nodule carbohydrate metabolism and polyols involvement in the response of medicago sativa to salt...
TRANSCRIPT
NM
FD
a
ARRA
KLMPS
1
cw11bpaioaauo
cT
0h
Environmental and Experimental Botany 85 (2013) 43– 49
Contents lists available at SciVerse ScienceDirect
Environmental and Experimental Botany
journa l h omepa g e: www.elsev ier .com/ locate /envexpbot
odule carbohydrate metabolism and polyols involvement in the response ofedicago sativa to salt stress
rancisco Palma ∗, Noel A. Tejera, Carmen Lluchepartamento de Fisiología Vegetal, Facultad de Ciencias, Universidad de Granada, Campus de Fuentenueva s/n, 18071 Granada, Spain
r t i c l e i n f o
rticle history:eceived 13 April 2012eceived in revised form 7 August 2012ccepted 19 August 2012
eywords:egumesedicago sativa
oliolsalt stress, Symbiosis
a b s t r a c t
Alterations of plant growth, chlorophyll fluorescence parameters, nodule carbon metabolism and polyolsconcentration as result of salt stress were examined in alfalfa (Medicago sativa). Plants, in symbiosiswith Sinorhizobium meliloti GR4 strain, were grown under controlled conditions for 35 days (DAS) andsubjected to 150 mM of NaCl stress. Plant biomass (PDW) and nitrogen fixation rate (NFR) were markedlyaffected by salt stress conditions; the highest reductions of PDW (50%) and NFR (40%) were registered at84 DAS and 56 DAS, respectively. In addition, salinity affected the chlorophyll fluorescence parameters,decreased initial chlorophyll fluorescence (F0) and increased the optimum quantum yield of PSII (Fv/Fm
ratio). The enzyme activities sucrose synthase activity and phosphoenolpyruvate carboxylase, responsiblefor the carbon supply to the bacteroids by the formation of dicarboxylates, were drastically inhibited bysalinity, mainly at 56 DAS with the beginning of flowering. The content of total soluble sugars and prolineincreased under salt stress, and these concentrations were higher in nodule than in leaf. This last result
suggests that the nodule is an organ specially protected in order to maintain its functioning, even understress conditions. Besides, the content of myoinositol and pinitol in leaves and nodules changed with theplant growth stage and the saline treatment. Under salinity stress, the concentrations of pinitol in nodulewere higher than in leaf, which supports the central function of this molecule in the adaptive responseof nodules to salt stress. The increase of pinitol synthesis in nodule of M. sativa under salt stress could beone of the adaptive features used by the plant.. Introduction
Salinity is one of the major abiotic factors limiting global agri-ultural productivity, and it is estimated that one-third of theorld’s irrigated land are unsuitable for crops (Frommer et al.,
999). Salt stress drastically affects the photosynthesis (Soussi et al.,998), the nitrogen metabolism (Cordovilla et al., 1994), the car-on metabolism (Delgado et al., 1993; Balibrea et al., 2003) and thelant nutrition (Mengel and Kirkby, 2001). Legumes are classifieds salt-sensitive crop species (Läuchli, 1984) and their productions particularly affected by salt stress because these plants dependn symbiotic N2 fixation for their nitrogen requirement (Elsheikhnd Wood, 1995). The limitation of productivity is associated with
lower growth of the host plant, poor development of the root nod-
les (Georgiev and Atkins, 1993) and consequently with a reductionf the nitrogen-fixation capacity (Ben Salah et al., 2011).∗ Corresponding author at: Departamento de Fisiología Vegetal, Facultad de Cien-ias, Universidad de Granada, Campus de Fuentenueva s/n, 18071 Granada, Spain.el.: +34 958 243382; fax: +34 958 248995.
E-mail address: [email protected] (F. Palma).
098-8472/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.envexpbot.2012.08.009
© 2012 Elsevier B.V. All rights reserved.
Under the variation of saline environments, plants develop dif-ferent adaptive mechanisms (Rhodes et al., 2002; Sairam et al.,2006), some of which include the synthesis and accumulationof low-molecular weight organic compounds in the cytosol andorganelles (Ashraf and Harris, 2004; Sairam et al., 2006). Thesecompounds, collectively called compatible osmolytes, are sim-ple sugars, disaccharides, sugar alcohols or polyols, amino acidsand sulfonium compounds (Ashraf and Harris, 2004; Bartels andSunkar, 2005; Ashraf and Foolad, 2007). A major function of theaccumulation of the compatible osmolytes is the osmotic adjust-ment to counteract the high concentrations of inorganic salts inthe vacuole and in the root medium (Zhu, 2001; Rhodes et al.,2002). Another function of the compatible osmolytes is the osmo-protection which may occur at lower salt concentrations. This roleinvolves the protection of thylakoids and the plasma membrane, aswell as the stabilization of proteins. Under salinity stress these com-pounds also act as a sink of energy or reducing power, as a source ofcarbon and nitrogen, or scavenging reactive oxygen species (Bartelsand Sunkar, 2005; Sairam et al., 2006).
Soluble carbohydrates and their polyol derivatives are the mostcommon osmolytes accumulating in plants in response to lowwater potentials. Cyclic polyols involve myoinositol, ononitol andpinitol. Myoinositol is derived from glucose-6-phosphate and can

4 d Exp
bicdsgtipaaac
iagpa
2
2
iatvew(br(c
2
tmgtgdnafgo
2
emwflroF(bm
4 F. Palma et al. / Environmental an
e further methylated to sequoyitol or ononitol, which are epimer-zed to d-pinitol (Loewus and Loewus, 1980). An increase in pinitolontent has been shown to occur in plants subjected to a watereficit (Streeter et al., 2001; Matos et al., 2010) as well as to a highalinity (Sengupta et al., 2008). Salinity induces the expression ofenes, one of which has been shown to be involved in pinitol syn-hesis (Vernon and Bohnert, 1992). Thus, the introduction of genesnvolved in the synthesis of proline (P5CF127A), betaines (betA) andolyols (mt1D, imt1) into plants contributes to abiotic stress toler-nce (Rathinasabapathi, 2000; Chen and Murata, 2002). It has beenlso suggested that exogenous application of compatible solutes isn alternative approach to improve crop productivity under salineonditions (Makela et al., 1999; Chen and Murata, 2002).
