neural stem cell transplant-induced effect on neurogenesis and
TRANSCRIPT

Research ArticleNeural Stem Cell Transplant-Induced Effect on Neurogenesisand Cognition in Alzheimer Tg2576 Mice Is Inhibited byConcomitant Treatment with Amyloid-Lowering or Cholinergic1205727 Nicotinic Receptor Drugs
Anna M Lilja1 Linn Malmsten1 Jennie Roumljdner1 Larysa Voytenko1 Alexei Verkhratsky2
Sven Ove Oumlgren23 Agneta Nordberg14 and Amelia Marutle1
1Department of Neurobiology Care Sciences and Society Center for Alzheimer Research Division of Translational AlzheimerNeurobiology Karolinska Institutet Karolinska University Hospital Huddinge 141 57 Huddinge Sweden2Faculty of Life Sciences The University of Manchester Manchester M13 9PT UK3Department of Neuroscience Karolinska Institutet 171 77 Stockholm Sweden4Department of Geriatric Medicine Karolinska University Hospital Huddinge 141 57 Huddinge Sweden
Correspondence should be addressed to Anna M Lilja annaliljakise Linn Malmsten linnmalmstenkiseand Amelia Marutle ameliamarutlekise
Received 15 March 2015 Revised 9 June 2015 Accepted 25 June 2015
Academic Editor Lucas Pozzo-Miller
Copyright copy 2015 Anna M Lilja et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Stimulating regeneration in the brain has the potential to rescue neuronal networks and counteract progressive pathological changesinAlzheimerrsquos disease (AD)This study investigatedwhether drugswith differentmechanisms of action could enhance neurogenesisand improve cognition in mice receiving human neural stem cell (hNSC) transplants Six- to nine-month-old AD Tg2576 micewere treated for five weeks with the amyloid-modulatory and neurotrophic drug (+)-phenserine or with the partial 1205727 nicotinicreceptor (nAChR) agonist JN403 combined with bilateral intrahippocampal hNSC transplantation We observed improved spatialmemory in hNSC-transplanted non-drug-treated Tg2576 mice but not in those receiving drugs and this was accompanied by anincreased number of Doublecortin- (DCX-) positive cells in the dentate gyrus a surrogate marker for newly generated neuronsTreatment with (+)-phenserine did however improve graft survival in the hippocampus An accumulation of 1205727 nAChR-expressingastrocyteswas observed around the injection site suggesting their involvement in repair and scarring processes Interestingly JN403treatment decreased the number of 1205727 nAChR-expressing astrocytes correlating with a reduction in the number of DCX-positivecells in the dentate gyrusWe conclude that transplanting hNSCs enhances endogenous neurogenesis and prevents further cognitivedeterioration in Tg2576 mice while simultaneous treatments with (+)-phenserine or JN403 result in countertherapeutic effects
1 Introduction
In recent years expectations have been raised that stem cellreplacement therapy could be used as a therapeutic approachto compensate for lost or damaged neuronal cells and torestore functionality in brain areas affected by chronic neuro-degenerative diseases such as Alzheimer disease (AD) Theaccumulation of amyloid-120573 (A120573) and the formation of neu-rofibrillary tangles during AD pathogenesis are accompaniedby cholinergic deficits involving a significant loss of nico-tinic acetylcholine receptors (nAChRs) neuroinflammatory
changes and reduced growth factor production in the brainThese neurodegenerative processes alter the brain microen-vironment resulting in neuronal dysfunction and ultimatelyin cognitive decline [1]
The hippocampus is affected early in the disease and thisregion is one of only two known neurogenic niches of theadult brain [2] where new neurons are generated from neuralstemprogenitor cells [3] We have earlier proposed that thepathophysiological environment in the AD brain could haveadverse effects on neurogenesis [3 4] and it is suggestedthat the memory deficits observed in AD may be linked to
Hindawi Publishing CorporationNeural PlasticityVolume 2015 Article ID 370432 13 pageshttpdxdoiorg1011552015370432
2 Neural Plasticity
