normal human kidney hla-dr–expressing renal …jasn.asnjournals.org/content/14/5/1336.full.pdf ·...
TRANSCRIPT

Normal Human Kidney HLA-DR–Expressing RenalMicrovascular Endothelial Cells: Characterization, Isolation,and Regulation of MHC Class II Expression
KIMBERLY A. MUCZYNSKI,*† DAVID M. EKLE,*† DAVID M. CODER,‡ andSUSAN K. ANDERSON*†
Departments of *Medicine, †Pediatrics, and ‡Immunology, University of Washington, Seattle, Washington.
Abstract. Human, but not murine, renal peritubular and glo-merular capillaries constitutively express class II major histo-compatibility (MHC) proteins at high levels in normal humankidney. Expression of class II proteins on renal microvascularendothelial cells (RMEC) makes it available to circulatinglymphocytes and imparts a surveillance capacity to RMEC forcontrolling inflammatory responses. In this report, the co-expression of HLA-DR and the endothelial marker CD31 areused to identify RMEC as a distinct population of cells withina standard renal biopsy using flow cytometry. A three-laser,multicolor flow cytometry analysis using Alexa dyes, devel-oped for characterizing the expression of cell surface antigens,identifies RMEC as a population separate from HLA-DR-expressing leukocytes. HLA-DR RMEC co-express HLA-DPand HLA-DQ. RMEC also express the T cell costimulatory
factor CD58 but not CD80, CD86, or CD40. On the basis ofhigh HLA-DR expression, RMEC are isolated for culture usingfluorescence-activated cell sorting and magnetic beads. Cul-tured RMEC require normal basal physiologic concentrationsof gamma interferon (�IFN) to maintain HLA protein expres-sion. This expression is regulated by CIITA, the MHC classII-specific transcription factor. Four tissue-specific promotershave been described for CIITA. In freshly isolated RMEC,RT-PCR and hybridization using specific oligonucleotideprobes to CIITA promoter sequences identify only the statin-sensitive �IFN-induced promoter IV of CIITA. Therefore, theconstitutive expression of HLA-DR on RMEC in normal hu-man kidney is located in a position for immune surveillance,depends on basal physiologic concentrations of �IFN, and maybe amenable to regulation with statins.
MHC proteins are of two classes distinguished on the basis ofstructure and function. Class I molecules, composed of a poly-morphic subunit complexed with �-2-microglobulin, are foundon all nucleated cells and present antigenic peptides to CD8�T lymphocytes; class II molecules, composed of polymorphic� and � chains, are constitutively expressed on a limitednumber of cell types (dendritic cells, macrophages, B lympho-cytes) and present antigenic peptides to CD4� T lymphocytes.We recently described an unusual expression of MHC class IIproteins in normal human kidneys that is not found in murinekidneys (1). The human MHC class II protein HLA-DR isabundantly expressed on peritubular and glomerular capillaryendothelial cells but not on endothelial cells of larger bloodvessels of normal kidney. Antibodies to HLA-DR and CD31, aprotein highly expressed on endothelial cells, co-localize onperitubular and glomerular cells within sections of kidneytissue, indicating capillary endothelial cell location of HLA-DR. HLA-DR has also been identified on rare scattered circu-
lating leukocytes found within the kidney, but over 98% of theDR identified by immunofluorescence microscopy in kidneycortex is located on capillary endothelial cells (1). We refer tothese cells co-expressing HLA-DR and CD31 as renal micro-vascular endothelial cells (RMEC).
HLA class II molecules with their bound peptides play apivotal role in directing the immune responses of CD4� Tlymphocytes. Presentation of peptides to unprimed CD4� Tcells in the absence of CD80 and CD86 on the antigen-presenting cell renders T cells unresponsive to bound peptide(2–4); in the presence of these T cell costimulatory factors, theT lymphocytes become activated. Further control of CD4� Tlymphocyte responses comes from the regulated expression ofHLA class II molecules. Although constitutive class II expres-sion is limited to a few cell types, most other cells can beinduced to express class II with high concentrations of gammainterferon (�IFN; 100 to 500 units/ml).
Regulation of HLA class II expression occurs at the level oftranscription. The three HLA class II isotypes, DR, DP, andDQ, are coordinately transcribed due to common promotersand transcription factors. Transcription factors required forexpression of class II genes include CIITA (5–7), RFX-5 (8),RFX-B (9,10), and RFXAP (11). CIITA, the master switch forclass II transcription, is a non-DNA binding protein that isspecific for class II expression (12). The other transcriptionfactors are DNA binding proteins ubiquitously present in mostcells regardless of whether or not they express class II proteins.
Received May 25, 2002. Accepted January 30, 2003.Correspondence to Kimberly A. Muczynski, University of Washington, Depart-ment of Medicine and Division of Nephrology, Box 356521, Seattle, WA 98195.Phone: 206-598-6190; Fax: 425-255-5318; E-mail: [email protected]
1046-6673/1405-1336Journal of the American Society of NephrologyCopyright © 2003 by the American Society of Nephrology
DOI: 10.1097/01.ASN.0000061778.08085.9F
J Am Soc Nephrol 14: 1336–1348, 2003

CIITA, which can be induced with �IFN, is the specific �IFN-inducible factor responsible for induced class II expression(6,7). Regulation of constitutive class II expression on RMEClikely depends on CIITA, although a CIITA-independentmechanism for class II expression on endothelial cells thatinvolves NK cells has been described (13).
CIITA itself is highly regulated. Four different promotersand first exons have been sequenced for CIITA that are utilizedin a tissue specific manner (14). Constitutive HLA class IIexpression in dendritic cells utilizes promoter I; B cells expressclass II genes using CIITA promoter III; and �IFN-induction ofclass II genes is associated with CIITA promoter IV expression(14,15). Recent reports also describe constitutive class II ex-pression on melanoma and cortical thymic epithelial cells thatis mediated by CIITA promoter IV (16–19).
Characterization of the regulation of RMEC CIITA requiresisolating the cells. Classic capillary endothelial cell isolationprocedures involved separating the microvasculature from tis-sue and then allowing cells to grow from the vessels (20).Other investigators have isolated microvascular endothelialcells on the basis of induced expression of cell surface activa-tion markers (21). In this report, we isolate RMEC based onnormal basal level co-expression of DR and CD31, a marker ofendothelial cells, using fluorescence-activated cell sorting(FACS) and magnetic beads. RMEC lose DR expression after1 to 2 wk in vitro, but DR expression can be maintained withlow concentrations of �IFN. Similar findings have been re-ported for microvascular endothelial cells isolated from humanheart (22). To characterize RMEC as they exist in vivo withoutchanges that occur in culture, a triple laser flow cytometryanalysis is devised to evaluate expression of surface proteins inviable cells from normal kidney tissue. We find that RMEC invivo express DR via CIITA promoter IV without the T cellcostimulatory factors CD80, CD86, and CD40. These obser-vations suggest that RMEC have the potential for mediatingperipheral tolerance through DR expression and that statinsmay provide a pharmacologic means of regulating this immuneactivity by inhibiting the expression of CIITA promoter IV(23,24).
Materials and MethodsAntibodies
Monoclonal antibodies that recognize monomorphic HLA class IIdeterminants of the following specificities were purified and fluoro-phore-labeled in the laboratory: L243 recognizes DR (25), B7/21recognizes DP (26), and SPVL3 recognizes DQ (27). FITC conjuga-tion was performed in 0.29 M carbonate/bicarbonate buffer, pH 9.3,and labeled antibody was separated from free FITC using a SephadexG-25 column. Conjugation to Alexa Fluor dyes (Alexa 568, Alexa633, Alexa 680) was done with Alexa Fluor Protein Labeling Kitsfrom Molecular Probes (Eugene, OR) according to manufacturer’sinstructions. Two monoclonal antibodies to CD31, BD PharMingen(San Diego, CA) clone WM59 and Caltag (Burlingame, CA) cloneMBC78.2, both directly conjugated to the same fluorophore (FITC orPE) were used in combination for flow cytometry. Anti-CD45, CD14,CD19, CD80, CD86, CD58, CD141, and HLA-DR (L243) phyco-erythrin (PE) conjugates and CD40-FITC were obtained from BDPharMingen. CD105-PE was obtained from Caltag. L243, SPVL3,
anti-CD14, and anti-CD58 are IgG2a antibodies; the other antibodiesused are all IgG1. Anti-von Willebrand factor (vWF) rabbit serum andFITC or tetramethylrhodamine isothiocyanate (TRITC) conjugatedsecondary goat anti-rabbit antibodies were obtained from Sigma (St.Louis, MO).
CellsHUVEC and lung microvascular endothelial cells (LMEC) were
purchased from Clonetics and grown in EGM-2MV BulletKit medium(BioWhittaker, Walkersville, MD). Jurkat, a T cell line that does notexpress CD80, CD86, or CD40; EBV-transformed B-lymphoblastoidcell lines, which express high levels of HLA class II proteins, CD80,CD86, CD40, and CD58; T2, a T and B lymphoblastoid hybrid that isDR-negative on the basis of class II gene deletion (28); ThM, amelanoma line that does not express DR unless induced with �IFN(29); and HK-2, a human kidney proximal tubule cell line that doesnot express DR unless �IFN-induced (30), were grown in RPMIcontaining 10% bovine calf serum.