The objective of the present work was to investigate changesnduced by salinity on the content of some compatible osmolytesnd enzyme activities of carbon metabolism in Medicago sativarown under symbiotic conditions. In addition, the content of theolyols myoinositol and pinitol, as well as their relation with thedaptation of the symbiosis to salt stress, was also evaluated.
. Materials and methods
.1. Biological material and growth conditions
Seeds of M. sativa (var. Aragon) were surface sterilized bymmersion in 5% NaClO for 3 min and germinated in 0.8% water-gar plates at 28 ◦C in darkness. Two days after, seedlings wereransferred to individual pots of about 200 ml containing aermiculite–perlite mixture (3:1) and watered with N-free nutri-nt solution (Rigaud and Puppo, 1975). Each seedling, inoculatedith 1 ml of a stationary culture of Sinorhizobium meliloti GR4 strain
ca. 109 cell ml−1), was grown in a controlled environmental cham-er with a 16/8 h light–dark cycle, 23/18 ◦C day–night temperature,elative humidity 55/65% and photosynthetic photon flux density400–700 nm) of 450 �mol m−2 s−1 supplied by combined fluores-ent and incandescent lamps.
.2. Treatments and harvest
When plants were 35 days old (symbiosis established) they werereated with sodium chloride (150 mM) by adding it to the growth
edium. Ten plants were harvested from both treated and controlroup on days 42, 56, 70 and 84 respectively. A fresh sample ofhe root, containing nodules of each plant, was used for the nitro-enase assay, and afterward nodules were removed, weighed, andried at 70 ◦C for 48 h to calculate dry weight. Other samples ofodules from each plant, leaves, and roots were pooled and storedt −80 ◦C for the enzyme assays and analytical determinations. Theresh weight (FW) of roots (including the portion used for the nitro-enase assay), stems, and leaves were recorded, whereupon all thergans were dried at 70 ◦C for 48 h and their dry weight calculated.
.3. Chlorophyll fluorescence measurement
Chlorophyll fluorescence, of the first full expanded leaf ofach plant, was measured on each plant using a portable pulse-odulated fluorometer (OS5-FL, Opti-Sciences). After the leavesere dark-adapted for 30 min, the fluorescence parameters initialuorescence (F0), maximal fluorescence (Fmax) and variable fluo-escence (Fv) were analyzed (Schreiber et al., 1994). The F0 wasbtained with modulated low intensity light (<0.1 �mol m−2 s−1),
max was determined by a 0.8 s long saturating light pulse180 �mol m−2 s−1) and Fv was estimated by the differenceetween F0 and Fmax. The ratio of variable fluorescence to maxi-al fluorescence (Fv/Fmax), represents the relative state of PSII anderimental Botany 85 (2013) 43– 49
was used to assess the functional damage to the plants (Schreiberet al., 1994).
2.4. Nitrogen fixation assay
Nitrogenase activity (EC 1.7.9.92) was measured as the repre-sentative H2 evolution in an open-flow system (Witty and Minchin,1998) using an electrochemical H2 sensor (Qubit System Inc.,Canada). For nitrogenase measurements, pots maintained in a con-trolled environmental chamber (as described above) were sealedand H2 production was recorded. Apparent nitrogenase activity(ANA, rate of H2 production in air) was determined under N2:O2(80%:20%) with a total flow of 0.4 l min−1. After reaching steady-state conditions, total nitrogenase activity (TNA) was determinedunder Ar:O2 (79%:21%). The nitrogen fixation rate (NFR) was calcu-lated as (TNA − ANA)/3. Standards of high-purity H2 were used tocalibrate the detector.
2.5. Preparation of extracts and enzyme assays
Extracts were prepared homogenizing 0.2 g of nodules in amortar with 33% (w/w) polyvinyl–polypyrrolidone and 1.5 ml of50 mM phosphate K buffer (pH 8) containing 1 mM EDTA and 20%(v/v) ethylene glycol for sucrose synthase activity and 100 mMmaleic–KOH buffer (pH 6.8) containing 100 mM sucrose, 2% (v/v)�-mercaptoethanol and 20% (v/v) ethylene glycol for phospho-enolpyruvate carboxylase activity. Extracts were centrifuged at30 000 × g for 20 min and desalted supernatants were used to deter-mine enzyme activities. All operations were carried out at 4 ◦C.
Sucrose synthase activity (E.C.2.4.1.13) was measured accordingto (Morell and Copeland, 1985). The production of UDP–glucosewas coupled to the reduction of NAD+ in the presence of excessUDP–glucose dehydrogenase. Reaction mixtures were containedin a volume of 1 ml, 100 mM bicine KOH buffer (pH 8.5), 100 mMsucrose, 2 mM UDP, 0.025 U UDP–glucose dehydrogenase and1.5 mM NAD+. The assay was started by the addition of the enzymeextract. The activity was spectrophotometrically measured byfollowing the NAD+ reduction at 340 nm. Phosphoenolpyruvatecarboxylase activity (E.C.4.1.1.31) assays were optimized from the(Soussi et al., 1998) method. The reaction mixtures contained100 mM bicine KOH (pH 8.5), 5.0 mM MgCl2, 0.2 mM NADH, 10 mMNaHCO3 and 2.0 mM PEP. The activity was followed by the oxidationof NADH at 340 nm.
2.6. Proline, total soluble sugars, malate and protein analysis
Proline, soluble sugars and malate were extracted using 1 g ofsample (leaves and nodules) and 12 ml of extraction medium. Bothsoluble sugars and proline were quantified by means of a colori-metric reaction with anthrone and ninhydrin reagents, respectively(Irigoyen et al., 1992). Standard curves, prepared with l-prolineand glucose, were used to estimate concentrations. Malate contentwas determined enzymatically monitoring the production of NADHat 340 nm (Delhaize et al., 1993). The sample was incubated withbuffer (0.5 M glycine, 0.4 M hydrazine, pH 9.0), NAD (40 mM) andmalate dehydrogenase (5 mg/ml). Protein concentration was mea-sured at 660 nm by the method of (Lowry et al., 1951) using theFolin–Ciocalteau reagent with bovine-serum albumin as a standard.
