nitrogen and sugar concentrations in the xylem exudate of field-grown maize at different growth...
TRANSCRIPT

Eur. J. Agron., 1994, 3(3), 197-204
Nitrogen and sugar concentrations in the xylem exudate of field-grown maize at different growth stages and levels of nitrogen fertilization
C. Engels*, B. Buerkert and H. Marschner
Institut fiir Pflanzenerndhrung ( 330 ), 70593 Stuttgart, Germany.
Received 11 February 1994 ; accepted 16 March 1994.
* To whom correspondence should be addressed.
Abstract In field-grown maize (Zea mays L.) at different growth stages xylem exudate was collected and analyzed for nitrogen compounds and sugars. The plants were grown at two levels of nitrogen (N) fertilization (low N, high N). In an additional experiment in some plants the cobs were removed after anthesis.
The concentrations of nitrate- and reduced-N in the exudate were greater at high than at low N fertilization. At both levels of N fertilization, during plant development the concentrations of nitrate-N declined but not those of reduced-N, leading to an increase in the percentage of reduced-N up to 80-90 per cent of the total N in the exudate. This pattern may reflect either an increase in the proportion of nitrate reduction in the roots, or of N cycling between the shoot and the roots, or both. Increased internal cycling of N was considered to be the main cause of the significant increase in the concentrations of reduced N in the exudate after cob removal.
The concentrations of reducing sugars and sucrose in the exudate, at both levels of N fertilization, increased after anthesis to more than 20 mM, and after ear removal to more than 50 mM. Cycling of sugars between shoot and roots and mobilization of sugars previously stored in the roots are discussed as possible causes for the high sugar concentrations in the exudate, which might significantly contribute to the osmotic driving force for the volume flow in the xylem.
Key-words : cob removal, cycling, growth stage, maize nitrogen, sugar, xylem exudate.
INTRODUCTION
Transport of nitrate and reduced N compounds from the roots to the shoot can be measured in xylem exudate of decapitated plants. The relative content of nitrate and reduced N in the xylem exudate has been taken as indicator for the location of nitrate reduction (shoot or roots) in different plant species (Keltjens et al., 1986), its dependence of the time of the day (Keltjens and Nijenstein, 1987), or the rate of N transport through the roots and xylem loading (Rufty et al., 1981). Furthermore, from N fluxes in the xylem exudate conclusions have been drawn on the importance of retranslocation of reduced N from the shoot to the roots (Simpson et al., 1982) and on cycling of N between shoot and roots (Cooper and Clarkson, 1989).
The above mentioned experiments were conducted with seedlings under controlled environmental conditions. Information on the exudate composition of field-grown plants during plant development is relatively scarce. In the exudate of field-grown maize
ISSN 1161-0301/94!03/$ 4.001 © Gauthier-Villars - ESAg
plants, equal concentrations of nitrate and amino acids were measured at flowering (Canny and McCully, 1988). In the present study, which was part of a research program on the time course of N uptake in maize at different levels of N fertilization (Geyer, 1991 ), the concentrations of nitrate and reduced N in the xylem exudate were determined at different growth stages and at low and high levels of N fertilization.
Xylem transport of carbohydrates is common in spring in perennial plants such as grapevine (Hardy and Possingham, 1969) but is usually absent in herbaceous plants (Pate, 1975). In the xylem exudate of white lupin, recently photosynthetically fixed carbon has been found, but the nature of the carbon compounds (organic acids, sugars etc.) has not been identified (Minchin and McNaughton, 1987). In maize, malate is a common and important component in the xylem exudate (Butz and Long, 1979; Teyker et al., 1991), but surprisingly, sugar concentrations of about 5 mM were found at flowering (Canny and McCully, 1988).

198
The aim of the present study was to investigate whether sugar concentrations in the xylem exudate of field-grown maize are dependent on plant developmental stage, on the level of N fertilization or on the availability of carbohydrates in the shoot. For examination of the latter aspect the effect of cob removal on xylem exudate composition was studied.
MATERIALS AND METHODS
Maize plants (Zea mays L., cv. Olymp) were grown in two years ( 1987 and 1991) on a Gleyic Luvisol at the Agricultural Experimental Station of the University of Hohenheim, Stuttgart. In 1987, the weather was characterized by cool temperatures in May and June and high rainfall in May, June and July (see Figure 1). In 1991, the weather was characterized by high temperatures and extremely low rainfall in July and August. In both years the plants were grown at two levels of N fertilization (low N and high N) as indicated in Table 1. Nitrogen fertilizer was applied on 7 May in 1987 and on 27 May (80 kg ha- 1) and 22 July (58 kg ha- 1) in 1991. In 1987, plants were grown in plots of 135m2, at a density of 11 plants per m2 in a randomized block design with three replicates. In 1991, plants were grown at the same density in two plots (one low N and one high N plot) with a size of 270 m2 each.
