o3-diversité morpho-fonctionnelle des organismes -...
TRANSCRIPT

O3-Diversité morpho-fonctionnelle des
organismesNutrition
& croissance

Ce cours ne traite ni des Angiospermes ni des animaux,
car ils sont étudiés en détail dans des cours spécifiques.

1. Les organismes pluricellulaires sont formés de cellules spécialisées,
dont celles qui assurent la croissance et la nutrition
1.1. Des cellules différenciées ou non assurent la croissance des pluricellulaires

(Ulva lactuca)
http://www.alternaturel.fr/les-champignons/
11.1. Quand toutes les cellules différenciéesparticipent à la croissance, on parle decroissance diffuse

11.2. Une cellule apicale indifférenciée assure la croissance indéfinie chez les autres algues* pluricellulaires

11.3. Un ensemble de cellules indifférenciéesassure la croissance indéfinie desAngiospermes (voir cours Angiospermes), etla croissance finie des Métazoaires (pourinfo)

1.2. Le mode de croissance détermine le plan d’organisation, qui est en lien avec le mode de nutrition des pluricellulaires
12.1. La croissance diffuse ne permet pas la formation d’organes,mais étale l’appareil végétatif dans le milieu (Ulve, champignons*)
http://www.alternaturel.fr/les-champignons/

12.2. Une croissance parméristèmes forme des organes ettissus spécialisés dans chaqueétape de la nutrition (voir coursAngiospermes)
La nutrition des Angiospermes se fait par des échanges inter-organes


• Les organismes à croissance diffuse peuvent explorer le milieu par leur croissance dans le milieu. C’est surtout le cas des champignons, qui absorbent les nutriments du milieu.
• Une croissance par une apicale ou un méristème permet de construire un appareil végétatif plus élaboré, bien étalé dans le milieu, ce qui augmente la surface en contact avec la lumière.
Le mode de croissance des organismes détermine leur plan d’organisation et, dans une certaine limite, leur mode de nutrition.

1. La croissance des pluricellulaires est peut être assurée par
a) des cellules différenciées
b) des cellules indifférenciées
2. les modalités de la croissance détermine
a) la morphologie de l'organisme
b) le volume de milieu qu'il peut explorer pour sa nutrition
c) son mode de nutrition

Photosynthèse
Source d’énergie = lumière
Source d’azote = nitrate de
l’eau ou N2 de l’air
Production de molécules organiques
(sucres, aa, nt)
Source d’électrons = eau
Source de carbone = CO2
2. En milieu aquatique, la nutrition des Autotrophespluricellulaires repose sur la diffusion entre le milieuextérieur et intérieur, et parfois sur une diffusioninterne
MILIEU
DIFFUSION INTERNE
DIFFUSION
Autotrophe pluricellulaire aquatique

2.1.Tous les autotrophes aquatiques possèdent des surfaces photosynthétiques
21.1. Tout organisme photosynthétique possède de la chlorophylle a
Autres pigments

21.2. Les pigments associés à la chlorophylle a dépendent des groupes et donnent la couleur aux algues
http://www.ac-rennes.fr/pedagogie/svt/travaux/algues/pages/lumiere4.htm

Légendez les photos ci-contre de l’algue brune-jaune, et
établissez la particularité des organites de ce groupe
Noyau
Pyrénoïde (RubisCO)
http://www.snv.jussieu.fr/bmedia/Chloroplaste/giraud.htm
21.3. Les chloroplastes diffèrent par le nombre de membranes externes, l’agencement des thylakoïdes et la présence ou non de pyrénoïde

