neuronal activation of the hypothalamic magnocellular system in response to oropharyngeal stimuli in...
TRANSCRIPT

Journal of Neuroendocrinology, 1997, Vol. 9, 329–340
Neuronal Activation of the Hypothalamic Magnocellular System inResponse to Oropharyngeal Stimuli in the Rat
Naceur Naımi*, Serge Rivest†, Ilie Racotta‡, and Denis Richard**Departement de Physiologie, Faculte de Medecine, Universite Laval, Quebec (Que), G1K 7P4, Canada.†Laboratoire d’Endocrinologie moleculaire, Centre hospitalier de l’Universite Laval, 2705 boulevard Laurier, Quebec, G1V 4G2.‡Departamento de Fisiologia, Escuela Nacional de Ciencias Biologicas, Prolongation Carpio y Plan de Ayala, Mexico, DF 11340.
Key words: arginine-vasopressin, oxytocin brain, c-fos mRNA, hypothalamus, immediate-early genes, in situ hybridization,paraventricular nucleus of the hypothalamus, supraoptic nucleus.
Abstract
The present study was designed to delineate the neuronal site, the nature, and the gastrointestinal origin of the stimulation of thehypothalamic magnocellular system induced by the ingestion of sweetened condensed milk. Concomitant localization of the c-fos protein(Fos) with either arginine-vasopressin (AVP) mRNA or oxytocin (OT) mRNA in the paraventricular nucleus of the hypothalamus (PVH) andthe supraoptic nucleus (SON) revealed that the hypothalamic neurons containing AVP and OT were activated following ingestion ofsweetened condensed milk. Expression of c-fos mRNA was also determined in rats implanted with a gastric cannula that allowed for real,sham, and gastric feeding of sweetened condensed milk. The results provide evidence that the stimulation of the PVH and SON inducedby sweetened condensed milk originate from oropharyngeal stimuli. Indeed, in real-and sham-fed rats, the postprandial levels of c-fosmRNA in the PVH and SON were significantly higher than the preprandial values, whereas there was no early postprandial rise in c-fosmRNA levels within the magnocellular division of the PVH and SON after gastric feeding. The results of this study also suggested thatthe stimulation of the PVH and SON induced by sweetened condensed milk was related to the hypertonicity of the milk. Indeed, ingestionof an hypertonic solution of sucrose with a carbohydrate content close to that of sweetened condensed milk led to a stimulation of thePVH and SON that was comparable to that induced by the milk, whereas ingestion of an isotonic solution of sucrose did not trigger anysignificant activation of the PVH and SON. Taken together, the present results indicate that magnocellular neurosecretory neurons aresensitive to oropharyngeal stimuli and fur ther support the view of the existence of oropharyngeal osmoreceptors.
In a recent series of experiments undertaken to determine the the hypothalamic magnocellular neurosecretory system translatesinto activations of the AVP and OT neuronal systems. To achievebrain sites involved in the thermogenic and satiating responses to
food, we made the observation that the intake of sweetened this objective, concomitant measurements of Fos and either AVPmRNA and OT mRNA were carried out on brain sections ofcondensed milk may lead to a rapid and transient expression of
c-fos and NGFI-B genes in the magnocellular division of the rats killed following ingestion of sweetened condensed milk. Thee�ects of the milk were also examined in rats implanted with aparaventricular nucleus of the hypothalamus (PVH) and in the
supraoptic nucleus (SON ) (1). Being very low in most brain gastric cannula that allows for real, sham and gastric feeding.These preparations, which have been widely used in studies onstructures under basal resting conditions and transiently induced
in activated brain areas by a variety of stimuli (2–10), the feeding physiology, (15–18), were used as a strategy to furtherdelineate the nature and gastrointestinal origin of the stimuliexpression of c-fos protein (Fos), c-fos mRNA (11–13) and
NGFI-B mRNA (4) have emerged in recent years as reliable triggering the activity of hypothalamic magnocellular neurosecre-tory cells in response to sweetened condensed milk. Finally, itmarkers for neuronal activity.
The large neurons of the SON and PVH that project to the was also one of the intents of the present study to investigate thee�ect of the hypertonicity of sweetened condensed milk on theneural lobe of the hypophysis have traditionally been referred to
as the hypothalamic magnocellular neurosecretory cells (14). hypothalamic neurosecretory system, osmotic stimuli (19, 20)being classified as among the most potent activators of the AVPGiven the fact that virtually all the magnocellular neurosecretory
cells synthesize and secrete either arginine-vasopressin (AVP) or and OT cells of the PVH and SON. To achieve this objective and,because hypertonicity of sweetened condensed milk is primarilyoxytocin (OT ) (14), it was one of the objectives of this study to
examine whether the influence of sweetened condensed milk on determined by its mono/disaccharide content, experiments were
Correspondence to: Denis Richard, Departement de Physiologie, Faculte de Medecine, Universite Laval, Quebec, Canada, G1K 7P4.
© 1997 Blackwell Science Ltd
330 Sensory activation of the vasopressinergic and oxytocinergic neurons
carried out to compare sweetened condensed milk with hypertonicand isotonic sucrose solutions in their e�ects in inducing c-fosexpressions in the PVH and SON.
Results
Figure 1 illustrates the e�ects of real, sham, and gastric feedingof sweetened condensed milk on the pre- and postprandial plasmalevels of glucose. ANOVA revealed a significant ‘feeding’בtime’interaction on plasma glucose. Whereas postprandial elevationsin glucose were observed after real and gastric feeding, there wasno significant postprandial increase in plasma glucose in sham-fed rats, indicating that the sham-procedure used in this studywas adequate in regard to preventing milk from entering thesmall intestine. A posteriori comparisons revealed that the post-prandial elevation seen after gastric feeding was higher than thatseen after real feeding, this insuring that the milk injected in thestomach e�ciently reached the gut.
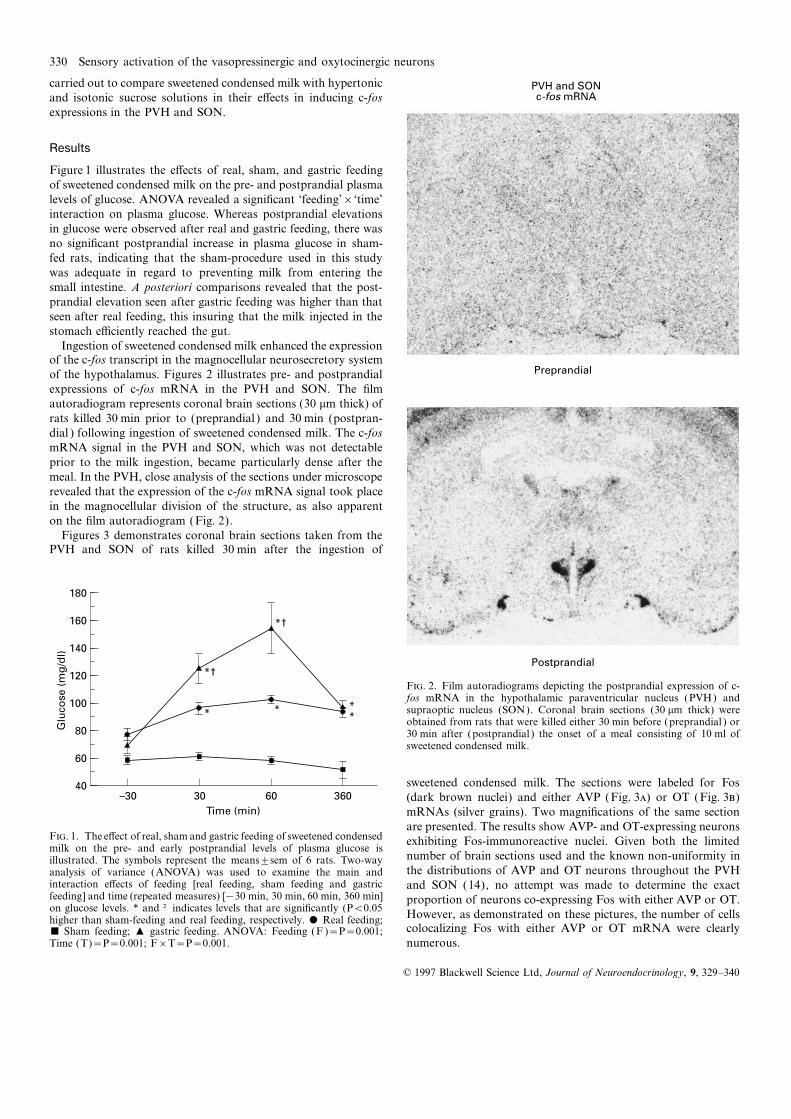
Ingestion of sweetened condensed milk enhanced the expressionof the c-fos transcript in the magnocellular neurosecretory systemof the hypothalamus. Figures 2 illustrates pre- and postprandialexpressions of c-fos mRNA in the PVH and SON. The filmautoradiogram represents coronal brain sections (30 mm thick) ofrats killed 30 min prior to (preprandial ) and 30 min (postpran-dial ) following ingestion of sweetened condensed milk. The c-fosmRNA signal in the PVH and SON, which was not detectableprior to the milk ingestion, became particularly dense after themeal. In the PVH, close analysis of the sections under microscoperevealed that the expression of the c-fos mRNA signal took placein the magnocellular division of the structure, as also apparenton the film autoradiogram (Fig. 2).