Xylem exudate was collected from plants of both N treatments at several dates during the growing period (Table 2). For the collection of xylem exudate the plants were decapitated in the morning (9.00 to 11.00 a.m.) about 2 em above the soil surface. The cut surface was cleaned with a paper towel and mois-
22 A
20 ~---4 - ' (.) 18 L. Q) 16 .... ::J 1a 14 .... ~ 12
~ 10 8 A
6 M J J A s 0
Month
C. Engels et al.
Table 1. Nitrogen fertilizer applied (kg N ha- 1 as calcium ammonium nitrate) and contents of mineral N (Nmin = kg N ha- 1
in 0-90 em soil depth measured before sowing + N fertilized) in 1987 and 1991.
Low N High N
1987 N fertil.
0 124
Nmin
76 200
N fertil.
0 138
1991 Nmin
102 240
Table 2. Harvest dates for the collection of xylem exudate ; DAS = days after sowing (sowing dates: 25 May, 1987 and 22 May, 1991); PDS =plant developmental stage.
1987 1991 DAS PDS DAS PDS
43 3-4 leaf stage 33 5 leaf stage 52 stem elongation 43 stem elongation 79 panicle initiation 59 panicle initiation 92 anthesis 70 anthesis
106 milk ripeness 94 milk ripeness 123 wax-ripe stage 113 wax-ripe stage
tened with a drop of Micropur (Roth Company, Karlsruhe, Germany) to reduce bacterial growth during exudation. Thereafter a balloon was mounted over the cut stem and sealed at the base with Prestic (Bostic GmbH, Oberursel, Germany). After 3 h the exudate was collected with a syringe and stored in vials on ice until returned to the laboratory for weighing and storing at - 18 oc until analysis.
140 B
120 ·---·- -. -100 E E 80 -s 60 c: "(ij , a: 40 \ ,
4 , , .... . .. , 20 -~
0 M J J A s 0
Month Figure 1. Meteorological data of the experimental site in Hohenheim; A) Mean monthly air temperatures: •: 1987; •: 1991; 0 long-term average. B) Total rainfall per month: • : 1987; • : 1991; 0 long-term average.
Eur. J. ARran.
Nitrogen and sugar in xylem exudate of maize
In 1991, as an additional treatment in several rows of the high N plot, the developing cobs were removed after an thesis (-cob). Furthermore, several rows received an additional late N fertilization (50 kg N ha- 1 as calcium ammonium nitrate, dissolved in water and applied with 15 mm of irrigation water) 75 days after sowing ( + N). Exudate was collected from plants of these plots and compared with control plants (Con, intact plants with no additional late N fertilization) at 93 and 105 days after sowing (DAS).
