norfloxacin binds to human fecal material
TRANSCRIPT

Vol. 32, No. 12ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Dec. 1988, p. 1869-18740066-4804/88/121869-06$02.00/0Copyright © 1988, American Society for Microbiology
Norfloxacin Binds to Human Fecal MaterialCHARLOTTA EDLUND, LENNART LINDQVIST, AND CARL ERIK NORD*
Department of Microbiology, Karolinska Institute, Huddinge University Hospital, and National BacteriologicalLaboratory,* Stockholm, Sweden
Received 28 June 1988/Accepted 21 September 1988
Earlier studies have reported very high (120 to 2,700 mg/kg) concentrations of norfloxacin in feces aftertherapeutic doses. MICs for fecal microorganisms are with few exceptions far below these levels. Nevertheless,clinical investigations show that the main part of the aerobic gram-positive and the anaerobic microfloraremains unaffected after norfloxacin administration. In this study, the binding of [14Cjnorfloxacin to fecalmaterial was analyzed. The binding of a group of nonlabeled quinolones to feces and the interactions betweenEnterococcus faecium, Bacteroides fragilis, and norfloxacin were also investigated. The results showed thatnorfloxacin has the ability to bind to feces. The specific binding was reversible, saturated after 90 min ofincubation at 37°C, and increased linearly with fecal concentration. Scatchard plots and nonlinear regressioncomputer analyses revealed two different binding classes. The primary specific binding had a dissociationconstant (KD) of 1.0 ,uM and a maximal binding capacity (Bma.) of 0.12 ,umol/g of feces. The KD and Bmax ofthe secondary, more unspecific binding were 450 ,uM and 11.8 p,mol/g of feces, respectively. The binding ofunlabeled ciprofioxacip, enoxacin, ofloxacin, pefloxacin, and norfloxacin to feces was comparable to that of['4C]norfloxacin. The results of norfloxacin binding to suspensions of B. fragilis suggested that the main partof the binding is to the bacterial fraction of feces. In the presence of 8.0 g (dry weight) of B. fragilis per liter,the MBC of norfloxacin for E. faecium increased from 8 to 256 p,g/ml. The findings of the present studyindicated that binding of norfloxacin to feces may explain the paradox of high fecal concentrations ofnorfloxacin versus the actual effect on the normal gastrointestinal microflora.
Previous studies on the newer quinolones have shown thatadministration of these agents causes selective ecologicaldisturbances in the gastrointestinal flora (11, 20). Severalinvestigations including healthy volunteers as well as dif-ferent categories of patients on the impact of norfloxacin onthe normal gastrointestinal microflora showed uniform re-sults (2, 9, 17, 18). The aerobic gram-negative microorgan-isms are eliminated or strongly suppressed during adminis-tration, while the aerobic gram-positive and the anaerobicmicrofloras are unaffected. These favorable ecological prop-erties of norfloxacin contribute to a preservation of "colo-nization resistance" (25) and thus make norfloxacin suitablefor long-term prophylactic treatment of urinary tract infec-tions, selective decontamination in immunocompromisedpatients, and treatment of bacterial intestinal infections.However, very high concentrations of norfloxacin are
achieved in feces. Peak fecal concentrations of norfloxacinof 120 to 2,700 mg/kg after therapeutic doses have beenreported (5, 9, 18). MICs for both aerobic and anaerobicmicroorganisms are with few exceptions far below theselevels (10, 19, 22). Thus, despite the high levels of norflox-acin in feces, the main part of the aerobic gram-positive andthe anaerobic flora remains unaffected after norfloxacinadministration. Different theories have been raised to ex-plain this paradox. Differences in anaerobiosis of the colonenvironment in vivo compared with the in vitro conditionshave been suggested to influence the sensitivity of anaerobicbacteria. An inoculum effect on the activity of norfloxacinhas been shown for anaerobic strains (12). At an inoculum of109 CFU/ml, norfloxacin concentrations greater than 512 ,ug/ml are required to reduce most gram-positive and gram-negative fecal anaerobes to less than 103 CFU/ml. The third
* Corresponding author.
theory suggests that there is a binding between norfloxacinand fecal material. Reversible binding to feces has been seenfor other antimicrobial agents (13, 14). The influence of feceson the antimicrobial activity of norfloxacin has been studied(26), but to our knowledge no study has characterized thebinding of quinolones to feces.The aim of the present study was to analyze the binding of
[14C]norfloxacin to fecal material and Bacteroidesfragilis; todetermine the binding of a group of unlabeled quinolones tofeces; and to investigate interactions between Enterococcusfaecium, B. fragilis, and norfloxacin.
