nuclear transfer into mouse zygotes
TRANSCRIPT
108 nature genetics • volume 24 • february 2000
We have previously reported a novel nuclear-transfer technique to clone
mice from adult somatic cells1,2 and late-passage embryonic stem cells3. Themethod entails the injection of a somatic-cell nucleus into an enucleated, unfertil-ized oocyte using a piezo-electricallydriven micromanipulator. Embryonicdevelopment is activated by incubationwith strontium chloride one to six hoursafter nuclear transfer.
Here we sought to determine if thiscloning method could be adapted to usezygotes as recipients for somatic cellnuclei. In parallel, we have investigated arelated issue: the developmental effect ofthe timing of oocyte activation relative tonuclear transfer. We microinjected nucleifrom cumulus cells of adult femaleB6D2F1 mice into enucleated, unfertilizedB6D2F1 oocytes1. Reconstructed oocyteswere activated with strontium chloride atone of the following time points: (i) one tothree hours after nuclear transfer (post-activation); (ii) immediately after nucleartransfer (immediate activation); or (iii)one hour before nuclear transfer (pre-activation). Additionally, other nucleiwere injected into zygotes after removal ofboth pronuclei (iv), which we estimate tobe five to six hours after the zygotes wereproduced following coital fertilization;this is the earliest point at which pronucleican be readily visualized for removal. Thefour experimental conditions were carriedout concurrently.
In a first series of experiments, chromo-somes of reconstructed oocytes (n=580)were examined morphologically byGiemsa staining4 just before the firstmitotic division, approximately 16 hoursafter nuclear transfer. The percentages ofreconstructed eggs lacking evident chro-
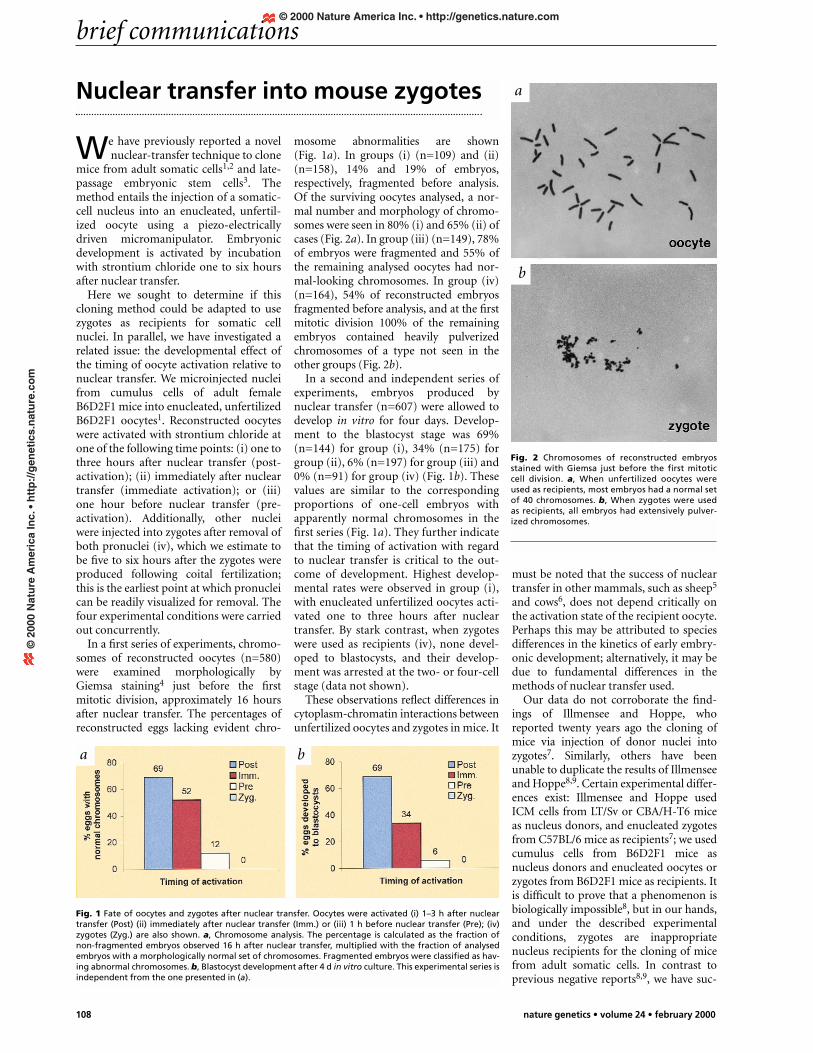
mosome abnormalities are shown(Fig. 1a). In groups (i) (n=109) and (ii)(n=158), 14% and 19% of embryos,respectively, fragmented before analysis.Of the surviving oocytes analysed, a nor-mal number and morphology of chromo-somes were seen in 80% (i) and 65% (ii) ofcases (Fig. 2a). In group (iii) (n=149), 78%of embryos were fragmented and 55% ofthe remaining analysed oocytes had nor-mal-looking chromosomes. In group (iv)(n=164), 54% of reconstructed embryosfragmented before analysis, and at the firstmitotic division 100% of the remainingembryos contained heavily pulverizedchromosomes of a type not seen in theother groups (Fig. 2b).