The concentration of nitrate N in the xylem sap was analysed with a Technicon autoanalyser according to a method of Britt (1962). Total N was determined after oxidation with potassium persulfate (K20 8S2) in a UV -digester. Reduced N was calculated from the difference between total and nitrate-N. Reducing sugars were measured using the p-hydroxybenzoic acid hydroazide reaction (Blakeney and Mutton, 1980), and sucrose from the difference of reducing sugar concentrations with and without incubation of the xylem exudate for 2 hours at 37 oc with invertase (100 units ml- 1 exudate E.C. 3.2.1.26 from SIGMA).
From each treatment four samples of exudate (each representing the exudate of 1-3 plants) were collected at each harvest. All data were statistically evaluated by analysis of variance (ANOV A). Standard errors of the means (SEM) and degrees of freedom for the error variance or the least significant difference for p = 0.05 (LSD 0.05) are indicated in the Figures and in Table 3.
RESULTS
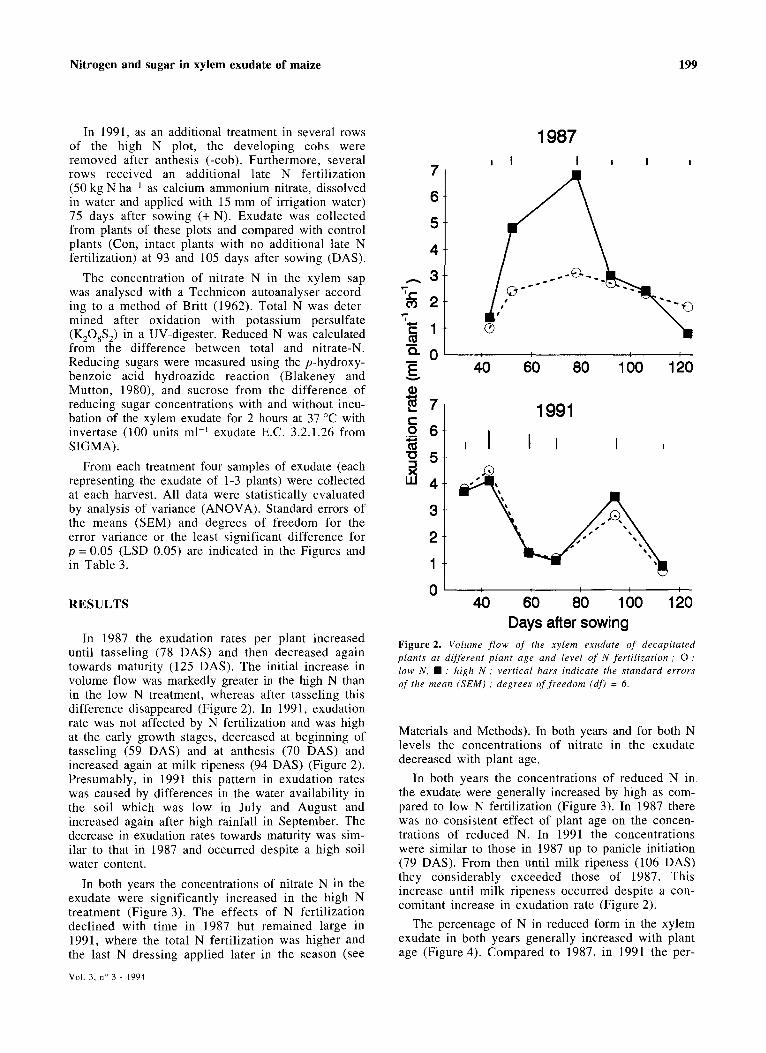
In 1987 the exudation rates per plant increased until tasseling (78 DAS) and then decreased again towards maturity (125 DAS). The initial increase in volume flow was markedly greater in the high N than in the low N treatment, whereas after tasseling this difference disappeared (Figure 2). In 1991, exudation rate was not affected by N fertilization and was high at the early growth stages, decreased at beginning of tasseling (59 DAS) and at anthesis (70 DAS) and increased again at milk ripeness (94 DAS) (Figure 2). Presumably, in 1991 this pattern in exudation rates was caused by differences in the water availability in the soil which was low in July and August and increased again after high rainfall in September. The decrease in exudation rates towards maturity was similar to that in 1987 and occurred despite a high soil water content.
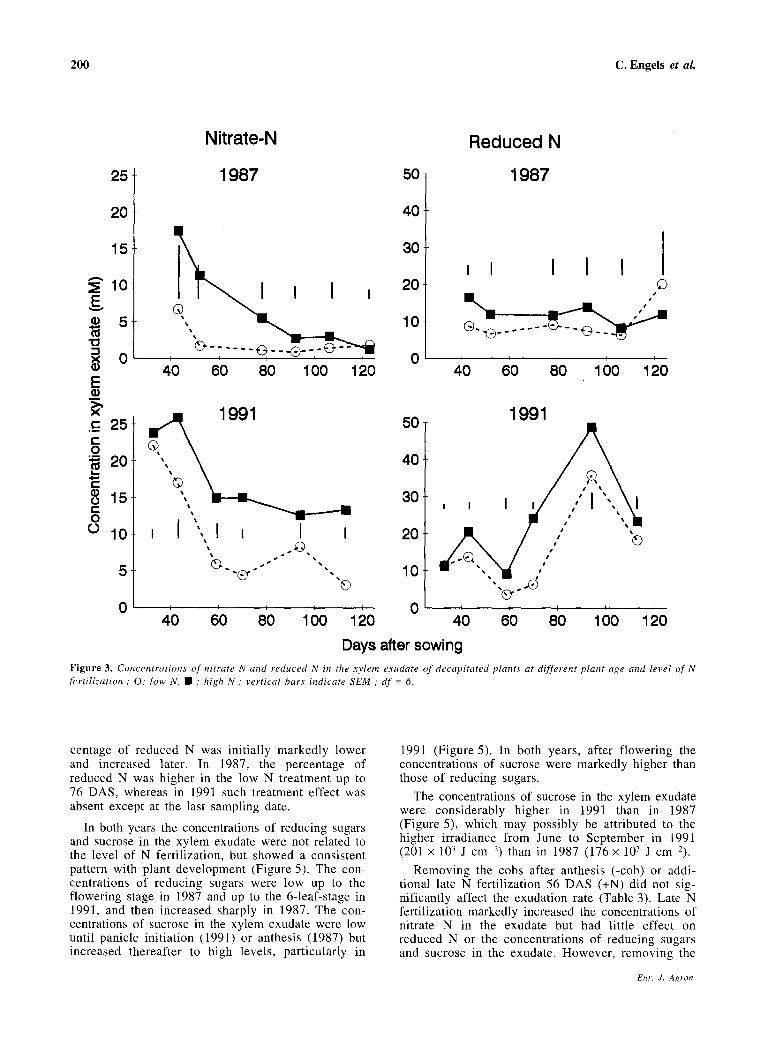
In both years the concentrations of nitrate N in the exudate were significantly increased in the high N treatment (Figure 3). The effects of N fertilization declined with time in 1987 but remained large in 1991, where the total N fertilization was higher and the last N dressing applied later in the season (see
Vol. 3, 0° 3- 1994
199
1987
7
6
5
4
- 3 ";'.s:::
2 C') ";' - 1 fa a. 0 E 40 60 80 100 120 -Q)
1a 7 1991 .... c: 0 ~
6 I
"C 5 ~ 4
3
2
1
0 40 60 80 100 120
Days after sowing Figure 2. Volume flow of the xylem exudate of decapitated plants at different plant age and level of N fertilization ; 0 ; low N, • : high N; vertical bars indicate the standard errors of the mean (SEM); degrees of freedom (df) = 6.