MATERIALS AND METHODSChemicals. Norfloxacin (batch 69588-01) was obtained
from Astra Alab AB, Sodertalje, Sweden. Ciprofloxacin wasobtained from Bayer AG, Wuppertal, Federal Republic ofGermany; enoxacin was from Warner Lambert, Ann Arbor,Mich.; ofloxacin was from Hoechst, Frankfurt, FederalRepublic of Germany; and pefloxacin was from LaboratoireRoger Bellon, Paris, France. Piperazine-[U-_4C]norfloxacinwas synthesized by Merck Sharp & Dohme, Rahway, N.J.The labeled drug had a specific radioactivity of 14.85 mCi/mmol. Stock solutions were prepared in 0.1 M sodiumphosphate buffer (pH 7.2) and stored at -70°C.
Feces. Feces from one female volunteer (age, 30 years)were used throughout the study (Fl). In two additionalexperiments, feces from two other volunteers (males aged 35and 48 years, respectively) were used (F2 and F3). Freshfecal samples were diluted in 0.1 M sodium phosphate buffer(pH 7.2) to a 5% (wt/vol) suspension, mixed by rnagneticstirring at 4°C for 1 h, and then filtered through two layers ofcoarse gauze and stored at -700C until assayed. The dryweights of Fl, F2, and F3 were 16.3, 11.2, and 16.9% of thewet weight, respectively. The dry weights of the filtered
1869
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 24
Oct
ober
202
1 by
280
4:30
c:81
e:46
01:6
d4f:
c763
:cac
9:c0
4b.
ANTIMICROB. AGENTS CHEMOTHER.
TABLE 1. Microbial composition of feces from three volunteers(Fl, F2, and F3)
Log1o bacteria/g of fecesBacteria
Fl F2 F3
Enterococci 5.1 5.6 5.4Alpha-streptococci 5.8 5.5 5.7Bacillus sp. 4.4 4.0 2.7Escherichia coli 4.6 4.8 4.6Klebsiella pneumoniae 5.6 2.6Citrobacter freundii 4.6
Anaerobic cocci 8.0 3.0 5.7Lactobacilli 5.2 6.1 7.5Bifidobacteria 6.3 8.0 6.2Clostridia 7.3 4.8 4.7Bacteroides sp. 9.9 9.8 9.4Veillonella sp. 3.0 5.8 4.8
samples were all 60% of the dry weight of correspondingunfiltered feces. The microbial compositions of samples Fl,F2, and F3 are shown in Table 1.
Bacterial strains. B. fragilis (MIC, 128 ,ug/ml), was isolatedfrom Fl and incubated anaerobically on blood agar plates at37°C for 48 h. The colonies were suspended in 0.1 M sodiumphosphate buffer (pH 7.2) to a dry weight of 4.7 g/liter whenused for the binding assay, and they were suspended inprereduced peptone-yeast extract medium when used for theMBC assay.A clinical strain of E. faecium (MIC, 4 ,ug/ml) diluted in
brain heart infusion broth to a final concentration of 106CFU/ml was used for MBC determinations.
Binding experiments. Norfloxacin binding was measuredby a membrane filter technique. The reaction mixture con-tained 0.32 ml of the filtered fecal suspension, unless other-wise stated, and 1.5 ,uM [14C]norfloxacin in a total volume of3.0 ml. The specific binding was determined by the differ-ence between total radioactivity found in the absence orpresence of excess of unlabeled norfloxacin (560 ,uM). Bind-ing studies were also done with various concentrations of[14C]norfloxacin (0.1 to 500 ,uM).The binding of five nonlabeled quinolones (ciprofloxacin,
enoxacin, ofloxacin, pefloxacin, and norfloxacin) was deter-mined by using their ability to compete with binding of 1.5,uM [14C]norfloxacin to feces.
Binding of norfloxacin to B. fragilis, one of the predomi-nating microorganisms in feces, was studied with[14C]norfloxacin (0.1 to 500 ,M) and 0.32 ml of the B. fragilissuspension. The final concentration of B. fragilis was 0.5 g(dry weight) per liter.The reaction mixtures were incubated for various times at
37°C during end-over-end rotation before filtration. All de-terminations were made in triplicate samples. From eachsample, three portions (0.5 to 1.0 ml) were filtered undervacuum (Uni-Pore model 25 filter holder; Bio-Rad Labora-tories, Richmond, Calif.) through 1.0-,m-pore-size glassfiber filters (GF/B; Whatman, Ltd., Maidstone, England).After being washed three times with 2 ml of 0.1 M sodiumphosphate buffer (4°C, pH 7.2) to remove unbound labeleddrug, the filters were counted for radioactivity in a liquidscintillation spectrometer at 87% efficiency (counts perminute/disintegrations per minute). The coefficient of varia-tion of the triplicate samples was within 10% in all experi-ments.