Figures 3 demonstrates coronal brain sections taken from thePVH and SON of rats killed 30 min after the ingestion of
Postprandial
PVH and SONc-fos mRNA
Preprandial
F. 2. Film autoradiograms depicting the postprandial expression of c-fos mRNA in the hypothalamic paraventricular nucleus (PVH) andsupraoptic nucleus (SON). Coronal brain sections (30 mm thick) wereobtained from rats that were killed either 30 min before (preprandial ) or30 min after (postprandial ) the onset of a meal consisting of 10 ml ofsweetened condensed milk.
sweetened condensed milk. The sections were labeled for Fos(dark brown nuclei) and either AVP (Fig. 3) or OT (Fig. 3)mRNAs (silver grains). Two magnifications of the same section
360
180
40
Time (min)
Glu
co
se
(m
g/d
l)
160
80
60
60–30 30
100
140
120
**
**
*†
*†
are presented. The results show AVP- and OT-expressing neuronsF. 1. The e�ect of real, sham and gastric feeding of sweetened condensed exhibiting Fos-immunoreactive nuclei. Given both the limitedmilk on the pre- and early postprandial levels of plasma glucose is
number of brain sections used and the known non-uniformity inillustrated. The symbols represent the means±sem of 6 rats. Two-waythe distributions of AVP and OT neurons throughout the PVHanalysis of variance (ANOVA) was used to examine the main and
interaction e�ects of feeding [real feeding, sham feeding and gastric and SON (14), no attempt was made to determine the exactfeeding] and time (repeated measures) [−30 min, 30 min, 60 min, 360 min] proportion of neurons co-expressing Fos with either AVP or OT.on glucose levels. * and † indicates levels that are significantly (P<0.05 However, as demonstrated on these pictures, the number of cellshigher than sham-feeding and real feeding, respectively. $ Real feeding;
colocalizing Fos with either AVP or OT mRNA were clearly& Sham feeding; + gastric feeding. ANOVA: Feeding (F )=P=0.001;Time (T)=P=0.001; F×T=P=0.001. numerous.
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340

Sensory activation of the vasopressinergic and oxytocinergic neurons 331
Fos-ir/AVP mRNA
Fos-ir/OT mRNA
PVH SON
SONPVH
F. 3. Photomicrographs of brain sections of the hypothalamic paraventricular nucleus (PVH) and supraoptic nucleus (SON) illustrating c-fos protein(brown nuclei) and either arginine-vasopressin (AVP) (Fig. ) or oxytocin (OT ) (Fig. ) mRNA (silver grains within cytoplasm). Brain sections (30 mmthick) were obtained from rats that were killed 30 min after the onset of a meal consisting of 10 ml of sweetened condensed milk. Some of theco-localizations are indicated by arrows. Magnification, ×50 (top panels) and ×250 (bottom panels).

332 Sensory activation of the vasopressinergic and oxytocinergic neurons
In this study, intronic in situ hybridization proved to be very (4), dehydration (5), salt loading (4, 6), interleukin-1 (7, 23, 24),useful in allowing visualization of a transcriptional activity that immune challenge (8), and fenfluramine (9, 10). With respect towould have been otherwise not detectable using exonic in situ the physiology of feeding, Fos immunoreactivity, seen mostly inhybridization histochemistry. Indeed, enhanced transcriptional the brainstem, has been observed in response to cholecystokininactivation of the AVP gene was detected using an intron-specific (25–28), bombesin (29), glucoprivic (2-deoxy-D-glucose) andcRNA probe which allows hybridization to nucleotide sequences lipoprivic (2-mercaptoacetate) drugs (30), taste-aversive sub-of the hnRNA before the processing to mRNA (Fig. 4). In stances (25, 31–33), serotoninergic anorectic agents (9, 34, 35)contrast, comparison of basal mRNA levels of either AVP or OT and gastric distention (32, 36).(data not shown) with postprandial levels did not reveal any An apparently large proportion of PVH and SON neuronschanges in the transcriptional activity of these genes. co-localized Fos with either AVP or OT mRNAs following