Materials and Methods). In both years and for both N levels the concentrations of nitrate in the exudate decreased with plant age.
In both years the concentrations of reduced N in the exudate were generally increased by high as compared to low N fertilization (Figure 3). In 1987 there was no consistent effect of plant age on the concentrations of reduced N. In 1991 the concentrations were similar to those in 1987 up to panicle initiation (79 DAS). From then until milk ripeness (106 DAS) they considerably exceeded those of 1987. This increase until milk ripeness occurred despite a concomitant increase in exudation rate (Figure 2).
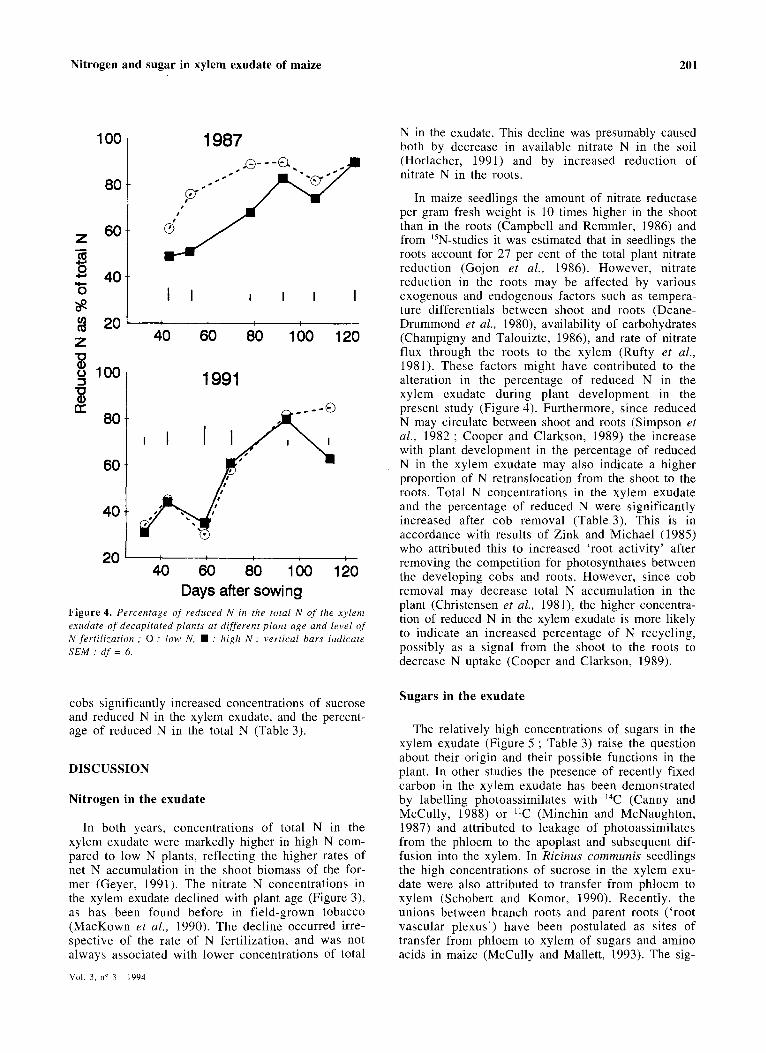
The percentage of N in reduced form in the xylem exudate in both years generally increased with plant age (Figure 4). Compared to 1987, in 1991 the per-
200 C. Engels et al.
Nitrate-N Reduced N
25 1987 50 1987
20 40
15 30
~ 10 20 0 E Q -Q) 5 \ 10 \
1ti \ \
"C 0------0- --G---0--:::::J 0 0 li1 40 60 80 100 120 40 60 80 100 120 E Q)
S<' 1991 1991 .£: 25 50 c:: 0
40 1a 20 \
\ .... \ - Q c:: Q) 15 \ 30 0 \
c:: \ 0 \ , u 10 20 ,
\ ,Q , \ - ' '
, 5 0, ' ' 10 '12! ' ' ,
'0 ' k) '0'
0 0 40 60 80 100 120 40 60 80 100 120
Days after sowing Figure 3. Concentrations of nitrate N and reduced N in the xylem exudate of decapitated plants at different plant age and level of N fertilization; 0: low N, • : high N; vertical bars indicate SEM; df = 6.
centage of reduced N was initially markedly lower and increased later. In 1987, the percentage of reduced N was higher in the low N treatment up to 76 DAS, whereas in 1991 such treatment effect was absent except at the last sampling date.