In a second and independent series ofexperiments, embryos produced bynuclear transfer (n=607) were allowed todevelop in vitro for four days. Develop-ment to the blastocyst stage was 69%(n=144) for group (i), 34% (n=175) forgroup (ii), 6% (n=197) for group (iii) and0% (n=91) for group (iv) (Fig. 1b). Thesevalues are similar to the correspondingproportions of one-cell embryos withapparently normal chromosomes in thefirst series (Fig. 1a). They further indicatethat the timing of activation with regardto nuclear transfer is critical to the out-come of development. Highest develop-mental rates were observed in group (i),with enucleated unfertilized oocytes acti-vated one to three hours after nucleartransfer. By stark contrast, when zygoteswere used as recipients (iv), none devel-oped to blastocysts, and their develop-ment was arrested at the two- or four-cellstage (data not shown).
These observations reflect differences incytoplasm-chromatin interactions betweenunfertilized oocytes and zygotes in mice. It
must be noted that the success of nucleartransfer in other mammals, such as sheep5
and cows6, does not depend critically onthe activation state of the recipient oocyte.Perhaps this may be attributed to speciesdifferences in the kinetics of early embry-onic development; alternatively, it may bedue to fundamental differences in themethods of nuclear transfer used.
Our data do not corroborate the find-ings of Illmensee and Hoppe, whoreported twenty years ago the cloning ofmice via injection of donor nuclei intozygotes7. Similarly, others have beenunable to duplicate the results of Illmenseeand Hoppe8,9. Certain experimental differ-ences exist: Illmensee and Hoppe usedICM cells from LT/Sv or CBA/H-T6 miceas nucleus donors, and enucleated zygotesfrom C57BL/6 mice as recipients7; we usedcumulus cells from B6D2F1 mice asnucleus donors and enucleated oocytes orzygotes from B6D2F1 mice as recipients. Itis difficult to prove that a phenomenon isbiologically impossible8, but in our hands,and under the described experimentalconditions, zygotes are inappropriatenucleus recipients for the cloning of micefrom adult somatic cells. In contrast toprevious negative reports8,9, we have suc-
Nuclear transfer into mouse zygotes
brief communications
Fig. 1 Fate of oocytes and zygotes after nuclear transfer. Oocytes were activated (i) 1–3 h after nucleartransfer (Post) (ii) immediately after nuclear transfer (Imm.) or (iii) 1 h before nuclear transfer (Pre); (iv)zygotes (Zyg.) are also shown. a, Chromosome analysis. The percentage is calculated as the fraction ofnon-fragmented embryos observed 16 h after nuclear transfer, multiplied with the fraction of analysedembryos with a morphologically normal set of chromosomes. Fragmented embryos were classified as hav-ing abnormal chromosomes. b, Blastocyst development after 4 d in vitro culture. This experimental series isindependent from the one presented in (a).