• Les thylacoïdes sont associées par trois.
• Un thylacoïde traverse le pyrénoïde.
• L'enveloppe du chloroplaste est formée de quatre membranes. Les deux plus internes (en
bleu) correspondent à l'enveloppe directe du chloroplaste, les deux plus externes (en rouge)
sont en relation avec l'enveloppe nucléaire. Entre ces deux groupes de deux membranes.
• Cette particularité est mise en relation avec une origine endosymbiotique (endosymbiose
secondaire du chloroplaste).
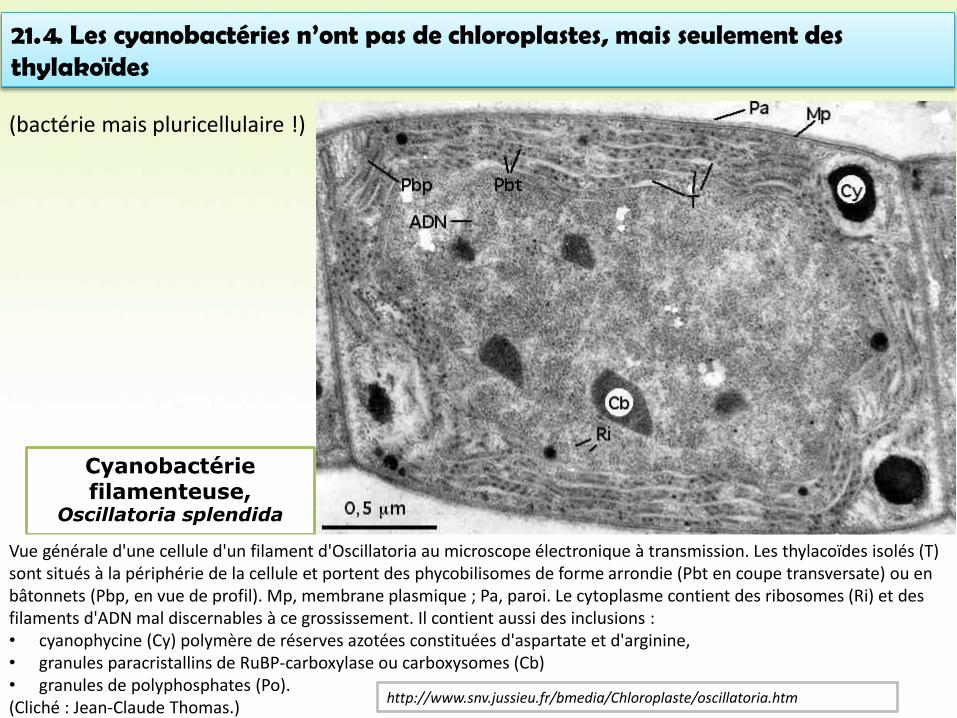
Cyanobactérie filamenteuse,
Oscillatoria splendida
Vue générale d'une cellule d'un filament d'Oscillatoria au microscope électronique à transmission. Les thylacoïdes isolés (T) sont situés à la périphérie de la cellule et portent des phycobilisomes de forme arrondie (Pbt en coupe transversate) ou en bâtonnets (Pbp, en vue de profil). Mp, membrane plasmique ; Pa, paroi. Le cytoplasme contient des ribosomes (Ri) et des filaments d'ADN mal discernables à ce grossissement. Il contient aussi des inclusions :• cyanophycine (Cy) polymère de réserves azotées constituées d'aspartate et d'arginine,• granules paracristallins de RuBP-carboxylase ou carboxysomes (Cb)• granules de polyphosphates (Po). (Cliché : Jean-Claude Thomas.)
http://www.snv.jussieu.fr/bmedia/Chloroplaste/oscillatoria.htm
21.4. Les cyanobactéries n’ont pas de chloroplastes, mais seulement des thylakoïdes
(bactérie mais pluricellulaire !)