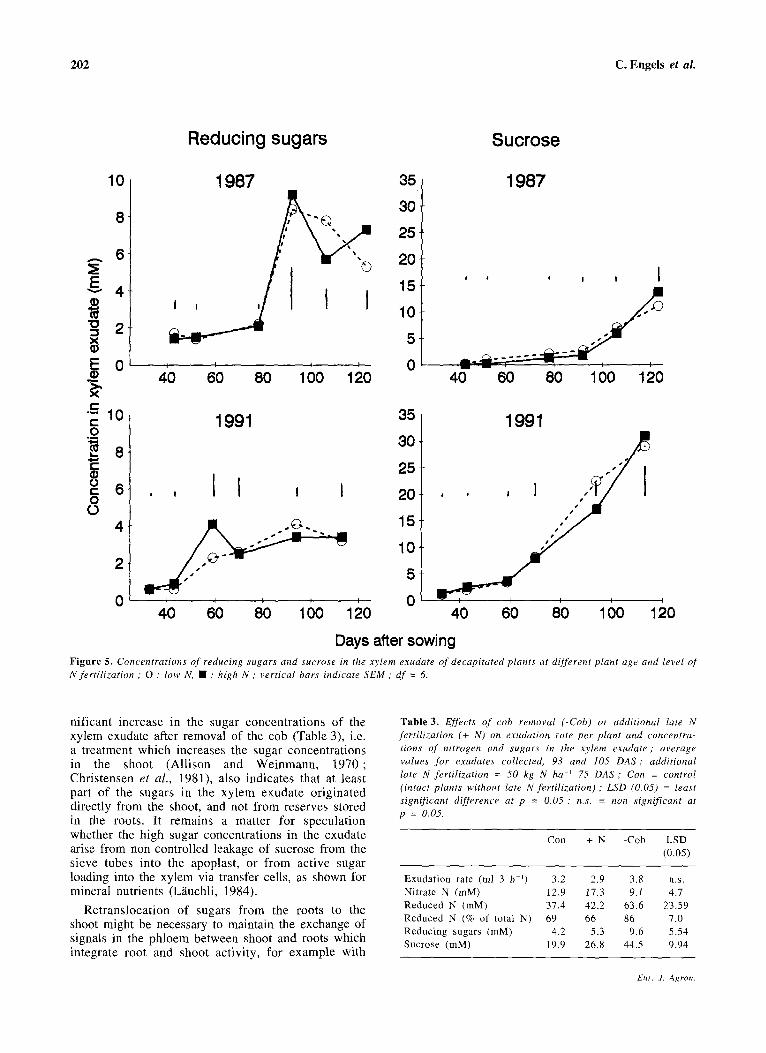
In both years the concentrations of reducing sugars and sucrose in the xylem exudate were not related to the level of N fertilization, but showed a consistent pattern with plant development (Figure 5). The concentrations of reducing sugars were low up to the flowering stage in 1987 and up to the 6-leaf-stage in 1991, and then increased sharply in 1987. The concentrations of sucrose in the xylem exudate were low until panicle initiation (1991) or an thesis ( 1987) but increased thereafter to high levels, particularly in
1991 (Figure 5). In both years, after flowering the concentrations of sucrose were markedly higher than those of reducing sugars.
The concentrations of sucrose in the xylem exudate were considerably higher in 1991 than in 1987 (Figure 5), which may possibly be attributed to the higher irradiance from June to September in 1991 (201 x 103 J cm-2) than in 1987 (176 x 103 J cm-2).
Removing the cobs after an thesis (-cob) or additiona] late N fertilization 56 DAS (+N) did not significantly affect the exudation rate (Table 3). Late N fertilization markedly increased the concentrations of nitrate N in the exudate but had little effect on reduced N or the concentrations of reducing sugars and sucrose in the exudate. However, removing the
Eur. J. ARran.
Nitrogen and sugar in xylem exudate of maize
100 1987
80 0 I
I I
I
z 60 0
a:s -0 40 --0
'* ~ 20 z 40 60 80 100 120 "C Q)
100 0 1991 ::J -g
----8 a: 80
60
40
20~-4----+----+----~--~ 40 60 80 100 120
Days after sowing Figure 4. Percentage of reduced N in the total N of the xylem exudate of decapitated plants at different plant age and level of N fertilization ; 0 ; low N, • : high N; vertical bars indicate SEM; df = 6.
cobs significantly increased concentrations of sucrose and reduced N in the xylem exudate, and the percentage of reduced N in the total N (Table 3).
DISCUSSION
Nitrogen in the exudate
In both years, concentrations of total N in the xylem exudate were markedly higher in high N compared to low N plants, reflecting the higher rates of net N accumulation in the shoot biomass of the former (Geyer, 1991). The nitrate N concentrations in the xylem exudate declined with plant age (Figure 3), as has been found before in field-grown tobacco (MacKown et al., 1990). The decline occurred irrespective of the rate of N fertilization, and was not always associated with lower concentrations of total
Vol. 3, 0° 3- 1994
201
N in the exudate. This decline was presumably caused both by decrease in available nitrate N in the soil (Horlacher, 1991) and by increased reduction of nitrate N in the roots.