Isolated RMEC were grown in Clonetics EGM-2-MV BulletKitmedium in a 5% CO2 humidified environment. RMEC grow equallywell in GibcoBRL human endothelial-SFM medium (Life Technolo-gies, Gaithersburg, MD) containing either 5% fetal calf serum or 5 to10% pooled human AB sera.
RMEC IsolationRMEC were isolated from normal areas of human kidney cortex
recovered at the time of nephrectomy for renal cell carcinoma. Donorshad normal renal function, no proteinuria, and grossly normal appear-ing parenchyma in areas unaffected by tumor. Tissue could be storedat 4°C overnight in Hank’s balanced salt solution (HBSS; GibcoBRLcatalog number 24020–125) supplemented with 100 U/ml penicillinand 50 �g/ml streptomycin (HBSS) before cell isolation withoutaffecting results.
Cell suspensions used for flow cytometric sorting and magneticbead isolation were prepared in the same manner using sterile tech-niques. Excised grossly normal kidney cortex (1 to 10 g) was mincedwith a scalpel and digested for 1 h at 37°C in 30 ml of 0.2%collagenase P (Roche Molecular Biochemicals, Indianapolis, IN) inHBSS on an orbital shaker. This preparation was centrifuged at lowspeed (600 to 1000 � g), resuspended in 10 to 20 ml phosphate-buffered saline (PBS) containing 0.05% trypsin and 0.53 mM EDTA(GibcoBRL), and agitated for 10 to 15 min at 37°C. An equal volumeof Clonetics EGM-2-MV BulletKit medium was added to neutralizetrypsin, and the resulting cell suspension was passed through a 20-mesh sieve to remove remaining large particulates. Recovered cellswere then labeled with antibody as described below for isolation ofRMEC using flow cytometry or magnetic beads. Cells could also bestored at 4°C overnight before cytometry without loss of viability orchange in phenotype.
RMEC were isolated by flow cytometry sorting as follows. Kidneycell suspensions were labeled on ice for 1 h with saturating concen-trations of two anti-CD31 antibodies that recognize different CD31epitopes, both conjugated with the same fluorophore, and with FITC-conjugated or PE-conjugated L243 (anti-HLA-DR) in RPMI 1640containing 25 mM HEPES, 5% FBS, pH 8.0 (diluent). L243-FITCwas used with anti-CD31-PE, and L243-PE with anti-CD31-FITC.Cells were washed with diluent and passed through a nylon strainer(40-�m pore size; Falcon 352340, Becton Dickinson, San Jose, CA;available from Fisher, Pittsburgh, PA). Propidium iodide (PI) wasadded at a concentration of 5 �g/ml just before cell sorting as aviability indicator. Cell sorting was performed with a FACSVantage
J Am Soc Nephrol 14: 1336–1348, 2003 HLA-DR–Expressing RMEC 1337

SE (Becton Dickinson) equipped with a 70-�m-diameter nozzle tipand an argon laser-emitting 150 mW of 488 nm light. PI-negative cellswith the highest level of CD31 and DR expression were collectedunder sterile conditions in a 15 ml conical polypropylene tube con-taining Clonetics EGM-2-MV BulletKit medium, centrifuged, andplaced in culture with fresh medium.
RMEC were isolated using magnetic beads as follows. Kidneycortical cells prepared above were incubated on ice with saturatingconcentrations of magnetic anti-CD14- and anti-CD19-coated Dyna-beads (Dynal, Oslo, Norway) in diluent for 30 to 60 min and thendropped onto a tilted cell culture dish placed over a magnet. Cells thatdid not adhere to the magnet were collected, washed, and labeled withL243 in diluent on ice for 30 min. A trace amount of L243-FITC orL243-Alexa 633 was added so the isolation procedure could beassessed by immunofluorescence microscopy. Excess unbound L243was removed by washing the cells. L243-bound cells were isolatedusing goat anti-mouse IgG-coated Dynabeads as described above orby using goat anti-mouse Miltenyi microbeads (Miltenyi Biotec, Au-burn, CA) according to the manufacturer’s specifications. Cells ad-herent to the magnetic beads were placed in culture in CloneticsEGM-2-MV BulletKit medium.
Cell Suspensions from Renal BiopsiesCell suspensions for multicolor flow cytometric analysis were
prepared from 16-gauge needle biopsy tissue cores obtained underdirect visualization of surgically removed longitudinally bisected kid-neys from grossly normal cortex. Four to eight biopsies were takenfrom each kidney in different areas of normal cortex. Each biopsycontained enough cells for labeling with 6 to 8 different sets ofantibodies, with each set consisting of 15,000 to 20,000 cells. Singlebiopsy cores were placed in 1.5-ml microfuge tubes, digested for 1 hat 37°C with 0.2% collagenase P in HBSS on an orbital shaker,followed by 10 min at 37°C with 0.05% trypsin-0.53 mM EDTA inPBS. Trypsin was neutralized with an equal volume of CloneticsEGM-2-MV BulletKit medium. Cells could be stored overnight at4°C before labeling with antibody without affecting results. Flowcytometry results of different biopsy tissue cores from the same donorwere identical.
Antibody Labeling of Cells for FlowCytometric Analysis
Cells from renal biopsies or cells prepared for RMEC isolationwere labeled with the desired combination of antibodies, each conju-gated to a different fluorophore (FITC, PE, Alexa Fluor 633, or AlexaFluor 680) in RPMI 1640 or diluent containing 0.02% sodium azideon ice. Excess antibody was removed by washing cells. 4',6-dia-midino-2-phenylindole (DAPI) was added at a concentration of 1�g/ml in RPMI as a viability indicator just before analysis with a LSRbench top flow cytometer (Becton Dickinson Biosciences). PI wasadded at a concentration of 5 �g/ml as a viability indicator with single488-nm laser instruments.
Flow CytometrySingle laser flow cytometry was performed as described previously
(29).A three-laser multicolor flow cytometry analysis was developed to
characterize cell surface antigens on freshly isolated RMEC to gaininformation about the in vivo phenotype of the cells. A BectonDickinson LSR bench top analyzer equipped with HeCd (325 nm), Ar(488 nm), and HeNe (633 nm) lasers was configured to allow dis-crimination of DAPI, FITC, PE, Alexa 633, and Alexa 680 fluoro-
phores. Specific description of LSR configuration and filters areavailable upon request. Compensation was adjusted using B lympho-blastoid cells labeled with single L243 conjugates: L243-FITC, -PE,-Alexa 633, or -Alexa 680. A B lymphoblastoid cell line in whichHLA class II genes are deleted, T2, was used as a negative control forL243 binding. After compensation, each fluorophore detected thesame level of DR expression. Conjugation of purified antibody toFITC, Alexa 633, and Alexa 680 dyes is a straightforward procedureand increases the flexibility of antibody-fluorophore combinationsavailable for use with this 5-color flow cytometry analysis. Only PEconjugates need to be purchased from commercial suppliers.
Cell Quest and WinMDI software were used for data analysis.
Immunofluorescence MicroscopyImmunofluorescence microscopy on human kidney tissue was per-
formed as described previously (1). Cultured RMEC viewed by im-munofluorescence microscopy were treated with trypsin to removethem from culture dishes, labeled according to the procedure used forflow cytometry, and then fixed in 1% paraformaldehyde in phosphate-buffered saline. Trypsin does not affect the level of detectableHLA-DR expression. Cells were mounted in Vectashield MountingMedium with DAPI (Vector Laboratories).
RT-PCR and Southern BlotsSingle-stranded cDNA was made from RMEC using cesium chlo-
ride gradient isolated total RNA and a Superscript first-strand synthe-sis system (Invitrogen Life Technologies, Carlsbad, CA). Oligo dT orCIITA oligonucleotide 5'-TCTTGCTGCTGCTCCTCT-3' were usedto prime the reverse transcriptase. PCR using the oligo dT-primedcDNA and internal CIITA common primers 5'-CCTGATGCACAT-GTACTGGGC-3' and 5'-ACGTCCATCACCCGGAGGGAC-3' gen-erated a 711-bp product common to all CIITA. Promoter-specificRT-PCR was performed with cDNA template, a common reverseprimer (RP) 5'-GCACCTCACCATGGTAGATGA-3' and a specificforward primer (FP). CIITA III FP sequence used was 5'-GAG-GCTAGTGATGAGGCTGTG-3' and CIITA IV FP, 5'-ACTTGC-CGCGGCCCCAGAGCT-3'. PCR amplification was performed with55 cycles of 94°C � 30 s (denaturation), 50°C � 1 min (annealing),and 72°C � 2 min (extension). Glyceraldehyde-3-phosphate dehydro-genase (GADPH) was amplified as an internal control using Strat-agene (La Jolla, CA) primers that generated a 600-bp product.
Since CIITA is a 4.5-kb mRNA of low abundance, it is difficult todetect CIITA promoters when fewer than 1 � 106 RMEC are avail-able and cDNA is made from oligo dT primed reverse transcriptasereactions. Therefore, a 1500-bp cDNA PCR template correspondingto the 5' end of CIITA was generated. CIITA promoter RT-PCRproduct yields are greater using this shortened cDNA template, al-though identical results are obtained with cDNA generated from oligodT priming.