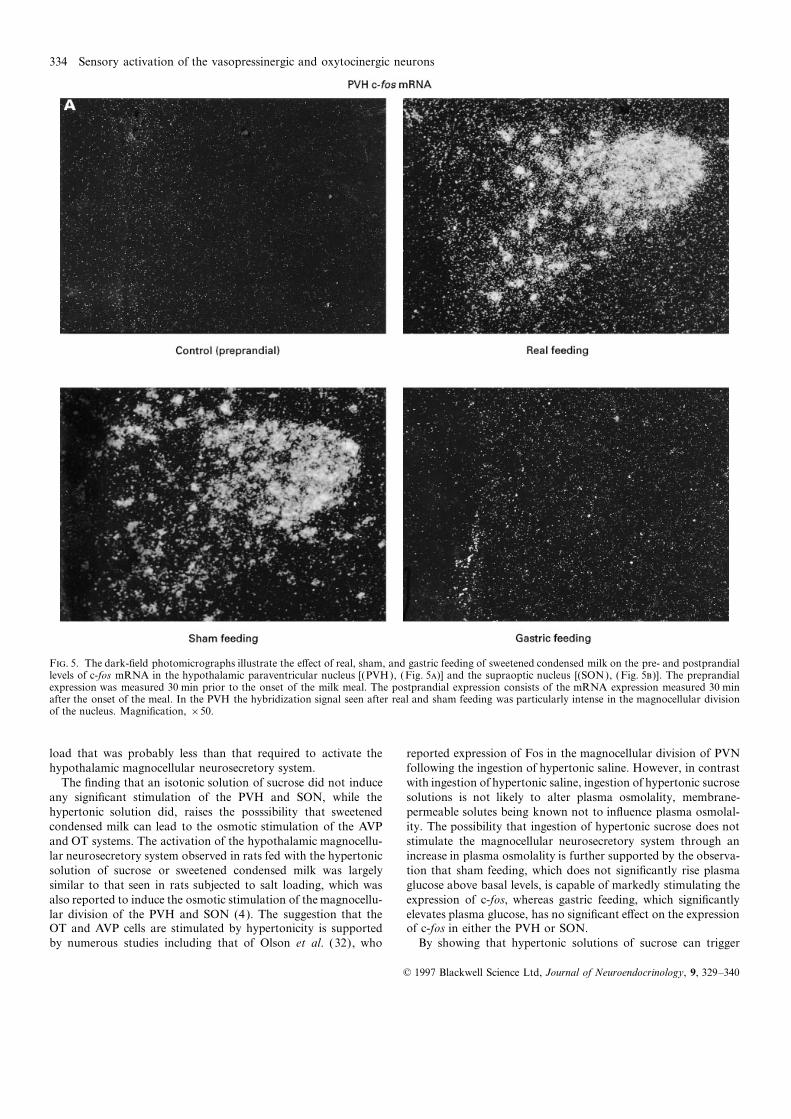
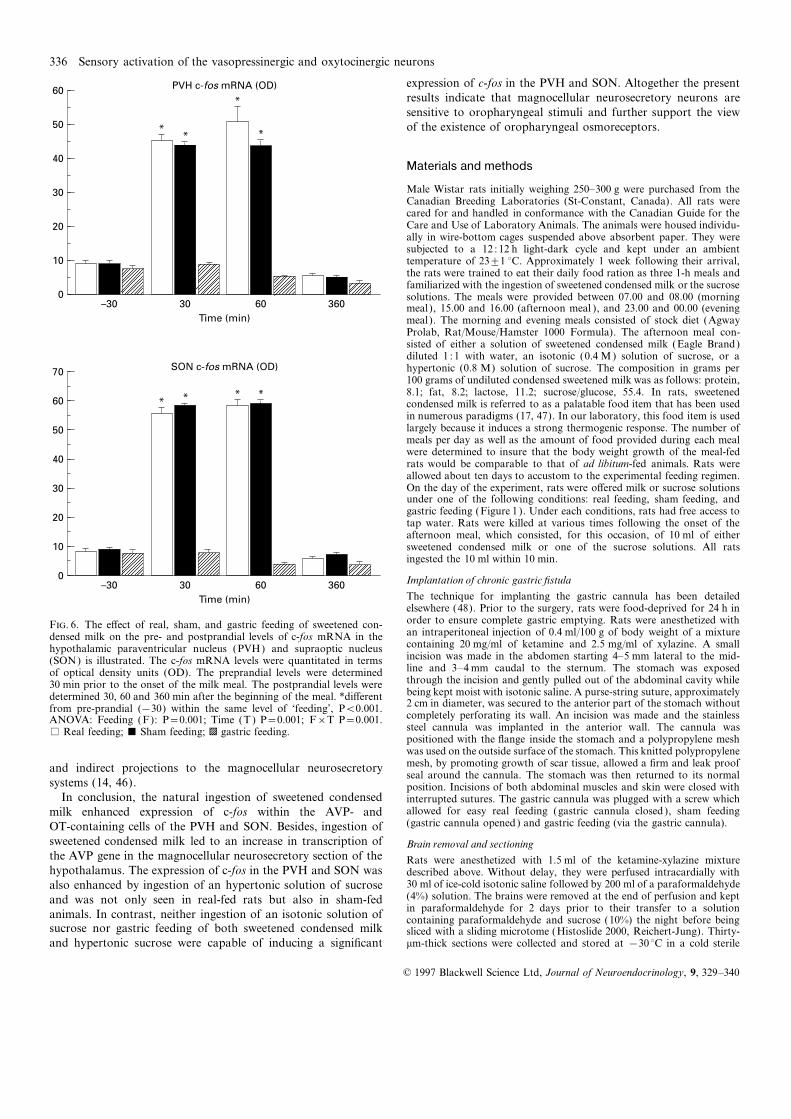
Figures 5 and 6 illustrate the e�ects of real, sham, and gastric ingestion of sweetened condensed milk. This suggests that inges-feeding of sweetened condensed milk on the pre- and postprandial tion of sweeteend condensed milk can activate AVP and OTlevels of c-fos mRNA in the PVH and SON. The hybridization neurons, which constitute the most prevalent neurons of thesignals detected 30 min after the onset of the meal are depicted hypothalamic magnocellular neurosecretory system. The numberin figures 5 and 5. ANOVA of the OD of the c-fos hybridization of cells colocalizing Fos with AVP mRNA following the naturalsignals (Fig. 6) revealed a significant ‘feeding’בtime’ interaction. ingestion of sweetened condensed milk appears to be particularlyIn sham- and real-fed rats, the mRNA levels measured 30 and considerable. In this regard, it is noteworthy that the intake of60 min after the onset of the meal were significantly higher than milk also stimulated the expression of AVP hnRNA in both SONthe preprandial values. In contrast, there was no postprandial and PVH, suggesting a direct involvement of Fos in the transcrip-rise in c-fos mRNA levels following gastric feeding. Sham feeding tion of the AVP gene. This gene comprises on its promoter regioninduced a postprandial increase in the expression of c-fos that consensus base sequences resembling to the aminopyridine-1was not di�erent from that observed in real-fed rats. The photo- (AP-1) binding site, upon which Fos, in the form of an heterodi-micrographs again indicate that the hybridization signal seen in merized complex with Jun (the protein product of the IEGthe PVH was largely concentrated in the magnocellular division c-jun), binds to modulate gene expression. That Fos can directlyof the structure. The postprandial increases in mRNA levels modulate AVP gene expression is also in line with the observationobserved in real- and sham-fed rats were transient; in all condi- that maximal expressions of Fos and AVP hnRNA in responsetions, the mRNA levels assessed 360 min after the onset of the to stress occurred as two consecutive events separated by a shortmeal were comparable to preprandial values. interval of time (37). Sweetened condensed milk also induced
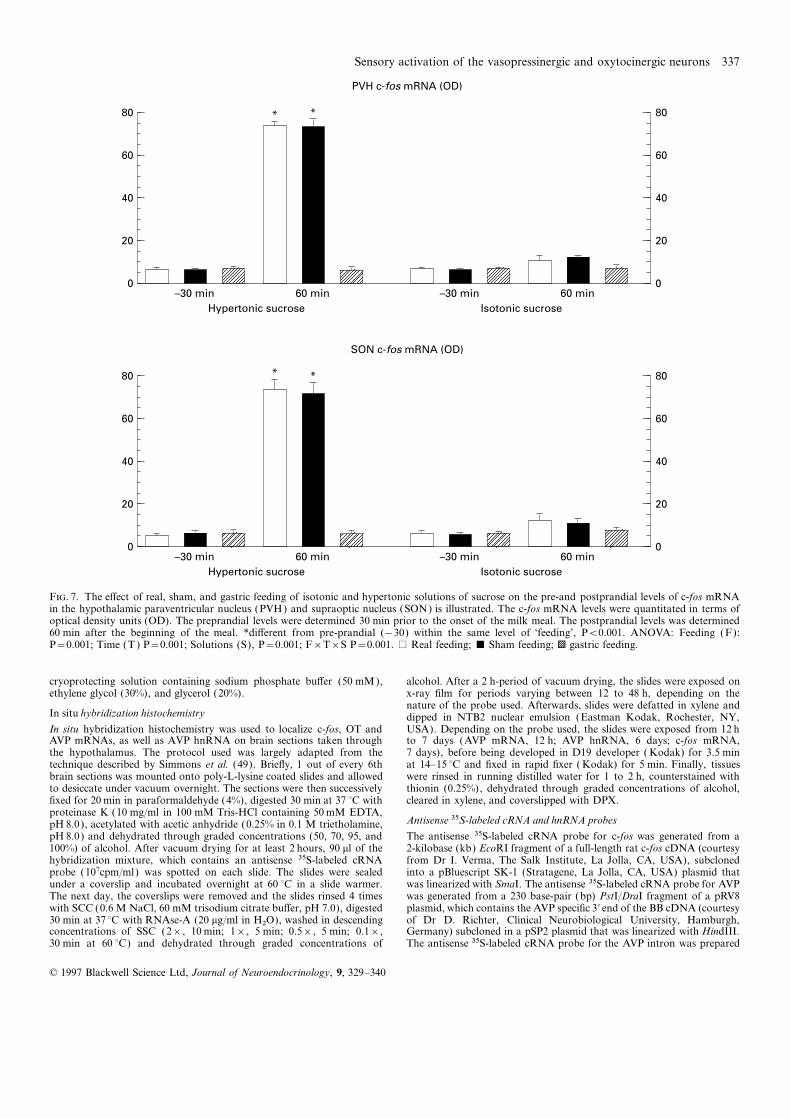
Figure 7 summarizes the e�ects of two sucrose solutions on the Fos expression in OT-containing cells. The OT gene, similar toOD of the c-fos mRNA signal in the PVN and SON. Figures 8 the AVP gene, contains non canonical AP-1 consensus sequencesillustrates the e�ects of real, sham, and gastric feeding of sucrose upon which the Fos/Jun heterodimer complex could act. However,solutions on the pre- and postprandial levels of c-fos mRNA in
we have so far been unable to detect any transcription of OTthe PVN. ANOVA revealed a significant ‘feeding’בtime’×hnRNA following ingestion of sweetened condensed milk. Thissolution’ interaction on the OD od the c-fos mRNA signal infailure, which does not ultimately preclude the possibility thatboth the PVH and SON. When the rats were real- and sham-fedsweetened condensed milk can influence the regulation of the OTwith the hypertonic sucrose solution, postprandial c-fos mRNAgene transcription, may be indicative of an action of Fos on otherlevels were significantly higher than the preprandial values. Thisgenes. Indeed, Fos can modulate the expression of many othermeal e�ect was not observed after gastric feeding and, moreover,genes including that of AVP, which has been reported to coexistnot seen in any feeding conditions when the rats were fed with awith OT in neurons of the PVH and SON (38).isotonic sucrose solution.