Evaluation of binding data. The equilibrium dissociationconstant (KD) and the total number of binding sites (Bmax)
z
4x0
mL-
0zaz00U
CO
100
90
80
70
60
50
40
30
20
10
0
5
4
m~30
-12c- I
0
-1I . I I
0 30 min 60 90
0 30 60 90 120 150 180TIME (min)
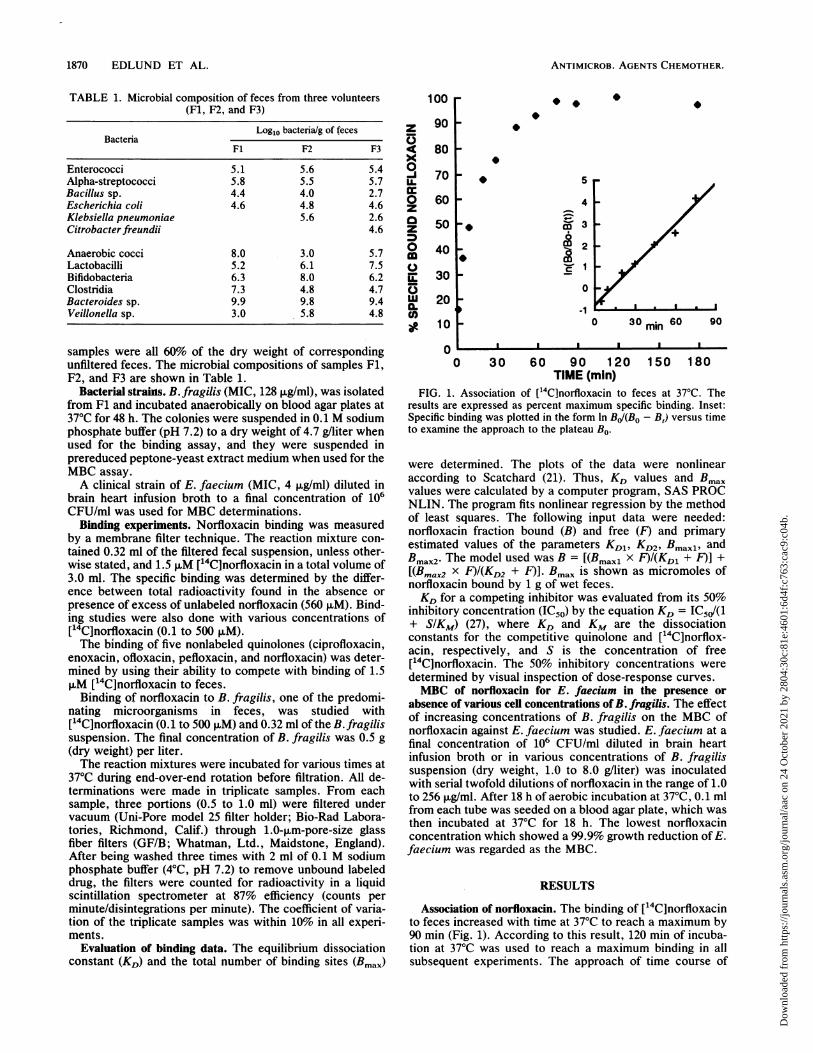
FIG. 1. Association of ['4C]norfloxacin to feces at 37°C. Theresults are expressed as percent maximum specific binding. Inset:Specific binding was plotted in the form In BJ/(Bo - B,) versus timeto examine the approach to the plateau Bo.