In maize seedlings the amount of nitrate reductase per gram fresh weight is 10 times higher in the shoot than in the roots (Campbell and Remmler, 1986) and from 15N-studies it was estimated that in seedlings the roots account for 27 per cent of the total plant nitrate reduction (Gojon et al., 1986). However, nitrate reduction in the roots may be affected by various exogenous and endogenous factors such as temperature differentials between shoot and roots (DeaneDrummond et al., 1980), availability of carbohydrates (Champigny and Talouizte, 1986), and rate of nitrate flux through the roots to the xylem (Rufty et al., 1981 ). These factors might have contributed to the alteration in the percentage of reduced N in the xylem exudate during plant development in the present study (Figure 4). Furthermore, since reduced N may circulate between shoot and roots (Simpson et al., 1982 ; Cooper and Clarkson, 1989) the increase with plant development in the percentage of reduced N in the xylem exudate may also indicate a higher proportion of N retranslocation from the shoot to the roots. Total N concentrations in the xylem exudate and the percentage of reduced N were significantly increased after cob removal (Table 3). This is in accordance with results of Zink and Michael (1985) who attributed this to increased 'root activity' after removing the competition for photosynthates between the developing cobs and roots. However, since cob removal may decrease total N accumulation in the plant (Christensen et al., 1981 ), the higher concentration of reduced N in the xylem exudate is more likely to indicate an increased percentage of N recycling, possibly as a signal from the shoot to the roots to decrease N uptake (Cooper and Clarkson, 1989).
Sugars in the exudate
The relatively high concentrations of sugars in the xylem exudate (Figure 5 ; Table 3) raise the question about their origin and their possible functions in the plant. In other studies the presence of recently fixed carbon in the xylem exudate has been demonstrated by labelling photoassimilates with 14C (Canny and McCully, 1988) or 11 C (Minchin and McNaughton, 1987) and attributed to leakage of photoassimilates from the phloem to the apoplast and subsequent diffusion into the xylem. In Ricinus communis seedlings the high concentrations of sucrose in the xylem exudate were also attributed to transfer from phloem to xylem (Schobert and Komor, 1990). Recently, the unions between branch roots and parent roots ('root vascular plexus') have been postulated as sites of transfer from phloem to xylem of sugars and amino acids in maize (McCully and Mallett, 1993). The sig-
202 C. Engels et al.
Reducing sugars Sucrose
10 1987 35 1987
e 30
25
- 6 20 ~ g 4 15 .m (U 10 '0 2 :::J 5 ~ E 0 0 G) 40 60 eo 100 120 ~
40 eo 120 100 60
.5 10 35 t: 1991 0 ~ e 30 '--t: 25 G)
1991
0 6 t: 20 0 (.)
4 15
10 2
5
0 0 40 60 eo 100 120 40
Days after sowing
60 eo 100 120
Figure 5. Concentrations of reducing sugars and sucrose in the xylem exudate of decapitated plants at different plant age and level of N fertilization ; 0 : low N, • : high N; vertical bars indicate SEM ; df = 6.
nificant increase in the sugar concentrations of the xylem exudate after removal of the cob (Table 3), i.e. a treatment which increases the sugar concentrations in the shoot (Allison and Weinmann, 1970; Christensen et al., 1981 ), also indicates that at least part of the sugars in the xylem exudate originated directly from the shoot, and not from reserves stored in the roots. It remains a matter for speculation whether the high sugar concentrations in the exudate arise from non-controlled leakage of sucrose from the sieve tubes into the apoplast, or from active sugar loading into the xylem via transfer cells, as shown for mineral nutrients (Uiuchli, 1984).
Retranslocation of sugars from the roots to the shoot might be necessary to maintain the exchange of signals in the phloem between shoot and roots which integrate root and shoot activity, for example with
Table 3. Effects of cob removal (-Cob) or additional late N fertilization ( + N) on exudation rate per plant and concentrations of nitrogen and sugars in the xylem exudate ; average values for exudates collected, 93 and 105 DAS ; additional late N fertilization = 50 kg N ha- 1 75 DAS; Con = control (intact plants without late N fertilization) ; LSD (0.05) = least significant difference at p = 0.05 ; n.s. = non significant at p = 0.05.
Con + N -Cob LSD (0.05)
Exudation rate (rnl 3 h- 1) 3.2 2.9 3.8 n.s. Nitrate N (rnM) 12.9 17.3 9.1 4.7 Reduced N (rnM) 37.4 42.2 63.6 23.59 Reduced N (% of total N) 69 66 86 7.0 Reducing sugars (mM) 4.2 5.3 9.6 5.54 Sucrose (mM) 19.9 26.8 44.5 9.94
Eur. J. A11ron.