The present results also revealed that the magnocellular neuro-secretory cells of the hypothalamus can be activated by oro-
Discussion pharyngeal stimuli. Indeed, the postprandial expression of c-fosin the PVH and SON, was induced by real and sham feeding butThe present results provide evidence that the ingestion ofnot in response to the gastric infusion of sweetened condensedsweetened condensed milk can be a potent stimulator of themilk. Because plasma glucose levels of sham-fed rats did notmagnocellular neurosecretory system in the rat. High levels of c-elevate above basal values during the postprandial period, it canfos mRNA were measured in the magnocellular neurons of PVHbe argued that the sham feeding preparation used in this studyand SON after the natural ingestion of the milk. Fos and c-foswas e�cient in preventing most of the ingested milk from reachingmRNA have emerged in recent years as reliable markers ofintestine. Sham feeding has previously been used to examine theneuronal activity (11, 12, 21). The c-fos gene, which is generallyinfluence of oro-pharyngeal factors in the absence of postingestivenot expressed in non-stimulated neurons, is quickly and transi-intestinal feedbacks (15–18). On the other hand, our finding thatently expressed in activated cells. Fos can be immuno-detectedgastric feeding did not activate c-fos expression in the PVH andduring the 0.5- to 5-hour period that follows neuronal activation.SON somewhat contrasts with that of Olson et al. (32), whoThe use of Fos and c-fos mRNA as markers for neuronaldemonstrated the presence of Fos in the OT-associated areas ofactivation is further warranted by the fact that c-fos expressionthe magnocellular PVH after gastric distension. It must beis induced by numerous extracellular stimuli and may occur in amentioned that Olson et al. (32) observed Fos immunoreactivitylarge number of brain structures. Use of Fos and c-fos mRNAin the PVH in response to the ingestion of a 28 ml load (32),as markers of activated systems has proved to be particularlywhich exceeds by far the volume ingested in this study. In thisrelevant in the delineation of neuroendocrine circuits stimulated
by immobilization (2, 22), treadmill running (3), hemorrhage work, gastric distention was induced by 10 ml of milk, a gastric
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340

Sensory activation of the vasopressinergic and oxytocinergic neurons 333
Pre
pra
nd
ial
Po
stp
ran
dia
l
AV
P h
nR
NA
PV
H
Pre
pra
nd
ial
Po
stp
ran
dia
l
AV
P h
nR
NA
SO
N
14
OD units
2
Op
tical d
en
sit
y
14
12
10 8 6 4 0
pre
*
20
OD units
8 4 0
pre
*
16
12
po
st
po
st
F
.4.
Dar
k-fie
ldph
otom
icro
grap
hsof
coro
nals
ectio
nsde
pict
ing
vaso
pres
sin(A
VP
)het
eron
ucle
ar(h
n)R
NA
inth
ehy
poth
alam
icpa
rave
ntric
ular
nucl
eus(
PVH
)and
supr
aopt
icnu
cleu
s(SO
N).
Brai
nse
ctio
ns(3
0mm
thic
k)w
ere
obta
ined
from
rats
that
wer
eki
lled
eith
er30
min
befo
re(p
repr
andi
al)
or30
min
afte
r(p
ostp
rand
ial)
the
onse
tof
am
eal
cons
istin
gof
10m
lof
swee
tene
dco
nden
sed
milk
.The
optic
alde
nsity
ofth
eAV
Phn
RN
Ahy
brid
izat
ion
signa
lwas
signi
fican
tlyhi
gher
afte
rth
anbe
fore
the
mea
l.*d
i�er
entf
rom
prep
rand
ial,
P<0.
01.M
agni
ficat
ion,
×50.
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340
334 Sensory activation of the vasopressinergic and oxytocinergic neurons
F. 5. The dark-field photomicrographs illustrate the e�ect of real, sham, and gastric feeding of sweetened condensed milk on the pre- and postprandiallevels of c-fos mRNA in the hypothalamic paraventricular nucleus [(PVH), (Fig. 5)] and the supraoptic nucleus [(SON), (Fig. 5)]. The preprandialexpression was measured 30 min prior to the onset of the milk meal. The postprandial expression consists of the mRNA expression measured 30 minafter the onset of the meal. In the PVH the hybridization signal seen after real and sham feeding was particularly intense in the magnocellular divisionof the nucleus. Magnification, ×50.
load that was probably less than that required to activate the reported expression of Fos in the magnocellular division of PVNfollowing the ingestion of hypertonic saline. However, in contrasthypothalamic magnocellular neurosecretory system.
The finding that an isotonic solution of sucrose did not induce with ingestion of hypertonic saline, ingestion of hypertonic sucrosesolutions is not likely to alter plasma osmolality, membrane-any significant stimulation of the PVH and SON, while the
hypertonic solution did, raises the posssibility that sweetened permeable solutes being known not to influence plasma osmolal-ity. The possibility that ingestion of hypertonic sucrose does notcondensed milk can lead to the osmotic stimulation of the AVP
and OT systems. The activation of the hypothalamic magnocellu- stimulate the magnocellular neurosecretory system through anincrease in plasma osmolality is further supported by the observa-lar neurosecretory system observed in rats fed with the hypertonic
solution of sucrose or sweetened condensed milk was largely tion that sham feeding, which does not significantly rise plasmaglucose above basal levels, is capable of markedly stimulating thesimilar to that seen in rats subjected to salt loading, which was
also reported to induce the osmotic stimulation of the magnocellu- expression of c-fos, whereas gastric feeding, which significantlyelevates plasma glucose, has no significant e�ect on the expressionlar division of the PVH and SON (4). The suggestion that the
OT and AVP cells are stimulated by hypertonicity is supported of c-fos in either the PVH or SON.By showing that hypertonic solutions of sucrose can triggerby numerous studies including that of Olson et al. (32), who
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340

Sensory activation of the vasopressinergic and oxytocinergic neurons 335
activation of the magnocellular neurosecretory cells of the hypo- through oropharyngeal osmoreceptors appears as conceivable asis the early suppression of vasopressin secretion in dehydratedthalamus before reaching the stomach, the present results further
support the existence of putative osmoreceptors in the oropharyn- subjects induced by the passage of water through the oropharynx(40, 41, 43). The anticipatory (prior to a change in plasmageal portion of the gastrointestinal tract (39–42). Appelgren et al.