alterations in hippocampal neurogenesis as reviewed in [5ndash7] How neurogenesis is linked to cognitive function andwhether stimulating regenerative mechanisms in the braincould restore or prevent further deterioration of cognition inthe disease are actively being investigated So far observationsfrom experimental studies in animal models of AD have pro-vided supportive evidence that transplanted murine neuralprecursor or stem cells demonstrate neurotrophic neuropro-tective and immunomodulatory potencies these cells sur-vive and migrate as normal differentiate into neurons astro-cytes and oligodendrocytes and ameliorate learning andmemory deficits in the recipient [8]
Moreover we and others have earlier shown that pharma-cological interventions can mobilize endogenous stempro-genitor cells in the hippocampus Treatment with the drugsgalantamine or memantine that are used to treat AD in clini-cal practice as well as the candidate drugs (minus)- and (+)-phen-serine increased neurogenesis in vitro in primary murinecortical cultures and in vivo in animal models of the disease[9ndash11]
In the current study we applied a combinational thera-peutical approach to investigate whether drugs with specificand distinct mechanisms of action on A120573 production and1205727 nAChRs respectively could enhance neurogenesis andimprove memory in human neural stem cell- (hNSC-) trans-planted transgenic mice overexpressing the human amyloidprecursor protein with the double Swedishmutation Tg2576This animalmodel is phenotypically similar to the prodromalstage of AD and at 6 to 9 months of age Tg2576 micedemonstrate elevated levels of A120573 oligomers in the brain butno A120573 plaques increased astrogliosis as well as impairmentof synaptic function and learning and memory deficits [12ndash15] The study mice underwent intrahippocampal transplan-tation with 5 weeksrsquo concurrent treatment with either (+)-phenserine the partial 1205727 nAChR agonist JN403 or vehicleWe hypothesized that drug treatment could potentiate theeffects of hNSC transplantation and the rationale for usingthese two drugs was based on previous reports of both amy-loid-lowering and neurotrophic effects for (+)-phenserine[16 17] and improvement in learning and memory followingthe stimulation of 1205727 nAChRs with selective cholinergic ago-nists including JN403 [18 19] Spatial learning and memorywere tested both before and after intervention with trans-plantation and drug treatment The subsequent effects onhNSC transplant survival endogenous neurogenesis and theinvolvement of a subpopulation of astroglia the 1205727 nAChR-expressing astrocytes in the neurogenic niche of the dentategyrus (DG) were determined
2 Materials and Methods
21 HumanNeural StemCell Culture hNSCswere purchasedfrom Lonza (Walkersville MD USA) and cultured in neuralprogenitor maintenance medium according to instructionsfrom the manufacturer These cells have previously beencharacterized in vitro [20] For cellular differentiation in vitroneurospheres were plated onto laminin-coated cover slipsand cultured in DMEM F12 and neurobasal medium (1 1)
supplemented with B27 (1 50) and 1 penicillinstrepto-mycin (Invitrogen La Jolla CAUSA)The cells were exposedweekly for a total of 28 days to extract from human ADautopsy frontal cortex (AD) containing 043 pM total A120573 orfrom a human healthy control (HC) diluted equivalently asthe extract with 043 pM A120573 or to vehicle In parallel exper-iments cells were pretreated with 10minus7M JN403 or 10minus7M(+)-phenserine 24 hours prior to the addition of AD extractFor in vitro differentiation experiments two independentexperiments with four replicate samples for each treatmentwere used for analysis
22 Tg2576 Mouse Cortical Primary Neurons Primary neu-rons were isolated from the cerebral cortex of Tg2576 mouseembryos at embryonic day E17 The tissues were triturated tosingle cell suspension and plated onto poly-D-lysine-coatedcover slips and cultured in neurobasal medium withoutglutamine andwere supplementedwith B27 (1 50) 100xGlu-tamax (1 400) and 1 penicillinstreptomycin (InvitrogenLa Jolla CA USA) The cells were exposed weekly to either10minus7M JN403 10minus7M (+)-phenserine or vehicle followingplating at the density 2 times 105 cellscm2 After 21 days in cul-ture the cells were washed with PBS and used for immuno-cytochemistry Three biological replicates of each treatmentwere used for analysis