Fig. 2 Chromosomes of reconstructed embryosstained with Giemsa just before the first mitoticcell division. a, When unfertilized oocytes wereused as recipients, most embryos had a normal setof 40 chromosomes. b, When zygotes were usedas recipients, all embryos had extensively pulver-ized chromosomes.
a
b
a b
© 2000 Nature America Inc. • http://genetics.nature.com©
200
0 N
atu
re A
mer
ica
Inc.
• h
ttp
://g
enet
ics.
nat
ure
.co
m

brief communications
nature genetics • volume 24 • february 2000 109
ceeded in reproducibly cloning mice bynuclear transfer from a variety of celltypes1–3. In reply to Illmensee’s recent res-urrection of this debate10, we concur withSolter11 that there is no firm reason forHineininterpretierung of the claim ofmouse cloning using zygotes.
AcknowledgementsWe thank T. Perry for critical and usefulcomments on the manuscript. R.Y. acknowledgesfinancial support from ProBio America.
Teruhiko Wakayama1,3, Hiroyuki Tateno1,Peter Mombaerts2 & Ryuzo Yanagimachi11University of Hawaii Medical School,Department of Anatomy and ReproductiveBiology, Honolulu, Hawaii, USA. 2TheRockefeller University, New York, New York,USA. 3Present address: The RockefellerUniversity, New York, NY, USA. Correspondenceshould be addressed to T.W. (e-mail: [email protected]).
1. Wakayama, T., Perry, A.C F., Zuccotti, M., Johnson,K.R. & Yanagimachi, R. Nature 394, 369–374 (1998).
2. Wakayama, T. & Yanagimachi, R. Nature Genet. 22,127–128 (1999).
3. Wakayama, T., Rodriguez, I., Perry, A.C.F.,Yanagimachi, R. & Mombaerts, P. Proc. Natl Acad.Sci. USA 96, 14984–14989 (1999).
4. Mikamo, K. & Kamiguchi, Y. in Radiation-InducedChromosome Damage in Man (eds Ishihara, T. &Sasaki, M.S.) 411–432 (Alan R. Liss, New York,1983).
5. Campbell, K.H.S., McWhir, J., Ritchie, W.A. &Wilmut, I. Nature 380, 64–66 (1996).
6. Stice, S.L., Keefer, C.L. & Matthews, L. Mol. Reprod.Dev. 38, 61–68 (1994).
7. Illmensee, K. & Hoppe, P.C. Cell 23, 9–18 (1981).8. McGrath, J. & Solter, D. Science 226, 1317–1319
(1984).9. Howlett, S.K., Barton, S.C. & Surani, M.A.
Development 101, 915–923 (1987).10. Illmensee, K. Nature 398, 19–20 (1999).11. Solter, D. Nature 399, 13 (1999).
Mammalian cloning has recently beendeveloped1–5, but a major obstacle
has been the low frequency of viableclones, as most clones die during gesta-tion or soon after birth. Parameters thataffect cloning efficiency may includegenetic background, passage number,cell-cycle stage of the donor cell6, loss ofimprints, accumulated genetic damage ofthe donor cells or the ability of the oocyteto epigenetically reprogram the donor-cell nucleus. Cloning in mice5,7,8, the bestmammalian model organism, has untilrecently been limited to freshly isolatedor primary cultures of somatic cells,which limits study of the parameters thataffect cloning efficiency. We have deter-mined that established and targetedembryonic stem (ES) cells can generatecloned embryos and animals, thus mak-ing possible the study of parametersimportant for cloning.
