemente, son aptos para la producción de enzimas lignocelulolíticas y
polisacáridos por fermentación en estado sólido, con las relaciones carbono/nitrógeno indicadas, según los
resultados esperados.
Uno de los aspectos importantes a tener en cuenta durante el diseño y operación en los procesos de FES
es la transferencia de masa. La difusión del oxígeno y de los nutrientes generan restricciones importantes en
el desarrollo fúngico de estas fermentaciones. La transferencia de masa intrapartícula depende de factores
como el tamaño de la partícula y el contenido de humedad de los sustratos. Esta transferencia está ligada
directamente a la transferencia de los nutrientes y de las enzimas con el sólido. Existen muchas limitaciones
acerca de la relación entre los hongos y los sustratos sólidos sobre los que se desarrolla la fermentación. El
desarrollo de la biomasa celular en las fermentaciones sumergidas se da generalmente en forma de esferas
(pellets) y en torno a ellas giran los procesos de difusión de oxígeno y nutrientes que han sido ampliamente
estudiados. En las FES los fenómenos de transferencia de masa son diferentes, ya que la biomasa (hifa de los
hongos filamentosos) se desarrolla sobre y entre el material sólido que sirve como sustrato (Chen et al.,
2002; Lonsane et al., 2005).
Figura 10. Esquema de un proceso a micro escala durante una FES, la hifa produce un trenzado micelial el cual se
observa sobre las partículas que componen el sustrato sólido. Del trenzado micelial sobresalen unas hifas aéreas por
entre las cuales muy probablemente ocurre el intercambio gaseoso (g) (O2, CO2 y otros). En los espacios del entramado
micelial horizontal hay líquido (l). Las actividades metabólicas muestran que se producen principalmente dentro de la
superficie del sustrato y dentro de los poros; sin embargo, las regiones expuestas del micelio (por ejemplo las hifas
aéreas) también muestran un metabolismo y el transporte de sustancias de la penetración de las mismas hifas aéreas.
Enzimas hidrolíticas (azul claro), que son producidos por el micelio se difunden por la matriz sólida y catalizan la
degradación de las macromoléculas en unidades más pequeñas (verde), los que más tarde son tomados por el hongo
como nutrientes. El O2 se consume y el CO2, H20, el calor y los productos bioquímicos de interés se producen durante la
fermentación. Por lo tanto, los gradientes desarrollados dentro de la biopelícula como las fuerzas de difusión de O2 para
difundirse desde la fase gaseosa hasta las zonas más profundas de la biopelícula (lila) y la difusión del CO2 desde estas
regiones hasta la zona de difusión de los gases (rojo). La producción de calor (Q; naranja) conduce a un rápido aumento
de la temperatura (T), que es un problema grave durante el FES. El calor es por lo tanto eliminado del sustrato no sólo a
través de la conducción, sino también por la evaporación, que es parte del balance de agua complejo del sistema (azul
oscuro). Junto a la evaporación, el balance de agua incluye la absorción de agua por el micelio durante el crecimiento, el
consumo de agua durante las reacciones de hidrólisis y la producción de agua a través de la respiración. Otro factor
importante, es el pH local, que podría ser cambiado debido a la liberación de los ácidos carboxílicos y el intercambio de
amoníaco (gris). Los productos bioquímicos de interés (magenta) que se liberan en la matriz sólida y el líquido fijado
entre los espacios durante la fermentación podrían ser absorbidos por la matriz sólida y tener que ser extraídos para su

uso posterior al final del proceso. Todos estos y muchos otros fenómenos pueden influir fuertemente en el rendimiento
del proceso durante la FES. Fuente: Hölker y Lenz (2005).
Tabla 11. Enzimas ligninolíticas producidas en procesos de FES.
Material Organismo Enzima ligninolítica Referencia
Bagazo de caña
Polyporus BH1, Polyporus BW1 Ligninasas Nigam et al. (1987)
Trametes versicolor MnP, Lacasa Pal et al. (1995)
Pleurotus ostreatus, Phanerochaete
chrysosporium
MnP, LiP, Lacasa Pradeep y Datta (2002)
Borra de café y
aserrín de roble
Grifola frondosa MnP, LiP, Lacasa Montoya (2008)
Desechos de
banano
Pleurotus ostreatus, P. sajor-caju Lacasa, LiP Reddy et al. (2003)
Maíz y sus
residuos
Lentinus edodes, cepa CS-495 Lacasa D’Annibale et al. (1996)
Sporotrichum sp. Lignocelulasas Li et al. (2000)
Pleurotus pulmonaris Lacasa Tychanowicz et al. (2004)
Algodón
Pleurotus ostreatus, Pleurotus
cystidiosus, Pleurotus pulmonarius,
Pholiota nameko.
Lacasas, MnP
Jaszek et al. (1998)
Phanerochaete chrysosporium,
Funalia trogii
Lacasas, peroxidasas, celulasas Ünyayar y Sik (1998)
Residuos de Uva
Coriolus versicolor, Panus tigrinus,
Phanerochaete chrysosporium
Ligninasas
Golovleva et al. (1987)
Daedaleopsis confragosa, Marasmius
allaceus, Phanerochaete
chrysosporium
Lacasa, MnP
Elisashvili et al. (2001b)
Aserrín Rigidoporus lignosus MnP Galliano et al. (1991)
Coriolus hirsutus Lacasa, MnP Elisashvili et al. (2001b)
Paja de trigo Pleurotus eryngii, Pleurotus
pulmonarius
MnP Martínez et al. (1996)
Salvado de trigo
Pleurotus pulmonarius Lacasa Marques de Souza et al.
(2002)
Pleurotus ostreatus, Phanerochaete
chrysosporium
Lacasa, MnP, LiP Shinners-Carnelley et al.
(2002)
Fomes sclerodermeus Lacasa, MnP Papinutti et al. (2003)
Granos de trigo
Coriolus versicolor, Panus tigrinus,
Phanerochaete chrysosporium
Ligninasas
Golovleva et al. (1987)
Phlebia radiata Lacasa, MnP, LiP Vares et al. (1995)
Pleurotus spp. Lacasa, MnP Lang et al. (1996)
Phanerochaete chrysosporium LiP, MnP Castillo et al. (1997)
Pleurotus sp.2, Trametes gallica MnP, Lacasa Xie et al. (2001)
Phanerochaete chrysosporium LiP, MnP, Lacasa Fujian et al. (2001)
Pleurotus ostreatus Lacasa Baldrian y Gabriel (2002)
Pleurotus pulmonarius Lacasa Marques de Souza et al.
(2002)
Maderas
Bjerkandera sp. cepa BOS55 LiP, MnP Mester et al. (1998)
Ceriporiopsis subvermispora
MnP, laccase, hemicellulase,
cellulase Ferraz et al. (2003)
La transferencia de calor es uno de los mayores problemas a solucionar cuando se trabaja con FES. El
factor más determinante en este aspecto, es el calor metabólico producido, el cual depende de una serie de
características físico-químicas y ambientales del proceso. Al inicio de la fermentación, generalmente la
temperatura de los sustratos es homogénea, así como la concentración de oxígeno; pero con el progreso de la
fermentación, la difusión de oxígeno se limita; hay liberación de calor con dificultad para disiparlo por la
pequeña conductividad térmica de los sustratos, la porosidad del sustrato disminuye y toda la transferencia
de calor y masa se dificulta (Raghavarao et al., 2003). En este campo hace falta mucha investigación y
desarrollo acerca del control de la transferencia de calor y masa en los procesos de FES, debido a la

heterogeneidad de los materiales utilizados para la elaboración de los sustratos, y a las dificultades ligadas a
la manipulación de los hongos filamentosos y al control de las velocidades de desarrollo de las reacciones
bioquímicas intra y extra celulares.
Las áreas de aplicación de la FES son: aplicaciones para el control ambiental, las cuales incluyen la
producción de compost, la producción de alimento para animales utilizando residuos sólidos, los procesos de
biorremediación y biodegradación de compuestos difíciles de degradar, detoxificaciones biológicas de
residuos industriales. Como segunda gran aplicación, está la adición de valor a diferentes productos, como
enriquecimiento nutricional a residuos de cereales por biotransformación, biopulpado, producción de
alimentos fermentados, enzimas, pigmentos. En la Tabla11 se enumeran una serie de hongos de pudrición
blanca actuando sobre diversos materiales sólidos orgánicos (soporte), en su mayoría residuos
lignocelulósicos, con la respectiva enzima ligninolítica producida. En particular, la producción de enzimas
ligninolíticas es reportada con mejores actividades enzimáticas en los procesos de fermentación en estado
sólido. Por ejemplo, la MnP de Lentinus edodes y Pleurotus ostreatus sobre sustratos a base de hojas de
árboles y paja de trigo fue tres veces mayor que la actividad de la misma enzima, en fermentación sumergida
con los mismos materiales lignocelulósicos pretratados (Elisashvili et al., 2008).
2.7. Modelamiento de las cinéticas de fermentación de hongos de pudrición blanca
La cinética del crecimiento microbiano se relaciona con el cálculo de las velocidades de reacción de
formación de biomasa celular, velocidad de formación de producto y de CO2. En la Tabla 12 se enumeran
algunas de las ecuaciones más utilizadas para perfiles cinéticos cuando se espera describir procesos de
fermentación sumergida y en fase sólida. De igual forma, estas ecuaciones han sido utilizadas para la
descripción de la producción de diversas biosustancias utilizando bacterias y hongos en dichos procesos de
fermentación. Los modelos más reportados son el modelo lineal, logístico y de dos fases (de fase de
aceleración y desaceleración) (Sangsurasak et al., 1996). Estos modelos no incluyen el efecto de la
concentración de nutrientes sobre el desarrollo. Las ecuaciones lineal, exponencial y logística han utilizado
ampliamente, mientras que la ecuación de dos fases es mas reciente (Ikasari y Mitchell, 2000). En el modelo
de dos fases, la fase exponencial es seguida por la fase de desaceleración, cuya expresión está representada
por la ecuación (4b) (ver Tabla 12), la cual está afectada por un decrecimiento, que se debe probablemente a
dos factores. Primero a que el instante en el que se inicia la fase de desaceleración es el tiempo ta, con el
parámetro L representado como la relación entre las velocidades específicas de desarrollo de las fases de
desaceleración y de aceleración, respectivamente. Segundo, la desaceleración se describe por un
decrecimiento exponencial reflejado en la velocidad específica de desarrollo, con una constante de velocidad
de primer orden k (ecuación (4b)). Una dificultad para aplicar este modelo es que el período exponencial es
típicamente corto, pero se ha indicado cuando se requieren ajustes de procesos con pocos datos
experimentales (Mitchell et al., 2004; Van de Lagemaat y Pyle, 2005). Asimismo, para la descripción de la
producción de biomasa en diversos tipos de procesos de fermentación se utilizan las expresiones tipo Monod
y Logística modificada, las cuales en la Tabla 12 corresponden a las ecuaciones (5) y (6) respectivamente
(Mitchell et al., 2004; Tavares et al., 2005). Las expresiones tipo Monod para describir la producción de
biomasa presentan múltiples variaciones de la representación del modo del desarrollo cinético con la
obtención de la velocidad específica de desarrollo de los hongos filamentosos macroscópicamente; ya que lo
que se busca es relacionar la velocidad de un segmento de biomasa, una hifa, antes de su ramificación,
buscando entender como después de pocas ramificaciones simétricas de la biomasa entre el sustrato sólido,
ésta se extiende activamente en segmentos de micelio equivalentes a la mitad de la biomasa anterior. Una
aproximación para describir este desarrollo del micelio debe estar relacionado con la concentración de
sustrato limitante, como los azúcares solubles o el oxígeno, pudiéndose incorporar este efecto de la
concentración de sustrato limitante en el modelo cinético, sólo si estas expresiones son acopladas con las
ecuaciones de transporte local de nutrientes (Mitchell et al., 2004; Tavares et al., 2005). La ecuación
logística modificada (ecuación (6)) ha sido propuesta para procesos de FES en los cuales el incremento en la

producción de biomasa se debe a la equivalencia entre el incremento total de biomasa y la disminución total
de masa seca; lo que infiere, como los diferentes tipos de microorganismos con los que se trabaja en procesos
de FES pueden presentar una aceleración temprana en su crecimiento, seguido de un período extendido en el
cual la desaceleración es lenta. Este perfil puede ser aproximado por el incremento de la relación (X/Xm)
elevado a un exponente n. El valor del coeficiente n de la ecuación (6) determina la sensibilidad relativa de la
auto-inhibición del cultivo por la alta densidad de biomasa (Mitchell et al., 2004). La ecuación (7) de la Tabla
12 fue propuesta por Luedeking y Piret (1959) inicialmente para la producción de lactato por bacterias. Sin
embargo, esta ecuación ha sido utilizada con éxito para describir la producción de enzimas y otras
biosustancias de hongos filamentosos en procesos de FS y FES tomando la misma expresión o haciéndole
modificaciones que permitan la descripción del proceso de producción de una biosustancia en particular
(Aguilar et al., 2001; Tavares et al., 2005; Shi et al., 2012).
Tabla 12. Formas de ecuaciones diferenciales que son aplicadas en sistemas de fermentación
Forma ecuación Ecuación
Lineal
(1)
Exponencial
(2)
Logística
(3)
Dos fases
(4a)
(4b)
Producción de biomasa en
fase exponencial tipo
Monod
(5)
Logística modificada
(6)
n<1 el organismos es relativamente sensible a ala auto-inhibición y ésta
ocurre para valores bastante bajos de X.
Para n=1 ecuación logística.
n>1 el organismo es relatovamente resistente a la auto-inhibición y ésta
ocurre solo cuando X≈Xm
Modelo Luedeking–Piret
(7)
*X es biomasa microbial, t es tiempo, k es la constante de velocidad de desarrollo lineal, µ es la velocidad específica de
desarrollo constante, Xo es la biomasa inicial, Xm es la máxima cantidad de biomasa posible, ta, L y k son parámetros
para la expresión de dos fase que son explicadas en el texto (Mitchell et al., 1999; Mitchell et al., 2004; Tavares et al.,
2005; Shi et al., 2012)
dXkX
dt
dXX
dt
1m
dX XX
dt X
, a
dXX t t
dt
,ak t t
aLe XdX
t tdt
max
s
SdXX
dt k S
1
n
m
dX XX
dt X
dXdP
dtX
dt

Las sustancias que se producen en un proceso de fermentación pueden pertenecer a diversos grupos de
metabolitos, los cuales pueden ser productos finales de metabolismo energético, productos intermedios de
metabolismo primario o productos de metabolismo secundario. Las expresiones matemáticas utilizadas para
la descripción de la formación de productos dependen de qué clase de metabolito se esté produciendo;
generalmente las expresiones más simples son para el primer grupo, ya que su formación se considera
directamente proporcional a la formación de la biomasa celular y se tienen en cuenta los parámetros de
mantenimiento celular y crecimiento microbiano, respecto de la formación de la misma sustancia; en el caso
de los tipos de sustancias que corresponden a las clases productos intermedios de metabolismo primario y
productos de metabolismo secundario, la cinética de formación de estos productos es más compleja que los
del primer grupo, y no existen hasta el momento expresiones simples bien definidas que abarquen a todas las
sustancias pertenecientes a estos grupos. Existen aún, varios interrogantes con relación a los mecanismos que
controlan la síntesis de los metabolitos secundarios y su interrelación con los metabolitos primarios.
Actualmente, no hay modelos matemáticos disponibles que permitan describir la producción de biosustancias
de hongos macromicetos, ni su crecimiento; probablemente porque cada uno de estos hongos se ve afectado
directamente por diversos factores intrínsecos y extrínsecos, como son las características físicas y la
composición química de los medios de cultivo, la procedencia de las cepas, la influencia de las condiciones
ambientales y el crecimiento lento asociado a la gran mayoría de las especies de macromicetos en
comparación con el tiempo de crecimiento de los hongos micromicetos.
La formación de los productos en procesos de fermentación tiene relación directa con la formación de
biomasa celular. En general la velocidad de formación de los productos es directamente proporcional a la
formación de biomasa celular y a su velocidad específica de formación. Esta relación, radica en que dichas
sustancias son metabolitos producidos durante el proceso de fermentación, metabolitos primarios y
secundarios. Sin embargo, lo relevante, no corresponde únicamente al conocimiento sobre si la sustancia de
interés es un metabolito primario o secundario por la necesidad de determinar las fases de producción de la
misma sustancia; sino porque, es muy importante conocer la relación o interrelación que existe entre el
término de la producción de metabolitos primarios, la cual está asociada directamente con el crecimiento y
desarrollo del organismo y el inicio de la fase estacionaria, que es la etapa en la que normalmente se forman
los metabolitos secundarios. Asimismo, para el conocimiento de formación de productos en procesos de
fermentación es necesario conocer varias de las sustancias que se forman durante el crecimiento y desarrollo
del organismo a fin de comprender el mecanismo de producción de la sustancias de interés y lograr plantear
expresiones matemáticas que permitan la descripción y escalamiento de su proceso productivo.
El método Plackett–Burman (PB) (Plackett y Burman, 1946), ha sido propuesto para la optimización de
las variables del procesos de fermentación sumergida, así como el uso de superficies de respuesta (SR) para
realizar optimizaciones de estos procesos (Liu et al., 2003). Utilizando estas técnicas no es posible
cuantificar cada una de las variables a medir en el proceso, pero si son una herramienta válida que indica la
tendencia de cada factor y su relativa influencia en el mismo (Xu et al., 2008). Por ejemplo, la producción de
polisacáridos extracelulares de Morchella esculenta As51620 se ve afectada por el tiempo de fermentación y
composición del medio; mientras que el desarrollo de la biomasa celular se ve influenciada por la
temperatura y la composición del medio. Pudiéndose tratar estos tres factores (temperatura, tiempo de
fermentación y composición del medio) como una sola variable utilizando herramientas de superficie de
respuesta (SR). Con esta técnica se puede dar explicación al efecto de la combinación de todos los factores
en los procesos de fermentación sumergida, utilizando los datos experimentales, los métodos matemáticos y
la inferencia estadística (Elibol, 2004; Xu et al., 2008).
La descripción de procesos de fermentación, bien sea FS o FES, con herramientas matemáticas que
permitan avanzar en el conocimiento de la formación de biomasa celular, de los productos y el consumo de
los sustratos no han sido completamente esclarecidos. Diferentes investigadores han propuesto diversos
modelos matemáticos a fin de aproximar la descripción del crecimiento y formación de biomasa, el consumo
de sustrato y la formación de productos. Sin embargo, los procesos de fermentación presentan gran
complejidad y diversidad en referencia a la composición de los sustratos, la selección de los
microorganismos y la formación de productos. Los sustratos utilizados en los procesos de fermentación en
general son multicomponentes, hecho que no ha posibilitado encontrar expresiones matemáticas que

permitan una descripción general del consumo de los sustratos; así como tampoco encontrar expresiones para
la formación de los productos. En referencia a la formulación de expresiones matemáticas para describir la
formación de productos en procesos de fermentación se dificulta por la gran cantidad de sustancias que se
forman durante el crecimiento y desarrollo del organismo como parte de su mecanismo para lograr que se
den las reacciones metabólicas que finalmente posibilitarán la obtención de las sustancias de interés.
No hay suficiente literatura disponible referente al planteamiento de modelos matemáticos que
describan la formación de biomasa celular, consumo de sustratos o formación de productos con hongos
macromicetos de pudrición blanca tanto en procesos de FS, como en procesos de FES. Los hongos
macromicetos son organismos que presentan dos fases de crecimiento bien definidas, una fase vegetativa y
una fase reproductiva, son filamentosos y hacen que las hifas se envuelvan entre sí formando pellets cuando
están en medio líquido y agitación y cuando están en medio sólido colonizan alrededor y entre los sustratos
sólidos. Investigadores como Tišma et al.(2010), Tavares et al.(2005), Shi et al. (2012) y otros han propuesto
modelos matemáticos para la descripción del crecimiento de hongos de pudrición blanca y la formación de
enzimas, polisacáridos y otras biosustancias en procesos de FS y FES. Estos modelos matemáticos han sido
propuestos a partir de modelos y ecuaciones tipo, como son la ecuación de Mitchaelis-Menten y varias de las
que se describen en la Tabla 12. Estas expresiones matemáticas han sido modificadas o aplicadas
directamente por los investigadores para el ajuste de datos experimentales, empleando como variables
diversos componentes de los sustratos, diferentes tipos de productos como enzimas, polisacáridos (intra y
exo), además de parámetros asociados a los procesos de fermentación como son el pH, la actividad de agua,
temperatura del proceso, humedad relativa, concentración de oxígeno, entre otros (Hormiga et al., 2008;
Rocha et al., 2009; Sriyudthsak y Shiraishi, 2010; Shi et al., 2012). Otra de las dificultades que se presentan
cuando se busca proponer modelos matemáticos con el fin de ajustar datos experimentales en estos procesos
de fermentación, es la ausencia de expresiones que permitan determinar la mejor aproximación del ajuste de
los datos experimentales con el modelo propuesto. Sin embargo, algunos investigadores han buscado aplicar
expresiones utilizadas en matemáticas financieras y estadística con fines de encontrar parámetros que
permitan comparar la aproximación de un modelo matemático propuesto para el ajuste de un grupo de datos
experimentales dado como lo proponen Rocha et al. (2009).
2.8. Evaluación de biomasa fúngica, enzimas lignocelulolíticas y polisacáridos
2.8.1. Determinación de biomasa celular
En los procesos de FES la estimación directa de la biomasa es usualmente imposible, por la dificultad
de la separación eficiente de la biomasa de la matriz sólida (sustrato). La biomasa de los hongos filamentosos
puede ser estimada indirectamente por la determinación de componentes celulares de las colonias fúngicas,
tales como la N-acetil-D-glucosamina (NAGA), presente en la quitina como componente de la pared celular
fúngica; el ergosterol presente en la membrana celular; por la determinación de proteínas o ácidos nucleicos;
o se puede estimar la biomasa por medio del balance estequiométrico cuando se conoce el valor del dióxido
de carbono formado (De Carvalho et al., 2005). El ergosterol es un componente celular que favorablemente
puede ser usado para la determinación de la biomasa fúngica por su fácil extracción y análisis. Este
compuesto es utilizado para la determinación de la biomasa fúngica en sustratos sólidos, porque es el esterol
más importante de la membrana celular de los hongos y algunas microalgas, estando prácticamente ausente
en plantas y animales (Gong et al., 2001; Mattila et al., 2002).
El análisis de ergosterol fue inicialmente propuesto para estimar contaminaciones de cereales por
hongos, lo que llevó este análisis a ser adoptado para la estimación de la biomasa fúngica en materiales
sólidos (Montgomery et al., 2000 ). Actualmente, el ergosterol puede ser tomado como una representación
adecuada de la cuantificación de biomasa total (células vivas) en procesos de FES, ya que permanece durante
todo el proceso (Mattila et al., 2002; Lindblom et al., 2004; Zhao et al., 2005). La determinación del
ergosterol se puede realizar utilizando cromatografía líquida de alta resolución (HPLC) con detector arreglo
de diodos (PDA) y columna C18, iniciándose la detección del ergosterol a 282 nm, con una fase móvil de
metanol puro y acetonitrilo. La extracción de las muestras en procesos de fermentación se realiza con etanol

grado analítico, hidróxido de sodio y n-hexano para obtener una fase liviana y una fase pesada, evaporándose
posteriormente la fase orgánica al vacío (Mattila et al., 2002; Lindblom et al., 2004; De Carvalho et al.,
2005; Zhao et al., 2005).
Los principales componentes de la pared celular fúngica son los polisacáridos, los cuales corresponden
entre el 80 y 90% de la masa seca. Entre los que se encuentra la quitina, polisacárido que se encuentra
formando parte del esqueleto fungal, responsable de la rigidez y la forma de la pared celular. La quitina es el
componente característico de los grupos taxonómicos Zygo-, Asco-, Basidio-, y Deuteromycetes, estando
ausentes en otros grupos como Oomycetes. Por lo tanto, el contenido de quitina es característico de las
especies de hongos verdaderos e independientes de la variedad. Asimismo, los niveles de quitina de los
hongos pueden estar regulados por el tipo de nutrición de cada especie (Vetter, 2007). El contenido de
quitina puede determinarse por el método de valoración de Glucosamina (N-acetil-D-glucosamina: NAGA)
(Plassard et al., 1982) en las muestras de sustratos colonizadas por los hongos filamentosos. Las muestras de
sustrato fermentadas se someten a hidrólisis ácida con HCl 6N, con posterior alcalinización y reacción del
hidrolizado alcalinizado con reactivos preparados a partir de acetilacetona y p-dimetilamino benzaldehído
por espectrofotometría a 530 nm. Se realiza una curva patrón con una solución de D-(+)-clorhidrato de
glucosamina. Es así como varios investigadores han publicado resultados del contenido de quitina de
diversas especies de hongos basidiomicetos entre el 1 y 25% presentes en la pared celular y cuantificada por
varios métodos (Ivshin et al., 2007; Nitschke et al., 2011).
La biomasa celular en procesos de FS se determina por peso seco cuando los medios de cultivo permiten
la separación completa entre la biomasa y el sustrato por filtración. Sin embargo, hay procesos de FS en los
que los medios de cultivo tienen material sólido en suspensión que hace parte del sustrato, por ejemplo
materiales lignocelulósicos pretratados, en estos casos es necesario determinar también el contenido de
quitina o ergosterol a fin de disminuir el error en el que se pueda incurrir cuando se determina el incremento
de la biomasa por peso seco.
2.8.2. Rastreo de cepas para producción de biosustancias
Cuando se está buscando que un organismo produzca una sustancia en particular, se debe seguir algunos
lineamientos para seleccionar el organismo: esta selección puede realizarse a través de búsquedas
bibliográficas especializadas según la sustancia que se requiera. Puede hacerse un rastreo a los organismos
con el mejor potencial para la producción de la sustancia de referencia. Existen métodos para rastrear
organismos, entre los que se encuentran los métodos para seleccionar hongos basidiomicetos de pudrición
blanca a fin de visualizar su capacidad de producción diversas sustancias. En el caso de ligninasas, se
conocen los métodos de Kirk y Farrel, (1987) modificado y Zhao et al., (1996), asímismo, se encuentran
métodos de rastreo para celulasas (Mandels y Weber, 1969; Montenecourt y Eveleigh, 1977; Zhang et al.,
2006a; Sohail et al., 2009) producidas especialmente por hongos. Estos métodos usualmente son cualitativos
o semicuantitativos, ya que una de las principales características que deben cumplir estos procedimientos es
que sean de fácil ejecución. La forma como se detectan las diferentes sustancias puede ser por
decoloraciones en las zonas de crecimiento, por formación de halos de diversos colores como reacción a
algún reactivo en particular, entre otros. Sin embargo, cuando se va a rastrear el contenido de intra o exo-
polisacáridos es necesario recurrir a métodos cuantitativos a tiempos fijos de crecimiento y producción del
polisacárido, bien sea por FS o por FES empleando métodos espectrofotómetricos como la determinación del
contenido de polisacáridos como carbohidratos totales por el método fenol-sulfúrico.
Los métodos cualitativos constituyen una buena herramienta para la búsqueda rápida (rastreo) de
enzimas que degraden el complejo lignocelulósico (Pointing, 1999; Nazareno et al., 2000). Estas pruebas
proporcionan una indicación certera de la presencia de estas enzimas con el fin de realizar una adecuada
selección de cepas de hongos respecto al grupo de enzimas que producen, disminuyendo la cantidad de
pruebas cuantitativas, que son mucho más costosas y demandan mayor tiempo. Sin embargo, estos métodos
tienen limitaciones especialmente de precisión, aunque juegan un rol importante en procesos de selección de
cepas (Montenecourt y Eveleigh, 1977; Takano, 2001).

Actualmente se emplean varios métodos cualitativos para la detección de celulasas, entre los que se
encuentran los medios sintéticos con el reactivo específico sobre el cual actúa la enzima. Por ejemplo, para la
detección de la endoglucanasa se emplea carboximetil celulosa (CMC), para exoglucanasas se usa celulosa
cristalina o agar azure celulosa. Estas dos enzimas o grupo de enzimas generalmente se detectan por la
formación de un halo blanco alrededor de la colonia. Para la β-glucosidasa se emplea agar esculina
(6,7dihidroxicumarino 6-glucósido) como fuente única de carbono. La enzima β-glucosidasa forma glucosa,
la cual es excretada al medio y forma un halo negro en torno a la colonia; este halo negro también puede
formarse cuando se realizan las pruebas para las enzimas celulolíticas, endoglucanasas y exoglucanasas
(Pointing, 1999; Zhang et al., 2006b). Para detectar ligninasas, se emplean reactivos que las óxido-reductasas
puedan degradar. Por ejemplo, para la detección de ligninasas como la enzima lignina peroxidasa (LiP) se
utiliza azure B, el cual se decolora por la acción de esta enzima (Archibald, 1992) y la detección de lacasa se
hace a través de la preparación de un medio con el reactivo ABTS (2,2'azino- bis (3-etilbenzo-tiazolin-6-
ácido sulfónico)), el cual se oxida en presencia de la enzima lacasa (Thurston, 1994; Eggert et al., 1996).
2.8.3. Determinación de actividades enzimáticas lignocelulolíticas
Las actividades enzimáticas lignocelulolíticas se determinan utilizando sustratos específicos para cada
una, en los tiempos y temperaturas de reacción necesarios para que la enzima ejerza su acción sobre el
sustrato. En la Tabla 13 se enumeran las enzimas con los sustratos específicos de reacción y las longitudes de
onda a las que deben ser leídas.
Tabla 13. Métodos para determinación de actividad de enzimas lignocelulolíticas
Actividad enzimática Sustrato Longitud de onda (nm) Referencia
Endoglucanasa
Carboximetilcelulosa
(determinación final de azúcares
reductores producidos)
540 Somogyi (1945); Nelson
(1944)
Exoglucanasa
Celulosa cristalina
(determinación final de azúcares
reductores producidos)
540 Somogyi (1945); Nelson
(1944)
β-glucosidasa p-nitrofenil ß-D-glucopiranósido 430 Nidetzky et al. (1994)
Endoxilanasa Xilano (determinación final de
azúcares reductores producidos) 540
Somogyi (1945); Nelson
(1944)
LiP Azure B 650 Archibald (1992)
Lacasa 2,2´-azinobis (3-etilbenztiazoline-
6-sulfonato) (ABTS) 420 Paszczczynski et al. (1988)
MnP Sulfato de manganeso y rojo
fenol 610 Paszczczynski et al. (1988)
2.8.4. Determinación de polisacáridos
Para determinar el contenido y la clase de polisacáridos presentes en los hongos hay una gran variedad
de métodos de extracción y técnicas para su determinación. Los métodos de extracción son seleccionados
dependiendo de la estructura que se espera identificar o cuantificar; estos métodos van desde una sola
extracción con un solvente como etanol cuando se requiere cuantificar la cantidad total de polisacáridos
como carbohidratos totales realizando algunas modificaciones al método fenol-sulfúrico de Dubois et al.
(1956), hasta completos esquemas de extracción que generalmente inician con un desengrasado de la muestra
empleando solventes orgánicos no polares o mezclas entre solvente polares y no polares como metanol,
hexano, éter de petróleo, tetracloruro de carbono, acetona, etc. La muestra sin grasa puede ser sometida una
serie de extracciones con agua y agua con bases (KOH, NaOH) y sometidos a ebullición por períodos de
tiempo que pueden estar entre las dos y cinco horas a ebullición con el fin de lograr extraer la mayor
cantidad de fracciones de polisacáridos tanto solubles como insolubles (Chihara et al., 1970; Amaral et al.,
2008; Nie y Xie, 2011). Los polisacáridos de los hongos hacen parte de su pared celular, varios de ellos son
solubles en agua, otros no, por lo que el uso de soluciones básicas como solventes busca solubilizar parte de

los polisacáridos insolubles en agua. Estas fracciones obtenidas en cada extracción deben ser sometidas a
procesos de lavado con exceso de agua destilada o bidestilada en diálisis abiertas o cerradas; además, de
someter las muestras a congelación, descongelación y posterior centrifugación a fin de retirar los excesos de
álcali y otras sustancias que pudieron ser extraídas durante los procesos de extracción y que no hacen parte
del grupo de sustancias bioactivas buscadas. Asimismo, cada una de las fracciones obtenidas debe ser
sometida a ultrafiltración a fin de obtener las sustancias puras que deben ser identificadas por algún método
cromatográfico, como HPLC con detector de índice de refracción, método con el cual se puede identificar la
sustancia y cuantificarla.
Respecto a la determinación de los β-glucanos, existen métodos de detección por su especificidad, como
los que emiten fluorescencia, FIA-calcofluor. Los β-glucanos se unen al colorante calcofluor, cuando esto
ocurre, la intensidad de los tintes aumenta en una medida proporcional a la cantidad de β-glucano en la
muestra. La fluorescencia se mide a una longitud de onda de exitación y emisión de 360 y 425 nm
respectivamente. Debido a que el calcofluor es específico para β-glucanos, la muestra no tiene que estar pura,
aunque el calcofluor si puede unirse a sustancias como xiloglucanos o algunos derivados de celulosa, por lo
que la muestra deberá estar libre de estos componentes (Izawa et al., 1995; Brummer y Cui, 2006).
REFERENCIAS
Adachi Y., Ohno N., Ohsawa M., Oikawa S., Yadomae T. (1990). Change of biological activities of (1-3)-B-D glucan from Grifola
frondosa upon molecular-weight reduction by heat-treatment. Chemical and Pharmaceutical Bulletin, 38: 477-481.
Adler E. (1977). Lignin chemistry. Past, present and future. Wood Science Technology, 11: 169-218.
Aguilar C., Augur C., Favela-Torres E., Viniegra-Gonzalez G. (2001). Production of tannase by Aspergillus niger Aa-20 in
submerged and solid-state fermentation: influence of glucose and tannic acid. Journal of Industrial Microbiology and
Biotechnology Advances, 26: 296-302.
Ali H. (2010). Biodegradation of Synthetic Dyes-A Review. Water Air Soil Pollut, 213: 251-273.
Alkorta I., Garbisu G., Llama M.J., Serra J.L. (1998). Industrial applications of pectic enzymes: a review. Process Biochemistry, 33:
21-28.
Allen D.G., Robinson C.W. (1989). Hydrodynamics and mass transfer in Aspergillus niger fermentations in buble column and loop
bioreactors. Biotechnology and Bioengineering 34: 731-740.
Alvira P., Tomás-Pejó E., Ballesteros M., Negro M.J. (2009). Pretreatment technologies for an efficient bioethanol production
process based on enzymatic hydrolysis: A review. Bioresource Technology, doi: 10.1016/j.biortech.2009.11.093.
Amaral A., Carbonero E., Simao R., Kadowaki M., Sassaki G., Osaku C., Gorin P., Lacomini M. (2008). An unusual water-soluble
B-glucan from the basidiocarp of the fungus Ganoderma resinaceum. Carbohydrate Polymers, 72: 473-478.
Amat-García E.C., Amat-García G.D., Henao L.G. (2001). Diversidad taxonómica y ecológica de la entomofauna micófaga en un
bosque alto-andino de la cordillera oriental de Colombia. Revista Academia Colombiana de Ciencia 1-21.
Ander P. (1994). The celloiose-oxidizing enzymes CBQ and CBO as related to lignin and cellulose degradation - a review. FEMS
Microbiology Reviews, 13: 297-312.
Ângelo A.S. (2004). Enzimas hidrolíticas. En: Fungos: Uma Introducâo à biologia, bioquimica e biotecnologia. Universidade. de
Caxias do Sul. pp. 263-285.
Arantes V., Milagres A.M.F. (2007a). The effect of a catecholate chelator as a redox agent in Fenton-based reactions on degradation
of lignin-model substrates and on COD removal from effluent of an ECF kraft pulp mill. Journal of Hazardous Materials, 141:
273-279.
Arantes V., Milagres A.M.F. (2007b). The synergistic action of ligninolytic enzymes (MnP and Laccase) and Fe3+-reducing activity
from white-rot fungi for degradation of Azure B. Enzyme and Microbial Technology, 42: 17-22.
Archibald F. (1992). A new assay for lignin-type peroxidases employing the dye Azure B. Applied Environmental Microbiology, 58:
3110-3116
Arias E.L., Piñeros P.A. (2008). Enzimas ligninolíticas fúngicas para fines ambientales.Trabajo de grado. Facultad de Ciencias,
Pontificia Universidad Javeriana: Bogotá. 202 p. p.
Arteaga-Hoyos A.B. (2002). Utilización de los desechos del banano para el cultivo del hongo comestible Pleurotus ostreatus. Augura
Carta Informativa, (212): 4-6.
Asatiani M.D., Kachlishvili E.T., Khardziani T.S., Metreveli E.M., Mikiashvili N.A., Songulashvili G.G., Tsiklauri N.D., Wasser
S.P., Elisashvili V.I. (2008). Basidiomycetes as a source of antioxidants, lectins, polysaccharides, and enzymes. Journal of
Biotechnology 136S: S717-S742.
Bajpai P. (1997). Microbial xylanolytic enzyme system: properties and applications. Advances in Applied Microbiology, 43: 141-194.
Baker R.A., Wicker L. (1996). Current and potential applications of enzyme infusion in the food industry. Trends Food Science
Technology, 7: 279-284.
Baldrian P., Gabriel J. (2002). Variability of laccase activity in the whiterot basidiomycete Pleurotus ostreatus. Folia Microbiology,
47: 385-390.

Bao W., Fukushima Y., Jensen K., Moen M., Hammel K. (1994). Oxidative degradation of non-phenolic lignin during lipid
peroxidation by fungal manganese peroxidase. FEBS Letters, 354: 297-300.
Barr B.K., HsieH Y.L., Ganem B., Wilson D.B. (1996). Identification of two functionally different classes of exocellulases.
Biochemistry, 35: 586-592.
Béguin P., Aubert J.P. (1994). The biological degradation of cellulose. FEMS Microbiology Reviews, 13: 25-58.
Beily P., Kratky Z., Vrsanska M. (1981). Substrate binding site of endo-1,4- β-xylanase of the yeast Cryptococcus albidus. Europe
Journal Biochemestry, 119: 559 -571.
Benavidez-Calvache O.L. (2004). Estudio Químico de la fracción insaponificable del hongo macromiceto Lentinula edodes
(Shiitake).Maestría en Ciencias-Química. Programa de Postgrados en Química, Universidad Nacional de Colombia: Bogotá. 98
p.
Blanch H.W., Clark D.S. (1997). Biochemical Engineering. Marcel Dekker: New York. 702 pp. p.
Blanchette R.A. (1995). Degradation of the lignocellulose complex in wood. Canadian Journal of Botany, 73: S999-S1010.
Blanchette R.A., Krueger E.W., Haight J.E., Akhtar M., Akin D.E. (1997). Cell wall alterations in loblolly pine wood decayed by the
white-rot fungus, Ceriporiopsis subvermispora. Journal of Biotechnology, 53 203-213.
Blánquez P., Casas N., Font X., Gabarrell M., Sarrá M., Caminal G. (2004). Mechanism of textile metal dye biotransformation by
Trametes versicolor. Water Resource, 38: 2166-2172.
Blánquez P., Sarrà M., Vicent M. (2006). Study of the cellular retention time and the partial biomass renovation in a fungal
decolourisation continuous process. Water Resource, 40: 1650-1656.
Blánquez P., Caminal G., Sarrà M., Vicent M. (2007). The effect of HRT on the decolourisation of the Grey Lanaset G textile dye by
Trametes versicolor. Chemistry Engineering Journal, 126: 163-169.
Boisset C., Fraschini C., Schülein M., Henriss B. (2000). Imaging the enzymatic digestion of bacterial cellulose ribbons reveals the
endo character of the cellobioydrolase Cel6A from Humicola insolens and its mode of synergy with cellobiohydrolse Cel 7A.
Environmental Microbiology, 66: 1444-1452.
Bommarius A.S., Riebel B.R. (2004a). Biocatalysis. Wiley-VCH Verlag GmbH & Co. KGaA: Atlanta, USA.
Bommarius A.S., Riebel B.R. (2004b). Biocatalysis. Wiley-VCH Verlag GmbH & Co. KGaA: Atlanta, USA. 611 p. p.
Borchers A., Keen C., Gerhwin M. (2004). Mushrooms, tumors, and immunity: an update
. Experiment in Biological Medical, 229: 393-406.
Bosell G.P., Jacobs H., Davidson F.A., G.M. G., Ritz K. (2003). Growth and Function of Fungal Mycelia in Heterogeneous
Environments. Bulletin of Mathematical Biology, 65: 447-477.
Brummer Y., Cui S.W. (2006). Detection and Determination of Polysaccharides in Foods. En: Food Polysaccharides and Their
Applications. Stephen A.M., Phillips G.O., Williams P.A. (Eds.). CRC Press Taylor & Francis Group: Boca Raton, FL. 676-706
p. pp. 676-706.
Burgstaller W., Schinner F. (1993). Leaching of metals with fungi. Journal of Biotechnology, 27: 91-116.
Burnett J.H. (1976). Fundamental of Micology. 2a ed Edward Arnold: London. 221 p. p.
Burns J.K. (1991). The polygalacturonases and lyases. En: The Chemistry and Technology of Pectin. Walter R.H. (Ed.). Academic
Press,: San Diego. pp. 165- 188.
Buswell J.A., Odier E. (1987). Lignin biodegradation. Review Biotechnology, 6: 1-59.
Buswell J.A., Cai Y., Chang S.T. (1995). Effect of nutrient nitrogen and manganese on manganese peroxidase and laccase production
by Lentinula (Lentinus) edodes. FEMS Microbiology Letters, 128: 32- 45.
Campbell M.K., Farrell S.O. (2004). Bioquímica. Cuarta ed International Thomson Editores: México. 725 p. p.
Cardona B.E., Saldarriaga Y., Guzmán L. (2005). Registro de Gimnopilus rugulosus (Agaricales, Cortinariaceae) de Colombia.
Revista Mexicana de Micología, 021: 55-58.
Cardona M., Osorio J., Quintero J. (2009). Degradación de colorantes industriales con hongos ligninolíticos. Revista Facultad de
Ingeniería de la Universidad de Antioquia, (48): 27-37.
Carle - Urioste J.C., Escobar - Vera J., El-Gogary S. (1997). Cellulase induction in Trichoderma reesei by cellulose requires its own
basal expression. Journal of Biological Chemistry, 272: 10169-10174.
Carlile M., Watkinson S., Gooday G. (2001). The fungi. Second ed Academic Press: London. 588 p. p.
Carreño N., Vargas A., Bernal A.J., Restrepo S. (2007). Problemas fitopatológicos en especies de la familia Solanaceae causados por
los géneros Phytophthora, Alternaria y Ralstonia en Colombia. Una revisión. Agronomía Colombiana, 25(2): 320-329.
Castillo M.d.P., Ander P., Stenstrom J. (1997). Lignin and manganese peroxidase activity in extracts from straw solid substrate
fermentation. Biotechnol. Technol. , 11 701-706.
Castro-Ríos K. (2006). Validación de deshidratación convencional para la conservación del hongo comestible Pleurotus sajor-caju
Revista Universidad de Caldas: 123-133.
Catley B.J. (1992). The biochemistry of some fungal polysaccharides with industrial potencial. En: Handbook of applied mycology:
fungal biotechnology. Mukerji K.G. (Ed.). Marcel Dekker,: New York. pp. 1091-1114.
Chandra M., Kalra A., Sharma P.K., Sangwan R.S. (2009). Cellulase production by six Trichoderma spp. fermented on medicinal
plant processings. Journal Industrial Microbiology Biotechnology, 36: 605-609.
Chandra R.P., Bura R., Mabee W.E., Berlin A., Pan X., Saddler J.N. (2007). Substrate pretreatment: the key to effective enzymatic
hydrolysis of lignocellulosics? Advance Biochemistry Engineering and Biotechnology, 108: 67-93.
Chang S.T., Miles P.G. (2004). Mushrooms Cultivation, Nutritional Value, Medicinal effect, and Enviromental Impact. Segunda ed
CRC Press: New York. 451 p.

Chang T.S., Siddiq M., Sinha N.K., Cash J.N. (1994). Plum juice quality affected by enzyme treatment and fining. Journal of Food
Science, 59: 1065-1069.
Chaparro D.F., Rosas D.C. (2006). Aislamiento y evaluación de la actividad enzimática de hongos descomponedores de madera en la
reserva natural La montaña del ocaso, Quimbaya-Quindío. Microbiología Industrial, Javeriana: Bogotá. 109 p. p.
Chaparro D.F., Rosas D.C., Varela A. (2009). Aislamiento y evaluación de la actividad enzimática de hongos descomponedores de
madera (Quindío, Colombia) Revista Iberoamericana de Micologia, 26(4): 238-243.
Chen H., Xu F., Li Z. (2002). Recent developments in biodegradation of industrial pollutants by white rot fungi and their enzyme
system. Bioresource Technology 81: 261.
Chen Y.J., Shiao M.S., Lee S.S., Wang S.Y. (1997). Effects of Cordyceps sinensis on the proliferation and differentiation of human
leukemic U937 cells. Life Science, 60(23): 49-59.
Chihara G., Maeda Y., Humuro J., Sasaki T., Fukuoka F. (1969). Inhibition of mouse sarcoma 180 by polysaccharides from Lentinus
edodes Nature, 222: 687-688.
Chihara G., Hamuro J., Maeda Y., Arai Y., Fukuoka F. (1970). Fractionation and Purification of the Polysaccharides with Marked
Antitumor Activity, Especially Lentinan, from Lentinus edodes (Berk.) Sing, (an Edible Mushroom). Cáncer Research, 30:
2776-2781.
Chitiva - Jaramillo A., Torrenegra – Guerrero R., Cabrera - Parada C., Díaz - Puentes N., Pineda - Parra V. (2009). Contribución al
estudio de microhongos filamentosos en los ecosistemas páramo de guasca y el tablazo.[1-10]. Disponible en:
http://www.javeriana.edu.co/Facultades/Ciencias/gifuj/hongos_ ecosistemas_ paramo.pdf. [Visitada en
marzo de 2010].
Chu K.H., Kim E.Y. (2006). Predictive modeling of competitive biosorption equilibrium data. Biotehchnology Bioprocess
Engineerong, 11: 67-71.
Comtat J., Joseleau J. (1981). Mode of action of a xylanase and its significance for the structural investigation of the branched L-
arabino-Dglucurono-D-xylan from redwood (Sequoia sempervirens). Carbohydrates Resources, 95: 101-112.
Corrales A., López C.A. (2005). Macrocybe titans (Bigelow y Kimbr.) Pegler, Lodge y Nakasone, un registro nuevo para Colombia.
Actual Biología, 27(82): 93-97
Corredor I.C., Caridad M., Restrepo S. (2007). Evaluación de la suceptibilidad de hongos endófitos aislados de rosa (rosa hybrida) a
insecticidas comerciales. Revista Colombiana de Biotecnología, 9(1): 59-74.
Cortés M.R., García A.S., Suarez H.M. (2007). Fortificación de hongos comestibles (Pleurotus ostreatus) con calcio, selenio y
vitamina C. VITAE, Revista de la Facultad de Química Farmacéutica, 14(1): 16-24.
Coughlan M.P., Ljungdahl L.G. (1988). Comparative biochemistry of fungal and bacterial cellulolytic enzyme systems. En:
Biochemistry and Genetics of Cellulose Degradation Millet J.P.A.P.B.J. (Ed.). Academic Press: London. pp. 34 p.
Coughlan M.P., Hazlewood G.P. (1993). β-1,4-D-xylan-degrading enzyme systems: biochemistry, molecular biology and
applications. Biotechnology and Applied Biochemistry, 17: 259-289.
Cui F., Chisti Y. (2003). Polysaccharopeptides of Coriolus versicolor: physiological activity, uses and production. Biotechnology and
Advances, 21: 109-122.
Cui S. (2005). Food Carbohydrates, Chemistry, Physical propierties and Applications. CRC Press, ed. Group T.F. Taylor & Francis
Group: Boca Raton Florida. 404 p.
D’Annibale A., Celleti D., Felici M., Di Mattia E., Giovannozzi-Sermani G. (1996). Substrate specificity of laccase from Lentinus
edodes. Acta
Biotechnol. Bioengineering, 16: 257-270.
De Carvalho J.C., Pandey A., Oishi B.O., Brand D., Rodriguez-León J.A., Soccol C.R. (2005). Relation between growth,
respirometric anaysis and biopigments production from Monascus by solid-state fermentation. Biochemical Engineering
Journal, 29: 262-269.
Dekker R.F.H., Richards G.N. (1975). Purification, properties and mode of action of hemicellulase I produced by Ceratocystis
paradoxa. Carbohydrates Resources 39: 97-114.
Dekker R.F.H. (1985). Biodegradation of the hemicelluloses. En: Biosynthesis and Biodegradation of wood components. Higuchi T.
(Ed.). Academic press,: Orlando. pp. 113-132.
Delgado F., López Y., Giraldo E.M. (2001). Actividad enzimática de hongos y su patogenicidad sobre Hypothenemus hampei.
Manejo Integrado de Plagas (Costa Rica), 60: 43-49.
Dhouib A., Hamza M., Zouari H., Mechichi T., H’midi R., Labat M., Martínez M.J., Sayadi S. (2005). Autochthonous fungal strains
with high ligninolytic activities from Tunisian biotopes. African Journal of Biotechnology, 4(5): 431-436.
Dobozi M.S., Szakacs G., Bruschi C.V. (1992). xylanase activity of Phanerochaete Chrysosporium. Applied and environmental
microbiology, 58(11): 3466-3471.
Dubois M., Gilles K.A., Hamilton J.K., Rebers P.A., Smith F. (1956). Colorimetric method for determination of sugars and related
substances. Analytical Chemistry Engineering Journal, 28(3): 350-356.
Eggert C., Temp U., Dean J.F., Eriksson K.L. (1996). A fungal metabolite mediates degradation of non-phenolic lignin structures and
synthetic lignin by laccase. FEBS Letters, 391: 144-148.
Ek M., Gellerstedt G., Henriksson G. (2009). Pulp and Paper Chemistry and Technology: Wood Chemistry and Wood Biotechnology.
ed. KG W.d.G.G.C. Vol. 1. THE GRUITER: Berlin. 320 p.

El Gogary S., Leite O., Criverallo D.E., Eveleigh D.E., El-Dorry H. (1989). Mechanism by which cellulose triggers
cellobiohydrolase I gene expression in Trichoderma reesei. Proceedings of the National academy of Sciences of the United
States of America, 86: 6138-6141.
Elibol M. (2004). Optimization of medium composition for actinorhodin production by Streptomyces coelicolor A3 (2) with response
surface methodology. Process Biochemistry, 39: 1057-1062.
Elisashvili V., Parlar H., Kachlishvili E., Chichua D., Kvesitadze G. (2001a). Ligninolytic activity of basidiomycetes grown under
submerged and solid-state fermentation on plant raw material (sawdust of grapevine cuttings). Advances Food Science, 23: 117-
123.
Elisashvili V., Parlar H., Kachlishvili E., Chihua D., Bakrazde M., Kokhreidze N., Kvesitadze G. (2001b). Potential of solid-state
fermentation for laccase production. Advance Food Science, 23: 117-123.
Elisashvili V., Penninckx M., Kachlishvili E., Tsiklauri N., Metreveli E., Kharziani T., Kvesitadze G. (2008). Lentinus edodes and
Pleurotus species lignocellulolytic enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes of
different composition. Bioresource Technology, 99: 457-462.
Enshasy H.E. (2010). Immunomodulators. En: The Mycota. Hofrichter M. (Ed.). Springer-Verlag: Berlin, 477 p. pp. 477.
Evans C., Dutton. M., Guillen. F., Veness R. (1994). Enzymes and small molecular mass agents involved with lignocellulose
degradation. FEMS Microbiology Letters, 13: 235-240.
Fang Q.H., Zhong J.J. (2002). Effect of initial pH on production of ganoderic acid and polysaccharide by submerged fermentation of
Ganoderma lucidum. Process Biochemistry, 37(7): 69-74.
Fermor T.R., Wood D.A. (1981). Degardation of bacteria by Agaricus bisporus and other fungi. Journal of General Microbiology,
126: 377-387.
Fernández J.A., Henao L.M., Pedroza-Rodríguez A.M., Quevedo-Hidalgo B. (2009). Inmovilización de hongos ligninolíticos para la
remoción del colorante negro reactivo 5. Revista Colombiana de Biotecnología, 11(1): 59-72.
Ferraz A., Cordova A.M., Machuca A. (2003). Wood biodegradation and enzyme production by Ceriporiopsis subvermispora during
solidstate fermentation of Eucalyptus grandis. Enzyme and Microbiology Technology, 32: 59-65.
Forchiassin F., Levin L. (en imprenta). Enzimas Extracelulares Degradación de Polímeros Vegetales. En: Introducción al reino de
los hongos y los grupos afines. Albertó E., Vadell E. (Eds.). Literature for Latin America: Buenos Aires.
Fujian X., Hongzhang C., Zuohu L. (2001). Solid-state production of lignin peroxidase (LiP) and manganese peroxidase (MnP) by
Phanerochaete chrysosporium using steam-exploded straw as substrate. Bioresource Technology, 80: 149-151.
Gadd G.M. (1999). Fungal production of citric and oxalic acid: importance in metal speciation, physiology and biogeochemical
processes. Advance in Microbiology and Physiology, 41: 47-92.
Gadd G.M. (2001). Fungi in Bioremediation. Cambridge University Press: Cambridge. 481 p.
Galliano H., Gas G., Seris J.L., Boudet A.M. (1991). Lignin degradation by Rigidoporus lignosus involves synergistic action of two
oxidizing enzymes: manganese peroxidase and laccase. Enzyme and Microbial Technology, 13: 478-482.
Gao Y., Zhou S. (2003). Cancer prevention and treatment by Ganoderma, a mushroom with medicinal properties. Food Research
International, 19: 275-325.
Garcés-Molina A.M., Velez-Cardona N., Ruiz-Alzate S., Serna-D´León J.G., Suarez-Holguín E. (2006). Revista Lasallista de
Investigación, 2(2): 15-20.
García A.M., Torres R.G. (2003). Producción de enzimas ligninolíticas por Basidiomycetes mediante la técnica de fermentación en
fase sólida. Revista Colombiana de Biotecnología, 5(001): 56-64.
Glen J.K., Gold M.H. (1985). Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-
degrading basidiomycetes Phanerochaete chrysosporium. Archives of Biochemistry and Biophysics, 242: 329-341.
Gold M.H., Alic M. (1993). Molecular Biology of the lignin-degrading Basidiomycete Phanerochaete Chrysosporium.
Microbiological Reviews, 57(3): 605-622.
Golovleva L.A., Malt’seva O.V., Leont’evskii A.A., Myasoedova N.M., Skryabin G.K. (1987). Ligninase biosynthesis by the fungus
Panus tigrinus during solid-state fermentation of straw. Doklady Akademii Nauk, 294: 992-995.
Gómez-Dorado C., Martinez-Salgado M., Nieto-Mosquera D., Pedrosa-Rodriguez A., Rodriguez-Vásquez R., Rosas-Acosta J.
(2005). Estudio del efecto de dos inductores y un protector enzimático sobre la actividad de las enzimas MnP y lacasa
producidas por Trametes versicolor y su acción en la decoloración. Revista de la Facultad de Ciencias Universidad Javeriana,
10(2): 37-45.
Gong P., Guan X., Witter E. (2001). A rapid method to extract ergosterol from soil by physical disruption. Applied Soil Ecology, 17:
285-289.
Góngora C.E. (2009). Avances en conocimiento y mejoramiento del hongo entomopatógenos Beauveria bassiana para el control de
la broca del café, Hypothenemus hampei. Cenicafé. Disponible en:
http://almacafe.com.co/modules/News/images/simposio2.pdf. [Visitada en marzo de 2010].
Goodenough P.W. (1996). Structural studies on cellulases, pectinases an xylanases. En: Enzymes for Carbohydrate Engineering.
Elsevier Science B.V,. pp. 83-107.
Gow N.A.R. (1995a). The Growing Fungus. ed. Hall C. N.A.R Gow & G. M. Gadd: London. 402 p. p.
Gow N.A.R. (1995b). The growing Fungus. ed. Hall C. N.A.R Gow & G. M. Gadd: London.
Granada R.D.M., Mejía G.A.I., Jimenez T.G.A. (2005). Utilización de residuos de plátano para la producción de metabolitos
secundarios por fermentación en estado sólido con el hongo Lentinus crinitus. Revista de la Facultad de Química Farmacéutica,
12(2): 13-20.

Guiseppin M.L.F. (1984). Effects of disolved oxygen concentration on lipase production by Rhizopus delemar. Applied Microbiology
and Biotechnology, 20: 161-173.
Gutiérrez Ramírez L., Pérez Bran J., Uribe M. (2012). Evaluación in vitro de celulasas producidas por cepas nativas de Trichoderma
reesei, Cladosporium herbarum y Aspergillus niger. Journal Agricultural and Animal Science, 1(1): 7-15.
Guzmán M.L., Angel J.C., Rivas A., Suarez L. (2003). Producción de hongos comestibles en residuos de caña de azúcar, Memorias
VI congreso Colombiano de la Asociación de Técnicos de la Caña de Azúcar: Colombia. 323-331.
Hamelinck C.N., van Hooijdonk G., Faaij A.P.C. (2005). Ethanol from lignocellulosic biomass: techno-economic performance in
short-, middle- and long-term. Biomass Bioenergy, 28: 384-410.
Hamlyn P.F., Schmidt R.J. (1994). Potential therapeutic application of fungal filaments in wound management. Mycologist, 8: 147-
152.
Hammel K.E., Kapich A.N., Jensen Jr. K.A., Ryan Z.C. ( 2002). Reactive oxygen species as agents of wood decay by fungi. Enzyme
Microbiology and Technology, 30: 445-453.
Henry W.P. ( 2003). Non-enzymatic iron, manganese and copper chemistry of potencial importance in wood decay. En: Wood
Deterioration and Preservation - Advances in our Changing World. Society A.C. (Ed.). Goodell, B., Nicholas, D.D., Schultz,
T.P. (Eds.): Washington. pp. 175-195.
Hobbs C. (2000). Medicinal values of Lentinus edodes (Berk.) Sing. (Agaricomyvetideae). A literature review. Interantional Journal
of Medicinal Mushrooms, 2: 287-297.
Hölker U., Höfer M., Lenz J. (2004). Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Applied
Microbiology Biotechnology, 64: 175-186.
Hölker U., Lenz J. (2005). Solid-state fermentation - are there any biotechnological advantages? Current Option in Microbiology, 8:
301-306.
Hormiga J.A., Verab J., Frías I., Torres Dariasa N.V. (2008). Growth and ligninolytic system production dynamics of the
Phanerochaete chrysosporium fungus A modelling and optimization approach. Journal of Biotechnology, 137: 50-58.
Hsieh C., Liu C.-J., Tseng M.-H., Lo C.-T., Yang Y.-C. (2006). Effect of olive oil on the production of mycelial biomass and
polysaccharides of Grifola frondosa under high oxygen concentration aeration. Enzyme and Microbial Technology, 39: 434-439.
Hsieh C., Wang H.-L., Chena C.-C., Hsub T.-H., Tseng M.-H. (2008). Effect of plant oil and surfactant on the production of mycelial
biomass and polysaccharides in submerged culture of Grifola frondosa. Biochemical Engineering Journal, 38: 198-205.
Hudson H. (1986). Fungal Biology. CPC press Edward Arnolds Eds: Maryland. 298 p. p.
Hyun H.H., Zeikus J.G. (1985). General Biochemical characterization of thermostable extracellular B-amylasa from Clostridium
thermosulfurogenes. Applied Environmental Microbiology, 49: 1162-1170.
Ikasari L., Mitchell D.A. (2000). Two-phase model of yhe kinetics of growth of Rhizopus oligosporus in membrana culture.
Biotechnology Bioengineering, 68: 619-627.
Ilmen M., Saloheimo A., Onnela M.L., Pentilla M. (1996). Rgulation of cellulase gen expression in the filamentous fungus
Trichoderma reesei. Applied Environmental Microbiology, 63: 1298-1306.
Ishikawa K., Majima T., Ebina T. (1992). Anti-tumour effect of a Coriolus preparation, PSK: Induction of macrophage chemotactic
factor (MCF) in spleens of tumour bearing mice. Biotherapy, 5: 251-258.
Ivshin V., Artamonova S., Ivshina T., Sharnina F. (2007). Methods for Isolation of Chitin–Glucan Complexes from Higher Fungi
Native Biomass. Polymer Science, Serie B., 49(11-12): 305-310.
Izawa M., Takashio M., Koshino S. (1995). Several new factors influencing the measurement of β-glucan content using calcofluor
flow-injection analysis method. Journal of the Institute of Brewing, 101: 371.
Jaszek M., Malarczyk E., Leonowicz A. (1998). Investigation of ligninolytic enzymes during solid state fermentation of cotton wastes
by selected strains of basidiomycetes. En: Proceedings of the 7th International Conference on Biotechnology in the Pulp and
Paper Industry. Vancouver. pp. 16-18.
Jennings D.H. (1995). The Physiology of Fungal Nutrition, Cambridge, UK: Cambridge University press.
Johnson E.A., Sakajoh M., Halliwell G., Madia A., Demain A.L. (1982). Saccharification of complex cellulosic substrates by the
cellulase system from C. thermocellum. Applied and environmental microbiology, 43: 1125-1132.
Jong S.C., Birgmingham J.M. (1992). Medicinal benefits of the mushroom Ganoderma. Adv. Appl. Microbiol., 37: 101-134.
Joselau J.P., Ruel K. (1994). Wood polysaccharides and their degradation by fungi. En: Host Wall Alterations by Parasitic Fungi.
Ouellette P. (Ed.). APS Press: Minnesota. pp. 334-387.
Joshi V.K., Pandey A. (1999a). Biotechnology. Food fermentation Vol II. Educational Publishers & distributors: New Delhi. 998 p p.
Joshi V.K., Pandey A. (1999b). Biotechnology: Food Fermentation (Microbiology, Biochemistry and Technology) Vol I. Educational
Publishers & distributiors: New Delhi. 521 P. p.
Kalisz H.M., Wood D.A., Moore D. (1987). Production, regulation and release of extracellular proteinasa activity in basidiomycetes
fungi. Transactions of the British Mycological Society, 88: 221-227.
Kalisz H.M. (1988). Microbial Proteinases. Advances in Biochemical Engineering/Biotechnology, 36: 1-65.
Kalisz H.M., Wood D.A., Moore D. (1989). Some Characteristics of extracellular proteinases from Coprinus cinereus. Mycological
Research, 92: 278-285.
Karunanithy C., Muthukumarappan K., Julson J.L. (2008). Influence of high shear bioreactor parameters on carbohydrate release
from different biomasses American Society of Agricultural and Biological Engineers Annual International Meeting ASABE
084114. ASABE.
Kersten P., Cullen D. (2007). Extracellular oxidative systems of the lignin-degrading Basidiomycete Phanerochaete chrysosporium.
Fungal Genetics and Biology, 44: 77-87.

Kiho T., Ookubo K., Usui S., Ukai S., Hirano K. (1999). Structural features and hypoglycemic activity of a polysaccharides (CS-
F10) from the cultured mycelium of Cordyceps sinensis. Biological and Pharmaceutical Bulletin, 22(9): 66-70.
Kim F., Sakagami H., Tanuma S., Konno K. (1990). Stimulation of interferon-g-induced human myelogenous leukemic cell
differentiation by high molecular weight PSK subfraction. Anticancer Research, 10: 55-58.
Kim S., Holtzapple M.T. (2006). Delignification kinetics of corn stover in lime pretreatment. Bioresource Technology, 97: 778-785.
Kim T.H., Taylor F., Hicks K.B. (2008). Bioethanol production from barley hull using SAA (soaking in aqueous ammonia)
pretreatment. . Bioresouce Technology, 99: 5694-5702.
Kirk T.K., Connors W., Zeikus J. (1976). Requeriment for a growth substrate during lignin decomposition by two wood-roting fungi.
Applied Environmental Microbiology, 32: 192-194.
Kirk T.K., Farrel R.L. (1987). Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology 41: 465-
505.
Kobayashi H., Matsunaga K., Oguchi Y. (1995). Antimetastatic Effects of P5K (Krestin), a Protein-bound Polysaccharide Obtained
from Basidiomycetes: An Overview. Cancer Epidemiology, Biomarkers & Prevention, 4: 275-281.
Koduri R.S., Tien M. (1994). Kinetic analysis of lignin peroxidase-explanation for the mediation phenomen by veratryl alcohol.
Biochemistry, 33: 4225-4230.
Koh J.H., Kim J.M., Chang U.J., Suh H.J. (2003). Hypocholesterolemic effect of hotwater extract from mycelia of Cordyceps
sinensis. Biological and Pharmaceutical Bulletin, 26: 84-87.
Komatsu M., Okubo S., Kikumoto S., Kimura K., Saito G., Sakai S. (1969). Host-mediated antitumor action of Schizophyllan a
glucan produced by Schizophyllum commune. Gann, 60: 137-144.
Komura D.L., Ruthers A.C., Carbonero E.R., Alquini G., Rosa M.C., Sassaki G.L., Lacomini M. (2010). The origin of mannans
found in submerged culture of basidiomycetes. Carbohydrate polymers, 79: 1052-1056.
Kootstra A.M.J., Beeftink H.H., Scott E.L., Sanders J.P.M. (2009). Comparison of dilute mineral and organic acid pretreatment for
enzymatic hydrolysis of wheat straw. Biochemical Engineering Journal, 46: 126-131.
Kosaka A., Yamashita A. (1993). Lentinan as an effective inmmunomodulator against breast cancer, with special reference to clinical
applications. International Journal Immunotherapy, 9: 111-116.
Koutinas A.A., Wang R., Webb C. (2004). Restructuring Upstream Bioprocessing: Technological and Economical Aspects for
Production of a Generic Microbial Feedstock From Wheat. Published online in Wiley InterScience: 1-15.
Kumar R., Wyman C.E. (2009). Effects of cellulase and xylanase enzymes on the deconstruction of solids from pretreatment of
poplar by leading technologies. Biotechnology Progress, 25: 302-314.
Kumari J., Sahoo P.K. (2006). Non-specific immune response of healthy and immunocompromised Asia catfish (Clarias batrachus)
to several immunostimulants. Aquaculture, 255: 133-141.
Ladner W., Ditrich K. (1999). Biocatalytic production of chiral intermediates. Chimica Oggi 7/8: 51-54.
Lang E., Nerud F., Novotna E., Martens R. (1996). Production of ligninolytic exoenzymes and 14C-pyrene mineralization by
Pleurotus sp. in lignocellulose substrate. Folia Microbiology, 41: 489-493.
Lemaire M. (1996). The Cellulosome - an exocellular multiprotein complex specialised in cellulose degradation. Critical Reviews
and Biochemistry & Molecular Biology, 31: 201-236.
Lenz J., Höfer M., Krasenbrink J.B., Hölker U. ( 2004). Computational and physical methods in solid-state fermentation. Applied
Microbiology Biotechnology, 65: 9-17.
Leonowicz A., Matuszewska A., Luterek J., Ziegenhagen D., Wojtas-wasilewska M., Cho N.S., otros. (1999). Biodegradation of
lignin by white-rot fungi. Fungal Genetics and Biology 27: 175-185.
Leschine S.B. (1995). Cellulose degradation in anerobic environments. Annual Review Microbiology, 49: 399-410.
Leung M., Fung K., Choy Y. (1997). The isolation and characterization of an immunomodulatory and antitumor polysaccharide
preparation from Flammulina velutipes. Immunopharmacol 35: 255-263.
Leung M., Liu C., Koon J., Fung K. (2006). Polysaccharide biological response modifiers. Immunologycal Letters, 105: 101-114.
Levin L. (1998). Biodegradación de materiales lignocelulósicos por Trametes trogii (Aphyllophorales, Basidiomycetes).Tesis
Doctoral. Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires: Buenos Aires.
Levin L., Forchiassin F., Ramos A.M. (2002). Copper induction of lignin-modifying enzymes in the white-rot fungus Trametes
trogii. Mycologia, 94(3): 377-383.
Levin L., Papinutti L., Forchiassin F. (2004). Evaluation of Argentinean white rot fungi for their ability to produce lignin-modifying
enzymes and decolorize industrial dyes. Bioresource Technology, 94: 169-176.
Levin L., Herrmann C., Papinutti V.L. (2008). Optimization of lignocellulolytic enzyme production by the white-rot fungus Trametes
trogii in solid-state fermentation using response surface methodology. Biochemical Engineering Journal 39: 207-214.
Li Q., He Y.C., Xian M., Jun G., Xu X., Yang J.M., Li L.Z. (2009). Improving enzymatic hydrolysis of wheat straw using ionic
liquid 1-ethyl-3-methyl imidazolium diethyl phosphate pretreatment. Bioresource Technology, 100: 3570-3575.
Li X., Liun Y., Ji L. (2000). Production of lignocellulolytic enzymes in solid state fermentation using corn stalks as substrate.
Henong Xuebao, 14: 99-103.
Lia K., Azadic P., Collinsb R., Toland J., Kimc J.S., E. K.E.L. (2000). Relationships between activities of xylanases and xylan
structures. Enzyme and Microbial Technology, 27: 89-94.
Lindblom C.M., Wachenfeldt E.V., Tranvik L.J. (2004). Ergosterol as a measure of living fungal biomass: persistence in
environmental samples after fungal death. Journal of Microbiological Methods, 59 253- 262.
Lindequist U., Niedermeyer T., Ju¨lich W.-D. (2005). The pharmaceutical potential of mushrooms. Electronic centralised aircraf
monitor (eCAM), 2: 285-299.