23 Animals Tg2576mice expressing the APP Swedishmut-ation (APPSWE2576Kha) aged 5ndash7 months (3 females (F)3 males (M)) and 6ndash9 months (17 F 13M) were obtained bybackcrossing B6SJL (F1) females (Taconic) at the KarolinskaInstitutet animal care facility as previously described [15]Age-matched wild type littermates (3 F 3M) were used ascontrol animals in the pilot study assessing memory inTg2576mice with theMorris water maze (MWM) navigationtask All mice were housed in enriched cages with a 12-hourlight-dark cycle and access to food and water ad libitum Allexperimental procedures complied with the guidelines andregulations of the Swedish National Board for LaboratoryAnimals and the Regional Ethics and Animal Research Com-mittee at Karolinska Institutet approved the study protocol
24 Drug Treatment Tg2576 mice aged 6ndash9 months weredivided into three treatment groups and given intraperitonealinjections of 03mgkg JN403 (119899 = 5) or 25mgkg (+)-phenserine (119899 = 7) solubilized in physiological saline solutionor vehicle (physiological saline solution) (119899 = 18) once dailyfor 1 week To monitor potential adverse drug reactionsJN403 was initially administered at dosages of 001mgkg(days 1-2) and 01mgkg (days 3-4) before reaching the fulldose from day 5
25 hNSC Transplantation Cells were triturated countedand diluted with cell medium (vehicle) Tg2576 mice wereanesthetized using a constant flow of 4 isoflurane and keptwarm under a heating lamp throughout the transplantationprocedure The head of each mouse was fixed using ear andtooth bars before a skin incision into the skull bone wasmadeusing a 07mm steel burr (Meisinger Neuss Germany) with
Neural Plasticity 3
the following coordinates relative to the bregma AP minus206ML plusmn175 and DV minus175mm Using a 26-gauge microsyringe(ILS Microsyringes Stutzerbach Germany) approximately25000 cells per hemisphere were injected in a total volumeof 1 120583L into the hippocampal DG of each Tg2576 mouseand the needle was thereafter kept in the injection site forapproximately 30 seconds to allow the volume to diffuseThe wound was then sutured with metal clips Lidocaine wasused for local anesthesia during the procedure and 032Msucrose solution was injected subcutaneously after surgeryThe animalsweremonitored daily for bodyweight and properhealing of the incision site after surgery No immunosuppre-ssants were used and no symptoms indicative of a reactionto the transplant were observed within the time frame inves-tigated
Drug treatment was interrupted on the day of transplan-tation and the mice were allowed a 3-day recovery periodbefore drug treatment was continued for an additional 4weeks Demographic information of all the mice included inthe transplantation is provided in Supplementary Table 1 (seeSupplementary Material available online at httpdxdoiorg1011552015370432) and the study outline is illustrated inFigure 2(a)
26 Morris Water Maze First a pilot study was conductedin which 5ndash7-month-old Tg2576 mice and their age-matchedwild type littermates underwent testing for spatial learningand memory in the MWM task The escape latency (time toreach the hidden platform) for Tg2576mice and age-matchedwild type littermates was compared during acquisition trialsof 60 secondsrsquo duration which were performed 6 times a dayat 10-minute intervals for 4 days Both the latency to the cen-ter of the platform (time to reach the former platform loca-tion) and the number of platform crossings were measured
Later in the follow-up acquisition study the Tg2576mice(vehicle- or hNSC-transplanted and saline- or drug-treated)now aged 6ndash9 months underwent 60-second trials 4 times aday at 15-second intervals for 5 days To assess retention ofspatial memory 24 hours after the last acquisition trial theplatform was removed and the mice were tracked in the poolfor 60 seconds in a probe trial Both the latency to the centerof the platform and the time spent in the target quadrant weremeasured The mice were randomly placed in one of fourfixed positions around thewall of a 1mdiameter round swim-ming pool During the acquisition period the mice learnedthe location of a hidden platform (9 times 9 cm) which wasplaced 1 cm under the water surface and 25 cm from the wallThe location of the platform was learned through visual cues(wall posters) on the walls around the pool The water tem-perature was maintained at 21∘C plusmn 2∘C and the water wasmade opaque (white) to contrast the dark color of the micefrom the black pool The mice were habituated a day beforethe first acquisition day by letting them swim in the pool for60 seconds without the platform The platform location wasfixed during the whole acquisition phase Mice that did notfind the platform within 60 seconds were aided to stay onthe platform for 10 seconds to assist the learning processThebehavior of the mice in the MWM task was recorded by anautomated video-tracking system (Ethovision) To evaluate