2.2. Le pyrénoïde des algues réduit la photorespiration, comme le métabolisme en C4 de certaines Angiospermes
• le CO2 diffuse 10 000 fois plus lentement dans l'eau que dans l'air,
• le CO2 dissous s'équilibre avec le bicarbonate HCO3–, qui n’est pas reconnu
par la RubisCO
Conséquence : La RuBisCO des algues marines sans pyrénoïde ne fonctionne
qu'au quart de sa vitesse maximum.
Le problème : en milieu aquatique, le CO2 est peu disponible pour la photosynthèse
La Spirogyre, algue d’eau douce
pluricellulaire

Solution :
Le pyrénoïde, qui est un amas de
grains d’amidon et de RubisCO autour
d’un thylakoïde central.
Le CO2 est concentré près de la
RubisCO par:
1. des transports actifs d’ions HCO3-
2. L’anhydrase carbonique, présente
dans le thylakoïde, qui retransforme
ces ions bicarbonates en CO2, qui
lui, est reconnu par la RubisCO

NB: ici une seule cellule est dessinée, mais le
pyrénoïde est présent chez des Algues
pluricellulaires (comme ci-dessus)
Les pyrénoïdes sont des structures très
plastiques dont l'apparence est fortement
corrélée à l'activité de concentration du dioxyde
de carbone : Dans un environnement riche en
CO2, les pyrénoïdes sont petits et leur matrice
manque de structure
Le pyrénoïde concentre le CO2 chez les algues aquatiques

C4 Hatch & Slack
PRINCIPE : • Réduction de la photorespiration, qui est l’oxydation de
la matière organique par la RubisCO (d’où son nom! Ribulose bi phosphate +Carboxylase+ Oxygénase ou oxydase)
• La photorespiration est proportionnelle au rapport O2/CO2, car ces deux molécules sont les substrats de la RubisCO
MODALITE :• Fixation du CO2 par une enzyme à très forte affinité
pour le CO2 (PEPcarboxylase)• Transport vers une autre cellule sous forme d’une
molécule à 4 carbones =C4 (malate)• Pas de production d’oxygène à proximité de la RubisCOECOLOGIE : • Vu le coût énergétique de la concentration du CO2 dans
le mésophylle, le métabolisme en C4 ne confère un avantage sélectif qu’en milieu chaud et sec = stomates souvent fermés
Expliquez rapidement le principe du métabolisme
en C4 (sans se perdre dans les détails!)

Le pyrénoïde a le même rôle que le métabolisme en C4

2.3. Tous les autotrophes aquatiques possèdent des surfaces d’échanges avec le milieu extérieur
23.1. Pour la plupart des autotrophes aquatiques, c’est toute la surface de l’organisme qui est la surface d’échange avec le milieu

23.2. Certaines algues accumulent les ions HCO3- grâce à des transports actifs et des enzymes (anhydrase carbonique)

Anabaena sp.
Nitrogénase
O2
La
nitrogénase
est inhibée
par l’O2
23.3. Les cyanobactéries ont deshétérocystes spécialisés dans lafixation de l’azote de l’air
Eau de mer du grand large, très pauvre en nitrates
cyclique
Les détails
ne sont pas
à connaître

stromatolites
Ces stromatolithes sont trouvés dans des
couches qui varient en âge de 3,5 Ga à
l'Actuel.

2.4. Certains autotrophes possèdent des structures spécialisées dans les échanges internes
24.1. Les cyanobactéries ont des plasmodesmes entre leurs cellules
http://garciajeanlouis9051.perso.neuf.fr/gb_photosynthetiques.htmlThylakoïdes

24.2. Les algues brunes possèdent des cellules facilitant conduction des sucres

Le métabolisme de l’autotrophie au carbone des pluricellulaires est la photosynthèse. Tous les autotrophes possèdent des thylakoïdes avec de la chlorophylle a et des enzymes de réduction de l’azote minéral.
• Il existe plusieurs structures pour les chloroplastes, qui dérivent de l’évolution. Cependantleur fonctionnement est similaire. Ce qui change ce sont les longueurs d’ondes captées parles pigments et la présence éventuelle d’un pyrénoïde.
• Le pyrénoïde est un amas de RubisCO et d’amidon autour d’un thylakoïde remplid’anhydrase carbonique. Celle-ci produit du CO2 à partir des ions HCO3- à proximité de laRubisCO, ce qui diminue son activité oxygénase et donc la photorespiration.
• Cependant l’oxygène produit par la photosynthèse détruit la matière organique et inhibe laréduction des nitrates. La séparation de la production d’oxygène et des deux autresmétabolismes dans des cellules différentes évite ce problème. On trouve cela tant chez leseucaryotes (C4 pour C + réduction des nitrates dans la racine) que chez les procaryotes(hétérocystes pour N).
• Les échanges avec le milieu extérieur se font par toute la surface des algues et descyanobactéries. Les cyanobactéries fixent en plus l’azote de l’air. Certaines algues ont destransporteurs actifs d’ions HCO3-, ce qui leur permet de . Elles possèdent aussi desanhydrases carboniques qui le transforment en CO2.