were determined. The plots of the data were nonlinearaccording to Scatchard (21). Thus, KD values and Bmaxvalues were calculated by a computer program, SAS PROCNLIN. The program fits nonlinear regression by the methodof least squares. The following input data were needed:norfloxacin fraction bound (B) and free (F) and primaryestimated values of the parameters KDl, KD2, Bmaxi, andBmax2. The model used was B = [(Bmaxi X F)/(KD1 + F)] +[(Bmax2 x F)I(KD2 + F)]. Bmax is shown as micromoles ofnorfloxacin bound by 1 g of wet feces.KD for a competing inhibitor was evaluated from its 50%
inhibitory concentration (IC50) by the equation KD = IC5d(1+ SIKM) (27), where KD and KM are the dissociationconstants for the competitive quinolone and ['4C]norflox-acin, respectively, and S is the concentration of free['4C]norfloxacin. The 50% inhibitory concentrations weredetermined by visual inspection of dose-response curves.MBC of norfloxacin for E. faecium in the presence or
absence of various cell concentrations of B. fragilis. The effectof increasing concentrations of B. fragilis on the MBC ofnorfloxacin against E. faecium was studied. E. faecium at afinal concentration of 106 CFU/ml diluted in brain heartinfusion broth or in various concentrations of B. fragilissuspension (dry weight, 1.0 to 8.0 g/liter) was inoculatedwith serial twofold dilutions of norfloxacin in the range of 1.0to 256 ,ug/ml. After 18 h of aerobic incubation at 37°C, 0.1 mlfrom each tube was seeded on a blood agar plate, which wasthen incubated at 37°C for 18 h. The lowest norfloxacinconcentration which showed a 99.9% growth reduction of E.faecium was regarded as the MBC.
RESULTS
Association of norfloxacin. The binding of [14C]norfloxacinto feces increased with time at 37°C to reach a maximum by90 min (Fig. 1). According to this result, 120 min of incuba-tion at 37°C was used to reach a maximum binding in allsubsequent experiments. The approach of time course of
1870 EDLUND ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 24
Oct
ober
202
1 by
280
4:30
c:81
e:46
01:6
d4f:
c763
:cac
9:c0
4b.

NORFLOXACIN BINDING TO FECES 1871
100 a a * i80 x
z X5 60 x4x940
0z0zM 200
0
*mLL
* *o oA A
x
0
aA
x
x
180 210 240
FIG. 2. Dissociation of 1.5 p.M ['4C]norfloxacin bound to fecesat various temperatures. The results are expressed as the percentageof the specific binding found at the time zero (logarithmic scale).Symbols: *, 4°C; O, 20°C; A, 37°C; x, 37°C. (Plus 560 ,uMnorfloxacin.)
specific binding (B,) to the steady-state level observed (BO)was-examined by plotting the function In BJ(BO - B,) againsttime (27). This gave a straight line consistent with a pseudofirst-order reaction, with an apparent half-time of 15 min.
Dissociation of norfloxacin. The dissociation of [14C]nor-floxacin bound to feces was examined by incubating 5.3 g offeces per liter with 1.5 ,uM [14C]norfloxacin for 120 min in thepresence or absence of 560 ,uM unlabeled norfloxacin. Thesamples were then centrifuged and washed three times with0.1 M sodium phosphate buffer (pH 7.2) at 4°C, and theresuspended samples were further incubated for up to 240min at various temperatures. Binding of norfloxacin to feceswas reversible and temperature dependent (Fig. 2). After 240min, 60 and 35% were dissociated in the presence andabsence, respectively, of unlabeled norfloxacin. At 4°C, only10% of the bound norfloxacin was dissociated after 240 min.Suspension of the samples in 0.1 M NaOH resulted in >95%dissociation within 3 min.
Binding of ['4C]norfloxacin to increasing concentrations offecal solutions. The binding of [14C]norfloxacin to humanfeces was found to be a linear function of feces concentration(Fig. 3). At a fecal concentration of 5.3 g/liter, the specificbinding represented greater than 80% of the total bindingobserved. The same assay was also done with 0.22-,um-pore-size filters (GS; Millipore Corp., Bedford, Mass.), whichyielded similar results. Owing to a shorter time needed forsamples to penetrate the 1.0-p.m-pore-size filters, thesefilters were used in subsequent assays. Binding to feceswhich were centrifuged and washed three times before theassay was also determined. Washed and unwashed fecesbound equal amounts of norfloxacin, which shows thatnorfloxacin binds to the solid part of feces.
Binding of increasing amounts of [14C]norfloxacin to feces.The ability of increasing concentrations of [V4C]norfloxacin(0.1 to 500 ,uM) to bind to feces was assayed. Scatchard plotsof the data were nonlinear, which means that more than oneclass of binding was involved (Fig. 4). Computer fitting of thedata assumed two binding classes, one with high affinity andlow capacity (class 1) and one with low affinity and highcapacity (class 2). This assay was also performed for twoadditional fecal samples, F2 and F3. The values of thenumerical estimates of the parameters KD1, KD2, B maxl, and
^ 04S
A .0O 0,3E
zX r)
X 0,20-J
0° 01z
0*.L O_ . _
cm0,0
0 2 4 6 8FECES (g/1)
FIG. 3. Binding of 1.5 ,uM [14C]norfloxacin to an increasingconcentration of feces (0 to 8.3 g/liter). Symbols: *, specificbinding; x, nonspecific binding.