(41) reported that the rapid fall in circulating AVP in dehydrated osmolality) regulation of fluid balance can be seen as an importantresponse in dehydrated subjects, in particular, as the intake ofdogs after the ingestion of water and solutions of NaCl was
followed by a rise in the AVP plasma levels that was largely water must preferably cease prior to normalisation of plasmaosmolality to avoid liquid ingestion in excess of the physiologicaldetermined by the osmolality of the solution ingested and inde-
pendent of the plasma osmolality (41). In addition, Akaishi & needs. Although the present study was not specifically aimed atdelineating the pathways activated by the ingestion of sweetenedHomma (42), by measuring the discharge rate of hypothalamic
AVP cells in rats after ligation of the oesophagus, reported an condensed milk that could convey impulses from oropharyngealsignals to the magnocellular neurosecretory neurons it is worthyoropharyngeal stimulation of AVP neurons that was proportional
to the molality of the saline solution applied in the oropharynx. of mention that the NST, which receives a�erents from theglossopharyngeal and the vagus nerves can be strongly activatedAlthough the presence of oropharyngeal osmoreceptors with
physiological functions remains speculative, it represents a plaus- by both real and sham feeding preparations (44, 45) (Naımi andRichard, unpublished results). The A2 noradrenergic cell groupsible issue considering the immense capacity of oropharynx in
generating metabolic cues following ingestion. The stimulation of of the NST as well as those from the ventrolateral medulla (A1noradrenergic cell group) have been reported to send both directthe magnocellular neurosecretory neurons by hypertonicity
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340
336 Sensory activation of the vasopressinergic and oxytocinergic neurons
expression of c-fos in the PVH and SON. Altogether the presentresults indicate that magnocellular neurosecretory neurons aresensitive to oropharyngeal stimuli and further support the viewof the existence of oropharyngeal osmoreceptors.
Materials and methods
Male Wistar rats initially weighing 250–300 g were purchased from theCanadian Breeding Laboratories (St-Constant, Canada). All rats werecared for and handled in conformance with the Canadian Guide for theCare and Use of Laboratory Animals. The animals were housed individu-ally in wire-bottom cages suspended above absorbent paper. They weresubjected to a 12512 h light-dark cycle and kept under an ambienttemperature of 23±1 °C. Approximately 1 week following their arrival,the rats were trained to eat their daily food ration as three 1-h meals andfamiliarized with the ingestion of sweetened condensed milk or the sucrosesolutions. The meals were provided between 07.00 and 08.00 (morningmeal), 15.00 and 16.00 (afternoon meal ), and 23.00 and 00.00 (eveningmeal). The morning and evening meals consisted of stock diet (AgwayProlab, Rat/Mouse/Hamster 1000 Formula). The afternoon meal con-sisted of either a solution of sweetened condensed milk (Eagle Brand)diluted 151 with water, an isotonic (0.4 M ) solution of sucrose, or ahypertonic (0.8 M) solution of sucrose. The composition in grams per100 grams of undiluted condensed sweetened milk was as follows: protein,8.1; fat, 8.2; lactose, 11.2; sucrose/glucose, 55.4. In rats, sweetenedcondensed milk is referred to as a palatable food item that has been usedin numerous paradigms (17, 47). In our laboratory, this food item is usedlargely because it induces a strong thermogenic response. The number ofmeals per day as well as the amount of food provided during each mealwere determined to insure that the body weight growth of the meal-fedrats would be comparable to that of ad libitum-fed animals. Rats wereallowed about ten days to accustom to the experimental feeding regimen.On the day of the experiment, rats were o�ered milk or sucrose solutionsunder one of the following conditions: real feeding, sham feeding, andgastric feeding (Figure 1 ). Under each conditions, rats had free access totap water. Rats were killed at various times following the onset of theafternoon meal, which consisted, for this occasion, of 10 ml of eithersweetened condensed milk or one of the sucrose solutions. All ratsingested the 10 ml within 10 min.
Implantation of chronic gastric fistula
The technique for implanting the gastric cannula has been detailedelsewhere (48). Prior to the surgery, rats were food-deprived for 24 h in
70
0
Time (min)
50
60
40
30
20
10
–30 30 60 360
*** *
SON c-fos mRNA (OD)
60
0
Time (min)
50
40
30
20
10
–30 30 60 360
**
*
*
PVH c-fos mRNA (OD)
order to ensure complete gastric emptying. Rats were anesthetized withF. 6. The e�ect of real, sham, and gastric feeding of sweetened con-an intraperitoneal injection of 0.4 ml/100 g of body weight of a mixturedensed milk on the pre- and postprandial levels of c-fos mRNA in thecontaining 20 mg/ml of ketamine and 2.5 mg/ml of xylazine. A smallhypothalamic paraventricular nucleus (PVH) and supraoptic nucleusincision was made in the abdomen starting 4–5 mm lateral to the mid-(SON) is illustrated. The c-fos mRNA levels were quantitated in termsline and 3–4 mm caudal to the sternum. The stomach was exposedof optical density units (OD). The preprandial levels were determinedthrough the incision and gently pulled out of the abdominal cavity while30 min prior to the onset of the milk meal. The postprandial levels werebeing kept moist with isotonic saline. A purse-string suture, approximatelydetermined 30, 60 and 360 min after the beginning of the meal. *di�erent2 cm in diameter, was secured to the anterior part of the stomach withoutfrom pre-prandial (−30) within the same level of ‘feeding’, P<0.001.completely perforating its wall. An incision was made and the stainlessANOVA: Feeding (F): P=0.001; Time (T ) P=0.001; F×T P=0.001.steel cannula was implanted in the anterior wall. The cannula was
% Real feeding; & Sham feeding; b gastric feeding.positioned with the flange inside the stomach and a polypropylene meshwas used on the outside surface of the stomach. This knitted polypropylenemesh, by promoting growth of scar tissue, allowed a firm and leak proofand indirect projections to the magnocellular neurosecretoryseal around the cannula. The stomach was then returned to its normal
systems (14, 46). position. Incisions of both abdominal muscles and skin were closed withIn conclusion, the natural ingestion of sweetened condensed interrupted sutures. The gastric cannula was plugged with a screw which
allowed for easy real feeding (gastric cannula closed ), sham feedingmilk enhanced expression of c-fos within the AVP- and(gastric cannula opened) and gastric feeding (via the gastric cannula).OT-containing cells of the PVH and SON. Besides, ingestion of
sweetened condensed milk led to an increase in transcription of Brain removal and sectioningthe AVP gene in the magnocellular neurosecretory section of the Rats were anesthetized with 1.5 ml of the ketamine-xylazine mixturehypothalamus. The expression of c-fos in the PVH and SON was described above. Without delay, they were perfused intracardially with
30 ml of ice-cold isotonic saline followed by 200 ml of a paraformaldehydealso enhanced by ingestion of an hypertonic solution of sucrose(4%) solution. The brains were removed at the end of perfusion and keptand was not only seen in real-fed rats but also in sham-fedin paraformaldehyde for 2 days prior to their transfer to a solutionanimals. In contrast, neither ingestion of an isotonic solution of containing paraformaldehyde and sucrose (10%) the night before being
sucrose nor gastric feeding of both sweetened condensed milk sliced with a sliding microtome (Histoslide 2000, Reichert-Jung). Thirty-mm-thick sections were collected and stored at −30 °C in a cold sterileand hypertonic sucrose were capable of inducing a significant
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340
Sensory activation of the vasopressinergic and oxytocinergic neurons 337
80
0
Hypertonic sucrose
60
40
20
–30 min
SON c-fos mRNA (OD)
80
0
60
40
20
60 min
* *
Isotonic sucrose
–30 min 60 min
80
0
Hypertonic sucrose
60
40
20
–30 min
PVH c-fos mRNA (OD)
80
0
60
40
20
60 min
* *
Isotonic sucrose
–30 min 60 min
F. 7. The e�ect of real, sham, and gastric feeding of isotonic and hypertonic solutions of sucrose on the pre-and postprandial levels of c-fos mRNAin the hypothalamic paraventricular nucleus (PVH) and supraoptic nucleus (SON) is illustrated. The c-fos mRNA levels were quantitated in terms ofoptical density units (OD). The preprandial levels were determined 30 min prior to the onset of the milk meal. The postprandial levels was determined60 min after the beginning of the meal. *di�erent from pre-prandial (−30) within the same level of ‘feeding’, P<0.001. ANOVA: Feeding (F):P=0.001; Time (T ) P=0.001; Solutions (S), P=0.001; F×T×S P=0.001. % Real feeding; & Sham feeding; b gastric feeding.