Southern blots of RT-PCR products were prepared by standardtechniques and hybridized to promoter-specific CIITA oligonucleo-tides labeled at the 5' end with T4 polynucleotide kinase (NewEngland Biolabs, Beverly, MA) and �32P-ATP. Oligonucleotideprobe for CIITA promoter III sequence was 5'-AGACTCCGG-GAGCTGCTGCCTGGCTGGGATT-3'; oligonucleotide probe forCIITA promoter IV was 5'-GGCCACCAGCAGCGCGCGCGGGA-GCCCGGGGAACAG-3'.
1338 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 1336–1348, 2003

ResultsMulticolor Flow Cytometry Analysis of FreshlyIsolated RMEC
A three-laser multicolor flow cytometry analysis was devel-oped to characterize cell surface antigens on freshly isolatedRMEC to gain information about the in vivo phenotype of thecells. The flow cytometric analysis allows the use of CD31 andHLA-DR antibodies to precisely define RMEC within thepopulation of kidney cortex cells while simultaneously usingantibodies conjugated to other fluorophores to evaluate theexpression of additional RMEC cell surface proteins. We de-vised a filter system that allows the discrimination of Alexa633 and Alexa 680 antibody conjugates excited by the far redlaser. These fluorophores are used in combination with stan-dard FITC and PE conjugates. DAPI, excited with a UV laserand excluded by cells that maintain membrane integrity, isused as a viability indicator.
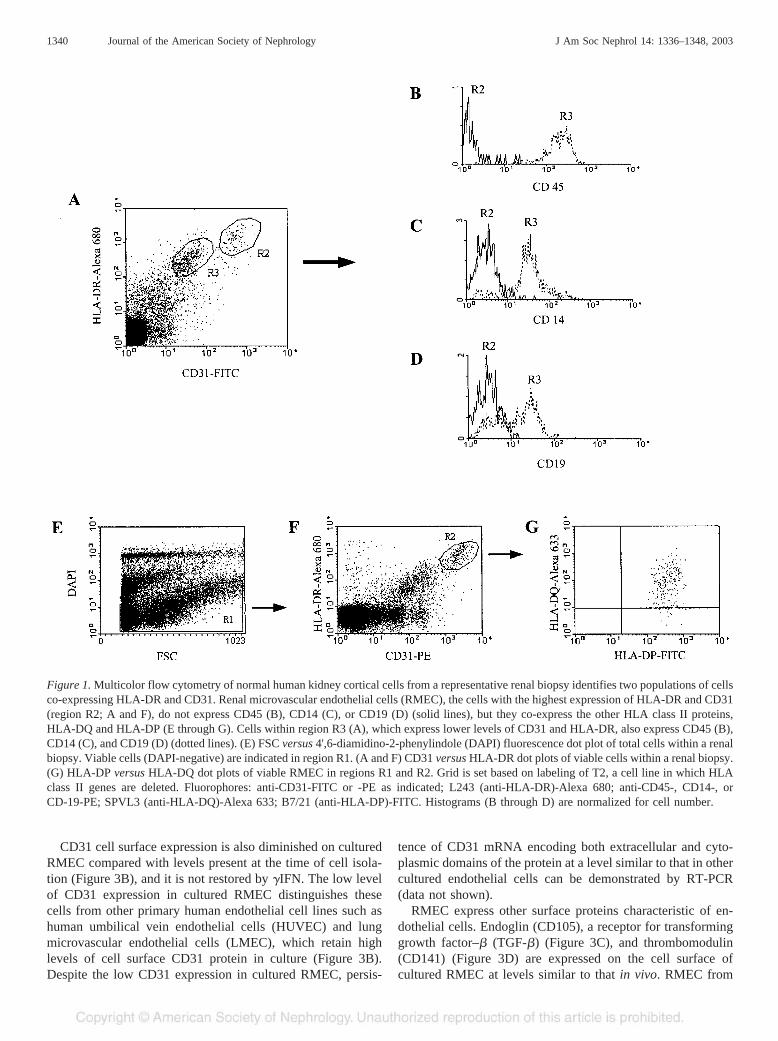
Multicolor flow cytometric analysis of viable cells containedwithin a renal biopsy of normal human kidney cortex revealstwo populations of cells expressing CD31 and HLA-DR (Fig-ure 1A). RMEC, which express high levels of CD31 andHLA-DR, do not express CD45, the leukocyte common anti-gen; the population of cells expressing lower levels of CD31and HLA-DR also express CD45, which identifies them asleukocytes (Figure 1B). Leukocytes that constitutively expressHLA-DR include monocytes/macrophages and B lymphocytes,which can be identified on the basis of CD14 and CD19expression, respectively. RMEC do not express CD14 andCD19, but a subset of the population of cells expressing lowlevels of CD31 and HLA-DR expresses CD14 and CD19,consistent with these cells being CD45� leukocytes of mono-cytic and B-lymphocytic origin (Figure 1, C and D). The twopopulations of CD31�/HLA-DR� cells are not evident byimmunofluorescence microscopy of kidney tissue sections(Figure 2A). This may be due to the loss of circulating CD45expressing leukocytes from tissue during preparation for mi-croscopy or it may be that the levels of CD31 and HLA-DRexpression on leukocytes are below the level of detection byimmunofluorescence microscopy. For example, flow cytom-etry detects cells with low level HLA-DR expression that areundetectable by immunofluorescence microscopy (Figure 3A).
Our initial immunofluorescence microscopy characteriza-tion of HLA class II proteins on RMEC described HLA-DRexpression without the other HLA class II isotypes, DP and DQ(1). This was unusual given that HLA class II proteins arecoordinately regulated through common promoters and tran-scription factors, including one transcription factor specific forclass II genes, CIITA. Among cells that express HLA class IIproteins on their surface, HLA-DP and HLA-DQ are generallyof lower abundance than HLA-DR. Therefore, one possibleexplanation for the apparent lack of HLA-DP and HLA-DQ onRMEC by immunofluorescence microscopy is that the level ofexpression is below the detectable limit of this technique.Using multicolor flow cytometric analysis, which is moresensitive for detecting low-level fluorescence, RMEC fromnormal kidney cortex are found to express HLA-DR, HLA-DP,
and HLA-DQ (Figure 1, E through G), although the level ofDQ expression has been variable for the donors studied. This isconsistent with HLA class II protein expression in normalhuman RMEC being regulated by the mechanisms reported insimpler cell culture systems and indicates that RMEC have thepotential for presenting antigen peptides via all three HLAclass II isotypes.
Isolation of RMEC Co-Expressing CD31 and HLA-DRThe abundant and relatively restricted expression of
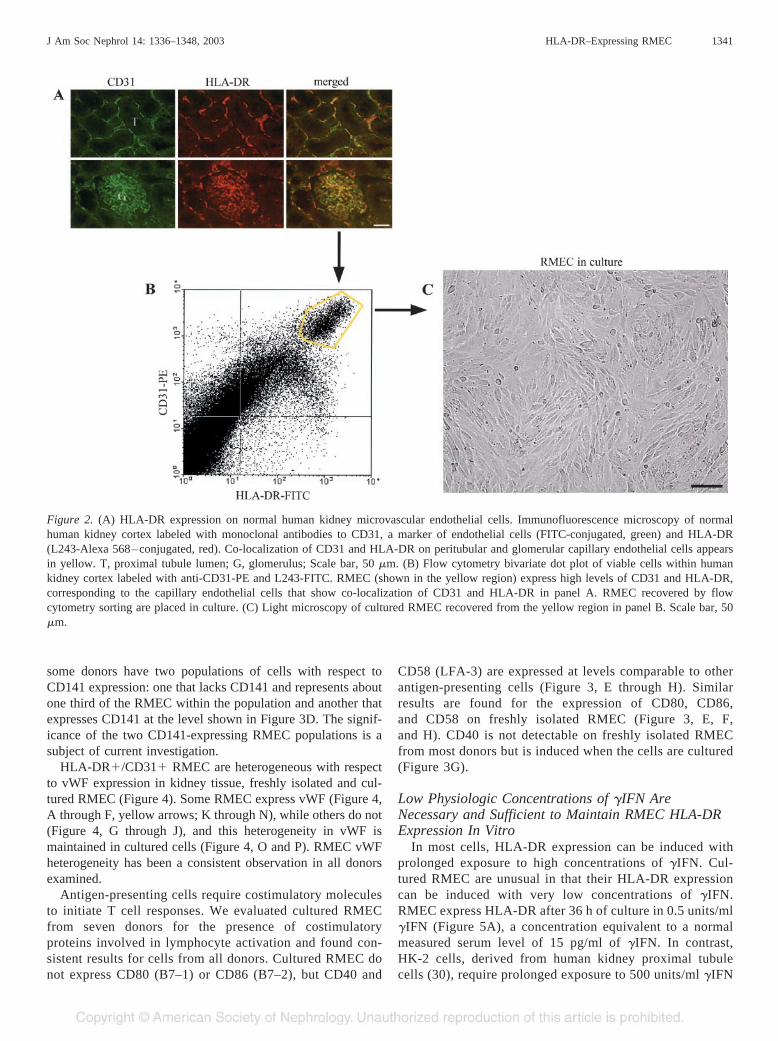
HLA-DR on RMEC in normal kidney allows isolation ofRMEC using antibodies to HLA-DR (Figure 2). Single cellsuspensions prepared from normal human kidney cortex usingcollagenase and trypsin are labeled with antibodies to HLA-DRand CD31, each conjugated with a different fluorophore. L243,a monoclonal antibody that recognizes an epitope common toall DR alleles, is used so that HLA-DR polymorphism does notaffect RMEC isolation. Viable cells with the highest expres-sion of both HLA-DR and CD31 are selected by flow cytom-etry for culture (Figure 2B). Cells are sorted at a rate of 5000to 8000 events per second. Two to three percent of the totalsorted cells are collected as RMEC, and purity is greater than98%. Isolated RMEC grow well in culture with commerciallyavailable media (Figure 2C).