To assess the most suitable genetic back-ground for cloning, we compared cloningefficiencies using F1 (129SvJae×C57BL/6)
ES cell lines and 129 (129SvJae) ES celllines as nuclear donors. Nuclear trans-plantation resulted in blastocyst-stageclones at a similar frequency for all ES-celldonors (Table 1). After transfer intopseudopregnant females, 7 of 34 (21%)F1-cloned blastocysts developed to termand into healthy adults. In comparison, 8of 76 (11%) inbred 129 donor ES cellsdeveloped to term, but all died within 24hours of birth, demonstrating an effect ofgenetic background on survival. The via-bility of the F1-derived clones was also5–70-fold greater than that reported forclones from other ES cell lines (R1, 4%,and E14, 0.3%; ref. 8). Thus, the F1 EScells were more efficient as donors fornuclear transfer than the inbred 129 (J1and E14) ES cells and the out-crossed 129R1 ES cells. The ability of ES cells to gen-erate viable cloned offspring might becorrelated with their degree of polymor-phism at 37 different SSLP markers (J1,0/37; E14, 0/37; R1, 12/37; our F1 line,28/37; ref. 9).
We also compared the developmentalpotential of clones derived from ES cellswith earlier studies of clones derived fromB6C3F1 and B6D2F1 somatic cells. Pre-implantation development of inbred 129and F1 ES-cell nuclei transferred into enu-cleated oocytes resulted in approximately15% blastocysts (Table 1), which was lessefficient than that reported for clonesderived from cumulus and tail-tip cells(50%–60%). The post-implantationdevelopment and survival to adulthood of21% of the F1 ES-cell–cloned blastocysts,however, was improved compared withthe 1.6% and 0.4% survival rate of clonesderived from cumulus and tail-tip donorblastocysts, respectively5,7.
The initial development of clones tothe blastocyst stage may be dependent onthe compatibility between the cell cyclesof the donor nucleus and the oocyte6.Thus the predominantly G0/G1 cumuluscells may be favoured over rapidly cyclingES cells and methods to purify or arrestES cells at specific stages of the cell cyclemay facilitate the generation of blasto-cysts. Wakayama et al.8 recently showedthat ES cells in either the G1 or G2/Mphase of the cell cycle could develop toterm after nuclear transfer. Therefore itmay be that only S-phase cells, which
Generation of mice from wild-type andtargeted ES cells by nuclear cloning
Table 1 Development of cloned embryos and mice from ES cells
Genetic Activated Embryosc transferred At term
background Cell line Passage oocytes (% activated) dead alive survived(% transferred)
F1 v 6.5 5–6 149 21 (14%) 0 4 4 (19%)F1 v6.5 sc84 8–9a 46 6 (13%) 2 2 2 (33%)F1 rtTA2∆SD-18 8–9a 32 7 (22%) 0 1 1 (14%)Total 227 34 (15%) 2 7 (21%) 7 (21%)
129 v 18.6 5–6 107 19 (19%) 1 4 0129 J1 11 145 26 (18%) 2 4 0129(high) J1 35b 92 23 (25%) 10 0 0129 LJG-13 8–9a 74 8 (11%) 4 0 0Total 418 76 (18%) 17 8 (11%) 0 (0%)
We produced embryos by transfer of nuclei from ES cells (cultured for 1–5 d without feeder cells in standard ES media containing 1,000 U/ml leukaemia inhibitoryfactor (LIF), but only 5% fetal calf serum) into enucleated B6D2F1 oocytes5. We activated the oocytes by exposure to Sr++-containing media, then cultured themin vitro for 3 d before transfer into 2.5-days post-coitum (d.p.c.) pseudopregnant Swiss females. We performed caesarean sections at 18.5–19.5 d.p.c. for all recip-ient females followed by fostering of any live pups. F1 refers to the 129SvJae×C57BL/6 genetic background. Surviving cloned animals were all derived from donorF1 ES-cell nuclei because all had agouti coat colour, whereas donor oocytes came from black, non-agouti B6D2F1 females and the cloned embryos had been trans-ferred into pseudopregnant white Swiss females. Targeted ES-cell lines are italicized. aES cells were targeted or subcloned at 4–5 passages and grown for another4 passages before transplantation. bHigh passage. cOnly embryos which had reached morula or blastocyst stage were transferred.
© 2000 Nature America Inc. • http://genetics.nature.com©
200
0 N
atu
re A
mer
ica
Inc.
• h
ttp
://g
enet
ics.
nat
ure
.co
m