Nitrogen and sugar in xylem exudate of maize
regard to N nutrition (Cooper and Clarkson, 1989). Furthermore, the high concentrations of sugars found in the present study might significantly contribute to the osmotic driving force for xylem volume flow, particularly late in the season when the nutrient availability in the rooting zone decreases. The maintenance of root pressure would be necessary to supply the developing ear with phloem-immobile nutrients like Ca or to 'repair' cavitated xylem vessels (Tyree et al., 1986).
The sugar concentrations in the exudate sharply increased after anthesis (Figure 5). During this time the relative sink strength for assimilates of the roots compared to that of the shoot decreases, as indicated by net losses of root weight and root length between anthesis and maturity (Mengel and Barber, 1974). Recently fixed assimilates possibly are not the sole source of sugars in the exudate. As shown by Canny and McCully (1988) for field-grown maize, in the xylem exudate large differences persisted between the distribution of recently fixed 14C and formerly fixed 12C into different fractions (neutral : cationic : anionic). This indicates different origins of the recently assimilated 14C-compounds and the nonlabelled 12C-carbon in the xylem exudate. Possibly, the sharp increase in sugar concentrations in the xylem exudate after anthesis (Figure 5) was mainly caused by remobilization of previously accumulated sugars in the roots. Accumulation of significant amounts of non-structural carbohydrates in the stalks and leaves until anthesis (Kiniry et al., 1992), and subsequent remobilization and utilization for grain growth (Daynard et al., 1969 ; McPherson and Boyer, 1977) is a well known phenomenon in maize. Also in the roots of maize significant amounts of sugars (Engels and Marschner, unpublished results) and phosphorus (Fusseder, 1987 ; Kraus et al., 1987) may accumulate. The remobilisation from the roots and translocation to the shoot of previously stored N (Cliquet et al., 1990 ; Weiland and Ta, 1992) and C (Cliquet et al., 1990) has been shown. Since in maize senescent cortical root cells are protected from microbial attack by a suberized exodermis (Peterson, 1989), the nodal roots as well as the stalks might serve as nutrient storing tissue. The possible impact on grain filling of C and N compounds remobilized from the roots needs further investigation.
ACKNOWLEDGEMENT
This research was supported in part by the Deutsche Forschungsgemeinschaft (SFB 183 ). The authors thank Hinrich Bremer and Hauke Heuwinkel for excellent technical assistance.
Vol. 3, 0° 3 ~ 1994
203
REFERENCES
Allison J. C. S. and Weinmann H. (1970). Effect of absence of developing grain on carbohydrate content and senescence of maize leaves. Plant Physiol., 46, 435-436.
Blakeney A. B. and Mutton L. L. (1980). A simple colorimetric method for the determination of sugars in fruit and vegetables. f. Sci. Food Agric., 31, 889-897.
Britt R. D. (1962). Precise automatic spectrophotometric analysis in the low parts per billion range. Anal. Chern., 34, 1728-1731.
Butz R. G. and Long R. C. (1979). L-malate as an essential component of the xylem fluid of corn seedling roots. Plant Physiol., 64, 684-689.
Campbell C. and Remmler K. (1986). Regulation of corn leaf nitrate reductase. I. Immunochemical methods for analysis of the enzymes protein component. Plant Physiol., 80, 435-441.
Canny M. J. and McCully M. E. (1988). The xylem sap of maize roots : its collection, composition and formation. Aust. f. Plant Physiol., 15, 557-566.
Champigny M. L. and Talouizte A. (1986). Dependence of nitrate reduction on root soluble carbohydrates in wheat seedlings. In : Lambers H., Neeteson J. J. and Stulen I. (Eds.) Fundamental, ecological and agricultural aspects of nitrogen metabolism in higher plants. Dordrecht : Martinus Nijhoff, 279-282.
Christensen L. E., Below F. E. and Hageman R. H. (1981). The effects of ear removal on senescence and metabolism of maize. Plant Physiol., 68, 1180-1185.
Cliquet J.-B., Deleens E. and Mariotti A. (1990). C and N mobilization from stalk and leaves during kernel filling by 13C and 15N tracing in Zea mays L. Plant Physiol., 94, 1547-1553.
Cooper H. D. and Clarkson D. T. (1989). Cycling of amino nitrogen and other nutrients between shoots and roots in cereals - a possible mechanism integrating shoot and root in the regulation of nutrient uptake. f. exp. Bot., 40, 753-762.
Daynard T. B., Tanner J. W. and Hume D. J. (1969). The contribution of stalk soluble carbohydrates to grain yield in corn (Zea mays L.). Crop Sci., 9, 831-834.
Deane-Drummond C. E., Clarkson D. T. and Johnson C. B. (1980). The effect of differential root and shoot temperature on the nitrate reductase activity, assayed in vivo and in vitro in roots of Hordeum vulgare (barley). Relationship with diurnal changes in endogenous malate and sugar. Planta, 148, 455-461.
Fusseder A. (1987). The longevity and activity of the primary root of maize. Plant Soil, 101, 257-265.