3. Un autotrophe peut être (trouvez la réponse fausse)a) une algue rouge, car elle a aussi de la chlorophylle verte.b) une bactérie qui a des thylakoïdesc) seulement un organisme à chloroplastesd) une algue avec un pyrénoïdee) une algue avec des chloroplastes à 4 membranes
4. la photorespirationa) est une voie d'oxydation de la matière organique, indépendante de la photosynthèseb) dépend directement de la température chez les plantesc) est limitée par le pyrénoïde de certaines algues
5. la teneur en CO2 est limitante pour la photosynthèsea) en milieu aquatique car le CO2 se transforme en HCO3-b) en milieu chaud et sec pour les plantes en C3
6. Les échanges internes chez les autotrophes pluricellulaires se fonta) par des cellules spécialisées chez les algues brunesb) par le xylème et le phloème chez les Angiospermesc) par des plasmodesmes chez des cyanobactéries et les Angiospermes

3. En milieu aérien, la nutrition hétérotrophe des champignons* repose sur un mycélium étendu et l’absorbotrophie de substrats morts ou vivants
Eumycètes
Oomycètes
Clades=groupes phylogénétiques
Champignons
Groupe écologique

3.1.Les champignons* explorent le milieu par une croissance importante de leur mycélium diffus et ramifié
Richard
Hyphe en croissance (Robert)

3.2. La paroi des champignons libres impose l’absorbotrophie, souvent précédée d’une exodigestion
L'absorbotrophie est une forme de régime alimentaire où l'organisme se nourrit en absorbant les nutriments à travers la paroi extérieure de son corps.
Champignons: absorption de petites molécules à travers la paroi de chitine puis la membrane plasmique (transporteurs)

L’exodigestion du Penicillium : le mode d’affinage des fromages bleus
Moisissure du seigle
Lait cru

Montrez que le Penicillium réalise une exodigestion qui change le goût et la texture
Souche de
Champignon


3.3. Les champignons peuvent digérer des substrats morts ou vivants
33.1. La nutrition à partir de substrats morts est de la saprotrophieou de la nécrotrophie parasite
• Un organisme est dit saprophyte (sapros: décomposé, phuton: plante) s'il est
capable de se nourrir de matière organique morte, par absorption après une
éventuelle digestion externe. (sinon ils ne seraient que des détritivores) Ex: humus,
arbre mort
• Beaucoup de saprophytes sont des décomposeurs présents dans le sol ->
dégradent la matière organique -> rôle clé dans les cycles de la matière
Pleurote bleu d'huître Pleurotus ostreatus, Pleurotacée sur un arbre
fruitier abattu (Woluwe Saint-Pierre, Province de Brabant, Belgique -
24/11/1993 - Diapositive originale réalisée par Eric Walravens).
Agaric sur meule avec fumier

nécrotrophie parasite
Un nécrotrophe désigne un parasitoïde, un organisme qui consomme son hôte une
fois mort. (On appelle « parasitoïdes » les organismes qui, au cours de leur développement,
tuent systématiquement leur hôte, ce qui les fait sortir du cadre du parasitisme au sens strict.)
les champignons nécrotrophes qui tuent leur hôte et se nourrissent des matières
mortes, comme par exemple Botrytis cinerea = pourriture grise de la vigne.
Sur les inflorescences : l'attaque commence par la libération de
toxines qui produisent une nécrose sur le pédoncule, qui peut
provoquer la chute de la grappe. Puis la grappe est décomposée
au sol. -> nuisible pour certains cépages
Utilité : Le champignon produit une
polyphénoloxydase, qui change le goût et
la couleur des vins. Cela permet d’obtenir
des vins plus liquoreux (sauternes,
coteaux-du-layon, monbazillac, etc.), on
parle alors de « pourriture noble ».