RMEC can also be isolated on the basis of HLA-DR expres-sion using magnetic beads coated with an anti-mouse IgGantibody. However, cell suspensions derived from whole kid-ney cortex also contain circulating HLA-DR expressing mono-cytes, macrophages, and B lymphocytes; therefore, these cellsmust be removed before labeling with L243. Anti-CD14 andanti-CD19 beads remove monocytes/macrophages and B lym-phocytes, respectively. Kidney cortical cell suspensionscleared of CD14- and CD19-expressing cells are labeled withL243 to isolate the remaining HLA-DR–expressing cells,which are the RMEC. Ig-coated magnetic beads from Dynaland Miltenyi work equally well for isolating RMEC. Thepurity of RMEC isolated by magnetic bead immunoselection isgreater than 95%. The morphology and phenotype of thecultured cells are identical to RMEC isolated by flow cytom-etry. Greater numbers of viable cells are recovered usingmagnetic bead isolation techniques.
Phenotype of Cultured versus Freshly Isolated RMECCultured RMEC isolated by either flow cytometry or mag-
netic beads maintain a uniform phenotype that varies littlebetween donors, although the expression of some surface an-tigens differ from that observed on tissue and freshly isolatedRMEC. Cultured RMEC lose expression of HLA class IIproteins after 7 to 10 d unless �IFN is maintained in the media.RMEC that lose HLA-DR expression in culture can be inducedto re-express DR with �IFN (Figure 3A). The superior sensi-tivity of flow cytometry over immunofluorescence microscopyfor detecting low levels of surface protein expression is alsoapparent in Figure 3A; low levels of HLA-DR are readilydetected by flow cytometry 12 h after addition of �IFN,whereas immunofluorescence microscopy does not identifyHLA-DR expression until levels are higher at 24 h.
J Am Soc Nephrol 14: 1336–1348, 2003 HLA-DR–Expressing RMEC 1339
CD31 cell surface expression is also diminished on culturedRMEC compared with levels present at the time of cell isola-tion (Figure 3B), and it is not restored by �IFN. The low levelof CD31 expression in cultured RMEC distinguishes thesecells from other primary human endothelial cell lines such ashuman umbilical vein endothelial cells (HUVEC) and lungmicrovascular endothelial cells (LMEC), which retain highlevels of cell surface CD31 protein in culture (Figure 3B).Despite the low CD31 expression in cultured RMEC, persis-
tence of CD31 mRNA encoding both extracellular and cyto-plasmic domains of the protein at a level similar to that in othercultured endothelial cells can be demonstrated by RT-PCR(data not shown).
RMEC express other surface proteins characteristic of en-dothelial cells. Endoglin (CD105), a receptor for transforminggrowth factor–� (TGF-�) (Figure 3C), and thrombomodulin(CD141) (Figure 3D) are expressed on the cell surface ofcultured RMEC at levels similar to that in vivo. RMEC from
Figure 1. Multicolor flow cytometry of normal human kidney cortical cells from a representative renal biopsy identifies two populations of cellsco-expressing HLA-DR and CD31. Renal microvascular endothelial cells (RMEC), the cells with the highest expression of HLA-DR and CD31(region R2; A and F), do not express CD45 (B), CD14 (C), or CD19 (D) (solid lines), but they co-express the other HLA class II proteins,HLA-DQ and HLA-DP (E through G). Cells within region R3 (A), which express lower levels of CD31 and HLA-DR, also express CD45 (B),CD14 (C), and CD19 (D) (dotted lines). (E) FSC versus 4',6-diamidino-2-phenylindole (DAPI) fluorescence dot plot of total cells within a renalbiopsy. Viable cells (DAPI-negative) are indicated in region R1. (A and F) CD31 versus HLA-DR dot plots of viable cells within a renal biopsy.(G) HLA-DP versus HLA-DQ dot plots of viable RMEC in regions R1 and R2. Grid is set based on labeling of T2, a cell line in which HLAclass II genes are deleted. Fluorophores: anti-CD31-FITC or -PE as indicated; L243 (anti-HLA-DR)-Alexa 680; anti-CD45-, CD14-, orCD-19-PE; SPVL3 (anti-HLA-DQ)-Alexa 633; B7/21 (anti-HLA-DP)-FITC. Histograms (B through D) are normalized for cell number.
1340 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 1336–1348, 2003
some donors have two populations of cells with respect toCD141 expression: one that lacks CD141 and represents aboutone third of the RMEC within the population and another thatexpresses CD141 at the level shown in Figure 3D. The signif-icance of the two CD141-expressing RMEC populations is asubject of current investigation.
HLA-DR�/CD31� RMEC are heterogeneous with respectto vWF expression in kidney tissue, freshly isolated and cul-tured RMEC (Figure 4). Some RMEC express vWF (Figure 4,A through F, yellow arrows; K through N), while others do not(Figure 4, G through J), and this heterogeneity in vWF ismaintained in cultured cells (Figure 4, O and P). RMEC vWFheterogeneity has been a consistent observation in all donorsexamined.
Antigen-presenting cells require costimulatory moleculesto initiate T cell responses. We evaluated cultured RMECfrom seven donors for the presence of costimulatoryproteins involved in lymphocyte activation and found con-sistent results for cells from all donors. Cultured RMEC donot express CD80 (B7–1) or CD86 (B7–2), but CD40 and
CD58 (LFA-3) are expressed at levels comparable to otherantigen-presenting cells (Figure 3, E through H). Similarresults are found for the expression of CD80, CD86,and CD58 on freshly isolated RMEC (Figure 3, E, F,and H). CD40 is not detectable on freshly isolated RMECfrom most donors but is induced when the cells are cultured(Figure 3G).
Low Physiologic Concentrations of �IFN AreNecessary and Sufficient to Maintain RMEC HLA-DRExpression In Vitro
In most cells, HLA-DR expression can be induced withprolonged exposure to high concentrations of �IFN. Cul-tured RMEC are unusual in that their HLA-DR expressioncan be induced with very low concentrations of �IFN.RMEC express HLA-DR after 36 h of culture in 0.5 units/ml�IFN (Figure 5A), a concentration equivalent to a normalmeasured serum level of 15 pg/ml of �IFN. In contrast,HK-2 cells, derived from human kidney proximal tubulecells (30), require prolonged exposure to 500 units/ml �IFN
Figure 2. (A) HLA-DR expression on normal human kidney microvascular endothelial cells. Immunofluorescence microscopy of normalhuman kidney cortex labeled with monoclonal antibodies to CD31, a marker of endothelial cells (FITC-conjugated, green) and HLA-DR(L243-Alexa 568–conjugated, red). Co-localization of CD31 and HLA-DR on peritubular and glomerular capillary endothelial cells appearsin yellow. T, proximal tubule lumen; G, glomerulus; Scale bar, 50 �m. (B) Flow cytometry bivariate dot plot of viable cells within humankidney cortex labeled with anti-CD31-PE and L243-FITC. RMEC (shown in the yellow region) express high levels of CD31 and HLA-DR,corresponding to the capillary endothelial cells that show co-localization of CD31 and HLA-DR in panel A. RMEC recovered by flowcytometry sorting are placed in culture. (C) Light microscopy of cultured RMEC recovered from the yellow region in panel B. Scale bar, 50�m.