2.7. Myoinositol, pinitol and trehalose analysis
Sugar alcohols and trehalose were separated and quantified by
isocratic ion chromatography with pulsed amperometric detectionaccording to Cataldi et al. (2000) with modifications, conducted ona Dionex ICS-3000 chromatograph (Dionex Corp., Sunnyvale, CA).Leaf and nodule samples (0.2 g) were homogenizing in mortars with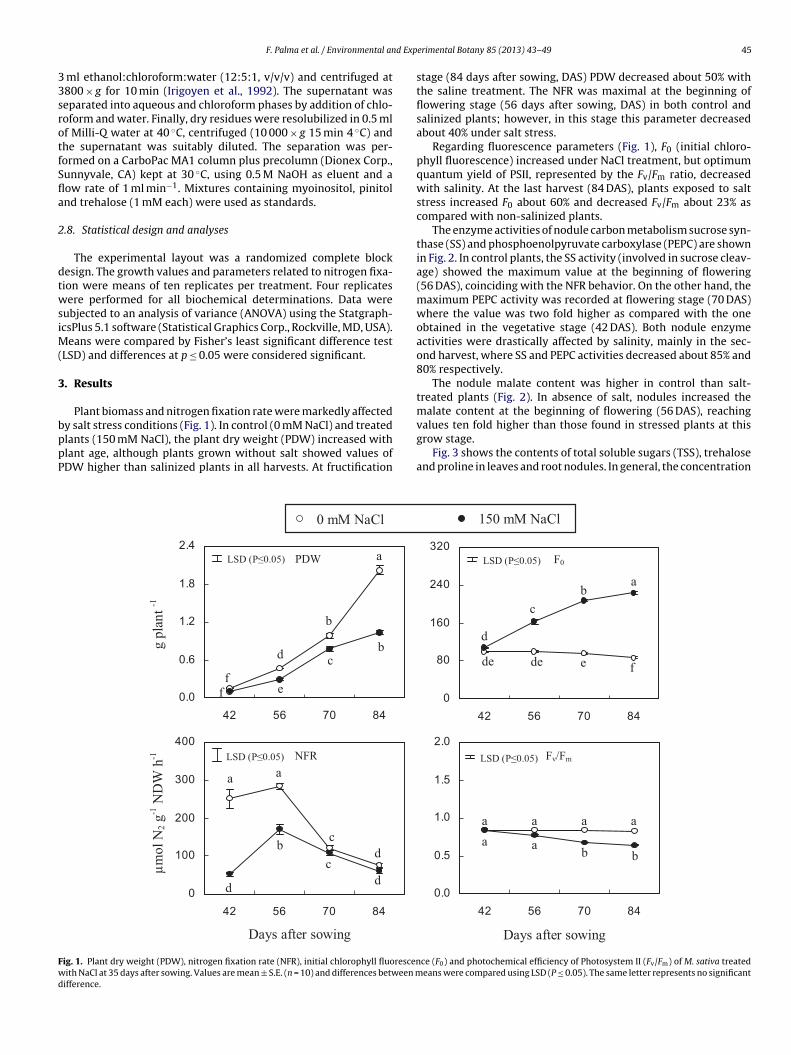
d Exp
33srotfSfla
2
dtwsiM(
3
bppP
Fwd
F. Palma et al. / Environmental an
ml ethanol:chloroform:water (12:5:1, v/v/v) and centrifuged at800 × g for 10 min (Irigoyen et al., 1992). The supernatant waseparated into aqueous and chloroform phases by addition of chlo-oform and water. Finally, dry residues were resolubilized in 0.5 mlf Milli-Q water at 40 ◦C, centrifuged (10 000 × g 15 min 4 ◦C) andhe supernatant was suitably diluted. The separation was per-ormed on a CarboPac MA1 column plus precolumn (Dionex Corp.,unnyvale, CA) kept at 30 ◦C, using 0.5 M NaOH as eluent and aow rate of 1 ml min−1. Mixtures containing myoinositol, pinitolnd trehalose (1 mM each) were used as standards.
.8. Statistical design and analyses
The experimental layout was a randomized complete blockesign. The growth values and parameters related to nitrogen fixa-ion were means of ten replicates per treatment. Four replicatesere performed for all biochemical determinations. Data were
ubjected to an analysis of variance (ANOVA) using the Statgraph-csPlus 5.1 software (Statistical Graphics Corp., Rockville, MD, USA).
eans were compared by Fisher’s least significant difference testLSD) and differences at p ≤ 0.05 were considered significant.
. Results
Plant biomass and nitrogen fixation rate were markedly affected
y salt stress conditions (Fig. 1). In control (0 mM NaCl) and treatedlants (150 mM NaCl), the plant dry weight (PDW) increased withlant age, although plants grown without salt showed values ofDW higher than salinized plants in all harvests. At fructification0.0
0.6
1.2
1.8
2.4
42 56 70 84
0
100
200
300
400
42 56 70 84
PDW
NFR
g pl
ant -1
μmol
N2
g-1N
DW
h-1
Days after sowing
0 mM NaCl
LSD (P≤0.05) a
b
b
c d
e f
f
a LSD (P≤0.05)
a
b c
c d
d d
ig. 1. Plant dry weight (PDW), nitrogen fixation rate (NFR), initial chlorophyll fluorescenith NaCl at 35 days after sowing. Values are mean ± S.E. (n = 10) and differences between mifference.
erimental Botany 85 (2013) 43– 49 45
stage (84 days after sowing, DAS) PDW decreased about 50% withthe saline treatment. The NFR was maximal at the beginning offlowering stage (56 days after sowing, DAS) in both control andsalinized plants; however, in this stage this parameter decreasedabout 40% under salt stress.