nécrotrophie parasite
Le cordyceps tue systématiquement son hôte insecte en détruisant son
cerveau, avant de se reproduire en digérant son hôte. (Forêt tropicale humide)
http://www.photomonde.fr/cordyceps-unilateralis-le-
champignon-qui-controle-le-cerveau-des-fourmis/

33.2. La nutrition à partir de substrats vivants estde la biotrophie, du mutualisme ou du parasitisme
1) Biotrophie= mode de nutrition caractéristique
de certains champignons et parasites qui se
nourrissent de cellules sans les détruire,
comme les mildious et rouilles (Oomycètes)
Plasmopara viticola, Midiou de la vigne
Pourquoi met-on
des rosiers près des
vignes?

2) MutualismeLe mutualisme est une interaction entre deux (ou plusieurs) espèces, dans laquelle
les organismes impliqués tirent tous les deux profit de cette relation. On parle alors
d'une interaction à bénéfices réciproques.
Le mutualisme peut être une relation obligatoire entre les individus impliqués
(symbiose) ou non (coopération)
Exemples de symbioses (voir plus loin)
Exemple de coopération: fourmi qui élève des champignons (Amérique du sud)
VégétauxMilieu humide et ventilé
Excréments de fourmis= engraisBactéries = lutte contre un autre
champignon pathogène
Enzymes de digestion de la cellulose absorbées par les fourmis
Renflements mangés par les fourmis
http://www.futura-sciences.com/magazines/nature/infos/dossiers/d/zoologie-fourmi-
secrets-fourmiliere-1404/page/12/
Leucoagaricus
gongylophorus

3) Parasitisme (sens strict = non biotrophe)
Le parasitisme est une relation biologique dont un des protagonistes (le parasite)
tire profit (en se nourrissant, en s'abritant ou en se reproduisant) aux dépens d'un
hôte.
Exemple de parasitisme avec destruction cellulaire : la tavelure du pommier-poirier
Elle est causée par un champignon ascomycète, causant des lésions noires ou brunes à la
surface des feuilles, des bourgeons ou des fruits. La maladie est favorisée par un climat humide
au moment du débourrement. La maladie tue rarement son hôte mais peut réduire
significativement la qualité et la production des fruits en l'absence de traitement par fongicide.Pb: les formes résistantes au traitement ; variétés très sensibles (Golden); 7 à 12 traitements de fongicides /an

3.4. Certains champignons forment des structures végétatives complexes qui permettent des symbioses
34.1. Un lichen est une association symbiotique entre une algue verte et unascomycète


endomycorhize à arbuscules et vésicules (herbacées)
endomycorhize à pelotons
ectomycorhize d’Angiospermes(arbres)
Ectomycorhize de Pinophyte
ectendomycorhize
Racine non mycorhizée (plantule ou Brassicacée!)
(Duhoux) : Différentes mycorhizes et
principaux types d’interfaces
34.2. Une ectomycorhize est une association symbiotique entre une racine d’arbreet un mycète

Duhoux
Manteau fongique
25 à 40% de la masse
totale des hyphes)
Réseau de Hartig
(entre les cellules végétales)
Hyphes extra-racinaires
Méristème
Apical
racinaire
Tissus conducteurs
L’ectomycorhize modifie la structure externe de la racine**

Association peu spécifique:
• 1 champi peut mycorhizer différents arbres
• 350 espèces de champi pour hêtre