Bmax2 are presented in Table 2. The parameters were used togenerate a curve relating fraction of bound drug to totalnorfloxacin concentration, extrapolating to 100% feces, as-suming that all binding material passed the gauze (Fig. 5).
Competitive binding studies. The KD values of high-affinitybinding to feces for five nonlabeled quinolones (ciproflo-xacin, enoxacin, ofloxacin, pefloxacin, and norfloxacin)were 3.4, 2.4, 2.3, 2.9, and 1.4 1iM, respectively, obtained bythe indirect competition method. All unlabeled quinolonesalso had a secondary, unspecific binding corresponding withthe binding of [14C]norfloxacin obtained in the present studyby the direct method.
Binding of norfloxacin to B. fragilis. The apparent KD andBmax values were determined for B. fragilis with [14C]norfloxacin (0.1 to 500 ,uM). KD1 and KD2 were estimated tobe 2.4 and 1,140 ,iM, respectively, and B,max, and Bmax2 wereestimated to be 0.16 and 19.6 ,umol/g, respectively.
0,14
0,12
0,10'
0,08IL
0,06
0,04
0,02
0,000 1 2 3 4 5 6
BOUND NORFLOXACIN (pmoVg feces)7
FIG. 4. Scatchard plot for [14C]norfloxacin binding to feces (Fl).The asymptotes as well as the curve combining the experimentaldata points were drawn by the computer program SAS PROC NLINand represent the best fit. B, Norfloxacin fraction bound; F,norfloxacin fraction free.
0 30 60 90 120 150TIME (mln)
VOL. 32, 1988
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 24
Oct
ober
202
1 by
280
4:30
c:81
e:46
01:6
d4f:
c763
:cac
9:c0
4b.

ANTIMICROB. AGENTS CHEMOTHER.
TABLE 2. Dissociation constants (KD) and maximal bindingcapacities (Bmax) for binding of ["4C]norfloxacin to three different
fecal specimens
i±m LmoUgSample
KDI KD2 Bxl Bmax2
Fl 1.0 450 0.12 11.8F2 0.6 830 0.03 17.2F3 1.4 450 0.10 10.7
MBC of norfloxacin for E. faecium in the presence orabsence of various ceil concentrations of B. fragiis. The effectof various cell concentrations of B. fragilis on the MBC forE. faecium was studied. The MBCs were strongly affectedby the presence of B. fragilis (Table 3).
DISCUSSION
The results presented in this report show that norfloxacinhas the ability to bind to feces. The specific binding wasreversible, saturated after 90 min of incubation at 37°C, andincreased in a linear relationship with fecal concentration.Scatchard plots and nonlinear regression computer analysesrevealed at least two different binding classes, one primary,specific binding and one secondary, more unspecific binding.When the binding of norfloxacin was studied directly orindirectly by a competitive method, very similar dissociationconstants were obtained, which supports the accuracy of themethods used. However, there seems to be a certain varia-tion between different fecal samples (Table 2). The resultsobtained by extrapolating to undiluted feces may give a hintof the clinical situation. At a norfloxacin concentration of1,000 mg/kg, which is common in clinical situations, morethan 95% of the norfloxacin is bound to feces, mainly byunspecific binding, and thus only 50 mg/kg is available asfree drug (Fig. 5). Furthermore, with such a high KD2 value,the low-affinity binding may be underestimated because offast dissociation, which may occur during filtration andwashing procedures.The binding of norfloxacin to feces might in part reflect its
mechanism of action. It has been shown that norfloxacin,
100
z
U
0-j
UI.
z
z
0
918
96
94
92100 101 1i2 10
NORFLOXACIN (mg /kg fces)FIG. 5. Fraction of bound norfloxacin related to total norfloxacin
concentration in 100% feces (extrapolated values). The experimen-tal data points and the predicted curves for each binding class derivefrom studies on Fl.