Regarding fluorescence parameters (Fig. 1), F0 (initial chloro-phyll fluorescence) increased under NaCl treatment, but optimumquantum yield of PSII, represented by the Fv/Fm ratio, decreasedwith salinity. At the last harvest (84 DAS), plants exposed to saltstress increased F0 about 60% and decreased Fv/Fm about 23% ascompared with non-salinized plants.
The enzyme activities of nodule carbon metabolism sucrose syn-thase (SS) and phosphoenolpyruvate carboxylase (PEPC) are shownin Fig. 2. In control plants, the SS activity (involved in sucrose cleav-age) showed the maximum value at the beginning of flowering(56 DAS), coinciding with the NFR behavior. On the other hand, themaximum PEPC activity was recorded at flowering stage (70 DAS)where the value was two fold higher as compared with the oneobtained in the vegetative stage (42 DAS). Both nodule enzymeactivities were drastically affected by salinity, mainly in the sec-ond harvest, where SS and PEPC activities decreased about 85% and80% respectively.
The nodule malate content was higher in control than salt-treated plants (Fig. 2). In absence of salt, nodules increased themalate content at the beginning of flowering (56 DAS), reaching
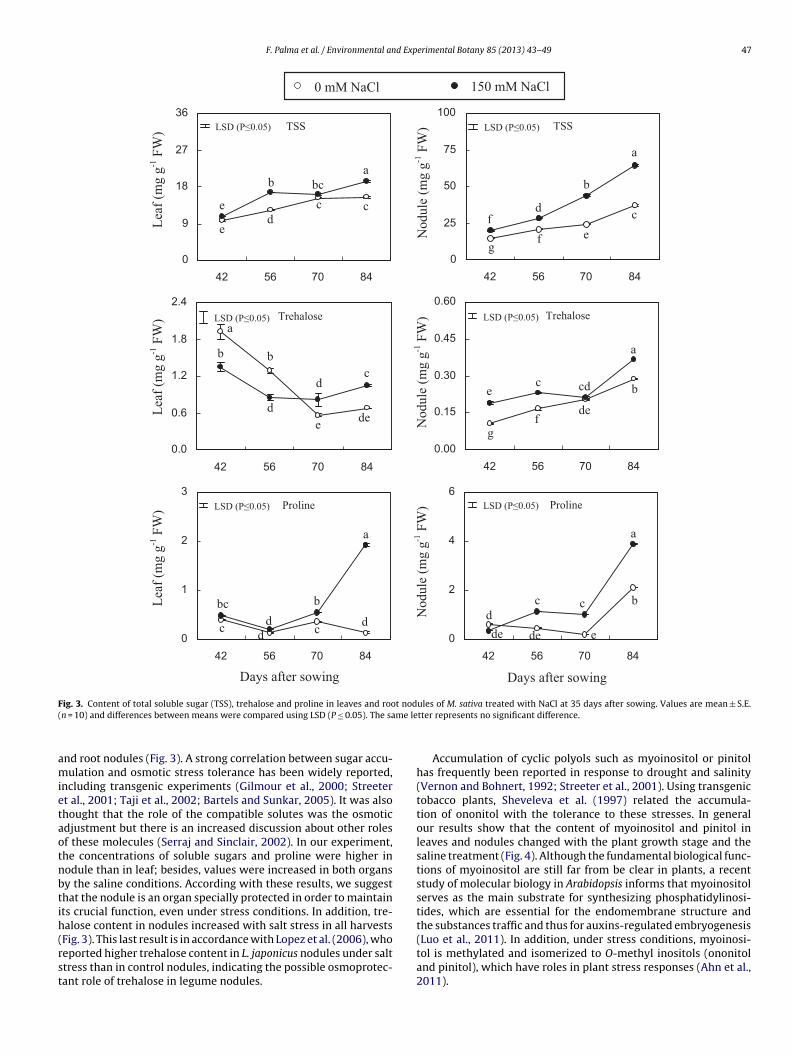
values ten fold higher than those found in stressed plants at thisgrow stage.Fig. 3 shows the contents of total soluble sugars (TSS), trehaloseand proline in leaves and root nodules. In general, the concentration
0
80
160
240
320
42 56 70 84
0.0
0.5
1.0
1.5
2.0
42 56 70 84
F0
Fv/F m
Days after sowing
150 mM NaCl
a
LSD (P≤0.05)
b c
d
de de e f
a
LSD (P≤0.05)
a a a a
a b b
ce (F0) and photochemical efficiency of Photosystem II (Fv/Fm) of M. sativa treatedeans were compared using LSD (P ≤ 0.05). The same letter represents no significant

46 F. Palma et al. / Environmental and Exp
0
5
10
15
20
42 56 70 84
mg
g-1FW
Malate
0.0000
0.0015
0.0030
0.0045
0.0060
42 56 70 84
0.0
0.1
0.2
0.3
0.4
42 56 70 84
SS
μmol
NA
D+ m
g-1 p
rotm
in-1
PEPC
0 mM NaCl 150 mM NaCl
Days after sowing
μmol
NA
DH
mg-1
pro
t min
-1
LSD (P≤0.05)
a
b b
c cd
cdd e
a
LSD (P≤0.05)
b c c
cdde e
f
LSD (P≤0.05)
a
b c c
d e f f
Fig. 2. Sucrose synthase activity (SS), phosphoenolpyruvate carboxylase activity(ac
otU4settAcflti
a
PEPC), and content of malate in root nodules of M. sativa treated with NaCl at 35 daysfter sowing. Values are mean ± S.E. (n = 10) and differences between means wereompared using LSD (P ≤ 0.05). The same letter represents no significant difference.