cryoprotecting solution containing sodium phosphate bu�er (50 mM ), alcohol. After a 2 h-period of vacuum drying, the slides were exposed onx-ray film for periods varying between 12 to 48 h, depending on theethylene glycol (30%), and glycerol (20%).nature of the probe used. Afterwards, slides were defatted in xylene and
In situ hybridization histochemistry dipped in NTB2 nuclear emulsion (Eastman Kodak, Rochester, NY,USA). Depending on the probe used, the slides were exposed from 12 hIn situ hybridization histochemistry was used to localize c-fos, OT and
AVP mRNAs, as well as AVP hnRNA on brain sections taken through to 7 days (AVP mRNA, 12 h; AVP hnRNA, 6 days; c-fos mRNA,7 days), before being developed in D19 developer ( Kodak) for 3.5 minthe hypothalamus. The protocol used was largely adapted from the
technique described by Simmons et al. (49). Briefly, 1 out of every 6th at 14–15 °C and fixed in rapid fixer ( Kodak) for 5 min. Finally, tissueswere rinsed in running distilled water for 1 to 2 h, counterstained withbrain sections was mounted onto poly-L-lysine coated slides and allowed
to desiccate under vacuum overnight. The sections were then successively thionin (0.25%), dehydrated through graded concentrations of alcohol,cleared in xylene, and coverslipped with DPX.fixed for 20 min in paraformaldehyde (4%), digested 30 min at 37 °C with
proteinase K (10 mg/ml in 100 mM Tris-HCl containing 50 mM EDTA,Antisense 35S-labeled cRNA and hnRNA probespH 8.0), acetylated with acetic anhydride (0.25% in 0.1 M trietholamine,
pH 8.0) and dehydrated through graded concentrations (50, 70, 95, and The antisense 35S-labeled cRNA probe for c-fos was generated from a2-kilobase (kb) EcoRI fragment of a full-length rat c-fos cDNA (courtesy100%) of alcohol. After vacuum drying for at least 2 hours, 90 ml of the
hybridization mixture, which contains an antisense 35S-labeled cRNA from Dr I. Verma, The Salk Institute, La Jolla, CA, USA), subclonedinto a pBluescript SK-1 (Stratagene, La Jolla, CA, USA) plasmid thatprobe (107cpm/ml) was spotted on each slide. The slides were sealed
under a coverslip and incubated overnight at 60 °C in a slide warmer. was linearized with SmaI. The antisense 35S-labeled cRNA probe for AVPwas generated from a 230 base-pair (bp) PstI/DraI fragment of a pRV8The next day, the coverslips were removed and the slides rinsed 4 times
with SCC (0.6 M NaCl, 60 mM trisodium citrate bu�er, pH 7.0), digested plasmid, which contains the AVP specific 3∞ end of the BB cDNA (courtesyof Dr D. Richter, Clinical Neurobiological University, Hamburgh,30 min at 37 °C with RNAse-A (20 mg/ml in H2O), washed in descending
concentrations of SSC (2×, 10 min; 1×, 5 min; 0.5×, 5 min; 0.1×, Germany) subcloned in a pSP2 plasmid that was linearized with HindIII.The antisense 35S-labeled cRNA probe for the AVP intron was prepared30 min at 60 °C) and dehydrated through graded concentrations of
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340

338 Sensory activation of the vasopressinergic and oxytocinergic neurons
PV
H c
-fo
s m
RN
A
Iso
ton
ic
Co
ntr
ol (p
rep
ran
dia
l)
Hyp
ert
on
icIs
oto
nic
Real fe
ed
ing
Hyp
ert
on
ic
Iso
ton
ic
Sh
am
feed
ing
Hyp
ert
on
icIs
oto
nic
Gastr
ic f
eed
ing
Hyp
ert
on
ic
F
.8.
heda
rk-fi
eld
phot
omic
rogr
aphs
illus
trat
eth
ee�
ecto
frea
l,sh
am,a
ndga
stric
feed
ing
ofiso
toni
can
dhy
pert
onic
solu
tions
ofsu
cros
eon
the
pre-
and
post
pran
dial
leve
lsof
c-fo
sm
RN
Ain
the
hypo
thal
amic
para
vent
ricul
arnu
cleu
s(P
VH
).Th
epr
epra
ndia
lexp
ress
ion
was
mea
sure
d30
min
prio
rto
the
onse
toft
hem
ilkm
eal.
The
post
pran
dial
expr
essio
nco
nsist
sof
the
mR
NA
expr
essio
nm
easu
red
60m
inaf
ter
the
onse
tof
the
mea
l.Th
ehy
brid
izat
ion
signa
lsee
naf
ter
real
and
sham
feed
ing
ofth
ehy
pert
onic
solu
tion
was
inte
nse
inth
em
agno
cellu
lar
divi
sion
ofth
ePV
Hco
mpa
red
toth
issig
nalm
easu
red
afte
rre
alan
dsh
amfe
edin
gof
the
isoto
nic
solu
tion.
Mag
nific
atio
n,×5
0.
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340

Sensory activation of the vasopressinergic and oxytocinergic neurons 339
from a 700 bp PvuII fragment of AVP intron I (courtesy of Dr black and white video camera (Sony, XC-77) coupled to a Macintoshcomputer (Power PC 7100/66) using Image software (version 1.55 non-T. J. Sherman, University of Pittsburgh, Pittsburgh) subcloned in a
pGEM3 plasmid that was linearized with HindIII. The antisense 35S- FPU, Wayne Rasband, NIH, Bethesda, MD, USA). The optical density(OD) for the hybridization signal was measured under dark-field illumina-labeled cRNA probe for OT was generated from a 190 bp of a
EcoRI/HindIII fragment of a pRO4 plasmid, which contains exon C and tion at a magnification of 10×. Brain sections for all groups were matchedfor rostrocaudal levels as closely as possible. When no hybridizationpart of the intron 2 of the rat OT gene (courtesy of Dr D. Richter,
Clinical Neurobiological University, Hamburgh, Germany) subcloned in signal was visible under dark-field illumination, the brain structures ofinterest were digitized under bright field illumination and then subjecteda pSP10 plasmid that was linearized with EcoRI. Radioactive antisense
riboprobes were synthesized by incubation of 250 ng of the linearized to densitometric analysis under dark field illumination. The OD for eachspecific region was corrected for the average background signal whichplasmids in 6 mM MgCl2, 36 mM Tris (pH 7.5), 2 mM spermidine,
10 mM dithiothreitol, 0.2 mM of ATP/GTP/CTP and [a-35S]UTP, 40U was determined by sampling cells located outside of the areas of interest.RNAse inhibitor (Promega, Madison, WI, USA), and 20 U of either T7
Blood glucose determination(AVP hnRNA and c-fos mRNA) or SP6 (AVP mRNA and OT mRNA)MNA polymerase. Unincorporated nucleotides were removed using the Pre- and postprandial concentrations of glucose were determined for theammonium acetate method; 100 ml of DNAse solution (1 ml DNAse, 5 ml three feeding conditions of this study. This measure was principally usedof 5 mg/ml tRNA, 94 ml of 10 mM Tris containing 10 mM MgCl2) was to establish that the sham-fed milk did not massively reach the gutadded, and 10 min later an extraction was accomplished using a phenol- whereas the gastric load e�ciently did. Fifty ml of blood were collectedchloroform solution. The cRNA was precipitated for 20 min on dry ice 30 min prior to or 30, 60 and 360 min following the onset of the milkwith 80 ml of 5 M ammonium acetate and 500 ml of ethanol (100%). The meal. Blood samples were obtained from the tail vein. Plasma glucosepellet was washed with 500 ml of ethanol, dried, and resuspended in 100 ml was determined by the glucose oxydase method using a glucose analyzerof 10 mM Tris/1 mM EDTA (pH 8.0). A concentration of 107 cpm was (Beckman, Palo Alto, CA, USA). The rats used for glucose measurementsmixed into 1 ml of the hybridization solution, which consisted of 500 ml were not employed in the histochemistry experiments.formamide, 60 ml 5 M NaCl, 10 ml 1 M Tris (pH 8.0), 2 ml 0.5 M EDTA