J Am Soc Nephrol 14: 1336–1348, 2003 HLA-DR–Expressing RMEC 1341
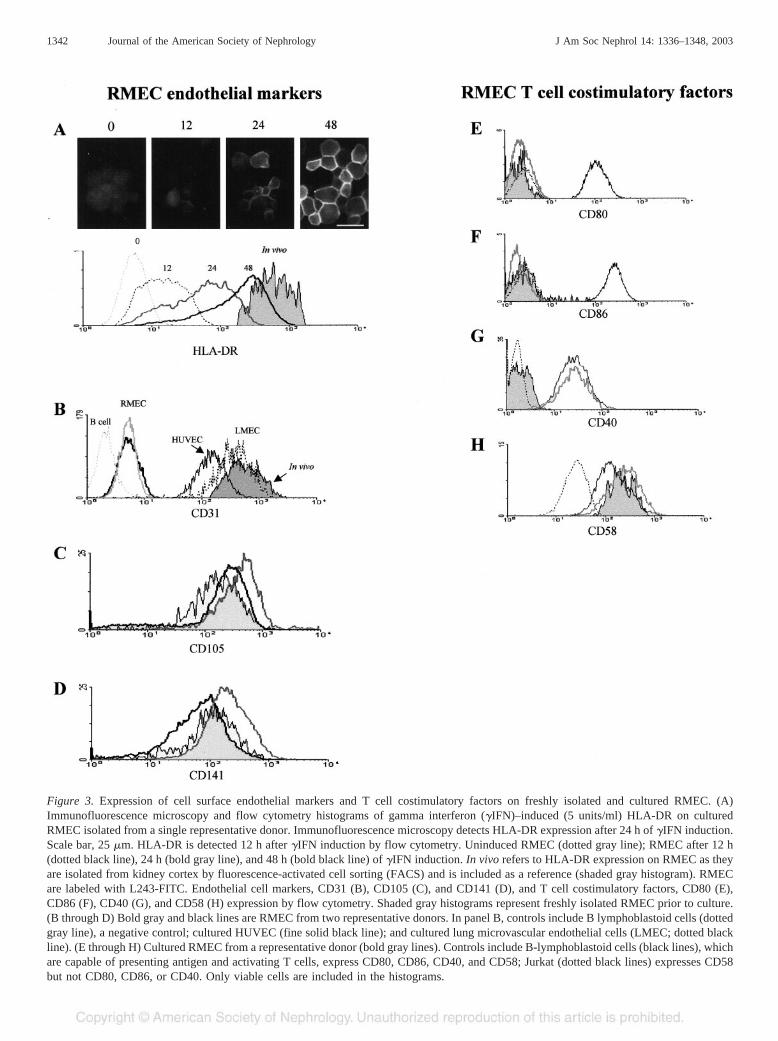
Figure 3. Expression of cell surface endothelial markers and T cell costimulatory factors on freshly isolated and cultured RMEC. (A)Immunofluorescence microscopy and flow cytometry histograms of gamma interferon (�IFN)–induced (5 units/ml) HLA-DR on culturedRMEC isolated from a single representative donor. Immunofluorescence microscopy detects HLA-DR expression after 24 h of �IFN induction.Scale bar, 25 �m. HLA-DR is detected 12 h after �IFN induction by flow cytometry. Uninduced RMEC (dotted gray line); RMEC after 12 h(dotted black line), 24 h (bold gray line), and 48 h (bold black line) of �IFN induction. In vivo refers to HLA-DR expression on RMEC as theyare isolated from kidney cortex by fluorescence-activated cell sorting (FACS) and is included as a reference (shaded gray histogram). RMECare labeled with L243-FITC. Endothelial cell markers, CD31 (B), CD105 (C), and CD141 (D), and T cell costimulatory factors, CD80 (E),CD86 (F), CD40 (G), and CD58 (H) expression by flow cytometry. Shaded gray histograms represent freshly isolated RMEC prior to culture.(B through D) Bold gray and black lines are RMEC from two representative donors. In panel B, controls include B lymphoblastoid cells (dottedgray line), a negative control; cultured HUVEC (fine solid black line); and cultured lung microvascular endothelial cells (LMEC; dotted blackline). (E through H) Cultured RMEC from a representative donor (bold gray lines). Controls include B-lymphoblastoid cells (black lines), whichare capable of presenting antigen and activating T cells, express CD80, CD86, CD40, and CD58; Jurkat (dotted black lines) expresses CD58but not CD80, CD86, or CD40. Only viable cells are included in the histograms.
1342 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 1336–1348, 2003

to induce similar levels of DR expression (Figure 5B). Theinduction of HLA-DR expression on cultured RMEC withbasal physiologic concentrations of �IFN suggests that theconstitutive in vivo expression of HLA-DR observed onRMEC in human kidney tissue may depend upon circulatingor locally produced �IFN.
Statin-Sensitive CIITA Promoter IV Transcription inFreshly Isolated RMEC
The loss of HLA-DR expression in cultured RMEC and itsrestoration by basal physiologic concentrations of �IFN sug-gested that the �IFN-inducible CIITA promoter IV might me-diate in vivo RMEC HLA-DR expression. RMEC, freshly
Figure 4. Expression of von Willebrand factor (vWF) is heterogeneous in tissue (A through F), freshly isolated RMEC (G through N), andcultured RMEC (O and P). Yellow arrows indicate RMEC expressing HLA-DR and vWF (A through C) or CD31 and vWF (D through F) intissue. RMEC freshly isolated by magnetic beads co-express HLA-DR and CD31 (G, I, and J; K, M, and N), and some also express vWF (Land N). Cultured RMEC that express vWF (O and P) can have a diffuse intracellular pattern or discrete intracellular structures resemblingWeibel-Palade bodies (P). Pseudocolors used: A, D, G, and K are in green channel; B, E, H, and L are in red channel; I and M are in bluechannel; DAPI nuclear label in O and P are in red channel. Scale bars: 50 �m in A through F; 20 �m in G through N; 20 �m in O; 30 �min P.
J Am Soc Nephrol 14: 1336–1348, 2003 HLA-DR–Expressing RMEC 1343
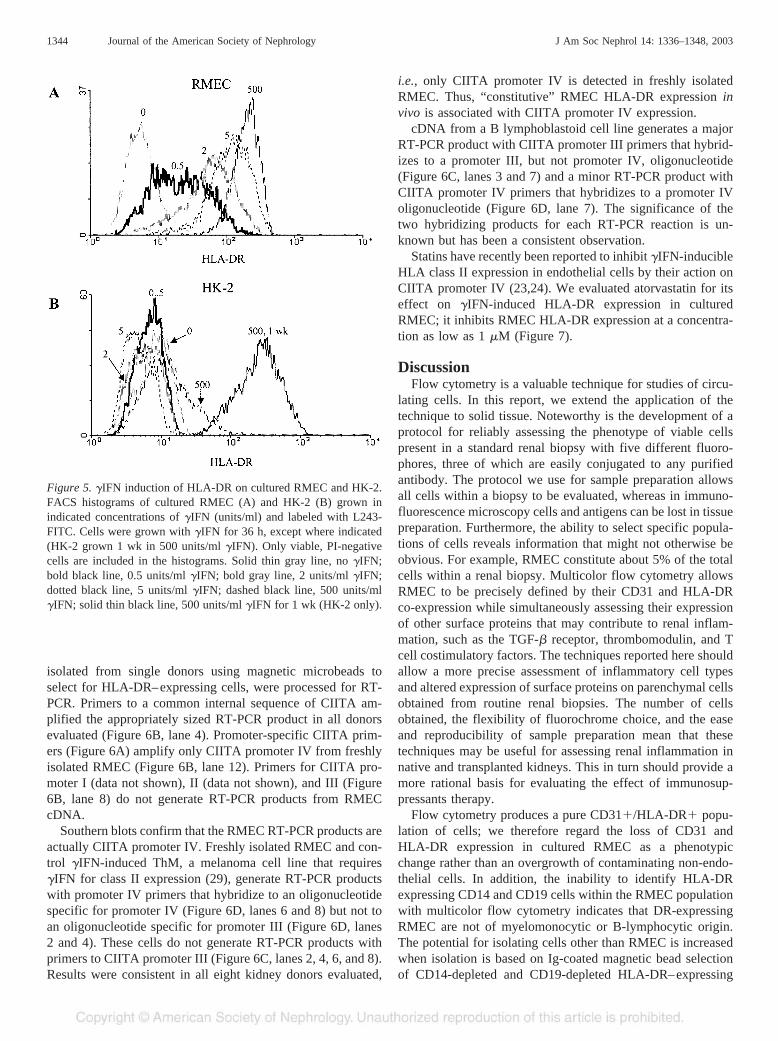
isolated from single donors using magnetic microbeads toselect for HLA-DR–expressing cells, were processed for RT-PCR. Primers to a common internal sequence of CIITA am-plified the appropriately sized RT-PCR product in all donorsevaluated (Figure 6B, lane 4). Promoter-specific CIITA prim-ers (Figure 6A) amplify only CIITA promoter IV from freshlyisolated RMEC (Figure 6B, lane 12). Primers for CIITA pro-moter I (data not shown), II (data not shown), and III (Figure6B, lane 8) do not generate RT-PCR products from RMECcDNA.
Southern blots confirm that the RMEC RT-PCR products areactually CIITA promoter IV. Freshly isolated RMEC and con-trol �IFN-induced ThM, a melanoma cell line that requires�IFN for class II expression (29), generate RT-PCR productswith promoter IV primers that hybridize to an oligonucleotidespecific for promoter IV (Figure 6D, lanes 6 and 8) but not toan oligonucleotide specific for promoter III (Figure 6D, lanes2 and 4). These cells do not generate RT-PCR products withprimers to CIITA promoter III (Figure 6C, lanes 2, 4, 6, and 8).Results were consistent in all eight kidney donors evaluated,
i.e., only CIITA promoter IV is detected in freshly isolatedRMEC. Thus, “constitutive” RMEC HLA-DR expression invivo is associated with CIITA promoter IV expression.
cDNA from a B lymphoblastoid cell line generates a majorRT-PCR product with CIITA promoter III primers that hybrid-izes to a promoter III, but not promoter IV, oligonucleotide(Figure 6C, lanes 3 and 7) and a minor RT-PCR product withCIITA promoter IV primers that hybridizes to a promoter IVoligonucleotide (Figure 6D, lane 7). The significance of thetwo hybridizing products for each RT-PCR reaction is un-known but has been a consistent observation.