f TSS and proline was higher in nodules than in leaves; besides,he values were increased in both organs by the saline conditions.nder the effect of salinity, TSS increased two fold in leaf and about2% in nodule, in the last harvest. Proline content also increasedignificantly by the salinity in both organs; at 84 DAS, the valuesnhanced 93% and 45% in leaf and nodule, respectively. Regardingrehalose content in leaves, the behavior was different in the con-rol treatment and stressed plants along the whole experiment.t the stages vegetative and beginning of the flowering, trehaloseoncentrations were higher in leaves of unstressed plants, but atowering and fructification stages, the values were higher in NaCl-
reated plants. Salinity also induced an accumulation of trehalosen nodules, which was found in all harvests.In general, the content of myoinositol and pinitol in leavesnd nodules changed with the plant growth stage and the saline
erimental Botany 85 (2013) 43– 49
treatment (Fig. 4). The concentrations of myoinositol decreasedsignificantly in leaf under salt effect, however, was higher in thenodules, especially two weeks after treatment. In nodules, the high-est value was recorded 7 days after NaCl application (42 DAS);reaching a six folds higher concentration than non-salinized plants.On the other hand, pinitol content increased with salt stress andplant age in both organs (leaf and nodule). Under salt stress, theincreases of pinitol content were more important in nodules, being1.5-fold higher at 42 DAS and about 2–3-fold higher at the rest ofharvests. In leaf, the increase of pinitol resulted from the salt stresswas about 20–25% along the whole experiment.
4. Discussion
Several papers have been written regarding the effect of saltstress on nitrogen fixation and on the enzymes of the carbonmetabolism in nodules (Lopez et al., 2008; Lopez and Lluch, 2008) aswell as on the accumulation of sugars and other compatible solutes(Bartels and Sunkar, 2005; Khadri et al., 2007), but the mechanismsimplicated in these processes remain unclear. In the present work,we examined the changes in the content of polyols (myoinositoland pinitol), the presence of soluble carbohydrates and the carbonmetabolism in nodules of M. sativa plants under salt stress.
Our study showed a significant decrease of PDW and NFR ofM. sativa under salt stress (Fig. 1). The highest reductions of PDW(50%) and NFR (40%) were registered at 84 DAS and 56 DAS, respec-tively. These results are in agreement with those reported in grainlegumes, such as Phaseolus vulgaris (Tejera et al., 2005) and Cicerarietinum (Soussi et al., 1998), and forage legumes as Lotus japoni-cus and Medicago truncatula (Lopez et al., 2008). In addition, salinityaffected the chlorophyll fluorescence parameters, decreased initialchlorophyll fluorescence (F0) and increased the optimum quantumyield of PSII (Fv/Fm ratio). This last result might be due to the lesionsin the reaction center of PSII, as it was previously pointed out insorghum (Masojidek and Hall, 1992), or indirectly by an acceleratedsenescence as it happens in rice (Moradi and Ismail, 2007).
The enzyme activities SS and PEPC, responsible of the carbonsupply to the bacteroids by the formation of dicarboxylates, weredrastically inhibited by salinity, mainly at 56 DAS in the beginningof flowering (Fig. 2). These results agree with the experiments ofBen Salah et al. (2009), who reported a decrease of SS activity inMedicago ciliaris subjected to salt stress. The sucrose synthase activ-ity of legumes ensures the supply of hexoses for the demands ofenergy and reducing power of nodules during the active stage ofnitrogen fixation and this enzyme may regulate carbon metabolismand nitrogen fixation (Lopez and Lluch, 2008). In this regard, nod-ules of non-stressed plants showed the maximum value of SSactivity at the beginning of flowering stage, coinciding with thepeak of nitrogenase activity (Fig. 1).
It has been previously hypothesized that there is a close rela-tion between nitrogen fixation and the capacity of the nodules tometabolize sucrose (Lopez and Lluch, 2008). Our results showedthat malate content in nodules of non stressed plants (control) washigher than in salt treated plants; besides, in absence of NaCl themalate concentration was more elevated at the beginning of flow-ering stage (Fig. 2). The decrease in SS activity by salt stress couldlead to a reduction of malate content in nodules, and this declinecould result a shortage of substrates for bacteroid respiration andprobably a disturbance in the regulation of the oxygen permeabil-ity (Gálvez et al., 2000). Consequently, it may explain the observeddecline in the nitrogen fixation rate and also could provide addi-
tional evidences on the central role of SS in the regulation of nodulecarbon metabolism and nitrogen fixation.In general, ours results showed that NaCl treatment increasedthe content of total soluble sugars, trehalose and proline in leaves
F. Palma et al. / Environmental and Experimental Botany 85 (2013) 43– 49 47
0.0
0.6
1.2
1.8
2.4
42 56 70 84
0
9
18
27
36
42 56 70 84
TSS
Leaf
(mg
g-1 F
W)
0.00
0.15
0.30
0.45
0.60
42 56 70 84
0
25
50
75
100
42 56 70 84
0
2
4
6
42 56 70 840
1
2
3
42 56 70 84
TSS
Trehalose Trehalose
Proline Proline
Nod
ule
(mg
g-1 F
W)
Leaf
(mg
g-1 F
W)
Leaf
(mg
g-1 F
W)
Nod
ule
(mg
g-1 F
W)
Nod
ule
(mg
g-1 F
W)
Days after sowing Days after sowing
0 mM NaCl 150 mM NaCl
LSD (P≤0.05)a
b b c
d
d dee
LSD (P≤0.05)
a b bc
c c d
e
e
LSD (P≤0.05)
a
b
c d
e f f
g
LSD (P≤0.05)
a
b c cd
dee
g f
LSD (P≤0.05)
a
b bc
c c d d
d
LSD (P≤0.05)
a
b c c d
de e de
F t nod( me le
amietaotnbtih(rst
ig. 3. Content of total soluble sugar (TSS), trehalose and proline in leaves and roon = 10) and differences between means were compared using LSD (P ≤ 0.05). The sa
nd root nodules (Fig. 3). A strong correlation between sugar accu-ulation and osmotic stress tolerance has been widely reported,
ncluding transgenic experiments (Gilmour et al., 2000; Streetert al., 2001; Taji et al., 2002; Bartels and Sunkar, 2005). It was alsohought that the role of the compatible solutes was the osmoticdjustment but there is an increased discussion about other rolesf these molecules (Serraj and Sinclair, 2002). In our experiment,he concentrations of soluble sugars and proline were higher inodule than in leaf; besides, values were increased in both organsy the saline conditions. According with these results, we suggesthat the nodule is an organ specially protected in order to maintaints crucial function, even under stress conditions. In addition, tre-alose content in nodules increased with salt stress in all harvests
Fig. 3). This last result is in accordance with Lopez et al. (2006), whoeported higher trehalose content in L. japonicus nodules under salttress than in control nodules, indicating the possible osmoprotec-ant role of trehalose in legume nodules.ules of M. sativa treated with NaCl at 35 days after sowing. Values are mean ± S.E.tter represents no significant difference.