Statistics(pH 8.0), 20 ml 50× Denhart’s solution, 200 ml 50% dextran sulfate, 50 ml10 mg/ml tRNA, 10 ml 1 M DTT. This solution was mixed and heated The analysis of variance (ANOVA) was used as statistical analysis. Afor 5 min at 65 °C before being spotted on slides. Radioactive sense one-way ANOVA was used to analyse the e�ects of sweetened condensed(control) cRNA copies were also prepared to verify the specificity of each milk on the pre- and postprandial levels of AVP mRNA and AVPprobe. Hybridization with these probes did not reveal any positive signal hnRNA. The e�ects of sweetened condensed milk on the plasma glucosein the brain of rats killed pre- or postprandially. as well as on the optical density (OD) of the c-fos mRNA hybridization
signal were examined using a 3×4 factorial design with feeding [realCombination of immunocytochemistry with in situ hybridization feeding, sham feeding and gastric feeding] and time [−30 min, 30 min,
60 min, 360 min] as factors. The e�ects of two sucrose solutions on theImmunocytochemical detection of Fos was combined with in situ hybrid-OD of the c-fos hybridization signal were analysed using a 3×2×2ization histochemical detection of AVP mRNA or OT mRNA to determinefactorial design with feeding [real feeding, sham feeding, and gastricwhether AVP and OT cells were activated following the ingestion offeeding], time [−30 min, 60 min] and solution [isotonic, hypertonic] assweetened condensed milk. Brain sections were first processed for immuno-factors. In the case of significant interactions, a posteriori comparisonschemical detection for Fos using a conventional avidin-biotin-were performed using the Bonferoni/Dunn multiple-comparison-immunoperoxidase method. Briefly, brain slices were washed in sterileprocedure.0.05 M potassium phosphate-bu�ered saline (KPBS) that was treated
with DEPC water. They were then incubated for 24 h at 4 °C with a Fosantibody (rabbit polyclonal IgG, Oncogene Science, NY, USA). The Fosantibody was used at a 151,500 dilution in KPBS (50 mM ) with normal Acknowledgementsgoat serum (1%), triton X-100 (0.4%) and bovine serum albumin (1%).Twenty-four h following incubation at 4 °C with the primary antibody, This work was supported by the Medical Research and Natural Sciences
and Engineering Research Councils of Canada. Dr Serge Rivest holds athe brain slices were rinsed in sterile KPBS and incubated with a mixtureof KPBS, triton-X, heparin, and biotinylated goat antirabbit IgG (151,500 Scholarship from the Medical Research Council of Canada.dilution; Vector Laboratories, CA, USA) for 90 min. Sections were thenrinsed with KPBS and incubated at room temperature for 60 min with Accepted 5 November 1996an avidin-biotin-peroxidase complex (Vectastain ABC Elite Kit, VectorLaboratories, CA, USA), followed by a second incubation with a mixtureof KPBS, triton-X-100, heparin and biotinylated goat antirabbit IgG with Referencesthe ABC Elite solution. After several rinses in sterile KPBS, the brainslices were allowed to react in a mixture containing sterile KPBS, the 1 Richard D, Naımi N, Rivier C, Rivest S. E�ects of meal intake onchromagen 3,3∞-diaminobenzidine tetrahydrochloride (DAB, 0.05%), and the expressions of the c-fos, NGFI-B and CRF genes in the rat brain.1% hydrogen peroxide. Thereafter, tissues were rinsed in sterile KPBS, Soc Neurosci Abs 1993; 19: 761.mounted onto poly-L-lysine-coated slides, desiccated overnight under 2 Kononen J, Honkaniemi J, Alho H, Koistinaho J, Iadarola M,vacuum, fixed in paraformaldehyde (4%) for 30 min, and digested for Peltohuikko M. Fos-like immunoreactivity in the rat hypothalamic-25 min at 37 °C with proteinase K (10 mg/ml in 100 mM tris HCl, pH 8.0, pituitary axis after immobilization stress. Endocrinology 1992; 130:and 50 mM EDTA). Prehybridization, hybridization, and post- 3041–3047.hybridization steps were performed as described above except for the 3 Rivest S, Rivier C. The role of corticotropin-releasing factor anddehydration step, which was shortened to avoid decolorization of Fos interleukin-1 in the regulation of neurons controlling reproductivecells. After vacuum drying for 2 h, sections were exposed onto x-ray film, functions. Endocrine Rev 1995; 16: 177–199.defatted in xylene, and dipped in the NTB2 nuclear emulsion. Slides were 4 Chan RKW, Brown ER, Ericsson A, Kovacs KJ, Sawchenko PE. Aexposed for 6 to 8 days, developed in D19 developer for 3.5 min at 15 °C, comparison of two immediate-early genes, c-fos and NGFI- B, asand fixed in rapid fixer for 5 min. Thereafter, tissues were rinsed in markers for functional activation in stress-related neuroendocrinerunning distilled water for 1 to 2 h, rapidly dehydrated through graded circuitry. J Neurosci 1993; 13: 5126–5138.concentrations of alcohol, cleared in xylene, and coverslipped with DPX. 5 Ding JM, Carver WC, Terracio L, Buggy J. Proto-oncogene c-fosThe detection of Fos protein immunoreactivity was revealed as dark and the regulation of vasopressin gene expression during dehydration.brown stains whereas AVP and OT mRNAs were revealed as silver grains. Mol Brain Res 1994; 21: 247–255.
6 Lafarga M, Martinezguijarro FJ, Berciano MT, Blascoibanez JM,Quantitative analysis Andres MA, Mellstrom B, Lopezgarcia C, Naranjo JR. Nuclear fos
domains in transcriptionally activated supraoptic nucleus neurons.The c-fos, AVP, and OT mRNAs as well as the AVP hnRNA wererevealed on NTB2 dipped nuclear emulsion slides and were analyzed and Neuroscience 1993; 57: 353–364.
7 Rivest S, Torres G, Rivier C. Di�erential e�ects of central andquantified under a light microscope (Olympus, BX50) equipped with a
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340

340 Sensory activation of the vasopressinergic and oxytocinergic neurons
peripheral injection of interleukin-1b on brain c-fos expression and 28 Li BH, Rowland NE. Cholecystokinin-induced and dexfenfluramine-induced anorexia compared using devazepide and c-fos expression inneuroendocrine functions. Brain Res 1992; 587: 13–23.
8 Rivest S, Laflamme N, Nappi RE. Immune challenge and immobiliza- the rat brain. Regul Pept 1994; 50: 223–233.29 Bonaz B, Degiorgio R, Tache Y. Peripheral bombesin induces c-fostion stress induce transcription of the gene encoding the CRF
receptor in selective nuclei of the rat hypothalamus. J Neurosci 1995; protein in the rat brain. Brain Res 1993; 600: 353–357.30 Ritter S, Dinh TT. 2-Mercaptoacetate and 2-deoxy-D-glucose induce15: 2680–2695.