Liu C., Liu Y., Liao W. (2003). Application of statistically based experimental designs for the optimization of nisin production from
whey. Biotechnology Letters, 25: 877-882.
Liu F., Fung M.C., Ooi V.E.C., Chang S.T. (1996). Induction in the mouse of gene expression of immunomodulating cytokines by
mushroom polysaccharide-protein complexes. Life Science, 58(1): 795-803.
Lockington R.A., Rodbourn L., Barnett S., Carter C.J., Kelly J.M. (2002). Regulation by carbon and nitrogen sources of family of
cellulases in Aspergillus nidulans. Fungal Genetics and Biology, 37: 190-196.
Lonsane B.K., Ghildyal N.P., Budiatman S., Ramakrishna S.V. (2005). Solid state fermentation for production of α-amylase by
Bacillus megaterium 16M Enzyme Microbiology Technology, 7: 258-265.
Lozano J.C. (1989). Producción comercial del Champiñon /Pleurotus ostreatus/ en pulpa de café, CONGRESO ASCOLFI, X. Cali
(Colombia): Manizales. 60 p.
Luedeking R., Piret E. (1959). Transient and steady states in continuous fermentation. Theory and experiment. Journal of
Biochemical and Microbiological Technology and Engineering, 1: 431-459.
Lull C., Wichers H., Savelkoul H. (2005). Antiinflammatory and immunomodulating properties of fungal metabolites. Mediator
Inflammation, 2: 63-80.
Lynd L.R., Weimer P.J., Zyl W.H.v., Pretorius I.S. (2002). Microbial cellulose utilization: Fundamentals and Biotechnology.
Microbiological and Molecular Biology Reviews, 66(3): 506-577.
Maganhotto de Souza-Silva C.M., Soares de Melo I., de Oliveira P.R. (2005). Ligninolytic enzyme production by Ganoderma spp.
Enzyme and Microbial Technology 37: 324-329.
Mandels M., Reese E. (1956). Induction of cellulase in Trichoderma viride as influenced by carbon sources and metals. Biology
Branch: 269-277.
Mandels M., Weber J. (1969). The production of cellulases and their applications. Advances Chemistry, 95: 391-414.
Mansel P.W.A. (1994). Polysaccharides in skin care. Cosmetics Toilet, 109: 7-72.
Mansfield S.D., Mooney C., Sanddler J.N. (1999). Substrate and enzyme characteristics that limit cellulose hydrolysis.
Biotechnological progress, 15: 804-816.
Markham P. (1998). Occlusions of septal pores in filamentous fungi. Mycological Research, 98: 1089-1106.
Marques de Souza C.G., Zilly A., Peralta R.M. (2002). Production of laccase as the sole phenoloxidase by a Brazilian strain of
Plerotus pulmonarius in solid state fermentation. Journal Basic Microbiology, 42: 83-90.
Martinez A. (2002). Molecular Biology and structure-function of lignin-degradin heme peroxidases. Enzyme Microbiology
Technology, 30: 425-444.
Martinez M., Ruiz-Dueñas F., Guillen F., Martinez A. (1996). Purifacation and catalytic properties of two manganese peroxidase
isoenzymes from Pleurotus eryngii. European Journal Biochemistry, 237: 424-432.
Martínez M.J., Ruíz-Dueñas F.J., Guillén F., Martínez A.T. (1996). Purification and catalytic properties of two manganese-
peroxidase isoenzymes from Pleurotus eryngii. European Journal of Biochemistry, 237: 424-432.
Mattila P., Lampi A.M., Ronkainen R., Toivo J., Piironen V. (2002). Sterol and Vitamin D2 contents in some wild and cultivated
mushroomn. Food Chemistry, 76: 293-298.
Mayer A.M., Staples R.C. (2002). Laccase: new functions for an old enzyme. Phytochemistry, 60: 551-565.
Maziero R., Cavazzoni V., Ramos-Bononi V.L. (1999). Screening of Basidiomycetes for the production of exopolisaccharides and
biomass in submerged culture. Revista de Microbiologia, 30: 77-84.
Meagher M.M., Tao B.Y., Chow J.M., Reilly P.J. (1988). Kinetics and subsite mapping of β -D-xylobiose and D-xylose producing
Aspergillus niger endo-β-1,4-D-xylanase. Carbohydrates Resources, 173: 273-283.
Meier H. (1955). Decomposition of the cell wall by wood-destroying fungi and the submicroscopic structure of tracheids of spruce
and wood fibers of birch (translation Nº 92: Department of the Secretary of State of Canada, February 1956) Holz. Roh-Werkst.,
13: 323-338.
Mester T., Sierra-Alvarez R., Holzforschung. (1998). Peroxidase and aryl metabolite production by the white rot fungus Bjerkandera
sp. strain BOS55 during solid state fermentation of lignocellulosic substrates. J. Field, 52: 351-358.
Meyer H.P., Kiener A., Imwinkelried R., Shaww N. (1997). Biotransformations for fine chemicals production. Chimia, 51: 287-289.
Mitchell D., Stuart D., Tanner R. (1999). Solid-State fermentation - Microbial growth kinetics, in The Encyclopedia of Bioprocess
Technology: Fermentation, Biocatalysis and bioseparation, Flickinger M., Drew S., EditorsWiley: New York. 2407-2429
Mitchell D.A., Von meien O.F., Krieger N., Dalsenter F.D. (2004). A review of recent developments in modeling of microbial
growth kinetics and intraparticle phenomena in solid-state fermentation. Biochemical Engineering Journal 17: 15-26.
Mizuno T. (1992). Antitumor active polysaccharides isolated from the fruiting body of Hericium erinaceum, and edible, and
medicinal mushroom called Yamabushitake or Houtou. Bioscience Biotechnology and Biochemistry, 56: 349-357.
Mizuno T., Saito H., Nishitoba T., Kawagishi H. (1995). Antitumor-active substances from mushrooms. Food review international,
11: 23-61.
Mizuno T. (1999). The extraction and development of antitumouractive polysaccharides from medicinal mushrooms in Japan.
International Journal Medicine Mushrooms, 1: 9-29.
Mohammad-Fata M., Hossein M., Shirin G., Ghorban-Ali H. (2007). Immunomodulating and anticancer agents in the realm of
macromycetes fungi (macrofungi). International Immunopharmacology 7: 701-724.
Moncada B., Forero E. (2006). El género Pseudocyphellaria VAIN. (Lobariaceae - ascomycetes liquenizados) en Colombia.
Caldasia, 28(2): 197-215.
Montenecourt B.S., Eveleigh D.E. (1977). Semiquantitative plate assay for determination of cellulase production by Trichoderma
viride. Applied and Environmental Microbiology, 33(1): 178-183.

Montes B., Restrepo A., McEwen J.G. (2003). Nuevos aspectos sobre la clasificación de los hongos y su posible aplicación médica.
Biomédica, 23: 213-237.
Montgomery H.J., Monrealb C.M., Young J.C., Seifert K.A. (2000 ). Determinination of soil fungal biomass from soil ergosterol
analyses. Soil Biology & Biochemistry, 32: 1207±1217.
Montoya B.S., Varon L.M., Levin L. (2008). Effect of culture parameter on the production of the edible mushroom Grifola frondosa
(maitake) in tropical weathers. World J. Microbiol. Biotechnol., 24: 1361-1366.
Montoya B.S., Orrego C.A., Levin L. (2011a). Growth, fruiting and lignocellulolytic enzyme production by the edible mushroom
Grifola frondosa (maitake). World Journal of Microbiology and Biotechnology, ISSN 0959-3993: DOI 10.1007/s11274-11011-
10957-11272.
Montoya S., Restrepo G.M. (2006). Evaluación de la síntesis de proteína a partir del crecimiento vegetativo de Pleurotus sp. sobre
residuos de algarrobo y uva pasa. Revista Universidad de Caldas, 26(1-2): 23-32.
Montoya S. (2008). Actividad enzimática, degradación de residuos sólidos orgánicos y generación de biomasa útil por el
macromiceto Grifola frondosa.Investigación. Ingeniería Química, Universidad Nacional de Colombia Sede Manizales:
Manizales. 95 p. p.
Montoya S., Gallego J., Sucerquia A., Pelaez B., Betancourt O., Arias D. (2010). Macromicetos observados en bosques del
Departamento de Caldas: su influencia en el equilibrio y la conservación de la biodiversidad. Boletín Científico Centro de
Museos de Historia Naural, 14(2): 57-76.
Montoya S.B., Orrego C.A., Levin L. (2011b). Modeling Grifola frondosa fungal growth during solid-state fermentation.
Engineering in Life Science, 11(316-321).
Moon S.H., Park J.M., Chun H.Y., Kim S.J. (2006). Comparisons of physical properties of bacterial cellulose produced in different
culture conditions using saccharified food wastes. Biotechnology Bioprocess Engineering, 11: 26-31.
Moore D. (1998a). Fungal Morphogenesis. Primera ed Cambrige University Press.: New York. 469 p. p.
Moore D. (1998b). Fungal Morphogenesis. Cambrige University Press: New York, United States of America.
Moreira M.T., Feijoo G., Lema J.M. (2003). Fungal bioreactors: Applications to white-rot fungi. Reviews in Environmental Science
and Bio/Technology 2: 247-259.
Mosier N., Hendrickson R., Ho N., Sedlak M., Ladisch M.R. (2005a). Optimization of pH controlled liquid hot water pretreatment of
corn stover. Bioresource Technology, 96: 1986-1993.
Mosier N., Wyman C.E., Dale B.D., Elander R.T., Lee Y.Y., Holtzapple M., Ladisch C.M. (2005b). Features of promising
technologies for pretreatment of lignocellulosic biomass. Bioresource Technology, 96: 673-686.
Mouso N., Papinutti L., Forchiassin F. (2003). Efecto combinado del cobre y pH inicial del medio de cultivo sobre la producción de
lacasa y manganeso peroxidasa por Stereum hirsutum (Willd) Pers. Revista Iberoamericana de Micología, 20: 176-178.
Mukhamedzhanova T.G., Bezborodov A.M. (1982). Effect of nitrogen and carbon sources on lipase accumulation by the fungus
Rhizopus oryzae 1414. Mikrobiologia, 18: 16-22.
Muller dos Santos M., Souza das Rosa A., Dal’Boit, Mitchell D., Krieger N. (2004). Thermal denaturation: is solid-state fermentation
really a good technology for the production of enzymes? Bioresource Technology 93: 261-268.
Munoz G.A., Agosin E., Cotoras M., San Martin R., Volpe D. (1995). Comparison of aerial and submerged spore properties for
Trichoderma harzianum. FEMS Microbiology Letter, 125: 63-70.
Naeem S. (2002). Autotrophic-Hecterotrophic Interactions and their Impacts on Biodiversity and Ecosystem Functioning. En:
Functional Consequences of Biodiversity. Princeton University Press,: New Jersy. pp. 96-119.
Nakamura K., Yamaguchi Y., Kagota S., Shinozuka K., Kunitomo M. (1999). Activation of in vivo Kupffer cell function by oral
administration of Cordyceps sinensis in rats. . Japan Journal Pharmacology, 79: 508-508.
Nazareno M.C., Bucsinszky A.M.M., Tournier H.A., Cabello M.N., Arambarri A.M. (2000). Extracellular ABTS-oxidizing activity
of autochthonous fungal strains from Argentina in solid medium. Revista Iberoamericana de Micología, 17: 64-68.
Nelson N.J. (1944). A photometric adaptation of the Somogyi method for the detrmination of glucose. Journal Biochemistry, 153:
375-380.
Nidetzky B., Steiner W., Hayn M., Claeyssens M. (1994). Cellulose hydrolysis by the cellulases from Trichoderma reesei: a new
model for synergistic interaction. Bichemistry Journal, 298: 705-710.
Nie S.-P., Xie M.-Y. (2011). A review on the isolation and structure of tea polysaccharides and their bioactivities. Food
Hydrocolloids, 25: 144-149.
Nieto I., Chegwin C. (2010). Influencia del sustrato utilizado para el crecimiento de hongos comestibles sobre sus características
nutraceúticas. Revista Colombiana de Biotecnología, 12(1): 169-178.
Nieto I.J., Cucaita R.E. (2007). Ácidos grasos, ésteres y esteroles del cuerpo fructífero del hongo Laccaria laccata. Revista
Comobiana de Química, 36(3): 277-284.
Nieto I.J., Ávila I.M. (2008). Determinación de ácidos grasos y compuestos triterpenoides del cuerpo fructífero de Suillus luteus.
Revista Colombiana de Química, 37(3): 45-52.
Nieto-Ramirez I.J., Chegwin-Angarita C., Osorio-Zuluaga H.J. (2007). Incorporación de cafeína en el hongo Pleurotus sajor-caju
cultivado sobre pulpa de café. Revista Iberoamericana de Micología 24: 72-74.
Nigam P., Pandey A., Prabhu K.A. (1987). Cellulase and ligninase production by basidiomycete culture in solid-state fermentation.
Biology Wastes, 20: 1-9.
Niku-Paavola M., Raaska M., Itavaara M. (1990). Detection of white-rot fungi by non-toxic stain. Mycology and Resource, 94: 27-
31.

Nitschke J., Altenbach H.-J., Malolepszy T., Mölleken H. (2011). A new method for the quantification of chitin and chitosan in
edible mushrooms. Carbohydrate Research, 346: 1307-1310.
Ooi V., Liu F. (2000). Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Current Medicine
Chemistry, 7: 715-728.
Ooi V.E.C., Liu F. (1999). A review of a farmacological activities of mushroom polysaccharides. International Journal Medicinal
Mushrooms, 1: 195-206.
Ortiz-Moreno M. (2010). Macromicetos en Zona Rural de Villavicencio. Orinoquía, 14(2): 125-132.
Pal M., Calvo A., Terron M.C., Gonzalez A.E. (1995). Solid-state fermentation of sugarcane bagasse with Flammulina velutipes and
Trametes versicolor. World Journal Microbiology Biotechnology, 11: 541-545.
Palonen H., Thomsen A.B., Tenkanen M., Schmidt A.S., Viikari L. (2004). Evaluation of wet oxidation pretreatment for enzymatic
hydrolysis of softwood. Applied Biochemistry and Biotechnology, 117: 1-17.
Papinutti V.L., Dorio L.A., Forchiassin F. (2003). Degradación de madera de álamo por Fomes sclerodermeus: Producción de
enzimas ligninolíticas en aserrín de álamo y cedro. Revista Iberoamericana de Micología, 20: 16-20.
Park J.-P., Kim S.-W., Hwang H.-J., Cho Y.-J., Yun J.-W. (2002). Stimulatory effect of plant oils and fatty acids on the exo-
biopolymer production in Cordyceps militaris. Enzyme and Microbial Technology, 31: 250-255.
Pasquel A. (2001). Gomas: una aproximación a la Industria de alimentos. Revista Amazónica de Investigación Alimentaria, 1(1): 1-8.
Paszczczynski A., Crawford R., Huyn V. (1988). Manganese peroxidase of Phanerochaete chrysosporium purification. Methods
enzymology, 161: 264-270.
Pelaez F., Martinez M.J., Martinez A.T. (1995). Screening of 68 species of basidiomycetes for enzymes involved in lignin
degradation. Mycology Resources, 99: 37-42.
Peña-Serna C., Sierra-Cadavid A., Sáez A., Rojano B.A. (2006). Establecimiento de un medio de cultivo para una cepa nativa de un
hongo Poliporal. Revista Colombiana de Biotecnología, 8(001): 5-13.
Peña-Venegas C.P., Cardona G.I., Arguelles J.H., Arcos A.L. (2007). Micorrizas Arbusculares del Sur de la Amazonia Colombiana y
su Relación con Algunos Factores Fisicoquímicos y Biológicos del Suelo. Acta Amazónica, 37(3): 327-336.
Pérez J., Jeffries T.W. (1992). Roles of Manganese and organic acid chelators in regulating lignin degradation and biosynthesis of
peroxidases by Phanerochaete chrysosporium. App Environl Microbiol, 58: 2402-2409.
Plackett R.L., Burman J.P. (1946). The design of optimum multifactorial experiments. Biometrika, 33: 305-325.
Plassard C., Mousain D., Salsac L. (1982). Estimation of mycelial growth of basidiomycetes by means of chitin determination.
Phytochemistry, 21: 345-348.
Pointing S.B. (1999). Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal
Diversity, 2: 17-33.
Pointing S.B. (2001). Feasibility of bioremediation by white-rot fungi. Applied Microbiology Biotechnology, 57: 20-33.
Pradeep V., Datta M. (2002). Production of ligninolytic enzymes for decolorization by cocultivation of white-rot fungi Pleurotus
ostreatus and Phanerochaete chrysosporium under solid-state fermentation. Applied Biochemistry Biotechnology, 102-103: 109-
118.
Prosser J.I., Tough A.J. (1991). Growth mechanism and growth kinetics of filamentous microorganisms. Critical Reviews in
Biotechnology, 10: 253-274.
Quintero J.C., Moreira M.T., Feijoo G., Lema J.M. (2008). Screening of white rot fungal species for their capacity to degrade lindane
and other isomers of hexachlorocyclohexane (HCH). Ciencia e Investigación Agraria, 35(2): 159-167.
Quintero J.D., Feijoo G.C., Lema J.M. (2006). Producción de enzimas ligninolíticas con hongos basidiomicetos cultivados sobre
materiales lignocelulósicos. Revista de la Facultad de química Farmacéutica, 13(2): 61-67.
Raghavarao K.S., Ranganathan T.V., Karanth N.G. (2003). Some engineering aspects of solid-state fermentation. Biochemical
Engineering Journal, 13: 127-135.
Reddy C.A. (1995). The potential for white-rot fungi in the treatment of pollutants. Current Opinion in Biotechnology, 6: 320-328.
Reddy G.V., Ravindra B.P., Komaraiah P., Roy R.M., Kothari I.L. (2003). Utilization of banana waste for the production of
lignolytic and cellulolytic enzymes by solid substrate fermentation using two Pleurotus species (P. ostreatus and P. sajor-caju).
Process Biochemistry, 38: 1457-1462.
Reid I.D. (1995). Biodegradation of lignin. Canadian Journal of Botany, 73: S1011-S1018.
Rivera A., Laverde C.M., Ríos-Motta J., Nieto I.J., Osorio H.J. (2005a). Dehidroergosterol: un artefacto generado durante el proceso
de extracción de esteroles en el hongo Pleurotus sajor-caju. Revista Colombiana de Química, 34(2): 117-125.
Rivera A., Osorio H.J., Ríos-Motta J., Nieto I.J., Laverde C.M. (2005b). Dehidroergosterol: un artefacto generado durante el proceso
de extracción de esteroles en el hongo Pleurotus sajor-caju. Revista Colombiana de Química, 34(2): 117-125.
Rocha O., Nobre C., Dominguez A., Torres A., Faria N., Rodrigues L., Teixeira J.A., Ferreira E.C., Rocha I. (2009). A Dynamical
Model for the rmentative production of fructooligosaccharides, in 10th International Symposium on Process Systems
Engineering - PSE2009
, Rita Maria de Brito Alves C.A.O.d.N.a.E.C.B.J.E., Editor© 2009 Elsevier B.V. All rights reserved: Braga, Portugal. 1827-1833.
Rodríguez I., Piñeros Y. (2007). Producción de complejos enzimáticos celulolíticos mediante el cultivo en fase sólida de
Trichoderma sp. sobre los racimos vacíos de palma de aceite como sustrato. Revista de la Facultad de química farmacéutica,
14(2): 35-42.

Rodríguez J., Ferraz A., Mello M.P. (2003). Role of metals in wood biodegradation. En: Wood Deterioration and Preservation -
Advances in our Changing World. Series A.S. (Ed.). Goodell, B., Nicholas, D.D., Schultz, T.P. (Eds.): Washington. pp. 154-
174.
Rodríguez N. (2003). Aprovechamiento de los residuos sólidos generados en el cultivo e industrialización del café para la
producción de hongos comestibles y medicinales.Postgrado. Universidad Politécnica de Valencia: Valencia. 140 p.
Rodríguez N., Jaramillo C. (2005). Cultivo de hongos comestibles del género Pleurotus sobre residuos agrícolas de la zona cafetera.
Centro Nacional de Investigaciones de Café, 23: 34-40.
Rodríguez V.N. (1993). El cultivo de hongos comestibles sobre desechos agroindustriales, Cenicafé: Chinchiná. 1-2 p.
Rodriguez-Couto S., Toca-Herrera J.L. (2007). Laccase production at reactor scale by filamentous fungi. Biotechnology advances,
25: 558.569.
Rodríguez-Couto S., Sanromán M.A. (2005). Application of solid-state fermentation to ligninolytic enzyme production. Biochemical
Engineering Journal 22 211-219.
Rodríguez-Couto S., Toca-Herrera J.L. (2007). Laccase production at reactor scale by filamentous fungi. Biotechnology Advances,
25: 558-569.
Rodríguez-Palenzuela P., García J., de Blas C., Madrid. D.B.U.P.d., Madrid. D.d.P.A.U.P.d. (2000). XIV Curso de Especialización:
Avances en nutrición y alimentación animal. En: Fibra soluble y su implicación en nutrición animal: Enzimas y probióticos.
Madrid. pp. 17p.
Ruíz A., Varela A. (2006). Nuevos registros de Aphyllophorales (basidiomicota) en bosque montano húmedo y de niebla de
Colombia. Caldasia 28(2): 259-266.
Russell R., Paterson M. (2008). Cordyceps – A traditional Chinese medicine and another fungal therapeutic biofactory.
Phytochemistry, 69: 1469-1495.
Sáez A., Florez L., Cadavid A. (2002). Caracterización de una cepa nativa de Aspergillus niger y evaluación de la producción de
ácido cítrico. Revista Universidad EAFIT, (128): 33-42.
Sánchez O.J., Cardona C.A. (2007). Producción de Alcohol Carburante: Una Alternativa para el Desarrollo Agroindustrial.
Universidad Nacional de Colombia: Manizales. 386 p.
Sánchez O.J., Cardona C.A. (2008). Trends in biotechnological production of fuel ethanol from different feedstocks. Bioresource
Technology, 99: 5270-5295.
Sangsurasak P., Nopharatana M., Mitchell D. (1996). Mathematical modeling of the growth of filamentous fungi in solid-state
fermentation. Journal Science Industrial Resources 55: 333-342.
Sarangi I., Ghosh D., Bhutia S., Mallick S., Maiti T. (2006). Anti-tumor and immunomodulating effects of Pleurotus ostreatus
mycelia-derived proteoglycans. International Immunopharmacology, 6: 1287-1297.
Schulze B., Wubbolts M.G. (1999). Biocatalysis for industrial production of fine chemicals. Chemical Biotechnology, 10: 609-615.
Shen Q., Royse D. (2002). Effects of genotipes of maitake (Grifola frondosa) on biological efficiency, quality and crop cycle time.
Applied Microbiology Biotechnology, 58: 178-182.
Shi J., Sharm-Shivappa R.R., Chinn M., Howell N. (2009). Effect of microbial pretreatment one nzymatic hydrolysis and
fermentation of cotton stalks for ethanol production. Biomass and Bioenergy, 33: 88-96.
Shi J., Sharma-Shivappa R.R., Chinn M.S. (2012). Interactions between fungal growth, substrate utilization and enzyme production
during shallow stationary cultivation of Phanerochaete chrysosporium on cotton stalks. Enzyme and Microbial Technology, 51:
1- 8.
Shinners-Carnelley T.C., Szpacenko A., Tewari J.P., Palcic M.M., Cho N.S. (2002). Enzymatic activity of Cyathus olla during solid
state fermentation of canola roots. Phytoprotection 83: 31-40.
Soares G., De Amorin M., Costa-Ferreira M. (2001). Use of laccase together with redox mediators to decolourize Remazol Brilliant
Blue R. Journal Biotechnology, 89: 123-129.
Sohail M., Siddiqi R., Ahmad A., Khan S.A. (2009). Cellulase production from Aspergillus niger MS82: effect of temperature and
pH. New Biotecchnology, 25(6): 437-441.
Somogyi M. (1945). A new reagent for the determination of sugars. Journal Biology Chemistry, 160: 61-73.
Sriyudthsak K., Shiraishi F. (2010). Investigation of the performance of fermentation processes using a mathematical model
including effects of metabolic bottleneck and toxic product on cells. Mathematical Biosciences, 228: 1-9.
Stames P. (1993). Growing gourmet and medicinal fungi Ten Speed Press and Mycomedia: Hong Kong. 456 p. p.
Stead P., Marley H., Mahmoudian M., Webb G., Noble D., Ip Y., Piga E., Rossi T., Roberts S. (1996). Efficient procedures for the
largescale preparation of (1S,2S)-trans-2-methoxycyclohexanol, a key chiral intermediate in the synthesis of tricyclic b-lactam
antibiotics. Dawson MJ: Tetrahedron - Asymmetry, 7: 2247-2250.
Sun Y., Cheng J. (2002). Hydrolysis of lignocellulosic materials for ethanol production: a review. Bioresource Technology, 83: 1-11.
Takano M., Nishida, A., Nakamura, M. (2001). Screening of wood-rotting fungi for kraft pulp bleaching by the Poly R decolorization
test and biobleaching of hardwood kraft pulp by Phanerochaete crassa WD 1694. Journal of Wood Science, 47: 63-68.
Takasaki Y. (1976). Production and utilization of B-amylase and pullulanase from B. cereus var. mycoides. Agriculture Biology
Chemistry, 40: 1515-1525.
Tang Y.J., Zhong J.J. (2002). Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma
lucidum, grown on lactose in a bioreactor. Biotechnology Letters, 24: 1023-1026.
Tao Y., Zhang L., Cheung P. (2006). Physicochemical properties and antitumor activities of watersoluble native and sulfated
hyperbranched mushroom polysaccharides. Carbohydrates Resources, 341: 2261-2269.

Tavares A., Coelho M., Coutinho J., Xavier A. (2005). Laccase improvement in submerged cultivation: induced production and
kinetic modelling. Journal of Chemical Technology and Biotechnology, 80: 669-676.
Thurston C. (1994). The structure and function of fungal laccases. Microbiology-UK, 140: 19-26.
Tišma M., Sudar M., Raĉki S., Zelic B. (2010). Mathematical model for Trametes versicolor growth in submerged cultivation.
Bioprocess and Biosystems Engineering, 33: 749-758.
Tobin J.M., White C., Gadd C.M. (1994). Metal accumulation by fungi—applications in environmental biotechnology. Journal of
Industrial and Microbiology 13: 126-130.
Tolan J., Olson D., Dines R. (1996). Survey of mill usage of xylanase. En: Enzymes for pulp and paper processing (Jeffries, TW.
Viikari L). Society A.C. (Ed.). Washington DC. pp. 25-35.
Tomme P.H., Van Tilbeurgh G., Petterson J., Van Damme J., Vanderker-Ckhove J., Knowles T., Teeri, Claeyssens M. (1988).
Studies of cellulolityc system of Trichoderma reesei QM 9414. Analysis of domain function in two cellobiohydrolases by
limited proteolysis. Europe Journal Biochemestry, 170: 575-581.
Tsukagoshi S., Hashimoto Y., Fujii G., Kobayashi H., Nomoto K., Orita K. (1984). Krestin (PSK). Cancer Treatment review, 11:
131-155.
Tuor U., Wariishi H., Schoemaker H., Gold M.H. (1992). Oxidation of phenolic arylglycerol beta-aryl ether lignin model compounds
by manganese peroxidase from Phanerochete chrysosporium: oxidative cleavage of an alpha-carbonyl mode compoundl.
Biochemistry, 31: 4986-4995.
Tychanowicz G.K., Zilly A., Marques de Souza C.G., Peralta R.M. (2004). Decolourisation of industrial dyes by solid-state cultures
of Pleurotus pulmonarius. Process Biochemistry, 39: 855-859.
Ünyayar A., Sik S. (1998). Phanerochaete chrysosporium and Funalia trogii for the degradation of cotton stalk and their laccase,
peroxidase, ligninase and cellulase enzyme activities under semi-solid state conditions. Turkish Jornal Biology, 22: 287-298.
Uribe-López A.M. (1983). Hongos comestibles: otro producto de las plantaciones de coníferas no explotado en Colombia
[Agaricales, Aphyllophorales, Lycoperdales, Pezizales]. ed. Agronomía F.d. Universidad Nacional de Colombia, Sede
Medellín: Medellín (Colombia). 73 p.
Valbuena D.C., Alzate C.J. (2007). Evaluación de la patogenicidad de los hongos Bauveria basiana y Metarhizium anisopliae en el
control de la garrapata del ganado Rhipicephalus micraplus en su fase parasítica en su estado larval y ninfal.Trabajo de grado.
Microbiología Agrícola y Veterinaria, Javeriana: Bogotá. 144 p. p.
Van de Lagemaat J., Pyle D.L. (2005). Modelling the uptake and growth kinetics of Penicillium glabrum in a tannic acid-containing
solid-state fermentation for tannase production. Process Biochemistry, 40: 1773-1782.
Vares T., Kalsi M., Hatakka A. (1995). Lignin peroxidases, manganese peroxidases and other ligninolytic enzymes produced by
Phlebia radiata during solid state fermentation of wheat straw. Applied and Environment Microbiology 61: 3515-3520.
Vasco-Palacios A.M., Franco-Molano A.E., López-Quintero C.A., Bohechout T. (2005). Macromicetes (ascomicota, basidiomicota)
de la región del medio Caquetá, departamentos de Caquetá y Amazonas (Colombia). Biota Colombiana, 6(1): 127-140.
Vera D.F., Pérez H., Valencia H. (2002). Aislamiento de hongos solubilizadores de fosfatos de la rizosfera de arazá (Eugenia
stipitata, Myrtaceae). Acta Biológica Colombiana 7(1): 33-39.
Vetter J. (2007). Chitin content of cultivated mushrooms Agaricus bisporus, Pleurotus ostreatus and Lentinula edodes. Food
Chemistry, 102: 6-9.
Visser J., Beldman G., Kusters–van Someran M.A., Voragen A.G.J. (1991). International Conference of Xylans and Xylanases
Netherlands: Elsevier. pp. 89-94.
Vroemen A.J. (2002). Glucose oxidase. En: Handbook of food enzymology. Witaker J.R., Voragen A.G., Wong D.W.S. (Eds.).
Marcel Dekker, INC: New York. pp. 425-432.
Wade L.G. (1993). Química Orgánica. 2 ed, ed. Educación P. Debra Wechsler: México. 1312 p.
Wainwright M. (1988). Metabolic diversity of fungi in relation to growth and mineral cycling in soil—a review. Transactions of the
British Society of Mycology, 90: 159-170.
Wasser S. (2002). Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Applied Microbiology
and Biotechnology, 60: 258-274.
Wasser S.P., Weis A.L. (1999). Medicinal properties of substances occurring in higher Basidiomycete mushrooms: current
perspective (review). International Journal Medicine Mushrooms, 1: 31-62.
Weinberg Z.G. (2000). Biotechnology in developing countries: opportunities in solid state fermentation applied in the agricultural
industry. Internation Journal of Biotechnology, 2(4): 364-373.
Whittaker R.H. (1969). New Concepts of Kingdoms of Organisms. Science, 163: 150-160.
Wong D.W.S. (2009). Structure and Action Mechanism of Ligninolytic Enzymes. Applied Biochemestry and Biotechnology, 157:
174-209.
Wood T.M., García C.V. (1994). Enzymes and mechanisms involved in microbial cellulolysis. Kluwer Academic publishers:
Netherlands.
Wu J., Ding Z.-Y., Zhang K.-C. (2006). Improvement of exopolysaccharide production by macro-fungus Auricularia auricula in
submerged culture. Enzyme and Microbial Technology, 39: 743-749.
Wyman C.E., Dale B.E., Elander R.T., Holtzapple M., Ladisch M.R., Lee Y.Y. (2005). Coordinated development of leading biomass
pretreatment technologies. Bioresource Technology, 96: 1959-1966.
Xie J., Sun X., Ren L., Zhang Y.Z., Wu Kung S. (2001). Studies in lignocellulolytic enzymes production and biomass degradation of
Pleurotus sp.2 and Trametes gallica in wheat straw cultures. Chemical Engineering Journal and the Biochemical Engineering
Journal, 17: 575-578.

Xu H., Sun L.-P., Shi Y.Z., Wu Y.W., Zhang B., Zhao D.Q. (2008). Optimization of cultivation conditions for extracellular
polysaccharide and mycelium biomass by Morchella esculenta As51620 Biochemical Engineering Journal, 39: 66-73.
Yachmenev V., Condon B., Klasson T., Lambert A.J. (2009). Acceleration of the enzymatic hydrolysis of corn stover and sugar cane
bagasse celluloses by low intensity uniform ultrasound. Biobased Material Bioenergy, 3: 25-31.
Yalin W., Ishurd O., Cuirong S., Yuanjiang P. (2005). Structure analysis and anti-tumor activity of (1→3)-β-D-glucans
(cordyglucans) from the mycelia of Cordyceps sinensis. Planta Medica, 71: 381-384.
Yang B.-K., Gu Y.-A., Jeong Y.-T., Jeong H., C-H. S. (2007). Chemical characteristics and immune-modulating activities of exo-
biopolymers produced by Grifola frondosa during submerged fermentation process. International Journal of Biological
Macromolecules, 41: 327-333.
Yang F.-C., Ke Y.-F., Kuo S.-S. (2000). Effect of fatty acids on the mycelial growth and polysaccharide formation by Ganoderma
lucidum in shake flask cultures. Enzyme and Microbial Technology, 27: 295-301.
Zabel R.A., Morrel J.J. (1992a). Decay and its Prevention. Academic Press: San Diego. 534 p. p.
Zabel R.A., Morrel J.J. (1992b). Decay and its Prevention. Academic Press: San Diego, California.
Zaks A. (2001). Industrial biocatalysis. Chemical Biology, 5: 130-136.
Zanchetta P., Lagarde P.N., Guezennec J. (2003). A new bone-healing material: A hyaluronic acid-like bacterial exopolysaccharide.
Calcified Tissue International, 72: 74-79.
Zapata P., Rojas D., Carlos Fernández C., Ramírez D., Restrepo G., Orjuela V., Arroyave M., Gómez T., Atehortúa L. (2007).
Producción de biomasa y exopolisacáridos de Grifola frondosa bajo cultivo sumergido utilizando fuentes de carbono no
convencionales. Revista EIA, ISSN 1794-1237, (7): 137-144.
Zapata P., Rojas D., Atehortúa L. (2012). Production of Biomass, Polysaccharides, and Ganoderic Acid using Non-conventional
Carbon. International Journal of Medicinal Mushrooms, 14(2): 197-203.
Zhang P., Himmel M.E., Mielenz J.R. (2006a). Outlook for cellulase improvement: Screening and selection strategies. Biotechnology
Advances 24: 452-481.
Zhang Y.-H.P., Himmel M.E., Mielenz J.R. (2006b). Outlook for cellulase improvement: Screening and selection strategies.
Biotechnology Advances, 24: 452-481.
Zhao J., Janse B.J. (1996). Comparison of H2O2-producing enzymes in selected white rot fungi. FEMS Microbiology Letters, 139:
215-221.
Zhao J., Koker T., Janse B.J. (1996). Comparative studies of lignin peroxidases and manganese-dependent peroxidases produced by
selected white rot fungi in solid media. FEMS Microbiology Letters, 143: 393-399.
Zhao X.R., Lin Q., Brookes P.C. (2005). Does soil ergosterol concentration provide a reliable estimate of soil fungal biomass? Soil
Biology and Biochemistry, 37: 311-317.
Zhu S., Wu Y., Yu Z., Wang C., Yu F., Jin S., Ding Y., Chi R., Liao J., Zhang Y. (2006). Comparison of three microvawe/chemical
pretreatment processes for enzymaric hydrolyisis of rice straw. Biosystem Engineering, 93: 279-283.
Zhuang O., Mizuno T. (1999). Biological responses from Grifola frondosa (Dick.:Fr.) S.F. Gray-Maitake (Aphyllophoromycetideae).
Internation Journal medicinal mushrooms, 1: 317-324.

72
3. Evaluación de Actividades Endoglucanasa,
Exoglucanasa, Lacasa y Lignina Peroxidasa en Diez
Hongos de Pudrición Blanca

1 Magíster en Ingeniería-Ingeniería Química, Instituto de Biotecnología Agropecuaria, Universidad de Caldas, Manizales, Colombia.
2 Doctor en Ingeniería, Instituto de Biotecnología Agropecuaria, Universidad de Caldas, Manizales, Colombia.
3 Doctora en Ciencias, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Argentina.
Correspondencia: [email protected]
EVALUACIÓN DE ACTIVIDADES ENDOGLUCANASA, EXOGLUCANASA, LACASA Y LIGNINA PEROXIDASA EN DIEZ HONGOS DE PUDRICIÓN BLANCA
EVALUATION OF ENDOGLUCANASE, EXOGLUCANASE, LACCASE, AND LIGNIN
PEROXIDASE ACTIVITIES ON TEN WHITE-ROT FUNGI
AVALIAÇÃO DE ATIVIDADES ENDOGLUCANASE, EXOGLUCANASE, LACASE E LIGNINA PEROXIDASE EM DEZ FUNGOS DA PODRIDÃO BRANCA
SANDRA MONTOYA B. 1, ÓSCAR JULIÁN SÁNCHEZ T. 2, LAURA LEVIN 3
RESUMEN Este trabajo presenta una vía de rastreo de producción de enzimas lignocelulolíticas en diez especies de hongos de pudrición blanca: Lentinula edodes, Schizophyllum commune, Trametes trogii, Coriolus versicolor, Pycnoporus sanguineus, Ganoderma applanatum, Ganoderma lucidum, Grifola frondosa, Pleurotus ostreatus y Auricularia delicata. Estas especies primero fueron rastreadas sobre medios de cultivo sólido que contienen carboximetil celulosa, celulosa cristalina, ABTS (2,2´-azino-bis(3-etilbenzotiazolina-6-sulfonato)) y azure B, los cuales evidenciaron la producción de las enzimas endoglucanasa, exoglucanasa, lacasa y lignina peroxidasa (LiP). Las actividades celulolíticas fueron detectadas a los cinco días de incubación con el indicador rojo congo, formándose un halo claro-blanco en las zonas donde se degrada la celulosa. Para las ligninasas, este rastreo consistió en el seguimiento a la formación de halos verdes por oxidación del ABTS para lacasa y halos de decoloración sobre el azure B para la LiP durante 14 días de incubación. De este rastreo cualitativo, se seleccionaron cuatro cepas (G. lucidum, L. edodes, C. versicolor y T. Trogii), como las mejores productoras de enzimas celulolíticas y ligninolíticas. Estas cuatro especies fueron inoculadas sobre un sustrato de aserrín de roble, obteniéndose 51,8% de lignina degradada por L. edodes y 22% de celulosa degrada por C. versicolor. PALABRAS CLAVE: Celulasas, Ligninasas, Lentinula edodes, Coriolus versicolor, Celulosa. ABSTRACT This paper presents a way of tracking the production of lignocellulolytic enzymes in ten species of white rot fungi: Lentinula edodes, Schizophyllum commune, Trametes trogii, Coriolus versicolor, Pycnoporus sanguineus, Ganoderma applanatum, Ganoderma lucidum, Grifola frondosa, Pleurotus ostreatus and Auricularia delicata. These species were first screened on solid culture media containing carboxymethyl cellulose, crystalline cellulose, ABTS (2,2´-azino-bis(3-ethylbenzothiazoline-6-sulphonate)) and azure B, which showed the production of endoglucanase, exoglucanase, laccase and lignin peroxidase (LiP) enzymes. Cellulolytic activities were detected after five days of incubation with congo red indicator, forming a clear-white halo in areas where cellulose was degraded. For ligninases, the tracking consisted of the monitoring in the formation of green halos due to ABTS oxidation for laccase, and decolorization halos on azure B for LiP during 14 days of incubation. From this qualitative screening, four strains were selected (G. lucidum, L. edodes, C. versicolor and T. trogii) as the best producers of cellulolytic and ligninolytic enzymes. These four species were inoculated on a substrate of sawdust oak, yielding 51.8% of lignin degraded by L. edodes and 22% of cellulose degraded by C. versicolor.

74
KEYWORDS: Cellulases, Ligninases, Lentinula edodes, Coriolus versicolor, Cellulose. RESUMO Este trabalho apresenta uma maneira de seguir produção da enzimas lignocelulolíticas em 10 espécies de fungos da podridão branca: Lentinula edodes, Schizophyllum commune, Trametes trogii, Coriolus versicolor, Pycnoporus sanguineus, Ganoderma applanatum, Ganoderma lucidum, Grifola frondosa, Pleurotus ostreatus e Auricularia delicata. Estas espécies foram primeiramente rastreados em meio de cultura sólido contendo os reagentes de carboximetil celulose, celulose cristalina, ABTS (2,2 '-azinobis (3-etilbenztiazoline-6-sulfonato)) e azure B que mostrou a produção de enzimas de endoglucanase, exoglucanase lacase, peroxidase de lignina e (LiP). Actividades celulolíticas foram detectados após cinco dias de incubação com o indicador vermelho congo, formando um claro halo-branco em áreas degrada a celulose. Para ligninases, esta consistia de rastreamento faixa formação de halo verde por oxidação de ABTS a lacase e descoloração halos no lábio azure B, durante 14 dias de incubação. Esta triagem qualitativa, foram selecionados quatro cepas: G. lucidum, L. edodes, C. versicolor e T. trogii, cepas como os melhores produtores de enzimas celulolíticas e ligninolíticas. Estas quatro espécies foram inoculados sobre um substrato de madeira de carvalho serradura, obtendo-se 51,8% de lignina por L. edodes degradad e 22% de celulose degradada por C. versicolor.
PALAVRAS-CHAVE: Celulases, Ligninases, Lentinula edodes, Coriolus versicolor, Celulose.
INTRODUCCIÓN
Las enzimas lignocelulolíticas juegan un rol importante en la conversión del complejo lignocelulósico en azúcares fermentables para la producción de alcohol carburante en la actualidad [1]. Las enzimas celulolíticas son consideradas de gran importancia industrial. Por ejemplo, ellas han sido ampliamente utilizadas para reemplazar la hidrólisis ácida como método de conversión de celulosa en azúcares fermentables, especialmente glucosa para la producción de etanol y otros alcoholes de importancia industrial, asimismo, para la glucosa obtenida es utilizada como materia prima para la manufactura de diferentes alimentos para animales, humanos y productos químicos. Las celulasas también son utilizadas en diversos procesos de fermentación para la producción de sustancias de uso industrial como el ácido láctico, en procesos de fermentación aceto-
butílica; en el mejoramiento de la textura del papel (liso), biodesentintado para reciclado del papel utilizando una mezcla de celulasas, pectinasas, xilanasas y lipasas; en el biopulido de telas para el tratamiento superficial de las pelusas, en la elaboración de paños super absorbentes, en la extracción de jugos, aromas, aceites y pigmentos con agregados enzimáticos de celulasas, pectinasas y xilanasas, éstas rompen paredes celulares y hace más eficiente la extracción; en la elaboración de aditivos no calóricos [2-5]. De igual forma, las ligninasas son utilizadas para la deslignificación de los materiales lignocelulósicos dispuestos para alimentación de rumiantes. Por ejemplo, la industria de producción de carne bovina en el mundo presenta dificultades para mantener estable el suministro de alimentos para los animales. Uno de los principales inconvenientes es la variación del clima, lo que ha generado que este segmento de la

75
industria alimentaria busque nuevas alternativas para el mantenimiento de la alimentación animal [6]. En el planeta se desechan grandes cantidades de materiales lignocelulósicos que generan contaminación y que pueden ser utilizados para este fin utilizando complejos de enzimas ligninolíticas para la degradación de la lignina y posteriormente realizar los ensilajes o las transformaciones y suplementaciones requeridas por los animales [7, 6]. La producción de enzimas degradadoras del complejo lignocelulósico, se realiza a partir de microorganismos que las producen, entre los que se encuentran los hongos de pudrición blanca [8]. Los materiales lignocelulósicos en promedio se componen de 40-50% de celulosa, 25-40% de hemicelulosa y 25-35% de lignina y otros monómeros en menor proporción [9]. Estos materiales pueden ser degradados en medios naturales por los hongos de pudrición blanca a través de las enzimas que excretan, como el complejo de celulasas celulasas: endoglucanasas, exoglucanasas, otras que degradan oligosacáridos como la β-glucosidasa, las óxidorreductasas, como lacasas, manganeso peroxidasa y lignina peroxidasa, las cuales son las enzimas responsables de la degradación del complejo lignocelulósico [10, 11].
Los materiales lignocelulósicos son los sustratos indicados para el desarrollo de muchos hongos. Estos materiales lignocelulósicos constituyen el principal componente de los vegetales y sus residuos, así como de residuos agroindustriales, forestales y residuos sólidos urbanos, entre otros [5]. Los hongos de pudrición blanca presentan como característica especial la capacidad de degradar la lignina hasta su mineralización
a CO2 y agua gracias a las grandes cantidades de óxido-reductasas que excretan. Además tienen la capacidad de producir y transformar sustancias no fenólicas en fenólicas para su fácil degradación y la obtención de sustancias aromáticas de menor tamaño que la lignina y de menor toxicidad. Aunque estos hongos no pueden usar el carbono obtenido como fuente de energía, ellos son considerados como los mejores organismos degradadores de sustancias aromáticas que han sido encontrados hasta el momento [12]. Estos hongos también degradan celulosa y hemicelulosa, cuya hidrólisis provee una fuente verdadera de energía [13, 3]. Entre los hongos de pudrición blanca como potentes agentes productores de enzimas lignocelulolíticas, se encuentran especies de los géneros Auricularia, Schizophyllum, Lentinus, Polyporus, Pycnoporus y Trametes, entre otros [14]. Especies de estos géneros como Coriolus versicolor, Ganoderma lucidum, Ganoderma applanatum, Schizophyllum commune, Pycnoporus sanguineus, Pleurotus ostreatus, Grifola frondosa, Auricularia delicata, Trametes trogii y Lentinula edodes, en la actualidad son reconocidas por su gran capacidad de degradación de celulosa y lignina. Asimismo, estos hongos han sido utilizados en diferentes procesos con fines de biodegradación de diferentes polímeros: carbohidratos, aromáticos, hidrocarburos, colorantes sintéticos, etc. [15-17]. Los métodos cualitativos constituyen una buena herramienta para la búsqueda rápida (rastreo) de enzimas que degraden el complejo lignocelulósico [9, 18]. Estas pruebas proporcionan una indicación certera de la presencia de estas enzimas con el fin de realizar una adecuada selección de cepas de hongos respecto al grupo de enzimas que producen,

76
disminuyendo la cantidad de pruebas cuantitativas, que son mucho más costosas y demandan mayor tiempo. Sin embargo, estos métodos tienen limitaciones especialmente de precisión, aunque juegan un rol importante en procesos de selección de cepas [19, 20]. Actualmente se emplean varios métodos cualitativos para la detección de celulasas, entre los que se encuentran los medios sintéticos con el reactivo específico sobre el cual actúa la enzima. Por ejemplo, para la detección de la endoglucanasa se emplea carboximetil celulosa (CMC), para exoglucanasas se usa celulosa cristalina o agar azure celulosa. Estas dos enzimas o grupo de enzimas generalmente se detectan por la formación de un halo blanco alrededor de la colonia. Para la β-glucosidasa se emplea agar esculina (6,7dihidroxicumarino 6-glucósido) como fuente única de carbono. La enzima β-glucosidasa forma glucosa, la cual es excretada al medio y forma un halo negro en torno a la colonia; este halo negro también puede formarse cuando se realizan las pruebas para las enzimas celulolíticas, endoglucanasas y exoglucanasas [9, 21]. Para detectar ligninasas, se emplean reactivos que las óxido-reductasas puedan degradar. Por ejemplo, para la detección de ligninasas como la enzima lignina peroxidasa (LiP) se utiliza azure B, el cual se decolora por la acción de esta enzima [22] y la detección de lacasa se hace a través de la preparación de un medio con el reactivo ABTS (2,2'azino- bis (3-etilbenzo-tiazolin-6-ácido sulfónico)), el cual se oxida en presencia de la enzima lacasa [23, 24]. El objetivo de este trabajo fue determinar el microorganismo con mayor potencial para degradar celulosa y lignina sobre residuos lignocelulósicos mediante el rastreo de las enzimas lignocelulolíticas producidas por
diez especies de hongos macromicetos: L. edodes, S. commune, T. trogii, C. versicolor, P. sanguineus, G. applanatum, G. lucidum, G. frondosa, P. ostreatus y A. delicata.
MÉTODO Hongos Los siguientes hongos de pudrición blanca empleados para el rastreo fueron obtenidos de la colección del Laboratorio de Macromicetos de la Universidad de Caldas: L. edodes (CICL54), S. commune (UCS004), T. trogii (BAFC463), C. versicolor (PSUWC430), P. sanguineus (UCS003), G. applanatum (UCS001), G. lucidum (UCC002), G. frondosa (PSUMCC 922), P. ostreatus (UCC001) y A. delicata (UCS002). Las cepas fueron mantenidas en caja de Petri con agar papa dextrosa (PDA) a 4°C. Rastreo de celulasas Los medios de cultivo para el rastreo de las actividades endoglucanasa y exoglucanasa, se formularon con 1,7% de agar-agar, y 0,1% de CMC para las endoglucanasas y 0,1% de celulosa cristalina para las exoglucanasas como única fuente de carbono. El medio fue autoclavado durante 15 minutos a 121°C. Las diez cepas de hongos fueron inoculadas en cuatro cajas de Petri por especie y por triplicado y se incubaron por 5 días a 25°C. Las colonias fueron reveladas con el colorante azoico rojo congo al 0,3% por 15 minutos con posterior lavado del colorante empleando una solución 1 N de NaCl [9]. En todos los casos se realizó observación y medición de la formación de halos de decoloración y del crecimiento micelial de cada una de las especies.

77
Rastreo de ligninasas Para estimar la presencia de LiP se empleó el colorante heterocíclico azure B en una concentración de 50 µM en medio agar PDA. Para la enzima lacasa se empleó ABTS al 0,035% (p/v) en un medio heterogéneo con la siguiente composición en g/L: 10 glucosa, 2 K2HPO4, 0,5 MgSO4.7H2O, 0,2 extracto de levadura, 0,5 (NH4)2SO4, 0,1 Cl2Ca, 0,0064 CuSO4, 22 agar-agar y 0,35 de ABTS. Se inocularon las diez cepas preseleccionadas en cuatro cajas de Petri por triplicado. Se incubaron por 14 días a 25°C. Posteriormente se determinó el halo de decoloración para los medios con azure B y la formación de halo verde para los medios con ABTS, así como el tamaño de la colonia en todos los casos [9]. Determinación del contenido de fibra de los sustratos Para determinar el contenido de fibra fue necesario elaborar sustratos empleando 98% de aserrín de roble y 2% carbonato de calcio (porcentajes en base seca) al 60% de humedad y pH 5,5. El sustrato fue empacado en frascos de 125 mL con 30 g y llevados a esterilización a 121°C por 30 minutos. Posteriormente los sustratos fueron inoculados al 4% con cada una de las cepas (cuatro frascos por cada especie) y se llevaron a incubación por 30 días en penumbra y 25°C. Luego del proceso de inoculación, los frascos fueron tapados con interlón y película parafinada con el fin de filtrar el aire enurante el tiempo de incubación. Este experimento se realizó por triplicado. La determinación de la fibra se realizó a partir de la medida de fibra detergente neutra, fibra detergente ácida y fibra lignino-ácida, a partir de las cuales fueron calculados los contenidos de fibra
soluble, hemicelulosa, celulosa y lignina [25]. Determinación cuantitativa de actividades enzimáticas Los extractos para la determinación de las actividades enzimáticas (celulolíticas y ligninolíticas) fueron obtenidos por la adición de 12 mL de agua destilada estéril neutra a 1 g de medio sólido fresco, sometido a ultrasonido por 5 minutos y agitación por 10 minutos con posterior filtración y centrifugación. Hidrolasas. Para la determinación de la actividad endo-1,4-ß-D-glucanasa (E.C.3.2.1.4) se usó CMC como sustrato al 0,5% en buffer acetato de sodio pH 4,8 en reacción con 100 µL de extracto enzimático por 30 minutos a 50°C. La reacción enzimática se detuvo con la adición del reactivo DNS (ácido dinitrosalicílico) [26]. Se continuó con la reacción de DNS para la determinación de azúcares reductores, se leyó la absorbancia a una longitud de onda de 540 nm. Una unidad de actividad enzimática fue definida como la cantidad de enzima que produce 1 µmol de azúcares reductores en un minuto. Para cuantificar la actividad se elaboró una curva patrón de azúcares reductores con distintas concentraciones de glucosa. Para la actividad exo-1,4-ß-D-glucanasa (E.C.3.2.1.91) se usó celulosa cristalina al 1% en buffer acetato de sodio con pH de 4,8 como sustrato, en reacción con 100 µL de extracto enzimático a 50°C por 60 minutos. La reacción se detuvo adicionando el reactivo DNS. Se continuó con la determinación de azúcares reductores por el método DNS y se centrifugó antes de leer la absorbancia a una longitud de onda de 540 nm. Una unidad de actividad enzimática de exoglucanasa fue definida como la cantidad de enzima que produce 1 µmol de

78
azúcares reductores en un minuto. La actividad β-glucosidasa (E.C.3.2.1.21) se determinó por la reacción que se produce entre el sustrato, p-nitrofenil β-D-glucopiranosido al 0,02%, en buffer acetato de sodio con pH de 4,8 y 100 µL de extracto enzimático a 50°C por 30 minutos. La reacción se detuvo adicionando solución buffer Clark y Lubs (pH 9,8) y se leyó la absorbancia a una longitud de onda de 430 nm (ε430=18,5/mM cm) (Wood y Bhat, 1988). Una unidad de actividad enzimática β-glucosidasa se define como la cantidad de enzima que produce 1 µmol de producto (p-nitrofenol) en un minuto. Para cuantificar la actividad se elaboró una curva patrón con p-nitrofenol. La actividad endo-1,4-ß-D-xilanasa (E.C.3.2.1.8) se determinó mediante la reacción producida entre el sustrato xilano al 0,2% en buffer acetato de sodio con un pH de 4,8 y 100 µL de extracto enzimático a 50°C por 30 minutos. La reacción se detuvo adicionando DNS y se leyó la absorbancia a una longitud de onda de 540 nm. Una unidad de actividad enzimática de endo-1,4-ß-D-xilana se define como la cantidad de enzima que produce 1 µmol de azúcares reductores en un minuto. La curva patrón de azúcares reductores se elaboró con xilosa. Actividades ligninolíticas. Para la determinación de la actividad lacasa (E.C.1.10.3.2) se utilizaron 0,5 mM de ABTS en una solución 0,1M de buffer acetato de sodio a un pH de 3,6 como sustrato, según el método de Paszczynski y Crawford [27], leyendo el aumento de absorbancia a 420 nm (ε420=36/mM cm) luego de tres minutos de reacción a 30°C. Una unidad de actividad de lacasa fue definida como la cantidad de enzima requerida para oxidar 1 μmol de ABTS en 1 minuto. Para la cuantificación de la actividad lignina peroxidasa (E.C.1.11.1.14) se aplicó el método del azure B [22], que usa como
sustrato una solución 32 µM de azure B, 100 µM de H2O2 y 150 µL de extracto enzimático. La mezcla de reacción se realizó en buffer tartrato de sodio 50 mM a un pH de 4,5 y se inició la reacción con una solución de peróxido de hidrógeno 100 μM. La lectura de la disminución de absorbancia se hizo a 650 nm. Una unidad actividad enzimática es equivalente al decrecimiento de 0,1 unidades de absorbancia por minuto y por mL (U/min/mL). Análisis estadístico Para la selección de las cepas con mejor comportamiento en la producción de enzimas celulolíticas y ligninolíticas se realizaron dos experimentos sucesivos. El primero, consistió en el seguimiento de las 10 cepas aplicando técnicas semicuantitativas registrando el diámetro de los halos de decoloración /oxidación de los diferentes sustratos. El segundo experimento se deriva de los resultados del primero; a las cuatro cepas con mejor respuesta, se les realizó una prueba cuantitativa de degradación de fibra discriminada. En el primer caso, se realizó un diseño completamente aleatorizado a una vía donde la variable de respuesta fue la formación de los halos y el número de repeticiones fue de 12. Para el caso de las enzimas celulolíticas se tuvo en cuenta la formación del halo de degradación del colorante rojo congo (endoglucanasa y exoglucanasa). En el caso de las enzimas ligninolíticas se realizaron pruebas para la lacasa siendo su variable de respuesta la formación de halo de oxidación verdoso; también se hicieron pruebas para la LiP donde la variable de respuesta fue la formación de un halo de decoloración sobre el medio de cultivo con azure B. En el segundo experimento consistente en la determinación cuantitativa de la fibra, se realizó un análisis estadístico para

79
determinar la capacidad de degradación de lignina, celulosa y hemicelulosa sobre aserrín de madera, incubado a 25°C durante 30 días. Para la evaluación del rastreo de celulasas se realizó un análisis de varianza ANOVA a una vía, tomando como factor la cepa a 10 niveles y como variable de respuesta la formación de los halos. Se empleó el programa Statgraphics® Versión 5.1. También se evaluó la capacidad de degradación y las actividades enzimáticas lignocelulolíticas sobre sustrato lignocelulósico, de las cuatro especies de hongos preseleccionadas en las pruebas cualitativas. Para ello se realizó un ANOVA a una vía utilizando el mismo programa estadístico. RESULTADOS Rastreo en placa de enzimas celulolíticas A las diez cepas pre-seleccionadas (L. edodes, S. commune, T. trogii, C. versicolor, P. sanguineus, G. applanatum, G. lucidum, G. frondosa, P. ostreatus y A. delicata) se les realizó un rastreo a fin de seleccionar las cepas con mejor comportamiento en la producción de las enzimas celulolíticas endoglucanasa y exoglucanasa. Se evaluó, luego de cinco días de incubación, el tamaño de los halos de degradación formados y el tamaño de la colonia en mm. Como resultado del ANOVA se obtuvo un valor p < 0,5 (0,000), lo que mostró una diferencia estadísticamente significativa entre las cepas a un nivel de confianza del 95% para definir cuáles cepas fueron significativamente diferentes unas de otras. Se realizó una prueba de Tukey a fin de determinar que tratamientos fueron estadísticamente diferentes. Posteriormente, se realizó el análisis de los residuales de homocedasticidad y de
normalidad siempre y cuando la varianza fuera homogénea. En ningún caso los residuales pasaron ambas pruebas (homocedasticidad y normalidad) por lo que se debió realizar la prueba no paramétrica de Kruskal Wallis. Esta prueba mostró diferencia significativa con valor p < 0,5 (0,000) e indicó que las mejores cepas para la producción de enzimas celulolíticas fueron G. lucidum, L. edodes y C. versicolor. No todas las cepas evaluadas resultaron capaces de crecer en medios con celulosa como única fuente de carbono. Algunas cepas crecieron muy poco o no presentaron crecimiento sobre ninguno de los dos medios (G. frondosa, A. delicata, G. applanatum y S. commune); sin embargo, en todos los casos se observó secreción de enzimas celulolíticas ya que se apreció un halo decolorado alrededor del inóculo, no superior en promedio a los 12 mm (Cuadro 14). Las cepas de P. sanguineus, T. trogii y P. ostreatus crecieron y decoloraron el rojo congo, pero en menor medida que las cepas de L. edodes, G. lucidum y C. versicolor, Entre ellas se destacó la cepa C. versicolor. Cuadro 14. Diámetros del halo de crecimiento
micelial al quinto día de incubación.
Especie Diámetro halo/crecimiento micelial (mm)
A. delicata 8
S. commune 6,5
T. trogii 26
C. versicolor*
65
G. frondosa 10
G. lucidum*
59
G. applanatum 12
P. sanguineus 27
L. edodes*
62
P. ostreatus 31 *Estadísticamente diferentes a las demás especies en producción de celulasas

80
Rastreo en placa de enzimas ligninolíticas La evaluación del rastreo de ligninasas (lacasa y LiP) se realizó con un ANOVA a una vía, tomando como factor la cepa a 10 niveles y como variable de respuesta la formación de los halos. Como resultado del ANOVA se obtuvo una valor p < 0,05 (0,000), lo que mostró una diferencia estadísticamente significativa entre las cepas con un nivel de confianza del 95%. Para definir cuáles cepas fueron significativamente diferentes unas de otras se realizó una prueba de Tukey a fin de determinar que tratamientos fueron estadísticamente diferentes. Posteriormente, se realizó el análisis de los residuales de homocedasticidad y de normalidad, siempre y cuando la varianza fuera homogénea. En ningún caso los residuales pasaron ambas pruebas (homocedasticidad y normalidad) por lo que se debió realizar la prueba no paramétrica de Kruskal Wallis. Esta prueba arrojó diferencia significativa con valor p < 0,05 (0,000) e indicó que los mayores productores de enzimas ligninolíticas fueron T. trogii y C. versicolor. En el Cuadro 2 se muestran los resultados obtenidos de los ensayos con ABTS (para detectar lacasa) y azure B (para detectar LiP), donde (-) corresponde a un halo verdoso de oxidación del ABTS o blanquecino de decoloración del azure B menor a 1 cm de diámetro, (+) corresponde a un halo entre 1 y 2,9 cm de diámetro, y (++) corresponde a un halo mayor a 3 cm de diámetro a los 14 días de incubación. De las 10 cepas de hongos, solamente el S. commune dio negativa (-) en las tres repeticiones que se realizaron a cada cepa para las dos enzimas rastreadas (lacasa y LiP), lo que implica que para esta cepa debe realizarse una serie de pruebas adicionales a fin de corroborar su comportamiento frente a la producción de
enzimas ligninolíticas. En las cajas de ABTS con L. edodes, G. lucidum, C. versicolor y T. trogii se observó un halo de oxidación verde intenso mayor a 3 cm de diámetro (++) y en las cajas con azure B, T. trogii y C. versicolor produjeron halos de decoloración mayores a 3 cm (++). Luego de realizar la prueba no paramétrica de Kruskal Wallis a todos los datos obtenidos de estas pruebas cualitativas de rastreo de enzimas ligninolíticas se obtuvo diferencia significativa (valor p < 0,05) entre las especies de hongos, resultando C. versicolor y T. trogii como las mejores cepas productoras de ligninasas. Cuadro 15. Capacidad de oxidación del ABTS o
decoloración del azure B.
Cepa Oxidación del ABTS
Decoloración del azure B
L. edodes ++ -
A. delicata + -
G. frondosa + +
S. commune - -
G. applanatum + +
G. lucidum ++ +
C. versicolor*
++ ++
T. trogii*
++ ++
P. sanguineus + -
P. ostreatus + - *Estadísticamente diferentes a las demás especies en producción de ligninasas
Capacidad de degradación de celulosa y lignina De acuerdo con los resultados obtenidos del rastreo cualitativo realizado, en las cuatro cepas con mejor respuesta (L. edodes, C. versicolor, G. lucidum y T. trogii) se evaluó la capacidad de degradación de lignina y celulosa sobre un sustrato sólido a base de aserrín de roble durante 30 días de incubación (Figura 1). A los resultados obtenidos de la degradación de los componentes de la fibra en forma discriminada por cada una de las cuatro especies de hongos preseleccionadas en

81
las pruebas cualitativas, se les realizó un ANOVA a una vía arrojando como resultados que hubo diferencia significativa en la degradación de lignina con un valor-p < 0,05 (valor p = 0,0219). El hongo L. edodes degradó el mayor porcentaje de lignina (51,8%) correspondiente a una disminución de la cantidad de lignina del 29,86% en el sustrato crudo a 14,4% después de 30 días de incubación, seguido de T. trogii el cual a los 30 días de incubación dejó el sustrato con 14,94% de lignina. Asimismo, se realizó una prueba de comparación múltiple entre las cuatro especies de hongos y los datos finales de lignina obtenidos, ratificando que L. edodes fue la especie que degradó la mayor cantidad de lignina en 30 días de incubación con un nivel de significancia del 95%. En referencia a la degradación de celulosa, se encontró que hubo diferencia significativa (valor–p = 0,0362) entre las cuatro especies de hongos preseleccionadas corroborándose con una prueba de comparación múltiple a un nivel de significancia del 95%. C. versicolor degradó el mayor porcentaje de celulosa (22%) correspondiente a la disminución en la cantidad de celulosa de 59,56% en el sustrato crudo hasta 46,45% en el sustrato con 30 días de incubación, seguido de G. lucidum que redujo el porcentaje de celulosa en el sustrato hasta 46,95%. Sin embargo, dado que todas las cepas resultaron capaces de degradar porcentajes importantes de ambos polímeros, cualquiera de ellas podría utilizarse en la degradación de materiales lignocelulósicos; máxime teniendo en cuenta que los hongos de pudrición blanca pueden potenciar la acción de degradación de los componentes de la fibra variando las formulaciones de sustratos y las condiciones medio ambientales. Los resultados obtenidos en este trabajo tienen concordancia con
trabajos realizados por otros investigadores en referencia a los hongos basidiomicetos y en especial los de pudrición blanca, como potentes degradadores de lignina y celulosa [28, 29]. Figura 1. Componentes de la fibra en los sustratos
inoculados a los 30 días de incubación.
En el Cuadro 3 se detallan las actividades enzimáticas cuantitativas promedio (endoglucanasa, exoglucanasa, β- glucosidasa, endoxilanasa, lacasa y lignina peroxidasa) de los sustratos inoculados con las cuatro cepas preseleccionadas a los 30 días de incubación. C. versicolor y T. trogii exhibieron resultados significativamente diferentes de G. lucidum y L. edodes en cuanto a las actividades enzimáticas endoglucanasa (valor-p = 2,71×10-5) y exoglucanasa (valor-p = 9,16×10-5) se refiere, por lo que degradaron el mayor porcentaje de celulosa del sustrato (ver Figura 1). Sin embargo, respecto a la actividad de la β-glucosidasa las especies significativamente diferentes fueron L. edodes y C. versicolor con un valor-p = 0,0144. L. edodes mostró ser el hongo de mejor desempeño en cuanto a la degradación de lignina del sustrato utilizado (ver Figura 1) a pesar de no haberse encontrado diferencias significativas en la producción de enzimas ligninolíticas entre las cuatro cepas. No obstante, al analizar los resultados obtenidos debemos tener en cuenta que el método de extracción

82
utilizado puede no haber garantizado la extracción de todas las enzimas adsorbidas al sustrato. Cuadro 16. Actividades enzimáticas por gramo de
sólido seco (sustrato) inoculados con cuatro hongos durante 30 días de incubación. Cepa G.
lucidum L. edodes
C. versicolor
T. trogii
Endoglucanasa 3,872 3,371 9,798 10,514
Exoglucanasa
5,132 4,469 9,401 8,148
β- glucosidasa
1,896 4,436 3,496 4,024
Endoxilanasa
29,643 25,924 18,869 26,234
Lacasa 7,050 9,489 8,916 9,478
Lignina peroxidasa (LiP)
0,065 0,011 0,097 0,088
Las actividades enzimáticas están expresadas en [UE(µmol/g s.s min)], y la LIP en (U/mL/min)
CONCLUSIONES A partir del rastreo cualitativo en placa de la producción de celulasas y ligninasas en 10 cepas fúngicas causantes de pudrición blanca, se seleccionaron cuatro hongos: G. lucidum, L. edodes, C. versicolor y T. trogii y se evaluó su capacidad para degradar la lignina y la celulosa durante la fermentación en estado sólido de aserrín de roble. De los resultados obtenidos surge que cualquiera de estas cepas que se utilice con fines de degradación de materiales lignocelulósicos probablemente será efectiva. Entre ellas, sin embargo, L. edodes degradó el mayor porcentaje de lignina (51,8%) al cabo de 30 días, y C. versicolor el mayor porcentaje de celulosa (22%). Lo cual demuestra su potencialidad en procesos industriales de producción de enzimas a partir de materiales lignocelulósicos residuales.
AGRADECIMIENTOS Los autores agradecen a la Vicerrectoría de Investigaciones y Postgrados de la Universidad de Caldas (Colombia) por la financiación del presente trabajo y al Instituto de Biotecnología Agropecuaria de la misma institución por el apoyo material y de infraestructura. Asimismo, los autores expresan sus agradecimientos a la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires (Argentina) por su apoyo y asesoría durante la realización de este trabajo. REFERENCIAS [1] SÁNCHEZ, O.J. and CARDONA, C.A.
Trends in biotechnological production of fuel ethanol from different feedstocks. Bioresource Technology, 99, 2008, p. 5270-5295.
[2] JOSHI, V.K. and PANDEY, A. Biotechnology. Food fermentation Vol. II. New Delhi: Educational Publishers & Distributors, 1999, 998 p p.
[3] CARLILE, M., WATKINSON, S. and GOODAY, G. The fungi. Second ed. London: Academic Press, 2001, 588 p. p.
[4] BOMMARIUS, A.S. and RIEBEL, B.R. Biocatalysis. Atlanta, USA: Wiley-VCH Verlag GmbH & Co. KGaA, 2004, 611 p. p.
[5] SÁNCHEZ, O.J. y CARDONA, C.A. Producción de Alcohol Carburante: Una Alternativa para el Desarrollo Agroindustrial. Manizales: Universidad Nacional de Colombia, 2007, 386 p.
[6] KOUTINAS, A.A., WANG, R. and WEBB, C. Restructuring Upstream Bioprocessing: Technological and Economical Aspects for Production of a Generic Microbial Feedstock From Wheat. Published online in Wiley InterScience, 2004, p. 1-15.

83
[7] WEINBERG, Z.G. Biotechnology in developing countries: opportunities in solid state fermentation applied in the agricultural industry. Internation Journal of Biotechnology, 2 (4), 2000, p. 364-373.
[8] REDDY, C.A. The potential for white-rot fungi in the treatment of pollutants. Current Opinion in Biotechnology, 6, 1995, p. 320-328.
[9] POINTING, S.B. Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal Diversity, 2, 1999, p. 17-33.
[10] KIRK, T.K. and FARREL, R.L. Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology, 41, 1987, p. 465-505.
[11] ÂNGELO, A.S. Enzimas hidrolíticas. En: Fungos: Uma Introducâo à biologia, bioquimica e biotecnologia. Universidade de Caxias do Sul, 2004, p. 263-285.
[12] ARORA, D.S. and SHARMA, R.K. Ligninolytic fungal laccases and their biotechnological applications. Applied Biochemistry and Biotechnology, 160, 2010, p. 1760-1788.
[13] HUDSON, H. Fungal Biology. Maryland: Edward Arnolds Eds, 1986, 298 p. p.
[14] MONTOYA, S., GALLEGO, J., SUCERQUIA, A., PELAEZ, B., BETANCOURT, O. y ARIAS, D. Macromicetos observados en bosques del Departamento de Caldas: su influencia en el equilibrio y la conservación de la biodiversidad. Boletín Científico Centro de Museos de Historia Naural, 14 (2), 2010, p. 57-76.
[15] ARORA, D.S., CHANDER, M. and GILL, P.K. Involvement of lignin peroxidase, manganese peroxidase and laccase in degradation and selective ligninolysis of wheat straw. International Biodeterioration & Biodegradation, 50, 2002, p. 115-120.
[16] LITTHAUER, D., VAN VUUREN, M.J.,, VAN TONDER, A. and WOLFAARDT, F.W. Purification and kinetics of a thermostable laccase from Pycnoporus sanguineus (SCC 108). Enzyme and Microbial Technology, 40, 2007, p. 563-568.
[17] ELISASHVILI, V., KACHLISHVILI, E., KHARDZIANI, T. and AGATHOS, S.N. Effect of aromatic compounds on the production of laccase and manganese peroxidase by white-rot basidiomycetes. Journal of Industrial Microbiology and Biotechnology, 37, 2010, p. 1091-1096.
[18] NAZARENO, M.C., BUCSINSZKY, A.M.M., TOURNIER, H.A., CABELLO, M.N. and ARAMBARRI, A.M. Extracellular ABTS-oxidizing activity of autochthonous fungal strains from Argentina in solid medium. Revista Iberoamericana de Micología, 17, 2000, p. 64-68.
[19] MONTENECOURT, B.S. and EVELEIGH, D.E. Semiquantitative plate assay for determination of cellulase production by Trichoderma viride. Applied and environmental microbiology, 33 (1), 1977, p. 178-183.
[20] TAKANO, M., NISHIDA, A., NAKAMURA, M. Screening of wood-rotting fungi for kraft pulp bleaching by the Poly R decolorization test and biobleaching of hardwood kraft pulp by Phanerochaete crassa WD 1694. Journal of Wood Science, 47, 2001, p. 63-68.
[21] ZHANG, Y.-H.P., HIMMEL, M.E. and MIELENZ, J.R. Outlook for cellulase improvement: Screening and selection strategies. Biotechnology Advances, 24, 2006, p. 452-481.
[22] ARCHIBALD, F. A new assay for lignin-type peroxidases employing the dye Azure B. Applied Environmental Microbiology, 58, 1992, p. 3110-3116.

84
[23] THURSTON, C. The structure and function of fungal laccases. Microbiology-UK, 140, 1994, p. 19-26.
[24] EGGERT, C., TEMP, U., DEAN, J.F. and ERIKSSON, K.L. A fungal metabolite mediates degradation of non-phenolic lignin structures and synthetic lignin by laccase. FEBS Letters, 391, 1996, p. 144-148.
[25] LETERME, P. Análisis de alimentos y forrajes. (Protocolos de Laboratorio). Palmira: Universidad Nacional de Colombia, 2010, 66 pp. p.
[26] MILLER, G.L. Use of DinitrosaIicyIic acid reagent for determination of reducing sugar. Analytical Chemistry, 31 (3), 1959, p. 426-428.
[27] PASZCZYNSKI, A. and CRAWFORD, R.L. Degradation of azo compounds by
ligninases from Phanerochaete chrysosporium Involment of veratryl alcohol. Biochemistry Biophysics resources communications, 178, 1991, p. 1056-1063.
[28] BALDRIAN, P. and VALASKOVÁ, V. Degradation of cellulose by basidiomycetous fungi. FEMS Microbiology Review, 32, 2008, p. 501-521.
[29] MARTÍNEZ-ANAYA, C., BALCÁZAR-LÓPEZ, E., DANTÁN-GONZÁLEZ, E. y FOLCH-MALLOL, J., L. Celulasas fúngicas: Aspectos biológicos y aplicaciones en la industria energética. Revista Latinoamericana de Microbiología, 50 (3 y 4), 2008, p. 119-131.