Effet de la mycorhize sur la croissance de la plante


Une symbiose est une relation interspécifique à bénéfice mutuel



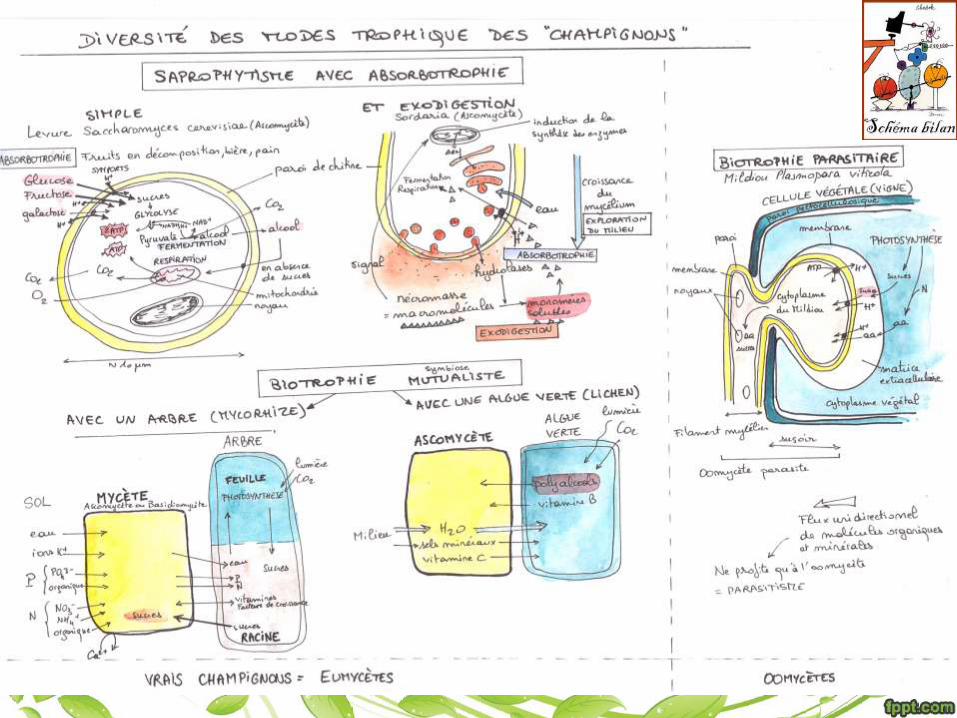

Les champignons sont de hétérotrophes dont la structure filamenteuse (en hyphes) leur permet d’explorer le milieu et de prélever de la matière organique à partir de substrats vivants ou morts. Ils réalisent souvent une digestion extracellulaire, puis une absorbotrophie.
Nous utilisons leur mode de nutrition :
- Pour la décomposition de la matière organique (gestion des déchets)
- Pour les enzymes libérées qui modifient leur environnement, en agroalimentaire (bière,
pain, fromages)
- Pour leur sous produits de fermentation : solvants de chimie!
• L’exodigestion et l’absorbotrophie sont dues à la présence d’une paroi.
• La croissance du mycélium dans le milieu assure le renouvellement des
substrats.
• Certains champignons forment des symbioses, avec des algues (lichen) ou des
plantes (mycorhizes)
• Bien que leur structure soit très simple, les champignons présentent une grande
diversité de mode trophique (libre/ parasite/ mutualiste; biotrophe/ nécrotrophe/
saprophyte)

7. Les champignons sont caractérisés par a) un appareil végétatif très étendu et en croissance permanenteb) l'absorption de petites molécules du milieuc) la possibilité de libérer des enzymes digestives dans le milieu
8. on peut qualifier de symbiosea) un lichenb) une mycorhizec) le mildiou

4. Les Unicellulaires assurent les mêmes fonctions que les pluricellulaires, mais dans une seule cellule
4.1.L’autotrophie est réalisable dans une seule cellule (Diatomée, euglène)

Les Euglènes existent sous deux formes
http://jean-jacques.auclair.pagesperso-
orange.fr/metabolisme/pigments.htm
Broyage +éthanol
A AB
B
En déduire le
mode trophique
des deux formes
d’Euglène