TABLE 3. Effect of various concentrations of B. fragilis on MBCof norfloxacin for E. faecium
B. fragilis (g MBC (JLg/ml)[dry wt]/liter)0 81.0 82.0 164.0 648.0 256
like other nalidixic acid analogs, inhibits the A subunit of thebacterial enzyme DNA gyrase (7, 16), although the details ofthe molecular interactions of norfloxacin are not yet fullyunderstood. The potency of quinolones as DNA gyraseinhibitors does not always correlate with antimicrobial ac-tivity (28), which means that other factors are important. Ithas been suggested that quinolones act on intact bacteriaprincipally by forming drug-enzyme-DNA complexes thatfunction as poisons rather than as simple inhibitors of DNAsupercoiling (16).
Several permeability barriers must be passed by the qui-nolones to reach the target site inside the bacteria. Lipopoly-saccharide structures in the gram-negative cell wall mayinteract with the drug and thus affect the uptake through theouter membrane. The porins of the outer membrane seem tobe the main route of entrance for quinolones with lowhydrophobicity such as norfloxacin, ciprofloxacin, enoxacin,fleroxacin, and ofloxacin, while more hydrophobic agentssuch as nalidixic acid may also penetrate through the lipidbilayer of the outer membrane (15). Intracellular compo-nents such as vesicles may also interfere with the quino-lones. Thus, many cell structures of the bacteria may inter-act with the quinolones, an interaction which could bereferred to as binding.Shen and Pernet (23) studied the binding of [3H]norflox-
acin to gyrase and substrate DNA. They showed that nor-floxacin binds to DNA but not to gyrase. The binding toplasmid DNA showed a primary process that was saturatedat a norfloxacin concentration about equal to the supercoil-ing Ki (1.8 ,uM) with an apparent KD value of 1 ,uM, which isin accordance with the KD1 found in the present study. Asecondary weaker binding followed, which corresponds toour findings. These results imply that the high-affinity bind-ing demonstrated in the present study is equivalent tobinding of norfloxacin to bacterial DNA. On the other hand,it is unclear whether the binding of norfloxacin to bacteriaactually is proportional to its antimicrobial activity. Diver etal. (8) studied the uptake of labeled ciprofloxacin, enoxacin,and pefloxacin by Escherichia coli KL16. A nonsaturableactive uptake across the cytoplasmic membrane was found.They demonstrated that the uptake of each drug did notcorrelate with antimicrobial activity. The uptake of quino-lones in E. coli has also been studied by two other groups (1,4), who concluded that the process was rapid, unsaturable,and occurred by simple energy-independent diffusion. Bed-ard et al. (1) stated that enoxacin is not bound firmly to itstarget(s) of action, since the antimicrobial effect of enoxacincould be reversed for up to 45 min by washing cells, evenafter exposure to high concentrations of enoxacin (80 ,ug/ml).These researchers further suggested that the bactericidaleffect resulting from inhibition of DNA gyrase is a processindependent of drug uptake and that the targets critical togrowth inhibition are saturated at low concentrations. Chap-man and Georgopapadakou (4) studied the association of
1872 EDLUND ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 24
Oct
ober
202
1 by
280
4:30
c:81
e:46
01:6
d4f:
c763
:cac
9:c0
4b.

NORFLOXACIN BINDING TO FECES 1873
fleroxacin to cell surfaces. A rapid binding was found whichthey referred to as fleroxacin chelating outer membrane-bound magnesium, which bridges adjacent lipopolysaccha-ride molecules. A slower, nonsaturable phase of uptake wassuggested to be the passive diffusion of fleroxacin throughboth porins and exposed lipid domains on the outer mem-brane. Cohen et al. (6) found that susceptible E. coli strainshave an energy-dependent reduced uptake of norfloxacin,which seems to be due to a carrier-mediated active effluxgenerated by proton motive force.The role of oxygen in the bactericidal effect of quinolones
was studied by Lewin et al. (C. S. Lewin, I. Morrissey, andJ. T. Smith, Abstr. 2nd Int. Symp. New Quinolones, abstr.4, 1988), and they reported that ofloxacin and ciprofloxacinexhibited only a bacteriostatic effect under anaerobic condi-tions in contrast to the bactericidal effect seen in the pres-ence of oxygen. These findings may also contribute to theminor impact on the fecal flora despite the high quinoloneconcentrations in the gut.
In the present study, we found that KD and Bmax values fora suspension of B. fragilis were in the same range as thosefor whole feces. Since it is assumed that 50% of the solid partof feces consists of bacteria (24), it seems likely that nor-floxacin binds mainly to the bacterial fraction of feces.The results of the competitive binding studies of four other
new quinolones showed that they have KD values compara-ble to that of norfloxacin. This implies that all new quinolonecompounds act in a similar way regarding their binding tofeces.