Accumulation of cyclic polyols such as myoinositol or pinitolhas frequently been reported in response to drought and salinity(Vernon and Bohnert, 1992; Streeter et al., 2001). Using transgenictobacco plants, Sheveleva et al. (1997) related the accumula-tion of ononitol with the tolerance to these stresses. In generalour results show that the content of myoinositol and pinitol inleaves and nodules changed with the plant growth stage and thesaline treatment (Fig. 4). Although the fundamental biological func-tions of myoinositol are still far from be clear in plants, a recentstudy of molecular biology in Arabidopsis informs that myoinositolserves as the main substrate for synthesizing phosphatidylinosi-tides, which are essential for the endomembrane structure andthe substances traffic and thus for auxins-regulated embryogenesis
(Luo et al., 2011). In addition, under stress conditions, myoinosi-tol is methylated and isomerized to O-methyl inositols (ononitoland pinitol), which have roles in plant stress responses (Ahn et al.,2011).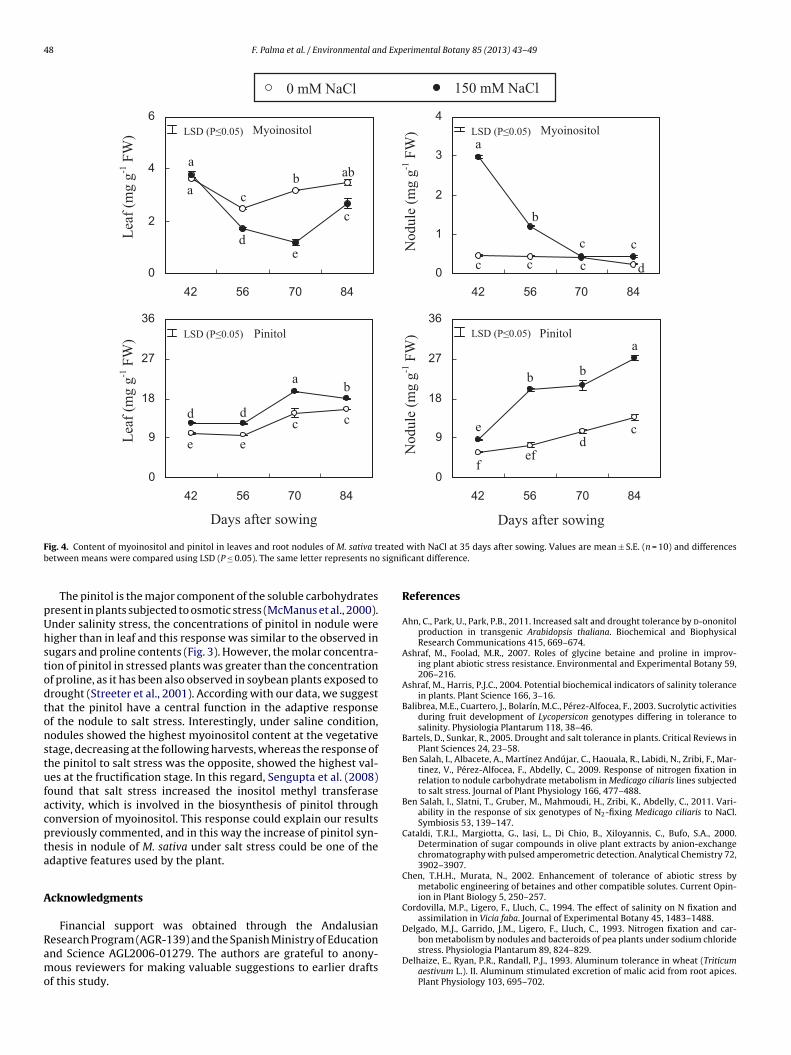
48 F. Palma et al. / Environmental and Experimental Botany 85 (2013) 43– 49
0
9
18
27
36
42 56 70 840
9
18
27
36
42 56 70 84
0
1
2
3
4
42 56 70 840
2
4
6
42 56 70 84
Myoinositol
Nod
ule
(mg
g-1 F
W)
Nod
ule
(mg
g-1 F
W) Pinitol
Myoinositol
Pinitol
Leaf
(mg
g-1 F
W)
Leaf
(mg
g-1 F
W)
Days after sowing Days after sowing
0 mM NaCl 150 mM NaCl
LSD (P≤0.05)
a
a abb
c c
d e
c c c c d
b
a LSD (P≤0.05)
c
a b
LSD (P≤0.05)
c c d d
e e e
LSD (P≤0.05)a
b b
c d
eff
F reatedb signifi
pUhstodtonstufacpta
A
Ramo
ig. 4. Content of myoinositol and pinitol in leaves and root nodules of M. sativa tetween means were compared using LSD (P ≤ 0.05). The same letter represents no
The pinitol is the major component of the soluble carbohydratesresent in plants subjected to osmotic stress (McManus et al., 2000).nder salinity stress, the concentrations of pinitol in nodule wereigher than in leaf and this response was similar to the observed inugars and proline contents (Fig. 3). However, the molar concentra-ion of pinitol in stressed plants was greater than the concentrationf proline, as it has been also observed in soybean plants exposed torought (Streeter et al., 2001). According with our data, we suggesthat the pinitol have a central function in the adaptive responsef the nodule to salt stress. Interestingly, under saline condition,odules showed the highest myoinositol content at the vegetativetage, decreasing at the following harvests, whereas the response ofhe pinitol to salt stress was the opposite, showed the highest val-es at the fructification stage. In this regard, Sengupta et al. (2008)ound that salt stress increased the inositol methyl transferasectivity, which is involved in the biosynthesis of pinitol throughonversion of myoinositol. This response could explain our resultsreviously commented, and in this way the increase of pinitol syn-hesis in nodule of M. sativa under salt stress could be one of thedaptive features used by the plant.
cknowledgments
Financial support was obtained through the Andalusianesearch Program (AGR-139) and the Spanish Ministry of Educationnd Science AGL2006-01279. The authors are grateful to anony-ous reviewers for making valuable suggestions to earlier drafts
f this study.
with NaCl at 35 days after sowing. Values are mean ± S.E. (n = 10) and differencescant difference.