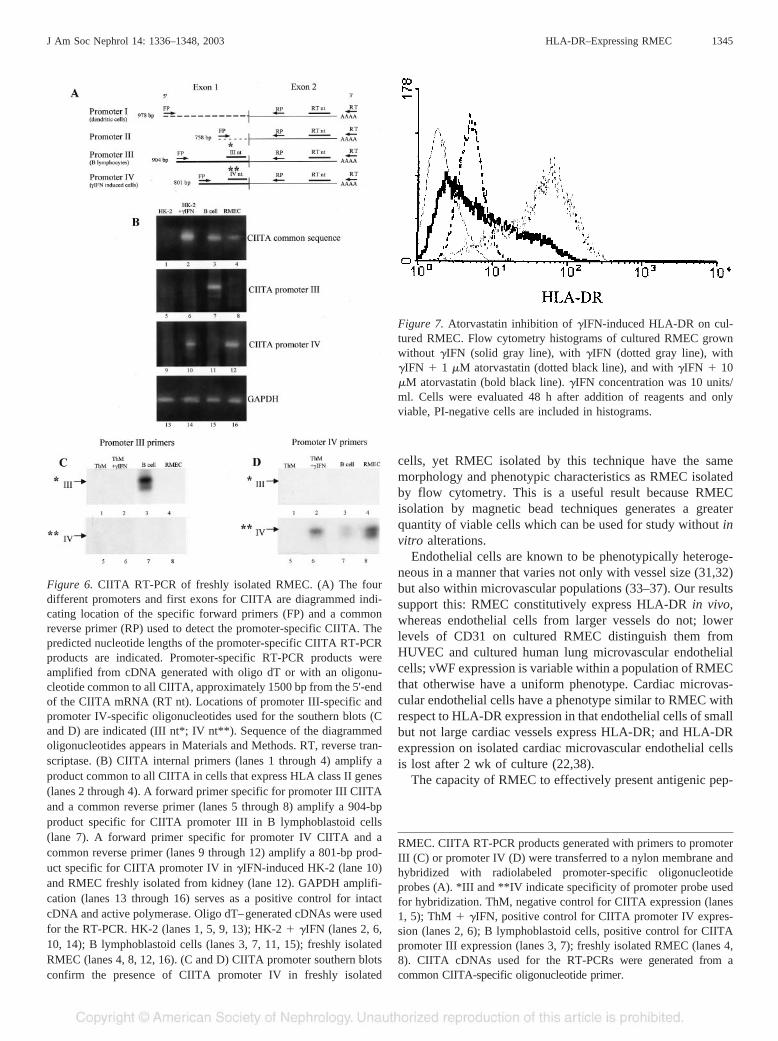
Statins have recently been reported to inhibit �IFN-inducibleHLA class II expression in endothelial cells by their action onCIITA promoter IV (23,24). We evaluated atorvastatin for itseffect on �IFN-induced HLA-DR expression in culturedRMEC; it inhibits RMEC HLA-DR expression at a concentra-tion as low as 1 �M (Figure 7).
DiscussionFlow cytometry is a valuable technique for studies of circu-
lating cells. In this report, we extend the application of thetechnique to solid tissue. Noteworthy is the development of aprotocol for reliably assessing the phenotype of viable cellspresent in a standard renal biopsy with five different fluoro-phores, three of which are easily conjugated to any purifiedantibody. The protocol we use for sample preparation allowsall cells within a biopsy to be evaluated, whereas in immuno-fluorescence microscopy cells and antigens can be lost in tissuepreparation. Furthermore, the ability to select specific popula-tions of cells reveals information that might not otherwise beobvious. For example, RMEC constitute about 5% of the totalcells within a renal biopsy. Multicolor flow cytometry allowsRMEC to be precisely defined by their CD31 and HLA-DRco-expression while simultaneously assessing their expressionof other surface proteins that may contribute to renal inflam-mation, such as the TGF-� receptor, thrombomodulin, and Tcell costimulatory factors. The techniques reported here shouldallow a more precise assessment of inflammatory cell typesand altered expression of surface proteins on parenchymal cellsobtained from routine renal biopsies. The number of cellsobtained, the flexibility of fluorochrome choice, and the easeand reproducibility of sample preparation mean that thesetechniques may be useful for assessing renal inflammation innative and transplanted kidneys. This in turn should provide amore rational basis for evaluating the effect of immunosup-pressants therapy.
Flow cytometry produces a pure CD31�/HLA-DR� popu-lation of cells; we therefore regard the loss of CD31 andHLA-DR expression in cultured RMEC as a phenotypicchange rather than an overgrowth of contaminating non-endo-thelial cells. In addition, the inability to identify HLA-DRexpressing CD14 and CD19 cells within the RMEC populationwith multicolor flow cytometry indicates that DR-expressingRMEC are not of myelomonocytic or B-lymphocytic origin.The potential for isolating cells other than RMEC is increasedwhen isolation is based on Ig-coated magnetic bead selectionof CD14-depleted and CD19-depleted HLA-DR–expressing
Figure 5. �IFN induction of HLA-DR on cultured RMEC and HK-2.FACS histograms of cultured RMEC (A) and HK-2 (B) grown inindicated concentrations of �IFN (units/ml) and labeled with L243-FITC. Cells were grown with �IFN for 36 h, except where indicated(HK-2 grown 1 wk in 500 units/ml �IFN). Only viable, PI-negativecells are included in the histograms. Solid thin gray line, no �IFN;bold black line, 0.5 units/ml �IFN; bold gray line, 2 units/ml �IFN;dotted black line, 5 units/ml �IFN; dashed black line, 500 units/ml�IFN; solid thin black line, 500 units/ml �IFN for 1 wk (HK-2 only).
1344 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 1336–1348, 2003
cells, yet RMEC isolated by this technique have the samemorphology and phenotypic characteristics as RMEC isolatedby flow cytometry. This is a useful result because RMECisolation by magnetic bead techniques generates a greaterquantity of viable cells which can be used for study without invitro alterations.
Endothelial cells are known to be phenotypically heteroge-neous in a manner that varies not only with vessel size (31,32)but also within microvascular populations (33–37). Our resultssupport this: RMEC constitutively express HLA-DR in vivo,whereas endothelial cells from larger vessels do not; lowerlevels of CD31 on cultured RMEC distinguish them fromHUVEC and cultured human lung microvascular endothelialcells; vWF expression is variable within a population of RMECthat otherwise have a uniform phenotype. Cardiac microvas-cular endothelial cells have a phenotype similar to RMEC withrespect to HLA-DR expression in that endothelial cells of smallbut not large cardiac vessels express HLA-DR; and HLA-DRexpression on isolated cardiac microvascular endothelial cellsis lost after 2 wk of culture (22,38).
The capacity of RMEC to effectively present antigenic pep-
Figure 6. CIITA RT-PCR of freshly isolated RMEC. (A) The fourdifferent promoters and first exons for CIITA are diagrammed indi-cating location of the specific forward primers (FP) and a commonreverse primer (RP) used to detect the promoter-specific CIITA. Thepredicted nucleotide lengths of the promoter-specific CIITA RT-PCRproducts are indicated. Promoter-specific RT-PCR products wereamplified from cDNA generated with oligo dT or with an oligonu-cleotide common to all CIITA, approximately 1500 bp from the 5'-endof the CIITA mRNA (RT nt). Locations of promoter III-specific andpromoter IV-specific oligonucleotides used for the southern blots (Cand D) are indicated (III nt*; IV nt**). Sequence of the diagrammedoligonucleotides appears in Materials and Methods. RT, reverse tran-scriptase. (B) CIITA internal primers (lanes 1 through 4) amplify aproduct common to all CIITA in cells that express HLA class II genes(lanes 2 through 4). A forward primer specific for promoter III CIITAand a common reverse primer (lanes 5 through 8) amplify a 904-bpproduct specific for CIITA promoter III in B lymphoblastoid cells(lane 7). A forward primer specific for promoter IV CIITA and acommon reverse primer (lanes 9 through 12) amplify a 801-bp prod-uct specific for CIITA promoter IV in �IFN-induced HK-2 (lane 10)and RMEC freshly isolated from kidney (lane 12). GAPDH amplifi-cation (lanes 13 through 16) serves as a positive control for intactcDNA and active polymerase. Oligo dT–generated cDNAs were usedfor the RT-PCR. HK-2 (lanes 1, 5, 9, 13); HK-2 � �IFN (lanes 2, 6,10, 14); B lymphoblastoid cells (lanes 3, 7, 11, 15); freshly isolatedRMEC (lanes 4, 8, 12, 16). (C and D) CIITA promoter southern blotsconfirm the presence of CIITA promoter IV in freshly isolated
Figure 7. Atorvastatin inhibition of �IFN-induced HLA-DR on cul-tured RMEC. Flow cytometry histograms of cultured RMEC grownwithout �IFN (solid gray line), with �IFN (dotted gray line), with�IFN � 1 �M atorvastatin (dotted black line), and with �IFN � 10�M atorvastatin (bold black line). �IFN concentration was 10 units/ml. Cells were evaluated 48 h after addition of reagents and onlyviable, PI-negative cells are included in histograms.