85
4. Producción de Polisacáridos por Cultivos
Sumergido y en Estado Sólido a partir de Diferentes
Basidiomicetos

86
Polysaccharide production by submerged and solid state cultures from several
basidiomycetes S. Montoya
(a), Ó.J. Sánchez
(a), L. Levin
(b), e-mail: [email protected], Tel/fax: 8781500x15661
(a) Institute of Agricultural Biotechnology, Universidad de Caldas, Calle 65 No. 26-10, Manizales,
Colombia (b)
Biodiversity and Experimental Biology Department, PRHIDEB-CONICET, FCEN, Ciudad
Universitaria, Pabellón II, Piso 4, Laboratorio 8 C1428 GBA, Universidad de Buenos Aires, Argentina
Abstract
Polysaccharides produced by microorganisms represent an industrially unexploited market. An important
number of polysaccharides have been isolated from fungi, especially mushrooms, with many interesting
biological functions, such as antitumor, hypoglycemic and immuno-stimulating activities. In the search of
new sources of fungal polysaccharides, the main goal of this research was to test the ability of several
species of basidiomycetes, among them various edible mushrooms, for production of both extracellular
(EPS) and intracellular (IPS) polysaccharides. Among 10 species screened for production of EPS in
submerged cultures with glucose, soy oil and yeast extract, the best results were obtained with Ganoderma
lucidum (0.79 g/L EPS) and Pleurotus ostreatus (0.75 g/L). Agitation strongly improved EPS production
in most of the studied strains. Eight out of ten species assayed could successfully developed basidiomes on
synthetic “bag-log” cultivation on a substrate consisting of oak sawdust and corn bran. This work
describes for the first time the environmental factors required for fruiting under such conditions for four
species: Schizophyllum commune, Coriolus versicolor, Ganoderma applanatum and Trametes trogii. IPS
were extracted from the carpophores. The content of IPS in the carpophores varied from 1.4% (G.
applanatum) up to 5.5% and 6% in G. lucidum and Grifola frondosa, respectively.
Keywords: basidiomycetes, submerged cultivation, solid state fermentation, exopolysaccharides,
fructification, endopolysaccharides
1. Introduction
Polysaccharides are industrially used as thickeners, stabilizers and gelling agents in food products.
They are also employed as depolluting agents and there is also a growing interest in their biological
functions like antitumor, antioxidant or prebiotic activities (Liu et al., 1996b). They are derived from a
wide variety of sources: bacterial, fungi, algae and plants. Despite the many sources of polysaccharides,
the world market is dominated by polysaccharides from algae and higher plants. Exopolysaccharides
(EPS) production from microorganisms, including bacteria, yeasts and moulds, demands shorter periods of
time. EPS produced by microorganisms represent an industrially untapped market (Donot et al., 2012).
Production of EPS is widely distributed among fungi (Papinutti, 2010). They fulfill different tasks
during the growth on natural substrates, such as adhesion to surfaces, immobilization of secreted enzymes,
prevention of hyphae from dehydration, and increased residence time of nutrients inside the mucilage
(Bolla et al., 2010). Interest has concentrated on polysaccharides produced by mushrooms, because of
their various biological and pharmacological activities. These include immunostimulating, antitumor,
antibacterial, antiviral, hypocholesterolemic, antioxidant and hypoglycemic activities, vitality and
performance enhancement effects and beneficial cosmetic effects on skin (Ooi y Liu, 2000b; Lee et al.,
2007a). Mushroom polysaccharides, as lentinan and schizophyllan, among others, are mainly glucans with
diferent structures. β-D-glucans are the most common polysaccharides of basidiomycetes having a

87
branched structure, varied degree of substitution, containing β(1→3) linkages in the main chain and
additional β(1→6) branch points (Amaral et al., 2008). To obtain bioactive polysaccharides from
mushrooms, different authors have cultivated edible or medicinal mushrooms on solid media (for fruiting
body production) or in submerged cultures. In addition to EPS excreted into the fermentation broth,
mycelial cultures also produce internal polysaccharides localized within the mycelia (endo or intracellular
polysaccharides, IPS). Submerged cultures of basidiomycetes can be used to produce polysaccharides in a
shorter time, when compared to the production and extraction process of mushroom fruiting bodies. This
encouraged many authors to develop a variety of culture media to optimize the production of these
polymers (Komura et al., 2010). Most reports have focused on EPS, but not on IPS (Lee et al., 2007a).
The IPS can be extracted from fruiting bodies and their mycelia by solid-liquid extraction using heated
water or mixtures of organic solvents (Komura et al., 2010). As the demand for fruiting bodies and/or
mycelium biomass is constantly increasing, artificial cultivation has become essential. Solid-state
cultivation is one way of meeting the rising demand for fungal mycelium and its bioactive metabolites
(Švagelj et al., 2008). In the search of new sources of fungal polysaccharides, the main goal of this
research was to screen the ability of several species of basidiomycetes for the production of EPS under
submerged cultivation and to evaluate the IPS content in the fruiting bodies produced by these fungi under
solid-state cultivation.
2. Materials and methods
2.1 Microorganisms
The fungi studied in this work were selected according to their commercial potential and their
attributes. Thus, fungi like Grifola frondosa, Lentinula edodes, Ganoderma lucidum and others are
becoming better known and their markets are growing because of their active components and usefulness.
On the other hand, fungi like Schyzophyllum commune, Ganoderma applanatum and Trametes trogii have
been studied by different researchers, which have found that their components are also important for
several segments of the industry, although their cultures have not been disseminated. In this way, they
represent both scientific and industrial interest.
Ten species of basidiomycetes, were screened: Lentinula edodes (CICL54), Schizophyllum commune
(UCS004), Trametes trogii (BAFC463), Coriolus versicolor (PSUWC430), Pycnoporus sanguineus
(UCS003), Ganoderma applanatum (UCF001), Ganoderma lucidum (UCC002), Grifola frondosa
(PSUMCC 922), Pleurotus ostreatus (UCC001) and Auricularia delicata (UCS002). Pure cultures are
deposited at Culture Collection of Macrofungi at University of Caldas (Manizales, Colombia). The species
were maintained on potato dextrose agar (PDA) at 4°C with periodic transfer.
2.2. Culture medium for production of EPS under submerged fermentation
Inocula consisted of a 50-mm2 surface agar plug from a 10-day-old culture grown on PDA.
Cultures were performed in 100 mL Erlenmeyer flasks containing 25 mL of liquid medium. The medium
consisted of: 1 g (NH4)2SO4, 1 g KH2PO4, 0,5 g MgSO4.7H2O, 0,1 g CaCl2, 30 g glucose, 14.15 g soy oil,
2 g yeast extract and distilled water up to 1 L. Initial pH of the medium was 5.5. The medium was
autoclaved at 121°C for 15 min. Incubation was carried out at 25°C. Static as well as stirring conditions
(100 rpm) were assayed. Cultures were harvested after 14 and 21 days. Six replicas of all the experiments
were performed.

88
2.3. EPS measurement
Cultures were harvested over time and mycelium was separated from the culture broth by vacuum
filtration using a pre-weighed glass microfiber filter (Whatman GF/F, Maidstone, England). To estimate
growth, mycelial weight was determined after drying overnight at 80ºC the mycelial mats obtained from
filtration. For both water- and alkali-soluble EPS determination, EPS were precipitated from the resulting
culture filtrate (0.5 mL mixed with 4 times the volume of ethanol 96%, during 45 min) and then separated
by centrifugation at 14000 rpm for 15 min. The insoluble components were re-suspended in 0.5 mL NaOH
1 M at 60 ºC for 1 h. After EPS solubilization, samples of 5-50 µL were taken (proper dilutions in distilled
water were done when necessary), the volume was completed to 0.5 mL with distilled water, then 0.5 mL
of 5% (w/v) phenol were added and the mixture was shaken thoroughly. Three mL of 98% sulphuric acid
were also added and stirred vigorously. The tubes were allowed to stand 30 min at room temperature. A
blank test was prepared by substituting the solubilised EPS with distilled water. Total carbohydrates were
determined measuring the absorbance at 490 nm (Dubois et al., 1956). The EPS content of supernatants
was calculated subtracting the amount of alcohol-insoluble carbohydrates from un-inoculated media. EPS
yields were expressed as grams of EPS per litre of culture medium (g/L) and specific yields as milligrams
of EPS per gram dry mycelium (mg/g mycelium).
2.4. Fruit body production
Spawn of all the species evaluated with the exception of G. frondosa was prepared on wheat
grains (40% moisture content) according to Chang and Miles (Chang y Miles, 2004). G. frondosa was
inoculated on corn grains according to Montoya et al. (Montoya et al., 2008b). The substrate consisted of
(dry weight basis) 73% oak sawdust (25% moisture content), 23% corn bran (13% moisture content), 2%
sucrose and 2% calcium carbonate. The substrates were packed in polypropylene bags and tyndallized.
The moisture content was adjusted according to the development requirements of each species L. edodes
(55%), S. commune (60%), T. trogii (65%), C. versicolor (60%), G. applanatum (65%), G. lucidum
(60%), G. frondosa (58%) and P. ostreatus (70%) (Chang y Quimio, 1982; Jaramillo et al., 1999;
Rodríguez, 2003a; Chang y Miles, 2004; Rodríguez y Jaramillo, 2005c; Montoya et al., 2008b; Montoya
et al., 2009a). Each bag of 20 cm diameter and 30 cm height contained 1 kg of substrate. One square hole
of 2.54 cm2 was made at the top of each bag and covered with a microporous breather strip to allow for
gas exchange. The bags containing the substrate were aseptically inoculated with 4% (wet basis) of spawn.
2.5. IPS measurement
Carpophores were dehydrated in a Deni® dehydrator tray, with forced convection of hot air, at a
temperature between 55 and 60°C up to 5% moisture content. Subsequently, the dried fruit bodies were
milled with a knife mill Tecator® (model Cyclotec 1093) to achieve particles size mesh 180-200. For IPS
determination, 1 g of dry powder (5% moisture content) was mixed with 50 mL of 80% (v/v) ethanol, and
extracted with an ultrasound-assisted extractor (Elmasonic E Elma®) for 30 min with an ulterior 20 h rest
contact between the fungus and the solvent. The extract was filtrated at reduced pressure by using a pre-
weighed glass microfiber filter (Whatman GF/F, Maidstone, England) and then centrifuged for 15 min at
14000 rpm. The precipitate was suspended in 1M NaOH solution at 60°C for 1h. Total carbohydrates were
determined in this suspension by the phenol-sulfuric method (Dubois et al., 1956).

89
2.6. Statistical analysis of data
To select the best EPS producing species among the evaluated ones, a multifactorial design with 3
factors was performed. The factors were: ten species, incubation day (14 or 21) and stirring conditions
(100 rpm or none). The response variable was the total carbohydrate concentration in g EPS/L. The data
presented are the average of the results of at least 3 replicates with a standard error of less than 5%.
Analysis of variance (ANOVA) and repeated measures analysis were tested by the software Statistica v. 7.
The significant differences between treatments were compared by Tukey’s test at 5% level of probability.
3. Results and discussion
3.1. Production of EPS in submerged cultivation
EPS production in submerged cultivation under stirred and static conditions was evaluated after 14
and 21 days of incubation in 10 species of basidiomycetes. Results are summarized in Table 1. The
ANOVA showed that the species and incubation conditions influenced EPS production while the
incubation time was not a significant factor in their EPS production. Among the species assayed, best
results were obtained with G. lucidum at 21 days of incubation under agitation (0.79 g/L of EPS) and P.
ostreatus (0.75 g/L) under the same conditions. Tukey’s test indicated that G. lucidum, produced higher
EPS concentrations on average than A. delicata, P. sanguineus, L. edodes, S. commune, G. frondosa, T.
trogii and G. applanatum, but no significant differences were obtained with P. ostreatus and C. versicolor
EPS production. Agitation strongly improved EPS production in most of the studied species. For example,
L. edodes and P. ostreatus EPS production at 21 days under static conditions was around 30% of that
produced under agitation. Nevertheless, for some of the species tested (T. trogii, G. applanatum, G.
frondosa and A. delicata) agitation and incubation time were not significant factors in EPS production.
Differences between EPS concentrations obtained under static and agitated conditions in both incubation
periods did not exceed 15%. Although the statistical analysis of experimental data showed no significant
differences in the incubation period, differences were observed in EPS production in some of the species
tracked, as was the case of L. edodes without stirring, which decreased from 0.21 g/L EPS at 14 days of
incubation down to 0.08 g/L after 21 days, T. trogii which rendered 0.28 g/L under agitated conditions
after 14 days but only 0.16 g/L at 21 days or P. sanguineus whose EPS production increased under static
conditions from 0.07 g/L at 14 days up to 0.20 g/L at 21 days. Stirring conditions may affect fungal
biomass production, and in turn, EPS production. Maintenance of high dissolved oxygen levels was
important for both cell growth and EPS formation in Tremella fuciformis (Cho et al., 2006). However,
Park et al. (Park et al., 2002a) reported an inhibitory effect of high aeration rates on EPS production by
Cordyceps militaris.
Submerged cultures showed different concentrations of EPS varying from 0.22 g/L to 0.79 g/L for
A. delicata and G. lucidum, respectively. Nevertheless culture medium and incubation conditions need to
be optimized for each strain. Komura et al. (Komura et al., 2010) recently compared EPS production
under submerged fermentation in various basidiomycetes and showed that EPS concentrations varied
between 0.24 and 0.91 g/L. Kim et al. (Kim et al., 2008b) examined the effect of different synthetic liquid
media on EPS production by shake flask cultures of 19 mushrooms. Best EPS production was attained by
G. lucidum and Phellinus linteus in potato malt peptone medium, with EPS concentrations of 1.17 and
1.52 g/L, respectively. Remarkably, high values were obtained recently by G. lucidum (15 g/L EPS) in an
optimized medium for EPS production under submerged fermentation (Papinutti, 2010). Undoubtedly, the
screening results shown in this work can be useful to address research efforts on optimization of culture
media for EPS production using different basidiomycetes.

90
Antrodia cinnamomea and G. frondosa exhibited EPS increase that paralleled growth response
(Bae et al., 2005; Lin y Sung, 2006). However, C. versicolor and Gloeophyllum trabeum showed an
inverse relationship between biomass and EPS production. Thus, low biomass production was associated
with high EPS production and vice versa. For this reason, specific EPS yield (mg EPS/g dry mycelium)
provides more information on how different culture conditions could influence the EPS enhancement
production (Papinutti, 2010). Table 1 also depicts specific EPS yields. The highest specific EPS yield was
attained by G. applanatum under static conditions after 21 days (204 mg/g). Specific EPS production
compares favorably to previous reports. A maximum of 139 mg EPS/g was obtained by G. lucidum in an
optimized medium for EPS production (Papinutti, 2010).
3.2. Basidiome development
For basidiome production of each 8 out of 10 species assayed (L. edodes, S. commune, T. trogii,
C. versicolor, G. applanatum, G. lucidum, G. frondosa and P. ostreatus) on synthetic “bag-log”
cultivation, several methods were modified and applied in this work (see Table 2). The morphological
properties of the carpophores obtained, are depicted in Figure 1 and Table 2. Figure 1 shows the fruit
bodies obtained from eight of ten isolates of study and Table 2 describes the details of how the
carpophores were obtained. Under the conditions tested neither A. delicata nor P. sanguineus developed
basidiomes. Although the environmental factors required for fruiting in synthetic “bag-log” cultivation of
some of the species screened in this work have already been described, such as for L. edodes and G.
lucidum (Chang y Miles, 2004), G. frondosa (Montoya et al., 2008b) and P. ostreatus (Rodríguez y
Jaramillo, 2005c), this work portrays for the first time the conditions required for synthetic “bag-log”
basididiome development for four species: S. commune, T. trogii, C. versicolor and G. applanatum (Table
2). As far as we know, only Badcock (Badcock, 1943) described a method for obtaining G. applanatum,
Polystictus versicolor (syn. of C. versicolor) and S. commune fructifications (after 11, 7 and 5.5 weeks,
respectively) in large tubes filled with a medium consisting of sawdust and various nutrients, and
Williams et al. (Williams et al., 1981) obtained morphologically typical fruit bodies of C. versicolor
within 8-12 weeks on cut logs of birch or hazel wood, placed with their bases submerged in water in
closed horticultural propagators.
3.3. Quantification of IPS in the basidiomes
The fruiting bodies obtained were subsequently subjected to drying and grinding for IPS
quantification. Figure 2 shows IPS content (percentage of the basidiome in dry basis). The content of IPS
in the carpophores varied from 1.4% (G. applanatum) to 5.5% and 6% of IPS in G. lucidum and G.
frondosa, respectively. Up to 4.5% of IPS were extracted by Lee et al. (Lee et al., 2007a) from the
mycelium of G. applanatum cultivated under submerged fermentation, 1.3% were obtained from
mycelium of G. frondosa grown in liquid cultures (Berovič et al., 2007), and 1.4% from its mycelium
grown under solid-state fermentation (Švagelj et al., 2008). Thus the results obtained, show the potential
of using some of the basidiomycetes evaluated in this work, in order to obtain significant amounts of IPS
from fruiting bodies.
4. Conclusions
This study contributes to expand the knowledge on exopolysaccharides production by
basidiomycetes, including scantily studied species. It also demonstrates that solid state basidiome

91
development under synthetic bag-log cultivation could be an alternative biotechnological process for
production and isolation of fungal intracellular polysaccharides.
5. Acknowledgements
The authors thank the Vice-Rectorate for Research and Graduate Studies of the Universidad de
Caldas (Colombia) for financial and material support. The authors also acknowledge the Universidad de
Buenos Aires and CONICET (Argentina) for scientific and material assistance.

92
Table 1. Exopolysaccharides (EPS) production under submerged cultivation in stirred and static conditions aSpecific exopolysaccaharides yield
bResults are the mean of six replicas of each experiment
Species
Shake flasks
14 days
Static conditions
14 days
Shake flasks
21 days
Static conditions
21 days
EPS
(g/L)
Biomass
(g/L)
sp EPSa
(mg/g)
EPS
(g/L)
Biomass
(g/L)
sp EPS
(mg/g)
EPS
(g/L)
Biomass
(g/L)
sp EPS
(mg/g)
EPS
(g/L)
Biomass
(g/L)
sp EPS
(mg/g)
L. edodes 0.19
b
1.92
96.35 0.21
1.73
118.50 0.28
3.41
82.35 0.08
2.03
41.38
S. commune 0.16
2.88
56.25 0.13
1.93
68.91 0.33
4.01
79.30 0.16
2.54
64.31
T. trogii 0.28
2.34
120.51 0.25
1.88
134.57 0.16
3.93
41.73 0.18
2.10
84.29
C. versicolor 0.35
2.98
116.11 0.25
2.32
109.05 0.59
4.67
126.12 0.24
2.12
112.74
P. sanguineus 0.24
1.87
128.34 0.07
1.23
56.91 0.19
3.05
61.31 0.20
1.98
100.51
G. lucidum 0.38
2.99
126.76 0.25
1.77
138.42 0.79
4.97
158.72 0.34
2.68
125.37
G.
applanatum 0.19
1.76
107.39 0.19
1.02
185.29 0.23
3.56
60.04 0.28
1.35
204.44
G. frondosa 0.17
2.54
65.75 0.17
1.45
120 0.24
4.38
55.71 0.28
3.01
93.36
P. ostreatus 0.48
2.97
160.61 0.19
2.00
96.5 0.75
4.87
154.62 0.24
3.77
64.72
A. delicata 0.15
1.54
98.70 0.13
0.98
131.63 0.22
2.85
78.60 0.19
1.79
107.26

93
Table 2. Development of basidiome on synthetic “bag-log” cultivation by several white rot fungi and environmental conditions applied
Species Spawn run (mycelial
growth)
Primordial formation Fruiting
development
Carpophores’ morphological properties
obtained in this work
Cultivation
methodology
adapted from:
L. edodes
25-28°C, darkness,
14.5% O2, 5.5% CO2,
60% RH, 50 days
Thermal shock at 8°C for 24 hs,
14.5% O2, 5.5% CO2, darkness; 7
days at 18-22°C, 800 luxes, 80%
RH
18% O2, 2.5% CO2,
18-20°C, 800 luxes,
80-85% RH, 7 days
Pileus 8-10 cm diameter, scaly, brown, with
well formed lamellae beige, stipe 5cm long, 1
cm in diameter
Chang & Miles
(2004)
S. commune 25-30°C, darkness, 60%
RH, 20 days
18-20°C, 18% O2, 2.5% CO2,
700-1000 luxes, 80-85% RH, 5
days
18-20°C, 18% O2,
2.5% CO2, 700-1000
luxes, 80-85% RH, 7
days
Fan-shaped pileus grayish-white, with white
lamellae, without stem, in clusters
Method developed
in this work
T. trogii
25-30°C, darkness, 11%
CO2, 9.5% O2, 60% RH,
30 days
18-20°C, 18% O2, 2.5% CO2,
400-500 luxes, 70-80% RH, 25
days
18-20°C, 18% O2,
2.5% CO2, 400-500
luxes, 70-80% RH, 7
days
Shelf-type pileus whitish yellow to honey,
corrugated and continues as vertical blinds, all
around the substrate block.
Method developed
in this work
C. versicolor
25-30°C, 17.2% O2, 7.3%
CO2 darkness, 60% RH,
30 days
18-20°C, 18% O2, 2.5% CO2,
500-800 luxes, 70-80% RH, 20
days
18-20°C, 18% O2,
2.5% CO2, 500-800
luxes, 70-80% RH, 14
days
Shelf-type pileus 4-6 cm wide showing typical
concentric zones of different colors light tan,
yellow to brown, individuals around the side of
the substrate block
Method developed
in this work
G. lucidum
27-30°C, darkness, 11%
CO2, 9.5% O2, 60% RH,
35 days
18-20°C, 18% O2, 2.5% CO2,
500-700 luxes, 70-75% RH, 40-
50 days
18-20°C, 18% O2,
2.5% CO2, 500-700
luxes, 70-80% RH, 15
days
Pileus 8-12 cm of diameter, shelf-type, light
yellow to red, with stipe 1-3 cm long, clustered
especially at the top of the substrate block
Chang & Miles
(2004)
G. applanatum
27-30°C, darkness, 8%
CO2, 12.5% O2, 60% RH,
25 days
18-20°C, 18% O2, 2.5% CO2,
300-500 luxes, 70% RH, 45-60
days
18-20°C, 18% O2,
2.5% CO2,300-500
luxes, 70-80% RH,
15 days
Pileus 4-7 cm de diámeter, shelf-type, beige to
dark brown, significant stipe, formed at the top
and around the substrate block
Method developed
in this work
G. frondosa
25°C, shadow, 14.5% O2,
5,5% CO2, 60% RH, 75
days
Thermal shock at 10°C for 24 hs
16-18°C, 19.5% O2, 0.7% CO2,
100-500 luxes, 70-80% RH, 7
days
16-18°C, 19,5% O2,
0,7% CO2, 100-500
luxes, 70-80% RH, 7
days
Pileus elongated, brown-dark gray to light
beige, mature fruiting clusters with overlapping
petals (caps and lateral stems) extending
outward resembling a cluster flower.
Montoya et al.
(2008a)
P. ostreatus
27-30°C, darkness, 9%
CO2, 11.5% O2, 60% RH,
15 days
18-20°C, 18% O2, 2.5% CO2,
700-1000 luxes, 90% RH, 5 days
18-20°C, 18% O2,
2.5% CO2, 700-1000
luxes, 90-95% RH, 7
days
Pileus 7-10 cm of diameter, oyster-shaped,
purplish to light beige, stipe 1-3 cm, gregarious.
Rodríguez &
Jaramillo (2005b);
Montoya et al.
(2009b)
RH: relative humidity

94
References
1. Liu F, Fung MC, V.E.C. O, Chang ST. Induction in the mouse of gene expression of
immunomodulating cytokines by mushroom polysaccharide-protein complexes. Life Sci
1996;58(1):795-803.
2. Donot F, Fontana A, Baccou J, Schorr-Galindo S. Microbial exopolysaccharides: Main examples of
synthesis, excretion, genetics and extraction. Carbohydr Polym 2012;87:951- 62.
3. Papinutti L. Effects of nutrients, pH and water potential on exopolysaccharides production by a
fungal strain belonging to Ganoderma lucidum complex. Bioresour Technol 2010;101:1941-6.
4. Bolla K, Gopinath B, Shaheen S, Charya M. Optimization of carbon and nitrogen sources of
submerged culture process for the production of mycelial biomass and exopolysaccharides by
Trametes versicolor. Int J Biotechnol Mol Biol Res 2010;1:15-21.
5. Lee WY, Park Y, Ahn JK, Ka KH, Park SY. Factors influencing the production of
endopolysaccharide and exopolysaccharide from Ganoderma applanatum. Enzyme Microb Technol
2007;40 249-54.
6. Ooi V, Liu F. Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Curr
Med Chem 2000;7:715-28.
7. Amaral A, Carbonero E, Simao R, Kadowaki M, Sassaki G, Osaku C, Gorin, PAJ Lacomini, M. An
unusual water-soluble B-glucan from the basidiocarp of the fungus Ganoderma resinaceum.
Carbohydr Polym 2008;72:473-8.
8. Komura DL, Ruthers AC, Carbonero ER, Alquini G, Rosa MC, Sassaki GL, et al. The origin of
mannans found in submerged culture of basidiomycetes. Carbohydr Polym 2010;79:1052-6.
9. Švagelj M, Berovič M, Boh B, Menard A, Simčič S, Wraber B. Solid-state cultivation of Grifola
frondosa (Dicks: Fr) S.F. Gray biomass and immunostimulatory effects of fungal intra- and
extracellular β-polysaccharides. New Biotechnol 2008;25:150-6.
10. Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of
sugars and related substances. Anal Chem Eng J 1956;28(3):350-6.
11. Chang ST, Miles PG. Mushrooms Cultivation, Nutritional Value, Medicinal effect, and Enviromental
Impact. Second ed. New York: CRC Press; 2004.
12. Montoya BS, Varon LM, Levin L. Effect of culture parameter on the production of the edible
mushroom Grifola frondosa (maitake) in tropical weathers. World J Microbiol Biotechnol
2008;24:1361-6.
13. Chang ST, Quimio TH. Tropical Mushrooms: Biological Nature and Cultivation Methods. T.H. QE,
editor. Hong Kong: Chinese University Press; 1982.
14. Jaramillo C, Rodriguez N, Gómez FA. Cultivo de hongos tropicales sobre residuos agroindustriales
presentes en la zona cafetera. Informe final experimento QIN-09-23 (Cultivation of tropical
mushrooms on agri-industrial wastes present in coffee-growing region. Final report of experiment
QIN-09-23, in Spanish). Chinchiná, Colombia: Cenicafé; 1999. p. 44.
15. Rodríguez N. Aprovechamiento de los residuos sólidos generados en el cultivo e industrialización del
café para la producción de hongos comestibles y medicinales (Utilization of solid wastes generated
during the cultivation and industrialization of coffee for production of edible and medicinal
mushrooms, in Spanish) [Ph.D. Thesis]. Valencia: Universidad Politécnica de Valencia; 2003.
16. Rodríguez N, Jaramillo C. Cultivo de hongos comestibles del género Pleurotus sobre residuos
agrícolas de la zona cafetera. Bol Téc Cenicafé 2005;23:34-40.
17. Montoya SB, Restrepo GM, Tabarez LA. Rendimientos en el cultivo de los hongos Pleurotus
ostreatus. Rev Inv UCM 2009;14:16-26.
18. Cho EJ, Oh JY, Chang HY, Yun JW. Production of exopolysaccharides by submerged mycelial
culture of a mushroom Tremella fuciformis. J Biotechnol 2006;127:129-40.
19. Park J-P, Kim S-W, Hwang H-J, Cho Y-J, Yun J-W. Stimulatory effect of plant oils and fatty acids
on the exo-biopolymer production in Cordyceps militaris. Enzyme Microb Technol 2002;31:250-5.

95
20. Kim TH, Taylor F, Hicks KB. Bioethanol production from barley hull using SAA (soaking in
aqueous ammonia) pretreatment. Bioresour Technol 2008;99:5694-702.
21. Bae J, Sim J, Lee B, Pyo H, Choe T, Yun J. Production of exopolysaccharide from mycelial culture
of Grifola frondosa and its inhibitory effect on matrix metalloproteinase-1 expression in UV-
irradiated human dermal fibroblasts. FEMS Microbiol Lett 2005;251:347-54.
22. Lin E, Sung S. Cultivating conditions influence exopolysaccharide production by the edible
Basidiomycete Antrodia cinnamomea in submerged culture. Int J Food Microbiol 2006;108:182-7.
23. Badcock E. Methods for obtaining fructifications of wood-rotting fungi in culture. Trans Br Mycol
Soc 1943;26:127-32.
24. Williams E, Todd N, Rayner A. Propagation and development of fruit bodies of Coriolus versicolor.
Trans Br Mycol Soc 1981;77:409-14.
25. Berovič M, Boh B, Wraber B, Pohleven F. Production of intra- and extracellular polysaccharides of
Grifola frondosa by solid state and submerged fermentation. Int J Med Mushrooms 2007;9:193-4.
26. Montoya BS, Varon LM, Levin L. Effect of culture parameter on the production of the edible
mushroom Grifola frondosa (maitake) in tropical weathers. World J Microbiol Biotechnol
2008;24:1361-6.
27. Rodríguez N, Jaramillo C. Cultivo de hongos comestibles del género Pleurotus spp. sobre residuos
agrícolas de la zona cafetera. Boletín Técnico Cenicafé 2005;27:54 pp.
28. Montoya SB, Restrepo GMF, Tabarez LAL. Rendimientos en el cultivo de los hongos Pleurotus
ostreatus. Revista de Investigaciones Universidad Católica de Manizales 2009;14:16-26.
Fig. 1. Fruiting body development under the conditions assayed in this work. (a) Grifola frondosa, (b)
Lentinus edodes, (c) Coriolus versicolor, (d) Ganoderma lucidum, (e) Schizophyllum commune, (f)
Pleurotus ostreatus, (g) Ganoderma applanatum, (h) Trametes trogii.

96
Fig. 2. Content of intracellular polysaccharides (IPS) in the carpophores (percentage in dry basis).

97
5. Selección de la Mejor Combinación Hongo –
Formulación para la Degradación de Sustratos
Lignocelulósicos mediante Fermentación en Estado
Sólido con Pleurotus ostreatus, Coriolus versicolor y
Lentinus edodes

98
Selección de la Mejor Combinación Hongo – Formulación para la
Degradación de Sustratos Lignocelulósicos mediante Fermentación en Estado
Sólido con Pleurotus ostreatus, Coriolus versicolor y Lentinus edodes
Sandra Montoya1, Óscar Julián Sánchez
2*, Laura Levin
3
RESUMEN
En esta investigación se realizó el cultivaron de tres especies de hongos de pudrición blanca: Coriolus
versicolor, Pleurotus ostreatus y Lentinus edodes sobre doce formulaciones de sustrato en estado sólido
empleando mezclas de diversos los materiales y sales carbonato de calcio y dos niveles de sulfato de cobre
(II). Los objetivos de este trabajo fueron i) evaluar los efectos de la relación C/N y del sulfato de cobre (II)
sobre las actividades enzimáticas endoglucanasa, exoglucanasa, endoxilanasa, β-glucosidasa, lacasa y
manganeso peroxidasa (MnP) durante 49 días de fermentación en estado sólido; así como el consumo de
celulosa y hemicelulosa y la degradación de lignina y ii) proponer un modelo matemático para la
descripción de la producción de biomasa, de las enzimas lignocelulolíticas, producción y consumo de
azúcares reductores, consumo de celulosa y hemicelulosa y degradación de lignina. Las tres especies de
hongos crecieron bien sobre todas las formulaciones de sustrato y se obtuvo respuesta en la medida de los
títulos de todas las actividades enzimáticas por combinación especie – fórmula. C. versicolor presentó la
mayor capacidad de degradación de lignina, celulosa y hemicelulosa para todas las combinaciones, con
65% como la máxima degradación de lignina para F1 y 43% de degradación de celulosa sobre F9. Se
propuso un modelo matemático compuesto de 11 ecuaciones diferenciales a fin de buscar la descripción
de los datos experimentales de todas las combinaciones resultantes para el C. versicolor, por haber sido el
hongo que mayor capacidad de degradación de los sustratos lignocelulósdicos mostró. En este trabajo se
presentan los resultados del modelamiento para la combinación especie de hongo fórmula (F9) de mejor
comportamiento respecto a la degradación de los sustratos basados en materiales lignocelulósicos.
Palabras clave: hongos de pudrición blanca, relación C/N, actividades lignocelulolíticas, degradación de
materiales lignocelulósicos.
1. INTRODUCCIÓN
Los hongos de pudrición blanca son hasta ahora los únicos organismos capaces de degradar
completamente todos los componentes de los materiales lignocelulósicos gracias a las enzimas
lignocelulolíticas que producen (Kirk y Farrel, 1987b). En general las enzimas degradadoras de
polisacáridos están sujetas a mecanismos de regulación de su síntesis. Es decir no se producen de modo
constante, sino que su síntesis es inducida por el sustrato adecuado y es reprimida por azúcares fácilmente
utilizables, en particular glucosa. El inductor más eficiente es el polímero-sustrato de las enzimas que
serán sintetizadas. Sin embargo, debido a su alto peso molecular, no son compuestos que puedan penetrar
en las células y ejercer su efecto (Béguin y Aubert, 1994). Algunas de las enzimas extracelulares que se
producen por los hongos son solubles y se encuentran libremente dispersas en la película de líquido que
rodea la hifa, pero otras están fijas en el espacio sujetas a la pared de la hifa, a la matriz extracelular o al
propio sustrato (Hudson, 1986a).
La producción de enzimas por hongos como Pleurotus ostreatus, Coriolus versicolor y Lentinus
edodes puede depender de diversos factores como las condiciones microambientales y nutricionales en el
sustrato que afecta la velocidad de crecimiento de cada una de las especies (Gutierrez-Correa y Tengerdy,
1997; Carabajal et al., 2012). La biodegradación de la celulosa es un proceso sinérgico que implica en
primera instancia las endoglucanasas y exocelobiohidrolasas, las cuales atacan las estructuras amorfa y
cristalina de la celulosa produciendo celotriosas y celobiosas que posteriormente son degradadas hasta

99
glucosa por la β-glucosidasa. La hidrólisis del xilano requiere principalmente de la acción de la
endoxilanasa y la β-xilosidasa, aunque para degradar completamente el xilano se requiere la acción de
otras enzimas para hidrolizar los xilanos sustituidos (Levin et al., 2008; Montoya et al., 2011a). La lignina
es degradada por la interacción de diversas enzimas extracelulares, principalmente fenoloxidasas (lacasas)
y peroxidasas (manganeso-peroxidasa (MnP) y lignina peroxidasa) (Martínez et al., 2005). Los
basidiomicetos de pudrición blanca se destacan por la producción de enzimas celulasas, xilanasas, lacasas
y MnP en procesos de fermentaciones en estado sólido utilizando sustratos lignocelulósicos. Se han
reportado perfiles de producción de enzimas lignocelulolíticas durante los ciclos de vida de cepas
comerciales de Grifola frondosa, P. ostreatus y L. edodes en fermentaciones en estado sólido mostrándose
que las actividades de las lacasas y MnP fueron altas al final de la fase de colonización y declinaron
rápidamente en las fases reproductivas. En contraste, las actividades enzimáticas de celulasas y xilanasas
se evidenciaron de manera medianamente estables durante la colonización para incrementarse durante la
fase reproductiva (Asatiani et al., 2008; Montoya et al., 2011a).
En el caso de los hongos de pudrición blanca cuando se cultivan en medios de composición
ligeramente diferente, se sintetizan diversas formas de una misma enzima (isoenzimas) que a pesar de
poseer propiedades catalíticas similares, difieren notablemente en sus características físico-químicas, por
lo que se deduce que hay formas múltiples que coexisten en determinado momento; también puede variar
la proporción de estas isoenzimas a lo largo del desarrollo del hongo (Carlile et al., 2001). La síntesis de
isoenzimas puede deberse a productos de genes diferentes, aunque también pueden deberse a cambios
postraduccionales como proteólisis, glicosilación o agregados cuaternarios. Se pueden sintetizar como
consecuencia de estados fisiológicos diferentes (composición del medio de cultivo, pH, temperatura, etc.)
a partir de genes regulados diferencialmente (Joselau y Ruel, 1994; Blanchette, 1995; Barr et al., 1996;
Carlile et al., 2001).
Reviste especial interés el modelamiento de la cinética de cultivo de los hongos de pudrición
blanca. Se han empleado varias expresiones matemáticas para la descripción de la producción de diversas
biosustancias utilizando hongos en procesos de fermentación. Los modelos más reportados son el modelo
lineal, logístico y, más recientemente, de dos fases (aceleración y desaceleración) (Sangsurasak et al.,
1996), aunque estos modelos no incluyen el efecto de la concentración de nutrientes sobre el desarrollo
(Ikasari y Mitchell, 2000; Mitchell et al., 2004; Van de Lagemaat y Pyle, 2005). Asimismo, para la
descripción de la producción de biomasa en diversos tipos de procesos de fermentación se utilizan las
expresiones tipo Monod y Logística modificada (Mitchell et al., 2004; Tavares et al., 2005). Las
expresiones matemáticas utilizadas para la descripción de la formación de productos dependen de qué
clase de metabolito se esté produciendo; generalmente, las expresiones más simples son para los
metabolitos primarios, ya que su formación se considera diractamente proporcional a la formación de la
biomasa celular teniéndose en cuenta los parámetros de matenimiento celular y crecimiento microbiano.
En el caso de los productos intermedios del metabolismo primario y productos del metabolismo
secundario, la descripción de su cinética de formación es más compleja que para los metabolitos
secundarios; no existen hasta el momento expresiones simples bien definidas que abarquen a todas las
sustancias pertenecientes a estos grupos. Actualmente no hay modelos matemáticos disponibles que
permitan describir la producción de biosustancias de hongos macromicetos, ni su crecimiento. Esta
dificultad se debe, probablemente, a que cada uno de estos hongos se ve afectado directamente por
factores intrínsecos y extrínsecos particulares como las características físicas y la composición química de
los medios de cultivo, la procedencia de las cepas, la influencia de las condiciones ambientales y el
crecimiento lento asociado a la gran mayoría de las especies de macromicetos en comparación con el
tiempo de crecimiento de los micromicetos. Asimismo, para el análisis de la formación de productos en
procesos de fermentación en estado sólido es necesario conocer varias de las sustancias que se forman
durante el crecimiento y desarrollo del organismo a fin de comprender el mecanismo de producción de las
sustancias de interés y lograr plantear expresiones matemáticas que permitan la descripción y
escalamiento de su proceso productivo.
El objetivo de este trabajo fue evaluar el efecto de la relación carbono nitrógeno y del sulfato de
cobre (II) sobre la producción de enzimas lignocelulolíticas por tres especies de hongos de pudrición

100
blanca (L. edodes, C. versicolor y P. ostreatus) empleando doce formulaciones diferentes de sustratos
lignocelulósicos. Estos hongos fueron seleccionados de entre diez hongos de pudrición blanca mediante
un rastreo basado en sus actividades enzimáticas celulolíticas, xilanolíticas y ligninolíticas, así como en su
capacidad de degradación de celulosa y lignina; los resultados de este rastreo se relacionan en el artículo
anterior. Igualmente este trabajo tuvo como objetivo adicional proponer y validar un modelo matemático
para la mejor combinación hallada especie de hongo - formulación a fin de realizar la descripción de la
cinética de crecimiento del organismo, la degradación de los materiales lignocelulósicos y la producción
de las enzimas lignocelulolíticas en condiciones de fermentación en estado sólido.
2. MATERIALES Y MÉTODOS
2.1. Microorganismos
Tres especies de basidiomicetos fueron empleados para la producción de enzimas
lignocelulolíticas en fermentación en estado sólido, tomadas de la colección de hongos de la Universidad
de Caldas (Manizales, Colombia): Lentinula edodes del Centro de Investigaciones del Café, Chinchiná,
Caldas, Colombia (CICL54), Coriolus versicolor de Pennsylvania State University, Estados Unidos de
Norte América (PSUWC430), y Pleurotus ostreatus de la Colección interna de la Universidad de Caldas
(UCC001). Las cepas fueron extendidas sobre agar papa dextrosa (PDA) y mantenidas en refrigeración a
4°C y transferencias periódicas.
2.2. Producción de semilla
La semilla para la inoculación de los medios de cultivo sólidos se realizó sobre trigo lavado,
hidratado con agua caliente hasta 40% de humedad. El trigo hidratado fue empacado en unidades de 1 kg
en bolsas de polipropileno biorientado, provistas con un filtro de interlón en el primer tercio de la parte
superior de la bolsa de una pulgada de lado. Posteriormente, el grano fue esterilizado a 121°C por 30
minutos aplicando el método Thindal. Finalmente el grano hidratado y esterilizado fue inoculado a una
tasa del 4% en base húmeda con cada especie de hongo en cabina de flujo laminar y llevadas a incubación
por 12 a 15 días a 25°C y penumbra hasta colonización completa.
2.3. Medios de cultivo
Se realizaron doce formulaciones de sustrato por triplicado. Los sustratos fueron formulados en
base seca, con 40% de aserrín de roble, 20% de capacho de coco y 2% de aceite de soya como materias
primas fijas en todas las formulaciones y variando los porcentajes de cascarilla de café y salvado de maíz
a fin de modificar la relación carbono nitrógeno (R C/N) (ver Tabla 1) y dos niveles de sulfato de cobre:
0,1596% para las fórmulas impares y 0,0798% para las fórmulas pares. Todos los sustratos fueron
formulados a 60% de humedad. Los sustratos fueron empacados en bolsas de polipropileno biorientado en
unidades de 200 g cada una y esterilizados a 121°C por 30 min aplicando el método Thindal.
Posteriormente fueron inoculados en cabina de flujo laminar con 4% de inóculo en base húmeda respecto
al sustrato. Éstos fueron llevados a incubación a 25°C en penumbra por 49 días. Se tomaron 15 muestras,
dos por semana, tomándose la primera muestra el día de la inoculación.
Tabla 1. Relaciones carbono nitrógeno (R C/N) en doce niveles (formulaciones)
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 F11 F12
142,640 91,77 114,61 93,64 100,92 91,79 85,34 84,87 55,11 51,55 114,55 111,05

101
2.4. Determinación de la relación carbono nitrógeno
La relación carbono nitrógeno (C/N) fue determinada a partir de la medida de la materia orgánica
de las doce formulaciones de sustratos empleando el método de Walkley y Black (1934), el cual utiliza
ácido sulfúrico concentrado y solución de dicromato de potasio. La muestra se lee a 585 nm. La cantidad
de carbono total corresponde al 58% de la materia orgánica. El nitrógeno orgánico total fue determinado
por el método de Kjeldahl (Kjeldahl, 1883).
2.5. Determinación cuantitativa de actividades enzimáticas
Los extractos para la determinación de las actividades enzimáticas (celulolíticas y ligninolíticas)
fueron obtenidos de 1 g de sustrato fresco en 12 mL de agua destilada estéril neutra sometido a ultrasonido
por 5 minutos y agitación por 10 minutos con posterior filtración y centrifugación.
2.5.2. Actividades celulolíticas y xilanolíticas
Endo-1,4-ß-D-glucanasa (E.C.3.2.1.4). Se usó carboximetil celulosa (CMC) como sustrato al
0,5% en buffer acetato de sodio pH 4,8 en reacción con 100 µL de extracto enzimático por 30 minutos a
50°C. La reacción se detuvo adicionando el reactivo DNS (ácido dinitrosalicílico) (Miller, 1959). Se
continuó con la reacción de DNS para la determinación de azúcares reductores, se leyó la absorbancia a
longitud de onda 540 nm. Una unidad de actividad enzimática (UE) fue definida como la cantidad de
enzima que produce 1 µmol de azúcares reductores en un minuto. Para cuantificar la actividad se elaboró
una curva patrón de azúcares reductores, con distintas cantidades de glucosa.
Exo-1,4-ß-D-glucanasa (E.C.3.2.1.91). Se usó celulosa cristalina 1% en buffer acetato de sodio pH
4,8 como sustrato, en reacción con 100 µL de extracto enzimático a 50°C por 60 minutos y agitación. La
reacción se detuvo adicionando el reactivo DNS. Se continuó la reacción de azúcares reductores por el
método DNS y se centrifugó antes de leer la absorbancia a longitud de onda 540 nm. Una unidad de
actividad enzimática (UE) fue definida como la cantidad de enzima que produce 1 µmol de azúcares
reductores en un minuto.
β-glucosidasa (E.C.3.2.1.21). La reacción se produce entre el sustrato de p-nitrofenil β-D-
glucopiranosido 0,02% en buffer acetato de sodio pH 4,8 con 100 µL de extracto enzimático a 50°C por
30 minutos. La reacción se detuvo adicionando solución buffer Clark y Lubs (pH 9,8) y se leyó la
absorbancia a longitud de onda 430 nm (ε430=18,5/mM cm) (Wood and Bhat, 1988). Una unidad de
actividad enzimática (UE) fue definida como la cantidad de enzima que produce 1 µmol de producto (p-
nitrofenol) en un minuto. Para cuantificar la actividad se confeccionó una curva patrón con p-nitrofenol.
Endo-1,4-ß-D-xilanasa (E.C.3.2.1.8). La reacción se produce entre el sustrato xilano al 0,2% en
buffer acetato de sodio pH 4,8 con 100 µL de extracto enzimático a 50°C por 30 minutos. La reacción se
detuvo adicionando DNS y se leyó la absorbancia a longitud de onda 540 nm. Una unidad de actividad
enzimática (UE) fue definida como la producción de 1 µmol de azúcares reductores en un minuto. La
curva patrón de azúcares reductores se confeccionó en este caso con xilosa.
2.5.3. Actividades ligninolíticas
Lacasa (E.C.1.10.3.2): Se utilizaron 0,5 mM de ABTS en solución 0,1M de buffer acetato de
sodio pH 3,6 como sustrato, según método de Paszczynski and Crawford (1991), leyendo el aumento de
absorbancia a 420 nm (ε420=36/mM cm) luego de tres minutos de reacción a 30°C. Una unidad de
actividad lacasa (UE) fue definida como la cantidad de enzima requerida para oxidar 1 μmol de ABTS en
1 minuto.
Manganeso peroxidasa (E.C.1.11.1.13) (MnP): Se utilizó como sustrato una solución 0,01% de
rojo fenol en buffer succinato de sodio 0,1 M y pH 4,5 y sulfato de manganeso (0,22 g/L), con peróxido de
hidrógeno 0,2 mM como iniciador de la reacción. La reacción fue detenida adicionando NaOH 5N

102
después de 10 minutos de reacción para leer el incremento de absorbancia a 610 nm (ε610=22/mM cm).
Una unidad de actividad enzimática (UE) fue definida como la cantidad de enzima necesaria para oxidar 1
μmol de rojo fenol en 1 minuto (Paszczczynski et al., 1988).
2.6. Cuantificación del contenido de N-acetil-D-glucosamina (NAGA) y biomasa
Las muestras de sustrato fueron colectadas en 15 tiempos de incubación, dos por semana, hasta el
día 49 de incubación para las tres especies de trabajo. El sustrato sólido incubado a diferentes tiempos fue
secado en estufa a 101°C hasta peso constante, molido y almacenado en lugar fresco y seco para la
determinación del contenido de quitina. El contenido de biomasa fúngica en los sustratos sólidos fue
indirectamente estimado por la determinación del contenido de N-acetil-D-glucosamina (NAGA) después
de la hidrólisis con HCl 6N grado analítico según el método de Plassard et al. (1982). Para ello se
cuantificó paralelamente el contenido de NAGA por gr de micelio en cada una de las tres especies,
cultivándolas en medio líquido. El contenido de NAGA del micelio de cada una de las especies de hongos
de trabajo: C. versicolor, P. ostreatus y L. edodes fue determinado del desarrollo del micelio en medio
líquido en matraces de 250 mL con 100 mL de medio de cultivo con la siguiente composición: glucosa (30
g/L), extracto de levadura (6 g/L), SO4Mg.5H2O (0,5 g/L), K2HPO4 (0,5 g/L) y CaCl2 (0,1 g/L). El tiempo
de fermentación para el crecimiento del hongo fue de 25 días y sometidos a 100 rpm de agitación. Los
compuestos solubles en agua fueron extraídos durante la hidrólisis con HCl 6N por tres horas.
2.7. Cuantificación de los componentes de la fibra y azúcares reductores
Se determinaron los componentes de la fibra (celulosa, hemicelulosa y lignina) de los sustratos de
cada unad e las formulaciones en 15 tiempos durante los 49 días de incubación, así como la fracción
soluble, empleando los resultados de la determinación de fibras detergente neutra, ácida y lignino-ácida.
La medición se realizó a una muestra seca y molida de sustrato; dicha muestra se sometió a tres hidrólisis
en serie (cada una de 75 min): i) hidrólisis con lauril sulfato de sodio y otros, ii) hidrólisis en solución de
bromuro de amonio en ácido sulfúrico 1N, y iii) hidrólisis con ácido sulfúrico al 72% (p/v). Al finalizar
cada hidrólisis, las muestras fueron lavadas y secadas a 105°C hasta peso constante (Leterme, 2010a). El
contenido de azúcares reductores como glucosa fue determinado por el método del ácido dinitrosalicílico
(DNS) (Miller, 1959).
2.7. Diseño experimental
Se planteó un diseño experimental bifactorial con la especie a tres niveles y doce formulaciones de
sustrato. Las formulaciones varían en su relación C/N y la concentración de sulfato de cobre (II). Las
variables independientes en este diseño experimental fueron la especie de hongo, la formulación del
sustrato y el tiempo de fermentación. Las variables de respuesta medidas fueron las actividades
enzimáticas: endoglucanasa, Exoglucanasa, β-glucosidasa, lacasa, manganeso peroxidasa y endoxilanasa a
fin de buscar la mejor combinación especie de hongo fórmula para la producción de celulasas y de
ligninasas en términos de la capacidad de degradación de los componentes de la fibra, celulosa,
hemicelulosa y lignina en 15 tiempos durante 49 días de fermentación.
2.6. Modelamiento matemático del crecimiento del hongo y la producción de enzimas
Para la solución de los modelos matemáticos del proceso de fermentación en estado sólido se
utilizó el software Matlab® 2010b (MathWorks, EUA) en un PC de 3 GB de memoria RAM y un
procesador Intel CORE-i3 de 2,13 GHz. Para los modelos matemáticos planteados en ecuaciones
diferenciales ordinarias se utilizó el comando ode45 que está basado en una fórmula explícita de Runge-
Kutta (4,5) usando un par de Dormand-Prince (Dormand y Prince, 1980) y el comando ode15s basado en
una fórmula de orden variable que funciona mediante fórmulas de diferenciación numérica.

103
2.8. Análisis estadístico
Para todos los análisis estadísticos del presente trabajo se utilizó el software Matlab ® 2010b. Este
análisis se realizó en tres etapas. Primero se realizó un ANOVA a todos los datos del diseño experimental
y sus posibles combinaciones: especie, fórmula y tiempo de fermentación mediante el comando anovan.
Luego se realizó un análisis comparativo (Kruskal Wallis) de la capacidad de degradación de celulosa,
hemicelulosa y lignina para cada especie de hongo estudiado y las doce formulaciones de sustratos
mediante los comandos kruskalwallis y multcompare. A los máximos obtenidos de cada especie se les
realizó un nuevo análisis comparativo a fin de encontrar la combinación de mejor desempeño con los
porcentajes de degradación de la fibra al final del período de fermentación (49 días) mediante los
comandos kruskalwallis y multcompare.
Para evaluar si el modelo matemático propuesto describe adecuadamente los datos
experimentales, se realizó una prueba F de una cola que parte de las siguientes hipótesis:
2
,
0 2
exp,
2
,
1 2
exp,
: 1
: 1
i
i
i
i
residuales F
datos F
residuales F
datos F
SH
S
SH
S
Donde: Fi está dado para i = 1,2,…,12, s2
residuales es la varianza de los residuales (desviaciones de los datos
experimentales con respecto a los valores calculados por el modelo matemático) y se calcula para cada
variable de respuesta medida y s2datos exp es la varianza de la serie experimental de cada variable de
respuesta medida. Para estos análisis se utilizaron los comandos vartest y vartest2 de Matlab. Con las
varianzas de los residuales obtenidos para cada serie experimental se calcularon los porcentajes de cada
una respecto a la serie que dio la mayor varianza de residuales a fin de mostrar el ajuste de los datos
experimentales al modelo matemático propuesto de forma más simple.
3. RESULTADOS Y DISCUSIÓN
3.1. Efecto de la relación C/N sobre la degradación de los residuos lignocelulósicos y la producción
de enzimas lignocelulolíticas
Las tres especies de hongos (C. versicolor, P. ostreatus y L. edodes) probadas en el diseño
experimental, fueron inoculadas e incubadas bajo las mismas condiciones ambientales en las doce
formulaciones de sustrato. El intervalo de la relación C/N en las doce formulaciones de sustrato evaluadas
fue de 50 a 140 con una variación en los contenidos de nitrógeno de 0,29 a 0,8% en base seca. Las tres
especies de hongos se desarrollaron bien en todas las formulaciones evaluadas y produjeron actividades
enzimáticas significativas en todas las relaciones C/N durante los 49 días de incubación, como se observa
en la Tabla 2. Todos los contenidos de nitrógeno de las formulaciones estimularon el crecimiento de los
macromicetos de trabajo durante la fase vegetativa, incrementando el contenido de biomasa (ver Figuras 1
y 2) en todos los casos desde el inicio de la fase de incubación. Empero se debe tener en cuenta que la
variación del contenido de nitrógeno y de la fuente de carbono se garantizó empleando los mismos
materiales y no modificando las fuentes de nitrógeno y de carbono. Sin embargo, sí hubo variabilidad
sobre las actividades enzimáticas medidas (endoglucanasa, exoglucanasa, β-glucosidasa, endoxilanasa,
lacasa y MnP), dependiendo de la especie y a diferentes tiempos de incubación como se observa en la
Tabla 2. Las actividades enzimáticas máximas fueron seleccionadas realizando análisis estadístico por
etapas. Primero se realizó un ANOVA a fin de determinar si hubo diferencias significativas entre los
resultados obtenidos respecto a las variables independientes especie, fórmula y tiempo; posteriormente, se
realizó una selección de los máximos de las actividades enzimáticas gráficamente. La mayor actividad de

104
β-glucosidasa fue detectada para L. edodes – F2 con 84,01 U/g ss (sustrato sólido) a los 14 de incubación.
En comparación con las actividades obtenidas de esta misma enzima para las otras dos especies de hongos
existe una gran diferencia con los datos obtenidos para P. ostreatus, siendo mucho menor la diferencia con
las registradas para C. versicolor - F5, cuyo valor máximo fue de 71,05 U/g ss. Para la enzima
endoglucanasa la mayor actividad fue detectada con P. ostreatus – F1 al día 11 de la fermentación con
128,75 U/g ss, seguida de 78,75 U/g ss para C. versicolor – F10 en el día 11 de la fermentación. Los
niveles más altos de la enzima exoglucanasa se obtuvieron para L. edodes – F6 y F5 con 106,44 U/g ss
(día 21) y 99,29 U/g ss (día 32) respectivamente; mientras que, la menor actividad de exoglucanasa
obtenida fue para P. ostreatus – F12 con 44,05 U/g ss (día 35). Respecto a la enzima endoxilanasa, la
mayor actividad obtenida se reporta para C. versicolor – F9 con 124,18 U/g ss en el día 21 de
fermentación, con el menor valor obtenido para P. ostreatus – F12 al día 28 de fermentación. Las
actividades enzimáticas máximas de lacasa y MnP fueron para C. versicolor – F7 y P. ostreatus – F5 con
106,76 U/g ss y 9,77 U/g ss respectivamente y ambas en el día 28 de la fermentación. Según los datos
obtenidos de las actividades enzimáticas lignocelulolíticas para las 12 formulaciones evaluadas, no hay un
aparente efecto directo de la relación C/N sobre la variación de dichas actividades enzimáticas,
probablemente porque los componentes del sustrato que provocan la variación de la relación C/N no se
modifican durante la formulación de los medios y los niveles evaluados no son suficientemente diferentes
para provocar un cambio significativo en estas actividades enzimáticas.
En este estudio se presentó la producción de cuatro actividades enzimáticas de hidrolasas y dos
actividades de ligninasas de tres especies de hongos de pudrición blanca, en fermentación en estado sólido
variando las mezclas de diversos materiales lignocelulósicos; entendiendo que para la bioconversión de
los materiales lignocelulósicos con hongos, las hidrolasas juegan un rol muy importante ya que son las que
proveen la fuente de energía a los microorganismos; por lo que la baja producción de estas enzimas
pueden limitar la velocidad de degradación de los polisacáridos en azúcares solubles. En este trabajo los
datos de las actividades de endoglucanasa y endoxilanasa obtenidos fueron mucho menores que las
obtenidas por otros investigadores como Kachlishvili et al. (2006) con diferentes relaciones C/N. Aunque
los títulos de exoglucanasa, β-glucosidasa, lacasa y MnP obtenidos fueron mayores en comparación con
los obtenidos para diferentes relaciones C/N y otras especies de basidiomicetos de pudrición blanca por
otros autores (Tsiklauri et al., 1999; Baldrian y Gabriel, 2002; Reddy et al., 2003). De todos modos los
títulos enzimáticos obtenidos pueden variar de acuerdo a la eficiencia del método utilizado para extraer las
enzimas a partir del sustrato sólido, ya que la metodología empleada no necesariamente garantiza la
extracción de todas las enzimas adsorbidas al sustrato.
3.2. Efecto del sulfato de cobre sobre la degradación de residuos lignocelulósicos y la producción de
enzimas lignocelulolíticas
Los dos niveles de CuSO4 utilizados en las doce formulaciones del presente estudio no generaron
inhibición aparente en el crecimiento de los tres organismos de trabajo. Como se observa en las Figuras 1
y 2 la biomasa de los tres hongos aumentó en todas las formulaciones durante los 49 días de fermentación.
Las enzimas lignocelulolíticas fueron determinadas dos veces por semana con muestras destructivas a
partir de la elaboración de un extracto líquido al cual se le determinó siete actividades enzimáticas, cuatro
hidrolasas y dos óxido-reductasas. Se observa una gran variación en los máximos de las actividades
enzimáticas medidas entre especies y formulaciones como se observa en la Tabla 2.
Son muchos los factores que pueden afectar la producción de enzimas lignocelulolíticas en hongos
de pudrición blanca. Además, las diferentes cepas responden de un modo particular a cada uno de los
factores intrínsecos y extrínsecos que deben enfrentar cuando son inoculados. Debido a esto, el estudio
combinado de dos o más factores puede generar mayor dificultad al momento de la interpretación de las
posibles interacciones entre los compuestos que afectan directamente la producción de enzimas
lignocelulolíticas. La variabilidad de las actividades enzimáticas de las hidrolasas es tan amplia entre las
especies de trabajo y las doce formulaciones probadas que no hay una evidencia clara de que las
cantidades de CuSO4 (0,1596% y 0,0796%) adicionadas a los sustratos hayan generado alguna inhibición

105
o activación directa a estas actividades. Adicionalmente, no hay reportes de que la adición de cobre al
medio de cultivo sólido en las cantidades adicionadas genere algún efecto importante sobre las actividades
enzimáticas de las hidrolasas.
Tabla 2. Máximas actividades enzimáticas lignocelulolíticas (U/gss) de tres especies de hongos de pudrición blanca
para doce niveles de relación carbono nitrógeno (R C/N) y dos niveles de CuSO4
Fórmula F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 F11 F12
CuSO4 (% p/p) 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798
R C/N 142,6 91,8 114,6 93,6 100,9 91,8 85,3 84,9 55,1 51,6 114,6 111
βG (U/g ss)
P. ostreatus 29,92
(35)
22,30
(7)
47,32
(46)
26,20
(32)
25,63
(21)
37,02
(21)
17,68
(28)
35,21
(39)
33,92
(25)
32,64
(28)
19,58
(35)
36,31
(32)
C. versicolor 45,93
(11)
51,7
(11)
61,71
(39)
69,71
(35)
71,05
(32)
57,99
(28)
55,71
(25)
28,30
(14)
21,96
(25)
63,99
(28)
43,41
(21)
26,21
(32)
L. edodes 64,28
(14)
84,01
(14)
67,28
(42)
56,75
(28)
63,66
(39)
69,24
(35)
69,71
(25)
69,19
(28)
61,90
(35)
76,00
(25)
63,42
(28)
61,42
(32)
ENG (U/g ss)
P. ostreatus 128,75
(11)
37,46
(11)
76,71
(42)
23,71
(4)
18,78
(4)
34,81
(4)
17,31
(4)
39,19
(4)
43,25
(4)
42,73
(4)
7,37
(4)
16,10
(35)
C. versicolor 52,15
(11)
22,85
(4)
73,11
(7)
41,17
(4)
54,75
(4)
59,82
(4)
20,89
(4)
33,48
(4)
78,75
(11)
48,26
(4)
18,61
(4)
16,31
(39)
L. edodes 44,29
(11)
38,15
(21)
66,85
(11)
18,44
(4)
25,61
(4)
42,91
(4)
10,18
(28)
11,68
(32)
12,73
(4)
13,43
(7)
12,21
(32)
14,22
(42)
EXG (U/g ss)
P. ostreatus 58,55
(18)
73,96
(11)
91,34
(39)
86,30
(42)
90,89
(46)
74,49
(7)
69,37
(28)
62,35
(25)
62,48
(21)
61,70
(32)
50,41
(21)
44,05
(35)
C. versicolor 47,72
(46)
56,77
(49)
55,86
(49)
59,6
(32)
74,50
(7)
57,82
(7)
60,45
(28)
61,3
(28)
64,45
(32)
70,10
(32)
53,03
(28)
50,10
(25)
L. edodes 57,43
(11)
89,12
(18)
77,64
(21)
89,12
(21)
99,29
(32)
106,44
(21)
61, 5
(25)
74,75
(25)
76,52
(28)
77,37
(32)
57,76
(35)
53,03
(7)
ENX (U/g ss)
P. ostreatus 39,89
(18)
37,14
(18)
40,77
(28)
37,67
(18)
60,52
(21)
80,88
(21)
33,43
(11)
35,91
(25)
62,29
(14)
61,23
(21)
27,58
(35)
19,70
(28)
C. versicolor 66,10
(28)
62,82
(32)
50,43
(21)
96,20
(25)
90,00
(7)
59,46
(11)
45,47
(28)
41,13
(7)
124,18
(21)
67,87
(14)
38,56
(28)
64,33
(35)
L. edodes 56,62
(18)
70,70
(28)
68,31
(28)
95,58
(35)
70,79
(32)
109,11
(21)
66,54
(32)
94,52
(18)
83,45
(28)
107,53
(21)
71,85
(32)
70,7
(28)
Lac (U/g ss)
P. ostreatus 80,81
(18)
40,23
(32)
90,42
(28)
28,80
(25)
43,33
(25)
40,15
(21)
78,53
(21)
40,40
(18)
78,83
(18)
44,65
(11)
62,31
(21)
66,56
(25)
C. versicolor 80,29
(28)
40,15
(25)
83,03
(28)
106,10
(39)
45,52
(21)
63,26
(14)
106,76
(28)
57,57
(11)
104
(25)
70,90
(32)
101,97
(25)
67,22
(25)
L. edodes 40,05
(35)
30,66
(28)
38,05
(21)
26,61
(35)
39,28
(21)
52,57
(18)
38,50
(21)
32,43
(18)
26,86
(18)
45,19
(25)
47,25
(18)
32,89
(21)
MnP (U/g ss)
P. ostreatus 5,51 (25) 4,66
(49)
6,64
(21)
7,69
(42)
9,57
(28)
6,42
(42)
4,98
(21)
5,02
(35)
6,14
(32)
4,49
(14)
4,50
(25)
4,48
(32)
C. versicolor 4,87 (46) 4,56
(32)
7,36
(28)
6,63
(32)
6,09
(35)
6,29
(21)
5,44
(18)
3,78
(11)
5,44
(25)
5,22
(25)
5,21
(14)
4,73
(18)
L. edodes 4,16 (39) 4,48
(35)
5,64
(28)
6,92
(25)
4,95
(42)
6,01
(28)
4,58
(25)
4,76
(28)
5,14
(21)
4,86
(32)
4,73
(39)
4,73
(49)
Nota: Los números entre paréntesis corresponden al día de incubación en el que se determinó la máxima actividad enzimática
(valor-p<0.05)
En referencia a las enzimas ligninolíticas, el efecto del CuSO4 como inductor de lacasa y
peroxidasas si ha sido estudiado por varios investigadores (Levin et al., 2002; Niladevi y Prema, 2008). La
actividad de lacasa para las tres especies de hongos evaluadas y las doce formulaciones de sustrato
diferentes si pudo haberse afectado por la presencia del CuSO4, ya que las actividades enzimáticas
máximas obtenidas de lacasa son mayores que las obtenidas en otros trabajos donde se trabajó con hongos
de pudrición blanca y sustratos a bases de mezclas de residuos lignocelulósicos, como en el trabajo de
Montoya et al. (2011a) en el que las actividades de lacasa obtenida para el hongo de pudrición blanca
Grifola frondosa no fueron superiores a 15 U/g ss. Asimismo, en el presente estudio se obtuvieron
actividades de lacasa del orden de 90,42 U/g ss (valor-p<0,05) para P. ostreatus y de 106,76 U/g ss (valor-
p < 0,05) para C. versicolor a los 28 días de incubación, comparables con las obtenidas por Gassara et al.