References
Ahn, C., Park, U., Park, P.B., 2011. Increased salt and drought tolerance by d-ononitolproduction in transgenic Arabidopsis thaliana. Biochemical and BiophysicalResearch Communications 415, 669–674.
Ashraf, M., Foolad, M.R., 2007. Roles of glycine betaine and proline in improv-ing plant abiotic stress resistance. Environmental and Experimental Botany 59,206–216.
Ashraf, M., Harris, P.J.C., 2004. Potential biochemical indicators of salinity tolerancein plants. Plant Science 166, 3–16.
Balibrea, M.E., Cuartero, J., Bolarín, M.C., Pérez-Alfocea, F., 2003. Sucrolytic activitiesduring fruit development of Lycopersicon genotypes differing in tolerance tosalinity. Physiologia Plantarum 118, 38–46.
Bartels, D., Sunkar, R., 2005. Drought and salt tolerance in plants. Critical Reviews inPlant Sciences 24, 23–58.
Ben Salah, I., Albacete, A., Martínez Andújar, C., Haouala, R., Labidi, N., Zribi, F., Mar-tinez, V., Pérez-Alfocea, F., Abdelly, C., 2009. Response of nitrogen fixation inrelation to nodule carbohydrate metabolism in Medicago ciliaris lines subjectedto salt stress. Journal of Plant Physiology 166, 477–488.
Ben Salah, I., Slatni, T., Gruber, M., Mahmoudi, H., Zribi, K., Abdelly, C., 2011. Vari-ability in the response of six genotypes of N2-fixing Medicago ciliaris to NaCl.Symbiosis 53, 139–147.
Cataldi, T.R.I., Margiotta, G., Iasi, L., Di Chio, B., Xiloyannis, C., Bufo, S.A., 2000.Determination of sugar compounds in olive plant extracts by anion-exchangechromatography with pulsed amperometric detection. Analytical Chemistry 72,3902–3907.
Chen, T.H.H., Murata, N., 2002. Enhancement of tolerance of abiotic stress bymetabolic engineering of betaines and other compatible solutes. Current Opin-ion in Plant Biology 5, 250–257.
Cordovilla, M.P., Ligero, F., Lluch, C., 1994. The effect of salinity on N fixation andassimilation in Vicia faba. Journal of Experimental Botany 45, 1483–1488.
Delgado, M.J., Garrido, J.M., Ligero, F., Lluch, C., 1993. Nitrogen fixation and car-
bon metabolism by nodules and bacteroids of pea plants under sodium chloridestress. Physiologia Plantarum 89, 824–829.Delhaize, E., Ryan, P.R., Randall, P.J., 1993. Aluminum tolerance in wheat (Triticumaestivum L.). II. Aluminum stimulated excretion of malic acid from root apices.Plant Physiology 103, 695–702.

d Exp
E
F
G
G
G
I
K
L
L
L
L
L
L
L
M
M
M
F. Palma et al. / Environmental an
lsheikh, E.A.E., Wood, M., 1995. Nodulation and N2 fixation by soybean inocu-lated with salt-tolerant rhizobia or salt-sensitive bradyrhizobia in saline soil.Soil Biology and Biochemistry 27, 657–661.
rommer, W.B., Ludewig, U., Rentsch, D., 1999. Taking transgenic plants with a pinchof salt. Science 285, 1222–1223.
álvez, S., Hirsch, A.M., Wycoff, K.L., Hunt, S., Layzell, D.B., Kondorosi, A., Crespi, M.,2000. Oxygen regulation of a nodule located carbonic anhydrase in alfalfa. PlantPhysiology 124, 1059–1068.
eorgiev, G.I., Atkins, C.A., 1993. Effects of salinity on N2 fixation, nitrogenmetabolism and export and diffusive conductance of cowpea root nodules. Sym-biosis 15, 239–255.
ilmour, S.J., Sebolt, A.M., Salazar, M.P., Everard, J.D., Thomashow, M.F., 2000. Over-expression of the Arabidopsis CBF3 transcriptional activator mimics multiplebiochemical changes associated with cold acclimation. Plant Physiology 124,1854–1865.
rigoyen, J.J., Emerich, D.W., Sánchez-Díaz, M., 1992. Water stress induced changes inconcentrations of proline and total soluble sugars in nodulated alfalfa (Medicagosativa) plants. Physiologia Plantarum 84, 55–60.
hadri, M., Tejera, N.A., Lluch, C., 2007. Sodium chloride–ABA interaction in twocommon bean (Phaseolus vulgaris) cultivars differing in salinity tolerance. Envi-ronmental and Experimental Botany 60, 211–218.