the differences in memory retention between the groups inthe probe trial the baseline values for latency were subtractedfrom the follow-up latency values (Δ latency)
Two mice (one in the hNSC-transplanted and saline-treated group as well as one in the hNSC-transplanted and(+)-phenserine treated group) could not be detected by theautomated video-tracking system because of their color andthese mice were therefore omitted from the MWM tests
27 Tissue Processing for Biochemical Analyses Within 12ndash16hours of completing the final behavioral assessments the ani-mals were anesthetized with a 1 1 ketaminexylazine mixture(100mgkg ketamine and 20mgkg xylazine) and sacrificedby transcardial perfusion with PBS buffer (pH 74) Thebrain from each animal was separated into hemispheres theleft hemisphere was snap frozen on dry-ice and the righthemisphere was postfixed in buffered 4 paraformaldehydeand then transferred to a 032M sucrose cryoprotectant solu-tion for 24 h at 4∘C The brain hemispheres were stored atminus80∘C until further analysisThe left-brain hemispheres weresubsequently used in the biochemical assays and the righthemispheres were cryostat sectioned and used in immuno-histochemistry studies
28 Immunohistochemistry For staining of differentiatedhNSCs in culture cells were incubated with rabbit anti-human glial fibrillary acidic protein (GFAP Dako GlostrupDenmark) mouse anti-120573III-tubulin (Sigma St Louis MOUSA) or mouse polyclonal anti-microtubule-associated pro-tein 2 (MAP2 Sigma St Louis MO USA) All primary anti-bodies were diluted 1 250 Thereafter cells were incubatedwith secondary Alexa Fluor conjugated antibodies (1 500donkey anti-mouse Alexa Fluor 546 or donkey anti-rabbitAlexa Fluor 488 Molecular Probes Eugene OR USA) Cellsimmunopositive for DAPI and GFAP or 120573III-tubulin or thenumber of dendritic branches perDAPI andMAP2 immuno-positive cell was counted in three random fields per cover slip(six fields per treatment under a Nikon E800 microscope)
In order to obtain comparable results between the differ-ent animals for each individual animal three coronal sectionsfrom the same hippocampal regions were selected for immu-nostaining Newly generated neurons in the hippocampuswere labeled with an antibody for themicrotubule-associatedprotein Doublecortin (DCX) (goat anti-DCX 1 200 SantaCruz Heidelberg Germany) according to the method des-cribed previously by Lilja et al [10] DCX immunoreactivitywas analyzed in three serial sections for each animal AllDCX+ cells in the DG were quantified blindly in which thetotal number of DCX+ cells the proportion of DCX+ cellswith dendrites and the number of branch points on theDCX+ cells with more pronounced dendritic branching werecounted
To investigate graft survival and the fate of transplantedhNSCs in the DG fluorescent immunohistochemical anal-ysis was carried out on serial sections from each animalusing primary antibodies polyclonal rabbit antiglial fibril-lary acidic protein (GFAP) (1 500 Dako Glostrup Den-mark) polyclonal rabbit anti-microtubule-associated protein2 (MAP2) (1 500 Millipore Temecula CA USA) and
4 Neural Plasticity
monoclonal mouse anti-human nuclei (hNuclei) (1 200Millipore Temecula CA USA) Secondary antibodies wereconjugated with Alexa Fluor (AF AF 488 donkey anti-rabbitand AF 546 donkey anti-mouse 1 500 Molecular ProbesEugene OR USA) In control experiments the primaryantibodywas omitted and a blocking bufferwas substituted toverify the specificity of the secondary antibodies The speci-ficity of hNuclei antibody staining was also assessed in micethat had not received any hNSCs implants Immunoreactivecells within the DG (including the subgranular zone andgranular and polymorph layers) were counted in three serialsections from each animal