In the presence of B. fragilis (8.0 g [dry weight] per liter),the MBC against E. faecium increased from 8 to 256 ,ug/ml(Table 3). In 100% feces, the dry weight of bacteria isapproximately 10-fold greater, which most certainly leads toan MBC above the concentrations of drug reached in thecolon after clinical doses. B. fragilis was used in this assaysince it is one of the predominating microorganisms in thenormal intestinal microflora and thus reflects the presence offeces. E. faecium was chosen because enterococci are oftenin the borderline of being affected in vivo after administra-tion of clinical doses of quinolone compounds, although invitro MICs and MBCs for enterococci are in the range of 1 to16 p,g/ml.The influence of feces on the activity of norfloxacin and
ciprofloxacin has been studied by van Saene et al. (26).MBCs for six aerobic gram-negative rods incubated withincreasing fecal concentrations were determined. It wasfound that practically all strains except Pseudomonas andAcinetobacter strains were inhibited by norfloxacin andciprofloxacin, even in the presence of 30% feces. Microor-ganisms for which MBCs are very low, such as mostenterobacteria, may be less influenced by feces comparedwith those for which MBCs are moderate or high. Thistheory is supported by Goldstein et al. (12), who studied theeffect of inoculum size on in vitro activity of norfloxacinagainst fecal anaerobic bacteria. These researchers found amarked inoculum effect at 109 CFU/ml for a range ofanaerobic bacterial strains, but not for two E. coli strainstested. They stated that fecal concentrations of norfloxacingreater than 512 Fxg/ml would be required to reduce mostanaerobic microorganisms to counts of less than 103 CFU/ml. The inoculum effect might be a result of binding ofnorfloxacin to bacteria. In contrast, Borobio and Perea (3)did not find any significant effect on the activity of ciproflo-xacin for various inocula (103 to 107 CFU/ml) of 30 strains ofanaerobic bacteria.
In conclusion, the results of the present study suggest that
binding of norfloxacin to feces, including or in combinationwith an inoculum effect, explains the paradox of high fecalconcentrations of norfloxacin versus the actual effect on thenormal gastrointestinal microflora.
LITERATURE CITED1. Bedard, J., S. Wong, and L. E. Bryan. 1987. Accumulation of
enoxacin by Escherichia coli and Bacillus subtilis. Antimicrob.Agents Chemother. 31:1348-1354.
2. Boerema, J. B. J., B. J. Olthof, and H. K. F. van Saene. 1986.Effects of norfloxacin on the faecal flora in patients withcomplicated urinary tract infections. Scand. J. Infect. Dis.Suppl. 48:27-31.
3. Borobio, V., and E. Perea. 1984. Effect of inoculum, pH, andmedium on the activity of ciprofloxacin against anaerobic bac-teria. Antimicrob. Agents Chemother. 25:342-343.
4. Chapman, J., and N. Georgopapadakou. 1988. Routes of quino-lone permeation in Escherichia coli. Antimicrob. Agents Che-mother. 32:438-442.
5. Cofsky, R., L. duBouchet, and S. Landesman. 1984. Recovery ofnorfloxacin in feces after administration of a single oral dose tohuman volunteers. Antimicrob. Agents Chemother. 26:110-111.
6. Cohen, S., J. Hooper, J. Wolfson, K. Souza, L. McMurry, and S.Levy. 1988. Endogenous active efflux of norfloxacin in suscep-tible Escherichia coli. Antimicrob. Agents Chemother. 32:1187-1191.
7. Crumplin, G. C., M. Kenwright, and T. Hirst. 1984. Investiga-tions into the mechanism of action of the antibacterial agentnorfloxacin. J. Antimicrob. Chemother. 13:9-23.
8. Diver, J. M., J. M. Andrews, L. J. V. Piddock, and R. Wise.1988. Investigations into the uptake of ciprofloxacin, enoxacinand pefloxacin by Escherichia coli. Rev. Infect. Dis. 10(Suppl.1):s22-s23.
9. Edlund, C., T. Bergan, K. Josefsson, R. Solberg, and C. E. Nord.1987. Effect of norfloxacin on human oropharyngeal and colonicmicroflora and multiple-dose pharmacokinetics. Scand. J. In-fect. Dis. 19:113-121.