106
(2010) para el hongo Phanerochaete chrysosporium en presencia de CuSO4 sobre diferentes sustratos a
base de diversos residuos (manzana, pulpa de papel, residuos de cerveceras y de pescaderías) y quienes no
obtuvieron actividad de lacasa cuando no utilizaron activadores. Adicionalmente, las actividades de lacasa
obtenidas en este estudio fueron también comparadas con las obtenidas por Rosale et al. (2007) donde se
presentó un incremento en la actividad de lacasa de Trametes hirsuta desarrollada sobre cáscara de naranja
con 1 mM de sulfato de cobre en cultivo sólido. Las lacasas pertenecen al grupo de oxidasas de cobre azul
y contienen cuatro átomos de cobre por molécula, distribuidos en tres diferentes sitios de unión de cobre
(Solomon et al., 1997); por lo tanto la presencia de cobre en los medios de cultivo siempre es importante
para la inducción y producción de las mismas. Además, es resaltable que con los hongos de pudrición
blanca se tiene una herramienta ambiental importante, ya que incrementando la actividad de lacasas
utilizando sulfato de cobre es posible utilizarlas en procesos de biorremediación de aguas residuales y
otros recalcitrantes, de modo que la concentración total de cobre en el medio es consumido o disminuido
por estos organismos durante el mismo proceso de biorremediación (Gassara et al., 2010).
Figura 1. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6
Figura 2. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12
En este trabajo las actividades de las enzimas MnP (ver Tabla 2) no mostraron una gran
variabilidad entre las especies y las formulaciones, las actividades obtenidas son comparables con las
reportadas en un trabajo previo (Montoya et al., 2011a) para G. frondosa sobre residuos lignocelulósicos
sin utilizar inductores o activadores y con los datos reportados por Gassara et al. (2010) cuando no
utilizaron activadores de peroxidasas. Varios investigadores han reportado efectos adversos sobre la
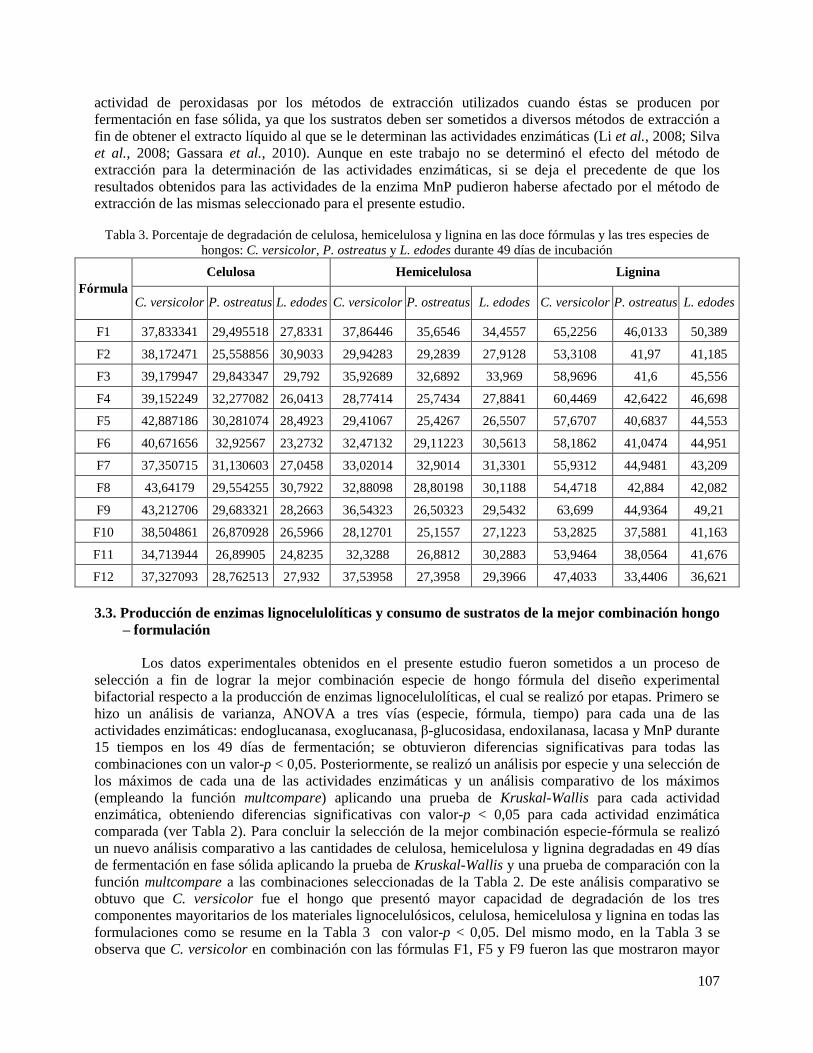
107
actividad de peroxidasas por los métodos de extracción utilizados cuando éstas se producen por
fermentación en fase sólida, ya que los sustratos deben ser sometidos a diversos métodos de extracción a
fin de obtener el extracto líquido al que se le determinan las actividades enzimáticas (Li et al., 2008; Silva
et al., 2008; Gassara et al., 2010). Aunque en este trabajo no se determinó el efecto del método de
extracción para la determinación de las actividades enzimáticas, si se deja el precedente de que los
resultados obtenidos para las actividades de la enzima MnP pudieron haberse afectado por el método de
extracción de las mismas seleccionado para el presente estudio.
Tabla 3. Porcentaje de degradación de celulosa, hemicelulosa y lignina en las doce fórmulas y las tres especies de
hongos: C. versicolor, P. ostreatus y L. edodes durante 49 días de incubación
Fórmula
Celulosa Hemicelulosa Lignina
C. versicolor P. ostreatus L. edodes C. versicolor P. ostreatus L. edodes C. versicolor P. ostreatus L. edodes
F1 37,833341 29,495518 27,8331 37,86446 35,6546 34,4557 65,2256 46,0133 50,389
F2 38,172471 25,558856 30,9033 29,94283 29,2839 27,9128 53,3108 41,97 41,185
F3 39,179947 29,843347 29,792 35,92689 32,6892 33,969 58,9696 41,6 45,556
F4 39,152249 32,277082 26,0413 28,77414 25,7434 27,8841 60,4469 42,6422 46,698
F5 42,887186 30,281074 28,4923 29,41067 25,4267 26,5507 57,6707 40,6837 44,553
F6 40,671656 32,92567 23,2732 32,47132 29,11223 30,5613 58,1862 41,0474 44,951
F7 37,350715 31,130603 27,0458 33,02014 32,9014 31,3301 55,9312 44,9481 43,209
F8 43,64179 29,554255 30,7922 32,88098 28,80198 30,1188 54,4718 42,884 42,082
F9 43,212706 29,683321 28,2663 36,54323 26,50323 29,5432 63,699 44,9364 49,21
F10 38,504861 26,870928 26,5966 28,12701 25,1557 27,1223 53,2825 37,5881 41,163
F11 34,713944 26,89905 24,8235 32,3288 26,8812 30,2883 53,9464 38,0564 41,676
F12 37,327093 28,762513 27,932 37,53958 27,3958 29,3966 47,4033 33,4406 36,621
3.3. Producción de enzimas lignocelulolíticas y consumo de sustratos de la mejor combinación hongo
– formulación
Los datos experimentales obtenidos en el presente estudio fueron sometidos a un proceso de
selección a fin de lograr la mejor combinación especie de hongo fórmula del diseño experimental
bifactorial respecto a la producción de enzimas lignocelulolíticas, el cual se realizó por etapas. Primero se
hizo un análisis de varianza, ANOVA a tres vías (especie, fórmula, tiempo) para cada una de las
actividades enzimáticas: endoglucanasa, exoglucanasa, β-glucosidasa, endoxilanasa, lacasa y MnP durante
15 tiempos en los 49 días de fermentación; se obtuvieron diferencias significativas para todas las
combinaciones con un valor-p < 0,05. Posteriormente, se realizó un análisis por especie y una selección de
los máximos de cada una de las actividades enzimáticas y un análisis comparativo de los máximos
(empleando la función multcompare) aplicando una prueba de Kruskal-Wallis para cada actividad
enzimática, obteniendo diferencias significativas con valor-p < 0,05 para cada actividad enzimática
comparada (ver Tabla 2). Para concluir la selección de la mejor combinación especie-fórmula se realizó
un nuevo análisis comparativo a las cantidades de celulosa, hemicelulosa y lignina degradadas en 49 días
de fermentación en fase sólida aplicando la prueba de Kruskal-Wallis y una prueba de comparación con la
función multcompare a las combinaciones seleccionadas de la Tabla 2. De este análisis comparativo se
obtuvo que C. versicolor fue el hongo que presentó mayor capacidad de degradación de los tres
componentes mayoritarios de los materiales lignocelulósicos, celulosa, hemicelulosa y lignina en todas las
formulaciones como se resume en la Tabla 3 con valor-p < 0,05. Del mismo modo, en la Tabla 3 se
observa que C. versicolor en combinación con las fórmulas F1, F5 y F9 fueron las que mostraron mayor

108
capacidad de degradación de los materiales lignocelulósicos, en los que la mejor combinación para
degradar celulosa y lignina fueron F1 y F9 y la que mejor degradó hemicelulosa fue la F9. A estas tres
combinaciones se les realizó un análisis comparativo a fin de seleccionar de entre las tres combinaciones
la de mejor comportamiento para aplicar un modelo matemático que describa los datos experimentales de
producción de enzimas lignocelulolíticas y biomasa y de consumo de los sustratos medidos. Sin embargo,
el análisis comparativo no presentó diferencia significativa (valor-p = 0.988), por lo que se seleccionó la
combinación C. versicolor – F9 ya que fue la que arrojó el mayor promedio de las medias.
3.4. Variación de las actividades enzimáticas durante la fermentación en fase sólida
La variación de las actividades enzimáticas durante los 49 días de fermentación en estado sólido
no presentó una tendencia definida para ninguna de las enzimas medidas para las tres especies de hongos
estudiados en las doce formulaciones de sustrato. Sin embargo, sí se conservó una tendencia general en los
datos experimentales obtenidos en este trabajo consistente en que todas las actividades de las enzimas
determinadas disminuyen luego de un tiempo específico; cada combinación arrojó un máximo de
actividad enzimática los cuales se consignan en la Tabla 2. Se hace la consideración que en adelante se
muestran solamente las figuras de la variación de las actividades enzimáticas medias. La actividad de las
enzimas celulolíticas medidas en este trabajo, endoglucanasa y exoglucanasa, fue variable sin tendencia
definida como se observa en las Figuras 3 y 4, pero en las que se perciben caídas de las actividades
enzimáticas luego del día 20 de incubación coincidiendo con el tiempo donde las tres especies han
terminado la colonización del sustrato. Sin embargo, algunas de las combinaciones (hongo-fórmula)
presentaron comportamiento diferentes, como es el caso de P. ostreatus – F3 donde la endoglucanasa
presenta una tendencia relativamente estable hasta el día 35 donde se observa que se inicia una alteración
de la actividad con ascenso y descenso hasta el término de 49 días de incubación. Asimismo, el
comportamiento de la exoglucanasa en F1 para las tres especies de hongos no presentó gran variación en
comparación con las demás combinaciones. De igual forma, el comportamiento de la actividad de la
endoglucanasa para las formulaciones F7 a F12 tampoco presentó una tendencia definida y si presentaron
descensos a partir del día 20 de incubación para las tres especies de trabajo. Mientras que la actividad de
la exoglucanasa para las formulaciones F7 a F12 presentó ascenso hasta el día 30 de incubación y una
tendencia al descenso a partir de allí hasta el final de las determinaciones, el día 49 de incubación. En
contraste, la actividad de la endoxilanasa se presenta un poco más dispersa que las dos hidrolasas
anteriores como se observa en la Figura 5, donde por ejemplo para la F5 es notable una gran dispersión de
los datos con actividades de hasta 90 U/gss. En referencia a la actividad de la β-glucosidasa, el
comportamiento de las combinaciones de cada uno de las especies de hongos entre las F7 y F12 se
observan en la Figura 6 en las que es notable la variación de los datos por combinación, pero si es marcada
la tendencia hacia un descenso de la actividad en todas las combinaciones. Las dos actividades de
ligninasas medidas en este trabajo mostraron comportamientos diferentes. La actividad de la lacasa varía
apreciablemente entre combinaciones como se puede observar en la Figura 7, dejando ver que el hongo C.
versicolor fue el que mayores títulos de esta actividad arrojó con actividades del orden de 80 a 100 U/gss
en varias de las combinaciones evaluadas como se consigna en la Tabla 2. Como se observa en la Figura 8
la variación de la actividad MnP fue escasa, y los títulos obtenidos de esta enzima fueron bajos durante los
49 días de incubación en la que fue determinada, como se consigna en la tabla 2.

109
Figura 3. Actividad enzimática de endoglucanasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 49 días de fermentación en estado
sólido
Muchos basidiomicetos tienen la capacidad de producir simultáneamente enzimas hidrolíticas y
oxidativas las cuales son necesarias para degradar los sustratos del complejo lignocelulósico (Kirk, 1983;
Wood y García, 1994). Estas enzimas son extracelulares e inducibles y según Buswell et al. (1996)
especialmente los hongos de pudrición blanca en general exhiben mayores títulos de ligninasas que de
celulasas; lo cual se convierte en una fortaleza de estos hongos ya que para lograr acceder a la celulosa
deben iniciar la degradación de la lignina. De hecho, según los datos experimentales obtenidos y
consignados en las Tablas 2 y 3 los datos de las actividades enzimáticas máximas de todas las
combinaciones y la capacidad de degradación de celulosa y lignina fueron obtenidos con las
combinaciones del hongo C. versicolor como el que más degradó lignina, por lo que probablemente debió
tener acceso más rápidamente a la celulosa, teniendo como resultado también una mayor degradación de
la celulosa. De forma que los títulos de las actividades enzimáticas obtenidas en este estudio indican que
los hongos de pudrición blanca como P. ostreatus, C. versicolor y L. edodes en investigaciones similares
obtuvieron resultados comparables, a pesar de la gran cantidad de factores que afectan los resultados de
las actividades enzimáticas, tanto de las hidrolasas como de las ligninasas (Mata y Savoie, 1998; Mata et
al., 2005; Gassara et al., 2010).
Figura 4. Actividad enzimática de exoglucanasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 49 días de fermentación en estado
sólido

110
Figura 5. Actividad enzimática de endoxilanasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 49 días de fermentación en estado
sólido
Figura 611. Actividad enzimática de β-glucosidasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 en 49 días de fermentación en
estado sólido
Figura 7. Actividad enzimática de lacasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 en 49 días de fermentación en estado sólido

111
Figura 8. Actividad enzimática de MnP de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 en 49 días de fermentación en estado sólido
Las actividades enzimáticas obtenidas presentaron descensos a diferentes tiempos del proceso de
fermentación en estado sólido, hecho que pudo atribuirse a aspectos como la eficiencia del método de
extracción empleado; a los cambios en la composición del sustrato durante el tiempo de fermentación, ya
que pueden producirse nuevas sustancias que inhiban las actividades enzimáticas; represión catabólica por
productos de las reacciones que se generan en la fermentación; producción de variantes isoenzimáticas de
las mismas enzimas a diferentes tiempos y diferentes condiciones del sustrato arrojan actividades
diferentes. Sin embargo, estos descensos en las actividades enzimáticas lignocelulolíticas también han
sido detectados por varios investigadores, quienes reportan algunas de estas hipótesis como posibles
causantes de los descensos de dichas actividades enzimáticas (Panagiotou et al., 2003; Mata et al., 2005;
Levin et al., 2008; Montoya et al., 2011a).
3.5. Modelamiento matemático
Basado en los datos experimentales obtenidos de actividades enzimáticas lignocelulolíticas
durante los 49 días de fermentación en estado sólido para las tres especies de hongos basidiomicetos de
pudrición blanca: C. versicolor, P. ostreatus y L. edodes, se propuso un modelo matemático compuesto de
11 ecuaciones diferenciales (ver Tabla 3) a fin de ajustar los datos experimentales de todas las
combinaciones resultantes para C. versicolor, ya que este fue el hongo que mayor capacidad de
degradación mostró de los materiales lignocelulósicos medidos en este trabajo (celulosa, hemicelulosa y
lignina). Sin embargo, aquí se presentan los resultados del modelamiento arrojados para la combinación
especie de hongo fórmula, F9, ya que ésta fue la que mostró el mejor comportamiento respecto de la
degradación de los sustratos basados en materiales lignocelulósicos. De igual forma, para comparar el
ajuste de los datos experimentales al modelo matemático propuesto se planteó una prueba F de una cola,
con la cual se obtuvieron las varianzas de los residuales entre los datos experimentales y el modelo,
individualmente para cada variable medida, pero haciendo la comparación de cada ecuación aplicada para
todas las formulaciones de F1 a F12 en la misma especies, en este caso C. versicolor, lo que permitió
probar la bondad del ajuste del modelo en términos de la mejor representación de los datos experimentales
obtenidos.
Con el modelo matemático propuesto se plantea la descripción de producción de biomasa, seis
enzimas lignocelulolíticas, azúcares reductores como producto intermedio consumido durante la misma
fermentación, el consumo de celulosa, hemicelulosa y la degradación de lignina. Asimismo, los datos
experimentales obtenidos en esta investigación serán ajustados al modelo planteado a fin de contribuir con
nuevas propuestas en el camino al planteamiento de modelos matemáticos más complejos que permitan la

112
descripción de los procesos de fermentación en fase sólida con fines industriales. Varias ecuaciones
cinéticas de complejidad variable, tales como la ecuación lineal, logística, de dos fases o la ecuación de
Monod han sido utilizadas por diferentes investigadores a fin de buscar la descripción de la producción de
biomasa en procesos de fermentación en estado sólido (Sangsurasak et al., 1996; Viccini et al., 2001). De
las expresiones matemáticas utilizadas, la más frecuente es la ecuación logística, debido probablemente a
la forma simple de la expresión. El modelo propuesto consignado en la Tabla 3 para la descripción de la
biomasa es la ecuación (1), la cual corresponde a la ecuación logística modificada por Mitchell et al.
(1999). Para describir la variación de los azúcares reductores en el tiempo se propuso la ecuación (2) que
correspondió a un factor de producción constante que afecta la derivada de la velocidad de producción de
biomasa, debido a que los azúcares reductores son considerados un producto de la fermentación que es
consumido por el mismo hongo como fuente de energía y su variación se ajustó más a la velocidad de
variación de la biomasa que a la variación de la biomasa misma. Para la degradación de lignina y los
consumos de hemicelulosa y celulosa en el tiempo de fermentación se plantearon las ecuaciones (3), (6) y
(8) cuyo planteamiento se realizó en función de las concentraciones de las enzimas específicas
responsables de la degradación de cada uno de los sustratos. La variación de las actividades enzimáticas
celulolíticas (endoglucanasa y exoglucanasa), xilanolítica (endoxilanasa) y las ligninasas (lacasa y MnP)
fueron propuestas como una producción que dependió de la concentración del sustrato específico y de la
velocidad de producción de la biomasa con un factor de inhibición provocado por la concentración de
azúcares reductores en el medio en el caso de las dos celulasas y la endoxilanasa, mientras que para las
dos ligninasas el factor de inhibición fue afectado por la misma concentración de lignina en el medio (ver
ecuaciones (4), (5), (7), (9) y (10)). La inhibición de las enzimas ligninolíticas puede deberse a sustancias
intermedias producidas durante la fermentación o probablemente por concentraciones muy altas de lignina
(Tengerdy y Szakacs, 2003). Aunque aún no hay certeza de ello, los datos experimentales obtenidos en
este trabajo mostraron una posible inhibición de las ligninasas y como no se determinaron productos
intermedios durante el desarrollo de la fermentación, se asumió una inhibición por sustrato, en este caso
por lignina. Finalmente para la variación de la enzima β-glucosidasa se propuso una ecuación cuya
producción dependió únicamente de la variación de la producción de biomasa con un factor de inhibición
causado por la concentración de los azúcares reductores. Los parámetros de cada una de las ecuaciones
consignadas en la Tabla 3 se describen en la Tabla 4.
La fermentación en estado sólido es un proceso heterogéneo que se ve afectado además por las
condiciones ambientales. La dificultad que representa la determinación del crecimiento de la biomasa, el
consumo de los sustratos y la imposibilidad de determinar todas las sustancias intermedias que pueden
generarse como producto de las reacciones bioquímicas durante las fases de incubación ha dificultado la
descripción de los procesos de fermentación en estado sólido y su escalamiento cuando se pretende aplicar
estos procesos en producción industriales de alguna sustancia de interés (Pal et al., 1995a; Sarikaya y
Ladisch, 1997; Pandey et al., 2000; Moreira et al., 2003). Las materias primas utilizadas para el
crecimiento y desarrollo de los hongos de pudrición blanca son bastante heterogéneas, compuestas
principalmente del complejo lignocelulósico (Viccini et al., 2001; Pradeep y Datta, 2002).
Los hongos basidiomicetos como los utilizados en el presente estudio contienen quitina en su
pared celular. Debido a que en los procesos de fermentación en estado sólido no es posible determinar
directamente la cantidad de biomasa, se determinó el contenido de quitina a través de la cuantificación de
N-acetil-D-glucosamina (NAGA) en todos los tiempos de la fermentación. Además, se le determinó el
contenido de NAGA al micelio seco obtenido de cultivo líquido a cada una de las tres especies de trabajo,
obteniendo como resultado 15,83% (p/p) en P. ostreatus, 14,15% (p/p) en C. versicolor y 10,08% (p/p) en
L. edodes. Estos porcentajes de NAGA fueron utilizados para calcular la cantidad de biomasa de los
sustratos sólidos en cada uno de los tiempos de fermentación; por lo que estos datos de biomasa fueron los
que se sometieron al modelamiento con la ecuación (1). El ajuste de los datos experimentales al modelo de
la biomasa y los azúcares reductores se muestran en la Figura 9, en la Tabla 5 se encuentran los valores de
los parámetros ajustados de la biomasa y los azúcares reductores y la bondad del ajuste obtenido para los
datos de la biomasa fue del 90% aproximadamente y para los azúcares reductores del 55% referidos a la
combinación que presentó el ajuste más pobre que corresponde a la varianza de los residuales más alta.

113
Tabla 3. Modelo matemático para la descripción de producción de biomasa, enzimas lignocelulolíticas y
consumo de la matriz lignocelulósica
Ecuación Descripción
. 1
n
b bm b
bm
dC CC
dt C
(1) Biomasa
. . . 1 1 .AR
n
b bp m
bm
dC Cq n
dt C
dC
dt
(2)
Azúcares
reductores
. .LL lac MnP
dCk C C
dt (3) Lignina
. . .lac blac L lac L
dC dCk C C
dt dt (4) Lacasa
. . .MnP bMnP L MnP L
dC dCk C C
dt dt
(5)
Manganeso
peroxidasa
(MnP)
.HMHM ENX
dCk C
dt (6) Hemicelulosa
. . .ENX bENX HM ENX AR
dC dCk C C
dt dt (7) Endoxilanasa
. .CC ENG EXG
dCk C C
dt
(8) Celulosa
. ..ENG bENG ARENG C
dC dCk C
dC
t dt
(9) Endoglucanasa
. ..EXG bEXG AREXG C
dC dCk C
dC
t dt (10) Exoglucanasa
.. BGBG b
BG AR
dC dCk
tC
d dt (11) β-glucosidasa
En la Figura 10 se presenta el comportamiento de los datos experimentales respecto al modelo
propuesto en las ecuaciones (3), (4) y (5); los parámetros de las ecuaciones se encuentran consignados en
la tabla 5. De igual forma, el ajuste para los datos experimentales de la lignina, lacasa y MnP al modelo
propuesto son del 90%, 43% y 87% respectivamente para la F9.
En la Figura 11 se muestra el ajuste de los datos experimentales de la hemicelulosa y la enzima
endoxilanasa al modelo (ecuaciones (6) y (7)) y los parámetros del modelamiento se encuentran en la
Tabla 5. Los porcentajes de ajuste al modelo para la hemicelulosa fueron del 86% y para la endoxilanasa
fue del 82% aproximadamente para F9. El modelamiento de los datos experimentales de celulosa,
endoglucanasa, exoglucanasa y β-glucosidasa se muestran en la Figura 12. Del mismo modo los
parámetros del modelo están consignados en la Tabla 5 y el ajuste de los datos experimentales al modelo
fue de 40% para celulosa, 88% endoglucanasa, 78% exoglucanasa y 96% β-glucosidasa para la F9.

114
Tabla 4. Parámetros de las ecuaciones del modelo matemático.
Parámetro Significado Parámetro Significado
µm Velocidad específica de crecimiento de la biomasa
(día-1) kENX
Coeficiente de producción de Endoxilanasa
(gss/mg)
Cbm Concentración de biomasa máxima (mg/gss) µENX Coeficiente de inhibición de Endoxilanasa (día-1)
N
n < 1 el organismo es relativamente sensible a la
auto-inhibición y ésta ocurre para valores muy bajos
de Cb
n = 1 ecuación logística
n > 1 el organismo es relativamente resistente a la
auto-inhibición y ésta ocurre solo cuando Cb ≈ Cbm
CHM0 Concentración inicial de hemicelulosa (mg/gss)
Cb0 Concentración de biomasa máxima (mg/gss)
CENX0 Actividad inicial de Endoxilanasa (U/gss)
qp Coeficiente de producción constante de azúcares
reductores (día) kC
Coeficiente de consumo de celulosa
(mg.gss/día.U2)
CAR0 Concentración inicial de azúcares reductores
(mg/gss) kENG
Coeficiente de producción de Endoglucanasa
(gss/mg)
kL Coeficiente de degradación de lignina
(mg.gss/día.U) µENG
Coeficiente de inhibición de Endoglucanasa (día-
1)
klac Coeficiente de producción de lacasa (gss/mg)
kEXG Coeficiente de producción de Exoglucanasa
(unidades gss/mg)
µlac Coeficiente de inhibición de lacasa (día-1) µEXG Coeficiente de inhibición de Exoglucanasa (día-1)
kMnP Coeficiente de producción de MnP (gss/mg) CC0 Concentración inicial de celulosa (mg/g ss)
µMnP Coeficiente de inhibición de MnP (día-1) CENG0 Actividad inicial de Endoglucanasa (U/g ss)
CL0 Concentración inicial de lignina (mg/g ss) CEXG0 Actividad inicial de Exoglucanasa (U/g ss)
Clac0
Actividad inicial de lacasa (U/gss) kBG
Coeficiente de producción de β-glucosidasa
(mg/mg)
CMnP0 Actividad inicial de MnP (U/gss) µBG Coeficiente de inhibición de β-glucosidasa (día-1)
kHM Coeficiente de consumo de hemicelulosa (mg/día.U) CBG0 Actividad inicial de β-glucosidasa (U/gss)
Figura 9. Perfil en el tiempo de la biomasa celular y los azúcares reductores para la combinación C. versicolor -
F9. Los puntos discretos corresponden a los datos experimentales; las líneas continuas son calculadas por el modelo
propuesto.

115
Figura 10. Ajuste de datos experimentales de lignina, lacasa, MnP para la combinación C. versicolor - F9
Figura 11. Ajuste de datos experimentales de hemicelulosa y endoxilanasa para la combinación C. versicolor -
F9
Figura 12. Ajuste de datos experimentales de celulosa, endoglucanasa, exoglucanasa y β-glucosidasa para la
combinación C. versicolor - F9
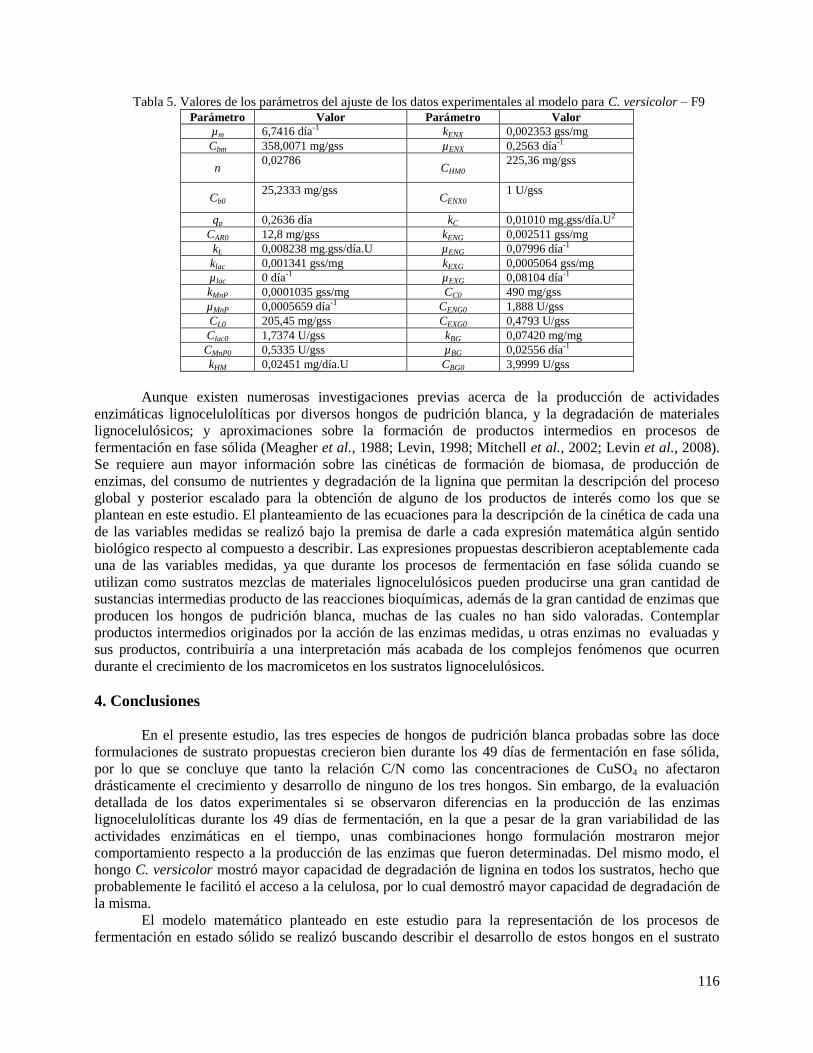
116
Tabla 5. Valores de los parámetros del ajuste de los datos experimentales al modelo para C. versicolor – F9
Parámetro Valor Parámetro Valor
µm 6,7416 día-1 kENX 0,002353 gss/mg
Cbm 358,0071 mg/gss µENX 0,2563 día-1
n 0,02786
CHM0 225,36 mg/gss
Cb0 25,2333 mg/gss
CENX0
1 U/gss
qp 0,2636 día kC 0,01010 mg.gss/día.U2
CAR0 12,8 mg/gss kENG 0,002511 gss/mg
kL 0,008238 mg.gss/día.U µENG 0,07996 día-1
klac 0,001341 gss/mg kEXG 0,0005064 gss/mg
µlac 0 día-1 µEXG 0,08104 día-1
kMnP 0,0001035 gss/mg CC0 490 mg/gss
µMnP 0,0005659 día-1 CENG0 1,888 U/gss
CL0 205,45 mg/gss CEXG0 0,4793 U/gss
Clac0 1,7374 U/gss kBG 0,07420 mg/mg
CMnP0 0,5335 U/gss µBG 0,02556 día-1
kHM 0,02451 mg/día.U CBG0 3,9999 U/gss
Aunque existen numerosas investigaciones previas acerca de la producción de actividades
enzimáticas lignocelulolíticas por diversos hongos de pudrición blanca, y la degradación de materiales
lignocelulósicos; y aproximaciones sobre la formación de productos intermedios en procesos de
fermentación en fase sólida (Meagher et al., 1988; Levin, 1998; Mitchell et al., 2002; Levin et al., 2008).
Se requiere aun mayor información sobre las cinéticas de formación de biomasa, de producción de
enzimas, del consumo de nutrientes y degradación de la lignina que permitan la descripción del proceso
global y posterior escalado para la obtención de alguno de los productos de interés como los que se
plantean en este estudio. El planteamiento de las ecuaciones para la descripción de la cinética de cada una
de las variables medidas se realizó bajo la premisa de darle a cada expresión matemática algún sentido
biológico respecto al compuesto a describir. Las expresiones propuestas describieron aceptablemente cada
una de las variables medidas, ya que durante los procesos de fermentación en fase sólida cuando se
utilizan como sustratos mezclas de materiales lignocelulósicos pueden producirse una gran cantidad de
sustancias intermedias producto de las reacciones bioquímicas, además de la gran cantidad de enzimas que
producen los hongos de pudrición blanca, muchas de las cuales no han sido valoradas. Contemplar
productos intermedios originados por la acción de las enzimas medidas, u otras enzimas no evaluadas y
sus productos, contribuiría a una interpretación más acabada de los complejos fenómenos que ocurren
durante el crecimiento de los macromicetos en los sustratos lignocelulósicos.
4. Conclusiones
En el presente estudio, las tres especies de hongos de pudrición blanca probadas sobre las doce
formulaciones de sustrato propuestas crecieron bien durante los 49 días de fermentación en fase sólida,
por lo que se concluye que tanto la relación C/N como las concentraciones de CuSO4 no afectaron
drásticamente el crecimiento y desarrollo de ninguno de los tres hongos. Sin embargo, de la evaluación
detallada de los datos experimentales si se observaron diferencias en la producción de las enzimas
lignocelulolíticas durante los 49 días de fermentación, en la que a pesar de la gran variabilidad de las
actividades enzimáticas en el tiempo, unas combinaciones hongo formulación mostraron mejor
comportamiento respecto a la producción de las enzimas que fueron determinadas. Del mismo modo, el
hongo C. versicolor mostró mayor capacidad de degradación de lignina en todos los sustratos, hecho que
probablemente le facilitó el acceso a la celulosa, por lo cual demostró mayor capacidad de degradación de
la misma.
El modelo matemático planteado en este estudio para la representación de los procesos de
fermentación en estado sólido se realizó buscando describir el desarrollo de estos hongos en el sustrato

117
lignocelulósico, a fin de buscar puntos generales de interpretación de estos procesos que por ser tan
heterogéneos se hacen complejos y carecen de expresiones matemáticas que permitan el escalamiento de
estos procesos con fines industriales, evitando hacer experimentación individual de cada proceso.
Agradecimientos
Los autores agradecen a la Vicerrectoría de Investigaciones y Postgrados de la Universidad de Caldas
(Colombia) por la financiación del presente trabajo y al Instituto de Biotecnología Agropecuaria de la
misma institución por el apoyo material y de infraestructura. Asimismo, los autores expresan sus
agradecimientos a la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires
(Argentina) por su apoyo y asesoría durante la realización de este trabajo.
Referencias Asatiani MD, Kachlishvili ET, Khardziani TS, Metreveli EM, Mikiashvili NA, Songulashvili GG, Tsiklauri ND, Wasser SP,
Elisashvili VI (2008) Basidiomycetes as a source of antioxidants, lectins, polysaccharides, and enzymes. Journal of
Biotechnology 136S: S717-S742.
Baldrian P, Gabriel J (2002) Variability of laccase activity in the whiterot basidiomycete Pleurotus ostreatus. Folia Microbiology
47: 385-390.
Barr BK, HsieH YL, Ganem B, Wilson DB (1996) Identification of two functionally different classes of exocellulases.
Biochemistry 35: 586-592.
Béguin P, Aubert JP (1994) The biological degradation of cellulose. FEMS Microbiology Reviews 13: 25-58.
Blanchette RA (1995) Degradation of the lignocellulose complex in wood. Canadian Journal of Botany 73: S999-S1010.
Buswell JA, Cai YJ, Chang ST (1996) Ligninolytic enzyme production and secretion in edible mushroom fungi. En Royse DJ
(Ed.) Mushroom Biology and Mushroom Products. The Pennsylvania State University pp. 113±122. Pennsylvania.
Carabajal M, Levin L, Abertó E, Lechner B (2012) Effect of co-cultivation of two Pleurotus species on lignocellulolytic enzyme
production and mushroom fructification. International Biodeterioration & Biodegradation 66: 71-76.
Carlile M, Watkinson S, Gooday G (2001) The fungi. Academic Press. London. 588 p. p.
Dormand JR, Prince PJ (1980) A family of embedded Runge-Kutta formulae. Journal of computational and applied mathematics
6: 19-26.
Gassara F, Brar S, Tyagi RD, Verma M, Surampalli RY (2010) Screening of agro-industrial wastes to produce ligninolytic
enzymes by Phanerochaete chrysosporium. Biochemical Engineering Journal 49: 388-394.
Gutierrez-Correa M, Tengerdy RP (1997) Production of cellulase on sugar cane bagasse by fungal mixed culture solid substrate
fermentation. Biotechnology Letters in Applied Microbiology 19: 665 - 667.
Hudson H (1986) Fungal Biology. Edward Arnolds Eds. Maryland. 298 p. p.
Ikasari L, Mitchell DA (2000) Two-phase model of yhe kinetics of growth of Rhizopus oligosporus in membrana culture.
Biotechnology Bioengineering 68: 619-627.
Joselau JP, Ruel K (1994) Wood polysaccharides and their degradation by fungi. En Ouellette P (Ed.) Host Wall Alterations by
Parasitic Fungi. APS Press. Minnesota.
Kachlishvili E, Penninckx M, Tsiklauri N, Elisashvili V (2006) Effect of nitrogen source on lignocellulolytic enzyme production
by white-rot basidiomycetes under solid-state cultivation. World Journal of Microbiology & Biotechnology 22(4): 391-397.
Kirk TK (1983) Degardation and convertion of lignocelluloses. En Smith JE, Berry DR, Kristiansen P (Eds.) The filamentous
fungi. London.
Kirk TK, Farrel RL (1987) Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology 41: 465-
505.
Kjeldahl J (1883) Neue Methode Zùr Bestimmung der Stickstoffs in Organichen Kòrpen. Zeitschrift fur Analytische Chemie 22:
366-382.
Leterme P. (2010). Análisis de alimentos y forrajes, in Protocolos de LaboratorioUniversidad Nacional de Colombia Sede
Palmira: Palmira. 66 pp.
Levin L (1998) Biodegradación de materiales lignocelulósicos por Trametes trogii (Aphyllophorales, Basidiomycetes).Tesis.
Universidad de Buenos Aires. Buenos Aires.
Levin L, Forchiassin F, Ramos AM (2002) Copper induction of lignin-modifying enzymes in the white-rot fungus Trametes
trogii. Mycologia 94(3): 377-383.
Levin L, Herrmann C, Papinutti VL (2008) Optimization of lignocellulolytic enzyme production by the white-rot fungus Trametes
trogii in solid-state fermentation using response surface methodology. Biochemical Engineering Journal 39: 207-214.
Li L, Li XZ, Tang WZ, Zhao J, Qu Y-B (2008) Screening of a fungus capable of powerful and selective delignification on wheat
straw. Letters in Applied Microbiology 47: 415-420.
Mata G, Savoie JM (1998) Extracellular enzyme activities in six Lentinula edodes strains during cultivation in wheat straw. World
J. Microbiol. Biotechnol. 14: 513-519.

118
Mata G, Murrieta Hernández DM, Iglesias Andreu LG (2005) Changes in lignocellulolytic enzyme activites in six Pleurotus spp.
strains cultivated on coffee pulp in confrontation with Trichoderma spp. World Journal of Microbiology & Biotechnology 21:
143-150.
Meagher MM, Tao BY, Chow JM, Reilly PJ (1988) Kinetics and subsite mapping of β -D-xylobiose and D-xylose producing
Aspergillus niger endo-β-1,4-D-xylanase. Carbohydrates Resources 173: 273-283.
Miller GL (1959) Use of DinitrosaIicyIic Acid Reagent for Determination of Reducing Sugar. Analytical Chemistry 31(3): 426-
428.
Mitchell D, Stuart D, Tanner R. (1999). Solid-State fermentation - Microbial growth kinetics, in The Encyclopedia of Bioprocess
Technology: Fermentation, Biocatalysis and bioseparation, Flickinger M, Drew S, EditorsWiley: New York. 2407-2429
Mitchell D, Von Meien O, Krieger N (2002) Recent developments in modeling of solid-state feremntation: heat and mass transfer
in bioreactors. BiochemicalEngineering Journal 13: 137-147.
Mitchell DA, Von meien OF, Krieger N, Dalsenter FD (2004) A review of recent developments in modeling of microbial growth
kinetics and intraparticle phenomena in solid-state fermentation. Biochemical Engineering Journal 17: 15-26.
Montoya BS, Orrego CA, Levin L (2011) Growth, fruiting and lignocellulolytic enzyme production by the edible mushroom
Grifola frondosa (maitake). World J. Microbiol. Biotechnol. ISSN 0959-3993: DOI 10.1007/s11274-11011-10957-11272.
Moreira MT, Feijoo G, Lema JM (2003) Fungal bioreactors: Applications to white-rot fungi. Reviews in Environmental Science
and Bio/Technology 2: 247-259.
Niladevi KN, Prema P (2008) Effect of inducers and process parameters on laccase production by Streptomyces psammoticus and
its application in dye decolourization. Bioresource Technology 99 4583-4589.
Pal M, Calvo A, Terron MC, Gonzalez AE (1995) Solid-state fermentation of sugarcane bagasse with Flammulina velutipes and
Trametes versicolor. World Journal Microbiology Biotechnology 11: 541-545.
Panagiotou G, Kekos D, Macris B, Christakopoulos P (2003) Production of cellulolytic and xylanolytic enzymes by Fusarium
oxysporum grown on corn stover in solid state fermentation. Industrial Crops and Products 18: 37-45.
Pandey A, Soccol CR, Mitchell D (2000) New developments in solid state fermentation: I-bioprocesses and products. Process
Biochemistry 35: 1153–1169.
Paszczczynski A, Crawford R, Huyn V (1988) Manganese peroxidase of Phanerochaete chrysosporium purification. Methods
enzymology 161: 264-270.
Paszczynski A, Crawford RL (1991) Degradation of azo compounds by ligninases from Phanerochaete chrysosporium Involment
of veratryl alcohol. Biochemistry Biophysics resources communications 178: 1056-1063.
Plassard C, Mousain D, Salsac L (1982) Estimation of mycelial growth of basidiomycetes by means of chitin determination.
Phytochemistry 21: 345-348.
Pradeep V, Datta M (2002) Production of ligninolytic enzymes for decolorization by cocultivation of white-rot fungi Pleurotus
ostreatus and Phanerochaete chrysosporium under solid-state fermentation. Applied Biochemistry Biotechnology 102-103:
109-118.
Reddy GV, Ravindra BP, Komaraiah P, Roy RM, Kothari IL (2003) Utilization of banana waste for the production of lignolytic
and cellulolytic enzymes by solid substrate fermentation using two Pleurotus species (P. ostreatus and P. sajor-caju). Process
Biochemistry 38: 1457-1462.
Rosale E, Couto SR, Sanromán A (2007) Increased laccase production by Trametes hirsuta grown on ground orange peelings.
Enzyme and Microbial Technology 40 1286-1290.
Sangsurasak P, Nopharatana M, Mitchell D (1996) Mathematical modeling of the growth of filamentous fungi in solid-state
fermentation. Journal Science Industrial Resources 55: 333-342.
Sarikaya A, Ladisch M (1997) An Unstructured Mathematical Model for Growth of Pleurotus ostreatus on Lignocellulosic
Material in Solid-State Fermentation Systems. Applied Biochemistry and Biotechnology 62: 72-85.
Silva EM, Martins SF, Milagres AMF (2008) Extraction of manganese peroxidase produced by Lentinula edodes. Bioresource
Technology 99: 2471-2475.
Solomon EI, Sundaram UM, Machonkin TE (1997) Multicopper oxidases and oxygenases. Chemical Reviews 96: 2563-2605.
Tavares A, Coelho M, Coutinho J, Xavier A (2005) Laccase improvement in submerged cultivation: induced production and
kinetic modelling. Journal of Chemical Technology and Biotechnology 80: 669-676.
Tengerdy RP, Szakacs G (2003) Bioconversion of lignocellulose in solid substrate fermentation. Biochemical Engineering
Journal 13: 169-179.
Tsiklauri ND, Khardziani TS, Kachlishvili ET, Elisashvili VI (1999) Cellulase and xylanase activities of higher basidiomycetes
during bioconversion of plant raw materials depending on the carbon source in the nutrient medium. Applied Biochemistry
and Microbiology 35: 291-295.
Van de Lagemaat J, Pyle DL (2005) Modelling the uptake and growth kinetics of Penicillium glabrum in a tannic acid-containing
solid-state fermentation for tannase production. Process Biochemistry 40: 1773-1782.
Viccini G, Mitchell D, Boit S, Gern J, da Rosa A, Costa R (2001) Analysis of growth kinetic profiles in solid-state fermentation.
Food Technology and Biotechnology Advances 39: 271-294.
Walkley A, Black I (1934) An estimation of the Degtjareft method for determining of soil organic matter and a proposed
modification of chromic acid titration method. Soil Science 37: 29-38.
Wood TM, García CV (1994) Enzymes and mechanisms involved in microbial cellulolysis. Kluwer Academic publishers.
Netherlands.

119
6. Selección de la Mejor Combinación Hongo –
Formulación para la producción de polisacáridos
mediante Fermentación en Estado Sólido con Pleurotus
ostreatus, Coriolus versicolor y Lentinus edodes

120
Selección de la Mejor Combinación Hongo – Formulación para la producción
de polisacáridos mediante Fermentación en Estado Sólido con Pleurotus
ostreatus, Coriolus versicolor y Lentinus edodes
Sandra Montoya1, Óscar Julián Sánchez
2*, Laura Levin
3
RESUMEN
En esta investigación se realizó el cultivo de tres especies de hongos de pudrición blanca: Coriolus
versicolor, Pleurotus ostreatus y Lentinus edodes sobre doce formulaciones de sustrato en estado sólido
empleando mezclas de diversos materiales y sales de carbonato de calcio y dos niveles de sulfato de cobre
(II). Los objetivos de este trabajo fueron i) evaluar los efectos de la relación C/N y del sulfato de cobre (II)
sobre la producción de polisacáridos durante 49 días de fermentación en estado sólido; así como el
consumo de celulosa y hemicelulosa y la degradación de lignina y ii) proponer un modelo matemático
para la descripción de la producción de biomasa, de los polisacáridos, producción y consumo de azúcares
reductores, consumo de celulosa y hemicelulosa y degradación de lignina. Las tres especies de hongos
crecieron bien sobre todas las formulaciones de sustrato. Empero, hubo combinaciones para los cuales la
producción de polisacáridos fue muy baja, como 23,282 mg/gss (g sustrato sólido) para la combinación P.
ostreatus – F12, mientras que la mayor producción la exhibió C. versicolor – F2 con 96,085 mg/gss. Se
propuso un modelo matemático compuesto de 6 ecuaciones diferenciales a fin de buscar la descripción de
los datos experimentales de todas las combinaciones resultantes para C. versicolor, por haber sido el
hongo que más cantidad de polisacáridos produjo; asimismo, se plantearon ecuaciones para describir la
degradación de los sustratos lignocelulósicos. En este trabajo se presentan los resultados del
modelamiento para las cuatro combinaciones especie de hongo-fórmula, F2, F4, F7 y F11 que arrojaron la
mayor producción de polisacáridos en los 49 días de fermentación en fase sólida.
Palabras clave: hongos de pudrición blanca, relación C/N, producción de polisacáridos, degradación de
materiales lignocelulósicos.
1. INTRODUCCIÓN
Los polisacáridos de los hongos macromicetos han sido reconocidos como excelentes
inmunoestimuladores debido al potencial de sus propiedades terapéuticas, a que no son tóxicos y son
escasos sus efectos secundarios en comparación con otros inmunoestimulantes. Estos polisacáridos
bioactivos son producidos en cantidades relevantes a partir de los cuerpos fructíferos de los hongos, su
micelio y además, de los filtrados obtenidos de sus cultivos en fermentación sumergida. En mayor medida
las estructuras de los polisacáridos procedentes de los macromicetos corresponden a homoglicanos,
heteroglicanos o complejos de gluco-proteínas. La fuente principal de polisacáridos antitumorales parecen
ser sus paredes celulares, las cuales están compuestas de polisacáridos. En general, la actividad
inmunomoduladora de los polisacáridos de los macromicetos está relacionada con las estructuras β-(1→3),
β-(1→6) glucanos, α-(1→3) glucanos y complejos de polisacáridos de galactomananos y glucuromananos
unidos a proteínas presentes en la pared celular de los macromicetos (Enshasy, 2010). El β-D-glucano es
un polisacárido que produce exclusivamente D-glucosa luego de la hidrólisis ácida (Mizuno, 1999). Los
polisacáridos obtenidos por fermentación sumergida con una adecuada selección del medio de cultivo, se
denominan exopolisacáridos (EPS), ya que se excretan en el medio de cultivo durante el curso de la
fermentación. Los intrapolisacáridos (IPS) pueden ser producidos a partir de los cuerpos fructíferos y su
micelio después de haber sido sometido a diferentes procesos de extracción sólido-líquido utilizando

121
diferentes disolventes, desde agua caliente hasta diferentes mezclas de disolventes orgánicos (Amaral et
al., 2008; Komura et al., 2010).
Los exopolisacáridos de basidiomicetos de pudrición blanca juegan un rol importante en los
procesos de crecimiento y desarrollo de estos organismos. Estos exopolisacáridos pueden inmovilizar las
exoenzimas que ellos mismo excretan (Tang y Zhong, 2002). Según Catley (1992) el gel formado por
estos biopolímeros previene la deshidratación de la hifa, permitiendo la adherencia de otras células a las
superficies de los sustratos, posibilitando la selección de las moléculas del ambiente que los rodea
(Maziero et al., 1999). Adicionalmente, estos biopolímeros tienen un amplio rango de aplicaciones
industriales.
Las diferentes especies de hongos producen diferentes tipos de polisacáridos con diversas
propiedades y la estructura del polisacárido influye sobre la actividad biológica que se obtiene. Del mismo
modo las estructuras químicas de los polisacáridos dependen de la fuente de donde provienen, el
carpóforo, el micelio o el medio de cultivo, estos compuestos varían en sus estructuras moleculares, la
unidad estructural que se repite (el monómero) y los pesos moleculares, entre otros factores (Tsukagoshi
et al., 1984; Ishikawa et al., 1992; Hamlyn y Schmidt, 1994; Kobayashi et al., 1995; Zhuang y Mizuno,
1999; Chang y Miles, 2004; Mohammad-Fata et al., 2007). Por ejemplo, el lentinan un β-(1→3)-D-
glucano extraído de L. edodes es una potente droga inmunoestimuladora licenciada en Japón para terapias
antitumorales (Kupfahl et al., 2006). De igual forma se ha reportado actividad antitumoral de los
complejos de polisacáridos extraídos del micelio o del cuerpo fructífero de hongos macromicetos, como el
complejo proteína-polisacárido PSK del hongo C. versicolor (Ooi y Liu, 2000a), el complejo polisacárido-
proteína del cultivo de Tricholoma lobayense (Liu et al., 1996a), exobiopolímeros de varias especies de
Cordyceps a las cuales se les ha demostrado actividad antitumoral (Chen et al., 1997),
inmunopotenciadora (Nakamura et al., 1999), actividad hipoglicémica (Kiho et al., 1999) y efecto
hipocolesterolémico (Koh et al., 2003). Estos ejemplos son una muestra de la potencialidad que tienen los
hongos como fuente de sustancias bioactivas para la obtención de potenciadores del sistema
inmunológico, teniendo en cuenta que de las 140.000 especies de macromicetos que se estima existen en
el planeta, solamente se conocen cerca del 10%, de las cuales sólo han sido examinadas desde su potencial
inmunomodulador el 5% (Lindequist et al., 2005b).
La suplementación de los sustratos es una estrategia para incrementar la producción de biomasa
celular y una forma de mantener constante la producción de polisacáridos (Tang y Zhong, 2002).
Asimismo, diferentes trabajos publicados en la última década evidencian la importancia de la presencia de
aceites vegetales ricos en ácido oleico como promotores de crecimiento de la biomasa fúngica y por ende
de sus polisacáridos constituyentes (Yang et al., 2000; Hsieh et al., 2008). Al momento de seleccionar el
tipo de aceite vegetal o ácido graso a utilizar como inductor de polisacáridos es necesario considerar la
composición de ácidos grasos del aceite; ya que ha sido reportado por varios investigadores que
cantidades importantes de ácido linoleico suprimen la producción de biomasa y polisacáridos, mientras
que la presencia de ácido oleico promueve su producción (Park et al., 2002a; Hsieh et al., 2006; Hsieh et
al., 2008). Los aceites de soya y de oliva han sido utilizados como promotores de crecimiento celular y de
polisacáridos en cultivos de macromicetos, tanto líquidos como sólidos. Al respecto, el aceite de oliva
contiene 84% de ácido oleico y entre 30 y 60% en aceite de soya con bajos contenidos de ácido linoleico
(Wade, 1993; Hsieh et al., 2008).
En general, el proceso de determinación de intra y exo-polisacáridos se inicia mediante la
medición de la cantidad de polisacáridos como carbohidratos totales presentes en una muestra
seleccionada. Estos polisacáridos corresponden a los contenidos en los cuerpos fructíferos de los hongos,
al micelio o a los presentes en los caldos de cultivo de fermentaciones sumergidas.
El objetivo de este trabajo fue evaluar el efecto de la relación carbono nitrógeno y del sulfato de cobre (II)
sobre la producción de polisacáridos por tres especies de hongos de pudrición blanca (L. edodes, C.
versicolor y P. ostreatus) empleando doce formulaciones diferentes de medios de cultivo por
fermentación en estado sólido en fase vegetativa. Estos hongos fueron seleccionados de entre diez hongos
de pudrición blanca mediante un rastreo basado en la medida de los polisacáridos como carbohidratos
totales; los resultados de este rastreo se relacionan en el artículo anterior. Igualmente este trabajo tuvo

122
como objetivo adicional proponer y validar un modelo matemático para la mejor combinación hallada
especie de hongo - formulación a fin de realizar la descripción de la cinética de crecimiento del
organismo, y la producción de polisacáridos en condiciones de fermentación en estado sólido.
2. MATERIALES Y MÉTODOS
2.1. Microorganismos
Tres especies de basidiomicetos fueron empleados para la producción de polisacáridos en
fermentación en estado sólido, tomadas de la colección de hongos de la Universidad de Caldas
(Manizales, Colombia): Lentinula edodes del Centro de Investigaciones del Café, Chinchiná, Caldas,
Colombia (CICL54), Coriolus versicolor de Pennsylvania State University, Estados Unidos de Norte
América (PSUWC430), y Pleurotus ostreatus de la Colección interna de la Universidad de Caldas
(UCC001). Las cepas fueron extendidas sobre agar papa dextrosa (PDA) y mantenidas en refrigeración a
4°C y transferencias periódicas.
2.2. Producción de semilla
La semilla para la inoculación de los medios de cultivo sólidos se realizó sobre trigo lavado,
hidratado con agua caliente hasta 40% de humedad. El trigo hidratado fue empacado en unidades de 1 kg
en bolsas de polipropileno biorientado, provistas con un filtro de interlón en el primer tercio de la parte
superior de la bolsa de una pulgada de lado. Posteriormente, el grano fue esterilizado a 121°C por 30
minutos aplicando el método Thindal. Finalmente el grano hidratado y esterilizado fue inoculado a una
tasa del 4% en base húmeda con cada especie de hongo en cabina de flujo laminar y llevadas a incubación
por 12 a 15 días a 25°C y penumbra hasta colonización completa.
2.3. Medios de cultivo
Se realizaron doce formulaciones de sustrato por triplicado. Los sustratos fueron formulados en
base seca, con 40% de aserrín de roble, 20% de capacho de coco y 2% de aceite de soya como materias
primas fijas en todas las formulaciones y variando los porcentajes de cascarilla de café y salvado de maíz
a fin de modificar la relación carbono nitrógeno (R C/N) (ver Tabla 1) y dos niveles de sulfato de cobre:
0,1596% para las fórmulas impares y 0,0798% para las fórmulas pares. Todos los sustratos fueron
formulados a 60% de humedad. Los sustratos fueron empacados en bolsas de polipropileno biorientado en
unidades de 200 g cada una y esterilizados a 121°C por 30 min aplicando el método Thindal.
Posteriormente fueron inoculados en cabina de flujo laminar con 4% de inóculo en base húmeda respecto
al sustrato. Éstos fueron llevados a incubación a 25°C en penumbra por 49 días. Se tomaron 15 muestras,
dos por semana, tomándose la primera muestra el día de la inoculación.
Tabla 1. Relaciones carbono nitrógeno (R C/N) en doce niveles (formulaciones)
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 F11 F12
142,640 91,77 114,61 93,64 100,92 91,79 85,34 84,87 55,11 51,55 114,55 111,05
2.4. Determinación de la relación carbono nitrógeno
La relación carbono nitrógeno (C/N) fue determinada a partir de la medida de la materia orgánica
de las doce formulaciones de sustratos empleando el método de Walkley y Black (1934), el cual utiliza
ácido sulfúrico concentrado y solución de dicromato de potasio. La muestra se lee a 585 nm. La cantidad

123
de carbono total corresponde al 58% de la materia orgánica. El nitrógeno orgánico total fue determinado
por el método de Kjeldahl (Kjeldahl, 1883).
2.5. Determinación de polisacáridos como carbohidratos totales
Los polisacáridos fueron extraídos de 0,1g de sólido seco en 5mL de etanol concentrado grado
reactivo, sometido a ultrasonido por 20 min, con posterior reposo por 24 horas. Luego se realizó una
filtración al vacío, el filtrado fue centrifugado por 20 min a 14.000 rpm descartando el sobrenadante. El
precipitado (la muestra) fue resuspendida en NaOH 1N a 60°C por 1 hora, posteriormente, la suspensión
fue sometida al método fenol-sulfúrico para determinar la cantidad de carbohidratos totales a 490 nm
(Dubois et al., 1956).
2.6. Cuantificación del contenido de N-acetil-D-glucosamina (NAGA) y biomasa
Las muestras de sustrato fueron colectadas en 15 tiempos de incubación, dos por semana, hasta el
día 49 de incubación para las tres especies de trabajo. El sustrato sólido incubado a diferentes tiempos fue
secado en estufa a 101°C hasta peso constante, molido y almacenado en lugar fresco y seco para la
determinación del contenido de quitina. El contenido de biomasa fúngica en los sustratos sólidos fue
indirectamente estimado por la determinación del contenido de N-acetil-D-glucosamina (NAGA) después
de la hidrólisis con HCl 6N grado analítico según el método de Plassard et al. (1982). Para ello se
cuantificó paralelamente el contenido de NAGA por g de micelio en cada una de las tres especies,
cultivándolas en medio líquido. El contenido de NAGA del micelio de cada una de las especies de hongos
de trabajo: C. versicolor, P. ostreatus y L. edodes fue determinado del desarrollo del micelio en medio
líquido en matraces de 250 mL con 100 mL de medio de cultivo con la siguiente composición: glucosa (30
g/L), extracto de levadura (6 g/L), SO4Mg.5H2O (0,5 g/L), K2HPO4 (0,5 g/L) y CaCl2 (0,1 g/L). El tiempo
de fermentación para el crecimiento del hongo fue de 25 días, en agitación a 100 rpm de agitación. Los
compuestos solubles en agua fueron extraídos durante la hidrólisis con HCl 6N por tres horas.
2.7. Cuantificación de de los componentes de la fibra y azúcares reductores
Se determinaron los componentes de la fibra (celulosa, hemicelulosa y lignina) de los sustratos de
cada una de las formulaciones en 15 tiempos durante los 49 días de incubación, así como de la fracción
soluble, empleando los resultados de la determinación de fibras detergente neutra, ácida y lignino-ácida.
La medición se realizó a una muestra seca y molida de sustrato; dicha muestra se sometió a tres hidrólisis
en serie (cada una de 75 min): i) hidrólisis con lauril sulfato de sodio y otros, ii) hidrólisis en solución de
bromuro de amonio en ácido sulfúrico 1N, y iii) hidrólisis con ácido sulfúrico al 72% (p/v). Al finalizar
cada hidrólisis, las muestras fueron lavadas y secadas a 105°C hasta peso constante (Leterme, 2010a). El
contenido de azúcares reductores como glucosa fue determinado por el método del ácido dinitrosalicílico
(DNS) (Miller, 1959).
2.8. Diseño experimental
Se planteó un diseño experimental bifactorial con la especie a tres niveles y doce formulaciones de
sustrato. Las formulaciones varían en su relación C/N y la concentración de sulfato de cobre (II). Las
variables independientes en este diseño experimental fueron la especie de hongo, la formulación del
sustrato y el tiempo de fermentación. Las variables de respuesta medidas fueron la producción de biomasa,
de polisacáridos, producción y consumo de azúcares reductores y la degradación de la celulosa en 15
tiempos durante 49 días de fermentación en estado sólido.

124
2.9. Modelamiento matemático del crecimiento del hongo y la producción de enzimas
Para la solución de los modelos matemáticos del proceso de fermentación en estado sólido se
utilizó el software Matlab® 2010b (MathWorks, EUA) en un PC de 3 GB de memoria RAM y un
procesador Intel CORE-i3 de 2,13 GHz. Para los modelos matemáticos planteados en ecuaciones
diferenciales ordinarias se utilizó el comando ode45 que está basado en una fórmula explícita de Runge-
Kutta (4,5) usando un par de Dormand-Prince (Dormand y Prince, 1980) y el comando ode15s basado en
una fórmula de orden variable que funciona mediante fórmulas de diferenciación numérica.
2.10. Análisis estadístico
Para todos los análisis estadísticos del presente trabajo se utilizó el software Matlab ® 2010b. Este
análisis se realizó en tres etapas. Primero se realizó un ANOVA a todos los datos del diseño experimental
y sus posibles combinaciones: especie, fórmula y tiempo de fermentación mediante el comando anovan.
Luego se realizó un análisis comparativo (Kruskal Wallis) de la producción de polisacáridos para cada
especie de hongo estudiado y las doce formulaciones de sustratos mediante los comandos kruskalwallis y
multcompare.
Para evaluar si el modelo matemático propuesto describe adecuadamente los datos
experimentales, se realizó una prueba F de una cola que parte de las siguientes hipótesis:
2
,
0 2
exp,
2
,
1 2
exp,
: 1
: 1
i
i
i
i
residuales F
datos F
residuales F
datos F
SH
S
SH
S
Donde: Fi está dado para i = 1,2,…,12, s2
residuales es la varianza de los residuales (desviaciones de los datos
experimentales con respecto a los valores calculados por el modelo matemático) y se calcula para cada
variable de respuesta medida y s2datos exp es la varianza de la serie experimental de cada variable de
respuesta medida. Para estos análisis se utilizaron los comandos vartest y vartest2 de Matlab. Con las
varianzas de los residuales obtenidos para cada serie experimental se calcularon los porcentajes de cada
una respecto a la serie que dio la mayor varianza de residuales a fin de mostrar el ajuste de los datos
experimentales al modelo matemático propuesto de forma más simple.
3. RESULTADOS Y DISCUSIÓN
3.1. Efecto de la relación C/N y sulfato de cobre sobre la producción de biomasa y polisacáridos
Las tres especies de hongos (C. versicolor, P. ostreatus y L. edodes) probadas en el diseño
experimental, fueron inoculadas e incubadas bajo las mismas condiciones ambientales en las doce
formulaciones de sustrato. El intervalo de la relación C/N en las doce formulaciones de sustrato evaluadas
fue de 50 a 140 con una variación en los contenidos de nitrógeno de 0,29 a 0,8% en base seca y dos
niveles de CuSO4. Las tres especies de hongos se desarrollaron bien en todas las formulaciones evaluadas,
sin aparente inhibición en el crecimiento de los tres organismos de trabajo en las cuales, probablemente
los contenidos de nitrógeno de las formulaciones estimularon el crecimiento de los macromicetos durante
la fase vegetativa y los dos niveles de sulfato de cobre no afectaron el crecimiento de los hongos, de hecho
es evidente el incremento en el contenido de biomasa como se observa en las Figuras 1 y 2 en todos los
casos desde el inicio de la fase de incubación. Empero se debe tener en cuenta que la variación del

125
contenido de nitrógeno y de la fuente de carbono se realizó empleando los mismos materiales y no
modificando las fuentes de nitrógeno y de carbono.
Las tres especies de hongos mostraron crecimiento homogéneo durante los 49 días de
fermentación sobre la mayoría de las formulaciones, con incremento sostenido para nueve de las 12
formulaciones, exceptuando las formulaciones F3, F4 y F5, las cuales presentaron un ascenso más lento.
Sin embargo, todas las combinaciones mostraron bajos períodos de adaptación y tendencia a la fase
estacionaria después del día 30 de incubación, tiempo en el que se había alcanzado la colonización
completa. Las combinaciones de las tres especies de hongos con la F3 fueron las que presentaron un
crecimiento diferente al de las demás, ya que como se observa en la Figura 1C la biomasa parece estar en
la fase logarítmica hasta el día 49 de incubación, especialmente en la combinación con L. edodes. Del
mismo modo, es importante tener en cuenta que las diferentes especies de hongos pueden tener
dependencia de las diferentes fuentes de carbono y esto provocar en ellos la variación en su crecimiento y
en la formación de los tipos de polisacáridos presentes en su pared celular y los exopolisacáridos que
excretan al medio. Según Xiao et al. (2010) la producción de polisacáridos y el crecimiento de la biomasa
de las diferentes especies de hongos dependen directamente de la fuente de carbono suministrada, lo que
sugiere que la composición del medio puede influir en la composición y el mecanismo de producción
tanto de los intra como de los exopolisacáridos para una determinada especie de hongo. Asimismo, la
cantidad y fuente de nitrógeno en la producción de los polisacáridos de los hongos y su influencia en su
crecimiento y desarrollo es definitivo, ya que el nitrógeno es un componente esencial de las proteínas y
ácidos nucleicos y su deficiencia afecta el crecimiento de los hongos y los metabolitos que producen
durante su crecimiento y desarrollo (Xiao et al., 2006). La fuente de nitrógeno utilizada en este trabajo fue
únicamente la proveniente de materiales orgánicos naturales, ya que variando los contenidos de cascarilla
de café y salvado de maíz en las formulaciones de sustrato se obtuvo la variación de las relaciones C/N,
con un intervalo en los contenidos de nitrógeno entre 0,29 y 0,8% en base seca, razón por la cual
probablemente las tres especies de hongos crecieron sobre todas las formulaciones evaluadas. Sin
embargo, el crecimiento de las tres especies de hongos no fue igual para las 12 formulaciones, ni la
producción de polisacáridos como se observa en las Figuras 1, 2, 3 y 4. De hecho, las combinaciones que
mostraron mejor producción de polisacáridos fueron para C. versicolor con F2, F4, F7 y F11, pero no fue
el C. versicolor el que mayor cantidad de biomasa produjo para estas mismas formulaciones; ya que fue el
L. edodes el hongo que mayor cantidad de biomasa produjo para esas mismas formulaciones. Empero, es
necesario tener en cuenta que en este trabajo se está determinando la cantidad total de polisacáridos de
sustratos sólidos colonizados con tres especies de hongos y no se está diferenciando entre los intra y los
exopolisacáridos, ni en la estructura de los mismos, siendo este aspecto relevante al momento de definir la
especie de hongo para la producción de polisacáridos.
En las combinaciones de P. ostreatus con F7, F9, F10 y F12 se observó una muy baja producción
de polisacáridos durante todo el tiempo de la fermentación como se observa en la Figura 3, del orden de
20 mg/gss o menos; también para estas cuatro combinaciones se detectó una menor producción de
biomasa en comparación con las demás combinaciones, del orden de 300 mg/gss (g sustrato sólido), lo
que permite inferir que sí hay una relación directa entre la producción de biomasa y la producción de
polisacáridos. Sin embargo, no hay un factor común bien definido al que se le pueda atribuir la baja
producción de biomasa y de polisacáridos para P. ostreatus en estas cuatro combinaciones, ya que las
relaciones C/N de cada una de estas formulaciones fueron F7:85, F9:55, F10:51 y F12:111, para las cuales
habría cierta relación entre los contenidos de F9 y F10, pero no podría hacerse la misma relación para F7 y
F12. De igual modo no fue claro el efecto de la sal de cobre, ya que para las formulaciones impares se
adicionó 0,1596% y para las formulaciones pares 0,0798% en base seca y en este caso las bajas
producciones se reflejaron en formulaciones con ambas participaciones de sulfato de cobre. No obstante,
debe considerarse la composición de los sustratos utilizados en este estudio, su procedencia y la alta
variabilidad en la calidad de las materias primas, ya que éstas fueron residuos lignocelulósicos, pudiendo
haber generado cambios en la composición química y características físicas de los sustratos; además de
tener en cuenta que durante la fermentación pudieron haberse producido sustancias intermedias que hayan

126
generado una disminución en la velocidad de crecimiento de este hongo sobre estas formulaciones de
sustrato y por ende la disminución en la velocidad de formación de los polisacáridos.
A pesar de todo el esfuerzo que se ha dado para explorar el efecto de las condiciones de cultivo en
la producción de polisacáridos de hongos, la mayoría de las investigaciones giran en torno a la producción
de exopolisacáridos por fermentación sumergida y muy pocas en torno a la producción de
intrapolisacáridos y son aún menos las referidas a la producción de polisacáridos contenidos en el micelio
de los sustratos sólidos colonizados por macromicetos (Lee et al., 1999; Shu y Lung, 2003; Lee et al.,
2004; Wu et al., 2004; Wu et al., 2006; Lee et al., 2007b). Sin embargo, es razonable esperar que la
producción de ambos tipos de polisacáridos, IPS y EPS podrían estar vinculados uno con otro, por la
misma dependencia con el crecimiento de la biomasa (Lama et al., 1996). Por lo tanto, las características
físico-químicas de los medios de cultivo y condiciones medioambientales siempre deberán ser tenidas en
cuenta a fin de satisfacer la producción de intra y exo-polisacáridos acorde con las necesidades
particulares de la especie de hongo a utilizar.
Figura 1. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6
Figura 2. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12

127
3.2. Producción de polisacáridos en la mejor combinación resultante del diseño experimental
Los datos experimentales obtenidos en el presente estudio fueron sometidos a un proceso de
selección a fin de lograr la mejor combinación especie de hongo fórmula del diseño experimental
bifactorial respecto a la producción de polisacáridos, el cual se realizó por etapas. Primero se hizo un
análisis de varianza, ANOVA a tres vías (especie, fórmula, tiempo) para los polisacáridos obtenidos
durante 15 tiempos en los 49 días de fermentación; se obtuvieron diferencias significativas para todas las
combinaciones con un valor-p < 0,05. Posteriormente, se realizó un análisis por especie y una selección de
los máximos de cada una de las cantidades de polisacáridos obtenidos y un análisis comparativo de los
máximos (empleando la función multcompare) y aplicando una prueba de Kruskal-Wallis, obteniéndose
diferencias significativas con valor-p < 0,05. De este análisis comparativo se obtuvo que la mejor
combinación para la producción de polisacáridos fue C. versicolor – F11 seguida de las combinaciones
con F2, F4 y F7. A estas cuatro combinaciones se les realizó un análisis comparativo a fin de seleccionar
de entre las cuatro combinaciones la de mejor comportamiento respecto a la producción de polisacáridos y
aplicar un modelo matemático que describa los datos experimentales de producción de biomasa, de
polisacáridos, de producción y consumo de azúcares reductores, de consumo de celulosa y de degradación
de lignina. Sin embargo, el análisis comparativo no presentó diferencia significativa (valor-p = 0.212), por
lo que se aplicó el modelo matemático a las cuatro combinaciones de C. versicolor.
Figura 3. Polisacáridos de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis formulaciones de
sustratos. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 49 días de fermentación en estado sólido
En la Tabla 2 se presentan los datos de las producciones de biomasa, polisacáridos y la relación de
la producción de polisacáridos con la producción de biomasa (producción específica de polisacáridos) a
los 49 días de fermentación (final del período), en la que se observó que la mayor producción de biomasa
por combinación fue de 377.133, 353.795 y 366.653 mg/gss para C. versicolor – F11, P. ostreatus – F10
y L. edodes – F6 respectivamente; la máxima producción de polisacáridos fue de 96.85 mg/gss, 90.78
mg/gss y 87.454 mg/gss para las combinaciones C. versicolor –F2, P. ostreatus – F6 y L. edodes – F3
respectivamente. Sin embargo, la producción específica de polisacáridos no correspondió estrictamente a
las combinaciones que arrojaron las máximas producciones de polisacáridos, por ejemplo para la
combinaciones de C. versicolor, la máxima producción de polisacáridos específica fue para F3 con 0.385,
mientras que para F2 se obtuvo 0.297, seguido de F11 con 0.247; de igual forma, para P. ostreatus la
mayor producción específica de polisacáridos fue para la F5 con 0.399, seguido de F6 con 0.259; para L.
edodes la mayor producción específica fue para F3 con 0.407 y F5 con 0.394. De lo anterior, se puede
inferir que además de la importancia que tiene la cantidad de polisacáridos producidos, otro importante
parámetro es la relación de producción de polisacáridos respecto a la cantidad de biomasa producida, al

128
momento de tomar decisiones en torno a la producción de polisacáridos por fermentación en fase sólida
buscando mejorar las condiciones de proceso y los rendimientos.
Como ya se mencionó, la producción de polisacáridos a partir del micelio obtenido de
fermentación en fase sólida no ha sido ampliamente estudiada, probablemente porque cuando se produce
un sustrato sólido para sembrar alguna especie de macromiceto se busca la producción de los cuerpos
fructíferos, en los que los contenidos de polisacáridos son bastante altos. Sin embargo, en este trabajo se
determinó el contenido de polisacáridos de los sustratos sólidos colonizados con tres especies de hongos
en 15 tiempos durante 49 días de fermentación a fin de mostrar la importancia que tienen los
basidiomicetos en la obtención de nuevos productos como son los alimentos para animales, en este caso en
particular, para rumiantes. Varios investigadores coinciden en que es necesario el desarrollo de nuevos
productos que mejoren la calidad de la alimentación de animales de cría, como los bovinos, y que además
estos alimentos deben ser coadyuvantes a fin de incrementar la salud del animal (Koutinas et al., 2004;
Graminha et al., 2008). Lo anterior demuestra la importancia de este estudio, ya que las materias primas
utilizadas para las formulaciones de los sustratos fueron residuos lignocelulósicos, los cuales son
potencialmente utilizados como alimento para rumiantes. Los sustratos fueron colonizados por las tres
especies de hongos de pudrición blanca obteniendo como resultado una conversión de los materiales
lignocelulósicos que componen los sustratos como se observa en las Figuras 5, 6 y 7. Asimismo, los
componentes de la fibra del material lignocelulósico presente en los sustratos fueron degradados. Los
polímeros de celulosa y hemicelulosa fueron degradados por acción de las enzimas hidrolíticas de los
hongos a azúcares reductores a fin de conseguir la fuente de energía necesaria para su crecimiento y
desarrollo (Ângelo, 2004). La lignina, fue degradada por óxidorreductasas excretadas por estos hongos
que, como agentes causantes de pudrición blanca, son los únicos hongos capaces de degradarla
completamente, facilitando en el proceso el acceso a la celulosa (Kirk y Farrel, 1987b). Sin embargo, estos
polímeros no fueron consumidos por el hongo en su totalidad luego de 49 días de fermentación (ver
Figuras 5, 6 y 7), hecho que se hace favorable para que los sustratos colonizados por estos hongos sean
potencialmente utilizados en alimentación de rumiantes, ya que en estos sustratos permanece una
apreciable cantidad de celulosa y hemicelulosa y un contenido bajo de lignina.
El aumento de la digestibilidad mediada por la acción enzimática del hongo, el incremento del
contenido de proteína con la colonización fúngica al cabo de 49 días de fermentación y la presencia de
polisacáridos fúngicos que pueden favorecer la salud de los animales, contribuyen a mejorar las
características de estos residuos lignocelulósicos para satisfacer las necesidades de alimentación bovina.
Según Weinberg (2000) el uso de sustratos agotados de la producción de hongos macromicetos, como
Pleurotus sp., L. edodes, Ganoderma sp., Agaricus bisporus y otros o el empleo de hongos de pudrición
blanca para deslignificar los materiales lignocelulósicos son una alternativa para la producción de
alimento para rumiantes, debido a que durante la fermentación en fase sólida con estos basidiomicetos, los
materiales lignocelulósicos exhiben una bioconversión favorable en términos de disminución de lignina y
producción de azúcares reductores que luego son utilizados por el animal en su digestión.
Tabla 2. Producción máxima de polisacáridos en doce fórmulas y las tres especies de hongos: C.
versicolor, P. ostreatus y L. edodes durante 49 días de incubación
Fórmula
C. versicolor P. ostreatus L. edodes
Bio
(mg/gss)
Pol
(mg/gss)
Rel.
(Pol/Bio)
Bio
(mg/gss)
Pol
(mg/gss)
Rel.
(Pol/Bio)
Bio
(mg/gss)
Pol
(mg/gss)
Rel.
(Pol/Bio)
F1 328,388 63,029 0,192 333,648 65,681 0,197 345,485 40,740 0,118
F2 323,785 96,085 0,297 332,333 70,669 0,213 346,142 63,147 0,182
F3 208,711 80,408 0,385 225,150 49,489 0,220 214,629 87,454 0,407
F4 351,902 91,057 0,259 334,604 88,207 0,264 309,318 57,763 0,187
F5 240,932 59,426 0,247 222,520 88,721 0,399 220,547 86,979 0,394
F6 360,808 76,726 0,213 350,711 90,780 0,259 366,653 75,063 0,205

129
F7 336,936 61,663 0,183 289,592 26,409 0,091 348,115 87,454 0,251
F8 346,910 61,083 0,176 345,485 47,589 0,138 329,703 37,534 0,114
F9 351,403 64,458 0,183 349,430 6,694 0,019 344,169 50,796 0,148
F10 341,424 40,911 0,120 353,795 24,430 0,069 323,127 73,599 0,228
F11 377,133 92,997 0,247 352,060 52,617 0,149 348,115 51,390 0,148
F12 362,138 61,801 0,171 331,671 23,282 0,070 350,986 78,705 0,224
Figura 4. Producción de polisacáridos de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes en seis
formulaciones de sustratos: A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 en 49 días de fermentación en estado sólido
Figura 5. Degradación de celulosa por tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 49 días de fermentación en estado
sólido

130
Figura 6. Degradación de hemicelulosa por tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre
seis formulaciones de sustratos. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 49 días de fermentación en estado sólido
Figura 7. Degradación delignina por tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 en 49 días de fermentación en estado sólido
3.5. Modelamiento matemático
Basado en los datos experimentales obtenidos de polisacáridos durante los 49 días de
fermentación en estado sólido para las tres especies de hongos basidiomicetos de pudrición blanca: C.
versicolor, P. ostreatus y L. edodes, se propuso un modelo matemático compuesto de 6 ecuaciones
diferenciales (ver Tabla 3) a fin de ajustar los datos experimentales de todas las combinaciones resultantes
para C. versicolor, ya que éste fue el hongo que mayor producción de polisacáridos mostró en todas las
combinaciones medidas. De hecho, en este trabajo se presentan los resultados del modelamiento arrojados
para las combinaciones C. versicolor con las formulaciones F2, F4, F7 y F11, ya que estas combinaciones
fueron las que presentaron la mayor producción de polisacáridos. De igual forma, para comparar el ajuste
de los datos experimentales al modelo matemático propuesto se planteó una prueba F de una cola, con la
cual se obtuvieron las varianzas de los residuales entre los datos experimentales y el modelo,
individualmente para cada variable medida, pero haciendo la comparación de cada ecuación aplicada para
todas las formulaciones de F1 a F12 en la misma especie, en este caso C. versicolor, lo que permitió
probar la bondad del ajuste del modelo en términos de la mejor representación de los datos experimentales
obtenidos.