äuchli, A., 1984. Salt exclusion: an adaptation of legumes for crops and pasturesunder saline conditions. In: Staples, R.C. (Ed.), Salinity Tolerance in Plants. Wiley-Interscience, New York, USA, pp. 171–188.
oewus, F.A., Loewus, M.W., 1980. Myoinosytol: biosynthesis and metabolism. In:Preiss, J. (Ed.), The Biochemistry of Plants. Carbohydrates: Structure and Func-tion. Academic Press, New York, USA, pp. 43–76.
opez, M., Herrera-Cervera, J.A., Iribarne, C., Tejera, N.A., Lluch, C., 2008. Growth andnitrogen fixation in Lotus japonicus and Medicago truncatula under NaCl stress:nodule carbon metabolism. Journal of Plant Physiology 165, 641–650.
opez, M., Herrera-Cervera, J.A., Lluch, C., Tejera, N.A., 2006. Trehalose metabolismin root nodules of the model legume Lotus japonicus in response to salt stress.Physiologia Plantarum 128, 701–709.
opez, M., Lluch, C., 2008. Nitrogen fixation is synchronized with carbon metabolismin Lotus japonicus and Medicago truncatula nodules under salt stress. Journal ofPlant Interactions 3, 137–144.
owry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin phenol reagent. Journal of Biological Chemistry 193, 265–275.
uo, Y., Qin, G., Zhang, J., Liang, Y., Song, Y., Zhao, M., Tsuge, T., Aoyama, T., Liu, J.,Gu, H., Qu, L.J., 2011. d-myo-Inositol-3-phosphate affects phosphatidylinositol-mediated endomembrane function in Arabidopsis and is essential forauxin-regulated embryogenesis. Plant Cell 23, 1352–1372.
akela, P., Kontturi, M., Pehu, E., Somersalo, S., 1999. Photosynthetic response ofdrought and salt stressed tomato and turnip rape plants to foliar applied glycine-betaine. Physiologia Plantarum 105, 45–50.
asojidek, J., Hall, D.O., 1992. Salinity and drought stresses are amplified by high
irradiance in sorghum. Photosynthetica 27, 159–171.atos, M.C., Campos, P.S., Passarinho, J.A., Semedo, J.N., Marques, N.M., Ramalho,J.C., Ricardo, C.P., 2010. Drought effect on photosynthetic activity, osmolyteaccumulation and membrane integrity of two Cicer arietinum genotypes. Pho-tosynthetica 48, 303–312.
erimental Botany 85 (2013) 43– 49 49
McManus, M.T., Bieleski, R.L., Caradus, J.R., Barker, D.J., 2000. Pinitol accumulationin mature leaves of white clover in response to a water deficit. Environmentaland Experimental Botany 43, 11–18.
Mengel, K., Kirkby, E.A., 2001. Principles of plant nutrition. Annals of Botany 93,479–480.
Moradi, F., Ismail, A.M., 2007. Responses of photosynthesis, chlorophyll fluorescenceand ROS-Scavenging systems to salt stress during seedling and reproductivestages in rice. Annals of botany 99, 1161–1173.
Morell, M., Copeland, L., 1985. Sucrose synthase of soybean nodules. Plant Physiology78, 149–154.
Rathinasabapathi, B., 2000. Metabolic engineering for stress tolerance: installingosmoprotectant synthesis pathways. Annals of botany 86, 709–716.
Rhodes, D., Nadolska-Orczyk, A., Rich, P.J., 2002. Salinity, osmolytes and compatiblesolutes. In: Läuchli, A., Lüttge, U. (Eds.), Salinity: Environment-Plants-Molecules.Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 181–204.
Rigaud, J., Puppo, A., 1975. Indole-3-acetic-acid catabolism by soybean bacteroids.Journal of General Microbiology 88, 223–228.
Sairam, R.K., Tyagi, A., Chinnusamy, V., 2006. Salinity tolerance: cellular mechanismsand gene regulation. In: Huang, B. (Ed.), Plant–Environment Interactions. , thirdedition. CRC Press, Boca Raton, FL, pp. 121–175.
Schreiber, U., Bilger, W., Newbauer, C., 1994. Chlrophyll fluorescence as a nonin-trusive indicator for rapid assessment of in vivo photosynthesis. In: Schulze,E.D., Caldwell, M.M. (Eds.), Ecophysiology and Photosynthesis. Springer Verlag,Heidelberg, pp. 49–70.
Sengupta, S., Patra, B., Ray, S., Majumder, A.L., 2008. Inositol methyl tranferase froma halophytic wild rice, Porteresia coarctata Roxb. (Tateoka): regulation of pinitolsynthesis under abiotic stress. Plant, Cell & Environment 31, 1442–1459.
Serraj, R., Sinclair, T.R., 2002. Osmolyte accumulation: can it really help increase cropyield under drought conditions? Plant, Cell & Environment 25, 333–341.
Sheveleva, E., Chmara, W., Bohnert, H.J., Jensen, R.G., 1997. Increased salt and droughttolerance by d-ononitol production in transgenic Nicotiana tabacum L. PlantPhysiology 115, 1211–1219.
Soussi, M., Ocana, A., Lluch, C., 1998. Effects of salt stress on growth, photosynthesisand nitrogen fixation in chick-pea (Cicer arietinum L.). Journal of ExperimentalBotany 49, 1329–1337.
Streeter, J.G., Lohnes, D.G., Fioritto, R.J., 2001. Patterns of pinitol accumulation insoybean plants and relationships to drought tolerance. Plant, Cell & Environment24, 429–438.
Taji, T., Ohsumi, C., Seki, M., Iuchi, S., Yamaguchi-Shinozaki, K., Shinozaki, K., 2002.Important roles of drought- and cold-inducible genes for galactinol synthase instress tolerance in Arabidopsis thaliana. Plant and Cell Physiology 43, S233–S233.
Tejera, N.A., Campos, R., Sanjuan, J., Lluch, C., 2005. Effect of sodium chloride ongrowth, nutrient accumulation, and nitrogen fixation of common bean plants insymbiosis with isogenic strains. Journal of Plant Nutrition 28, 1907–1921.
Vernon, D.M., Bohnert, H.J., 1992. A novel methyl transferase induced by osmoticstress in the facultative halophyte Mesembryanthemum crystallinum. EMBO Jour-
nal 11, 2077–2085.Witty, J.F., Minchin, F.R., 1998. Methods for the continuous measurement of O2
consumption and H2 production by nodulated legume root systems. Journal ofExperimental Botany 49, 1041–1047.
Zhu, J.K., 2001. Plant salt tolerance. Trends in Plant Science 6, 66–71.