The number of GFAP1205727 nAChR immunopositive cellswas evaluated on three serial coronal sections per animalafter consecutive incubations with primary antibodies (1 500rabbit anti-GFAP Dako 1 100 rat anti-1205727 nAChR Abcam)and secondary antibodies (1 200 goat anti-rabbit IgG-Fabhorseradish peroxidase- (HRP-) conjugated goat anti-ratIgG-Fab HRP-conjugated Invitrogen La Jolla CA USA)Immunoreactivity was visualized using peroxidase substratekits (DAB and SG Vector resp) The results were quanti-fied at 20x magnification in a prespecified area (325 120583m times435 120583m= 141375 120583m2) using software ImageJ (NIH BethesdaMD USA)
Nonfluorescent staining was evaluated under a light mic-roscope (Leica Wetzlar Germany) with an attached ProgResSpeedXT core5 video camera and an image analysis system(ProgRes Capture Pro288) Fluorescent staining was evalu-ated under a Nikon E800 microscope at 20x magnificationusing Adobe Photoshop CS5 software (San Jose CA USA)
29 A12057340 and A12057342 ELISA Frontal cortices and hippocampiwere extracted from the left hemisphere immersed in PBSbuffer (pH 74) and homogenized Samples were diluted 1 25in BSAT-DPBS (Dulbeccorsquos PBS with 5 BSA 003 Tween-20 and 1x protease inhibitor cocktail) A120573
40
and A12057342
levelswere measured using a sandwich ELISA assay (InvitrogenCamarillo CA USA) according to the manufacturersrsquo ins-tructions and the absorbance was read at 450 nm (TecanInfinite M1000)
210 Statistical Analysis GraphPad Prism 50 or 6 (GraphPadSoftware Inc) was used for all statistical analyses Datawere analyzed using the Mann-Whitney test for comparisonbetween two groups and theKruskal-Wallis one-wayANOVAtest by rank followed by Dunnrsquos post hoc test for compar-ison of gt2 groups for both in vitro and in vivo analysesRepeated measures ANOVA was applied for comparison ofwild type and Tg2576 mice in the acquisition tests in Morriswater maze Spearmanrsquos rank correlation was used in thecorrelation analysis which was visualized graphically usingsimple regression analysisThe data are presented as means plusmnstandard error of the mean (SEM) 119901 values lt 005 wereconsidered to be significant
3 Results
31 JN403 Exerts Neuroprotective Effects on hNSC-DerivedNeurons in Culture Neurotrophic actions of the 1205727 nAChR
partial agonist JN403 and the A120573 modulatory drug (+)-phenserine were assessed by examining the survival dif-ferentiation and maturation of progenitor cells in vitroJN403 protected hNSC-derived120573III-tubulinmarked neuronsagainst brain extract from a patient with AD containingvarious A120573 species (Figure S1A) On the other hand JN403 or(+)-phenserine did not significantly alter the number andorpattern of MAP2-positive Tg2576 cortical neurons in culture(Figure S1B)
32 Tg2576 Mice Demonstrate Impaired Spatial Memory Nav-igation Hippocampal-dependent spatial learning andmem-ory in Tg2576 mice (119899 = 6) and age-matched nontransgeniclittermates (wild type 119899 = 6) were tested in the MWM nav-igation task During the four days of acquisition training ofTg2576 mice resulted in an expected learning curve whereastraining of the wild type mice resulted in a very shallowlearning curve where escape latencies were similar over time(Figure 1(a)) The latency to the former platform location inthe probe trial 24 hours after the last day of training (day4) was also significantly longer for Tg2576 mice than forwild type mice (471 s compared to 119 s 119901 lt 005 Mann-Whitney test Figure 1(b))Wild typemice crossed the formerlocation of the platform more often than the Tg2576 mice(119901 lt 005 Mann-Whitney test Figure 1(c)) In contrast theswimming velocity and the total distance the mice swamduring the acquisition period (Figures S2A and S2B) did notdiffer significantly between the groups Differences in escapelatency between female andmalemice were also assessed andno significant differences appeared