Les plastes des diatomées renferment de la chlorophylle a et de la chlorophylle c, ainsi que des pigments caroténoides qui masquent plus ou moins la couleur verte des chlorophylles. A la différence de ceux des plantes terrestres, les chloroplastes des diatomées ne sont donc pas verts mais jaunes, vert olive ou bruns.Ils ont des pyrénoïdes.
algues brunes, microscopiques (dont la taille varie entre 5μm et 500 μm) et unicellulaires appartenant à l’embranchement des Chromophytes.
Diatomées


Pas explicitement au
programme


4.2. Des unicellulaires réalisent aussi une absorbotrophie, avec éventuellement une exodigestion
42.1. Absorbotrophie avec exo et endodigestion chez un Eucaryote libre (Mycète) : Saccharomyces cerevisiae

-Tube 1 : surnageant de levures portées à agitation douce pendant 1h + Saccharose.-Tube 2 : même surnageant que dans le Tube 1 + Maltose-Tube 3 : culot de levures, broyés avec du sable + Saccharose-Tube 4 : Le même culot que dans le Tube 3 + MaltoseAvec le test du glucose, on montre que les tubes 1, 3 et 4 sont positifs
Interprétez ces résultats

http://jean-jacques.auclair.pagesperso-orange.fr/metabolisme/respmetab.htm
Saccharase = exoenzyme
-> est présente dans le
surnageant
Maltase = enzyme interne
-> l’hydrolyse du glucose se
fait dans la cellule


42.2. Absorbotrophie chez une Bactérie commensale diauxique : Escherichia coli
La bactérie peut utiliser du glucose et du
lactose. En présence de glucose, elle
consomme préférentiellement ce substrat.
La disparition du glucose induit la
transcription d’un ensemble de gènes
(opéron lactose) qui produit des enzymes
et un transporteur permettant l’utilisation
du lactose. Le rendement est moins bon,
ce qui ralentit la croissance de la
population.

Le trypanosome se nourrit par
absorption des nutriments du plasma à
travers sa membrane plasmique. Il
réalise aussi une endocytose à
récepteur des LDL du sang, suivie
d’une protéolyse.
42.3.Absorbotrophie chez un eucaryoteparasite, le trypanosome

42.4. Hétérotrophie absorbotrophe dans une symbioseFixatrice d’azote : Rhizobium/Fabacée


Hétérotrophie absorbotrophe,
dans une symbiose

4.3. L’hétérotrophie phagotrophe d’un unicellulaire comme la paramécie est similaire au mode de nutrition animal

Les unicellulaires peuvent réaliser tous les modes trophiques étudiés précédemment chez des pluricellulaires : autotrophie, hétérotrophie absorbotrophe (avec exodigestion éventuelle),

9. un phagotrophe est a) un organisme qui se nourrit par autophagieb) Saccharomyces cerevisiaec) une paramécie

• Les organismes pluricellulaires sont formés de cellules différenciées ou non, organisées ou non en tissus, voire en organes.
• En milieu aquatique, il existe des autotrophes pluricellulaires dont la nutrition repose sur la diffusion entre le milieu extérieur et l’organisme et, pour certains, sur des échanges intercellulaires au sein de l’organisme.
• En milieu aérien principalement, certains hétérotrophes au carbone sont constitués de filaments pluricellulaires (champignons), dont la croissance permet l’exploration du milieu et le prélèvement de matière organique par absorbotrophie, voire exodigestion, à partir de substrats morts (saprotrophie ou nécrotrophie parasite) ou vivants (biotrophie, mutualiste ou parasite).
• Les organismes unicellulaires assurent l’ensemble des fonctions au niveau d’une seule cellule (vie unicellulaire).
Des fonctions du vivant, comme la nutrition et croissance, peuvent être assurées dans des milieux différents et/ou avec des plans d’organisation différents, y compris à l’état unicellulaire. La réalisation de ces fonctions dépend de l’adaptation au milieu et/ou aux contraintes du plan d’organisation.