9 Richard D, Rivest S, Rivier C. The 5-hydroxytryptamine agonist Fos-Like immunoreactivity in rat brain. Brain Res 1994; 641:111–120.fenfluramine increases Fos-like immunoreactivity in the brain. Brain
Res 1992; 594: 131–137. 31 Pezzone MA, Lee WS, Ho�man GE, Rabin BS. Induction of c-fosimmunoreactivity in the rat forebrain by conditioned and10 Laflamme N, Bovetto S, Richard D, Rivest S. Stimulation of the
serotoninergic system causes transcriptional activation of CRF and unconditioned aversive stimuli. Brain Res 1992; 597: 41–50.32 Olson BR, Freilino M, Ho�man GE, Stricker EM, Sved AF, Verbalisits receptor selectively within the paraventricular nucleus of the rat
hypothalamus. Br J Pharmacol 1995; in press. JG. c-fos expression in rat brain and brainstem nuclei in response totreatments that alter food intake and gastric motility. Mol Cell11 Doucet JP, Squinto SP, Bazan NG. Fos-Jun and the primary genomic
response in the nervous system. In: Bazan NG, ed. Molecular Neurosci 1993; 4: 93–106.33 Swank MW, Bernstein IL. c-fos induction in response to a condi-Neurobiology, Totowa, N.J.: Humana Press, 1990: 27–55.
12 Morgan JI, Curran T. Stimulus-transcription coupling in the nervous tioned stimulus after single trial taste aversion learning. Brain Res1994; 636: 202–208.system: involvement of the inducible proto-oncogenes fos and jun.
Annu Rev Neurosci 1991; 14: 421–451. 34 Torres G, Rivier C. Induction of c-fos in rat brain by acute cocaineand fenfluramine exposure: a comparison study. Brain Res 1994;13 Ho�man GE, Le WW, Abbud R, Lee WS, Smith MS. Use of fos-
related antigens (FRAs) as markers of neuronal activity: FRA 647: 1–9.35 Li BH, Rowland NE. Dexfenfluramine induces fos-like immunoreac-changes in dopamine neurons during proestrus, pregnancy and
lactation. Brain Res 1994; 654: 207–215. tivity in discrete brain regions in rats. Brain Res Bull 1993; 31: 43–48.36 Fraser KA, Raizada E, Davison JS. Oral-pharyngeal-esophageal and14 Armstrong WE. Hypothalamic supraoptic and paraventricular nuclei.
In: Paxinos G, ed. The Rat Nervous System, San Diego: Academic gastric cues contribute to meal-induced c-fos expression. AmerJ Physiol-Regul Integr C 1995; 37: R223–R230.Press, 1995: 377–390.
15 Weingarten HP, Watson SD. Sham feeding as a procedure for 37 Kovacs KJ, Sawchenko PE. Sequence of stress-induced alterationsin indices of synaptic and transcriptional activation in parvocellularassessing the influence of diet palatability on food intake. Physiol
Behav 1982; 28: 401–407. neurosecretory neurons. J Neurosci 1996; 16: 262–273.38 Mezey E, Kiss JZ. Coexpression of vasopressin and oxytocin in16 Conover KL, Shizgal P. Competition and summation between
rewarding e�ects of sucrose and lateral hypothalamic stimulation in hypothalamic supraoptic neurons of lactating rats. Endocrinology1991; 129: 1814–1820.the rat. Behav Neurosci 1994; 108: 537–548.
17 Davis JD, Smith GP, Kung TM. Abdominal vagotomy alters the 39 Nicolaidis S. Early systemic responses to orogastric stimulation inthe regulation of food and water balance: functional and electro-structure of the ingestive behavior of rats ingesting liquid diets. Behav
Neurosci 1994; 108: 767–779. physiological data. Ann N-Y Acad Sci 1969; 157: 1176–1203.40 Thrasher TN, Keil LC, Ramsay DJ. Drinking, oropharyngeal signals,18 Nissenbaum JW, Sclafani A. Sham-feeding response of rats to
plolycose and sucrose. Neurosci Biobehav Rev 1987; 11: 215–222. and inhibition of vasopressin secretion in dogs. Am J Physiol 1986;253: R509–R515.19 Renaud LP, Bourque CW. Neurophysiology and neuropharmacology
of hypothalamic magnocellular neurons secreting vasopressin and 41 Appelgren BH, Thrasher TN, Keil LC, Ramsay DJ. Mechanism ofdrinking-induced inhibition of vasopressin secretion in dehydratedoxytocin. Prog Neurobiol 1990; 36: 131–169.
20 Share L. Control of vasopressin release: an old but continuing story. dogs. Am J Physiol 1991; 261: R1226–R1233.42 Akaishi T, Homma S. Properties of oropharyngeal/laryngeal a�erentsNews Physiol Sci 1996; 11: 7–13.
21 Ho�man GE, Smith MS, Verbalis JG. c-Fos and related immediate regulating vasopressin release. Ann N-Y Acad Sci 1993; 689: 455–457.43 Salata RA, Verbalis JG, Robinson AG. Cold water stimulation ofearly gene products as markers of activity in neuroendocrine systems.
Front Neuroendocrinol 1993; 14: 173–213. oropharyngeal receptors in man inhibits release of vasopressin. J ClinEndocrinol Metab 1987; 65: 561–567.22 Imaki T, Shibasaki T, Hotta M, Demura H. Intracerebroventricular
administration of corticotropin-releasing factor induces c-fos messen- 44 Powley TL, Berthoud HR, Fox EA, Laughton W. The dorsal vagalcomplex forms a sensory-motor lattice: the circuitry of gastrointesti-ger RNA expression in brain regions related to stress responses—
comparison with pattern of c-fos messenger RNA induction after nal reflexes. In: Ritter S, Ritter RC, Barnes CD, eds. Neuroanatomyand Physiology of Abdominal Vagal A�erents, Boca Raton: CRC,stress. Brain Res 1993; 616: 114–125.
23 Rivest S, Rivier C. Interleukin-1b inhibits the endogenous expression 1992: 55–79.45 Ritter RC, Brenner L, Yox DP. Participation of vagal sensoryof the early gene c-fos located within the nucleus of LH-RH neurons
and interferes with hypothalamic LH-RH release during proestrus in neurons in putative satiety signals from the upper gastrointestinaltract. In: Ritter S, Ritter RC, Barnes CD, eds. Neuroanatomy andthe rat. Brain Res 1993; 613: 132–142.
24 Rivest S, Rivier C. Stress and interleukin-1b induced activation of c- Physiology of Abdominal Vagal A�erents, Boca Raton: CRC, 1992:222–247.fos, NGFI-B and CRF gene expression the hypothalamic PVN –
comparison between Sprague-Dawley, Fisher-344 and Lewis rats. 46 Bourque CW, Oliet SHR, Richard D. Osmoreceptors, osmoreception,and osmoregulation. Front Neuroendocrinol 1994; 15: 231–274.J Neuroendocrinol 194; 6: 101–117.
25 Olson BR, Ho�man GE, Sved AF, Stricker EM, Verbalis JG. 47 Sclafani A, Gorman AN. E�ects of age, sex, and prior body weighton the development of dietary obesity in adult rats. Physiol BehavCholecystokinin induces c-fos expression in hypothalamic oxytoci-
nergic neurons projecting to the dorsal vagal complex. Brain Res 1977; 18: 1021–1026.48 Waynforth HB. Experimental and Surgical Technique in the Rat.1992; 569: 238–248.
26 Verbalis JG, Stricker EM, Robinson AG, Ho�man GE. London: Academic Press, 1980.49 Simmons DM, Arriza JL, Swanson LW. A complete protocol for inCholecystokinin activates c-fos expression in hypothalamic oxytocin
and corticotropin-releasing hormone neurons. J Neuroendocrinol situ hybridization of messenger RNAs in brain and other tissues withradiolabeled single-stranded RNA probes. J Histotechnol 1989; 12:1991; 3: 205–213.
27 Chen DY, Deutsch JA, Gonzalez MF, Gu Y. The induction and 169–181.suppression of c-fos expression in the rat brain by cholecystokininand its antagonist L364,718. Neurosci Lett 1993; 149: 91–94.
© 1997 Blackwell Science Ltd, Journal of Neuroendocrinology, 9, 329–340