RMEC. CIITA RT-PCR products generated with primers to promoterIII (C) or promoter IV (D) were transferred to a nylon membrane andhybridized with radiolabeled promoter-specific oligonucleotideprobes (A). *III and **IV indicate specificity of promoter probe usedfor hybridization. ThM, negative control for CIITA expression (lanes1, 5); ThM � �IFN, positive control for CIITA promoter IV expres-sion (lanes 2, 6); B lymphoblastoid cells, positive control for CIITApromoter III expression (lanes 3, 7); freshly isolated RMEC (lanes 4,8). CIITA cDNAs used for the RT-PCRs were generated from acommon CIITA-specific oligonucleotide primer.
J Am Soc Nephrol 14: 1336–1348, 2003 HLA-DR–Expressing RMEC 1345

tides resulting in T cell activation cannot be predicted from thecostimulatory factors evaluated in this study. Whereas CD80and CD86, which provide the major costimulation forunprimed T cell responses, are lacking on RMEC, CD58,which has been associated with HLA-DR–initiated T cell ac-tivation in allogeneic assays with cardiac microvascular endo-thelial cells (39), is abundant on RMEC. CD40 is not normallyexpressed on RMEC in vivo, but a low level of CD40 isconsistently induced by unknown factors in our culturedRMEC. Studies are underway to evaluate the role of RMEC inpresentation of antigenic peptides. Such information will beuseful for determining whether HLA-DR, expressed so abun-dantly on RMEC, contributes to inflammation or tolerance inhuman kidneys. Knowledge of the role of RMEC HLA-DRwill influence how DR levels should be manipulated to curtailrenal inflammation.
Freshly isolated RMEC express all three HLA class II iso-types, DR, DP, and DQ, although the level of HLA-DQ ex-pression varies with the donor. For other cultured parenchymalcells we have evaluated that require �IFN induction to expressclass II (renal proximal tubular cells, mesangial cells, andmelanoma cells), HLA-DQ is detected only after prolongedhigh concentrations of �IFN; HLA-DR and HLA-DP are gen-erally expressed within 48 h. It may be that the variable levelsof HLA-DQ expressed on RMEC in vivo reflect variable con-centrations of circulating �IFN in the donors.
CIITA mRNA is present in freshly isolated human renalperitubular and glomerular capillary endothelial cells, where itsencoded protein likely mediates HLA-DR expression. There-fore, a CIITA-independent mechanism for HLA class II ex-pression, as had been described for endothelial cells (13), doesnot need to be invoked to explain the in vivo expression ofRMEC HLA-DR.
Although HLA-DR expression on RMEC appears constitu-tive in normal human kidneys, only the promoter IV form ofCIITA, associated with �IFN induction of class II genes, isamplified from freshly isolated RMEC-derived cDNA. Thissuggests that RMEC HLA-DR expression may require circu-lating or locally produced �IFN. We do not believe the tumorspresent in the kidneys used in this study provided this �IFN,because RMEC from kidneys with benign masses also ex-pressed HLA-DR. In addition, our studies with cultured RMECindicate that in vitro induction of CIITA and HLA-DR can beachieved with basal physiologic concentrations of �IFN thatare normally present in circulating blood.
Our studies examine the regulation of HLA-DR expressiononly in normal human kidneys. Under conditions of renalinflammation, including glomerulonephritis and acute rejectionof transplanted kidneys, HLA-DR is also expressed on renalproximal tubule cells (1,40–42). HLA-DR regulation has notbeen evaluated in these states of renal inflammation. Cellculture studies with the HK-2 proximal tubule cell line, andsimilar studies with primary cultures of renal proximal tubulecells, show that HLA class II expression can be induced withprolonged high concentrations of �IFN. It may be the level of�IFN that determines which cells in the kidney express class IIproteins. This would imply the existence of a �IFN sensing
“meter” to regulate class II transcription. The duration ofactivation of Stat1, which transduces the �IFN signal leadingto CIITA transcription, might be such a meter (43). Consistentwith the hypothesis that �IFN levels determine in vivoHLA-DR expression is the finding of increased �IFN mRNAlevels in rejecting transplanted kidneys where proximal tubulecells express class II (44).
Alternatively, HLA class II induction on renal parenchymalcells other than RMEC may require cytokines in addition to�IFN, e.g., tumor necrosis factor alpha (TNF-�). TNF-� syn-ergizes with �IFN to increase class II expression (45,46) buthas no effect on CIITA promoter activity (47). Elevated serumlevels of TNF-� appear 2 to 3 d before acute rejection oftransplanted kidneys (48). While the regulatory mechanism ofinduced HLA-DR expression under conditions of renal inflam-mation remains speculative, the absence of HLA-DR expres-sion on tubule cells in normal kidney tissue indicates that thelocal concentration of �IFN sensed in normal kidneys is low,yet adequate, for RMEC HLA-DR expression.
The in vivo use of CIITA promoter IV for RMEC HLA-DRexpression raises the possibility that statins could provide apharmacologic means for regulating RMEC class II. We dem-onstrate that the HMG-CoA reductase inhibitor atorvastatinreduces �IFN-induced RMEC DR expression in vitro; how-ever, our donor population did not allow us to answer thequestion of whether or not statins affect RMEC HLA-DRexpression in vivo, because none of the donors were takingstatins at the time of nephrectomy. Statins have been associatedwith a reduced rate of rejection in kidney transplants (49,50),although the mechanism for this effect has not been delineated.Although it is possible that statins may affect HLA class IIexpression on RMEC, the benefit of this under normal physi-ologic conditions depends on whether or not RMEC are capa-ble of promoting an inflammatory response. It is possible thatthe value of statins in reducing kidney rejection may be incontrolling the induced expression of HLA class II proteins onnon-RMEC that occurs with renal inflammation.
AcknowledgmentsFunding for this work was provided by grants to K. A. Muczynski
from the Northwest Kidney Centers (Seattle, WA) and the Atorvasta-tin Research Awards, and by NIH RO1 AI30527 awarded to TomCotner. We are indebted to the urologists of the University of Wash-ington Medical Centers for their assistance with procurement of renaltissue.
References1. Muczynski KA, Cotner T, Anderson SK: Unusual expression of
human lymphocyte antigen class II in normal renal microvascu-lar endothelium. Kidney Int 59: 488–497, 2001
2. Croft M, Dubey C: Accessory molecule and costimulation re-quirements for CD4 T cell response. Crit Rev Immunol 17:89–118, 1997
3. Hagerty DT, Evavold BD, Allen PM: Regulation of the costimu-lator B7, not class II major histocompatibility complex, restrictsthe ability of murine kidney tubule cells to stimulate CD4� Tcells. J Clin Invest 93: 1208–1215, 1994
1346 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 1336–1348, 2003

4. Singer GG, Yokoyama H, Bloom RD, Jevnikar AM, Nabavi N,Kelley VR: Stimulated renal tubular epithelial cells induce an-ergy in CD4� T cells [see comments]. Kidney Int 44: 1030–1035, 1993
5. Steimle V, Otten LA, Zufferey M, Mach B: Complementationcloning of an MHC class II transactivator mutated in hereditaryMHC class II deficiency (or bare lymphocyte syndrome). Cell75: 135–146, 1993
6. Steimle V, Siegrist CA, Mottet A, Lisowska-Grospierre B, MachB: Regulation of MHC class II expression by interferon-gammamediated by the transactivator gene CIITA. Science 265: 106–109, 1994
7. Chang CH, Fontes JD, Peterlin M, Flavell RA: Class II transac-tivator (CIITA) is sufficient for the inducible expression of majorhistocompatibility complex class II genes. J Exp Med 180: 1367–1374, 1994
8. Steimle V, Durand B, Barras E, Zufferey M, Hadam MR, MachB, Reith W: A novel DNA-binding regulatory factor is mutatedin primary MHC class II deficiency (bare lymphocyte syn-drome). Genes Dev 9: 1021–1032, 1995
9. Masternak K, Barras E, Zufferey M, Conrad B, Corthals G,Aebersold R, Sanchez JC, Hochstrasser DF, Mach B, Reith W: Agene encoding a novel RFX-associated transactivator is mutatedin the majority of MHC class II deficiency patients. Nat Genet20: 273–277, 1998