131
Con el modelo matemático propuesto se plantea la descripción de producción de biomasa, los
azúcares reductores como producto intermedio consumido durante la misma fermentación, el consumo de
celulosa, hemicelulosa y la degradación de lignina. Asimismo, los datos experimentales obtenidos en esta
investigación serán ajustados al modelo planteado a fin de contribuir con nuevas propuestas en el camino
al planteamiento de modelos matemáticos más complejos que permitan la descripción de los procesos de
fermentación en fase sólida con fines industriales. Varias ecuaciones cinéticas de complejidad variable,
tales como la ecuación lineal, logística, de dos fases o la ecuación de Monod han sido utilizadas por
diferentes investigadores a fin de buscar la descripción de la producción de biomasa en procesos de
fermentación en estado sólido (Sangsurasak et al., 1996; Viccini et al., 2001). De las expresiones
matemáticas utilizadas, la más frecuente es la ecuación logística, debido probablemente a la forma simple
de la expresión. El modelo propuesto consignado en la Tabla 3 para la descripción de la biomasa es la
ecuación (1), la cual corresponde a la ecuación logística modificada por Mitchell et al. (1999). Para
describir la variación de los azúcares reductores en el tiempo se propuso la ecuación (2) que correspondió
a un factor de producción constante que afecta la derivada de la velocidad de producción de biomasa,
debido a que los azúcares reductores son considerados un producto de la fermentación que es consumido
por el mismo hongo como fuente de energía y su variación se ajustó más a la velocidad de variación de la
biomasa que a la variación de la biomasa misma. Para la degradación de lignina y los consumos de
hemicelulosa y celulosa en el tiempo de fermentación se plantearon las ecuaciones (3), (4) y (5) cuyo
planteamiento se realizó en función de las concentraciones de las enzimas específicas responsables de la
degradación de cada uno de los sustratos. La producción de los polisacáridos fue expresada por la
ecuación de Luedeking y Piret (1959) dada por la ecuación (6) que representa la producción de
metabolitos en procesos de fermentación. Los parámetros de cada una de las ecuaciones consignadas en la
Tabla 3 se describen en la Tabla 4.
La fermentación en estado sólido es un proceso heterogéneo que se ve afectado además por las
condiciones ambientales. La dificultad que representa la determinación del crecimiento de la biomasa, el
consumo de los sustratos y la imposibilidad de determinar todas las sustancias intermedias que pueden
generarse como producto de las reacciones bioquímicas durante las fases de incubación ha dificultado la
descripción de los procesos de fermentación en estado sólido y su escalamiento cuando se pretende aplicar
estos procesos en producción industriales de alguna sustancia de interés (Pal et al., 1995b; Sarikaya y
Ladisch, 1997; Pandey et al., 2000; Moreira et al., 2003). Asimismo, afecta también, la heterogeneidad de
las materias primas utilizadas para el crecimiento y desarrollo de los hongos de pudrición blanca,
compuestas principalmente del complejo lignocelulósico (Viccini et al., 2001; Pradeep y Datta, 2002).
Los hongos basidiomicetos como los utilizados en el presente estudio contienen quitina en su
pared celular. Debido a que en los procesos de fermentación en estado sólido no es posible determinar
directamente la cantidad de biomasa, se determinó el contenido de quitina a través de la cuantificación de
N-acetil-D-glucosamina (NAGA) en todos los tiempos de la fermentación. Además, se le determinó el
contenido de NAGA al micelio seco obtenido de cultivo líquido a cada una de las tres especies de trabajo,
obteniendo como resultado 15,83% (p/p) en P. ostreatus, 14,15% (p/p) en C. versicolor y 10,08% (p/p) en
L. edodes. Estos porcentajes de NAGA fueron utilizados para calcular la cantidad de biomasa de los
sustratos sólidos en cada uno de los tiempos de fermentación; por lo que estos datos de biomasa fueron los
que se sometieron al modelamiento con la ecuación (1). Se aplicó el modelo matemático propuesto de seis
ecuaciones a todos las combinaciones de C. versicolor a fin de determinar la capacidad del modelo para
describir diferentes datos experimentales obtenidos para una misma especie. En las Figuras 8, 9, 10 y 11
se muestran el ajuste de los datos experimentales en contraste con el modelo matemático para las
combinaciones de C. versicolor con F2, F4, F7 y F9, las cuales fueron seleccionadas como las de mayor
producción de polisacáridos durante los 49 días de fermentación en estado sólido. Los porcentajes de
ajuste para la descripción del modelo matemático propuesto sobre los datos experimentales de las cuatro
combinaciones de C. versicolor con F2, F4, F7 y F11 se resumen en la Tabla 6; en la que se observa que el
mejor ajuste de los datos experimentales obtenidos para cada variable medida fue: biomasa, F7 (98%);
azúcares reductores, F7 (90%); polisacáridos, F7 (92%); hemicelulosa, F2 y F7 (60%); celulosa, F2
(95,7%) y lignina, F2 (88%). Por lo anterior, el modelo matemático propuesto no hace una descripción

132
apropiada para todas las variables medidas, ni para todas las series de datos experimentales, pero sí es una
aproximación a la búsqueda de modelos matemáticos que permitan la descripción de procesos de
fermentación en fase sólida para producir diferentes tipos de biosustancias o la trasformación de diversos
materiales con fines de escalamiento industrial.
Tabla 3. Modelo matemático para la descripción de producción de biomasa, consumo de la matriz lignocelulósica y
producción de polisacáridos
Ecuación Descripción
. 1
n
b bm b
bm
dC CC
dt C
(1) Biomasa
. . . 1 1 .AR
n
b bp m
bm
dC Cq n
dt C
dC
dt
(2)
Azúcares
reductores
. .LL lac MnP
dCk C C
dt (3) Lignina
.HMHM ENX
dCk C
dt (4) Hemicelulosa
. .CC ENG EXG
dCk C C
dt
(5) Celulosa
. .POL bPOL b
dC dCk b
dt dC
t (6) Polisacáridos
Tabla 4. Parámetros de las ecuaciones del modelo matemático.
Parámetro Significado
µm Velocidad específica de crecimiento de la biomasa (día-1)
Cbm Concentración de biomasa máxima (mg/gss)
N
n < 1 el organismo es relativamente sensible a la auto-inhibición y
ésta ocurre para valores muy bajos de Cb
n = 1 ecuación logística
n > 1 el organismo es relativamente resistente a la auto-inhibición
y ésta ocurre solo cuando Cb ≈ Cbm Cb0 Concentración de biomasa máxima (mg/gss)
qp Coeficiente de producción constante de azúcares reductores (día)
CAR0 Concentración inicial de azúcares reductores (mg/gss)
kL Coeficiente de degradación de lignina (mg.gss/día.U)
CL0 Concentración inicial de lignina (mg/g ss)
kHM Coeficiente de consumo de hemicelulosa (mg/día.U)
CHM0 Concentración inicial de hemicelulosa (mg/gss)
kC Coeficiente de consumo de celulosa (mg.gss/día.U2)
CC0 Concentración inicial de celulosa (mg/g ss)
kPOL Coeficiente de producción de polisacáridos (mg/mg)
B Velocidad específica de producción de polisacáridos (día-1)

133
Figura 8. Ajuste de datos experimentales de la producción de biomasa y de polisacáridos y la degradación de
celulosa, hemicelulosa y lignina para la combinación C. versicolor – F2 durante 49 días de fermentación en fase
sólida
Figura 9. Ajuste de datos experimentales de la producción de biomasa y de polisacáridos y la degradación de
celulosa, hemicelulosa y lignina para la combinación C. versicolor – F4 durante 49 días de fermentación en fase
sólida

134
Figura 10. Ajuste de datos experimentales de la producción de biomasa y de polisacáridos y la degradación de
celulosa, hemicelulosa y lignina para la combinación C. versicolor – F7 durante 49 días de fermentación en fase
sólida
Figura 11. Ajuste de datos experimentales de la producción de biomasa y polisacáridos y la degradación de celulosa,
hemicelulosa y lignina para la combinación C. versicolor – F11 durante 49 días de fermentación en fase sólida
Aunque existen numerosas investigaciones previas acerca de la producción de polisacáridos
fúngicos, éstas giran en torno a los procesos de fermentación sumergida y no a la producción de
polisacáridos de la fase vegetativa en procesos de fermentación en fase sólida, en la que además de
producir polisacáridos también ocurre la degradación de materiales lignocelulósicos. Se requiere aun
mayor información sobre las cinéticas de formación de biomasa, de producción de polisacáridos, del
consumo de nutrientes y degradación de la lignina que permitan la descripción del proceso global y
posterior escalado para la obtención de alguno de los productos de interés como los que se plantean en
este estudio. El planteamiento de las ecuaciones para la descripción de la cinética de cada una de las
variables medidas se realizó bajo la premisa de darle a cada expresión matemática algún sentido biológico
respecto al compuesto a describir. Las expresiones propuestas representaron aceptablemente cada una de
las variables medidas, ya que durante los procesos de fermentación en fase sólida cuando se utilizan como
sustratos mezclas de materiales lignocelulósicos pueden producirse una gran cantidad de sustancias
intermedias producto de las reacciones bioquímicas. Contemplar productos intermedios originados por la
acción del metabolismo de estos hongos, contribuiría a una interpretación más acabada de los complejos
fenómenos que ocurren durante el crecimiento de los macromicetos en los sustratos lignocelulósicos.
Tabla 5. Parámetros de ajuste de los datos experimentales al modelo para cuatro combinaciones de C. versicolor para
la producción de polisacáridos
Parámetro F2 F4 F7 F11
µm 41.664 43.2515 4.9462 28.6281
Cbm 314.1649 423.6859 348.12 357.8715
n 0.005335 0.002518 0.002282 0.004069
Cb0 47.8261 12.3485 34.7 27.9885
qp 0.1 0.7696 0.2686 0.7
CAR0 6.5 7.5 8 5.2
kL 0.02072 0.006017 0.007841 0.009598
CL0 211.62 194,14 200 207.25
kHM 0,04294 0.03174 0.05629 0.06197
CHM0 210.98 200.87 199.96 205.55
kC 0.005306 0.06209 0.01002 0.01059
CC0 493.29 475.65 499.35 490.041
kPOL 0.1413 0.06512 0.05862 0.1093
b 0.005059 0.0063 0.004031 0.0051

135
Tabla 6. Ajuste (%) de la descripción del modelo matemático a los datos experimentales de cuatro combinaciones de
C. versicolor con cuatro formulaciones para la producción de polisacáridos en fermentación en estado sólido
Fórmula Biomasa Azúcares
reductores Polisacáridos Celulosa Hemicelulosa Lignina
F2 91 55 25 95,7 60 88
F4 1 72,5 60 70 20 40,5
F7 98 90 92 85,7 60 64
F11 80 61 1 1 6,6 71,4
4. Conclusiones
En el presente estudio, las tres especies de hongos de pudrición blanca probadas sobre las doce
formulaciones de sustrato propuestas crecieron bien durante los 49 días de fermentación en fase sólida,
por lo que se concluye que tanto la relación C/N como las concentraciones de CuSO4 no afectaron
drásticamente el crecimiento y desarrollo de ninguno de los tres hongos. Sin embargo, de la evaluación
detallada de los datos experimentales sí se observaron diferencias en la producción de los polisacáridos y
en la producción específica de los mismos durante los 49 días de fermentación, en la que se observó que la
mayor producción de biomasa fue para la combinación C. versicolor – F11 (377.133 mg/gss), aunque esta
combinación no haya sido la de mayor producción de polisacáridos, pues la combinación C. versicolor –
F2 fue la que arrojó la mayor cantidad de polisacáridos con 96,085 mg/gss. Asimismo, la combinación que
mayor conversión mostró de polisacáridos referida a la cantidad de biomasa seca fue L. edodes – F2,
aspecto que puede ser de utilidad al momento de escalar los procesos.
El modelo matemático planteado en este estudio para la representación de los procesos de
fermentación en estado sólido se realizó buscando describir el desarrollo de tres hongos de pudrición
blanca sobre sustratos lignocelulósicos a fin de buscar puntos generales de interpretación de estos
procesos que por ser tan heterogéneos se hacen complejos y carecen de expresiones matemáticas que
permitan el escalamiento de los mismos con fines industriales, evitando hacer experimentación individual
de cada proceso.
Agradecimientos
Los autores agradecen a la Vicerrectoría de Investigaciones y Postgrados de la Universidad de
Caldas (Colombia) por la financiación del presente trabajo y al Instituto de Biotecnología Agropecuaria de
la misma institución por el apoyo material y de infraestructura. Asimismo, los autores expresan sus
agradecimientos a la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires
(Argentina) por su apoyo y asesoría durante la realización de este trabajo.
Referencias
Amaral A, Carbonero E, Simao R, Kadowaki M, Sassaki G, Osaku C, Gorin P, Lacomini M (2008) An unusual water-soluble B-
glucan from the basidiocarp of the fungus Ganoderma resinaceum. Carbohydrate Polymers 72: 473-478.
Ângelo AS (2004) Enzimas hidrolíticas. En Esposito, Azevedo JI (Eds.) Fungos: Uma Introducâo à biologia, bioquimica e
biotecnologia. Universidade de Caxias do Sul,.
Catley BJ (1992) The biochemistry of some fungal polysaccharides with industrial potencial. En Mukerji KG (Ed.) Handbook of
applied mycology: fungal biotechnology. Marcel Dekker,. New York.
Chang ST, Miles PG (2004) Mushrooms Cultivation, Nutritional Value, Medicinal effect, and Enviromental Impact. CRC Press.
New York. 451 p.
Chen YJ, Shiao MS, Lee SS, Wang SY (1997) Effects of Cordyceps sinensis on the proliferation and differentiation of human
leukemic U937 cells. Life Science 60(23): 49-59.
Dormand JR, Prince PJ (1980) A family of embedded Runge-Kutta formulae. Journal of Computational and Applied Mathematics
6: 19-26.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related
substances. Analytical Chemistry Engineering Journal 28(3): 350-356.
Enshasy HE (2010) Immunomodulators. En Hofrichter M (Ed.) The Mycota. Springer-Verlag. Berlin, 477 p.

136
Graminha EBN, Gonc¸alves AZL, Pirota RDPBB, M.A.A., Da Silva R, Gomes E (2008) Enzyme production by solid-state
fermentation: Application to animal nutrition. Animal Feed Science and Technology 144: 1-22.
Hamlyn PF, Schmidt RJ (1994) Potential therapeutic application of fungal filaments in wound management. Mycologist 8: 147-
152.
Hsieh C, Liu C-J, Tseng M-H, Lo C-T, Yang Y-C (2006) Effect of olive oil on the production of mycelial biomass and
polysaccharides of Grifola frondosa under high oxygen concentration aeration. Enzyme and Microbial Technology 39:
434-439.
Hsieh C, Wang H-L, Chena C-C, Hsub T-H, Tseng M-H (2008) Effect of plant oil and surfactant on the production of mycelial
biomass and polysaccharides in submerged culture of Grifola frondosa. Biochemical Engineering Journal 38: 198-205.
Ishikawa K, Majima T, Ebina T (1992) Anti-tumour effect of a Coriolus preparation, PSK: Induction of macrophage chemotactic
factor (MCF) in spleens of tumour bearing mice. Biotherapy 5: 251-258.
Kiho T, Ookubo K, Usui S, Ukai S, Hirano K (1999) Structural features and hypoglycemic activity of a polysaccharides (CS-F10)
from the cultured mycelium of Cordyceps sinensis. Biological and Pharmaceutical Bulletin 22(9): 66-70.
Kirk TK, Farrel RL (1987) Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology 41: 465-
505.
Kjeldahl J (1883) Neue Methode Zùr Bestimmung der Stickstoffs in Organichen Kòrpen. Zeitschrift fur Analytische Chemie 22:
366-382.
Kobayashi H, Matsunaga K, Oguchi Y (1995) Antimetastatic Effects of P5K (Krestin), a Protein-bound Polysaccharide Obtained
from Basidiomycetes: An Overview. Cancer Epidemiology, Biomarkers & Prevention 4: 275-281.
Koh JH, Kim JM, Chang UJ, Suh HJ (2003) Hypocholesterolemic effect of hotwater extract from mycelia of Cordyceps sinensis.
Biological and Pharmaceutical Bulletin 26: 84-87.
Komura DL, Ruthers AC, Carbonero ER, Alquini G, Rosa MC, Sassaki GL, Lacomini M (2010) The origin of mannans found in
submerged culture of basidiomycetes. Carbohydrate polymers 79: 1052-1056.
Koutinas AA, Wang R, Webb C (2004) Restructuring Upstream Bioprocessing: Technological and Economical Aspects for
Production of a Generic Microbial Feedstock From Wheat. Published online in Wiley InterScience: 1-15.
Kupfahl C, Geginat G, Hof H (2006) Lentinan has a stimulatory effect on innate and adaptive immunity against murine Listeria
monocytogenes infection. International Immunopharmacology 6: 686- 696.
Lama L, Nicolaus B, Calandrell V, Manca M, Romano I, Gambacorta AM, Arambarri (1996) Effect of growth conditions on
endo- and exopolymer biosynthesis in Anabaena cylindrica 10C. Phytochemistry 42: 655-664.
Lee B, Bae J, Pyo H, Choe T, Kim S, Hwang H, Yun J (2004) Submerged culture conditions for the production of mycelial
biomass and exopolysaccharides by the edible basidiomycete Grifola frondosa. Enzyme and Microbial Technology 35:
369-376.
Lee K, Lee S, Lee H (1999) Bistage control of pH for improving exopolysaccharide production from mycelia of Ganoderma
lucidum in an air-lift fermentor. Journal of Bioscience and Bioengineering 88: 646-650.
Lee WY, Park Y, Ahn JK, Ka KH, Park SY (2007) Factors influencing the production of endopolysaccharide and
exopolysaccharide from Ganoderma applanatum. Enzyme and Microbial Technology 40 249-254.
Leterme P. (2010). Análisis de alimentos y forrajes, in Protocolos de LaboratorioUniversidad Nacional de Colombia Sede
Palmira: Palmira. 66 pp.
Lindequist U, Niedermeyer T, Julich W-D (2005) The pharmaceutical potential of mushrooms. Electronic Centralised Aircraf
Monitor (eCAM) 2: 285-299.
Liu F, Fung MC, Ooi VEC, Chang ST (1996) Induction in the mouse of gene expression of immunomodulating cytokines by
mushroom polysaccharide-protein complexes. Life Science 58(1): 795-803.
Luedeking R, Piret E (1959) Transient and steady states in continuous fermentation. Theory and experiment. Journal of
Biochemical and Microbiological Technology and Engineering 1: 431-459.
Maziero R, Cavazzoni V, Ramos-Bononi VL (1999) Screening of Basidiomycetes for the production of exopolisaccharides and
biomass in submerged culture. Revista de Microbiologia 30: 77-84.
Miller GL (1959) Use of DinitrosaIicyIic Acid Reagent for Determination of Reducing Sugar. Analytical Chemistry 31(3): 426-
428.
Mitchell D, Stuart D, Tanner R. (1999). Solid-State fermentation - Microbial growth kinetics, in The Encyclopedia of Bioprocess
Technology: Fermentation, Biocatalysis and bioseparation, Flickinger M, Drew S, EditorsWiley: New York. 2407-
2429
Mizuno T (1999) The extraction and development of antitumouractive polysaccharides from medicinal mushrooms in Japan.
International Journal Medicine Mushrooms 1: 9-29.
Mohammad-Fata M, Hossein M, Shirin G, Ghorban-Ali H (2007) Immunomodulating and anticancer agents in the realm of
macromycetes fungi (macrofungi). International Immunopharmacology 7: 701-724.
Moreira MT, Feijoo G, Lema JM (2003) Fungal bioreactors: Applications to white-rot fungi. Reviews in Environmental Science
and Bio/Technology 2: 247-259.
Nakamura K, Yamaguchi Y, Kagota S, Shinozuka K, Kunitomo M (1999) Activation of in vivo Kupffer cell function by oral
administration of Cordyceps sinensis in rats. . Japan Journal Pharmacology 79: 508-508.
Ooi V, Liu F (2000) Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Current Medicine
Chemistry 7: 715-728.

137
Pal M, Calvo A, Terron MC, Gonzalez AE (1995) Solid-state fermentation of sugarcane bagasse with Flammulina velutipes and
Trametes versicolor. World Journal Microbiology and Biotechnology 11: 541-545.
Pandey A, Soccol CR, Mitchell D (2000) New developments in solid state fermentation: I-bioprocesses and products. Process
Biochemistry 35: 1153–1169.
Park J-P, Kim S-W, Hwang H-J, Cho Y-J, Yun J-W (2002) Stimulatory effect of plant oils and fatty acids on the exo-biopolymer
production in Cordyceps militaris. Enzyme and Microbial Technology 31: 250-255.
Plassard C, Mousain D, Salsac L (1982) Estimation of mycelial growth of basidiomycetes by means of chitin determination.
Phytochemistry 21: 345-348.
Pradeep V, Datta M (2002) Production of ligninolytic enzymes for decolorization by cocultivation of white-rot fungi Pleurotus
ostreatus and Phanerochaete chrysosporium under solid-state fermentation. Applied Biochemistry Biotechnology 102-
103: 109-118.
Sangsurasak P, Nopharatana M, Mitchell D (1996) Mathematical modeling of the growth of filamentous fungi in solid-state
fermentation. Journal Science Industrial Resources 55: 333-342.
Sarikaya A, Ladisch M (1997) An Unstructured Mathematical Model for Growth of Pleurotus ostreatus on Lignocellulosic
Material in Solid-State Fermentation Systems. Applied Biochemistry and Biotechnology 62: 72-85.
Shu C, Lung M (2003) Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia
camphorata in batch cultures
Process Biochemestry 39: 931-937.
Tang YJ, Zhong JJ (2002) Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma
lucidum, grown on lactose in a bioreactor. Biotechnology Letters 24: 1023-1026.
Tsukagoshi S, Hashimoto Y, Fujii G, Kobayashi H, Nomoto K, Orita K (1984) Krestin (PSK). Cancer Treatment Review 11: 131-
155.
Viccini G, Mitchell D, Boit S, Gern J, da Rosa A, Costa R (2001) Analysis of growth kinetic profiles in solid-state fermentation.
Food Technology and Biotechnology Advances 39: 271-294.
Wade LG (1993) Química Orgánica. Debra Wechsler. México. 1312 p.
Walkley A, Black I (1934) An estimation of the Degtjareft method for determining of soil organic matter and a proposed
modification of chromic acid titration method. Soil Science 37: 29-38.
Weinberg ZG (2000) Biotechnology in developing countries: opportunities in solid state fermentation applied in the agricultural
industry. Internation Journal of Biotechnology 2(4): 364-373.
Wu J, Cheung P, Wong K, Huang N (2004) Studies on submerged fermentation of (Fr.) Singer. Part 2. Effect of carbonto-nitrogen
ration of the culture medium on the content and composition of the mycelial dietary fibre. Food Chemistry 85: 101-105.
Wu J, Ding Z-Y, Zhang K-C (2006) Improvement of exopolysaccharide production by macro-fungus Auricularia auricula in
submerged culture. Enzyme and Microbial Technology 39: 743-749.
Xiao J-H, Xiao D-M, Xiong Q, Liang Z-Q, Zhong JJ (2010) Nutritional requirements for the hyperproduction of bioactive
exopolysaccharides by submerged fermentation of the edible medicinal fungus Cordyceps taii. Biochemical
Engineering Journal 49(2): 241-249.
Xiao JH, Chen DX, Wan WH, Hu XJ, Qi Y, Liang ZQ (2006) Enhanced simultaneous the production of mycelia and intracellular
polysaccharide in submerged cultivation of Cordyceps jiangxiensis using desirability functions. Process Biochemestry
41: 1887-1893.
Yang F-C, Ke Y-F, Kuo S-S (2000) Effect of fatty acids on the mycelial growth and polysaccharide formation by Ganoderma
lucidum in shake flask cultures. Enzyme and Microbial Technology 27: 295-301.
Zhuang O, Mizuno T (1999) Biological responses from Grifola frondosa (Dick.:Fr.) S.F. Gray-Maitake
(Aphyllophoromycetideae). Internation Journal Medicinal Mushrooms 1: 317-324.

138
7. Selección de la Mejor Combinación Hongo –
Formulación para la Degradación de Sustratos
Lignocelulósicos mediante Fermentación Sumergida
con Pleurotus ostreatus, Coriolus versicolor y Lentinus
edodes

139
Selección de la Mejor Combinación Hongo – Formulación para la
Degradación de Sustratos Lignocelulósicos mediante Fermentación Sumergida
con Pleurotus ostreatus, Coriolus versicolor y Lentinus edodes
Sandra Montoya1, Óscar Julián Sánchez
2*, Laura Levin
3
RESUMEN
En esta investigación se probaron tres especies de hongos de pudrición blanca: Coriolus versicolor,
Pleurotus ostreatus y Lentinus edodes sobre 12 formulaciones de medios de cultivo líquidos con cascarilla
de café pretratada como fuente de materiales lignocelulósicos y dos niveles de sulfato de cobre (II). Los
objetivos de este trabajo fueron i) evaluar los efectos de la relación C/N y del sulfato de cobre (II) sobre
las actividades enzimáticas endoglucanasa, exoglucanasa, endoxilanasa, β-glucosidasa, lacasa y
manganeso peroxidasa (MnP) durante 32 días de fermentación sumergida; así como el consumo de
celulosa y hemicelulosa y la degradación de lignina y ii) proponer un modelo matemático para la
descripción de la producción de las enzimas lignocelulolíticas, la producción de biomasa, producción y
consumo de azúcares reductores, consumo de celulosa y hemicelulosa y degradación de lignina. Las tres
especies de hongos crecieron bien sobre todas las formulaciones de los medios de cultivo y se obtuvieron
títulos de actividades enzimáticas de todas las enzimas. Del mismo modo, no hubo una gran variación
entre las combinaciones de las tres especies de hongos con las diversas formulaciones sobre la
degradación de los componentes de la fibra de los medios de cultivo, probablemente porque el material
lignocelulósico adicionado fue pretratado y esto debió generar un desorden en la matriz lignocelulósica de
modo que las hifas de los hongos pudieron acceder más fácilmente a la celulosa. P. ostreatus mostró una
mejor capacidad de degradación de los componentes de la fibra en las diferentes combinaciones, por lo
que este hongo fue seleccionado con las diferentes combinaciones para la aplicación del modelo
matemático compuesto por 11 ecuaciones diferenciales propuesto para describir los datos experimentales.
Palabras clave: hongos de pudrición blanca, relación C/N, actividades lignocelulolíticas, degradación de
materiales lignocelulósicos.
1. INTRODUCCIÓN
Los hongos de pudrición blanca son hasta ahora los únicos organismos capaces de degradar
completamente todos los componentes de los materiales lignocelulósicos gracias a las enzimas
lignocelulolíticas que producen (Kirk y Farrel, 1987b). En general las enzimas degradadoras de
polisacáridos están sujetas a mecanismos de regulación de su síntesis. Es decir no se producen de modo
constante, sino que su síntesis es inducida por el sustrato adecuado y es reprimida por azúcares fácilmente
utilizables, en particular glucosa. El inductor más eficiente es el polímero-sustrato de las enzimas que
serán sintetizadas. Sin embargo, debido a su alto peso molecular, no son compuestos que puedan penetrar
en las células y ejercer su efecto (Béguin y Aubert, 1994). Algunas de las enzimas extracelulares que se
producen por los hongos son solubles y se encuentran libremente dispersas en la película de líquido que
rodea la hifa, pero otras están fijas en el espacio sujetas a la pared de la hifa, a la matriz extracelular o al
propio sustrato (Hudson, 1986a).
Hay un mercado creciente en la producción de enzimas lignocelulolíticas y otras biosustancias por
fermentación sumergida empleando hongos macromicetos. En la actualidad, el cultivo sumergido de
hongos macromicetos es una alternativa promisoria a nivel mundial para la producción eficiente de
micelio y metabolitos. Sin embargo, a través de décadas la producción de metabolitos por fermentación
sumergida utilizando macromicetos continúa enfrentando limitaciones de carácter biológico, fisiológico y
de ingeniería. De igual forma el desarrollo de las hifas (especialmente de basidiomicetos) en cultivos

140
sumergidos resulta usualmente en el desarrollo incontrolado del micelio (conjunto de hifas). La extensión
de la biomasa fúngica tiene profundos efectos en la transferencia de masa, velocidad metabólica y
secreción de productos. El micelio fúngico puede enrollarse alrededor de las esferas (pellets) de la misma
biomasa, causando bloqueos y extendiéndose en todo el medio de cultivo, incrementando la viscosidad, la
cual resulta ser una limitante de la transferencia de masa y de oxígeno en el reactor. Todos estos
inconvenientes limitan el tiempo de operación para la producción de biosustancias de basidiomicetos por
fermentación sumergida. Asimismo, la falta de información sobre el cultivo sumergido de setas es
significativa en comparación con la gran cantidad de información acerca de la fermentación sumergida
empleando bacterias (estreptomicetos) y microhongos (Rodriguez-Couto y Toca-Herrera, 2007; Tang et
al., 2007).
Para producir enzimas lignocelulolíticas con hongos de pudrición blanca como Pleurotus
ostreatus, Coriolus versicolor y Lentinus edodes han sido empleados métodos de fermentación en estado
sólido, debido a que el medio sólido es la forma natural de crecimiento de los hongos. Sin embargo,
investigadores como Hölker et al. (2004) han alcanzado métodos sencillos y económicos de obtención de
enzimas lignocelulolíticas por fermentación sumergida con hongos de pudrición blanca. Empero
dificultades como la determinación de la biomasa y la descripción de la cinética de estos procesos, han
reducido la cantidad de información publicada que sirva como herramienta indispensable en la descripción
fisiológica y biotecnológica del proceso productivo de enzimas lignocelulolítcas por fermentación
sumergida (Kovarova-Kovar y Egli, 1998).
El modelado de procesos de fermentación sumergida para la producción de enzimas
lignocelulolíticas con hongos de pudrición blanca tiene un gran potencial en la búsqueda de las
condiciones óptimas del proceso, diseño y desarrollo de procesos, control, ampliación e identificación de
la estructura de costos (Vasic-Racki et al., 2003; Vrsalovic Presecki et al., 2006). Para ese propósito, se
han utilizado modelos matemáticos no-estructurados con el fin de describir el crecimiento celular y la
producción de las diferentes biosustancias en los diferentes procesos de producción discontinuos,
continuos o alimentados (Zelic et al., 2004). Los modelos más reportados para procesos de fermentación
por lotes son el modelo lineal, logístico y de dos fases (aceleración y desaceleración) (Sangsurasak et al.,
1996), aunque estos modelos no incluyen el efecto de la concentración de nutrientes sobre el desarrollo
(Ikasari y Mitchell, 2000; Mitchell et al., 2004; Van de Lagemaat y Pyle, 2005). Asimismo, para la
descripción de la producción de biomasa en diversos tipos de procesos de fermentación se utilizan las
expresiones tipo Monod y Logística modificada (Mitchell et al., 2004; Tavares et al., 2005). Las
expresiones matemáticas utilizadas para la descripción de la formación de productos dependen de qué
clase de metabolito se esté produciendo; generalmente, las expresiones más simples son para los
metabolitos primarios, ya que su formación se considera directamente proporcional a la formación de la
biomasa celular teniéndose en cuenta los parámetros de matenimiento celular y crecimiento microbiano.
Actualmente existen dificultades para describir la producción de biosustancias de hongos macromicetos y
su crecimiento con el uso de los modelos matemáticos disponibles (Tišma et al., 2010). Esta dificultad se
debe, probablemente, a que cada uno de estos hongos se ve afectado directamente por factores intrínsecos
y extrínsecos particulares como las características físicas y la composición química de los medios de
cultivo, la procedencia de las cepas, la influencia de las condiciones ambientales y una velocidad de
crecimiento menor asociada a la gran mayoría de las especies de macromicetos en comparación con el
tiempo de crecimiento de los micromicetos. Asimismo, para el análisis de la formación de productos en
procesos de fermentación sumergida es necesario conocer todas las interrelaciones que suceden entorno a
la formación de las diversas sustancias intermedias que se pueden formar durante el crecimiento y
desarrollo del organismo, que a su vez cambian las características fisico-químicas del medio de cultivo a
fin de comprender el mecanismo de producción de la biomasa y de las sustancias de interés y lograr
plantear expresiones matemáticas que permitan la descripción y escalamiento de estos procesos
productivos.
En el caso de los hongos de pudrición blanca cuando se cultivan en medios de composición
ligeramente diferente, se sintetizan diversas formas de una misma enzima (isoenzimas) que a pesar de
poseer propiedades catalíticas similares, difieren notablemente en sus características físico-químicas, por

141
lo que se deduce que hay formas múltiples que coexisten en determinado momento; también puede variar
la proporción de estas isoenzimas a lo largo del desarrollo del hongo (Carlile et al., 2001). La síntesis de
isoenzimas puede deberse a productos de genes diferentes, aunque también pueden deberse a cambios
postraduccionales como proteólisis, glicosilación o agregados cuaternarios. Se pueden sintetizar como
consecuencia de estados fisiológicos diferentes (composición del medio de cultivo, pH, temperatura, etc.)
a partir de genes regulados diferencialmente (Joselau y Ruel, 1994; Blanchette, 1995; Barr et al., 1996;
Carlile et al., 2001).
El objetivo de este trabajo fue evaluar el efecto de la relación carbono nitrógeno y del sulfato de
cobre (II) sobre la producción de enzimas lignocelulolíticas por tres especies de hongos de pudrición
blanca (L. edodes, C. versicolor y P. ostreatus) empleando doce formulaciones diferentes de medios de
cultivo por fermentación sumergida. Estos hongos fueron seleccionados de entre diez hongos de pudrición
blanca mediante un rastreo basado en sus actividades enzimáticas celulolíticas, xilanolíticas y
ligninolíticas, así como en su capacidad de degradación de celulosa y lignina; los resultados de este rastreo
se relacionan en el artículo anterior. Igualmente este trabajo tuvo como objetivo adicional proponer y
validar un modelo matemático para la mejor combinación hallada especie de hongo - formulación a fin de
realizar la descripción de la cinética de crecimiento del organismo, la degradación de los materiales
lignocelulósicos y la producción de las enzimas lignocelulolíticas en condiciones de fermentación
sumergida.
2. MATERIALES Y MÉTODOS
2.1. Microorganismos
Tres especies de basidiomicetos fueron empleados para la producción de enzimas
lignocelulolíticas en fermentación sumergida, tomadas de la colección de hongos de la Universidad de
Caldas (Manizales, Colombia): Lentinula edodes del Centro de Investigaciones del Café, Chinchiná,
Caldas, Colombia (CICL54), Coriolus versicolor de Pennsylvania State University, Estados Unidos de
Norte América (PSUWC430), y Pleurotus ostreatus de la Colección interna de la Universidad de Caldas
(UCC001). Las cepas fueron extendidas sobre agar papa dextrosa (PDA) y mantenidas en refrigeración a
4°C y transferencias periódicas.
2.2. Pretratamiento de cascarilla de café
La cascarilla de café seca y molida fue sumergida en H2SO4 al 3% en relación 1:4 sólido: líquido y
sometida a ultrasonido por una hora a 80°C con posterior tratamiento térmico a 121°C y 15 psig por una
hora con fines de modificar la matriz lignocelulósica. En seguida, la cascarilla fue lavada con exceso de
agua hasta obtener un pH = 6.8 en el agua de lavado del material para retirar el exceso de ácido. La
cascarilla lavada fue utilizada como fuente de carbono proveniente de material lignocelulósico en los
medios de cultivo para la fermentación sumergida del presente trabajo.
2.3. Medios de cultivo
Se realizaron doce formulaciones de medios de cultivo por triplicado con base en un litro de
medio de cultivo. La composición por cada litro de medio de cultivo fue: 100g de cascarilla de café
pretratada y seca, 2g KH2PO4, 0,5g MgSO4.7H2O, 0,1g CaCl2, 2g de extracto de levadura y tres
concentraciones de SO4(NH4)2 0,5g, 1g y 1,5g, dos concentraciones de aceite de soya 13,5g y 22,42g y
dos concentraciones de sulfato de cobre (II) 0,1592g (fórmulas pares) y 0,0798g (fórmulas impares), con
la variación de los niveles de SO4(NH4)2 y aceite de soya se modificó la relación carbono nitrógeno (R
C/N) (ver Tabla 1). Los medios de cultivo fueron servidos en matraces de 250 mL de capacidad con 100
mL de medio, esterilizados a 121°C por 20 minutos, los pH de los medios de cultivo variaron entre 5.6 y
5.8 para todas las formulaciones medido antes de la inoculación; seguidamente, los medios de cultivo

142
fueron inoculados en cabina de flujo laminar con un fragmento de micelio extendido en agar PDA de ½
cm de lado de cada especie de trabajo. Los matraces fueron llevados a incubación con agitación de 100
rpm, 25°C y penumbra por 32 días. Se tomaron 10 muestras, dos por semana y la primera muestra fue
tomada el día de la inoculación.
Tabla 1. Relaciones carbono nitrógeno (R C/N) en doce niveles (formulaciones)
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 F11 F12
24,68 24,13 28,40 26,84 23,98 22,06 23,41 23,79 20,81 20,61 20,70 21,04
2.4. Determinación de la relación carbono nitrógeno
La relación carbono nitrógeno (C/N) fue determinada a partir de la medida de la materia orgánica
de las doce formulaciones de sustratos empleando el método de Walkley y Black (1934), el cual utiliza
ácido sulfúrico concentrado y solución de dicromato de potasio. La muestra se lee a 585 nm. La cantidad
de carbono total corresponde al 58% de la materia orgánica. El nitrógeno orgánico total fue determinado
por el método de Kjeldahl (Kjeldahl, 1883).
2.5. Determinación cuantitativa de actividades enzimáticas
2.5.2. Actividades celulolíticas y xilanolíticas
Endo-1,4-ß-D-glucanasa (E.C.3.2.1.4). Se usó carboximetil celulosa (CMC) como sustrato al
0,5% en buffer acetato de sodio pH 4,8 en reacción con 100 µL de extracto enzimático (medio filtrado)
por 30 minutos a 50°C. La reacción se detuvo adicionando el reactivo DNS (ácido dinitrosalicílico)
(Miller, 1959). Se continuó con la reacción de DNS para la determinación de azúcares reductores, se leyó
la absorbancia a longitud de onda 540 nm. Una unidad de actividad enzimática (UE) fue definida como la
cantidad de enzima que produce 1 µmol de azúcares reductores en un minuto. Para cuantificar la actividad
se elaboró una curva patrón de azúcares reductores, con distintas cantidades de glucosa.
Exo-1,4-ß-D-glucanasa (E.C.3.2.1.91). Se usó celulosa cristalina 1% en buffer acetato de sodio pH
4,8 como sustrato, en reacción con 100 µL de extracto enzimático a 50°C por 60 minutos y agitación. La
reacción se detuvo adicionando el reactivo DNS. Se continuó la reacción de azúcares reductores por el
método DNS y se centrifugó antes de leer la absorbancia a longitud de onda 540 nm. Una unidad de
actividad enzimática (UE) fue definida como la cantidad de enzima que produce 1 µmol de azúcares
reductores en un minuto.
β-glucosidasa (E.C.3.2.1.21). La reacción se produce entre el sustrato de p-nitrofenil β-D-
glucopiranosido 0,02% en buffer acetato de sodio pH 4,8 con 100 µL de extracto enzimático a 50°C por
30 minutos. La reacción se detuvo adicionando solución buffer Clark y Lubs (pH 9,8) y se leyó la
absorbancia a longitud de onda 430 nm (ε430=18,5/mM cm) (Wood y Bhat, 1988). Una unidad de actividad
enzimática (UE) fue definida como la cantidad de enzima que produce 1 µmol de producto (p-nitrofenol)
en un minuto. Para cuantificar la actividad se confeccionó una curva patrón con p-nitrofenol.
Endo-1,4-ß-D-xilanasa (E.C.3.2.1.8). La reacción se produce entre el sustrato xilano al 0,2% en
buffer acetato de sodio pH 4,8 con 100 µL de extracto enzimático a 50°C por 30 minutos. La reacción se
detuvo adicionando DNS y se leyó la absorbancia a longitud de onda 540 nm. Una unidad de actividad
enzimática (UE) fue definida como la producción de 1 µmol de azúcares reductores en un minuto. La
curva patrón de azúcares reductores se confeccionó en este caso con xilosa.

143
2.5.3. Actividades ligninolíticas
Lacasa (E.C.1.10.3.2): Se utilizaron 0,5 mM de ABTS en solución 0,1M de buffer acetato de
sodio pH 3,6 como sustrato, según método de Paszczynski y Crawford (1991), leyendo el aumento de
absorbancia a 420 nm (ε420=36/mM cm) luego de tres minutos de reacción a 30°C. Una unidad de
actividad lacasa (UE) fue definida como la cantidad de enzima requerida para oxidar 1 μmol de ABTS en
1 minuto.
Manganeso peroxidasa (E.C.1.11.1.13) (MnP): Se utilizó como sustrato una solución 0,01% de
rojo fenol en buffer succinato de sodio 0,1 M y pH 4,5 y sulfato de manganeso (0,22 g/L), con peróxido de
hidrógeno 0,2 mM como iniciador de la reacción. La reacción fue detenida adicionando NaOH 5N
después de 10 minutos de reacción para leer el incremento de absorbancia a 610 nm (ε610=22/mM cm).
Una unidad de actividad enzimática (UE) fue definida como la cantidad de enzima necesaria para oxidar 1
μmol de rojo fenol en 1 minuto (Paszczczynski et al., 1988).
2.6. Cuantificación del contenido de N-acetil-D-glucosamina (NAGA) y biomasa
Las muestras de sustrato fueron colectadas en 10 tiempos de incubación hasta el final de la
fermentación (día 32) para las tres especies de trabajo. El medio de fermentación fue filtrado al vacío, con
posterior secado en estufa a 101°C hasta peso constante. A la muestra seca y molida se le determinó el
contenido de quitina. El contenido de biomasa fúngica en este medio fue estimado por la determinación
del contenido de N-acetil-D-glucosamina (NAGA) después de la hidrólisis con HCl 6N grado analítico
según el método de Plassard et al. (1982). Para ello se cuantificó paralelamente el contenido de NAGA
por gramo de micelio en cada una de las tres especies, cultivándolas en medio líquido. El contenido de
NAGA del micelio de cada una de las especies de hongos de trabajo: C. versicolor, P. ostreatus y L.
edodes fue determinado del desarrollo del micelio en medio líquido en matraces de 250 mL con 100 mL
de medio de cultivo con la siguiente composición: glucosa (30 g/L), extracto de levadura (6 g/L),
SO4Mg.5H2O (0,5 g/L), K2HPO4 (0,5 g/L) y CaCl2 (0,1 g/L). El tiempo de fermentación para el
crecimiento del hongo fue de 25 días y sometidos a 100 rpm de agitación. Los compuestos solubles en
agua fueron extraídos durante la hidrólisis con HCl 6N por tres horas.
2.7. Cuantificación de los componentes de la fibra y azúcares reductores
Se determinaron los componentes de la fibra (celulosa, hemicelulosa y lignina) de los medios de
cultivo de cada una de las formulaciones en 10 tiempos durante los 32 días de incubación, así como la
fracción soluble, empleando los resultados de la determinación de fibras detergente neutra, ácida y
lignino-ácida. La medición se realizó a una muestra seca y molida de sustrato; dicha muestra se sometió a
tres hidrólisis en serie (cada una de 75 min): i) hidrólisis con lauril sulfato de sodio y otros, ii) hidrólisis en
solución de bromuro de amonio en ácido sulfúrico 1N, y iii) hidrólisis con ácido sulfúrico al 72% (p/v). Al
finalizar cada hidrólisis, las muestras fueron lavadas y secadas a 105°C hasta peso constante (Leterme,
2010a). El contenido de azúcares reductores como glucosa fue determinado por el método del ácido
dinitrosalicílico (DNS) (Miller, 1959).
2.7. Diseño experimental
Se planteó un diseño experimental bifactorial con la especie a tres niveles y doce formulaciones de
sustrato. Las formulaciones varían en su relación C/N y la concentración de sulfato de cobre (II). Las
variables independientes en este diseño experimental fueron la especie de hongo, la formulación del
sustrato y el tiempo de fermentación. Las variables de respuesta medidas fueron las actividades
enzimáticas: endoglucanasa, exoglucanasa, β-glucosidasa, lacasa, MnP y endoxilanasa a fin de buscar la
mejor combinación especie de hongo fórmula para la producción de celulasas y de ligninasas en términos

144
de la capacidad de degradación de los componentes de la fibra, celulosa, hemicelulosa y lignina durante
32 días de fermentación sumergida en 10 tiempos.
2.6. Modelamiento matemático del crecimiento del hongo y la producción de enzimas
Para la solución de los modelos matemáticos del proceso de fermentación en estado líquido se
utilizó el software Matlab® 2010b (MathWorks, EUA) en un PC de 3 GB de memoria RAM y un
procesador Intel CORE-i3 de 2,13 GHz. Para los modelos matemáticos planteados en ecuaciones
diferenciales ordinarias se utilizó el comando ode45 que está basado en una fórmula explícita de Runge-
Kutta (4,5) usando un par de Dormand-Prince (Dormand y Prince, 1980) y el comando ode15s basado en
una fórmula de orden variable que funciona mediante fórmulas de diferenciación numérica.
2.8. Análisis estadístico
Para todos los análisis estadísticos del presente trabajo se utilizó el software Matlab ® 2010b. Este
análisis se realizó en tres etapas. Primero se realizó un ANOVA a todos los datos del diseño experimental
y sus posibles combinaciones: especie, fórmula y tiempo de fermentación mediante el comando anovan.
Luego se realizó un análisis comparativo (Kruskal Wallis) de la capacidad de degradación de celulosa,
hemicelulosa y lignina para cada especie de hongo estudiado y las doce formulaciones de sustratos
mediante los comandos kruskalwallis y multcompare. A los máximos obtenidos de cada especie se les
realizó un nuevo análisis comparativo a fin de encontrar la combinación de mejor desempeño con los
porcentajes de degradación de la fibra al final del período de fermentación (32 días) mediante los
comandos kruskalwallis y multcompare.
Para evaluar si el modelo matemático propuesto describe adecuadamente los datos
experimentales, se realizó una prueba F de una cola que parte de las siguientes hipótesis:
2
,
0 2
exp,
2
,
1 2
exp,
: 1
: 1
i
i
i
i
residuales F
datos F
residuales F
datos F
SH
S
SH
S
donde: Fi está dado para i = 1,2,…,12, s2residuales es la varianza de los residuales (desviaciones de los datos
experimentales con respecto a los valores calculados por el modelo matemático) y se calcula para cada
variable de respuesta medida y s2datos exp es la varianza de la serie experimental de cada variable de
respuesta medida. Para estos análisis se utilizaron los comandos vartest y vartest2 de Matlab. Con las
varianzas de los residuales obtenidos para cada serie experimental se calcularon los porcentajes de cada
una respecto a la serie que dio la mayor varianza de residuales a fin de mostrar el ajuste de los datos
experimentales al modelo matemático propuesto de forma más simple.
3. RESULTADOS Y DISCUSIÓN
3.1. Efecto de la relación C/N sobre la degradación de los residuos lignocelulósicos y la producción
de enzimas lignocelulolíticas
Las tres especies de hongos (C. versicolor, P. ostreatus y L. edodes) probadas en el diseño
experimental, fueron inoculadas e incubadas bajo las mismas condiciones ambientales en las doce
formulaciones de los medios de cultivo. El intervalo de la relación C/N en las doce formulaciones de
sustrato evaluadas fue de 20 a 28,4 con una variación en los contenidos de nitrógeno de 1.4 a 1,93% en
base seca. Las tres especies de hongos se desarrollaron bien en todas las formulaciones evaluadas y se

145
obtuvieron títulos de todas las actividades enzimáticas en todas las formulaciones durante los 32 días de
incubación, como se observa en la Tabla 2. Todos los contenidos de nitrógeno de las formulaciones
estimularon el crecimiento micelial de los macromicetos durante la fermentación, incrementando el
contenido de biomasa (ver Figuras 1 y 2) en todos los casos y con una marcada fase lag para las tres
especies en todos los medios de cultivo. Empero se debe tener en cuenta que la variación de la relación
C/N se provocó modificando los contenidos de (NH4)2SO4 y aceite de soya y no modificando las fuentes
de nitrógeno y de carbono en cada formulación. Sin embargo, sí hubo variabilidad sobre las actividades
enzimáticas medidas (endoglucanasa, exoglucanasa, β-glucosidasa, endoxilanasa, lacasa y MnP),
dependiendo de la especie y a diferentes tiempos de incubación como se observa en la Tabla 2. Las
actividades enzimáticas máximas fueron seleccionadas realizando análisis estadístico por etapas. Primero
se realizó un ANOVA a fin de determinar si hubo diferencias significativas entre los resultados obtenidos
respecto a las variables independientes especie, fórmula y tiempo; posteriormente, se realizó una selección
de los máximos de las actividades enzimáticas gráficamente. La mayor actividad de β-glucosidasa fue
detectada para L. edodes – F10 con 32.640 U/L.min a los 32 días de incubación, seguida por la
combinación C. versicolor – F5 con 30.560 U/L.min. Mientras que la máxima actividad de β-glucosidasa
para las combinaciones con P. ostreatus fue para la F3 con 15.721,6 U/L.min. Para la enzima
endoglucanasa la mayor actividad fue detectada para C. versicolor – F9 con 11.727,7 U/L.min en el día 32
de la fermentación, seguida de 11.403 U/L.min para P. ostreatus – F5 en el día 25 de la fermentación. Los
niveles más altos de la enzima exoglucanasa se obtuvieron para L. edodes – F6 y F5 con un títulos de
10.980 U/L.min (días 21) y 10.240 U/L.min (día 32) respectivamente. Para la enzima endoxilanasa, la
mayor actividad obtenida se reporta para L. edodes – F10 con 21880 U/L.min en el día 21 de
fermentación. Las máximas actividades enzimáticas de lacasa fueron obtenidas para C. versicolor – F11
día 25 de incubación y 260 U/L.min para P. ostreatus – F1 y F3 en los días 18 y 28 de la fermentación
respectivamente. En referencia a los títulos obtenidos de la MnP, los máximos detectados fueron para P.
ostretaus – F5 y F6 con 135,89 U/L.min y 106,2 U/L.min respectivamente. Detallando las actividades
enzimáticas máximas obtenidas para todas las formulaciones y visualizando especialmente los títulos
obtenidos para las formulaciones F3 cuya relación C/N fue la máxima y las formulaciones F10 y F11 que
tenían las menores relaciones C/N no se aprecia una diferencia notable entre ninguna de las actividades
enzimáticas medias como para ser atribuido a este factor, probablemente porque los componentes del
sustrato que provocan la variación de la relación C/N no se modifican durante la formulación de los
medios de cultivo y los niveles evaluados no son suficientemente diferentes para provocar un cambio
significativo en estas actividades enzimáticas.
En este estudio se presentó la producción de cuatro actividades enzimáticas celulolíticas y
xilanolíticas y dos actividades ligninolíticas de tres especies de hongos de pudrición blanca, en
fermentación sumergida con una fuente fija de material lignocelulósico, la cascarilla de café pretratada,
variando las cantidades de sulfato de amonio y aceite de soya adicionados por litro de medio de cultivo
para conseguir las diferentes relaciones C/N. La cascarilla de café pretratada se adicionó con el fin de
activar la producción de enzimas lignocelulolíticas por las especies de hongos utilizadas. Entendiendo que
para la bioconversión de los materiales lignocelulósicos con hongos, las enzimas que degradan
polisacáridos juegan un rol muy importante ya que son las que proveen la fuente de energía a los
microorganismos; por lo que la baja producción de estas enzimas pueden limitar la velocidad de
producción de azúcares solubles. Sin embargo, en este trabajo los organismos probablemente pudieron
aprovechar como fuentes de carbono, los contenidos en el aceite de soya y extracto de levadura para su
mantenimiento durante las fase lag prolongadas que se visualizan para todas las combinaciones hongo
formulaciones en las Figuras 1 y 2, mientras los mismos hongos inician la degradación del material
lignocelulósico a fin de tener la cantidad de nutrientes necesarios para su crecimiento y desarrollo.
Los títulos de las actividades de las enzimas lignocelulolíticas endoglucanasa, endoxilanasa,
exoglucanasa, lacasa y MnP obtenidas en este trabajo fueron similares a las obtenidas por Elisashvili et al.
(2008) cuando utilizaron hojas de árboles de eucalipto en los medios de cultivo para fermentación
sumergida con varias especies de L. edodes y P. ostreatus, ya que cuando ellos utilizaron como fuentes de
carbono en los medios de cultivo cáscaras de mandarinas, de manzana y de banano, los títulos de las

146
actividades enzimáticas fueron inferiores a las obtenidas en este trabajo, especialmente los títulos de la
MnP y lacasa. Asimismo, trabajos recientes han demostrado que diversos materiales lignocelulósicos
pueden estimular la producción de enzimas lignocelulolíticas por los basidiomicetos (Kapich et al., 2004;
Elisashvili et al., 2006). Acorde con lo anterior, los resultados obtenidos con las tres especies de hongos
de pudrición blanca L. edodes, C. versicolor y P. ostreatus utilizadas en este estudio para producir
enzimas lignocelulolíticas son concordantes con los obtenidos por otros autores respecto a las variaciones
cuantitativas de las actividades enzimáticas como las celulasas. Por ejemplo, en este estudio se obtuvieron
actividades de endoglucanasa máximas de 11.403 U/L.min, 11.727 U/L.min y 9.226 U/L.min en P.
ostreatus, C. versicolor y L. edodes respectivamente mayores a las actividades de endoglucanasa
obtenidas por Tsiklauri et al. (1999) utilizando aserrín de uva y cáscaras de mandarina con Funalia trogii
y Polyporus ciliatus. Esto significa que varios de estos hongos basidiomicetos son capaces de degradar
diversos materiales lignocelulósicos bajo diferentes relaciones C/N y en condiciones medioambientales
diversas.
3.2. Efecto del sulfato de cobre sobre la degradación de residuos lignocelulósicos y la producción
de enzimas lignocelulolíticas
Los dos niveles de CuSO4 utilizados en las doce formulaciones del presente estudio no generaron
inhibición aparente en el crecimiento de los tres organismos de trabajo. Como se observa en las Figuras 1
y 2 la biomasa de los tres hongos aumentó en todas las formulaciones durante los 32 días de fermentación.
Las enzimas lignocelulolíticas fueron medidas dos veces por semana tomando las muestras del medio
líquido en cabina de flujo laminar a las cuales se le determinó las actividades enzimáticas de cuatro
hidrolasas y dos óxido-reductasas. Se observa una gran variación en los máximos de las actividades
enzimáticas medidas entre especies y formulaciones como se observa en la Tabla 2.
Son muchos los factores que pueden afectar la producción de enzimas lignocelulolíticas de hongos
de pudrición blanca. Además, las diferentes cepas responden de un modo particular a cada uno de los
factores intrínsecos y extrínsecos que deben enfrentar cuando son inoculados. Debido a esto, el estudio
combinado de dos o más factores puede generar mayor dificultad al momento de la interpretación de las
posibles interacciones entre los compuestos que afectan directamente la producción de enzimas
lignocelulolíticas. La variabilidad de las actividades enzimáticas de las hidrolasas es tan amplia entre las
especies de trabajo y las doce formulaciones probadas que no hay una evidencia clara de que las
cantidades de CuSO4 (0,1596 g/L y 0,0796 g/L) adicionadas a los sustratos hayan generado alguna
inhibición o activación directa a estas actividades. Adicionalmente, no hay reportes de que la presencia de
cobre en el medio de cultivo líquido en las cantidades adicionadas genere algún efecto importante sobre
las actividades enzimáticas de las hidrolasas, de hecho en las Figuras 3, 4, 5 y 6 se muestran las
actividades enzimáticas hidrolíticas medidas para las tres cepas sobre las formulaciones F1 a F6 en las que
se observó una gran variación entre los diferentes tiempos.
Tabla 2. Máximas actividades enzimáticas lignocelulolíticas (U/L.min) de tres especies de hongos de pudrición
blanca para doce niveles de relación carbono nitrógeno (R C/N) y dos niveles de CuSO4 Fórmula F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 F11 F12
CuSO4 (g/L) 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798 0,1596 0,0798
R C/N 24,68 24,13 28,40 26,84 23,98 22,06 23,41 23,79 20,81 20,61 20,70 21,04
βG (U/g ss)
P. ostreatus 11950
(28) 9590 (7)
15721,6
(28)
12210
828)
11250
(28)
15930
(21)
7600
(28)
6720
(28)
14590
(25)
14040
(28)
7520
(28)
15620
(32)
C. versicolor 19760 (11)
22240 (11)
23770 (32)
28610 (32)
30560 (32)
24950 (28)
23960 (25)
12170 (14)
9450 (25)
27520 (28)
18670 (21)
11270 (32)
L. edodes 26710
(21)
24460
(18)
22690
(32)
24410
(28)
25600
(28)
26320
(32)
29990
(25)
29760
(28)
25840
(32)
30640
(32)
27280
(28)
26420
(32)
ENG (U/g ss)
P. ostreatus 4850 (14) 5860 (11)
5196,5 (21)
6089 (14)
8022 (21)
11403 (25)
10213,2 (25)
8480 (32)
8428,3 (32)
6575,2 (32)
2451,5 (32)
9631,7 (32)
C. versicolor 4310 (14) 3736,1
(14)
5237,1
(18)
7833,3
(25)
5845,6
(28)
5007
(28)
8184,9
(32)
8150
(32)
11727,7
(32)
10389
(28)
8360,7
(32)
9023,2
(32)
L. edodes 7250 (18) 5967,3 (21)
9307,2 (21)
10267,3 (21)
8658,2 (21)
8630 (28)
7995,6 (32)
8830 (32)
7752,2 (32)
6372,9 (28)
9226,1 (32)
8428,3 (32)

147
EXG (U/g ss)
P. ostreatus 6024 (28) 7630
(11) 6660 (7)
6690
(14) 5546 (7)
6270
(11)
7150
(28)
6430
(25)
6440
(21)
6360
(32)
5200
(21)
4320
(32)
C. versicolor 4560 (32) 5190
(32) 5720 (7)
6140
(32) 7680 (7)
5260
(32)
6230
(28)
6320
(28)
6640
(32)
7230
(32)
5470
(28)
5170
(25)
L. edodes 5930 (11) 9190
(18)
8000
(21)
9190
(21)
10240
(32)
10980
(21)
6340
(25)
7540
(28)
6270
(32)
7980
(32)
5940
(32)
4630
(32)
ENX (U/g ss)
P. ostreatus 7990 (14) 7560
(18)
8290
(28)
7670
(18)
12310
(21)
16460
(21)
6800
(11)
7300
(25)
10260
(21)
12406
(21)
11633,1
(32)
4001
(28)
C. versicolor 12980
(32)
12780
(32)
13040
(21)
19580
(25)
18320
(7)
12100
(11)
9250
(28) 8370 (7)
12670
(14)
9920
(18)
7804
(28)
7320
(32)
L. edodes 10170
(32)
12960
(21)
13110
(28)
12780
(25)
13320
(32)
12220
(21)
13540
(32)
15920
(32)
11740
(32)
21880
(21)
14602
(32)
11940
(25)
Lac (U/g ss)
P. ostreatus 260 (18) 150 (28) 260 (28) 84 (25) 155,4
(32)
117,1
(21) 229(21)
117,8
(18)
189,4
(21)
130,2
(11)
181,7
(21)
176,33
(32)
C. versicolor 240 (28) 130 (32) 240 (28) 260 (28) 135,5
(21)
311,3
(28) 311 (28)
167,9
(11)
272,6
(32)
206,7
(32)
297,4
(25) 196 (25)
L. edodes 120 (32) 110 (28) 110 (21) 76 (11) 120,1
(18)
161,2
(14)
112,3
(21) 95 (18) 78 (18)
131,8
(25)
137,8
(18) 95 (21)
MnP (U/g ss)
P. ostreatus 59 (32) 70 (14) 66,8
(25)
66,19
(18)
135,89
(32)
106,2
(25)
83,12
(21)
78,38
(28)
102,4
(32)
70,61
(28)
75,11
(25)
74,7
(32)
C. versicolor 70,7 (25) 76 (32) 75,2
(18)
65,5
(25)
93,76
(28)
100,6
(32)
77,4
(21) 60 (14)
90,9
(21)
87,05
(25)
72,24
(18) 106 (18)
L. edodes 68 (28) 69 (32) 69,6
(28) 99 (25)
78,21
(25)
100,3
(28)
76,5
(25)
81,81
(18)
85,74
(18)
81,08
(32)
72,444
(28)
77,8
(32)
Nota: Los números entre paréntesis corresponden al día de incubación en el que se determinó la máxima actividad enzimática
(valor-p<0.05)
El efecto del CuSO4 como inductor de lacasa y peroxidasas ha sido estudiado por varios
investigadores (Levin et al., 2002; Niladevi y Prema, 2008). Las actividades enzimáticas de MnP
obtenidas en las diferentes combinaciones evaluadas en este trabajo fueron mucho menores que las
obtenidas por Bonugli-Santos et al. (2012) de los hongos marinos Tictoperellus sp., Marasmiellus sp. y
Peniphora sp. sobre diferentes medios de cultivo con inductores 1mM de CuSO4, 1mM de guaiacol y
salvado de trigo, cuyos títulos enzimáticos encontrados fueron de 1.600 a 2.600U/L cuando emplearon
CuSO4 como activador para las tres especies y cuando utilizaron salvado de trigo la actividad de MnP fue
de 2.556 y 8.206 U/L para Marasmiellus sp. y Peniphora sp., pero la actividad de MnP reportada para
Tictoperellus sp. sobre medio de cultivo con salvado de trigo fue de 627x103 U/L. De igual forma, los
títulos de MnP obtenidos por Levin et al. (2002) para Trametes trogii utilizando 1mM de CuSO4 como
activador también fueron mayores que los obtenidos en todas las combinaciones del presente estudio. Lo
anterior probablemente se debió a que en el presente estudio se utilizaron concentraciones de CuSO4
menores a las utilizadas por estos investigadores, acción que pudo no haber generado el efecto de
activador de la enzima MnP de los hongos de pudrición blanca evaluados.

148
Figura 1. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de medios de cultivo. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 durante 32 días de fermentación
sumergida
Figura 2. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de medios de cultivo. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 durante 32 días de fermentación
sumergida
La actividad de lacasa obtenida en las diferentes combinaciones de las tres especies de hongos
evaluadas mostró relación con el comportamiento de la biomasa, ya que durante los primeros 10 días de
fermentación, en la cual todas las combinaciones valoradas se mantuvieron en una aparente fase lag, la
actividad de la enzima lacasa también se apreció con bajos títulos. Pasada esa fase, cuando los hongos
inician la fase logarítmica de crecimiento, la actividad de lacasa también se incrementó notablemente.
Diferentes especies de P. ostreatus, C. versicolor y L. edodes han sido reportados como buenos
productores de enzimas ligninolíticas empleando inductores o simplemente cuando son cultivados sobre
medios que tienen algún material lignocelulósico (Buswell et al., 1995; Baldrian y Gabriel, 2002). Los
resultados de este estudio indican que seleccionando las formulaciones y las condiciones de los medios de
cultivo para el crecimiento y desarrollo de hongos de pudrición blanca es posible aumentar la secreción de
las actividades enzimáticas como la de lacasa, como se observa en la Tabla 2, P. ostreatus con F1, F3 y F7
y C. versicolor con F1, F3, F6 y F7 arrojaron títulos de 260 y 229 U/L.min y de 240 a 311 U/L.min (valor-
p < 0.05) respectivamente, resultando promisorios para la producción de esta enzima. Además, se ha
demostrado que la presencia de la cascarilla de café pretratada en medios de cultivo líquidos para
fermentaciones sumergidas contribuyen al incremento de actividades enzimáticas ligninolíticas tales como
la de lacasa, hecho que también ha sido demostrado por otros investigadores (Elisashvili et al., 2006).
Finalmente, las lacasas pertenecen al grupo de oxidasas de cobre azul y contienen cuatro átomos de cobre

149
por molécula, distribuidos en tres diferentes sitios de unión de cobre (Solomon et al., 1997); por lo tanto la
presencia de cobre en los medios de cultivo siempre es importante para la inducción y producción de las
mismas. Además, cabe resaltar que con los hongos de pudrición blanca se tiene una herramienta ambiental
importante, ya que incrementando la actividad de lacasas utilizando sulfato de cobre es posible utilizarlas
en procesos de biorremediación de aguas residuales y otros recalcitrantes, de modo que la concentración
total de cobre en el medio es consumido o disminuido por estos organismos durante el mismo proceso de
biorremediación (Gassara et al., 2010).
Tabla 3. Porcentaje de degradación de celulosa, hemicelulosa y lignina en las doce fórmulas y las tres especies de
hongos: C. versicolor, P. ostreatus y L. edodes durante 32 días de fermentación sumergida (valor-p = 0,0656)
Fórmula
Celulosa Hemicelulosa Lignina
C. versicolor P. ostreatus L. edodes C. versicolor P. ostreatus L. edodes C. versicolor P. ostreatus L. edodes
F1 58,12 57,78 58,12 76,24 69,83 72,69 46,59 47,29 49,64
F2 57,74 58,12 58,09 58,12 73,86 67,53 45,83 47,57 48,06
F3 58,12 57,78 58,12 67,81 61,13 74,31 48,36 44,97 44,81
F4 56,50 57,10 58,34 76,79 69,83 72,69 43,96 43,24 47,29
F5 57,78 59,12 58,50 58,82 73,86 67,83 50,57 44,99 44,21
F6 56,79 57,71 58,21 67,81 61,63 74,01 43,58 45,34 43,19
F7 57,55 49,50 57,78 76,79 69,12 73,32 46,68 46,74 45,77
F8 58,12 57,78 57,18 57,85 74,46 67,83 44,86 46,95 52,99
F9 58,12 57,73 58,83 67,81 61,03 74,31 45,56 47,40 50,74
F10 57,06 58,12 57,15 76,79 69,12 73,32 45,42 47,38 46,73
F11 58,84 58,34 57,89 57,05 74,46 67,83 46,98 46,08 43,86
F12 57,11 58,84 58,34 67,81 61,32 74,31 44,44 45,62 44,81
3.3. Producción de enzimas lignocelulolíticas y consumo de sustratos de la mejor combinación
hongo – formulación
Los datos experimentales obtenidos en el presente estudio fueron sometidos a un proceso de
selección a fin de lograr la mejor combinación especie de hongo fórmula del diseño experimental
bifactorial respecto a la producción de enzimas lignocelulolíticas, el cual se realizó por etapas. Primero se
hizo un análisis de varianza, ANOVA a tres vías (especie, fórmula, tiempo) para cada una de las
actividades enzimáticas: endoglucanasa, exoglucanasa, β-glucosidasa, endoxilanasa, lacasa y MnP durante
10 tiempos en los 32 días de fermentación; se obtuvieron diferencias significativas para todas las
combinaciones con un valor-p < 0,05. Posteriormente, se realizó un análisis por especie y una selección de
los máximos de cada una de las actividades enzimáticas y un análisis comparativo de los máximos
(empleando la función multcompare) aplicando una prueba de Kruskal-Wallis para cada actividad
enzimática, obteniendo diferencias significativas con valor-p < 0,05 para cada actividad enzimática
comparada (ver Tabla 2). Para concluir la selección de la mejor combinación especie-fórmula se realizó
un nuevo análisis comparativo a las cantidades de celulosa, hemicelulosa y lignina degradadas en 32 días
de fermentación en fase sólida aplicando la prueba de Kruskal-Wallis y una prueba de comparación con la
función multcompare a las combinaciones seleccionadas de la Tabla 2. De este análisis comparativo se
detectó que las tres especies de hongos en todas las combinaciones degradaron de forma similar los tres
componentes mayoritarios de los materiales lignocelulósicos, celulosa, hemicelulosa y lignina como se
resume en la Tabla3.