33 Intrahippocampal hNSC Transplantation Prevents Mem-ory Deterioration in Tg2576 Mice To ensure that there wereno differences in learning and memory among the Tg2576mice that could be attributed to the age range (6ndash9 months)the mice were subjected to a baseline MWM test beforethe onset of treatment (an overview of the study design isprovided in Figure 1(d)) No significant differences in testresults were observed among the different treatment groups(at baseline or at follow-up) during the 5 days of acquisitiontraining In general improvements in escape latencies wereminor and resulted in similar shallow learning curves as thoseobserved during training of wild type mice in the pilot studyIn the 24-hour probe trial therewere no differences in latencybaseline values between groups (Figure 1(e) 119901 = 026 Dunnrsquostest) The cohort of hNSC-transplanted Tg2576 mice had amemory performance comparable to their baseline valuesat the MWM follow-up test whereas the other treatmentsgroups deteriorated compared to baseline (Figure 1(e)) Theprogression of pathology as well as extensive neurosurgerycould have influenced the performance at follow-up ThehNSC-transplanted mice found the former platform locationsignificantly faster than SHAM-transplanted mice in theMWM follow-up test (Δ latency 110 s and 399 s resp119901 lt 005 Dunnrsquos test Figure 1(f)) The hNSC-transplantedTg2576 mice treated with either JN403 or (+)-phenserineshowed no memory improvements compared to SHAM-transplanted mice (Figure 1(f))
Neural Plasticity 5
60
40
20
0
1 2 3 4
Esca
pe la
tenc
y (s
)
Time (days)
WtTg2576
(a)
Wt Tg2576
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
lowast
(b)
Wt Tg2576
3
2
1
0
Num
ber o
f pla
tform
cros
ses
lowast
(c)
Drug treatment
Was
hout
per
iod
Mor
ris w
ater
maz
e
Mor
ris w
ater
maz
e
Tran
spla
ntat
ion
Treatment groups
minusfo
llow
-up
minusba
selin
e
1W 1W 1W4W 1W
Vehicle injected + saline (SHAM + SAL) n = 9
hNSC + saline (hNSC + SAL) n = 9
hNSC + JN403 (hNSC + JN) n = 5
hNSC + (+)-phenserine (hNSC + PHEN) n = 7
(d)
80
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
BaselineFollow-up
SHAM + SAL + SALhNSC + JNhNSC + PHENhNSC
(e)
SHAM + SAL
60
40
20
0
Δ la
tenc
y (s
)
lowast
+ SALhNSC + JNhNSC + PHENhNSC
(f)
Figure 1 Continued
6 Neural Plasticity
SHAM + SAL
50
40
30
20
10
0
Tim
e in
targ
et q
uadr
ant (
)
+ SALhNSC + JNhNSC + PHENhNSC
(g)
Figure 1 Intrahippocampal hNSCs transplantation affects spatial navigation memory in Tg2576 mice Learning and memory were assessedin 5- to 7-month-old wild type (wt 119899 = 6) and Tg2576 (APPswe 119899 = 6) mice in the pilot Morris water maze task (a) Escape latency during 4days of acquisition training (b) Latency to the former location of the platform or (c) the number of platform crosses in a probe test 24 hourslater lowast119901 lt 005 compared to wild typeThe data are expressed as means plusmn SEM (d)The experimental design of the study with 6- to 9-month-old Tg2576 mice (119899 = 30) comprised (i) Morris water maze tests with 5 days of acquisition followed 24 hours later by a probe memory test(ii) daily intraperitoneal injections of 03mgkg JN403 or 25mgkg (+)-phenserine for 5 weeks in total (iii) bilateral hippocampal injectionsof sim25000 hNSCshemisphere after 1 week of drug treatment (iv) repetition of the Morris water maze tasks 5 weeks after transplantationand (v) sacrifice by transcardial perfusion with PBS (e) Time taken to find the former platform location in the baseline and follow-up test(f) Differences in time taken to find the former platform location (Δ latency follow-up probe test values minus baseline probe values) (g)Percent time spent in the target quadrant where the platform was located lowast119901 lt 005 compared to SHAM + SAL The data are expressed asmeans plusmn SEM
The standardmeasurement ofmemory is the percent timespent in the target quadrant compared with the other quad-rants rather than the latency to reach the former location ofthe platform The different groups of mice spent an equallyshort time in the target quadrant thus showing a low persis-tence which is not unusual for mice (Figure 1(g)) Swimmingvelocity or the total distance swum did not differ amongTg2576 mice in the various groups (Figures S2C and S2D)