10. Nagarajan UM, Louis-Plence P, DeSandro A, Nilsen R, BusheyA, Boss JM: RFX-B is the gene responsible for the most commoncause of the bare lymphocyte syndrome, an MHC class II im-munodeficiency [published erratum appears in Immunity 1999Mar;10(3):399]. Immunity 10: 153–162, 1999
11. Durand B, Sperisen P, Emery P, Barras E, Zufferey M, Mach B,Reith W: RFXAP, a novel subunit of the RFX DNA bindingcomplex is mutated in MHC class II deficiency. Embo J 16:1045–1055, 1997
12. Harton JA, Ting JP: Class II transactivator: Mastering the art ofmajor histocompatibility complex expression. Mol Cell Biol 20:6185–6194, 2000
13. Collinge M, Pardi R, Bender JR: Class II transactivator-indepen-dent endothelial cell MHC class II gene activation induced bylymphocyte adhesion. J Immunol 161: 1589–1593, 1998
14. Muhlethaler-Mottet A, Otten LA, Steimle V, Mach B: Expres-sion of MHC class II molecules in different cellular and func-tional compartments is controlled by differential usage of mul-tiple promoters of the transactivator CIITA. Embo J 16: 2851–2860, 1997
15. Lennon AM, Ottone C, Rigaud G, Deaven LL, Longmire J,Fellous M, Bono R, Alcaide-Loridan C: Isolation of a B-cell-specific promoter for the human class II transactivator. Immuno-genetics 45: 266–273, 1997
16. Deffrennes V, Vedrenne J, Stolzenberg MC, Piskurich J, BarbieriG, Ting JP, Charron D, Alcaide-Loridan C: Constitutive expres-sion of MHC class II genes in melanoma cell lines results fromthe transcription of class II transactivator abnormally initiatedfrom its B cell-specific promoter. J Immunol 167: 98–106, 2001
17. Goodwin BL, Xi H, Tejiram R, Eason DD, Ghosh N, Wright KL,Nagarajan U, Boss JM, Blanck G: Varying functions of specificmajor histocompatibility class II transactivator promoter III andIV elements in melanoma cell lines. Cell Growth Differ 12:327–335, 2001
18. Honey K, Rudensky A: The pIV-otal class II transactivatorpromoter regulates major histocompatibility complex class IIexpression in the thymus. J Exp Med 194: F15–F18, 2001
19. Waldburger JM, Suter T, Fontana A, Acha-Orbea H, Reith W:Selective abrogation of major histocompatibility complex class IIexpression on extrahematopoietic cells in mice lacking promoterIV of the class II transactivator gene. J Exp Med 194: 393–406,2001
20. Folkman J, Haudenschild CC, Zetter BR: Long-term culture ofcapillary endothelial cells. Proc Natl Acad Sci USA 76: 5217–5221, 1979
21. Gerritsen ME, Shen CP, McHugh MC, Atkinson WJ, Kiely JM,Milstone DS, Luscinskas FW, Gimbrone MA Jr: Activation-dependent isolation and culture of murine pulmonary microvas-cular endothelium. Microcirculation 2: 151–163, 1995
22. McDouall RM, Batten P, McCormack A, Yacoub MH, Rose ML:MHC class II expression on human heart microvascular endo-thelial cells: Exquisite sensitivity to interferon-gamma and nat-ural killer cells. Transplantation 64: 1175–1180, 1997
23. Kwak B, Mulhaupt F, Myit S, Mach F: Statins as a newlyrecognized type of immunomodulator. Nat Med 6: 1399–1402,2000
24. Sadeghi MM, Tiglio A, Sadigh K, O’Donnell L, Collinge M,Pardi R, Bender JR: Inhibition of interferon-gamma-mediatedmicrovascular endothelial cell major histocompatibility complexclass II gene activation by HMG-CoA reductase inhibitors.Transplantation 71: 1262–1268, 2001
25. Lampson LA, Levy R: Two populations of Ia-like molecules ona human B cell line. J Immunol 125: 293–299, 1980
26. Watson AJ, DeMars R, Trowbridge IS, Bach FH: Detection of anovel human class II HLA antigen. Nature 304: 358–361, 1983
27. Spits H, Borst J, Giphart M, Coligan J, Terhorst C, De Vries JE:HLA-DC antigens can serve as recognition elements for humancytotoxic T lymphocytes. Eur J Immunol 14: 299–304, 1984
28. Salter RD, Howell DN, Cresswell P: Genes regulating HLA classI antigen expression in T-B lymphoblast hybrids. Immunogenet-ics 21: 235–246, 1985
29. Muczynski KA, Anderson SK, Pious D: Discoordinate surfaceexpression of IFN-gamma-induced HLA class II proteins innonprofessional antigen-presenting cells with absence of DMand class II colocalization. J Immunol 160: 3207–3216, 1998
30. Ryan MJ, Johnson G, Kirk J, Fuerstenberg SM, Zager RA,Torok-Storb B: HK-2: An immortalized proximal tubule epithe-lial cell line from normal adult human kidney. Kidney Int 45:48–57, 1994
31. Page C, Rose M, Yacoub M, Pigott R: Antigenic heterogeneity ofvascular endothelium. Am J Pathol 141: 673–683, 1992
32. Kinjo T, Takashi M, Miyake K, Nagura H: Phenotypic hetero-geneity of vascular endothelial cells in the human kidney. CellTissue Res 256: 27–34, 1989
33. Belloni PN, Tressler RJ: Microvascular endothelial cell hetero-geneity: Interactions with leukocytes and tumor cells. CancerMetastasis Rev 8: 353–389, 1990
34. Belloni PN, Carney DH, Nicolson GL: Organ-derived microves-sel endothelial cells exhibit differential responsiveness to throm-bin and other growth factors. Microvasc Res 43: 20–45, 1992
35. Lehmann I, Brylla E, Sittig D, Spanel-Borowski K, Aust G:Microvascular endothelial cells differ in their basal and tumournecrosis factor-alpha-regulated expression of adhesion moleculesand cytokines. J Vasc Res 37: 408–416, 2000
36. Petzelbauer P, Bender JR, Wilson J, Pober JS: Heterogeneity ofdermal microvascular endothelial cell antigen expression andcytokine responsiveness in situ and in cell culture. J Immunol151: 5062–5072, 1993
J Am Soc Nephrol 14: 1336–1348, 2003 HLA-DR–Expressing RMEC 1347

37. Rupnick MA, Carey A, Williams SK: Phenotypic diversity incultured cerebral microvascular endothelial cells. In Vitro CellDev Biol 24: 435–444, 1988
38. McDouall RM, Yacoub M, Rose ML: Isolation, culture, andcharacterization of MHC class II-positive microvascular endo-thelial cells from the human heart. Microvasc Res 51: 137–152,1996
39. McDouall RM, Page CS, Hafizi S, Yacoub MH, Rose ML:Alloproliferation of purified CD4� T cells to adult human heartendothelial cells, and study of second-signal requirements. Im-munology 89: 220–226, 1996
40. Bishop GA, Hall BM, Duggin GG, Horvath JS, Sheil AG, TillerDJ: Immunopathology of renal allograft rejection analyzed withmonoclonal antibodies to mononuclear cell markers. Kidney Int29: 708–717, 1986
41. Hall BM, Bishop GA, Duggin GG, Horvath JS, Philips J, TillerDJ: Increased expression of HLA-DR antigens on renal tubularcells in renal transplants: Relevance to the rejection response.Lancet 2: 247–251, 1984
42. Muller CA, Markovic-Lipkovski J, Risler T, Bohle A, MullerGA: Expression of HLA-DQ, -DR, and -DP antigens in normalkidney and glomerulonephritis. Kidney Int 35: 116–124, 1989
43. Eason DD, Blanck G: High level class II trans-activator induc-tion does not occur with transient activation of the IFN-gammasignaling pathway. J Immunol 166: 1041–1048, 2001
44. Kirk AD, Bollinger RR, Finn OJ: Rapid, comprehensive analysisof human cytokine mRNA and its application to the study ofacute renal allograft rejection. Hum Immunol 43: 113–128, 1995
45. Leeuwenberg JF, Van Damme J, Meager T, Jeunhomme TM,Buurman WA: Effects of tumor necrosis factor on the interferon-gamma-induced major histocompatibility complex class II anti-gen expression by human endothelial cells. Eur J Immunol 18:1469–1472, 1988
46. Wedgwood JF, Hatam L, Bonagura VR: Effect of interferon-gamma and tumor necrosis factor on the expression of class I andclass II major histocompatibility molecules by cultured humanumbilical vein endothelial cells. Cell Immunol 111: 1–9, 1988
47. Nikcevich KM, Piskurich JF, Hellendall RP, Wang Y, Ting JP:Differential selectivity of CIITA promoter activation by IFN-gamma and IRF-1 in astrocytes and macrophages: CIITA pro-moter activation is not affected by TNF-alpha. J Neuroimmunol99: 195–204, 1999
48. Kutukculer N, Shenton BK, Clark K, Rigg KM, Forsythe JL,Kirby JA, Proud G, Taylor RM: Renal allograft rejection: thetemporal relationship and predictive value of plasma TNF (alphaand beta), IFN-gamma and soluble ICAM-1. Transpl Int 8:45–50, 1995
49. Katznelson S, Wilkinson AH, Kobashigawa JA, Wang XM, ChiaD, Ozawa M, Zhong HP, Hirata M, Cohen AH, Teraski PI, et al:The effect of pravastatin on acute rejection after kidney trans-plantation—A pilot study. Transplantation 61: 1469–1474, 1996
50. Tuncer M, Suleymanlar G, Ersoy FF, Yakupoglu G: Comparisonof the effects of simvastatin and pravastatin on acute rejectionepisodes in renal transplant patients. Transplant Proc 32: 622–625, 2000
1348 Journal of the American Society of Nephrology J Am Soc Nephrol 14: 1336–1348, 2003