150
Del mismo modo, en la Tabla 3 se observa que todas las combinaciones presentaron porcentajes
cercanos de degradación de cada uno de los componentes de la fibra adicionada; C. versicolor – F5 fue el
que mejor comportamiento presentó respecto a la capacidad de degradación de lignina con 50,57%, P.
ostreatus en combinación con las fórmulas F5 y F11 fueron las que mostraron mayor capacidad de
degradación de hemicelulosa con 73,86% y 74,46% respectivamente y P. ostreatus – F5 degradó 59,12%
de celulosa. A estas combinaciones se les realizó un análisis comparativo a fin de seleccionar de entre las
tres combinaciones la de mejor comportamiento para aplicar un modelo matemático que describa los datos
experimentales de producción de enzimas lignocelulolíticas y biomasa y de consumo de los sustratos
medidos. Sin embargo, el análisis comparativo no presentó diferencia significativa (valor-p = 0.677), por
lo que se seleccionó la combinación P. ostreatus – F5 ya que fue la que arrojó el mayor promedio de las
medias.
3.4. Variación de las actividades enzimáticas durante la fermentación en fase sólida
La variación de las actividades enzimáticas durante los 32 días de fermentación sumergida no
presentó una tendencia definida para ninguna de las enzimas medidas para las tres especies de hongos
estudiados en las doce formulaciones de medios de cultivo. La actividad de las enzimas celulolíticas
medidas en este trabajo, endoglucanasa y exoglucanasa, fue variable con tendencia ascendente hasta el día
20 de incubación sin descenso definido a partir de allí como se observa en las Figuras 3 y 4; exceptuando
las combinaciones de las tres especies con la F1 para endoglucanasa (ver Figura 3A) que sí presentan un
ascenso hasta el día 20 de incubación y posterior descenso sostenido hasta el final de la incubación (día
32). Del mismo modo las combinaciones de las tres especies con F2 para exoglucanasa mostraron
tendencia al ascenso hasta el día 20 de incubación con descenso a partir de allí. Asimismo, en contraste
con el comportamiento de las combinaciones de las tres especies de hongos de F1 a F6, las actividades de
la endoglucanasa y exoglucanasa de las tres especies de hongos con las formulaciones F7 a F12
presentaron tendencia hacia el ascenso hasta el día 32 de incubación. En contraste, la actividad de la
endoxilanasa presentó un ascenso leve y tendencia a estabilizarse como se observa en la Figura 5, salvo
para las combinaciones de las tres especies con F6 las cuales tendieron a tener un marcado ascenso hasta
el día 20 de incubación y un posterior descenso hasta el día 32 de incubación. De igual forma para las
combinaciones de las tres especies con las formulaciones F7 a F12 se presentó tendencia a la estabilidad
posterior al ascenso entre los días 10 y 20 de la incubación. En referencia a la actividad de la β-
glucosidasa, el comportamiento de las combinaciones de cada uno de las especies de hongos entre las F1 y
F3 mostraron una alta variabilidad como se observan en la Figura 6 las combinaciones de P. ostreatus con
F4 a F6 se mostraron estables, así como todas las combinaciones de las tres especies con las
formulaciones F7 a F12. Las dos actividades de ligninasas medidas en este trabajo mostraron
comportamientos similares tendientes al ascenso con posterior estabilización al final del período de
incubación (32 días). La actividad de lacasa mostró mayor variabilidad en las combinaciones de las tres
especies con F1, especialmente para el P. ostreatus alcanzando una actividad cercana a 300 U/L.min
después de los 20 días de incubación. Las actividades de MnP variaron hacia el ascenso con alguna
diferencia para las combinaciones con la F9 cuyas actividades parecen tender hacia el descenso luego de
los 25 días de incubación como se observa en la Figura 8.
Muchos basidiomicetos tienen la capacidad de producir simultáneamente enzimas hidrolíticas y
oxidativas las cuales son necesarias para degradar los sustratos del complejo lignocelulósico (Kirk, 1983;
Wood y García, 1994). Estas enzimas son extracelulares e inducibles y según Buswell et al. (1996)
especialmente los hongos de pudrición blanca en general exhiben mayores títulos de ligninasas que de
celulasas; lo cual se convierte en una fortaleza de estos hongos ya que para lograr acceder a la celulosa
deben iniciar la degradación de la lignina. Sin embargo, en este estudio el material lignocelulósico
empleado, cascarilla de café, como la principal fuente de carbono, fue sometido a un pretratamiento en el
que se desordenó la matriz lignocelulósica permitiendo el acceso más rápidamente a la celulosa por las
tres especies de hongos. De hecho, según los datos experimentales obtenidos y consignados en las Tablas
2 y 3 las actividades enzimáticas máximas de todas las combinaciones y la capacidad de degradación de

151
celulosa y lignina estuvieron muy próximos unos de otros para las diferentes combinaciones de las tres
especies de hongos de trabajo con las 12 formulaciones, lo que indica que los hongos de pudrición blanca
como P. ostreatus, C. versicolor y L. edodes degradan material lignocelulósico bajo condiciones de
fermentación sumergida sin grandes diferencias cuando se adiciona material lignocelulósico de mayor
acceso a la celulosa (pretratado). Empero, la cantidad de hemicelulosa presente en los medios de cultivo
fue bastante baja en comparación con la hemicelulosa que se encuentra presente en la cascarilla de café sin
pretatamiento, ya que durante el pretratamiento con ácido sulfúrico debió haberse alterado este
componente y sus productos arrastrados durante el lavado del material antes de ser utilizado como
componente de los medios de cultivo. Este hecho se corroboró comparando los títulos de las actividades
enzimáticas obtenidas en este estudio con las obtenidas en investigaciones similares en las que obtuvieron
resultados comparables, a pesar de la gran cantidad de factores que afectan los resultados de las
actividades enzimáticas, tanto de las hidrolasas como de las ligninasas (Tlecutil-Beristain et al., 112; Mata
y Savoie, 1998; Mata et al., 2005; Elisashvili et al., 2008; Gassara et al., 2010).
Figura 3. Actividad enzimática de endoglucanasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 32 días de fermentación
sumergida
Figura 4. Actividad enzimática de exoglucanasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6
en 32 días de fermentación sumergida

152
Figura 5. Actividad enzimática de endoxilanasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6
en 32 días de fermentación sumergida
Figura 6. Actividad enzimática de β-glucosidasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes
sobre seis formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6
en 32 días de fermentación sumergida
Figura 7. Actividad enzimática de lacasa de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12
en 32 días de fermentación sumergida

153
Figura 8. Actividad enzimática de MnP de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12
en 32 días de fermentación sumergida
En el tiempo de fermentación en el que se realizaron todas las determinaciones enzimáticas, de la
biomasa y los componentes de la fibra se evidenció la formación de pellets de la biomasa enrollándose en
las mismas hifas y en este caso en particular, también en las partículas del material lignocelulósico
utilizado como parte de la fuente de carbono, la cascarilla de café pretratada. Asimismo, hubo un
incremento considerable de la viscosidad, hasta que los medios de cultivo adquirieron apariencia de
gelatinización (tiempo superior a 32 días de incubación), probablemente por la secreción de otras
sustancias como biopolímeros que cumplen funciones fisiológicas para el crecimiento y desarrollo de
estos basidiomicetos. Aunque los títulos de las actividades enzimáticas obtenidas presentaron en su
mayoría tendencia a la estabilización en los diferentes tiempos del proceso de fermentación sumergida, los
apreciables cambios en de los medios de cultivo durante el tiempo de fermentación, debieron producir
nuevas sustancias que pudieron haber generado algunas perturbaciones en las actividades enzimáticas,
provocando las disminuciones de las actividades enzimáticas en algunas de las combinaciones o por otras
causas como represión catabólica por productos de las reacciones que se generan en la fermentación;
producción de variantes isoenzimáticas de las mismas enzimas a diferentes tiempos y diferentes
condiciones del sustrato arrojan actividades diferentes. Sin embargo, estos descensos en las actividades
enzimáticas lignocelulolíticas también han sido detectados por varios investigadores, quienes reportan
algunas de estas hipótesis como posibles causantes de los descensos de dichas actividades enzimáticas
(Panagiotou et al., 2003; Mata et al., 2005; Levin et al., 2008; Montoya et al., 2011a).
3.5. Modelamiento matemático
Basado en los datos experimentales obtenidos de actividades enzimáticas lignocelulolíticas
durante los 32 días de fermentación sumergida para las tres especies de hongos basidiomicetos de
pudrición blanca: C. versicolor, P. ostreatus y L. edodes, se propuso un modelo matemático compuesto de
11 ecuaciones diferenciales (ver Tabla 4) a fin de buscar la descripción de los datos experimentales de
todas las combinaciones resultantes para P. ostreatus. Empero, aquí se presentan los resultados del
modelamiento arrojados para la combinación especie de hongo fórmula, F5, ya que ésta fue la que mostró
el mejor comportamiento respecto de la degradación de los sustratos basados en materiales
lignocelulósicos. De igual forma, para comparar el ajuste de los datos experimentales al modelo
matemático propuesto se planteó una prueba F de una cola, con la cual se obtuvieron las varianzas de los
residuales entre los datos experimentales y el modelo, individualmente para cada variable medida, pero
haciendo la comparación de cada ecuación aplicada para todas las formulaciones de F1 a F12 en la misma
especies, en este caso P. ostreatus, lo que permitió probar cual combinación fue mejor descrita por el
modelo en términos de la mejor representación de los datos experimentales obtenidos.

154
Con el modelo matemático propuesto se plantea la descripción de producción de biomasa, seis
enzimas lignocelulolíticas, azúcares reductores como producto intermedio consumido durante la misma
fermentación, el consumo de celulosa, hemicelulosa y la degradación de lignina. Asimismo, los datos
experimentales obtenidos en esta investigación serán ajustados al modelo planteado a fin de contribuir con
nuevas propuestas en el camino al planteamiento de modelos matemáticos más complejos que permitan la
descripción de los procesos de fermentación sumergida con fines industriales empleando hongos de
pudrición blanca. Varias ecuaciones cinéticas de complejidad variable, tales como la ecuación lineal,
logística, de dos fases o la ecuación de Monod han sido utilizadas por diferentes investigadores a fin de
buscar la descripción de la producción de biomasa en procesos de fermentación sumergida y en estado
sólido (Sangsurasak et al., 1996; Viccini et al., 2001). De las expresiones matemáticas utilizadas, la más
frecuente es la ecuación logística, debido probablemente a la forma simple de la expresión. El modelo
propuesto consignado en la tabla 4 para la descripción de la biomasa es la ecuación (1), la cual
corresponde a la ecuación logística modificada por Mitchell et al. (1999). Para describir la variación de los
azúcares reductores en el tiempo se propuso la ecuación (2) que correspondió a un factor de producción
constante de azúcares reductores y un consumo de los mismos por el hongo como fuente de energía
durante la fermentación sumergida. Para la degradación de lignina y el consumo de celulosa en el tiempo
de fermentación se plantearon las ecuaciones (3) y (8) cuyo planteamiento se realizó en función de las
concentraciones de las concentraciones de los mismos sustratos, es decir de las concentraciones de lignina
y celulosa respectivamente; mientras que el consumo de hemicelulosa se planteó en términos de la
concentración de la endoxilanasa como se muestra en la ecuación (6). La variación de las actividades
enzimáticas celulolíticas fueron propuestas para la exoglucanasa un coeficiente de producción de la
actividad enzimática por la concentración de la celulosa y para la endoglucanasa se planteó la producción
de la actividad enzimática igual que para exoglucanasa afectado por un factor de inhibición regido por la
concentración de azúcares reductores, estas dos expresiones están dadas por las ecuaciones (10) y (9)
respectivamente. Para la descripción de la ligninasas se plantearon dos expresiones simples de producción
de las enzimas de un coeficiente de producción de cada una por la concentración del sustrato sobre el cual
actúan, la lignina como se muestra en las ecuaciones (4) y (5) para la lacasa y MnP respectivamente.
Finalmente para la variación de la enzima β-glucosidasa se propuso una ecuación cuya producción
dependió únicamente de la variación de la degradación de celulosa con un factor de inhibición causado
por la concentración de los azúcares reductores y está representada en la ecuación (11). Los parámetros de
cada una de las ecuaciones consignadas en la Tabla 4 se describen en la tabla 5.
La fermentación sumergida es considerada en general como un proceso homogéneo por la
agitación que se da en todo el tiempo de la fermentación. Sin embargo, en el proceso que se llevó a cabo
en este estudio, la fermentación sumergida no puede considerarse como un proceso completamente
homogéneo, así durante todo el tiempo de la fermentación se hubiera mantenido en agitación; debido a
que hubo varios factores que no permiten tal consideración, en especial las características del medio de
cultivo, el cual a pesar de ser inicialmente un medio líquido, contenía el 10% de la masa como cascarilla
de café pretratada, la cual hizo parte de la fase sólida del medio de cultivo, diferentes de la biomasa. Por lo
anterior, no se pudo determinar en forma directa el crecimiento de la biomasa como peso seco del micelio,
fue necesario determinar la biomasa indirectamente a través de la medición del contenido de NAGA y
realizar cálculos para la obtención de los datos de biomasa en el tiempo de fermentación. Asimismo, la
medida del consumo del sustrato fue realizada únicamente por la determinación de los componentes de la
fibra. En estos procesos de fermentación se producen una gran cantidad de sustancias intermedias que
pueden generarse como producto de las reacciones bioquímicas durante las fases de crecimiento y
desarrollo de estos basidiomicetos, lo cual hace aún más compleja la descripción de los procesos de
fermentación sumergida y su escalamiento cuando se pretende aplicar estos procesos en producción
industriales para la obtención de alguna sustancia de interés (Pal et al., 1995a; Sarikaya y Ladisch, 1997;
Pandey et al., 2000; Moreira et al., 2003; Patil y Dayanand, 2006; Rodríguez-Couto y Toca-Herrera, 2007;
Elisashvili et al., 2008).
Los hongos basidiomicetos como los utilizados en el presente estudio contienen quitina en su
pared celular. Debido a que en los procesos de fermentación sumergida del presente estudio no fue posible

155
determinar directamente la cantidad de biomasa, se determinó el contenido de quitina a través de la
cuantificación de N-acetil-D-glucosamina (NAGA) en todos los tiempos de la fermentación y se le
determinó el contenido de NAGA al micelio seco obtenido de cultivo líquido a cada una de las tres
especies de trabajo, obteniendo como resultado 15,83% (p/p) en P. ostreatus, 14,15% (p/p) en C.
versicolor y 10,08% (p/p) en L. edodes. Estos porcentajes de NAGA fueron utilizados para calcular la
cantidad de biomasa de los medios de cultivo en cada uno de los tiempos de fermentación; asimismo,
estos datos de biomasa fueron los que se sometieron al modelamiento con la ecuación (1). Para la
descripción de la producción de biomasa de la combinación P. ostreatus – F5 el coeficiente n que afecta la
relación entre la concentración de biomasa y la concentración de biomasa máxima fue mayor a 1 (7,23) lo
que sugiere que P. ostreatus en estos medios de cultivo se comporta como un organismo relativamente
resistente a la autoinhibición. El ajuste de los datos experimentales al modelo de la biomasa y los azúcares
reductores se muestran en la Figura 9, en la cual se observa una marcada fase lag y una rápida producción
de azúcares reductores, probablemente debido a que la concentración inicial de los azúcares reductores en
este medio de cultivo es bastante baja y el hongo debe iniciar su proceso de generación de fuente de
energía a fin de iniciar su crecimiento y desarrollo. En la Tabla 6 se encuentran los valores de los
parámetros ajustados de la biomasa y los azúcares reductores y la bondad del ajuste en términos de la
varianza de los residuales, obteniendo para los datos de la biomasa un ajuste aproximado del 77% y para
los azúcares reductores del 76% referidos a la combinación que presentó la descripción del menor ajuste
(o el ajuste más pobre) que corresponde a la varianza de los residuales más alta.
Tabla 4. Modelo matemático para la descripción de producción de biomasa, enzimas lignocelulolíticas y consumo de
la matriz lignocelulósica
Ecuación Descripción
. 1
n
b bm b
bm
dC CC
dt C
(1) Biomasa
.pAR q Cb
d
dtm
C (2) Azúcares reductores
.LL L
dCk C
dt (3) Lignina
.laclac L
dCk C
dt (4) Lacasa
.MnPMnP L
dCk C
dt (5) Manganeso peroxidasa (MnP)
.HMHM ENX
dCk C
dt (6) Hemicelulosa
.ENXENX HM
dCk C
dt (7) Endoxilanasa
.CC
dCk Cc
dt (8) Celulosa
. .ENGENG C ENG AR
dCk C C
dt (9) Endoglucanasa
.EXGEXG C
dCk C
dt (10) Exoglucanasa
. .BG CBG BG AR
dC dCk
dt dtC
(11) β-glucosidasa

156
En la Figura 10 se presenta el comportamiento de los datos experimentales respecto al modelo
propuesto para la descripción de la degradación de la lignina y de dos ligninasas, lacasa y MnP descrita
por las ecuaciones (3), (4) y (5); los parámetros de las ecuaciones se encuentran consignados en la Tabla
6. De igual forma, el ajuste para los datos experimentales de la lignina, lacasa y MnP al modelo propuesto
fueron aproximadamente del 97%, 99% y 54% respectivamente para la lignina, lacasa y MnP de la
combinación P. ostreatus - F5.
En la Figura 11 se muestra el ajuste de los datos experimentales de la hemicelulosa y la enzima
endoxilanasa al modelo (ecuaciones (6) y (7)) y los parámetros del modelamiento se encuentran en la
Tabla 6. Los porcentajes de ajuste al modelo para la hemicelulosa y endoxilanasa fueron
aproximadamente del 94% y 96% respectivamente para F5. El modelamiento de los datos experimentales
de celulosa, endoglucanasa, exoglucanasa y β-glucosidasa se muestran en la Figura 12. Del mismo modo
los parámetros del modelo están consignados en la ¡Error! No se encuentra el origen de la referencia. y
el ajuste aproximado de los datos experimentales al modelo fue de 50% para celulosa, 75%
endoglucanasa, 83% exoglucanasa y 93% β-glucosidasa para la combinación P. ostreatus – F5.
Tabla 5. Parámetros de las ecuaciones del modelo matemático.
Parámetro Significado Parámetro Significado
µm
Velocidad específica de crecimiento de la biomasa
(día-1) kHM Coeficiente de consumo de hemicelulosa (día-1)
Cbm Concentración de biomasa máxima (g/L)
kENX Coeficiente de producción de Endoxilanasa
(U/min.g/día)
N
n < 1 el organismo es relativamente sensible a la
auto-inhibición y ésta ocurre para valores muy bajos
de Cb
n = 1 ecuación logística
n > 1 el organismo es relativamente resistente a la
auto-inhibición y ésta ocurre solo cuando Cb ≈ Cbm
CHM0 Concentración inicial de hemicelulosa (g/L)
Cb0 Concentración de biomasa máxima (g/L) CENX0 Actividad inicial de Endoxilanasa (U/L.min)
qp Coeficiente de producción constante de azúcares
reductores (g/L/día) kC Coeficiente de consumo de celulosa (día-1)
M Coeficiente de mantenimiento celular (día-1)
kENG Coeficiente de producción de Endoglucanasa
(U/min.g/día)
CAR0 Concentración inicial de azúcares reductores (g/L)
µENG Coeficiente de inhibición de Endoglucanasa
(U/min.g/día)
kL Coeficiente de degradación de lignina (día-1)
kEXG Coeficiente de producción de Exoglucanasa
(U/min.g/día)
klac Coeficiente de producción de lacasa (U/min.g/día) CC0 Concentración inicial de celulosa (g/L)
kMnP Coeficiente de producción de MnP (U/min.g/día) CENG0 Actividad inicial de Endoglucanasa (U/L.min)
CL0 Concentración inicial de lignina (g/L) CEXG0 Actividad inicial de Exoglucanasa (U/L.min)
Clac0 Actividad inicial de lacasa (U/L.min)
kBG Coeficiente de producción de β-glucosidasa
(U/g.min)
CMnP0
Actividad inicial de MnP (U/L.min) µBG
Coeficiente de inhibición de β-glucosidasa
(U/g.min/día)
CBG0 Actividad inicial de β-glucosidasa (U/L.min)

157
Figura 9. Perfil en el tiempo de la biomasa celular y los azúcares reductores para la combinación P. ostreatus – F5.
Los puntos discretos corresponden a los datos experimentales; las líneas continuas son calculadas por el modelo
propuesto.
Figura 10. Ajuste de datos experimentales de lignina, lacasa, MnP para la combinación P. ostreatus – F5. Los puntos
discretos corresponden a los datos experimentales; las líneas continuas son calculadas por el modelo propuesto.
Figura 11. Ajuste de datos experimentales de hemicelulosa y endoxilanasa para la combinación P. ostreatus – F5.
Los puntos discretos corresponden a los datos experimentales; las líneas continuas son calculadas por el modelo
propuesto.

158
Figura 12. Ajuste de datos experimentales de celulosa, endoglucanasa, exoglucanasa y β-glucosidasa para la
combinación P. ostreatus – F5. Los puntos discretos corresponden a los datos experimentales; las líneas continuas
son calculadas por el modelo propuesto
Tabla 6. Valores de los parámetros del ajuste de los datos experimentales al modelo para P. ostreatus – F5
Parámetro Valor Parámetro Valor
µm 0.1523 día-1 kENX 347,7099 U/min.g/día
Cbm 76,1285 g/L CHM0 1,5859 g/L
n 7,2339 CENX0 1,578 U/L.min
Cb0 1,5001 g/L kC 0,035 día-1
qp 0,2608 g/L/día kENG 20,2676 U/min.g/día
m 0,0092 día-1 µENG 121,3156 U/min.g/día
CAR0 1,3916 g/L kEXG 4,4188 U/min.g/día)
kL 0,025 día-1 CC0 47,7708 g/L
klac 0,1433 U/min.g/día CENG0 1,1202 U/L.min
kMnP 0,1322 U/min.g/día) CEXG0 1750 U/L.min
CL0 38,66 g/L kBG 893,3498 U/g.min
Clac0 32 U/L.min µBG 167,3245 U/g.min/día)
CMnP0 7,3 U/L.min CBG0 817,99 U/L.min
kHM 6,39x10-6 día-1
Aunque existen numerosas investigaciones previas acerca de la producción de actividades
enzimáticas lignocelulolíticas por diversos hongos de pudrición blanca, y la degradación de materiales
lignocelulósicos; y aproximaciones sobre la formación de productos intermedios en procesos de
fermentación sumergida (Bizukojc y Ledakowicz, 2006; Rodríguez-Couto y Toca-Herrera, 2007). Se
requiere aun mayor información sobre las cinéticas de formación de biomasa, de producción de enzimas,
del consumo de nutrientes y degradación de la lignina que permitan la descripción del proceso global y
posterior escalado para la obtención de alguno de los productos de interés como los que se plantean en
este estudio empleando como materias primas residuos lignocelulósicos pretratados a fin de buscar bajar
los costos de producción de estas biosustancias. El planteamiento de las ecuaciones para la descripción de
la cinética de cada una de las variables medidas se realizó bajo la premisa de darle a cada expresión
matemática algún sentido biológico respecto al compuesto a describir. Las expresiones propuestas
describieron aceptablemente cada una de los compuestos determinados en el presente estudio. Sin
embargo, algunas de las ecuaciones describieron mejor los datos experimentales obtenidos que otras,
como en el caso de la descripción de la degradación de lignina y hemicelulosa, y producción de lacasa,
endoxilanasa y β-glucosidasa, las cuales dieron un porcentaje de ajuste en la descripción mayor al 90%,
mientras que para la degradación de celulosa y producción de MnP las ecuaciones propuestas arrojaron un
ajuste cercano al 50% lo que indica que deberá buscarse nuevas expresiones que mejoren los porcentajes
de ajuste a los datos experimentales. Por último, debe considerarse la posibilidad de medir algunos de los
productos intermedios originados por la acción de las enzimas medidas, u otras enzimas no evaluadas y

159
sus productos a fin de contribuir a una interpretación más acabada de los complejos fenómenos que
ocurren durante el crecimiento de los macromicetos en los procesos de fermentación sumergida.
4. Conclusiones
En el presente estudio, las tres especies de hongos de pudrición blanca probadas sobre las doce
formulaciones de sustrato propuestas crecieron bien durante los 32 días de fermentación sumergida, por lo
que se concluye que tanto la relación C/N como las concentraciones de CuSO4 no afectaron drásticamente
el crecimiento y desarrollo de ninguno de los tres hongos. Sin embargo, de la evaluación detallada de los
datos experimentales si se observaron diferencias en la producción de las enzimas lignocelulolíticas
durante los 32 días de fermentación, en la que a pesar de la gran variabilidad de las actividades
enzimáticas en el tiempo, unas combinaciones hongo formulación mostraron mejor comportamiento
respecto a la producción de las enzimas que fueron determinadas. Del mismo modo, el hongo P. ostreatus
mostró mejor comportamiento en cuanto a la capacidad de degradación de lignina y celulosa en la mayoría
de los medios de cultivo, hecho que probablemente ocurrió por el desorden generado a la matriz
lignocelulósica por el pretratamiento realizado a la cascarilla de café utilizada como fuente principal de
carbono en este proceso.
El modelo matemático planteado en este estudio para la representación de los procesos de
fermentación sumergida se realizó buscando describir el desarrollo de estos hongos de pudrición blanca
sobre medios de cultivo no convencionales, a fin de buscar puntos generales de interpretación de estos
procesos que por ser heterogéneos se hacen complejos y carecen de expresiones matemáticas directas que
permitan el escalamiento de los mismos con fines industriales, evitando hacer experimentación individual
de cada proceso.
Agradecimientos
Los autores agradecen a la Vicerrectoría de Investigaciones y Postgrados de la Universidad de Caldas
(Colombia) por la financiación del presente trabajo y al Instituto de Biotecnología Agropecuaria de la
misma institución por el apoyo material y de infraestructura. Asimismo, los autores expresan sus
agradecimientos a la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires
(Argentina) por su apoyo y asesoría durante la realización de este trabajo.
Referencias Baldrian P, Gabriel J (2002) Variability of laccase activity in the whiterot basidiomycete Pleurotus ostreatus. Folia
Microbiologica 47: 385-390.
Barr BK, HsieH YL, Ganem B, Wilson DB (1996) Identification of two functionally different classes of exocellulases.
Biochemistry 35: 586-592.
Béguin P, Aubert JP (1994) The biological degradation of cellulose. FEMS Microbiology Reviews 13: 25-58.
Bizukojc M, Ledakowicz S (2006) A kinetic model to predict biomass content for Aspergillus niger germinating spores in the
submerged culture. Process Biochemistry 41: 1063-1071.
Blanchette RA (1995) Degradation of the lignocellulose complex in wood. Canadian Journal of Botany 73: S999-S1010.
Bonugli-Santos R, Durrant L, Durães Sette L (2012) The production of ligninolytic enzymes by marine-derived basidiomycetes
and their biotechnological potential in the biodegradation of recalcitrant pollutants and the treatment of textile effluents.
Water Air Soil Pollution 223: 2333-2345.
Buswell JA, Cai Y, Chang ST (1995) Effect of nutrient nitrogen and manganese on manganese peroxidase and laccase production
by Lentinula (Lentinus) edodes. FEMS Microbiology Letters 128: 32- 45.
Buswell JA, Cai YJ, Chang ST (1996) Ligninolytic enzyme production and secretion in edible mushroom fungi. En Royse DJ
(Ed.) Mushroom Biology and Mushroom Products. The Pennsylvania State University pp. 113±122. Pennsylvania.
Carlile M, Watkinson S, Gooday G (2001) The fungi. Academic Press. London. 588 p. p.
Dormand JR, Prince PJ (1980) A family of embedded Runge-Kutta formulae. Journal of Computational and Applied Mathematics
6: 19-26.

160
Elisashvili V, Penninckx M, Kachlishvili E, Asatiani M, Kvesitadze G, Gadd G (2006) Use of Pleurotus dryinus for
lignocellulolytic enzymes production in submerged fermentation of mandarin peels and tree leaves. Enzyme and
Microbial Technology 38: 998-1004.
Elisashvili V, Penninckx M, Kachlishvili E, Tsiklauri N, Metreveli E, Kharziani T, Kvesitadze G (2008) Lentinus edodes and
Pleurotus species lignocellulolytic enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes
of different composition. Bioresource Technology 99: 457-462.
Gassara F, Brar S, Tyagi RD, Verma M, Surampalli RY (2010) Screening of agro-industrial wastes to produce ligninolytic
enzymes by Phanerochaete chrysosporium. Biochemical Engineering Journal 49: 388-394.
Hölker U, Höfer M, Lenz J (2004) Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Applied
Microbiology and Biotechnology 64: 175-186.
Hudson H (1986) Fungal Biology. Edward Arnolds Eds. Maryland. 298 p. p.
Ikasari L, Mitchell DA (2000) Two-phase model of yhe kinetics of growth of Rhizopus oligosporus in membrana culture.
Biotechnology Bioengineering 68: 619-627.
Joselau JP, Ruel K (1994) Wood polysaccharides and their degradation by fungi. En Ouellette P (Ed.) Host Wall Alterations by
Parasitic Fungi. APS Press. Minnesota.
Kapich AN, Prior BA, Botha A, Galkin S, Lundell T, Hatakka A (2004) Effect of lignocellulose-containing substrate on
production of ligninolytic peroxidases in submerged cultures of Phanerochaete chrysosporium ME-446. Enzyme and
Microbial Technology 34: 187-195.
Kirk TK (1983) Degardation and convertion of lignocelluloses. En Smith JE, Berry DR, Kristiansen P (Eds.) The filamentous
fungi. London.
Kirk TK, Farrel RL (1987) Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology 41: 465-
505.
Kjeldahl J (1883) Neue Methode Zùr Bestimmung der Stickstoffs in Organichen Kòrpen. Zeitschrift fur Analytische Chemie 22:
366-382.
Kovarova-Kovar K, Egli T (1998) Growth kinetics of suspended microbial cells: from single substrate-controlled growth to
mixed-substrate kinetics. Microbiology and Molecular Biology Reviews 62: 646-666.
Leterme P. (2010). Análisis de alimentos y forrajes, in Protocolos de Laboratorio Universidad Nacional de Colombia Sede
Palmira: Palmira. 66 pp.
Levin L, Forchiassin F, Ramos AM (2002) Copper induction of lignin-modifying enzymes in the white-rot fungus Trametes
trogii. Mycologia 94(3): 377-383.
Levin L, Herrmann C, Papinutti VL (2008) Optimization of lignocellulolytic enzyme production by the white-rot fungus Trametes
trogii in solid-state fermentation using response surface methodology. Biochemical Engineering Journal 39: 207-214.
Mata G, Savoie JM (1998) Extracellular enzyme activities in six Lentinula edodes strains during cultivation in wheat straw. World
Journal of Microbiology & Biotechnology 14: 513-519.
Mata G, Murrieta Hernández DM, Iglesias Andreu LG (2005) Changes in lignocellulolytic enzyme activites in six Pleurotus spp.
strains cultivated on coffee pulp in confrontation with Trichoderma spp. World Journal of Microbiology &
Biotechnology 21: 143-150.
Miller GL (1959) Use of DinitrosaIicyIic Acid Reagent for Determination of Reducing Sugar. Analytical Chemistry 31(3): 426-
428.
Mitchell D, Stuart D, Tanner R. (1999). Solid-State fermentation - Microbial growth kinetics, in The Encyclopedia of Bioprocess
Technology: Fermentation, Biocatalysis and bioseparation, Flickinger M, Drew S, EditorsWiley: New York. 2407-
2429
Mitchell DA, Von meien OF, Krieger N, Dalsenter FD (2004) A review of recent developments in modeling of microbial growth
kinetics and intraparticle phenomena in solid-state fermentation. Biochemical Engineering Journal 17: 15-26.
Montoya BS, Orrego CA, Levin L (2011) Growth, fruiting and lignocellulolytic enzyme production by the edible mushroom
Grifola frondosa (maitake). World Journal of Microbiology & Biotechnology ISSN 0959-3993: DOI 10.1007/s11274-
11011-10957-11272.
Moreira MT, Feijoo G, Lema JM (2003) Fungal bioreactors: Applications to white-rot fungi. Reviews in Environmental Science
and Bio/Technology 2: 247-259.
Niladevi KN, Prema P (2008) Effect of inducers and process parameters on laccase production by Streptomyces psammoticus and
its application in dye decolourization. Bioresource Technology 99 4583-4589.
Pal M, Calvo A, Terron MC, Gonzalez AE (1995) Solid-state fermentation of sugarcane bagasse with Flammulina velutipes and
Trametes versicolor. World Journal of Microbiology & Biotechnology 11: 541-545.
Panagiotou G, Kekos D, Macris B, Christakopoulos P (2003) Production of cellulolytic and xylanolytic enzymes by Fusarium
oxysporum grown on corn stover in solid state fermentation. Industrial Crops and Products 18: 37-45.
Pandey A, Soccol CR, Mitchell D (2000) New developments in solid state fermentation: I-bioprocesses and products. Process
Biochemistry 35: 1153–1169.
Paszczczynski A, Crawford R, Huyn V (1988) Manganese peroxidase of Phanerochaete chrysosporium purification. Methods
enzymology 161: 264-270.
Paszczynski A, Crawford RL (1991) Degradation of azo compounds by ligninases from Phanerochaete chrysosporium Involment
of veratryl alcohol. Biochemistry Biophysics Research Communications 178: 1056-1063.

161
Patil S, Dayanand A (2006) Optimization of process for the production of fungal pectinases from deseeded sunflower head in
submerged and solid-state conditions. Bioresource Technology 97: 2340-2344.
Plassard C, Mousain D, Salsac L (1982) Estimation of mycelial growth of basidiomycetes by means of chitin determination.
Phytochemistry 21: 345-348.
Rodriguez-Couto S, Toca-Herrera JL (2007) Laccase production at reactor scale by filamentous fungi. Biotechnology advances
25: 558.569.
Sangsurasak P, Nopharatana M, Mitchell D (1996) Mathematical modeling of the growth of filamentous fungi in solid-state
fermentation. Journal Science Industrial Resources 55: 333-342.
Sarikaya A, Ladisch M (1997) An unstructured mathematical model for growth of Pleurotus ostreatus on lignocellulosic material
in solid-state fermentation systems. Applied Biochemistry and Biotechnology 62: 72-85.
Solomon EI, Sundaram UM, Machonkin TE (1997) Multicopper oxidases and oxygenases. Chemical Reviews 96: 2563-2605.
Tang Y-J, Zhu L-W, Li H-M, Li D-S (2007) Submerged Culture of Mushrooms in Bioreactors – Challenges, Current State-of-the-
Art, and Future Prospects. Food Technology and Biotechnology 45(3): 221-229.
Tavares A, Coelho M, Coutinho J, Xavier A (2005) Laccase improvement in submerged cultivation: induced production and
kinetic modelling. Journal of Chemical Technology and Biotechnology 80: 669-676.
Tišma M, Sudar M, Raĉki S, Zelic B (2010) Mathematical model for Trametes versicolor growth in submerged cultivation.
Bioprocess and Biosystems Engineering 33: 749-758.
Tlecutil-Beristain S, Sánchez C, Loera O, Robson G, Díaz-Godínez G (112) Laccases of Pleurotus ostreatus observed at different
phases of its growth in submerged fermentation: production of a novel laccase isoform. Micologycal Research 1080 -
1084.
Tsiklauri ND, Khardziani TS, Kachlishvili ET, Elisashvili VI (1999) Cellulase and xylanase activities of higher basidiomycetes
during bioconversion of plant raw materials depending on the carbon source in the nutrient medium. Applied
Biochemistry and Microbiology 35: 291-295.
Van de Lagemaat J, Pyle DL (2005) Modelling the uptake and growth kinetics of Penicillium glabrum in a tannic acid-containing
solid-state fermentation for tannase production. Process Biochemistry 40: 1773-1782.
Vasic-Racki D, Kragl U, Liese A (2003) Benefits of enzyme kinetics modelling. Chemical and Biochemical Engineering
Quarterly 17: 7-18.
Viccini G, Mitchell D, Boit S, Gern J, da Rosa A, Costa R (2001) Analysis of growth kinetic profiles in solid-state fermentation.
Food Technology and Biotechnology Advances 39: 271-294.
Vrsalovic Presecki A, Findrik Z, Zelic B (2006) Modeling of the biotransformation processes. Chemical and Biochemical
Engineering Quarterly 20: 227-241.
Walkley A, Black I (1934) An estimation of the Degtjareft method for determining of soil organic matter and a proposed
modification of chromic acid titration method. Soil Science 37: 29-38.
Wood TM, García CV (1994) Enzymes and mechanisms involved in microbial cellulolysis. Kluwer Academic publishers.
Netherlands.
Zelic B, Vasic Racki D, Wandrey C, Takors R (2004) Modeling of the pyruvate production with Escherichia coli in a fed-batch
bioreactor. Bioprocess and Biosystems Engineering 26: 249-258.

162
8. Selección de la Mejor Combinación Hongo –
Formulación para la producción de exo-polisacáridos
mediante Fermentación sumergida con Pleurotus
ostreatus, Coriolus versicolor y Lentinus edodes

163
Selección de la Mejor Combinación Hongo – Formulación para la producción
de exo-polisacáridos mediante Fermentación sumergida con Pleurotus
ostreatus, Coriolus versicolor y Lentinus edodes
Sandra Montoya1, Óscar Julián Sánchez
2*, Laura Levin
3
RESUMEN
En esta investigación se probaron tres especies de hongos de pudrición blanca: Coriolus
versicolor, Pleurotus ostreatus y Lentinus edodes sobre 12 formulaciones de medios de cultivo líquidos
con cascarilla de café pretratada como fuente de materiales lignocelulósicos y dos niveles de sulfato de
cobre (II). Los objetivos de este trabajo fueron i) evaluar los efectos de la relación C/N y del sulfato de
cobre (II) sobre la producción de EPS durante 32 días de fermentación sumergida; así como el consumo de
celulosa y ii) proponer un modelo matemático para la descripción de la producción de EPS, la producción
de biomasa, producción y consumo de azúcares reductores y consumo de celulosa. Las tres especies de
hongos crecieron sobre todas las formulaciones de los medios de cultivo y se obtuvieron EPS, pero no en
las cantidades esperadas probablemente por la adición de sulfato de amonio a los medios de cultivo como
fuente de nitrógeno. Del mismo modo, no hubo una gran variación entre las combinaciones de las tres
especies de hongos con las diversas formulaciones sobre la degradación de la celulosa presente en los
medios de cultivo, posiblemente porque el material lignocelulósico adicionado fue pretratado y esto debió
generar un desorden en la matriz lignocelulósica de modo que las hifas de los hongos pudieron acceder
más fácilmente a la celulosa. Las combinaciones con C. versicolor arrojaron las mayores producciones de
EPS, por lo que este hongo fue seleccionado en las diferentes combinaciones para la aplicación del
modelo matemático compuesto por cuatro ecuaciones diferenciales propuesto para describir los datos
experimentales.
Palabras clave: hongos de pudrición blanca, relación C/N, producción de EPS, degradación de celulosa.
1. INTRODUCCIÓN
Los polisacáridos de los hongos macromicetos han sido reconocidos como excelentes
inmunoestimuladores debido al potencial de sus propiedades terapéuticas, a que no son tóxicos y son
escasos sus efectos secundarios en comparación con otros inmunoestimulantes. Estos polisacáridos
bioactivos son producidos en cantidades relevantes a partir de los cuerpos fructíferos de los hongos, su
micelio y además, de los filtrados obtenidos de sus cultivos en fermentación sumergida. En mayor medida
las estructuras de los polisacáridos procedentes de los macromicetos corresponden a homoglicanos,
heteroglicanos o complejos de gluco-proteínas. La fuente principal de polisacáridos antitumorales parecen
ser sus paredes celulares, las cuales están compuestas de polisacáridos. En general, la actividad
inmunomoduladora de los polisacáridos de los macromicetos está relacionada con las estructuras β-(1→3),
β-(1→6) glucanos, α-(1→3) glucanos y complejos de polisacáridos de galactomananos y glucuromananos
unidos a proteínas presentes en la pared celular de los macromicetos (Enshasy, 2010). El β-D-glucano es
un polisacárido que produce exclusivamente D-glucosa luego de la hidrólisis ácida (Mizuno, 1999). Los
polisacáridos obtenidos por fermentación sumergida con una adecuada selección del medio de cultivo, se
denominan exopolisacáridos (EPS), ya que se excretan en el medio de cultivo durante el curso de la
fermentación. Los intrapolisacáridos (IPS) pueden ser producidos a partir de los cuerpos fructíferos y su
micelio después de haber sido sometido a diferentes procesos de extracción sólido-líquido utilizando
diferentes disolventes, desde agua caliente hasta diferentes mezclas de disolventes orgánicos (Amaral et
al., 2008; Komura et al., 2010).

164
Los exopolisacáridos de basidiomicetos de pudrición blanca juegan un rol importante en los
procesos de crecimiento y desarrollo de estos organismos. Estos exopolisacáridos pueden inmovilizar las
exoenzimas que ellos mismo excretan (Tang y Zhong, 2002). Según Catley (1992) el gel formado por
estos biopolímeros previene la deshidratación de la hifa, permitiendo la adherencia de otras células a las
superficies de los sustratos, posibilitando la selección de las moléculas del ambiente que los rodea
(Maziero et al., 1999). Adicionalmente, estos biopolímeros tienen un amplio rango de aplicaciones
industriales.
Las diferentes especies de hongos producen diferentes tipos de polisacáridos con diversas
propiedades y la estructura del polisacárido influye sobre la actividad biológica que se obtiene. Del mismo
modo las estructuras químicas de los polisacáridos dependen de la fuente de donde provienen, el
carpóforo, el micelio o el medio de cultivo, estos compuestos varían en sus estructuras moleculares, la
unidad estructural que se repite (el monómero) y los pesos moleculares, entre otros factores (Tsukagoshi
et al., 1984; Ishikawa et al., 1992; Hamlyn y Schmidt, 1994; Kobayashi et al., 1995; Zhuang y Mizuno,
1999; Chang y Miles, 2004; Mohammad-Fata et al., 2007). Por ejemplo, el lentinan un β-(1→3)-D-
glucano extraído de L. edodes es una potente droga inmunoestimuladora licenciada en Japón para terapias
antitumorales (Kupfahl et al., 2006). De igual forma se ha reportado actividad antitumoral de los
complejos de polisacáridos extraídos del micelio o del cuerpo fructífero de hongos macromicetos, como el
complejo proteína-polisacárido PSK del hongo C. versicolor (Ooi y Liu, 2000a), el complejo polisacárido-
proteína del cultivo de Tricholoma lobayense (Liu et al., 1996a), exobiopolímeros de varias especies de
Cordyceps a las cuales se les ha demostrado actividad antitumoral (Chen et al., 1997),
inmunopotenciadora (Nakamura et al., 1999), actividad hipoglicémica (Kiho et al., 1999) y efecto
hipocolesterolémico (Koh et al., 2003). Estos ejemplos son una muestra de la potencialidad que tienen los
hongos como fuente de sustancias bioactivas para la obtención de potenciadores del sistema
inmunológico, teniendo en cuenta que de las 140.000 especies de macromicetos que se estima existen en
el planeta, solamente se conocen cerca del 10%, de las cuales sólo han sido examinadas desde su potencial
inmunomodulador el 5% (Lindequist et al., 2005b).
La suplementación de los sustratos es una estrategia para incrementar la producción de biomasa
celular y una forma de mantener constante la producción de polisacáridos (Tang y Zhong, 2002).
Asimismo, diferentes trabajos publicados en la última década evidencian la importancia de la presencia de
aceites vegetales ricos en ácido oleico como promotores de crecimiento de la biomasa fúngica y por ende
de sus polisacáridos constituyentes (Yang et al., 2000; Hsieh et al., 2008). Al momento de seleccionar el
tipo de aceite vegetal o ácido graso a utilizar como inductor de polisacáridos es necesario considerar la
composición de ácidos grasos del aceite; ya que ha sido reportado por varios investigadores que
cantidades importantes de ácido linoleico suprimen la producción de biomasa y polisacáridos, mientras
que la presencia de ácido oleico promueve su producción (Park et al., 2002a; Hsieh et al., 2006; Hsieh et
al., 2008). Los aceites de soya y de oliva han sido utilizados como promotores de crecimiento celular y de
polisacáridos en cultivos de macromicetos, tanto líquidos como sólidos. Al respecto, el aceite de oliva
contiene 84% de ácido oleico y entre 30 y 60% en aceite de soya con bajos contenidos de ácido linoleico
(Wade, 1993; Hsieh et al., 2008).
En general, el proceso de determinación de intra y exo-polisacáridos se inicia mediante la
medición de la cantidad de polisacáridos como carbohidratos totales presentes en una muestra
seleccionada. Estos polisacáridos corresponden a los contenidos en los cuerpos fructíferos de los hongos,
al micelio o a los presentes en los caldos de cultivo de fermentaciones sumergidas.
El objetivo de este trabajo fue evaluar el efecto de la relación carbono nitrógeno, del sulfato de cobre (II) y
de aceite de soya sobre la producción de exo-polisacáridos (EPS) por tres especies de hongos de pudrición
blanca (L. edodes, C. versicolor y P. ostreatus) empleando doce formulaciones diferentes de medios de
cultivo por fermentación sumergida. Estos hongos fueron seleccionados de entre diez hongos de pudrición
blanca mediante un rastreo basado en la medida de los polisacáridos como carbohidratos totales; los
resultados de este rastreo se relacionan en el artículo anterior. Igualmente este trabajo tuvo como objetivo
adicional proponer y validar un modelo matemático para la mejor combinación hallada especie de hongo -

165
formulación a fin de realizar la descripción de la cinética de crecimiento del organismo, y la producción
de EPS en condiciones de fermentación sumergida.
2. MATERIALES Y MÉTODOS
2.1. Microorganismos
Tres especies de basidiomicetos fueron empleados para la producción de polisacáridos en
fermentación en estado sólido, tomadas de la colección de hongos de la Universidad de Caldas
(Manizales, Colombia): Lentinula edodes del Centro de Investigaciones del Café, Chinchiná, Caldas,
Colombia (CICL54), Coriolus versicolor de Pennsylvania State University, Estados Unidos de Norte
América (PSUWC430), y Pleurotus ostreatus de la Colección interna de la Universidad de Caldas
(UCC001). Las cepas fueron extendidas sobre agar papa dextrosa (PDA) y mantenidas en refrigeración a
4°C y transferencias periódicas.
2.2. Pretratamiento de cascarilla de café
La cascarilla de café seca y molida fue sumergida en H2SO4 al 3% en relación 1:4 sólido: líquido y
sometida a ultrasonido por una hora a 80°C con posterior tratamiento térmico a 121°C y 15 psig por una
hora con fines de modificar la matriz lignocelulósica. En seguida, la cascarilla fue lavada con exceso de
agua hasta obtener un pH de 6.8 en el agua de lavado del material para retirar el exceso de ácido. La
cascarilla lavada fue utilizada como fuente de carbono proveniente de material lignocelulósico en los
medios de cultivo para la fermentación sumergida del presente trabajo.
2.3. Medios de cultivo
Se realizaron doce formulaciones de medios de cultivo por triplicado con base en un litro de
medio de cultivo. La composición por cada litro de medio de cultivo fue: 100g de cascarilla de café
pretratada y seca, 2g KH2PO4, 0,5g MgSO4.7H2O, 0,1g CaCl2, 2g de extracto de levadura y tres
concentraciones de SO4(NH4)2 0,5g, 1g y 1,5g, dos concentraciones de aceite de soya 13,5g y 22,42g y
dos concentraciones de sulfato de cobre (II) 0,1592g (fórmulas pares) y 0,0798g (fórmulas impares), con
la variación de los niveles de SO4(NH4)2 y aceite de soya se modificó la relación carbono nitrógeno (R
C/N) (ver Tabla 1). Los medios de cultivo fueron servidos en matraces de 250 mL de capacidad con 100
mL de medio, esterilizados a 121°C por 20 minutos, los pH de los medios de cultivo variaron entre 5.6 y
5.8 para todas las formulaciones medido antes de la inoculación; seguidamente, los medios de cultivo
fueron inoculados en cabina de flujo laminar con un fragmento de micelio extendido en agar PDA de ½
cm de lado de cada especie de trabajo. Los matraces fueron llevados a incubación con agitación de 100
rpm, 25°C y penumbra por 32 días. Se tomaron 10 muestras, dos por semana y la primera muestra fue
tomada el día de la inoculación.
Tabla 1. Relaciones carbono nitrógeno (R C/N) en doce niveles (formulaciones)
F1 F2 F3 F4 F5 F6 F7 F8 F9 F10 F11 F12
24,68 24,13 28,40 26,84 23,98 22,06 23,41 23,79 20,81 20,61 20,70 21,04
2.4. Determinación de la relación carbono nitrógeno
La relación carbono nitrógeno (C/N) fue determinada a partir de la medida de la materia orgánica
de las doce formulaciones de sustratos empleando el método de Walkley y Black (1934), el cual utiliza
ácido sulfúrico concentrado y solución de dicromato de potasio. La muestra se lee a 585 nm. La cantidad

166
de carbono total corresponde al 58% de la materia orgánica. El nitrógeno orgánico total fue determinado
por el método de Kjeldahl (Kjeldahl, 1883).
2.5. Determinación de polisacáridos como carbohidratos totales
La precipitación de los EPS se provocó a partir de una fracción de 150 µL en 2mL de etanol
concentrado grado reactivo con agitación por 1 horas. Luego se realizó una centrifugación por 20 min a
14.000 rpm descartando el sobrenadante. El precipitado (la muestra) fue resuspendida en NaOH 1N a
60°C por 1 hora, posteriormente, la suspensión fue sometida al método fenol-sulfúrico para determinar la
cantidad de carbohidratos totales a 490 nm (Dubois et al., 1956).
2.6. Cuantificación del contenido de N-acetil-D-glucosamina (NAGA) y biomasa
Las muestras de sustrato fueron colectadas en 10 tiempos de incubación hasta el final de la
fermentación (día 32) para las tres especies de trabajo. El medio de fermentación fue filtrado al vacío, con
posterior secado en estufa a 101°C hasta peso constante. A la muestra seca y molida se le determinó el
contenido de quitina. El contenido de biomasa fúngica en este medio fue estimado por la determinación
del contenido de N-acetil-D-glucosamina (NAGA) después de la hidrólisis con HCl 6N grado analítico
según el método de Plassard et al. (1982). Para ello se cuantificó paralelamente el contenido de NAGA
por gramo de micelio en cada una de las tres especies, cultivándolas en medio líquido. El contenido de
NAGA del micelio de cada una de las especies de hongos de trabajo: C. versicolor, P. ostreatus y L.
edodes fue determinado del desarrollo del micelio en medio líquido en matraces de 250 mL con 100 mL
de medio de cultivo con la siguiente composición: glucosa (30 g/L), extracto de levadura (6 g/L),
SO4Mg.5H2O (0,5 g/L), K2HPO4 (0,5 g/L) y CaCl2 (0,1 g/L). El tiempo de fermentación para el
crecimiento del hongo fue de 25 días y sometidos a 100 rpm de agitación. Los compuestos solubles en
agua fueron extraídos durante la hidrólisis con HCl 6N por tres horas.
2.7. Cuantificación de de los componentes de la fibra y azúcares reductores
Se determinaron los componentes de la fibra (celulosa, hemicelulosa y lignina) de los medios de
cultivo de cada una de las formulaciones en 10 tiempos durante los 32 días de incubación, así como la
fracción soluble, empleando los resultados de la determinación de fibras detergente neutra, ácida y
lignino-ácida. La medición se realizó a una muestra seca y molida de sustrato; dicha muestra se sometió a
tres hidrólisis en serie (cada una de 75 min): i) hidrólisis con lauril sulfato de sodio y otros, ii) hidrólisis en
solución de bromuro de amonio en ácido sulfúrico 1N, y iii) hidrólisis con ácido sulfúrico al 72% (p/v). Al
finalizar cada hidrólisis, las muestras fueron lavadas y secadas a 105°C hasta peso constante (Leterme,
2010a). El contenido de azúcares reductores como glucosa fue determinado por el método del ácido
dinitrosalicílico (DNS) (Miller, 1959).
2.8. Diseño experimental
Se planteó un diseño experimental bifactorial con la especie a tres niveles y doce formulaciones de
sustrato. Las formulaciones varían en su relación C/N y la concentración de sulfato de cobre (II). Las
variables independientes en este diseño experimental fueron la especie de hongo, la formulación del
sustrato y el tiempo de fermentación. Las variables de respuesta medidas fueron la producción de biomasa,
de polisacáridos, producción y consumo de azúcares reductores y la degradación de la celulosa en 10
tiempos durante 32 días de fermentación en estado sólido.
2.9. Modelamiento matemático del crecimiento del hongo y la producción de enzimas

167
Para la solución de los modelos matemáticos del proceso de fermentación sumergida se utilizó el
software Matlab® 2010b (MathWorks, EUA) en un PC de 3 GB de memoria RAM y un procesador Intel
CORE-i3 de 2,13 GHz. Para los modelos matemáticos planteados en ecuaciones diferenciales ordinarias
se utilizó el comando ode45 que está basado en una fórmula explícita de Runge-Kutta (4,5) usando un par
de Dormand-Prince (Dormand y Prince, 1980) y el comando ode15s basado en una fórmula de orden
variable que funciona mediante fórmulas de diferenciación numérica.
2.10. Análisis estadístico
Para todos los análisis estadísticos del presente trabajo se utilizó el software Matlab ® 2010b. Este
análisis se realizó en tres etapas. Primero se realizó un ANOVA a todos los datos del diseño experimental
y sus posibles combinaciones: especie, fórmula y tiempo de fermentación mediante el comando anovan.
Luego se realizó un análisis comparativo (Kruskal Wallis) de la producción de polisacáridos para cada
especie de hongo estudiado y las doce formulaciones de sustratos mediante los comandos kruskalwallis y
multcompare.
Para evaluar si el modelo matemático propuesto describe adecuadamente los datos
experimentales, se realizó una prueba F de una cola que parte de las siguientes hipótesis:
2
,
0 2
exp,
2
,
1 2
exp,
: 1
: 1
i
i
i
i
residuales F
datos F
residuales F
datos F
SH
S
SH
S
donde: Fi está dado para i = 1,2,…,12, s2residuales es la varianza de los residuales (desviaciones de los datos
experimentales con respecto a los valores calculados por el modelo matemático) y se calcula para cada
variable de respuesta medida y s2datos exp es la varianza de la serie experimental de cada variable de
respuesta medida. Para estos análisis se utilizaron los comandos vartest y vartest2 de Matlab. Con las
varianzas de los residuales obtenidos para cada serie experimental se calcularon los porcentajes de cada
una respecto a la serie que dio la mayor varianza de residuales a fin de mostrar el ajuste de los datos
experimentales al modelo matemático propuesto de forma más simple.
3. RESULTADOS Y DISCUSIÓN
3.1. Efecto de la relación C/N y sulfato de cobre sobre la producción de biomasa y polisacáridos
Las tres especies de hongos (C. versicolor, P. ostreatus y L. edodes) probadas en el diseño
experimental, fueron inoculadas e incubadas bajo las mismas condiciones ambientales en las doce
formulaciones de los medios de cultivo. El intervalo de la relación C/N en las doce formulaciones de
sustrato evaluadas fue de 20 a 28,4 con una variación en los contenidos de nitrógeno de 1.4 a 1,93% en
base seca. Las tres especies de hongos se desarrollaron bien en todas las formulaciones evaluadas durante
los 32 días de incubación, como se observa en las Figuras 1 y 2. Todos los contenidos de nitrógeno de las
formulaciones estimularon el crecimiento micelial de los macromicetos durante la fermentación,
incrementando el contenido de biomasa (ver Figuras 1 y 2) en todos los casos y con una marcada fase lag
para las tres especies en todos los medios de cultivo. Empero se debe tener en cuenta que la variación de la
relación C/N se provocó modificando los contenidos de (NH4)2SO4 y aceite de soya y no modificando las
fuentes de nitrógeno y de carbono en cada formulación. Sin embargo, sí hubo variabilidad sobre la
producción de EPS, ya que no para la especie que produjo mayor cantidad de biomasa, fue la que mas

168
excretó EPS. Por ejemplo, C. versicolor – F2 produjo 241,93 mg/L, la mayor cantidad de EPS no fue el
que formó la mayor cantidad de biomasa como se observa en la Tabla 2. Con el fin de seleccionar la
combinación de mejor comportamiento en cuanto a la producción de EPS se realizó un análisis estadístico
por etapas. Primero se realizó un ANOVA a fin de determinar si hubo diferencias significativas entre los
resultados obtenidos respecto a las variables independientes especie, fórmula y tiempo, obteniendo
diferencia significativa para las tres combinaciones evaluadas (especie, formulación y tiempo de
fermentación) con valor–p < 0.05; posteriormente, se realizó una selección de los máximos de las
producciones de EPS, obteniendo que C. versicolor en varias de las combinaciones evaluadas fue la
especie que mayor cantidad de EPS produjo, en especial las combinaciones F2, F5 y F11 (ver Tabla 2).
En este estudio se presentó la producción de EPS de tres especies de hongos de pudrición blanca,
en fermentación sumergida con una fuente fija de material lignocelulósico, la cascarilla de café pretratada,
variando las cantidades de sulfato de amonio y aceite de soya adicionados por litro de medio de cultivo
para conseguir las diferentes relaciones C/N. La cascarilla de café pretratada se adicionó con el fin de
activar la producción de enzimas lignocelulolíticas por las especies de hongos utilizadas. Entendiendo que
para la bioconversión de los materiales lignocelulósicos con hongos, las enzimas que degradan
polisacáridos juegan un rol muy importante ya que son las que proveen la fuente de energía a los
microorganismos; por lo que la baja producción de estas enzimas pueden limitar la velocidad de
producción de azúcares solubles. Sin embargo, en este trabajo los organismos probablemente pudieron
aprovechar como fuentes de carbono, los contenidos en el aceite de soya y extracto de levadura para su
mantenimiento durante las prolongadas fases lag que se visualizan para todas las combinaciones hongo
formulaciones en las Figuras 1 y 2, mientras los mismos hongos inician la degradación del material
lignocelulósico a fin de tener la cantidad de nutrientes necesarios para su crecimiento y desarrollo.
Del mismo modo, es importante tener en cuenta que las diferentes especies de hongos pueden
tener dependencia de las diferentes fuentes de carbono y esto provocar en ellos la variación en su
crecimiento y en la formación de los tipos de polisacáridos presentes en su pared celular y los
exopolisacáridos que excretan al medio. Según Xiao et al. (2010) la producción de polisacáridos y el
crecimiento de la biomasa de las diferentes especies de hongos dependen directamente de la fuente de
carbono suministrada, lo que sugiere que la composición del medio puede influir en la composición y el
mecanismo de producción tanto de los intra como de los exopolisacáridos para una determinada especie de
hongo. Asimismo, la cantidad y fuente de nitrógeno en la producción de los polisacáridos de los hongos y
su influencia en su crecimiento y desarrollo es definitivo, ya que el nitrógeno es un componente esencial
de las proteínas y ácidos nucleicos y su deficiencia afecta el crecimiento de los hongos y los metabolitos
que producen durante su crecimiento y desarrollo (Xiao et al., 2006). La fuente de nitrógeno utilizada en
este trabajo fue suministrada por extracto de levadura y el SO4(NH4)2 básicamente, se variaron tres niveles
de sulfato de amonio y dos niveles de aceite en las formulaciones de sustrato a fin de obtener la variación
de las relaciones C/N, con un intervalo en los contenidos de nitrógeno entre 1.4 a 1,93% en base seca
como ya se mencionó arriba. Sin embargo, el crecimiento de las tres especies de hongos no fue igual para
las 12 formulaciones, ni la producción de EPS como se observa en las Figuras 1, 2, 3 y 4. Se presentaron
grandes variaciones, como en el caso de la combinación C. versicolor – F9 que para el final del período de
fermentación se obtuvo 26,9428 mgEPS/L; en este caso en particular, el hongo pudo haber entrado en una
fase diferente de su crecimiento, que le haya generado una caída en la secreción de los EPS. Según se
observa en los datos experimentales obtenidos para producción de EPS, si parece haberse generado una
influencia de la relación C/N sobre la producción de los mismos, ya que las formulaciones F1, F2, F3, F4
y F5 fueron las de mayor relación C/N y para esas combinaciones se obtuvieron los mayores contenidos
de EPS, mientras que para las formulaciones de menor relación C/N (F9, F10, F11 y F12) se obtuvieron
menores cantidades de EPS, con excepción de la combinación C. versicolor – F11 con 183,2586
mgEPS/L.
Hay disponible gran cantidad de investigaciones en referencia a los efectos de las fuentes de
carbono y nitrógeno sobre la producción de EPS; en las que se han probado diversas fuentes de carbono,
como monosacáridos y disacáridos, y varias fuentes de nitrógeno de origen orgánico e inorgánico. Se
reportó por Park et al. (2002b) un incremento de producción de EPS de 2,3 a 7,5 g/L cuando se adicionó el

169
4% de aceite de oliva al medio de cultivo; Wu et al. (2006) reportaron las concentraciones de EPS
obtenidas utilizando diferentes monosacáridos y disacáridos como fuentes de carbono, de los que
reportaron como la mejor fuente productora de EPS, la glucosa. Asimismo, varios investigadores
coinciden en expresar que la adición de fuentes de nitrógeno inorgánico a los medios de cultivo produce
inhibición en la producción de EPS, hecho que pudo haber ocurrido en este estudio, ya que se utilizó como
parte de la fuente de nitrógeno, sulfato de amonio. De la misma manera, los doce medios de cultivo
utilizados en este trabajo contenían 100 g/L de cascarilla de café pretratada, la cual pudo haber generado
alguna interrupción en el crecimiento y desarrollo de los hongos y por ende afecta la producción de EPS.
Respecto a la producción específica de EPS (mg EPS/g biomasa seca) es un factor de importancia
a tener en cuenta debido a que la evaluación de este tipo de procesos en términos de eficiencia sobre una
variable de referencia es lo indicado, para lo cual la biomasa es una buena elección, ya que ella es la que
produce los EPS y en general siempre es determinada. Lo que se observa en la Tabla 2 es que no
necesariamente la combinación que produjo la mayor cantidad de biomasa no fue la que produjo la mayor
cantidad de EPS, como fue el caso de la combinación L. edodes – F12 para la cuales e obtuvo 3.099
mgEPS/g biomasa seca.
El efecto de la sal de cobre no fue claro, ya que para las formulaciones impares se adicionó 0,1596
g/L y para las formulaciones pares 0,0798 g/L y en este caso las bajas producciones no presnetarón
tendencia por las formulaciones pares o impares. No obstante, debe considerarse la composición de la
cascarilla de café utilizada en este estudio, su procedencia y el pretratameinto al que fue sometida antes de
iniciar la fermentación. Con los pretratamientos a los materiales lignocelulósicos se busca generar
cambios en la composición química y características físicas de los sustratos a fin de que sean más
exequibles para los hongos; además, se debe tener en cuenta que durante la fermentación pudieron haberse
producido sustancias intermedias que hayan generado una disminución en la velocidad de crecimiento de
este hongo sobre las diversas combinaciones que no dieron una respuesta satisfactoria respecto a la
producción de EPS. Por lo anterior, las condiciones de cultivo y su composición química si parecen un
factor determinante en la producción de EPS. La cantidad de EPS obtenidos en este trabajo en todas las
combinaciones especie de hongo fórmula fueron comparables con los resultados más bajos obtenidos por
Xiao et al. (2010) cuando probaron medios de cultivo con fuentes de carbono como galactosa y maltosa.
A pesar de todo el esfuerzo que se ha dado para explorar el efecto de las condiciones de cultivo en
la producción de polisacáridos de hongos, aún hace falta explorar el uso de una gran variedad de materias
primas procedentes de residuos industriales y agroindustriales a fin de disminuir los costos de producción
con miras hacia la industrialización (Lee et al., 1999; Shu y Lung, 2003; Lee et al., 2004; Wu et al., 2004;
Wu et al., 2006; Lee et al., 2007b). Finalmente, es razonable esperar que la producción de ambos tipos de
polisacáridos, IPS y EPS podrían estar vinculados uno con otro, por la misma dependencia con el
crecimiento de la biomasa (Lama et al., 1996). Por lo tanto, las características físico-químicas de los
medios de cultivo y condiciones medioambientales siempre deberán ser tenidas en cuenta a fin de
satisfacer la producción de intra y exo-polisacáridos acorde con las necesidades particulares de la especie
de hongo a utilizar.

170
Figura 1. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6
Figura 2. Producción de biomasa de tres especies de hongos C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 en 32 días de fermentación
sumergida
3.2. Producción de polisacáridos en la mejor combinación resultante del diseño experimental
Los datos experimentales obtenidos en el presente estudio fueron sometidos a un proceso de
selección a fin de lograr la mejor combinación especie de hongo fórmula del diseño experimental
bifactorial respecto a la producción de polisacáridos, el cual se realizó por etapas. Primero se hizo un
análisis de varianza, ANOVA a tres vías (especie, fórmula, tiempo) para los polisacáridos obtenidos
durante 10 tiempos en los 32 días de fermentación; se obtuvieron diferencias significativas para todas las
combinaciones con un valor-p < 0,05. Posteriormente, se realizó un análisis por especie y una selección de
los máximos de cada una de las cantidades de polisacáridos obtenidos y un análisis comparativo de los
máximos (empleando la función multcompare) y aplicando una prueba de Kruskal-Wallis, obteniéndose
diferencias significativas con valor-p < 0,05. De este análisis comparativo se obtuvo que la mejor
combinación para la producción de polisacáridos fue C. versicolor – F2 seguida de las combinaciones con
F5 y F11. A estas tres combinaciones se les realizó un análisis comparativo a fin de seleccionar de entre
las cuatro combinaciones la de mejor comportamiento respecto a la producción de polisacáridos y aplicar
un modelo matemático que describa los datos experimentales de producción de biomasa, de polisacáridos,

171
de producción y consumo de azúcares reductores, y de consumo de celulosa. Sin embargo, el análisis
comparativo no presentó diferencia significativa (valor-p = 0.131), por lo que se aplicó el modelo
matemático a las tres combinaciones de C. versicolor.
En la Tabla 2 se presentan los datos de las producciones de biomasa, polisacáridos y la relación de
la producción de polisacáridos con la producción de biomasa (producción específica de polisacáridos) a
los 32 días de fermentación (final del período), en la que se observó que la mayor producción de biomasa
por combinación fue de 81.514, 100.04 y 83.052 g/L para P. osteratus – F7, C. versicolor – F10 y L.
edodes – F10 respectivamente; mientras que la máxima producción de polisacáridos fue de 241.9377
mg/L, 183.282 mg/gss y 194.922 mg/L para las combinaciones C. versicolor –F2, P. ostreatus – F4 y L.
edodes – F10 respectivamente, de lo que se concluye que las combinaciones que más crecieron no fueron
las que más EPS produjeron. En concordancia con lo anterior, la producción específica de polisacáridos
para la combinación C. versicolor – F2 también fue la que arrojó la mayor relación 2,5937 mgEPS/g
biomasa seca; aunque no fue así para todas las combinaciones. De lo anterior, se puede inferir que además
de la importancia que tiene la cantidad de polisacáridos producidos, otro importante parámetro es la
relación de producción de polisacáridos respecto a la cantidad de biomasa producida, al momento de
tomar decisiones en torno a la producción de EPS por fermentación sumergida buscando mejorar las
condiciones de proceso y los rendimientos.
Figura 3. Producción de EPS de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F1, B: F2, C: F3, D: F4, E: F5, F: F6 en 32 días de fermentación sumergida
En las formulaciones probadas en este estudio, la principal fuente de carbono empleada fue la
cascarilla de café pretratada, por lo que en este trabajo se describe el consumo de celulosa. Los polímeros
de celulosa y hemicelulosa fueron degradados por acción de las enzimas hidrolíticas de los hongos a
azúcares reductores a fin de conseguir la fuente de energía necesaria para su crecimiento y desarrollo
(Ângelo, 2004). La lignina, fue degradada por óxidorreductasas excretadas por estos hongos que, como
agentes causantes de pudrición blanca, son los únicos hongos capaces de degradarla completamente,
facilitando en el proceso el acceso a la celulosa (Kirk y Farrel, 1987b). En esta fermentación los tres
hongos degradaron el material, pero no hasta el consumo total de la celulosa en los 32 días de
fermentación sumergida, la cantidad de hemicelulosa presente en los medios de cultivo fue muy baja
debido al pretratamiento del material antes de la fermentación, por lo que aquí solo se mostrará el
consumo de celulosa en la próxima sección.