34 hNSCTransplantation Increases EndogenousNeurogenesisin the Dentate Gyrus The early neuronal marker DCX istransiently expressed during a period that extends fromapro-liferative progenitor cell stage to a postmitotic phase at whichDCX-positive cells differentiate into mature neurons withincreasing dendritic arborization in the DG as illustrated inFigure 2(a) We thus characterized neural stemprogenitorcells DCX+ cells as an index of neurogenesisThese cells mayderive from the resident pool of progenitor cells in the hippo-campal DG or from the exogenous source of transplantedhNSCs DCX+ cells were detected exclusively in the subgran-ular layer while cells positive for the marker human nuclei(hNuclei+ cells) as described in the following section wereobservedmainly in the polymorph layer of the DGThis indi-cates that the DCX+ cells detected were mainly derived fromthe endogenous stem cell pool Immunohistochemical analy-sis demonstrated that hNSC transplantation in Tg2576 miceaugmented endogenous neurogenesis as increases in boththe total number of DCX+ cells (56 higher) and the numberof maturing DCX+ cells (82 higher) were observed in theDG compared to SHAM transplantation (both 119901 lt 005Dunnrsquos test Figures 2(b) and 2(c))
Treatment with JN403 and (+)-phenserine howevercounteracted the beneficial effects of hNSC transplantationon endogenous neurogenesis Moreover JN403 reduced thetotal number of DCX+ cells (43 lower 119901 lt 001 Dunnrsquostest) as well as the number of DCX+ cells with dendrites (55lower 119901 lt 005 Dunnrsquos test) compared to vehicle-treatedhNSC-transplanted mice Treatment of hNSC-transplantedmice with (+)-phenserine did not significantly alter thenumber of DCX+ cells (Figures 2(b) and 2(c))There were nosignificant differences in the number of dendritic branchesper DCX+ cell among the different groups (Figure 2(d))
The number of DCX+ cells with dendrites inversely cor-related with measures of the Δ latency obtained in the probetrial (119901 lt 001 119903 = minus061 Spearmanrsquos rank Figure 3(e))indicating that the increasedmaturation of newborn neuronsin the DG following hNSC transplantation was related toan improvement in hippocampus-dependent memory Rep-resentative images of DCX+ cells in the DG of hNSC- andSHAM-transplanted Tg2576 mice are shown in Figures 2(f)and 2(g)
35 (+)-Phenserine Treatment Increases the Survival of hNSC-Derived Neurons in the Hippocampus of Tg2576 Mice Chro-nic treatment with (+)-phenserine increased the survival ofengrafted hNSCs in the DG of Tg2576 mice as demonstratedby human nuclei antibody labeling The number of hNuclei+cells was larger in Tg2576 mice treated with (+)-phenserinethan in those receiving saline (43 increase 119901 lt 005 Dunnrsquostest) while there were no differences in numbers of hNuclei+cells between JN403- and saline-treated mice (Figure 3(a))Costaining hNuclei+ cells with MAP2 or GFAP markers for
Neural Plasticity 7
Proliferation Maturation Maturation with dendritic arborization
(a)
50
40
30
20
10
0SHAM + SAL
lowast
Num
ber o
f DCX
+ce
lls in
den
tate
gyr
us
+ SALhNSC + JNhNSC + PHENhNSC
(b)
SHAM + SAL
lowast
10
15
20
25
0
5
Num
ber o
f DCX
+ce
lls w
ith d
endr
ites
+ SALhNSC + JNhNSC + PHENhNSC
(c)
SHAM + SAL0
5
1
2
3
4
Den
driti
c bra
nche
sD
CX+
cells
with
den
drite
s
+ SALhNSC + JNhNSC + PHENhNSC
(d)
0
20
0 10 20 30 40
40
60
80
minus20
Prob
eΔla
tenc
y
r = minus061lowastlowastp lt 001
Number of DCX+ cells with dendrites
(e)
SHAM + SAL + SALhNSC
(f)
SHAM + SAL + SALhNSC
(g)
Figure 2 hNSC transplantation increases the number and the maturation of new neurons in the dentate gyrus (a) Doublecortin- (DCX-)labeled cells in the dentate gyrus (DG) of Tg2576micewere characterized according to their numbers and the level ofmaturation and dendriticarborization (b)The total number of DCX+ cells (c) the number of DCX+ cells possessing dendrites and (d) the average number of dendriticbranches on DCX+ cells in the DG inmice receiving SHAM (vehicle only) transplantation and saline (SHAM+ SAL) or mice receiving hNSCtransplantation and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (e) Correlation between the number ofDCX+ cells with dendrites and the difference in time taken to find the position of the platform (Δ latency follow-up probe test values minusbaseline acquisition values) in theMorris watermaze task in groups of Tg2576mice treatedwith hNSC+ SAL hNSC+ JN and hNSC+PHEN(119903 = minus061 lowastlowast119901 lt 001 119899 = 18) Representative images of immunostaining with DCX+ cells in the DG of TG2576 mice at (f) 10x and (g) 20xmagnification lowast119901 lt 005 compared to SHAM + SAL 119901 lt 005 and
119901 lt 001 compared to hNSC + SALThe data are expressed as means plusmnSEM
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