10. Edlund, C., and C. E. Nord. 1986. Comparative in vitro activi-ties of ciprofloxacin, enoxacin, norfloxacin, ofloxacin and pe-floxacin against Bacteroides fragilis and Clostridium difficile.Scand. J. Infect. Dis. 18:149-151.
11. Edlund, C., and C. E. Nord. 1988. A review on the impact of4-quinolones on the normal oropharyngeal and intestinal humanmicroflora. Infection 16:8-12.
12. Goldstein, E., D. Citron, and M. Corrado. 1987. Effect ofinoculum size on in vitro activity of norfloxacin against fecalanaerobic bacteria. Am. J. Med. 82:84-87.
13. Hazenberg, M. P., A. M. Pennock-Schroder, and J. P. van deMerwe. 1985. Binding to and antibacterial effect of aztreonam,temocillin, gentamicin and tobramycin on human faeces. J.Hyg. 95:255-263.
14. Hazenberg, M. P., A. M. Pennock-Schroder, and J. P. van deMerwe. 1986. Reversible binding of polymyxin B and neomycinto the solid part of faeces. J. Antimicrob. Chemother. 17:333-339.
15. Hirai, K., H. Aoyama, T. Irikura, S. lyobe, and S. Mitsuhashi.1986. Differences in susceptibility to quinolones of outer mem-brane mutants of Salmonella typhimurium and Escherichia coli.Antimicrob. Agents Chemother. 29:535-538.
16. Hooper, D., and J. Wolfson. 1988. Mode of action of thequinolone antimicrobial agents. Rev. Infect. Dis. 10(Suppl. 1):s14-s21.
17. Karp, J., W. Merz, C. Hendricksen, B. Laughon, T. Redden, B.Bamberger, J. Bartlett, R. Saral, and P. Burke. 1987. Oralnorfloxacin for prevention of gram-negative bacterial infectionsin patients with acute leukemia and granulocytopenia. Ann.Intern. Med. 106:1-7.
18. Meckenstock, R., E. Haralambie, G. Linzenmeier, and F. Wendt.1985. Die Beeinflussung der Darmflora durch Norfloxacin beigesunden Menschen. Z. Antimikrob. Antineoplast. Chemother.1:27-34.
19. Norrby, R., and M. Jonsson. 1983. Antibacterial activity ofnorfloxacin. Antimicrob. Agents Chemother. 23:15-18.
VOL. 32, 1988
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 24
Oct
ober
202
1 by
280
4:30
c:81
e:46
01:6
d4f:
c763
:cac
9:c0
4b.

1874 EDLUND ET AL. ANTIMICROB. AGENTS CHEMOTHER.
20. Reeves, D. S. 1986. The effect of quinolone antibacterials on thegastrointestinal flora compared with that of other antibacterials.J. Antimicrob. Chemother. 18:89-102.
21. Scatchard, G. 1949. The attractions of proteins for small mole-cules and ions. Ann. N.Y. Acad. Sci. 51:660-672.
22. Shannon, K., and I. Phillips. 1985. The antimicrobial spectrumof the quinolones. Res. Clin. Forum 7:29-36.
23. Shen, L., and A. Pernet. 1985. Mechanism of inhibition of DNAgyrase by analogues of nalidixic acid: the target of the drugs isDNA. Proc. Natl. Acad. Sci. USA 82:307-311.
24. Stephen, A., and J. Cummings. 1980. The microbial contributionto human faecal mass. J. Med. Microbiol. 13:45-56.
25. van der WaaQ, D. 1983. Colonization pattern of the digestive
tract by potentially pathogenic microorganisms: colonization-controlling mechanisms and consequences for antibiotic treat-ment. Infection ll(Suppl. 2):90-92.
26. van Saene, J., H. van Saene, C. Stoutenbeek, and C. Lerk. 1985.Influence of faeces on the activity of antimicrobial agents usedfor decontamination of the alimentary canal. Scand. J. Infect.Dis. 17:295-300.
27. Williams, L. T., and R. J. Lefkowitz. 1978. Theory of ligand-receptor interactions, p. 27-41. In Receptor binding studies inadrenergic pharmacology. Raven Press, New York.
28. Zweerink, M., and A. Edison. 1986. Inhibition of Micrococcusluteus DNA gyrase by norfloxacin and 10 other quinolonecarboxylic acids. Antimicrob. Agents Chemother. 29:598-601.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
ac o
n 24
Oct
ober
202
1 by
280
4:30
c:81
e:46
01:6
d4f:
c763
:cac
9:c0
4b.