172
Tabla 2. Producción máxima de polisacáridos de tres especies de hongos: C. versicolor, P. ostreatus y L.
edodes sobre doce formulaciones y durante 32 días de incubación
Fórmula
Biomasa (g/L) EPS (mg/L) Fracción (mg EPS/g biomasa seca)
P. ostreatus C. versicolor L. edodes P. ostreatus C. versicolor L. edodes P. ostreatus C. versicolor L. edodes
F1 79,488076 96,1549 68,3441 137,521 122,0016 84,5715 1,73009 1,268803 1,23744
F2 78,96429 93,27701 76,2122 139,918 241,9377 115,269 1,77191 2,593755 1,51247
F3 74,319357 94,87854 74,7643 143,912 176,8915 183,396 1,9364 1,864399 2,45299
F4 77,16561 93,75387 74,8809 183,282 140,1461 110,818 2,37518 1,49483 1,47993
F5 76,641824 93,33484 74,3362 179,402 187,6184 183,054 2,34079 2,010165 2,46251
F6 74,319357 92,3518 75,4801 146,537 146,6507 168,903 1,97171 1,587957 2,23772
F7 81,51408 92,08865 79,4033 74,4152 118,2358 146,308 0,91291 1,283934 1,8426
F8 67,737463 86,21135 65,0756 108,992 96,21134 74,4152 1,60904 1,115994 1,14352
F9 65,422958 83,68695 61,7939 26,9428 116,524 110,134 0,41183 1,39238 1,78227
F10 77,719356 100,0393 83,052 81,8327 157,0353 145,966 1,05293 1,569736 1,75753
F11 67,965624 80,73473 57,956 86,1691 183,8526 110,704 1,26783 2,277243 1,91014
F12 65,642319 84,53524 62,8967 72,2469 74,30104 194,922 1,10062 0,878936 3,09908
Figura 4. Producción de EPS de tres especies de hongos: C. versicolor, P. ostreatus, L. edodes sobre seis
formulaciones de sustratos diferentes. A: F7, B: F8, C: F9, D: F10, E: F11, F: F12 en 32 días de fermentación
sumergida
3.5. Modelamiento matemático
Basado en los datos experimentales obtenidos de EPS durante los 32 días de fermentación
sumergida para las tres especies de hongos basidiomicetos de pudrición blanca: C. versicolor, P. ostreatus
y L. edodes, se propuso un modelo matemático compuesto de 4 ecuaciones diferenciales (ver Tabla 3) a
fin de ajustar los datos experimentales de todas las combinaciones resultantes para C. versicolor, ya que
éste fue el hongo que mayor producción de EPS mostró en todas las combinaciones medidas. De hecho, en
este trabajo se presentan los resultados del modelamiento arrojados para las combinaciones C. versicolor
con las formulaciones F2, F5 y F11, ya que estas combinaciones fueron las que presentaron la mayor
producción de EPS durante los 32 días de fermentación. De igual forma, para comparar el ajuste de los
datos experimentales al modelo matemático propuesto se planteó una prueba F de una cola, con la cual se

173
obtuvieron las varianzas de los residuales entre los datos experimentales y el modelo, individualmente
para cada variable medida, pero haciendo la comparación de cada ecuación aplicada para todas las
formulaciones de F1 a F12 en la misma especie, en este caso C. versicolor, lo que permitió probar la
bondad del ajuste del modelo en términos de la mejor representación de los datos experimentales
obtenidos.
Con el modelo matemático propuesto se plantea la descripción de producción de biomasa, los
azúcares reductores como producto intermedio consumido durante la misma fermentación, el consumo de
celulosa y la producción de EPS. Asimismo, los datos experimentales obtenidos en esta investigación
serán ajustados al modelo propuesto a fin de contribuir con nuevas propuestas en el camino al
planteamiento de modelos matemáticos más complejos que permitan la descripción de los procesos de
fermentación sumergida con fines industriales. Varias ecuaciones cinéticas de complejidad variable, tales
como la ecuación lineal, logística, de dos fases o la ecuación de Monod han sido utilizadas por diferentes
investigadores a fin de buscar la descripción de la producción de biomasa en procesos de fermentación en
estado sólido (Sangsurasak et al., 1996; Viccini et al., 2001). De las expresiones matemáticas utilizadas, la
más frecuente es la ecuación logística, debido probablemente a la forma simple de la expresión. El modelo
propuesto consignado en la Tabla 3 para la descripción de la biomasa es la ecuación (1), la cual
corresponde a la ecuación logística modificada por Mitchell et al. (1999). Para describir la variación de los
azúcares reductores en el tiempo se propuso la ecuación (2) que correspondió a un primer factor de
producción constante de azúcares y un segundo factor de consumo de azúcares para el crecimiento y
desarrollo de los hongos. Para el consumo de celulosa en el tiempo de fermentación se planteó la ecuación
(4), cuyo planteamiento se realizó en función de la concentración de la celulosa y un parámetro que afecta
el consumo de la misma. La producción de los EPS fue expresada por la ecuación (3) que representa la
producción de metabolitos primarios en procesos de fermentación. Los parámetros de cada una de las
ecuaciones consignadas en la Tabla 3 se describen en la Tabla 4.
La fermentación sumergida para la producción de EPS con basidiomicetos es un proceso que
potencialmente puede ser utilizado con fines industriales, pero el éxito a escala comercial depende de los
costos que arrojen las nuevas tecnologías en comparación con las tecnologías existentes y si la industria
encuentra alguna ventaja. La expansión de las nuevas tecnologías para la producción de metabolitos como
los EPS se verá facilitada por el incremento en los rendimientos de los bioproductos y el desarrollo de
procesos con tecnologías exequibles. El cultivo sumergido con macromicetos se caracteriza por un
marcado incremento en la viscosidad en el caldo de cultivo con el curso de la fermentación, el cual es
debido al incremento de la biomasa, los cambios de morfología del crecimiento y la producción de
productos extracelulares que alteran el carácter reológico del cultivo; el aumento de la viscosidad es
generalmente considerada como un fenómeno indeseable pero inevitable, ya que causan dificultades en el
suministro de oxígeno, en la eliminación del dióxido de carbono y en la provisión de la agitación (Wagner
et al., 2003; Zhong y Tang, 2004; Tang et al., 2007). Todos estos aspectos han disminuido las
posibilidades de industrialización de muchos procesos de obtención EPS por fermentación sumergida
empleando setas, debido a que el planteamiento de modelos matemáticos que contribuyan con la
descripción de los procesos se dificulta y no hay una gran disponibilidad de propuestas que posibiliten la
descripción de procesos de producción de múltiples sustancias útiles.
La fermentación sumergida desde un esquema muy general es considerada un proceso
homogéneo, pero realmente cuando se trabaja con hongos macromcietos bajo condiciones de fermentación
sumergida y se adicionan materiales lignocelulóscios como parte de los componentes del medio de
cultivo, se convierte en un proceso heterogéneo que se ve afectado además por las condiciones
ambientales. Por lo anterior, la determinación del crecimiento de la biomasa y el consumo de los sustratos
se dificulta y es necesario recurrir a métodos indirectos de medida en varias ocasiones (Pal et al., 1995b;
Sarikaya y Ladisch, 1997; Pandey et al., 2000; Moreira et al., 2003).
En los procesos de fermentación sumergida del presente estudio no fue posible determinar
directamente la cantidad de biomasa de las tres especies de hongo, por lo que fue necesario determinarles
el contenido de quitina a través de la cuantificación de N-acetil-D-glucosamina (NAGA) en todos los
tiempos de la fermentación. Asimismo, se le determinó el contenido de NAGA al micelio seco obtenido

174
de cultivo líquido a cada una de las tres especies de trabajo, obteniendo como resultado 15,83% (p/p) en P.
ostreatus, 14,15% (p/p) en C. versicolor y 10,08% (p/p) en L. edodes. Estos porcentajes de NAGA fueron
utilizados para calcular la cantidad de biomasa de los medios de cultivo en cada uno de los tiempos de
fermentación; de igual forma, estos datos de biomasa fueron los que se sometieron al modelamiento con la
ecuación (1). Para la descripción de la producción de biomasa de las tres combinaciones de C. versicolor
que se presentan en esta trabajo, el coeficiente n que afecta la relación entre la concentración de biomasa y
la concentración de biomasa máxima fue mayor a 1 en los tres casos (ver Tabla 5) lo que sugiere que C.
versicolor en estos medios de cultivo se comportó como un organismo relativamente resistente a la
autoinhibición. En las Figuras 5, 6 y 7 se muestra el ajuste de los datos experimentales al modelo para las
tres combinaciones seleccionadas, en las cuales se observa una marcada fase lag y una rápida producción
de azúcares reductores, probablemente debido a que la concentración inicial de los azúcares reductores en
estos medio de cultivo es bastante baja y el hongo debe iniciar su proceso de generación de fuente de
energía a fin de iniciar su crecimiento y desarrollo. De igual forma, el consumo de celulosa y la
producción de EPS conservan la misma tendencia para las tres combinaciones, aunque la combinación de
F11 demuestra un menor ajuste de los datos experimentales con el modelo. En la Tabla 5 están
consignados los valores de los parámetros ajustados de cada una de las ecuaciones diferenciales
propuestas luego de solucionar el sistema de ecuaciones y los porcentajes que representaron la bondad del
ajuste en términos de la varianza de los residuales se encuentran consignados en la Tabla 6, obteniendo
para las variables de la combinación C. versicolor – F11 un ajuste aproximado para la biomasa del 60%,
azúcares reductores del 72%, celulosa 92,5% y los EPS 50%.
Tabla 3. Modelo matemático para descripción de producción de biomasa, consumo de celulosa y producción de EPS
Ecuación Descripción
. 1
n
b bm b
bm
dC CC
dt C
(1) Biomasa
.p bAR q C
dC
tm
d (2) Azúcares reductores
.CC C
dCk C
dt (3)
Celulosa
.EPS bEPS
dC dCk
dt dt (4) Polisacáridos
Tabla 4. Parámetros de las ecuaciones del modelo matemático. Parámetro Significado
µm Velocidad específica de crecimiento de la biomasa (día-1)
Cbm Concentración de biomasa máxima (g/L)
n
n < 1 el organismo es relativamente sensible a la auto-inhibición y
ésta ocurre para valores muy bajos de Cb
n = 1 ecuación logística
n > 1 el organismo es relativamente resistente a la auto-inhibición
y ésta ocurre solo cuando Cb ≈ Cbm Cb0 Concentración de biomasa máxima (g/L)
qp Coeficiente de producción constante de azúcares reductores
(g/L/día)
m Coeficiente de consumo de azúcares reductores (día-1)
CAR0 Concentración inicial de azúcares reductores (g/L)
kC Coeficiente de consumo de celulosa (día-1)
CC0 Concentración inicial de celulosa (g/L)
kEPS Coeficiente de producción de polisacáridos (mg/mg)
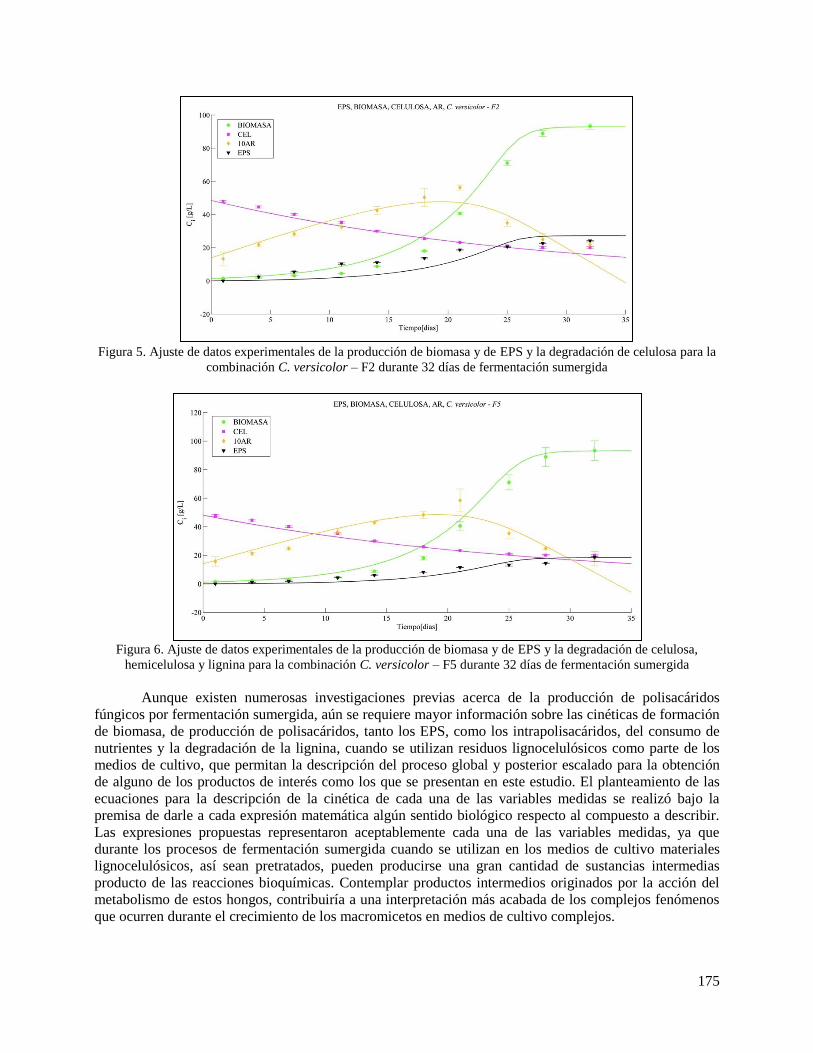
175
Figura 5. Ajuste de datos experimentales de la producción de biomasa y de EPS y la degradación de celulosa para la
combinación C. versicolor – F2 durante 32 días de fermentación sumergida
Figura 6. Ajuste de datos experimentales de la producción de biomasa y de EPS y la degradación de celulosa,
hemicelulosa y lignina para la combinación C. versicolor – F5 durante 32 días de fermentación sumergida
Aunque existen numerosas investigaciones previas acerca de la producción de polisacáridos
fúngicos por fermentación sumergida, aún se requiere mayor información sobre las cinéticas de formación
de biomasa, de producción de polisacáridos, tanto los EPS, como los intrapolisacáridos, del consumo de
nutrientes y la degradación de la lignina, cuando se utilizan residuos lignocelulósicos como parte de los
medios de cultivo, que permitan la descripción del proceso global y posterior escalado para la obtención
de alguno de los productos de interés como los que se presentan en este estudio. El planteamiento de las
ecuaciones para la descripción de la cinética de cada una de las variables medidas se realizó bajo la
premisa de darle a cada expresión matemática algún sentido biológico respecto al compuesto a describir.
Las expresiones propuestas representaron aceptablemente cada una de las variables medidas, ya que
durante los procesos de fermentación sumergida cuando se utilizan en los medios de cultivo materiales
lignocelulósicos, así sean pretratados, pueden producirse una gran cantidad de sustancias intermedias
producto de las reacciones bioquímicas. Contemplar productos intermedios originados por la acción del
metabolismo de estos hongos, contribuiría a una interpretación más acabada de los complejos fenómenos
que ocurren durante el crecimiento de los macromicetos en medios de cultivo complejos.

176
Figura 7. Ajuste de datos experimentales de la producción de biomasa y de EPS y la degradación de celulosa,
hemicelulosa y lignina para la combinación C. versicolor – F11 durante 32 días de fermentación sumergida
Tabla 5. Parámetros de ajuste de los datos experimentales al modelo matemático para la producción de EPS de tres
combinaciones de C. versicolor
Parámetro F2 F5 F11
µm 0.1626 día-1 0,1726 día-1 0,1799 día-1
Cbm 928713 g/L 93,2247 g/L 80,8324 g/L
N 5,9475 4,7975 4,1732
Cb0 1,4660 g/L 1,2 g/L 0,90 g/L
qp 0.25045 g/L/día 0,2545 g/L 0,2789 g/L
CAR0 1,3925 g/L 1,3925 g/L 0,9799 g/L
M 0.007214 día-1 0,007699 día-1 0.0088 día-1
kC 0.035 día-1 0.035 día-1 0,045 día-1
CC0 48,4907 g/L 48,1267 g/L 48,12 g/L
kEPS 0.002982 mg/mg 0,001991 mg/mg 0,00235 mg/mg
Tabla 6. Ajuste (%) de la descripción del modelo matemático a los datos experimentales de tres combinaciones de C.
versicolor con tres formulaciones para la producción de EPS por fermentación sumergida
Fórmula Biomasa Azúcares
reductores Polisacáridos Celulosa
F2 52 67 1 95
F5 40 64 66,6 95
F11 60 72 50 92,5
4. Conclusiones
En el presente estudio, el crecimiento de las tres especies de hongos de trabajo y la producción de
EPS pudieron haber estado afectadas más que por la relación C/N, por la adición de sulfato de amonio, ya
que en otras investigaciones se ha probado que la presencia de fuentes inorgánicas de nitrógeno en los
medios de cultivo disminuyen la producción de EPS. Por el contrario, las concentraciones de CuSO4 no
parecen haber afectado drásticamente el crecimiento y desarrollo de ninguno de los tres hongos, ya que no
se observó tendencia en la disminución o incremento de la biomasa o de los EPS para los dos niveles
utilizados.
El modelo matemático planteado en este estudio para la representación de los procesos de
fermentación sumergida se realizó buscando describir el desarrollo de tres hongos de pudrición blanca
sobre sustratos lignocelulósicos a fin de buscar puntos generales de interpretación de estos procesos que
por ser tan heterogéneos se hacen complejos y no hay disponibilidad de muchas expresiones matemáticas

177
que permitan el escalamiento de los mismos con fines industriales, evitando hacer experimentación
individual de cada proceso.
Agradecimientos
Los autores agradecen a la Vicerrectoría de Investigaciones y Postgrados de la Universidad de
Caldas (Colombia) por la financiación del presente trabajo y al Instituto de Biotecnología Agropecuaria de
la misma institución por el apoyo material y de infraestructura. Asimismo, los autores expresan sus
agradecimientos a la Facultad de Ciencias Exactas y Naturales de la Universidad de Buenos Aires
(Argentina) por su apoyo y asesoría durante la realización de este trabajo.
Referencias
Amaral A, Carbonero E, Simao R, Kadowaki M, Sassaki G, Osaku C, Gorin P, Lacomini M (2008) An unusual water-soluble B-
glucan from the basidiocarp of the fungus Ganoderma resinaceum. Carbohydrate Polymers 72: 473-478.
Ângelo AS (2004) Enzimas hidrolíticas. En Esposito, Azevedo JI (Eds.) Fungos: Uma Introducâo à biologia, bioquimica e
biotecnologia. Universidade de Caxias do Sul,.
Catley BJ (1992) The biochemistry of some fungal polysaccharides with industrial potencial. En Mukerji KG (Ed.) Handbook of
applied mycology: fungal biotechnology. Marcel Dekker,. New York.
Chang ST, Miles PG (2004) Mushrooms Cultivation, Nutritional Value, Medicinal effect, and Enviromental Impact. CRC Press.
New York. 451 p.
Chen YJ, Shiao MS, Lee SS, Wang SY (1997) Effects of Cordyceps sinensis on the proliferation and differentiation of human
leukemic U937 cells. Life Science 60(23): 49-59.
Dormand JR, Prince PJ (1980) A family of embedded Runge-Kutta formulae. Journal of Computational and Applied Mathematics
6: 19-26.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related
substances. Analytical Chemistry Engineering Journal 28(3): 350-356.
Enshasy HE (2010) Immunomodulators. En Hofrichter M (Ed.) The Mycota. Springer-Verlag. Berlin, 477 p.
Hamlyn PF, Schmidt RJ (1994) Potential therapeutic application of fungal filaments in wound management. Mycologist 8: 147-
152.
Hsieh C, Liu C-J, Tseng M-H, Lo C-T, Yang Y-C (2006) Effect of olive oil on the production of mycelial biomass and
polysaccharides of Grifola frondosa under high oxygen concentration aeration. Enzyme and Microbial Technology 39:
434-439.
Hsieh C, Wang H-L, Chena C-C, Hsub T-H, Tseng M-H (2008) Effect of plant oil and surfactant on the production of mycelial
biomass and polysaccharides in submerged culture of Grifola frondosa. Biochemical Engineering Journal 38: 198-205.
Ishikawa K, Majima T, Ebina T (1992) Anti-tumour effect of a Coriolus preparation, PSK: Induction of macrophage chemotactic
factor (MCF) in spleens of tumour bearing mice. Biotherapy 5: 251-258.
Kiho T, Ookubo K, Usui S, Ukai S, Hirano K (1999) Structural features and hypoglycemic activity of a polysaccharides (CS-F10)
from the cultured mycelium of Cordyceps sinensis. Biological and Pharmaceutical Bulletin 22(9): 66-70.
Kirk TK, Farrel RL (1987) Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology 41: 465-
505.
Kjeldahl J (1883) Neue Methode Zùr Bestimmung der Stickstoffs in Organichen Kòrpen. Zeitschrift fur Analytische Chemie 22:
366-382.
Kobayashi H, Matsunaga K, Oguchi Y (1995) Antimetastatic Effects of P5K (Krestin), a Protein-bound Polysaccharide Obtained
from Basidiomycetes: An Overview. Cancer Epidemiology, Biomarkers & Prevention 4: 275-281.
Koh JH, Kim JM, Chang UJ, Suh HJ (2003) Hypocholesterolemic effect of hotwater extract from mycelia of Cordyceps sinensis.
Biological and Pharmaceutical Bulletin 26: 84-87.
Komura DL, Ruthers AC, Carbonero ER, Alquini G, Rosa MC, Sassaki GL, Lacomini M (2010) The origin of mannans found in
submerged culture of basidiomycetes. Carbohydrate polymers 79: 1052-1056.
Kupfahl C, Geginat G, Hof H (2006) Lentinan has a stimulatory effect on innate and adaptive immunity against murine Listeria
monocytogenes infection. International Immunopharmacology 6: 686- 696.
Lama L, Nicolaus B, Calandrell V, Manca M, Romano I, Gambacorta AM, Arambarri (1996) Effect of growth conditions on
endo- and exopolymer biosynthesis in Anabaena cylindrica 10C. Phytochemistry 42: 655-664.
Lee B, Bae J, Pyo H, Choe T, Kim S, Hwang H, Yun J (2004) Submerged culture conditions for the production of mycelial
biomass and exopolysaccharides by the edible basidiomycete Grifola frondosa. Enzyme and Microbial Technology 35:
369-376.
Lee K, Lee S, Lee H (1999) Bistage control of pH for improving exopolysaccharide production from mycelia of Ganoderma
lucidum in an air-lift fermentor. Journal of Bioscience and Bioengineering 88: 646-650.

178
Lee WY, Park Y, Ahn JK, Ka KH, Park SY (2007) Factors influencing the production of endopolysaccharide and
exopolysaccharide from Ganoderma applanatum. Enzyme and Microbial Technology 40 249-254.
Leterme P. (2010). Análisis de alimentos y forrajes, in Protocolos de LaboratorioUniversidad Nacional de Colombia Sede
Palmira: Palmira. 66 pp.
Lindequist U, Niedermeyer T, Julich W-D (2005) The pharmaceutical potential of mushrooms. Electronic Centralised Aircraf
Monitor (eCAM) 2: 285-299.
Liu F, Fung MC, Ooi VEC, Chang ST (1996) Induction in the mouse of gene expression of immunomodulating cytokines by
mushroom polysaccharide-protein complexes. Life Science 58(1): 795-803.
Maziero R, Cavazzoni V, Ramos-Bononi VL (1999) Screening of Basidiomycetes for the production of exopolisaccharides and
biomass in submerged culture. Revista de Microbiologia 30: 77-84.
Miller GL (1959) Use of DinitrosaIicyIic Acid Reagent for Determination of Reducing Sugar. Analytical Chemistry 31(3): 426-
428.
Mitchell D, Stuart D, Tanner R. (1999). Solid-State fermentation - Microbial growth kinetics, in The Encyclopedia of Bioprocess
Technology: Fermentation, Biocatalysis and bioseparation, Flickinger M, Drew S, EditorsWiley: New York. 2407-
2429
Mizuno T (1999) The extraction and development of antitumouractive polysaccharides from medicinal mushrooms in Japan.
International Journal Medicine Mushrooms 1: 9-29.
Mohammad-Fata M, Hossein M, Shirin G, Ghorban-Ali H (2007) Immunomodulating and anticancer agents in the realm of
macromycetes fungi (macrofungi). International Immunopharmacology 7: 701-724.
Moreira MT, Feijoo G, Lema JM (2003) Fungal bioreactors: Applications to white-rot fungi. Reviews in Environmental Science
and Bio/Technology 2: 247-259.
Nakamura K, Yamaguchi Y, Kagota S, Shinozuka K, Kunitomo M (1999) Activation of in vivo Kupffer cell function by oral
administration of Cordyceps sinensis in rats. . Japan Journal Pharmacology 79: 508-508.
Ooi V, Liu F (2000) Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Current Medicine
Chemistry 7: 715-728.
Pal M, Calvo A, Terron MC, Gonzalez AE (1995) Solid-state fermentation of sugarcane bagasse with Flammulina velutipes and
Trametes versicolor. World Journal Microbiology and Biotechnology 11: 541-545.
Pandey A, Soccol CR, Mitchell D (2000) New developments in solid state fermentation: I-bioprocesses and products. Process
Biochemistry 35: 1153–1169.
Park J-P, Kim S-W, Hwang H-J, Cho Y-J, Yun J-W (2002a) Stimulatory effect of plant oils and fatty acids on the exo-biopolymer
production in Cordyceps militaris. Enzyme and Microbial Technology 31: 250-255.
Park JP, Kim SW, Hwang HJ, Cho YJ, Yun JW (2002b) Stimulatory effect of plant oils and fatty acids on the exo-biopolymer
production in Cordyceps militaris. Enzyme and Microbial Technology 31: 250-255.
Plassard C, Mousain D, Salsac L (1982) Estimation of mycelial growth of basidiomycetes by means of chitin determination.
Phytochemistry 21: 345-348.
Pradeep V, Datta M (2002) Production of ligninolytic enzymes for decolorization by cocultivation of white-rot fungi Pleurotus
ostreatus and Phanerochaete chrysosporium under solid-state fermentation. Applied Biochemistry Biotechnology 102-
103: 109-118.
Sangsurasak P, Nopharatana M, Mitchell D (1996) Mathematical modeling of the growth of filamentous fungi in solid-state
fermentation. Journal Science Industrial Resources 55: 333-342.
Sarikaya A, Ladisch M (1997) An Unstructured Mathematical Model for Growth of Pleurotus ostreatus on Lignocellulosic
Material in Solid-State Fermentation Systems. Applied Biochemistry and Biotechnology 62: 72-85.
Shu C, Lung M (2003) Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia
camphorata in batch cultures
Process Biochemestry 39: 931-937.
Tang Y-J, Zhu L-W, Li H-M, Li D-S (2007) Submerged Culture of Mushrooms in Bioreactors – Challenges, Current State-of-the-
Art, and Future Prospects. Food Technology and Biotechnology 45(3): 221-229.
Tang YJ, Zhong JJ (2002) Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma
lucidum, grown on lactose in a bioreactor. Biotechnology Letters 24: 1023-1026.
Tsukagoshi S, Hashimoto Y, Fujii G, Kobayashi H, Nomoto K, Orita K (1984) Krestin (PSK). Cancer Treatment Review 11: 131-
155.
Viccini G, Mitchell D, Boit S, Gern J, da Rosa A, Costa R (2001) Analysis of growth kinetic profiles in solid-state fermentation.
Food Technology and Biotechnology Advances 39: 271-294.
Wade LG (1993) Química Orgánica. Debra Wechsler. México. 1312 p.
Wagner R, Mitchell DA, Sassaki GL, Lopes de Almeida A, Berovic M (2003) Current techniques for the cultivation of
Ganoderma lucidum for the production of biomass, ganoderic acid and polysaccharides. Food Technology and
Biotechnology 41: 371-382.
Walkley A, Black I (1934) An estimation of the Degtjareft method for determining of soil organic matter and a proposed
modification of chromic acid titration method. Soil Science 37: 29-38.
Wu J, Cheung P, Wong K, Huang N (2004) Studies on submerged fermentation of (Fr.) Singer. Part 2. Effect of carbonto-nitrogen
ration of the culture medium on the content and composition of the mycelial dietary fibre. Food Chemistry 85: 101-105.

179
Wu J, Ding Z-Y, Zhang K-C (2006) Improvement of exopolysaccharide production by macro-fungus Auricularia auricula in
submerged culture. Enzyme and Microbial Technology 39: 743-749.
Xiao J-H, Xiao D-M, Xiong Q, Liang Z-Q, Zhong JJ (2010) Nutritional requirements for the hyperproduction of bioactive
exopolysaccharides by submerged fermentation of the edible medicinal fungus Cordyceps taii. Biochemical
Engineering Journal 49(2): 241-249.
Xiao JH, Chen DX, Wan WH, Hu XJ, Qi Y, Liang ZQ (2006) Enhanced simultaneous the production of mycelia and intracellular
polysaccharide in submerged cultivation of Cordyceps jiangxiensis using desirability functions. Process Biochemestry
41: 1887-1893.
Yang F-C, Ke Y-F, Kuo S-S (2000) Effect of fatty acids on the mycelial growth and polysaccharide formation by Ganoderma
lucidum in shake flask cultures. Enzyme and Microbial Technology 27: 295-301.
Zhong JJ, Tang YJ (2004) Submerged cultivation of medicinal mushrooms for production of valuable bioactive metabolites.
Advances in Biochemical Engineering/Biotechnology 87: 25-59.
Zhuang O, Mizuno T (1999) Biological responses from Grifola frondosa (Dick.:Fr.) S.F. Gray-Maitake
(Aphyllophoromycetideae). Internation Journal Medicinal Mushrooms 1: 317-324.

180
9. Biorreactor para la Obtención de Sustancias
Bioactivas por Fermentación en Estado Sólido
Empleando Hongos Macromicetos (Solicitud de
Patente)
Durante el desarrollo de la presente investigación se realizó el diseño, construcción y puesta en
marcha de un biorreactor de lecho fijo con convección natural y tiro forzado con el fin de realizar las
pruebas piloto con las mejores combinaciones “especie de hongo – formulación” que arrojara el dieño
experimental. Sin embargo, una vez el biorreactor fue , se consideró que éste podía ser sometido a proceso
de solicitud de patente en Colombia y otros países. Este proceso de solicitud de patente en Colombia se
inició con las pruebas preliminares realizadas al biorreactor, las cuales se discuten en el siguiente párrafo.
La patente solicitada ante la Superintendencia de Industria y Comercio tiene la siguienjte denominación:
“BIORREACTOR PARA LA OBTENCIÓN DE SUSTANCIAS BIOACTIVAS POR
FERMENTACIÓN EN ESTADO SÓLIDO EMPLEANDO HONGOS MACROMICETOS” (ver
Anexos 1, 2 y 3). Es importante aclarar que toda la información consignada en esta sección es confidencial
debido a que, además de haber realizado la solicitud de patente ante la superintendencia mencionada, se
está realizando la exploración y análisis de ampliación de esta solicitud en otros países. Adicionalmente,
se está evaluando la solicitud de nuevas patentes en otros terriotrios donde se tenga conocimiento del
potencial de uso y comercialización del desarrollo adquirido con los nuevos datos obtenidos de la presente
investigación para la obtención de sustancias empleando hongos macromicetos en procesos de
fermentación en fase sólida. Por lo anterior, los datos experimentales obtenidos sobre el biorreactor de
lecho fijo con convección natural y tiro forzado de las mejores combinaciones especie-formulación
arrojadas por el diseño experimental realizado en este trabajo no son consignados aquí; estos datos
experimentales hacen parte de la confidencialidad que se requiere cuando se está en la búsqueda de
patentes de equipos, proceso de producción de sustancias y productos.
Como fase experimental inicial y con fines de prueba y puesta en marcha del biorreactor de lecho
fijo, se realizaron pruebas con varias especies de hongos de pudrición blanca. De estas pruebas, se
obtuvieron varias características particulares del procesos de fermentación en fase sólida en el biorreactor
objeto del patentamiento como el incremento de la velocidad de colonización y producción de exudados
de los hongos basidiomicetos de pudrición blanca. A continuación se relacionan dos de los procesos
probados en el biorreactor con sendos hongos de pudrición blanca:
i) Ganoderma lucidum aumentó en un 68% su velocidad de crecimiento sobre sustrato a base de mezclas
de materiales lignocelulósicos, como aserrín de madera, cascarillas de café, cacao y arroz,
suplementados con salvado de maíz, arroz o trigo, carbonato de calcio y sulfato de calcio. La velocidad
de colonización se incrementó de 0,67 kg de sustrato colonizado/dia a 2,14 kg de sustrato
colonizado/dia. La producción de exudado como extracto crudo de enzimas lignocelulolíticas y
polisacáridos fúngicos se incrementó de 10% (p/p) a 15% (p/p), expresada como masa de exudado
producido por masa de sustrato húmedo al momento de la inoculación. La determinación de los
incrementos en velocidad de crecimiento del hongo y producción de exudado fue referida a la
inoculación del hongo en bolsas de tres kg de capacidad, siguiendo el mismo tratamiento al sustrato
que cuando se trabajó en el biorreactor.
ii) Coriolus versicolor aumentó en un 72% su velocidad de crecimeiento sobre un sustrato formulado con
base en los mismos materiales lignocelulósicos enunciados para el hongo G. lucidum. La velocidad de
colonización se incrementó de 0,75 kg de sustrato colonizado/dia a 2,0 kg de sustrato colonizado/dia.

181
La producción de exudado como extracto crudo de enzimas lignocelulolíticas y polisacáridos fúngicos
se incrementó de 7% (p/p) a 16,7% (p/p), como masa de exudado producido por masa de sustrato
húmedo al momento de la inoculación. La determinación de los incrementos en velocidad de
crecimiento del hongo y la producción de exudado fue referida a la inoculación del hongo en bolsas de
tres kg de capacidad, siguiendo el mismo tratamiento al sustrato que cuando se trabajó en el
biorreactor.
Se desarrolló el proceso de producción de las biosustancias (enzimas lignocelulolíticas y
polisacáridos fúngicos) por fermentación en fase sólida con hongos macromicetos de pudrición blanca y
utilizando residuos lignocelulósicos como materias primas. Este proceso se resume en la Figura 1 en la
que se especifican los puntos donde es necesario controlar cada una de las variables físico-químicas de los
sustratos, las condiciones medioambientales y la condiciones del biorreactor, las cuales dependen
directamente de la especie del hongo macromiceto seleccionado.
Las biosustancias obtenidas en el biorreactor de lecho fijo con convección natural y tiro forzado
fueron evaluadas de los exudados y del material sólido colonizado tanto para los sustratos inoculados con
Ganoderma lucidum, como para Coriolus versicolor. Los resultados promedio obtenidos para ambas
especies de hongos se muestran en la Tabla 1. Con la información anterior fue solicitada la patente de
invención para el biorreactor, el proceso y los productos obtenidos.
Tabla 1. Actividades enzimáticas y cantidad de polisacáridos y NAGA obtenidos en los sustratos sólidos colonizados
con G. lucidum y C. versicolor y sus exudados producidos en Biorreactor de lecho fijo
Bioproducto Sustrato colonizado Exudado
Endoglucanasa 8 - 86 U/gss 6 - 18 U/mL
Exoglucanasa 8 – 74 U/gss 4,5 – 36
Endoxilanasa 12 – 25 U/gss 12 – 27 U/mL
β-glucosidasa 2 – 9 u/gss 2.1 – 60 U/mL
Lacasa 12 – 97 U/gss 15 – 36 U/mL
Manganeso peroxidasa 25 – 65 U/gss 4,5 – 20 U/mL
Lignina peroxidasa*
9-18 U/gss 1 – 2 U/mL
Polisacáridos como
carbohidratos totales
0,03 - 0,13 g/gss 0,2 - 0,8 g/L
N-acetil-D-
glucosamina (NAGA)
0,04 - 0,13 mg/gss Nd
*Positiva unicamente para el hongo C. versicolor

182
Figura 1. Diagrama de proceso para la obtención de sustancias bioactivas por fermentación en fase sólida
en biorreactor bajo condiciones controladas, empleando hongos macromicetos
Selección de materia prima
Adecuación física de
materia prima (tp)
Formulación de sustratos
(%H, R C/N)
Esterilización de sustratos
(T°, P, tiempo)
Inoculación de sustratos
(% inóculo)
Incubación: Inicio procesos de fermentación
en fase sólida en biorreactor
(T°, HR, CO2, O2, Intensidad de luz, mezclado)
Producción de biosustancias:
(Enzimas, polisacáridos, oligosacáridos, etc.)
Materiales lignocelulóscios
(Residuos agroindustriales)
Insumos
(sales minerales,
suplementos nutricionales)
Semilla de
macromicetos
Producción de carpóforos
Subproducto con valor: Sustrato agotado

183
10. Conclusiones Generales
10.1. Contribuciones de la Tesis
En esta sección se relacionan las conclusiones del trabajo de tesis doctoral en cumplimento con el
planteamiento hecho en los objetivos específicos. El planteamiento general de la tesis correspondió a un
rastreo de cepas de entre diez especies preseleccionadas a fin de buscar las de mejor rendimiento en la
producción de celulasas, ligninasas y polisacáridos. Posteriormente se desarrolló un diseño experimental
bifactorial para fermentación sumergida y otro para fermentación en estado sólido con el fin de obtener la
mejor combinación especie-formulación en la producción de enzimas lignocelulolíticas y polisacáridos,
para lo cual se variaron 12 niveles de la relación C/N y tres niveles del bioagente correspondientes a las
tres especies de hongos con mejor comporatmiento en la etapa de rastreo. Finalmente, se llevó a cabo el
diseño, montaje y puesta en marcha de un biorreactor de lecho fijo a nivel piloto para la fermentación en
estado sólido con los macromicetos estudiados.
En este trabajo se realizaron pruebas de rastreo cualitativas para las enzimas celulolíticas
endoglucanasa y exoglucanasa, y para las enzimas ligninolíticas lacasa y lignina peroxidasa, a fin de
seleccionar de entre diez especies de hongos de pudrición blanca (Lentinula edodes, Schizophyllum
commune, Trametes trogii, Coriolus versicolor, Pycnoporus sanguineus, Ganoderma applanatum,
Ganoderma lucidum, Grifola frondosa, Pleurotus ostreatus y Auricularia delicata) las cepas de mejor
desempeño en cuanto a las actividades enzimáticas mencionadas (ver Capítulo 3). Se concluyó que las
especies de hongos con mejor producción de celulasas y ligninasas fueron las cepas de Lentinula edodes y
Coriolus versicolor. Asimismo, se realizó un rastreo cuantitativo a las mismas diez especies de hongos de
pudrición blanca determinando su contenido de exopolisacáridos e intrapolisacáridos, expresados como
carbohidratos totales (ver Capítulo 4). De ambos rastreos se obtuvo que las especies Lentinula edodes,
Coriolus versicolor y Pleurotus ostreatus fueron las cepas de mejor comportamiento respecto a la
producción de celulasas, ligninasas y exopolisacáridos, mientras que para la producción de
intrapolisacáridos de los cuerpos fructíferos de los hongos, se seleccionó el hongo Grifola frondosa.
Se realizaron procesos de fermentación en estado sólido para la obtención de enzimas
lignocelulolíticas de hongos de pudrición blanca empleando como materias primas para la elaboración de
los sustratos mezclas de residuos lignocelulósicos. Se determinaron las actividades de las enzimas
celulolíticas (endoglucanasa, exoglucanasa, endoxilanasa y β-glucosidasa) y de las enzimas ligninolíticas
(lacasa y manganeso peroxidasa). Asimismo, fueron determinadas la producción de biomasa, la
producción y consumo de azúcares reductores, consumo de celulosa y hemicelulosa y la degradación de la
lignina en 15 tiempos durante 49 días de fermentación. Este proceso fue evaluado en el marco de un
diseño experimental las siguientes tres especies de hongos: Lentinula edodes, Coriolus versicolor y
Pleurotus ostreatus (Capítulo 5). De esta evaluación se concluye que las actividades enzimáticas
obtenidas presentaron descensos a diferentes tiempos del proceso de fermentación en estado sólido, hecho
que pudo atribuirse a aspectos como la eficiencia del método de extracción empleado; a los cambios en la
composición del sustrato durante el tiempo de fermentación, ya que pueden producirse nuevas sustancias
que inhiban las actividades enzimáticas; a la represión catabólica por productos de las reacciones que se
generan en la fermentación; a la producción de variantes isoenzimáticas de las mismas enzimas a
diferentes tiempos; y a las diferentes condiciones de utilización del sustrato que arrojan actividades
diferentes.
Con este trabajo se hace un aporte al estudio de las cinéticas de formación de biomasa,
producción de enzimas, consumo de nutrientes y degradación de la lignina que permiten dar un paso
adelante en la descripción del proceso global y posterior escalado para la obtención mediante
fermentación en estado sólido de los productos de interés estudiados (Capítulo 5). Uno de los
inconvenientes más frecuentes en el planteamiento de las ecuaciones para la descripción de la cinética de
cada una de las variables mencionadas en procesos de fermentación en estado sólido consiste en conferirle
a cada expresión matemática algún sentido biológico respecto al metabolito a describir. Las expresiones

184
propuestas describieron aceptablemente cada una de las variables medidas según se desprende del análisis
estadístico realizado; cabe destacar a este respecto que durante los procesos de fermentación en fase
sólida, cuando se utilizan como sustratos mezclas de materiales lignocelulósicos, puede producirse una
gran cantidad de sustancias intermedias producto de las reacciones bioquímicas, que en la mayoría de los
casos no se pueden valorar, lo cual dificulta el modelamiento matemático.
La producción de polisacáridos fúngicos de la fase vegetativa de los macromicetos en procesos de
fermentación en fase sólida, en la que además de producir polisacáridos también ocurre la degradación de
materiales lignocelulósicos, es uno de los aportes que respresentan novedad en este trabajo. Lo anterior,
considerando que en el contexto general de los estudios sobre producción de polisacáridos por
fermentación en estado sólido, se enfatiza la producción de los cuerpos fructíferos de los hongos como
fuente de polisacáridos y no el uso del micelio desarrollado dentro del sustrato como fuente de los mismos
(Capítulo 6). Aunque se requiere una mayor información sobre las cinéticas de formación de biomasa, de
producción de polisacáridos, de consumo de nutrientes y de degradación de lignina, este trabajo hace un
aporte a esta temática con el planteamiento y validación de las expresiones matemáticas que describen la
cinética de cada una de las variables medidas. De esta manera, los resultados obtenidos tienen el potencial
de contribuir al mejor entendimiento del proceso global de cultivo de los hongos con miras al posterior
escalado a nivel comercial de la obtención de alguno de los productos de interés que se plantean en este
estudio.
Se presenta un avance en referencia a la obtención de enzimas lignocelulíticas por fermentación
sumergida empleando hongos macromicetos, especialmente en cuanto a las cinéticas de formación de
biomasa, de producción de enzimas, del consumo de nutrientes y de degradación de la lignina se refiere
(Capítulo 7). Los modelos matemáticos planteados para la descripción del proceso global de obtención de
enzimas lignocelulolíticas empleando como materias primas residuos lignocelulósicos pretratados,
representan una valiosa herramienta que contribuirá a cualificar nuestro entendimiento del cultivo
sumergido de los hongos de pudrición blanca y de su actividad enzimática, lo cual a su vez permitirá
establecer estrategias que disminuyan los costos de producción de estas biosustancias. Es evidente que se
requiere mayor información sobre la dinámica de la biosíntesis y consumo de estos metabolitos, pero el
planteamiento y validación de las ecuaciones cinéticas de cada una de las variables medidas permitieron la
descripción de cada una de los compuestos determinados en el presente estudio. A pesar de que algunas de
las ecuaciones describieron de una manera más adecuada los datos experimentales que otras, el hecho de
disponer de nuevos modelos cinéticos validados, se convierte en una oportunidad para continuar
desarrollando trabajos que profundicen en este tópico. En particular, la descripción matemática de algunos
de los productos intermedios originados por la acción de las enzimas estudiadas, o de otras enzimas no
evaluadas, así como de sus productos, contribuirá a una interpretación más acabada de los complejos
fenómenos que ocurren durante el crecimiento de los macromicetos en los procesos de fermentación
sumergida.
La producción de polisacáridos fúngicos por fermentación sumergida empleando residuos
lignocelulósicos como parte de los medios de cultivo es un proceso no muy estudiado (Capítulo 8), a pesar
de las valiosas actividades biológicas que exhiben estos metabolitos cuando son obtenidos a partir de
hongos de pudrición blanca (ver Capítulo 2). En este trabajo se plantearon ecuaciones para la descripción
de la cinética de formación de biomasa, de producción de exopolisacáridos y de consumo de nutrientes
bajo la premisa de conferirle a cada expresión matemática algún sentido biológico respecto al compuesto
cuya dinámica se desea describir. Las expresiones propuestas representaron aceptablemente cada una de
las variables medidas; la dificultad del modelamiento matemático de este proceso radica en la complejidad
de la determinación experimental de la gran cantidad de sustancias intermedias que participan en las vías
metabólicas de síntesis de los exopolisacáridos cuando se utilizan en los medios de cultivo en fase líquida
materiales lignocelulósicos, así éstos sean pretratados. Es de anotar que la descripción del proceso global
de obtención de los productos de interés estudiados, requiere de planteamientos matemáticos complejos
que permitan describir eficientemente estos procesos metabólicos de tal manera que los resultados
obtenidos sean funcionales y útiles para el escalamiento de la producción de polisacáridos fúngicos
bioactivos a nivel industrial con bajos costos de producción.

185
El presente trabajo doctoral contempló el cultivo a nivel piloto de hongos macromicetos
productores de enzimas lignocelulolíticas y polisacáridos empleando biomasa lignocelulósica (residuos
agroindustriales) como materia prima. Se desarrolló el cultivo de hongos macromicetos en un biorreactor
de lecho fijo con convección natural y tiro forzado bajo condiciones controladas. El biorreactor fue
diseñado, construido y puesto en marcha como resultado de esta tesis (Capítulo 9). Este biorreactor fue
puesto en operación con las especies de hongos Ganoderma lucidum (especie incluida en las diez cepas
preseleccionadas para los rastreos iniciales) y Coriolus versicolor sobre diferentes mezclas de residuos
lignocelulósicos. Posteriormente, se consideró que este equipo podía ser sometido a proceso de solicitud
de patente en Colombia y otros países. La solicitud de patente en Colombia se inició con las pruebas
preliminares realizadas en el biorreactor empleando las dos especies de hongos mencionadas. La solicitud
de patente denominada “Bioreactor para la obtención de sustancias bioactivas por fermentación en
estado sólido empleando hongos macromicetos” se realizó ante la Superintendencia de Industria y
Comercio de Colombia (ver Anexos XXX). Actualmente se está realizando la exploración y análisis de
ampliación de solicitud de esta patente en otros países, así como la solicitud de nuevas patentes en otros
terriotrios donde se tenga conocimiento del potencial de uso y comercialización tanto del equipo diseñado
como del proceso desarrollado en la presente investigación.
10.2. Impactos Potenciales de la Tesis
El aprovechamiento y tratamiento de los diferentes tipos de residuos generados durante las
actividades agrícolas y agropindustriales en Colombia y el mundo son un reto para la humanidad, ya que
éstos se han constituido en uno de los problemas más álgidos (y a la vez, en una de las oportunidades más
claras) para las sociedades presentes y futuras. En este trabajo se desarrollaron procesos de obtención de
enzimas lignocelulolíticas y polisacáridos empleando como materias primas residuos lignocelulósicos con
hongos de pudrición blanca. En los procesos desarrollados se realizó la descripción de las cinéticas de
crecimiento de la biomasa, la producción y consumo de los azúcares reductores, el consumo de celulosa y
hemicelulosa y la degradación de lignina en procesos de fermentación sumergida y en estado sólido. Con
este trabajo se generan múltiples posibilidades para el uso adecuado de residuos agrícolas y
agroindustriales con miras a la producción de productos de uso industrial en el segmento agrícola,
agroindustrial y farmacológico.
El sector agrícola colombiano requiere del desarrollo de nuevos productos que mejoren la calidad
de la alimentación de animales de cría, como los bovinos, y que además estos alimentos sirvan como
coadyuvantes en el incremento de la salud del animal. Los residuos lignocelulósicos se pueden utilizar
potencialmente como alimento para rumiantes, pero con problemas de digestibilidad además de su bajo
contenido proteico. Los sustratos basados en materiales lignocelulósicos colonizados por estos hongos de
pudrición blanca, resultan ser una alternativa promisoria para este sector ya que el sustrato experimenta un
aumento de la digestibilidad mediada por la acción enzimática del hongo y un incremento del contenido
de proteína con la colonización fúngica al final de la fermentación. Adicionalmente, la ocurrencia de los
polisacáridos fúngicos contribuye a mejorar las características de estos residuos lignocelulósicos para
satisfacer las necesidades de alimentación bovina y el incremento de la salud animal. De hecho el uso de
sustratos agotados de la producción de hongos macromicetos como Pleurotus sp., L. edodes, Ganoderma
sp., Agaricus bisporus y otros, así como el empleo de hongos de pudrición blanca para deslignificar los
materiales lignocelulósicos, son una alternativa para la producción de alimento de rumiantes. Lo anterior,
debido a que durante la fermentación en fase sólida con estos basidiomicetos, los materiales
lignocelulósicos exhiben una bioconversión favorable en términos de disminución de lignina y producción
de azúcares reductores que luego son utilizados por las bacterias ruminales que, finalmente, suministran
los nutrientes requeridos por el animal para su desarrollo.
Con los resultados que se iban obteniendo durante la ejecución de esta tesis doctoral, se
complementó el conocimiento y la experiencia necesarios para la participación de la autora de esta tesis
(en conjunto con el director de la misma y otro colega) en la Convocatoria Colciencias 523-2011 para la
conformación de un banco de proyectos de creación de empresas o unidades de negocio de base

186
tecnológica. El grupo proponente participó con el proyecto Desarrollo y evaluación de los parámetros de
cultivo a escala semi industrial del hongo Grifola frondosa en climas tropicales y su transformación con
fines comerciales como productos nutracéutico. Este proyecto fue aprobado para ser desarrollado en dos
etapas, de la cual actualmente se está desarrollando la primera que corresponde especialmente al estudio
de mercado de los productos nutracéuticos obtenidos a partir del hongo Grifola frondosa (maitake). La
formulación de este proyecto partió de una investigación terminada en la Universidad de Caldas sobre el
desarrollo del cultivo del hongo Grifola frondosa en climas tropicales a escala de laboratorio cuyops
resultados fueron complementados con los hallazgos encontrados en esta tesis. Entre estos hallazgos se
cuenta la exploración y refinación de los métodos de extracción de las diferentes fracciones de
polisacáridos y otros carbohidratos de interés farmacológico a partir de los cuerpos fructíferos del hongo
G. frondosa. En el Anexo 4 se muestra el esquema de extracción y la composición química global de las
fracciones obtenidas hasta el momento, así como un perfil general del balance de masa del cuerpo
fructífero del hongo. La purificación de las fracciones no se alcanzó a realizar, pero se definieron los
métodos para su desarrollo, los cuales consisten en una serie de diálisis abiertas y cerradas con diferentes
tamaños de membranas para eliminar el exceso de hidróxido de potasio y separar cada una las fracciones
obtenidas de oligosacáridos y monosacáridos. Asimismo, se definieron los métodos de determinación de
glucanos por HPLC con detector de fluorescencia y el perfil de carbohidratos por HPLC con detector de
índice de refracción.
10.3. Trabajos Futuros
Actualmente están en desarrollo tres trabajos de grado a nivel de maestría derivados de esta tesis
doctoral, los cuales se relacionan a continuación:
Trabajo de grado de Maestría en Ingeniería de alimentos (Universidad de Caldas): Estudio cinético
para la producción de polisacáridos de Ganoderma applanatum.
Trabajo de grado de Maestría en Sistemas de Producción Agropecuaria (Universidad de Caldas):
Aplicación de cocteles enzimáticos producidos con el macromiceto Coriolus versicolor como
coadyuvantes en los procesos de compostaje y ensilaje.
Trabajo de grado de Maestría en Microbiología Agroindustrial (Universidad Católica de Manizales):
Evaluación del efecto de las ligninasas (lacasa y manganeso peroxidasa) en la fructificación y
eficiencia biológica del hongo Ganoderma lucidum.
Adicionalmente y como aplicación directa de los resultados de la presente tesis, se está
participando en la Convocatoria del Fondo de Ciencia, Tecnología e Innovación del Sistema Nacional de
Regalías Vigencia 2012 con el proyecto Diseño e implementación de una estrategia integral para el
tratamiento y aprovechamiento de residuos en el sector agrícola y agroindustrial de Caldas mediante
innovación en procesos biotecnológicos y la formación del capital humano pertinente para la generación
de valor en el sector agrícola y agroindustrial de Caldas. En este proyecto se propone el desarrollo de dos
paquetes tecnológicos derivados de la tesis doctoral para la producción de cocteles enzimáticos con
actividad lignocelulolítica y de polisacáridos por fermentación sumergida y en estado sólido, así como el
inicio de la implementación de la cadena productiva de hongos macromicetos en el Departamento de
Caldas. A la fecha, este proyecto ha pasado todos los filtros respectivos a nivel departamental (Comisión
de Ciencia y Tecnología –CODECTI– de Caldas, Gobernación de Caldas) y la revisión nacional por parte
de la Secretaría Técnica de Colciencias en donde fue declarado viable; el proyecto espera su posible
aprobación por parte del Órgano Colegiado de Decisión –OCAD– en las próximas semanas. Todos estos
desarrollos se realizarían en la Planta de Bioprocesos de la Universidad de Caldas.

187
Se contempla el inicio de las siguientes propuestas de investigaciones derivadas de la tesis
doctoral a corto y mediano plazo en las siguientes temáticas:
Síntesis de enzimas lignocelulolíticas por fermentación en estado sólido empleando mezclas de
residuos lignocelulósicos en donde se determine una mayor cantidad de sustancias intermedias
producto del metabolismo de hongos de pudrición blanca a fin de lograr dilucidar las técnicas de
ataque que utilizan estos hongos para degradar la matriz lignocelulósica.
Producción de polisacáridos por fermentación en estado sólido en fase vegetativa en biorreactores bajo
condiciones controladas en la que se diferencien y caractericen las fracciones de intrapolisacáridos y
los exopolisacáridos en los exudados del cultivo fúngico.
Evaluación del efecto de la viscosidad en los procesos de fermentación sumergida para la producción
de enzimas lignocelulolíticas y polisacáridos empleando hongos macromicetos en biorreactor de
tanque agitado.
Caracterización química y evaluación de la actividad biológica de las fracciones obtenidas del proceso
de extracción de polisacáridos de los cuerpos fructíferos de los macromicetos.
Evaluación de los hongos de pudrición blanca utilizados en este estudio como agentes biológicos
degradadores de residuos recalcitrantes, por ejemplo, en las aguas provenientes de los lixiviados de
rellenos sanitarios, y en aguas contaminadas con compuestos organoclorados y organofosforados. Se
plantea la utilización como inóculo de los sustratos colonizados por los hongos en los biorreactores en
estado sólido luego de haber producido enzimas o polisacáridos.
Proponer nuevas pruebas de los modelos matemáticos planteados en esta tesis para la descripción de
los procesos de fermentación sumergida y en estado sólido utilizando formulaciones de sustratos
diferentes, así como otras especies de hongos a fin de examinar la universalidad de los modelos
matemáticos propuestos.

188
Adachi Y., Ohno N., Ohsawa M., Oikawa S., Yadomae T. (1990). Change of biological activities of (1-3)-B-D glucan from
Grifola frondosa upon molecular-weight reduction by heat-treatment. Chemical and Pharmaceutical Bulletin, 38: 477-481.
Adler E. (1977). Lignin chemistry. Past, present and future. Wood Science Technology, 11: 169-218.
Aguilar C., Augur C., Favela-Torres E., Viniegra-Gonzalez G. (2001). Production of tannase by Aspergillus niger Aa-20 in
submerged and solid-state fermentation: influence of glucose and tannic acid. Journal of Industrial Microbiology and
Biotechnology Advances, 26: 296-302.
Ali H. (2010). Biodegradation of Synthetic Dyes-A Review. Water Air Soil Pollut, 213: 251-273.
Alkorta I., Garbisu G., Llama M.J., Serra J.L. (1998). Industrial applications of pectic enzymes: a review. Process Biochemistry,
33: 21-28.
Allen D.G., Robinson C.W. (1989). Hydrodynamics and mass transfer in Aspergillus niger fermentations in buble column and
loop bioreactors. Biotechnology and Bioengineering 34: 731-740.
Alvira P., Tomás-Pejó E., Ballesteros M., Negro M.J. (2009). Pretreatment technologies for an efficient bioethanol production
process based on enzymatic hydrolysis: A review. Bioresource Technology, doi: 10.1016/j.biortech.2009.11.093.
Amaral A., Carbonero E., Simao R., Kadowaki M., Sassaki G., Osaku C., Gorin P., Lacomini M. (2008). An unusual water-
soluble B-glucan from the basidiocarp of the fungus Ganoderma resinaceum. Carbohydrate Polymers, 72: 473-478.
Amat-García E.C., Amat-García G.D., Henao L.G. (2001). Diversidad taxonómica y ecológica de la entomofauna micófaga en un
bosque alto-andino de la cordillera oriental de Colombia. Revista Academia Colombiana de Ciencia 1-21.
Ander P. (1994). The celloiose-oxidizing enzymes CBQ and CBO as related to lignin and cellulose degradation - a review. FEMS
Microbiology Reviews, 13: 297-312.
Ângelo A.S. (2004). Enzimas hidrolíticas. En: Fungos: Uma Introducâo à biologia, bioquimica e biotecnologia. Esposito,
Azevedo J.I. (Eds.). Universidade de Caxias do Sul,. pp. 263-285.
Arantes V., Milagres A.M.F. (2007a). The effect of a catecholate chelator as a redox agent in Fenton-based reactions on
degradation of lignin-model substrates and on COD removal from effluent of an ECF kraft pulp mill. Journal of Hazardous
Materials, 141: 273-279.
Arantes V., Milagres A.M.F. (2007b). The synergistic action of ligninolytic enzymes (MnP and Laccase) and Fe3+-reducing
activity from white-rot fungi for degradation of Azure B. Enzyme and Microbial Technology, 42: 17-22.
Archibald F. (1992a). A new assay for lignin-type peroxidases employing the dye Azure B. Applied Environmental Microbiology,
58: 3110-3116
Archibald F. (1992b). A new assay for lignin-type peroxidases employing the dye Azure B. Applied Environmental Microbiology,
58: 3110-3116.
Arias E.L., Piñeros P.A. (2008). Enzimas ligninolíticas fúngicas para fines ambientales.Trabajo de grado. Facultad de Ciencias,
Pontificia Universidad Javeriana: Bogotá. 202 p. p.
Arora D.S., Chander M., Gill P.K. (2002). Involvement of lignin peroxidase, manganese peroxidase and laccase in degradation
and selective ligninolysis of wheat straw. International Biodeterioration & Biodegradation, 50: 115-120.
Arora D.S., Sharma R.K. (2010). Ligninolytic fungal laccases and their biotechnological applications. Applied Biochemistry and
Biotechnology, 160: 1760-1788.
Arteaga-Hoyos A.B. (2002). Utilización de los desechos del banano para el cultivo del hongo comestible Pleurotus ostreatus.
Augura Carta Informativa, (212): 4-6.
Asatiani M.D., Kachlishvili E.T., Khardziani T.S., Metreveli E.M., Mikiashvili N.A., Songulashvili G.G., Tsiklauri N.D., Wasser
S.P., Elisashvili V.I. (2008). Basidiomycetes as a source of antioxidants, lectins, polysaccharides, and enzymes. Journal of
Biotechnology 136S: S717-S742.
Badcock E. (1943). Methods for obtaining fructifications of wood-rotting fungi in culture. Transactions of the British Mycological
Society, 26: 127-132.
Bae J., Sim J., Lee B., Pyo H., Choe T., Yun J. (2005). Production of exopolysaccharide from mycelial culture of Grifola
frondosa and its inhibitory effect on matrix metalloproteinase-1 expression in UV-irradiated human dermal fibroblasts.
FEMS Microbiology Letters, 251: 347-354.
Bajpai P. (1997). Microbial xylanolytic enzyme system: properties and applications. Advances in Applied Microbiology, 43: 141-
194.
Baker R.A., Wicker L. (1996). Current and potential applications of enzyme infusion in the food industry. Trends Food Science
Technology, 7: 279-284.
Baldrian P., Gabriel J. (2002). Variability of laccase activity in the whiterot basidiomycete Pleurotus ostreatus. Folia
Microbiology, 47: 385-390.
Baldrian P., Valasková V. (2008). Degradation of cellulose by basidiomycetous fungi. FEMS Microbiology Review, 32: 501-521.
Bao W., Fukushima Y., Jensen K., Moen M., Hammel K. (1994). Oxidative degradation of non-phenolic lignin during lipid
peroxidation by fungal manganese peroxidase. FEBS Letters, 354: 297-300.
Barr B.K., HsieH Y.L., Ganem B., Wilson D.B. (1996). Identification of two functionally different classes of exocellulases.
Biochemistry, 35: 586-592.
Béguin P., Aubert J.P. (1994). The biological degradation of cellulose. FEMS Microbiology Reviews, 13: 25-58.
Beily P., Kratky Z., Vrsanska M. (1981). Substrate binding site of endo-1,4- β-xylanase of the yeast Cryptococcus albidus.
Europe Journal Biochemestry, 119: 559 -571.

189
Benavidez-Calvache O.L. (2004). Estudio Químico de la fracción insaponificable del hongo macromiceto Lentinula edodes
(Shiitake).Maestría en Ciencias-Química. Programa de Postgrados en Química, Universidad Nacional de Colombia: Bogotá.
98 p.
Berovič M., Boh B., Wraber B., Pohleven F. (2007). Production of intra- and extracellular polysaccharides of Grifola frondosa by
solid state and submerged fermentation. International Journal for Medicinal Mushrooms, 9: 193-194.
Bizukojc M., Ledakowicz S. (2006). A kinetic model to predict biomass content for Aspergillus niger germinating spores in the
submerged culture. Process Biochemistry, 41: 1063-1071.
Blanch H.W., Clark D.S. (1997). Biochemical Engineering. Marcel Dekker: New York. 702 pp. p.
Blanchette R.A. (1995). Degradation of the lignocellulose complex in wood. Canadian Journal of Botany, 73: S999-S1010.
Blanchette R.A., Krueger E.W., Haight J.E., Akhtar M., Akin D.E. (1997). Cell wall alterations in loblolly pine wood decayed by
the white-rot fungus, Ceriporiopsis subvermispora. Journal of Biotechnology, 53 203-213.
Blánquez P., Casas N., Font X., Gabarrell M., Sarrá M., Caminal G. (2004). Mechanism of textile metal dye biotransformation by
Trametes versicolor. Water Resource, 38: 2166-2172.
Blánquez P., Sarrà M., Vicent M. (2006). Study of the cellular retention time and the partial biomass renovation in a fungal
decolourisation continuous process. Water Resource, 40: 1650-1656.
Blánquez P., Caminal G., Sarrà M., Vicent M. (2007). The effect of HRT on the decolourisation of the Grey Lanaset G textile dye
by Trametes versicolor. Chemistry Engineering Journal, 126: 163-169.
Boisset C., Fraschini C., Schülein M., Henriss B. (2000). Imaging the enzymatic digestion of bacterial cellulose ribbons reveals
the endo character of the cellobioydrolase Cel6A from Humicola insolens and its mode of synergy with cellobiohydrolse Cel
7A. Environmental Microbiology, 66: 1444-1452.
Bolla K., Gopinath B., Shaheen S., Charya M. (2010). Optimization of carbon and nitrogen sources of submerged culture process
for the production of mycelial biomass and exopolysaccharides by Trametes versicolor. International Journal for
Biotechnology and Molecular Biology Research, 1: 15-21.
Bommarius A.S., Riebel B.R. (2004a). Biocatalysis. Wiley-VCH Verlag GmbH & Co. KGaA: Atlanta, USA.
Bommarius A.S., Riebel B.R. (2004b). Biocatalysis. Wiley-VCH Verlag GmbH & Co. KGaA: Atlanta, USA. 611 p. p.
Bonugli-Santos R., Durrant L., Durães Sette L. (2012). The Production of Ligninolytic Enzymes by Marine-Derived
Basidiomycetes and Their Biotechnological Potential in the Biodegradation of Recalcitrant Pollutants and the Treatment of
Textile Effluents. Water Air Soil Pollut, 223: 2333-2345.
Borchers A., Keen C., Gerhwin M. (2004). Mushrooms, tumors, and immunity: an update
. Experiment in Biological Medical, 229: 393-406.
Bosell G.P., Jacobs H., Davidson F.A., G.M. G., Ritz K. (2003). Growth and Function of Fungal Mycelia in Heterogeneous
Environments. Bulletin of Mathematical Biology, 65: 447-477.
Brummer Y., Cui S.W. (2006). Detection and Determination of Polysaccharides in Foods. En: Food Polysaccharides and Their
Applications. Stephen A.M., Phillips G.O., Williams P.A. (Eds.). CRC Press Taylor & Francis Group: Boca Raton, FL. 676-
706 p. pp. 676-706.
Burgstaller W., Schinner F. (1993). Leaching of metals with fungi. Journal of Biotechnology, 27: 91-116.
Burnett J.H. (1976). Fundamental of Micology. 2a ed Edward Arnold: London. 221 p. p.
Burns J.K. (1991). The polygalacturonases and lyases. En: The Chemistry and Technology of Pectin. Walter R.H. (Ed.).
Academic Press,: San Diego. pp. 165- 188.
Buswell J.A., Odier E. (1987). Lignin biodegradation. Review Biotechnology, 6: 1-59.
Buswell J.A., Cai Y., Chang S.T. (1995). Effect of nutrient nitrogen and manganese on manganese peroxidase and laccase
production by Lentinula (Lentinus) edodes. FEMS Microbiology Letters, 128: 32- 45.
Buswell J.A., Cai Y.J., Chang S.T. (1996). Ligninolytic enzyme production and secretion in edible mushroom fungi. En:
Mushroom Biology and Mushroom Products. Royse D.J. (Ed.). The Pennsylvania State University pp. 113±122:
Pennsylvania.
Campbell M.K., Farrell S.O. (2004). Bioquímica. Cuarta ed International Thomson Editores: México. 725 p. p.
Carabajal M., Levin L., Abertó E., Lechner B. (2012). Effect of co-cultivation of two Pleurotus species on lignocellulolytic
enzyme production and mushroom fructification. International Biodeterioration & Biodegradation, 66: 71-76.
Cardona B.E., Saldarriaga Y., Guzmán L. (2005). Registro de Gimnopilus rugulosus (Agaricales, Cortinariaceae) de Colombia.
Revista Mexicana de Micología, 021: 55-58.
Cardona M., Osorio J., Quintero J. (2009). Degradación de colorantes industriales con hongos ligninolíticos. Revista Facultad de
Ingeniería de la Universidad de Antioquia, (48): 27-37.
Carle - Urioste J.C., Escobar - Vera J., El-Gogary S. (1997). Cellulase induction in Trichoderma reesei by cellulose requires its
own basal expression. Journal of Biological Chemistry, 272: 10169-10174.
Carlile M., Watkinson S., Gooday G. (2001). The fungi. Second ed Academic Press: London. 588 p. p.
Carreño N., Vargas A., Bernal A.J., Restrepo S. (2007). Problemas fitopatológicos en especies de la familia Solanaceae causados
por los géneros Phytophthora, Alternaria y Ralstonia en Colombia. Una revisión. Agronomía Colombiana, 25(2): 320-329.
Castillo M.d.P., Ander P., Stenstrom J. (1997). Lignin and manganese peroxidase activity in extracts from straw solid substrate
fermentation. Biotechnol. Technol. , 11 701-706.
Castro-Ríos K. (2006). Validación de deshidratación convencional para la conservación del hongo comestible Pleurotus sajor-
caju Revista Universidad de Caldas: 123-133.

190
Catley B.J. (1992). The biochemistry of some fungal polysaccharides with industrial potencial. En: Handbook of applied
mycology: fungal biotechnology. Mukerji K.G. (Ed.). Marcel Dekker,: New York. pp. 1091-1114.
Chandra M., Kalra A., Sharma P.K., Sangwan R.S. (2009). Cellulase production by six Trichoderma spp. fermented on medicinal
plant processings. Journal Industrial Microbiology Biotechnology, 36: 605-609.
Chandra R.P., Bura R., Mabee W.E., Berlin A., Pan X., Saddler J.N. (2007). Substrate pretreatment: the key to effective
enzymatic hydrolysis of lignocellulosics? Advance Biochemistry Engineering and Biotechnology, 108: 67-93.
Chang S.T., Quimio T.H. (1982). Tropical Mushrooms: Biological Nature and Cultivation Methods. ed. T.H. Q.E. Chinese
University Press: Hong Kong. 493 p.
Chang S.T., Miles P.G. (2004). Mushrooms Cultivation, Nutritional Value, Medicinal effect, and Enviromental Impact. Segunda
ed CRC Press: New York. 451 p.
Chang T.S., Siddiq M., Sinha N.K., Cash J.N. (1994). Plum juice quality affected by enzyme treatment and fining. Journal of
Food Science, 59: 1065-1069.
Chaparro D.F., Rosas D.C. (2006). Aislamiento y evaluación de la actividad enzimática de hongos descomponedores de madera
en la reserva natural La montaña del ocaso, Quimbaya-Quindío. Microbiología Industrial, Javeriana: Bogotá. 109 p. p.
Chaparro D.F., Rosas D.C., Varela A. (2009). Aislamiento y evaluación de la actividad enzimática de hongos descomponedores
de madera (Quindío, Colombia) Revista Iberoamericana de Micologia, 26(4): 238-243.
Chen H., Xu F., Li Z. (2002). Recent developments in biodegradation of industrial pollutants by white rot fungi and their enzyme
system. Bioresource Technology 81: 261.
Chen Y.J., Shiao M.S., Lee S.S., Wang S.Y. (1997). Effects of Cordyceps sinensis on the proliferation and differentiation of
human leukemic U937 cells. Life Science, 60(23): 49-59.
Chihara G., Maeda Y., Humuro J., Sasaki T., Fukuoka F. (1969). Inhibition of mouse sarcoma 180 by polysaccharides from
Lentinus edodes Nature, 222: 687-688.
Chihara G., Hamuro J., Maeda Y., Arai Y., Fukuoka F. (1970). Fractionation and Purification of the Polysaccharides with Marked
Antitumor Activity, Especially Lentinan, from Lentinus edodes (Berk.) Sing, (an Edible Mushroom). Cáncer Research, 30:
2776-2781.
Chitiva - Jaramillo A., Torrenegra – Guerrero R., Cabrera - Parada C., Díaz - Puentes N., Pineda - Parra V. (2009). Contribución
al estudio de microhongos filamentosos en los ecosistemas páramo de guasca y el tablazo.[1-10]. Disponible en:
http://www.javeriana.edu.co/Facultades/Ciencias/gifuj/hongos_ ecosistemas_ paramo.pdf. [Visitada en
marzo de 2010].
Cho E.J., Oh J.Y., Chang H.Y., Yun J.W. (2006). Production of exopolysaccharides by submerged mycelial culture of a
mushroom Tremella fuciformis. Journal of Biotechnology, 127: 129-140.
Chu K.H., Kim E.Y. (2006). Predictive modeling of competitive biosorption equilibrium data. Biotehchnology Bioprocess
Engineerong, 11: 67-71.
Comtat J., Joseleau J. (1981). Mode of action of a xylanase and its significance for the structural investigation of the branched L-
arabino-Dglucurono-D-xylan from redwood (Sequoia sempervirens). Carbohydrates Resources, 95: 101-112.
Corrales A., López C.A. (2005). Macrocybe titans (Bigelow y Kimbr.) Pegler, Lodge y Nakasone, un registro nuevo para
Colombia. Actual Biología, 27(82): 93-97
Corredor I.C., Caridad M., Restrepo S. (2007). Evaluación de la suceptibilidad de hongos endófitos aislados de rosa (rosa
hybrida) a insecticidas comerciales. Revista Colombiana de Biotecnología, 9(1): 59-74.
Cortés M.R., García A.S., Suarez H.M. (2007). Fortificación de hongos comestibles (Pleurotus ostreatus) con calcio, selenio y
vitamina C. VITAE, Revista de la Facultad de Química Farmacéutica, 14(1): 16-24.
Coughlan M.P., Ljungdahl L.G. (1988). Comparative biochemistry of fungal and bacterial cellulolytic enzyme systems. En:
Biochemistry and Genetics of Cellulose Degradation Millet J.P.A.P.B.J. (Ed.). Academic Press: London. pp. 34 p.
Coughlan M.P., Hazlewood G.P. (1993). β-1,4-D-xylan-degrading enzyme systems: biochemistry, molecular biology and
applications. Biotechnology and Applied Biochemistry, 17: 259-289.
Cui F., Chisti Y. (2003). Polysaccharopeptides of Coriolus versicolor: physiological activity, uses and production. Biotechnology
and Advances, 21: 109-122.
Cui S. (2005). Food Carbohydrates, Chemistry, Physical propierties and Applications. CRC Press, ed. Group T.F. Taylor &
Francis Group: Boca Raton Florida. 404 p.
D’Annibale A., Celleti D., Felici M., Di Mattia E., Giovannozzi-Sermani G. (1996). Substrate specificity of laccase from Lentinus
edodes. Acta
Biotechnol. Bioengineering, 16: 257-270.
De Carvalho J.C., Pandey A., Oishi B.O., Brand D., Rodriguez-León J.A., Soccol C.R. (2005). Relation between growth,
respirometric anaysis and biopigments production from Monascus by solid-state fermentation. Biochemical Engineering
Journal, 29: 262-269.
Dekker R.F.H., Richards G.N. (1975). Purification, properties and mode of action of hemicellulase I produced by Ceratocystis
paradoxa. Carbohydrates Resources 39: 97-114.
Dekker R.F.H. (1985). Biodegradation of the hemicelluloses. En: Biosynthesis and Biodegradation of wood components. Higuchi
T. (Ed.). Academic press,: Orlando. pp. 113-132.
Delgado F., López Y., Giraldo E.M. (2001). Actividad enzimática de hongos y su patogenicidad sobre Hypothenemus hampei.
Manejo Integrado de Plagas (Costa Rica), 60: 43-49.