Geyer B. (1991). Wachstum, Stickstoffaufnahme und Nitratspeicherung bei Mais (Zea mays L.) nach Dlingung mit Glille und Mineralstickstoff. Doctoral Thesis, Institut flir Pflanzenernahrung, Universitat Hohenheim.
Gojon A., Soussana J. F., Passama L. and Robin P.

204
(1986). Nitrate reduction in roots and shoots of barley (Hordeum vulgare L.) and corn (Zea mays L.) seedlings. I. 15N study. Plant Physiol., 82, 254-260.
Hardy P. J. and Possingham J. V. (1969). Studies on translocation of metabolites in the xylem of grapevine shoots. J. exp. Bot., 20, 325-335.
Horlacher D. (1991). Einfluj3 organischer und mineralischer N-Diinger auf Sproj3wachstum und Nitratauswaschung bei Silomais sowie Quantifizierung der Ammoniakverluste nach Ausbringung von Fliissigmist. Doctoral Thesis, lnstitut fiir Pflanzenernahrung, Universitat Hohenheim.
Keltjens W. G. and Nijenstein J. H. (1987). Diurnal variations in uptake, transport and assimilation of N03- and efflux of OH- in maize plants. J. Plant Nutr., 10, 887-900.
Keltjens W. G., Nieuwenhuis J. W. and Nelemans J. A. (1986). Nitrogen retranslocation in plants of maize, lupin, and cocklebur. Plant Soil, 91, 323-328.
Kiniry J. R., Tischler C. R., Rosenthal W. D. and Gerik T. J. (1992). Nonstructural carbohydrate utilization by sorghum and maize shaded during grain growth. Crop Sci., 32, 131-137.
Kraus M., Fusseder A. and Beck E. (1987). Development and replenishment of the P-depletion zone around the primary root of maize during the vegetation period. Plant Soil, 101, 247-255.
Lauchli A. (1984). Mechanisms of nutrient fluxes at membranes of the root surface and their regulation in the whole plant. In : Barber S. A. and Bouldin D. R. (Eds.) Roots, Nutrient and Water Influx, and Plant Growth. ASA Special Publication No 49, Soil Sci. Soc. Am., 1-25.
McCully M. E. and Mallett J. E. (1993). The branch roots of Zea. 3. Vascular connections and bridges for nutrient cycling. Ann. Bot., 71, 327-341.
McKown C. T., Sutton T. G. and Bush L. P. ( 1990). Nitrogen compositional changes in xylem exudate and leaves of burley tobacco. Crop Sci., 30, 133-138.
McPherson H. G. and Boyer J. S. (1977). Regulation of grain yield by photosynthesis in maize subjected to a water deficiency. Agron. J., 69, 714-718.
Mengel D. B. and Barber S. A. (1974). Development and
C. Engels et al.
distribution of the corn root system under field conditions. Agron. J., 66, 341-344.
Minchin P. E. H. and McNaughton G. S. (1987). Xylem transport of recently fixed carbon within Jupin. Aust. J. Plant Physiol., 14, 325-329.
Pate J. S. (1975). Exchange of solutes between phloem and xylem and circulation in the whole plant. In : Zimmermann M. H. and Milburn J. A. (Eds.) Transport in Plants. I. Phloem Transport. Encycl. Plant Physiol. New Series, Vol. 1. Berlin : SpringerVerlag, 451-473.
Peterson C. A. (1989). Significance of the exodermis in root function. In: Loughman B. C., Gasparikova 0. and Kolek J. (Eds.) Structural and functional aspects of transport in roots. Dordrecht : Kluwer Academic Publishers, 35-40.
Rufty T. W., Jr., Jackson W. A. and Raper Jr., C. D. (1981). Nitrate reduction in roots as affected by the presence of potassium and by flux of nitrate through the roots. Plant Physiol., 68, 605-609.
Schobert C. and Komar E. (1990). Transfer of amino acids anti nitrate from the roots into the xylem of Ricinus communis seedlings. Planta, 181, 85-90.
Simpson R. J., Lambers H. and Dalling M. J. (1982). Translocation of nitrogen in a vegetative wheat plant. Physiol. Plant., 56, 11-17.
Teyker R. H., Galerani P.R. and Nafziger E. D. (1991). Analysis of xylem exudate by ion chromatography : influence of nitrogen and residue management on corn exudate composition. Commun. Soil Sci. Plant Anal., 22, 785-793.
Tyree M. T., Fiscus E. L., Wullschleger S.D. and Dixon M. A. (1986). Detection of xylem cavitation in corn under field conditions. Plant Physiol., 82, 597-599.
Weiland R. T. and Ta T. C. (1992). Allocation and retranslocation of 15N by maize (Zea mays L.) hybrids under field conditions of low and high N fertility. Aust. J. Plant Physiol., 19, 77-88.
Zink F. and Michael G. (1985). The effect of reduced number of kernels and of leaves per plant on the nitrogen storage characteristics of the kernels of two maize hybrids with different grain-N-content. J. Agron. Crop Sci., 154, 203-212.
Eur. J. A.Rrcm.