191
Dhouib A., Hamza M., Zouari H., Mechichi T., H’midi R., Labat M., Martínez M.J., Sayadi S. (2005). Autochthonous fungal
strains with high ligninolytic activities from Tunisian biotopes. African Journal of Biotechnology, 4(5): 431-436.
Dobozi M.S., Szakacs G., Bruschi C.V. (1992). xylanase activity of Phanerochaete Chrysosporium. Applied and environmental
microbiology, 58(11): 3466-3471.
Donot F., Fontana A., Baccou J., Schorr-Galindo S. (2012). Microbial exopolysaccharides: Main examples of synthesis, excretion,
genetics and extraction. Carbohydrate Polymers, 87: 951- 962.
Dormand J.R., Prince P.J. (1980). A family of embedded Runge-Kutta formulae. Journal of Computational and Applied
Mathematics, 6: 19-26.
Dubois M., Gilles K.A., Hamilton J.K., Rebers P.A., Smith F. (1956). Colorimetric method for determination of sugars and
related substances. Analytical Chemistry Engineering Journal, 28(3): 350-356.
Eggert C., Temp U., Dean J.F., Eriksson K.L. (1996). A fungal metabolite mediates degradation of non-phenolic lignin structures
and synthetic lignin by laccase. FEBS Letters, 391: 144-148.
Ek M., Gellerstedt G., Henriksson G. (2009). Pulp and Paper Chemistry and Technology: Wood Chemistry and Wood
Biotechnology. ed. KG W.d.G.G.C. Vol. 1. THE GRUITER: Berlin. 320 p.
El Gogary S., Leite O., Criverallo D.E., Eveleigh D.E., El-Dorry H. (1989). Mechanism by which cellulose triggers
cellobiohydrolase I gene expression in Trichoderma reesei. Proceedings of the National academy of Sciences of the United
States of America, 86: 6138-6141.
Elibol M. (2004). Optimization of medium composition for actinorhodin production by Streptomyces coelicolor A3 (2) with
response surface methodology. Process Biochemistry, 39: 1057-1062.
Elisashvili V., Parlar H., Kachlishvili E., Chichua D., Kvesitadze G. (2001a). Ligninolytic activity of basidiomycetes grown under
submerged and solid-state fermentation on plant raw material (sawdust of grapevine cuttings). Advances Food Science, 23:
117-123.
Elisashvili V., Parlar H., Kachlishvili E., Chihua D., Bakrazde M., Kokhreidze N., Kvesitadze G. (2001b). Potential of solid-state
fermentation for laccase production. Advance Food Science, 23: 117-123.
Elisashvili V., Penninckx M., Kachlishvili E., Asatiani M., Kvesitadze G., Gadd G. (2006). Use of Pleurotus dryinus for
lignocellulolytic enzymes production in submerged fermentation of mandarin peels and tree leaves. Enzyme and Microbial
Technology, 38: 998-1004.
Elisashvili V., Penninckx M., Kachlishvili E., Tsiklauri N., Metreveli E., Kharziani T., Kvesitadze G. (2008). Lentinus edodes and
Pleurotus species lignocellulolytic enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes of
different composition. Bioresource Technology, 99: 457-462.
Elisashvili V., Kachlishvili E., Khardziani T., Agathos S.N. (2010). Effect of aromatic compounds on the production of laccase
and manganese peroxidase by white-rot basidiomycetes. Journal of Industrial Microbiology and Biotechnology, 37: 1091-
1096.
Enshasy H.E. (2010). Immunomodulators. En: The Mycota. Hofrichter M. (Ed.). Springer-Verlag: Berlin, 477 p. pp. 477.
Evans C., Dutton. M., Guillen. F., Veness R. (1994). Enzymes and small molecular mass agents involved with lignocellulose
degradation. FEMS Microbiology Letters, 13: 235-240.
Fang Q.H., Zhong J.J. (2002). Effect of initial pH on production of ganoderic acid and polysaccharide by submerged fermentation
of Ganoderma lucidum. Process Biochemistry, 37(7): 69-74.
Fermor T.R., Wood D.A. (1981). Degardation of bacteria by Agaricus bisporus and other fungi. Journal of General Microbiology,
126: 377-387.
Fernández J.A., Henao L.M., Pedroza-Rodríguez A.M., Quevedo-Hidalgo B. (2009). Inmovilización de hongos ligninolíticos para
la remoción del colorante negro reactivo 5. Revista Colombiana de Biotecnología, 11(1): 59-72.
Ferraz A., Cordova A.M., Machuca A. (2003). Wood biodegradation and enzyme production by Ceriporiopsis subvermispora
during solidstate fermentation of Eucalyptus grandis. Enzyme and Microbiology Technology, 32: 59-65.
Forchiassin F., Levin L. (en imprenta). Enzimas Extracelulares Degradación de Polímeros Vegetales. En: Introducción al reino
de los hongos y los grupos afines. Albertó E., Vadell E. (Eds.). Literature for Latin America: Buenos Aires.
Fujian X., Hongzhang C., Zuohu L. (2001). Solid-state production of lignin peroxidase (LiP) and manganese peroxidase (MnP) by
Phanerochaete chrysosporium using steam-exploded straw as substrate. Bioresource Technology, 80: 149-151.
Gadd G.M. (1999). Fungal production of citric and oxalic acid: importance in metal speciation, physiology and biogeochemical
processes. Advance in Microbiology and Physiology, 41: 47-92.
Gadd G.M. (2001). Fungi in Bioremediation. Cambridge University Press: Cambridge. 481 p.
Galliano H., Gas G., Seris J.L., Boudet A.M. (1991). Lignin degradation by Rigidoporus lignosus involves synergistic action of
two oxidizing enzymes: manganese peroxidase and laccase. Enzyme and Microbial Technology, 13: 478-482.
Gao Y., Zhou S. (2003). Cancer prevention and treatment by Ganoderma, a mushroom with medicinal properties. Food Research
International, 19: 275-325.
Garcés-Molina A.M., Velez-Cardona N., Ruiz-Alzate S., Serna-D´León J.G., Suarez-Holguín E. (2006). Revista Lasallista de
Investigación, 2(2): 15-20.
García A.M., Torres R.G. (2003). Producción de enzimas ligninolíticas por Basidiomycetes mediante la técnica de fermentación
en fase sólida. Revista Colombiana de Biotecnología, 5(001): 56-64.
Gassara F., Brar S., Tyagi R.D., Verma M., Surampalli R.Y. (2010). Screening of agro-industrial wastes to produce ligninolytic
enzymes by Phanerochaete chrysosporium. Biochemical Engineering Journal 49: 388-394.

192
Glen J.K., Gold M.H. (1985). Purification and characterization of an extracellular Mn(II)-dependent peroxidase from the lignin-
degrading basidiomycetes Phanerochaete chrysosporium. Archives of Biochemistry and Biophysics, 242: 329-341.
Gold M.H., Alic M. (1993). Molecular Biology of the lignin-degrading Basidiomycete Phanerochaete Chrysosporium.
Microbiological Reviews, 57(3): 605-622.
Golovleva L.A., Malt’seva O.V., Leont’evskii A.A., Myasoedova N.M., Skryabin G.K. (1987). Ligninase biosynthesis by the
fungus Panus tigrinus during solid-state fermentation of straw. Doklady Akademii Nauk, 294: 992-995.
Gómez-Dorado C., Martinez-Salgado M., Nieto-Mosquera D., Pedrosa-Rodriguez A., Rodriguez-Vásquez R., Rosas-Acosta J.
(2005). Estudio del efecto de dos inductores y un protector enzimático sobre la actividad de las enzimas MnP y lacasa
producidas por Trametes versicolor y su acción en la decoloración. Revista de la Facultad de Ciencias Universidad
Javeriana, 10(2): 37-45.
Gong P., Guan X., Witter E. (2001). A rapid method to extract ergosterol from soil by physical disruption. Applied Soil Ecology,
17: 285-289.
Góngora C.E. (2009). Avances en conocimiento y mejoramiento del hongo entomopatógenos Beauveria bassiana para el control
de la broca del café, Hypothenemus hampei. Cenicafé. Disponible en:
http://almacafe.com.co/modules/News/images/simposio2.pdf. [Visitada en marzo de 2010].
Goodenough P.W. (1996). Structural studies on cellulases, pectinases an xylanases. En: Enzymes for Carbohydrate Engineering.
Elsevier Science B.V,. pp. 83-107.
Gow N.A.R. (1995a). The Growing Fungus. ed. Hall C. N.A.R Gow & G. M. Gadd: London. 402 p. p.
Gow N.A.R. (1995b). The growing Fungus. ed. Hall C. N.A.R Gow & G. M. Gadd: London.
Graminha E.B.N., Gonc¸alves A.Z.L., Pirota R.D.P.B., Balsalobre M.A.A., Da Silva R., Gomes E. (2008). Enzyme production by
solid-state fermentation: Application to animal nutrition. Animal Feed Science and Technology, 144: 1-22.
Granada R.D.M., Mejía G.A.I., Jimenez T.G.A. (2005). Utilización de residuos de plátano para la producción de metabolitos
secundarios por fermentación en estado sólido con el hongo Lentinus crinitus. Revista de la Facultad de Química
Farmacéutica, 12(2): 13-20.
Guiseppin M.L.F. (1984). Effects of disolved oxygen concentration on lipase production by Rhizopus delemar. Applied
Microbiology and Biotechnology, 20: 161-173.
Gutiérrez Ramírez L., Pérez Bran J., Uribe M. (2012). Evaluación in vitro de celulasas producidas por cepas nativas de
Trichoderma reesei, Cladosporium herbarum y Aspergillus niger. Journal Agricultural and Animal Science, 1(1): 7-15.
Gutierrez-Correa M., Tengerdy R.P. (1997). Production of cellulase on sugar cane bagasse by fungal mixed culture solid substrate
fermentation. Biotechnology Letters in Applied Microbiology, 19: 665 - 667.
Guzmán M.L., Angel J.C., Rivas A., Suarez L. (2003). Producción de hongos comestibles en residuos de caña de azúcar,
Memorias VI congreso Colombiano de la Asociación de Técnicos de la Caña de Azúcar: Colombia. 323-331.
Hamelinck C.N., van Hooijdonk G., Faaij A.P.C. (2005). Ethanol from lignocellulosic biomass: techno-economic performance in
short-, middle- and long-term. Biomass Bioenergy, 28: 384-410.
Hamlyn P.F., Schmidt R.J. (1994). Potential therapeutic application of fungal filaments in wound management. Mycologist, 8:
147-152.
Hammel K.E., Kapich A.N., Jensen Jr. K.A., Ryan Z.C. ( 2002). Reactive oxygen species as agents of wood decay by fungi.
Enzyme Microbiology and Technology, 30: 445-453.
Henry W.P. ( 2003). Non-enzymatic iron, manganese and copper chemistry of potencial importance in wood decay. En: Wood
Deterioration and Preservation - Advances in our Changing World. Society A.C. (Ed.). Goodell, B., Nicholas, D.D., Schultz,
T.P. (Eds.): Washington. pp. 175-195.
Hobbs C. (2000). Medicinal values of Lentinus edodes (Berk.) Sing. (Agaricomyvetideae). A literature review. Interantional
Journal of Medicinal Mushrooms, 2: 287-297.
Hölker U., Höfer M., Lenz J. (2004). Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Applied
Microbiology Biotechnology, 64: 175-186.
Hölker U., Lenz J. (2005). Solid-state fermentation - are there any biotechnological advantages? Current Option in Microbiology,
8: 301-306.
Hormiga J.A., Verab J., Frías I., Torres Dariasa N.V. (2008). Growth and ligninolytic system production dynamics of the
Phanerochaete chrysosporium fungus A modelling and optimization approach. Journal of Biotechnology, 137: 50-58.
Hsieh C., Liu C.-J., Tseng M.-H., Lo C.-T., Yang Y.-C. (2006). Effect of olive oil on the production of mycelial biomass and
polysaccharides of Grifola frondosa under high oxygen concentration aeration. Enzyme and Microbial Technology, 39: 434-
439.
Hsieh C., Wang H.-L., Chena C.-C., Hsub T.-H., Tseng M.-H. (2008). Effect of plant oil and surfactant on the production of
mycelial biomass and polysaccharides in submerged culture of Grifola frondosa. Biochemical Engineering Journal, 38: 198-
205.
Hudson H. (1986a). Fungal Biology. CPC press Edward Arnolds Eds: Maryland. 298 p. p.
Hudson H. (1986b). Fungal Biology. Edward Arnolds Eds: Maryland. 298 p. p.
Hyun H.H., Zeikus J.G. (1985). General Biochemical characterization of thermostable extracellular B-amylasa from Clostridium
thermosulfurogenes. Applied Environmental Microbiology, 49: 1162-1170.
Ikasari L., Mitchell D.A. (2000). Two-phase model of yhe kinetics of growth of Rhizopus oligosporus in membrana culture.
Biotechnology Bioengineering, 68: 619-627.

193
Ilmen M., Saloheimo A., Onnela M.L., Pentilla M. (1996). Rgulation of cellulase gen expression in the filamentous fungus
Trichoderma reesei. Applied Environmental Microbiology, 63: 1298-1306.
Ishikawa K., Majima T., Ebina T. (1992). Anti-tumour effect of a Coriolus preparation, PSK: Induction of macrophage
chemotactic factor (MCF) in spleens of tumour bearing mice. Biotherapy, 5: 251-258.
Ivshin V., Artamonova S., Ivshina T., Sharnina F. (2007). Methods for Isolation of Chitin–Glucan Complexes from Higher Fungi
Native Biomass. Polymer Science, Serie B., 49(11-12): 305-310.
Izawa M., Takashio M., Koshino S. (1995). Several new factors influencing the measurement of β-glucan content using
calcofluor flow-injection analysis method. Journal of the Institute of Brewing, 101: 371.
Jaramillo C., Rodriguez N., Gómez F.A. (1999). Cultivo de hongos tropicales sobre residuos agroindustriales presentes en la
zona cafetera. Informe final experimento QIN-09-23 (Cultivation of tropical mushrooms on agri-industrial wastes present in
coffee-growing region. Final report of experiment QIN-09-23, in Spanish), Cenicafé: Chinchiná, Colombia. 44.
Jaszek M., Malarczyk E., Leonowicz A. (1998). Investigation of ligninolytic enzymes during solid state fermentation of cotton
wastes by selected strains of basidiomycetes. En: Proceedings of the 7th International Conference on Biotechnology in the
Pulp and Paper Industry. Vancouver. pp. 16-18.
Jennings D.H. (1995). The Physiology of Fungal Nutrition, Cambridge, UK: Cambridge University press.
Johnson E.A., Sakajoh M., Halliwell G., Madia A., Demain A.L. (1982). Saccharification of complex cellulosic substrates by the
cellulase system from C. thermocellum. Applied and environmental microbiology, 43: 1125-1132.
Jong S.C., Birgmingham J.M. (1992). Medicinal benefits of the mushroom Ganoderma. Adv. Appl. Microbiol., 37: 101-134.
Joselau J.P., Ruel K. (1994). Wood polysaccharides and their degradation by fungi. En: Host Wall Alterations by Parasitic Fungi.
Ouellette P. (Ed.). APS Press: Minnesota. pp. 334-387.
Joshi V.K., Pandey A. (1999a). Biotechnology. Food fermentation Vol II. Educational Publishers & distributors: New Delhi. 998 p
p.
Joshi V.K., Pandey A. (1999b). Biotechnology: Food Fermentation (Microbiology, Biochemistry and Technology) Vol I.
Educational Publishers & distributiors: New Delhi. 521 P. p.
Joshi V.K., Pandey A. (1999c). Biotechnology. Food fermentation Vol. II. Educational Publishers & Distributors: New Delhi. 998
p p.
Kachlishvili E., Penninckx M., Tsiklauri N., Elisashvili V. (2006). Effect of nitrogen source on lignocellulolytic enzyme
production by white-rot basidiomycetes under solid-state cultivation. World Journal of Microbiology & Biotechnology,
22(4): 391-397.
Kalisz H.M., Wood D.A., Moore D. (1987). Production, regulation and release of extracellular proteinasa activity in
basidiomycetes fungi. Transactions of the British Mycological Society, 88: 221-227.
Kalisz H.M. (1988). Microbial Proteinases. Advances in Biochemical Engineering/Biotechnology, 36: 1-65.
Kalisz H.M., Wood D.A., Moore D. (1989). Some Characteristics of extracellular proteinases from Coprinus cinereus.
Mycological Research, 92: 278-285.
Kapich A.N., Prior B.A., Botha A., Galkin S., Lundell T., Hatakka A. (2004). Effect of lignocellulose-containing substrate on
production of ligninolytic peroxidases in submerged cultures of Phanerochaete chrysosporium ME-446. Enzyme and
Microbial Technology, 34: 187-195.
Karunanithy C., Muthukumarappan K., Julson J.L. (2008). Influence of high shear bioreactor parameters on carbohydrate release
from different biomasses American Society of Agricultural and Biological Engineers Annual International Meeting ASABE
084114. ASABE.
Kersten P., Cullen D. (2007). Extracellular oxidative systems of the lignin-degrading Basidiomycete Phanerochaete
chrysosporium. Fungal Genetics and Biology, 44: 77-87.
Kiho T., Ookubo K., Usui S., Ukai S., Hirano K. (1999). Structural features and hypoglycemic activity of a polysaccharides (CS-
F10) from the cultured mycelium of Cordyceps sinensis. Biological and Pharmaceutical Bulletin, 22(9): 66-70.
Kim F., Sakagami H., Tanuma S., Konno K. (1990). Stimulation of interferon-g-induced human myelogenous leukemic cell
differentiation by high molecular weight PSK subfraction. Anticancer Research, 10: 55-58.
Kim S., Holtzapple M.T. (2006). Delignification kinetics of corn stover in lime pretreatment. Bioresource Technology, 97: 778-
785.
Kim T.H., Taylor F., Hicks K.B. (2008a). Bioethanol production from barley hull using SAA (soaking in aqueous ammonia)
pretreatment. . Bioresouce Technology, 99: 5694-5702.
Kim T.H., Taylor F., Hicks K.B. (2008b). Bioethanol production from barley hull using SAA (soaking in aqueous ammonia)
pretreatment. Bioresource Technology, 99: 5694-5702.
Kirk T.K., Connors W., Zeikus J. (1976). Requeriment for a growth substrate during lignin decomposition by two wood-roting
fungi. Applied Environmental Microbiology, 32: 192-194.
Kirk T.K. (1983). Degardation and convertion of lignocelluloses. En: The filamentous fungi. Smith J.E., Berry D.R., Kristiansen
P. (Eds.). London. pp. 266-295.
Kirk T.K., Farrel R.L. (1987a). Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology, 41:
465-505.
Kirk T.K., Farrel R.L. (1987b). Enzymatic combustion: the microbial degradation of lignin. Annual Reviews of Microbiology 41:
465-505.
Kjeldahl J. (1883). Neue Methode Zùr Bestimmung der Stickstoffs in Organichen Kòrpen. Zeitschrift fur Analytische Chemie 22:
366-382.

194
Kobayashi H., Matsunaga K., Oguchi Y. (1995). Antimetastatic Effects of P5K (Krestin), a Protein-bound Polysaccharide
Obtained from Basidiomycetes: An Overview. Cancer Epidemiology, Biomarkers & Prevention, 4: 275-281.
Koduri R.S., Tien M. (1994). Kinetic analysis of lignin peroxidase-explanation for the mediation phenomen by veratryl alcohol.
Biochemistry, 33: 4225-4230.
Koh J.H., Kim J.M., Chang U.J., Suh H.J. (2003). Hypocholesterolemic effect of hotwater extract from mycelia of Cordyceps
sinensis. Biological and Pharmaceutical Bulletin, 26: 84-87.
Komatsu M., Okubo S., Kikumoto S., Kimura K., Saito G., Sakai S. (1969). Host-mediated antitumor action of Schizophyllan a
glucan produced by Schizophyllum commune. Gann, 60: 137-144.
Komura D.L., Ruthers A.C., Carbonero E.R., Alquini G., Rosa M.C., Sassaki G.L., Lacomini M. (2010). The origin of mannans
found in submerged culture of basidiomycetes. Carbohydrate polymers, 79: 1052-1056.
Kootstra A.M.J., Beeftink H.H., Scott E.L., Sanders J.P.M. (2009). Comparison of dilute mineral and organic acid pretreatment
for enzymatic hydrolysis of wheat straw. Biochemical Engineering Journal, 46: 126-131.
Kosaka A., Yamashita A. (1993). Lentinan as an effective inmmunomodulator against breast cancer, with special reference to
clinical applications. International Journal Immunotherapy, 9: 111-116.
Koutinas A.A., Wang R., Webb C. (2004). Restructuring Upstream Bioprocessing: Technological and Economical Aspects for
Production of a Generic Microbial Feedstock From Wheat. Published online in Wiley InterScience: 1-15.
Kovarova-Kovar K., Egli T. (1998). Growth kinetics of suspended microbial cells: from single substrate-controlled growth to
mixed-substrate kinetics. Microbiology and Molecular Biology Reviews, 62: 646-666.
Kumar R., Wyman C.E. (2009). Effects of cellulase and xylanase enzymes on the deconstruction of solids from pretreatment of
poplar by leading technologies. Biotechnology Progress, 25: 302-314.
Kumari J., Sahoo P.K. (2006). Non-specific immune response of healthy and immunocompromised Asia catfish (Clarias
batrachus) to several immunostimulants. Aquaculture, 255: 133-141.
Kupfahl C., Geginat G., Hof H. (2006). Lentinan has a stimulatory effect on innate and adaptive immunity against murine
Listeria monocytogenes infection. International Immunopharmacology, 6: 686- 696.
Ladner W., Ditrich K. (1999). Biocatalytic production of chiral intermediates. Chimica Oggi 7/8: 51-54.
Lama L., Nicolaus B., Calandrell V., Manca M., Romano I., Gambacorta A.M., Arambarri. (1996). Effect of growth conditions on
endo- and exopolymer biosynthesis in Anabaena cylindrica 10C. Phytochemistry, 42: 655-664.
Lang E., Nerud F., Novotna E., Martens R. (1996). Production of ligninolytic exoenzymes and 14C-pyrene mineralization by
Pleurotus sp. in lignocellulose substrate. Folia Microbiology, 41: 489-493.
Lee B., Bae J., Pyo H., Choe T., Kim S., Hwang H., Yun J. (2004). Submerged culture conditions for the production of mycelial
biomass and exopolysaccharides by the edible basidiomycete Grifola frondosa. Enzyme and Microbial Technology, 35: 369-
376.
Lee K., Lee S., Lee H. (1999). Bistage control of pH for improving exopolysaccharide production from mycelia of Ganoderma
lucidum in an air-lift fermentor. Journal of Bioscience and Bioengineering, 88: 646-650.
Lee W.Y., Park Y., Ahn J.K., Ka K.H., Park S.Y. (2007a). Factors influencing the production of endopolysaccharide and
exopolysaccharide from Ganoderma applanatum. Enzyme and Microbial Technology, 40 249-254.
Lee W.Y., Park Y., Ahn J.K., Ka K.H., Park S.Y. (2007b). Factors influencing the production of endopolysaccharide and
exopolysaccharide from Ganoderma applanatum. Enzyme and Microbial Technology 40 249-254.
Lemaire M. (1996). The Cellulosome - an exocellular multiprotein complex specialised in cellulose degradation. Critical Reviews
and Biochemistry & Molecular Biology, 31: 201-236.
Lenz J., Höfer M., Krasenbrink J.B., Hölker U. ( 2004). Computational and physical methods in solid-state fermentation. Applied
Microbiology Biotechnology, 65: 9-17.
Leonowicz A., Matuszewska A., Luterek J., Ziegenhagen D., Wojtas-wasilewska M., Cho N.S., otros. (1999). Biodegradation of
lignin by white-rot fungi. Fungal Genetics and Biology 27: 175-185.
Leschine S.B. (1995). Cellulose degradation in anerobic environments. Annual Review Microbiology, 49: 399-410.
Leterme P. (2010a). Análisis de alimentos y forrajes, in Protocolos de LaboratorioUniversidad Nacional de Colombia Sede
Palmira: Palmira. 66 pp.
Leterme P. (2010b). Análisis de alimentos y forrajes. Protocolos de Laboratorio Universidad Nacional de Colombia: Palmira. 66
pp. p.
Leung M., Fung K., Choy Y. (1997). The isolation and characterization of an immunomodulatory and antitumor polysaccharide
preparation from Flammulina velutipes. Immunopharmacol 35: 255-263.
Leung M., Liu C., Koon J., Fung K. (2006). Polysaccharide biological response modifiers. Immunologycal Letters, 105: 101-114.
Levin L. (1998). Biodegradación de materiales lignocelulósicos por Trametes trogii (Aphyllophorales, Basidiomycetes).Tesis
Doctoral. Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires: Buenos Aires.
Levin L., Forchiassin F., Ramos A.M. (2002). Copper induction of lignin-modifying enzymes in the white-rot fungus Trametes
trogii. Mycologia, 94(3): 377-383.
Levin L., Papinutti L., Forchiassin F. (2004). Evaluation of Argentinean white rot fungi for their ability to produce lignin-
modifying enzymes and decolorize industrial dyes. Bioresource Technology, 94: 169-176.
Levin L., Herrmann C., Papinutti V.L. (2008). Optimization of lignocellulolytic enzyme production by the white-rot fungus
Trametes trogii in solid-state fermentation using response surface methodology. Biochemical Engineering Journal 39: 207-
214.

195
Li L., Li X.Z., Tang W.Z., Zhao J., Qu Y.-B. (2008). Screening of a fungus capable of powerful and selective delignification on
wheat straw. Letters in Applied Microbiology, 47: 415-420.
Li Q., He Y.C., Xian M., Jun G., Xu X., Yang J.M., Li L.Z. (2009). Improving enzymatic hydrolysis of wheat straw using ionic
liquid 1-ethyl-3-methyl imidazolium diethyl phosphate pretreatment. Bioresource Technology, 100: 3570-3575.
Li X., Liun Y., Ji L. (2000). Production of lignocellulolytic enzymes in solid state fermentation using corn stalks as substrate.
Henong Xuebao, 14: 99-103.
Lia K., Azadic P., Collinsb R., Toland J., Kimc J.S., E. K.E.L. (2000). Relationships between activities of xylanases and xylan
structures. Enzyme and Microbial Technology, 27: 89-94.
Lin E., Sung S. (2006). Cultivating conditions influence exopolysaccharide production by the edible Basidiomycete Antrodia
cinnamomea in submerged culture. International Journal of Food Microbiology, 108: 182-187.
Lindblom C.M., Wachenfeldt E.V., Tranvik L.J. (2004). Ergosterol as a measure of living fungal biomass: persistence in
environmental samples after fungal death. Journal of Microbiological Methods, 59 253- 262.
Lindequist U., Niedermeyer T., Ju¨lich W.-D. (2005a). The pharmaceutical potential of mushrooms. Electronic centralised aircraf
monitor (eCAM), 2: 285-299.
Lindequist U., Niedermeyer T., Julich W.-D. (2005b). The pharmaceutical potential of mushrooms. Electronic Centralised
Aircraf Monitor (eCAM), 2: 285-299.
Litthauer D., van Vuuren, M.J.,, van Tonder A., Wolfaardt F.W. (2007). Purification and kinetics of a thermostable laccase from
Pycnoporus sanguineus (SCC 108). Enzyme and Microbial Technology, 40: 563-568.
Liu C., Liu Y., Liao W. (2003). Application of statistically based experimental designs for the optimization of nisin production
from whey. Biotechnology Letters, 25: 877-882.
Liu F., Fung M.C., Ooi V.E.C., Chang S.T. (1996a). Induction in the mouse of gene expression of immunomodulating cytokines
by mushroom polysaccharide-protein complexes. Life Science, 58(1): 795-803.
Liu F., Fung M.C., V.E.C. O., Chang S.T. (1996b). Induction in the mouse of gene expression of immunomodulating cytokines by
mushroom polysaccharide-protein complexes. Life Sciences, 58(1): 795-803.
Lockington R.A., Rodbourn L., Barnett S., Carter C.J., Kelly J.M. (2002). Regulation by carbon and nitrogen sources of family of
cellulases in Aspergillus nidulans. Fungal Genetics and Biology, 37: 190-196.
Lonsane B.K., Ghildyal N.P., Budiatman S., Ramakrishna S.V. (2005). Solid state fermentation for production of α-amylase by
Bacillus megaterium 16M Enzyme Microbiology Technology, 7: 258-265.
Lowry O.H., Rosebrough N.J., Farr A.L., Randall R.J. (1951). Protein measuremnet with the folin phenol reagent. Department of
Pharmacology, Washington University School oj Medicine, St. Louis, Missouri: 265 - 275.
Lozano J.C. (1989). Producción comercial del Champiñon /Pleurotus ostreatus/ en pulpa de café, CONGRESO ASCOLFI, X.
Cali (Colombia): Manizales. 60 p.
Luedeking R., Piret E. (1959). Transient and steady states in continuous fermentation. Theory and experiment. Journal of
Biochemical and Microbiological Technology and Engineering, 1: 431-459.
Lull C., Wichers H., Savelkoul H. (2005). Antiinflammatory and immunomodulating properties of fungal metabolites. Mediator
Inflammation, 2: 63-80.
Lynd L.R., Weimer P.J., Zyl W.H.v., Pretorius I.S. (2002). Microbial cellulose utilization: Fundamentals and Biotechnology.
Microbiological and Molecular Biology Reviews, 66(3): 506-577.
Maganhotto de Souza-Silva C.M., Soares de Melo I., de Oliveira P.R. (2005). Ligninolytic enzyme production by Ganoderma
spp. Enzyme and Microbial Technology 37: 324-329.
Mandels M., Reese E. (1956). Induction of cellulase in Trichoderma viride as influenced by carbon sources and metals. Biology
Branch: 269-277.
Mandels M., Weber J. (1969). The production of cellulases and their applications. Advances Chemistry, 95: 391-414.
Mansel P.W.A. (1994). Polysaccharides in skin care. Cosmetics Toilet, 109: 7-72.
Mansfield S.D., Mooney C., Sanddler J.N. (1999). Substrate and enzyme characteristics that limit cellulose hydrolysis.
Biotechnological progress, 15: 804-816.
Markham P. (1998). Occlusions of septal pores in filamentous fungi. Mycological Research, 98: 1089-1106.
Marques de Souza C.G., Zilly A., Peralta R.M. (2002). Production of laccase as the sole phenoloxidase by a Brazilian strain of
Plerotus pulmonarius in solid state fermentation. Journal Basic Microbiology, 42: 83-90.
Martinez A. (2002). Molecular Biology and structure-function of lignin-degradin heme peroxidases. Enzyme Microbiology
Technology, 30: 425-444.
Martinez M., Ruiz-Dueñas F., Guillen F., Martinez A. (1996). Purifacation and catalytic properties of two manganese peroxidase
isoenzymes from Pleurotus eryngii. European Journal Biochemistry, 237: 424-432.
Martínez M.J., Ruíz-Dueñas F.J., Guillén F., Martínez A.T. (1996). Purification and catalytic properties of two manganese-
peroxidase isoenzymes from Pleurotus eryngii. European Journal of Biochemistry, 237: 424-432.
Martínez-Anaya C., Balcázar-López E., Dantán-González E., Folch-Mallol J., L. (2008). Celulasas fúngicas: Aspectos biológicos
y aplicaciones en la industria energética. Revista Latinoamericana de Microbiología, 50(3 y 4): 119-131.
Mata G., Savoie J.M. (1998). Extracellular enzyme activities in six Lentinula edodes strains during cultivation in wheat straw.
World Journal of Microbiology and Biotechnology, 14: 513-519.
Mata G., Murrieta Hernández D.M., Iglesias Andreu L.G. (2005). Changes in lignocellulolytic enzyme activites in six Pleurotus
spp. strains cultivated on coffee pulp in confrontation with Trichoderma spp. World Journal of Microbiology &
Biotechnology, 21: 143-150.

196
Mattila P., Lampi A.M., Ronkainen R., Toivo J., Piironen V. (2002). Sterol and Vitamin D2 contents in some wild and cultivated
mushroomn. Food Chemistry, 76: 293-298.
Mayer A.M., Staples R.C. (2002). Laccase: new functions for an old enzyme. Phytochemistry, 60: 551-565.
Maziero R., Cavazzoni V., Ramos-Bononi V.L. (1999). Screening of Basidiomycetes for the production of exopolisaccharides and
biomass in submerged culture. Revista de Microbiologia, 30: 77-84.
Meagher M.M., Tao B.Y., Chow J.M., Reilly P.J. (1988). Kinetics and subsite mapping of β -D-xylobiose and D-xylose
producing Aspergillus niger endo-β-1,4-D-xylanase. Carbohydrates Resources, 173: 273-283.
Meier H. (1955). Decomposition of the cell wall by wood-destroying fungi and the submicroscopic structure of tracheids of
spruce and wood fibers of birch (translation Nº 92: Department of the Secretary of State of Canada, February 1956) Holz.
Roh-Werkst., 13: 323-338.
Mester T., Sierra-Alvarez R., Holzforschung. (1998). Peroxidase and aryl metabolite production by the white rot fungus
Bjerkandera sp. strain BOS55 during solid state fermentation of lignocellulosic substrates. J. Field, 52: 351-358.
Meyer H.P., Kiener A., Imwinkelried R., Shaww N. (1997). Biotransformations for fine chemicals production. Chimia, 51: 287-
289.
Miller G.L. (1959). Use of DinitrosaIicyIic Acid Reagent for Determination of Reducing Sugar. Analytical Chemistry, 31(3): 426-
428.
Mitchell D., Stuart D., Tanner R. (1999). Solid-State fermentation - Microbial growth kinetics, in The Encyclopedia of Bioprocess
Technology: Fermentation, Biocatalysis and bioseparation, Flickinger M., Drew S., EditorsWiley: New York. 2407-2429
Mitchell D., Von Meien O., Krieger N. (2002). Recent developments in modeling of solid-state feremntation: heat and mass
transfer in bioreactors. BiochemicalEngineering Journal, 13: 137-147.
Mitchell D.A., Von meien O.F., Krieger N., Dalsenter F.D. (2004). A review of recent developments in modeling of microbial
growth kinetics and intraparticle phenomena in solid-state fermentation. Biochemical Engineering Journal 17: 15-26.
Mizuno T. (1992). Antitumor active polysaccharides isolated from the fruiting body of Hericium erinaceum, and edible, and
medicinal mushroom called Yamabushitake or Houtou. Bioscience Biotechnology and Biochemistry, 56: 349-357.
Mizuno T., Saito H., Nishitoba T., Kawagishi H. (1995). Antitumor-active substances from mushrooms. Food review
international, 11: 23-61.
Mizuno T. (1999). The extraction and development of antitumouractive polysaccharides from medicinal mushrooms in Japan.
International Journal Medicine Mushrooms, 1: 9-29.
Mohammad-Fata M., Hossein M., Shirin G., Ghorban-Ali H. (2007). Immunomodulating and anticancer agents in the realm of
macromycetes fungi (macrofungi). International Immunopharmacology 7: 701-724.
Moncada B., Forero E. (2006). El género Pseudocyphellaria VAIN. (Lobariaceae - ascomycetes liquenizados) en Colombia.
Caldasia, 28(2): 197-215.
Montenecourt B.S., Eveleigh D.E. (1977). Semiquantitative plate assay for determination of cellulase production by Trichoderma
viride. Applied and Environmental Microbiology, 33(1): 178-183.
Montes B., Restrepo A., McEwen J.G. (2003). Nuevos aspectos sobre la clasificación de los hongos y su posible aplicación
médica. Biomédica, 23: 213-237.
Montgomery H.J., Monrealb C.M., Young J.C., Seifert K.A. (2000 ). Determinination of soil fungal biomass from soil ergosterol
analyses. Soil Biology & Biochemistry, 32: 1207±1217.
Montoya B.S., Varon L.M., Levin L. (2008a). Effect of culture parameter on the production of the edible mushroom Grifola
frondosa (maitake) in tropical weathers. World J. Microbiol. Biotechnol., 24: 1361-1366.
Montoya B.S., Varon L.M., Levin L. (2008b). Effect of culture parameter on the production of the edible mushroom Grifola
frondosa (maitake) in tropical weathers. World Journal of Microbiology and Biotechnology, 24: 1361-1366.
Montoya B.S., Orrego C.A., Levin L. (2011a). Growth, fruiting and lignocellulolytic enzyme production by the edible mushroom
Grifola frondosa (maitake). World Journal of Microbiology and Biotechnology, ISSN 0959-3993: DOI 10.1007/s11274-
11011-10957-11272.
Montoya S., Restrepo G.M. (2006). Evaluación de la síntesis de proteína a partir del crecimiento vegetativo de Pleurotus sp. sobre
residuos de algarrobo y uva pasa. Revista Universidad de Caldas, 26(1-2): 23-32.
Montoya S. (2008). Actividad enzimática, degradación de residuos sólidos orgánicos y generación de biomasa útil por el
macromiceto Grifola frondosa.Investigación. Ingeniería Química, Universidad Nacional de Colombia Sede Manizales:
Manizales. 95 p. p.
Montoya S., Gallego J., Sucerquia A., Pelaez B., Betancourt O., Arias D. (2010). Macromicetos observados en bosques del
Departamento de Caldas: su influencia en el equilibrio y la conservación de la biodiversidad. Boletín Científico Centro de
Museos de Historia Naural, 14(2): 57-76.
Montoya S.B., Restrepo G.M., Tabarez L.A. (2009a). Rendimientos en el cultivo de los hongos Pleurotus ostreatus. Revista de
Investigaciones Universidad Católica de Manizales, 14: 16-26.
Montoya S.B., Restrepo G.M.F., Tabarez L.A.L. (2009b). Rendimientos en el cultivo de los hongos Pleurotus ostreatus. Revista
de Investigaciones Universidad Católica de Manizales, 14: 16-26.
Montoya S.B., Orrego C.A., Levin L. (2011b). Modeling Grifola frondosa fungal growth during solid-state fermentation.
Engineering in Life Science, 11(316-321).
Moon S.H., Park J.M., Chun H.Y., Kim S.J. (2006). Comparisons of physical properties of bacterial cellulose produced in
different culture conditions using saccharified food wastes. Biotechnology Bioprocess Engineering, 11: 26-31.
Moore D. (1998a). Fungal Morphogenesis. Cambrige University Press: New York, United States of America.

197
Moore D. (1998b). Fungal Morphogenesis. Primera ed Cambrige University Press.: New York. 469 p. p.
Moreira M.T., Feijoo G., Lema J.M. (2003). Fungal bioreactors: Applications to white-rot fungi. Reviews in Environmental
Science and Bio/Technology 2: 247-259.
Mosier N., Hendrickson R., Ho N., Sedlak M., Ladisch M.R. (2005a). Optimization of pH controlled liquid hot water pretreatment
of corn stover. Bioresource Technology, 96: 1986-1993.
Mosier N., Wyman C.E., Dale B.D., Elander R.T., Lee Y.Y., Holtzapple M., Ladisch C.M. (2005b). Features of promising
technologies for pretreatment of lignocellulosic biomass. Bioresource Technology, 96: 673-686.
Mouso N., Papinutti L., Forchiassin F. (2003). Efecto combinado del cobre y pH inicial del medio de cultivo sobre la producción
de lacasa y manganeso peroxidasa por Stereum hirsutum (Willd) Pers. Revista Iberoamericana de Micología, 20: 176-178.
Mukhamedzhanova T.G., Bezborodov A.M. (1982). Effect of nitrogen and carbon sources on lipase accumulation by the fungus
Rhizopus oryzae 1414. Mikrobiologia, 18: 16-22.
Muller dos Santos M., Souza das Rosa A., Dal’Boit, Mitchell D., Krieger N. (2004). Thermal denaturation: is solid-state
fermentation really a good technology for the production of enzymes? Bioresource Technology 93: 261-268.
Munoz G.A., Agosin E., Cotoras M., San Martin R., Volpe D. (1995). Comparison of aerial and submerged spore properties for
Trichoderma harzianum. FEMS Microbiology Letter, 125: 63-70.
Naeem S. (2002). Autotrophic-Hecterotrophic Interactions and their Impacts on Biodiversity and Ecosystem Functioning. En:
Functional Consequences of Biodiversity. Princeton University Press,: New Jersy. pp. 96-119.
Nakamura K., Yamaguchi Y., Kagota S., Shinozuka K., Kunitomo M. (1999). Activation of in vivo Kupffer cell function by oral
administration of Cordyceps sinensis in rats. . Japan Journal Pharmacology, 79: 508-508.
Nazareno M.C., Bucsinszky A.M.M., Tournier H.A., Cabello M.N., Arambarri A.M. (2000). Extracellular ABTS-oxidizing
activity of autochthonous fungal strains from Argentina in solid medium. Revista Iberoamericana de Micología, 17: 64-68.
Nelson N.J. (1944). A photometric adaptation of the Somogyi method for the detrmination of glucose. Journal Biochemistry, 153:
375-380.
Nidetzky B., Steiner W., Hayn M., Claeyssens M. (1994). Cellulose hydrolysis by the cellulases from Trichoderma reesei: a new
model for synergistic interaction. Bichemistry Journal, 298: 705-710.
Nie S.-P., Xie M.-Y. (2011). A review on the isolation and structure of tea polysaccharides and their bioactivities. Food
Hydrocolloids, 25: 144-149.
Nieto I., Chegwin C. (2010). Influencia del sustrato utilizado para el crecimiento de hongos comestibles sobre sus características
nutraceúticas. Revista Colombiana de Biotecnología, 12(1): 169-178.
Nieto I.J., Cucaita R.E. (2007). Ácidos grasos, ésteres y esteroles del cuerpo fructífero del hongo Laccaria laccata. Revista
Comobiana de Química, 36(3): 277-284.
Nieto I.J., Ávila I.M. (2008). Determinación de ácidos grasos y compuestos triterpenoides del cuerpo fructífero de Suillus luteus.
Revista Colombiana de Química, 37(3): 45-52.
Nieto-Ramirez I.J., Chegwin-Angarita C., Osorio-Zuluaga H.J. (2007). Incorporación de cafeína en el hongo Pleurotus sajor-caju
cultivado sobre pulpa de café. Revista Iberoamericana de Micología 24: 72-74.
Nigam P., Pandey A., Prabhu K.A. (1987). Cellulase and ligninase production by basidiomycete culture in solid-state
fermentation. Biology Wastes, 20: 1-9.
Niku-Paavola M., Raaska M., Itavaara M. (1990). Detection of white-rot fungi by non-toxic stain. Mycology and Resource, 94:
27-31.
Niladevi K.N., Prema P. (2008). Effect of inducers and process parameters on laccase production by Streptomyces psammoticus
and its application in dye decolourization. Bioresource Technology, 99 4583-4589.
Nitschke J., Altenbach H.-J., Malolepszy T., Mölleken H. (2011). A new method for the quantification of chitin and chitosan in
edible mushrooms. Carbohydrate Research, 346: 1307-1310.
Ooi V., Liu F. (2000a). Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Current Medicine
Chemistry, 7: 715-728.
Ooi V., Liu F. (2000b). Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Current Medicinal
Chemistry, 7: 715-728.
Ooi V.E.C., Liu F. (1999). A review of a farmacological activities of mushroom polysaccharides. International Journal Medicinal
Mushrooms, 1: 195-206.
Ortiz-Moreno M. (2010). Macromicetos en Zona Rural de Villavicencio. Orinoquía, 14(2): 125-132.
Pal M., Calvo A., Terron M.C., Gonzalez A.E. (1995a). Solid-state fermentation of sugarcane bagasse with Flammulina velutipes
and Trametes versicolor. World Journal Microbiology Biotechnology, 11: 541-545.
Pal M., Calvo A., Terron M.C., Gonzalez A.E. (1995b). Solid-state fermentation of sugarcane bagasse with Flammulina velutipes
and Trametes versicolor. World Journal Microbiology and Biotechnology, 11: 541-545.
Palonen H., Thomsen A.B., Tenkanen M., Schmidt A.S., Viikari L. (2004). Evaluation of wet oxidation pretreatment for
enzymatic hydrolysis of softwood. Applied Biochemistry and Biotechnology, 117: 1-17.
Panagiotou G., Kekos D., Macris B., Christakopoulos P. (2003). Production of cellulolytic and xylanolytic enzymes by Fusarium
oxysporum grown on corn stover in solid state fermentation. Industrial Crops and Products, 18: 37-45.
Pandey A., Soccol C.R., Mitchell D. (2000). New developments in solid state fermentation: I-bioprocesses and products. Process
Biochemistry, 35: 1153–1169.
Papinutti L. (2010). Effects of nutrients, pH and water potential on exopolysaccharides production by a fungal strain belonging to
Ganoderma lucidum complex. Bioresource Technology, 101: 1941-1946.

198
Papinutti V.L., Dorio L.A., Forchiassin F. (2003). Degradación de madera de álamo por Fomes sclerodermeus: Producción de
enzimas ligninolíticas en aserrín de álamo y cedro. Revista Iberoamericana de Micología, 20: 16-20.
Park J.-P., Kim S.-W., Hwang H.-J., Cho Y.-J., Yun J.-W. (2002a). Stimulatory effect of plant oils and fatty acids on the exo-
biopolymer production in Cordyceps militaris. Enzyme and Microbial Technology, 31: 250-255.
Park J.P., Kim S.W., Hwang H.J., Cho Y.J., Yun J.W. (2002b). Stimulatory effect of plant oils and fatty acids on the exo-
biopolymer production in Cordyceps militaris. Enzyme and Microbial Technology, 31: 250-255.
Pasquel A. (2001). Gomas: una aproximación a la Industria de alimentos. Revista Amazónica de Investigación Alimentaria, 1(1):
1-8.
Paszczczynski A., Crawford R., Huyn V. (1988). Manganese peroxidase of Phanerochaete chrysosporium purification. Methods
enzymology, 161: 264-270.
Paszczynski A., Crawford R.L. (1991). Degradation of azo compounds by ligninases from Phanerochaete chrysosporium
Involment of veratryl alcohol. Biochemistry Biophysics resources communications, 178: 1056-1063.
Patil S., Dayanand A. (2006). Optimization of process for the production of fungal pectinases from deseeded sunXower head in
submerged and solid-state conditions. Bioresource Technology, 97: 2340-2344.
Pelaez F., Martinez M.J., Martinez A.T. (1995). Screening of 68 species of basidiomycetes for enzymes involved in lignin
degradation. Mycology Resources, 99: 37-42.
Peña-Serna C., Sierra-Cadavid A., Sáez A., Rojano B.A. (2006). Establecimiento de un medio de cultivo para una cepa nativa de
un hongo Poliporal. Revista Colombiana de Biotecnología, 8(001): 5-13.
Peña-Venegas C.P., Cardona G.I., Arguelles J.H., Arcos A.L. (2007). Micorrizas Arbusculares del Sur de la Amazonia
Colombiana y su Relación con Algunos Factores Fisicoquímicos y Biológicos del Suelo. Acta Amazónica, 37(3): 327-336.
Pérez J., Jeffries T.W. (1992). Roles of Manganese and organic acid chelators in regulating lignin degradation and biosynthesis of
peroxidases by Phanerochaete chrysosporium. App Environl Microbiol, 58: 2402-2409.
Plackett R.L., Burman J.P. (1946). The design of optimum multifactorial experiments. Biometrika, 33: 305-325.
Plassard C., Mousain D., Salsac L. (1982). Estimation of mycelial growth of basidiomycetes by means of chitin determination.
Phytochemistry, 21: 345-348.
Pointing S.B. (1999). Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal
Diversity, 2: 17-33.
Pointing S.B. (2001). Feasibility of bioremediation by white-rot fungi. Applied Microbiology Biotechnology, 57: 20-33.
Pradeep V., Datta M. (2002). Production of ligninolytic enzymes for decolorization by cocultivation of white-rot fungi Pleurotus
ostreatus and Phanerochaete chrysosporium under solid-state fermentation. Applied Biochemistry Biotechnology, 102-103:
109-118.
Prosser J.I., Tough A.J. (1991). Growth mechanism and growth kinetics of filamentous microorganisms. Critical Reviews in
Biotechnology, 10: 253-274.
Quintero J.C., Moreira M.T., Feijoo G., Lema J.M. (2008). Screening of white rot fungal species for their capacity to degrade
lindane and other isomers of hexachlorocyclohexane (HCH). Ciencia e Investigación Agraria, 35(2): 159-167.
Quintero J.D., Feijoo G.C., Lema J.M. (2006). Producción de enzimas ligninolíticas con hongos basidiomicetos cultivados sobre
materiales lignocelulósicos. Revista de la Facultad de química Farmacéutica, 13(2): 61-67.
Raghavarao K.S., Ranganathan T.V., Karanth N.G. (2003). Some engineering aspects of solid-state fermentation. Biochemical
Engineering Journal, 13: 127-135.
Reddy C.A. (1995). The potential for white-rot fungi in the treatment of pollutants. Current Opinion in Biotechnology, 6: 320-
328.
Reddy G.V., Ravindra B.P., Komaraiah P., Roy R.M., Kothari I.L. (2003). Utilization of banana waste for the production of
lignolytic and cellulolytic enzymes by solid substrate fermentation using two Pleurotus species (P. ostreatus and P. sajor-
caju). Process Biochemistry, 38: 1457-1462.
Reid I.D. (1995). Biodegradation of lignin. Canadian Journal of Botany, 73: S1011-S1018.
Rivera A., Laverde C.M., Ríos-Motta J., Nieto I.J., Osorio H.J. (2005a). Dehidroergosterol: un artefacto generado durante el
proceso de extracción de esteroles en el hongo Pleurotus sajor-caju. Revista Colombiana de Química, 34(2): 117-125.
Rivera A., Osorio H.J., Ríos-Motta J., Nieto I.J., Laverde C.M. (2005b). Dehidroergosterol: un artefacto generado durante el
proceso de extracción de esteroles en el hongo Pleurotus sajor-caju. Revista Colombiana de Química, 34(2): 117-125.
Rocha O., Nobre C., Dominguez A., Torres A., Faria N., Rodrigues L., Teixeira J.A., Ferreira E.C., Rocha I. (2009). A Dynamical
Model for the rmentative production of fructooligosaccharides, in 10th International Symposium on Process Systems
Engineering - PSE2009
, Rita Maria de Brito Alves C.A.O.d.N.a.E.C.B.J.E., Editor© 2009 Elsevier B.V. All rights reserved: Braga, Portugal. 1827-1833.
Rodríguez I., Piñeros Y. (2007). Producción de complejos enzimáticos celulolíticos mediante el cultivo en fase sólida de
Trichoderma sp. sobre los racimos vacíos de palma de aceite como sustrato. Revista de la Facultad de química farmacéutica,
14(2): 35-42.
Rodríguez J., Ferraz A., Mello M.P. (2003). Role of metals in wood biodegradation. En: Wood Deterioration and Preservation -
Advances in our Changing World. Series A.S. (Ed.). Goodell, B., Nicholas, D.D., Schultz, T.P. (Eds.): Washington. pp. 154-
174.
Rodríguez N. (2003a). Aprovechamiento de los residuos sólidos generados en el cultivo e industrialización del café para la
producción de hongos comestibles y medicinales (Utilization of solid wastes generated during the cultivation and

199
industrialization of coffee for production of edible and medicinal mushrooms, in Spanish).Ph.D. Thesis. Universidad
Politécnica de Valencia: Valencia. 140 p.
Rodríguez N. (2003b). Aprovechamiento de los residuos sólidos generados en el cultivo e industrialización del café para la
producción de hongos comestibles y medicinales.Postgrado. Universidad Politécnica de Valencia: Valencia. 140 p.
Rodríguez N., Jaramillo C. (2005a). Cultivo de hongos comestibles del género Pleurotus sobre residuos agrícolas de la zona
cafetera. Centro Nacional de Investigaciones de Café, 23: 34-40.
Rodríguez N., Jaramillo C. (2005b). Cultivo de hongos comestibles del género Pleurotus spp. sobre residuos agrícolas de la zona
cafetera. Boletín Técnico Cenicafé, 27: 54 pp.
Rodríguez N., Jaramillo C. (2005c). Cultivo de hongos comestibles del género Pleurotus sobre residuos agrícolas de la zona
cafetera. Boletín Técnico Cenicafé, 23: 34-40.
Rodríguez V.N. (1993). El cultivo de hongos comestibles sobre desechos agroindustriales, Cenicafé: Chinchiná. 1-2 p.
Rodriguez-Couto S., Toca-Herrera J.L. (2007). Laccase production at reactor scale by filamentous fungi. Biotechnology advances,
25: 558.569.
Rodríguez-Couto S., Sanromán M.A. (2005). Application of solid-state fermentation to ligninolytic enzyme production.
Biochemical Engineering Journal 22 211-219.
Rodríguez-Couto S., Toca-Herrera J.L. (2007). Laccase production at reactor scale by filamentous fungi. Biotechnology
Advances, 25: 558-569.
Rodríguez-Palenzuela P., García J., de Blas C., Madrid. D.B.U.P.d., Madrid. D.d.P.A.U.P.d. (2000). XIV Curso de
Especialización: Avances en nutrición y alimentación animal. En: Fibra soluble y su implicación en nutrición animal:
Enzimas y probióticos. Madrid. pp. 17p.
Rosale E., Couto S.R., Sanromán A. (2007). Increased laccase production by Trametes hirsuta grown on ground orange peelings.
Enzyme and Microbial Technology, 40 1286-1290.
Ruíz A., Varela A. (2006). Nuevos registros de Aphyllophorales (basidiomicota) en bosque montano húmedo y de niebla de
Colombia. Caldasia 28(2): 259-266.
Russell R., Paterson M. (2008). Cordyceps – A traditional Chinese medicine and another fungal therapeutic biofactory.
Phytochemistry, 69: 1469-1495.
Sáez A., Florez L., Cadavid A. (2002). Caracterización de una cepa nativa de Aspergillus niger y evaluación de la producción de
ácido cítrico. Revista Universidad EAFIT, (128): 33-42.
Sánchez O.J., Cardona C.A. (2007). Producción de Alcohol Carburante: Una Alternativa para el Desarrollo Agroindustrial.
Universidad Nacional de Colombia: Manizales. 386 p.
Sánchez O.J., Cardona C.A. (2008). Trends in biotechnological production of fuel ethanol from different feedstocks. Bioresource
Technology, 99: 5270-5295.
Sangsurasak P., Nopharatana M., Mitchell D. (1996). Mathematical modeling of the growth of filamentous fungi in solid-state
fermentation. Journal Science Industrial Resources 55: 333-342.
Sarangi I., Ghosh D., Bhutia S., Mallick S., Maiti T. (2006). Anti-tumor and immunomodulating effects of Pleurotus ostreatus
mycelia-derived proteoglycans. International Immunopharmacology, 6: 1287-1297.
Sarikaya A., Ladisch M. (1997). An Unstructured Mathematical Model for Growth of Pleurotus ostreatus on Lignocellulosic
Material in Solid-State Fermentation Systems. Applied Biochemistry and Biotechnology, 62: 72-85.
Schulze B., Wubbolts M.G. (1999). Biocatalysis for industrial production of fine chemicals. Chemical Biotechnology, 10: 609-
615.
Shen Q., Royse D. (2002). Effects of genotipes of maitake (Grifola frondosa) on biological efficiency, quality and crop cycle
time. Applied Microbiology Biotechnology, 58: 178-182.
Shi J., Sharm-Shivappa R.R., Chinn M., Howell N. (2009). Effect of microbial pretreatment one nzymatic hydrolysis and
fermentation of cotton stalks for ethanol production. Biomass and Bioenergy, 33: 88-96.
Shi J., Sharma-Shivappa R.R., Chinn M.S. (2012). Interactions between fungal growth, substrate utilization and enzyme
production during shallow stationary cultivation of Phanerochaete chrysosporium on cotton stalks. Enzyme and Microbial
Technology, 51: 1- 8.
Shinners-Carnelley T.C., Szpacenko A., Tewari J.P., Palcic M.M., Cho N.S. (2002). Enzymatic activity of Cyathus olla during
solid state fermentation of canola roots. Phytoprotection 83: 31-40.
Shu C., Lung M. (2003). Effect of pH on the production and molecular weight distribution of exopolysaccharide by Antrodia
camphorata in batch cultures
. Process Biochemestry, 39: 931-937.
Silva E.M., Martins S.F., Milagres A.M.F. (2008). Extraction of manganese peroxidase produced by Lentinula edodes.
Bioresource Technology, 99: 2471-2475.
Soares G., De Amorin M., Costa-Ferreira M. (2001). Use of laccase together with redox mediators to decolourize Remazol
Brilliant Blue R. Journal Biotechnology, 89: 123-129.
Sohail M., Siddiqi R., Ahmad A., Khan S.A. (2009). Cellulase production from Aspergillus niger MS82: effect of temperature and
pH. New Biotecchnology, 25(6): 437-441.
Solomon E.I., Sundaram U.M., Machonkin T.E. (1997). Multicopper oxidases and oxygenases. Chemical Reviews, 96: 2563-
2605.
Somogyi M. (1945). A new reagent for the determination of sugars. Journal Biology Chemistry, 160: 61-73.

200
Sriyudthsak K., Shiraishi F. (2010). Investigation of the performance of fermentation processes using a mathematical model
including effects of metabolic bottleneck and toxic product on cells. Mathematical Biosciences, 228: 1-9.
Stames P. (1993). Growing gourmet and medicinal fungi Ten Speed Press and Mycomedia: Hong Kong. 456 p. p.
Stead P., Marley H., Mahmoudian M., Webb G., Noble D., Ip Y., Piga E., Rossi T., Roberts S. (1996). Efficient procedures for
the largescale preparation of (1S,2S)-trans-2-methoxycyclohexanol, a key chiral intermediate in the synthesis of tricyclic b-
lactam antibiotics. Dawson MJ: Tetrahedron - Asymmetry, 7: 2247-2250.
Sun Y., Cheng J. (2002). Hydrolysis of lignocellulosic materials for ethanol production: a review. Bioresource Technology, 83: 1-
11.
Švagelj M., Berovič M., Boh B., Menard A., Simčič S., Wraber B. (2008). Solid-state cultivation of Grifola frondosa (Dicks: Fr)
S.F. Gray biomass and immunostimulatory effects of fungal intra- and extracellular β-polysaccharides. New Biotechnology,
25: 150-156.
Takano M., Nishida, A., Nakamura, M. (2001). Screening of wood-rotting fungi for kraft pulp bleaching by the Poly R
decolorization test and biobleaching of hardwood kraft pulp by Phanerochaete crassa WD 1694. Journal of Wood Science,
47: 63-68.
Takasaki Y. (1976). Production and utilization of B-amylase and pullulanase from B. cereus var. mycoides. Agriculture Biology
Chemistry, 40: 1515-1525.
Tang Y.-J., Zhu L.-W., Li H.-M., Li D.-S. (2007). Submerged Culture of Mushrooms in Bioreactors – Challenges, Current State-
of-the-Art, and Future Prospects. Food Technology and Biotechnology, 45(3): 221-229.
Tang Y.J., Zhong J.J. (2002). Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma
lucidum, grown on lactose in a bioreactor. Biotechnology Letters, 24: 1023-1026.
Tao Y., Zhang L., Cheung P. (2006). Physicochemical properties and antitumor activities of watersoluble native and sulfated
hyperbranched mushroom polysaccharides. Carbohydrates Resources, 341: 2261-2269.
Tavares A., Coelho M., Coutinho J., Xavier A. (2005). Laccase improvement in submerged cultivation: induced production and
kinetic modelling. Journal of Chemical Technology and Biotechnology, 80: 669-676.
Tengerdy R.P., Szakacs G. (2003). Bioconversion of lignocellulose in solid substrate fermentation. Biochemical Engineering
Journal, 13: 169-179.
Thurston C. (1994). The structure and function of fungal laccases. Microbiology-UK, 140: 19-26.
Tišma M., Sudar M., Raĉki S., Zelic B. (2010). Mathematical model for Trametes versicolor growth in submerged cultivation.
Bioprocess and Biosystems Engineering, 33: 749-758.
Tlecutil-Beristain S., Sánchez C., Loera O., Robson G., Díaz-Godínez G. (112). Laccases of Pleurotus ostreatus observed at
different phases of its growth in submerged fermentation: production of a novel laccase isoform. Micologycal Research 1080
- 1084.
Tobin J.M., White C., Gadd C.M. (1994). Metal accumulation by fungi—applications in environmental biotechnology. Journal of
Industrial and Microbiology 13: 126-130.
Tolan J., Olson D., Dines R. (1996). Survey of mill usage of xylanase. En: Enzymes for pulp and paper processing (Jeffries, TW.
Viikari L). Society A.C. (Ed.). Washington DC. pp. 25-35.
Tomme P.H., Van Tilbeurgh G., Petterson J., Van Damme J., Vanderker-Ckhove J., Knowles T., Teeri, Claeyssens M. (1988).
Studies of cellulolityc system of Trichoderma reesei QM 9414. Analysis of domain function in two cellobiohydrolases by
limited proteolysis. Europe Journal Biochemestry, 170: 575-581.
Tsiklauri N.D., Khardziani T.S., Kachlishvili E.T., Elisashvili V.I. (1999). Cellulase and xylanase activities of higher
basidiomycetes during bioconversion of plant raw materials depending on the carbon source in the nutrient medium. Applied
Biochemistry and Microbiology, 35: 291-295.
Tsukagoshi S., Hashimoto Y., Fujii G., Kobayashi H., Nomoto K., Orita K. (1984). Krestin (PSK). Cancer Treatment review, 11:
131-155.
Tuor U., Wariishi H., Schoemaker H., Gold M.H. (1992). Oxidation of phenolic arylglycerol beta-aryl ether lignin model
compounds by manganese peroxidase from Phanerochete chrysosporium: oxidative cleavage of an alpha-carbonyl mode
compoundl. Biochemistry, 31: 4986-4995.
Tychanowicz G.K., Zilly A., Marques de Souza C.G., Peralta R.M. (2004). Decolourisation of industrial dyes by solid-state
cultures of Pleurotus pulmonarius. Process Biochemistry, 39: 855-859.
Ünyayar A., Sik S. (1998). Phanerochaete chrysosporium and Funalia trogii for the degradation of cotton stalk and their laccase,
peroxidase, ligninase and cellulase enzyme activities under semi-solid state conditions. Turkish Jornal Biology, 22: 287-298.
Uribe-López A.M. (1983). Hongos comestibles: otro producto de las plantaciones de coníferas no explotado en Colombia
[Agaricales, Aphyllophorales, Lycoperdales, Pezizales]. ed. Agronomía F.d. Universidad Nacional de Colombia, Sede
Medellín: Medellín (Colombia). 73 p.
Valbuena D.C., Alzate C.J. (2007). Evaluación de la patogenicidad de los hongos Bauveria basiana y Metarhizium anisopliae en
el control de la garrapata del ganado Rhipicephalus micraplus en su fase parasítica en su estado larval y ninfal.Trabajo de
grado. Microbiología Agrícola y Veterinaria, Javeriana: Bogotá. 144 p. p.
Van de Lagemaat J., Pyle D.L. (2005). Modelling the uptake and growth kinetics of Penicillium glabrum in a tannic acid-
containing solid-state fermentation for tannase production. Process Biochemistry, 40: 1773-1782.
Vares T., Kalsi M., Hatakka A. (1995). Lignin peroxidases, manganese peroxidases and other ligninolytic enzymes produced by
Phlebia radiata during solid state fermentation of wheat straw. Applied and Environment Microbiology 61: 3515-3520.

201
Vasco-Palacios A.M., Franco-Molano A.E., López-Quintero C.A., Bohechout T. (2005). Macromicetes (ascomicota,
basidiomicota) de la región del medio Caquetá, departamentos de Caquetá y Amazonas (Colombia). Biota Colombiana, 6(1):
127-140.
Vasic-Racki D., Kragl U., Liese A. (2003). Benefits of enzyme kinetics modelling. Chemical and Biochemical Engineering
Quarterly 17: 7-18.
Vera D.F., Pérez H., Valencia H. (2002). Aislamiento de hongos solubilizadores de fosfatos de la rizosfera de arazá (Eugenia
stipitata, Myrtaceae). Acta Biológica Colombiana 7(1): 33-39.
Vetter J. (2007). Chitin content of cultivated mushrooms Agaricus bisporus, Pleurotus ostreatus and Lentinula edodes. Food
Chemistry, 102: 6-9.
Viccini G., Mitchell D., Boit S., Gern J., da Rosa A., Costa R. (2001). Analysis of growth kinetic profiles in solid-state
fermentation. Food Technology and Biotechnology Advances, 39: 271-294.
Visser J., Beldman G., Kusters–van Someran M.A., Voragen A.G.J. (1991). International Conference of Xylans and Xylanases
Netherlands: Elsevier. pp. 89-94.
Vroemen A.J. (2002). Glucose oxidase. En: Handbook of food enzymology. Witaker J.R., Voragen A.G., Wong D.W.S. (Eds.).
Marcel Dekker, INC: New York. pp. 425-432.
Vrsalovic Presecki A., Findrik Z., Zelic B. (2006). Modeling of the biotransformation processes. Chemical and Biochemical
Engineering Quarterly 20: 227-241.
Wade L.G. (1993). Química Orgánica. 2 ed, ed. Educación P. Debra Wechsler: México. 1312 p.
Wagner R., Mitchell D.A., Sassaki G.L., Lopes de Almeida A., Berovic M. (2003). Current techniques for the cultivation of
Ganoderma lucidum for the production of biomass, ganoderic acid and polysaccharides. Food Technology and
Biotechnology, 41: 371-382.
Wainwright M. (1988). Metabolic diversity of fungi in relation to growth and mineral cycling in soil—a review. Transactions of
the British Society of Mycology, 90: 159-170.
Walkley A., Black I. (1934). An estimation of the Degtjareft method for determining of soil organic matter and a proposed
modification of chromic acid titration method. Soil Science, 37: 29-38.
Wasser S. (2002). Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Applied Microbiology
and Biotechnology, 60: 258-274.
Wasser S.P., Weis A.L. (1999). Medicinal properties of substances occurring in higher Basidiomycete mushrooms: current
perspective (review). International Journal Medicine Mushrooms, 1: 31-62.
Weinberg Z.G. (2000). Biotechnology in developing countries: opportunities in solid state fermentation applied in the agricultural
industry. Internation Journal of Biotechnology, 2(4): 364-373.
Whittaker R.H. (1969). New Concepts of Kingdoms of Organisms. Science, 163: 150-160.
Williams E., Todd N., Rayner A. (1981). Propagation and development of fruit bodies of Coriolus versicolor. Transactions of the
British Mycological Society, 77: 409-414.
Wong D.W.S. (2009). Structure and Action Mechanism of Ligninolytic Enzymes. Applied Biochemestry and Biotechnology, 157:
174-209.
Wood T.M., García C.V. (1994). Enzymes and mechanisms involved in microbial cellulolysis. Kluwer Academic publishers:
Netherlands.
Wu J., Cheung P., Wong K., Huang N. (2004). Studies on submerged fermentation of (Fr.) Singer. Part 2. Effect of carbonto-
nitrogen ration of the culture medium on the content and composition of the mycelial dietary fibre. Food Chemistry, 85: 101-
105.
Wu J., Ding Z.-Y., Zhang K.-C. (2006). Improvement of exopolysaccharide production by macro-fungus Auricularia auricula in
submerged culture. Enzyme and Microbial Technology, 39: 743-749.
Wyman C.E., Dale B.E., Elander R.T., Holtzapple M., Ladisch M.R., Lee Y.Y. (2005). Coordinated development of leading
biomass pretreatment technologies. Bioresource Technology, 96: 1959-1966.
Xiao J.-H., Xiao D.-M., Xiong Q., Liang Z.-Q., Zhong J.J. (2010). Nutritional requirements for the hyperproduction of bioactive
exopolysaccharides by submerged fermentation of the edible medicinal fungus Cordyceps taii. Biochemical Engineering
Journal, 49(2): 241-249.
Xiao J.H., Chen D.X., Wan W.H., Hu X.J., Qi Y., Liang Z.Q. (2006). Enhanced simultaneous the production of mycelia and
intracellular polysaccharide in submerged cultivation of Cordyceps jiangxiensis using desirability functions. Process
Biochemestry, 41: 1887-1893.
Xie J., Sun X., Ren L., Zhang Y.Z., Wu Kung S. (2001). Studies in lignocellulolytic enzymes production and biomass degradation
of Pleurotus sp.2 and Trametes gallica in wheat straw cultures. Chemical Engineering Journal and the Biochemical
Engineering Journal, 17: 575-578.
Xu H., Sun L.-P., Shi Y.Z., Wu Y.W., Zhang B., Zhao D.Q. (2008). Optimization of cultivation conditions for extracellular
polysaccharide and mycelium biomass by Morchella esculenta As51620 Biochemical Engineering Journal, 39: 66-73.
Yachmenev V., Condon B., Klasson T., Lambert A.J. (2009). Acceleration of the enzymatic hydrolysis of corn stover and sugar
cane bagasse celluloses by low intensity uniform ultrasound. Biobased Material Bioenergy, 3: 25-31.
Yalin W., Ishurd O., Cuirong S., Yuanjiang P. (2005). Structure analysis and anti-tumor activity of (1→3)-β-D-glucans
(cordyglucans) from the mycelia of Cordyceps sinensis. Planta Medica, 71: 381-384.

202
Yang B.-K., Gu Y.-A., Jeong Y.-T., Jeong H., C-H. S. (2007). Chemical characteristics and immune-modulating activities of exo-
biopolymers produced by Grifola frondosa during submerged fermentation process. International Journal of Biological
Macromolecules, 41: 327-333.
Yang F.-C., Ke Y.-F., Kuo S.-S. (2000). Effect of fatty acids on the mycelial growth and polysaccharide formation by Ganoderma
lucidum in shake flask cultures. Enzyme and Microbial Technology, 27: 295-301.
Zabel R.A., Morrel J.J. (1992a). Decay and its Prevention. Academic Press: San Diego. 534 p. p.
Zabel R.A., Morrel J.J. (1992b). Decay and its Prevention. Academic Press: San Diego, California.
Zaks A. (2001). Industrial biocatalysis. Chemical Biology, 5: 130-136.
Zanchetta P., Lagarde P.N., Guezennec J. (2003). A new bone-healing material: A hyaluronic acid-like bacterial
exopolysaccharide. Calcified Tissue International, 72: 74-79.
Zapata P., Rojas D., Carlos Fernández C., Ramírez D., Restrepo G., Orjuela V., Arroyave M., Gómez T., Atehortúa L. (2007).
Producción de biomasa y exopolisacáridos de Grifola frondosa bajo cultivo sumergido utilizando fuentes de carbono no
convencionales. Revista EIA, ISSN 1794-1237, (7): 137-144.
Zapata P., Rojas D., Atehortúa L. (2012). Production of Biomass, Polysaccharides, and Ganoderic Acid using Non-conventional
Carbon. International Journal of Medicinal Mushrooms, 14(2): 197-203.
Zelic B., Vasic Racki D., Wandrey C., Takors R. (2004). Modeling of the pyruvate production with Escherichia coli in a fed-
batch bioreactor. Bioprocess and Biosystems Engineering, 26: 249-258.
Zhang P., Himmel M.E., Mielenz J.R. (2006a). Outlook for cellulase improvement: Screening and selection strategies.
Biotechnology Advances 24: 452-481.
Zhang Y.-H.P., Himmel M.E., Mielenz J.R. (2006b). Outlook for cellulase improvement: Screening and selection strategies.
Biotechnology Advances, 24: 452-481.
Zhao J., Janse B.J. (1996). Comparison of H2O2-producing enzymes in selected white rot fungi. FEMS Microbiology Letters, 139:
215-221.
Zhao J., Koker T., Janse B.J. (1996). Comparative studies of lignin peroxidases and manganese-dependent peroxidases produced
by selected white rot fungi in solid media. FEMS Microbiology Letters, 143: 393-399.
Zhao X.R., Lin Q., Brookes P.C. (2005). Does soil ergosterol concentration provide a reliable estimate of soil fungal biomass?
Soil Biology and Biochemistry, 37: 311-317.
Zhong J.J., Tang Y.J. (2004). Submerged cultivation of medicinal mushrooms for production of valuable bioactive metabolites.
Advances in Biochemical Engineering/Biotechnology, 87: 25-59.
Zhu S., Wu Y., Yu Z., Wang C., Yu F., Jin S., Ding Y., Chi R., Liao J., Zhang Y. (2006). Comparison of three
microvawe/chemical pretreatment processes for enzymaric hydrolyisis of rice straw. Biosystem Engineering, 93: 279-283.
Zhuang O., Mizuno T. (1999). Biological responses from Grifola frondosa (Dick.:Fr.) S.F. Gray-Maitake
(Aphyllophoromycetideae). Internation Journal medicinal mushrooms, 1: 317-324.