2 Neural Plasticity
alterations in hippocampal neurogenesis as reviewed in [5ndash7] How neurogenesis is linked to cognitive function andwhether stimulating regenerative mechanisms in the braincould restore or prevent further deterioration of cognition inthe disease are actively being investigated So far observationsfrom experimental studies in animal models of AD have pro-vided supportive evidence that transplanted murine neuralprecursor or stem cells demonstrate neurotrophic neuropro-tective and immunomodulatory potencies these cells sur-vive and migrate as normal differentiate into neurons astro-cytes and oligodendrocytes and ameliorate learning andmemory deficits in the recipient [8]
Moreover we and others have earlier shown that pharma-cological interventions can mobilize endogenous stempro-genitor cells in the hippocampus Treatment with the drugsgalantamine or memantine that are used to treat AD in clini-cal practice as well as the candidate drugs (minus)- and (+)-phen-serine increased neurogenesis in vitro in primary murinecortical cultures and in vivo in animal models of the disease[9ndash11]
In the current study we applied a combinational thera-peutical approach to investigate whether drugs with specificand distinct mechanisms of action on A120573 production and1205727 nAChRs respectively could enhance neurogenesis andimprove memory in human neural stem cell- (hNSC-) trans-planted transgenic mice overexpressing the human amyloidprecursor protein with the double Swedishmutation Tg2576This animalmodel is phenotypically similar to the prodromalstage of AD and at 6 to 9 months of age Tg2576 micedemonstrate elevated levels of A120573 oligomers in the brain butno A120573 plaques increased astrogliosis as well as impairmentof synaptic function and learning and memory deficits [12ndash15] The study mice underwent intrahippocampal transplan-tation with 5 weeksrsquo concurrent treatment with either (+)-phenserine the partial 1205727 nAChR agonist JN403 or vehicleWe hypothesized that drug treatment could potentiate theeffects of hNSC transplantation and the rationale for usingthese two drugs was based on previous reports of both amy-loid-lowering and neurotrophic effects for (+)-phenserine[16 17] and improvement in learning and memory followingthe stimulation of 1205727 nAChRs with selective cholinergic ago-nists including JN403 [18 19] Spatial learning and memorywere tested both before and after intervention with trans-plantation and drug treatment The subsequent effects onhNSC transplant survival endogenous neurogenesis and theinvolvement of a subpopulation of astroglia the 1205727 nAChR-expressing astrocytes in the neurogenic niche of the dentategyrus (DG) were determined
2 Materials and Methods
21 HumanNeural StemCell Culture hNSCswere purchasedfrom Lonza (Walkersville MD USA) and cultured in neuralprogenitor maintenance medium according to instructionsfrom the manufacturer These cells have previously beencharacterized in vitro [20] For cellular differentiation in vitroneurospheres were plated onto laminin-coated cover slipsand cultured in DMEM F12 and neurobasal medium (1 1)
supplemented with B27 (1 50) and 1 penicillinstrepto-mycin (Invitrogen La Jolla CAUSA)The cells were exposedweekly for a total of 28 days to extract from human ADautopsy frontal cortex (AD) containing 043 pM total A120573 orfrom a human healthy control (HC) diluted equivalently asthe extract with 043 pM A120573 or to vehicle In parallel exper-iments cells were pretreated with 10minus7M JN403 or 10minus7M(+)-phenserine 24 hours prior to the addition of AD extractFor in vitro differentiation experiments two independentexperiments with four replicate samples for each treatmentwere used for analysis
22 Tg2576 Mouse Cortical Primary Neurons Primary neu-rons were isolated from the cerebral cortex of Tg2576 mouseembryos at embryonic day E17 The tissues were triturated tosingle cell suspension and plated onto poly-D-lysine-coatedcover slips and cultured in neurobasal medium withoutglutamine andwere supplementedwith B27 (1 50) 100xGlu-tamax (1 400) and 1 penicillinstreptomycin (InvitrogenLa Jolla CA USA) The cells were exposed weekly to either10minus7M JN403 10minus7M (+)-phenserine or vehicle followingplating at the density 2 times 105 cellscm2 After 21 days in cul-ture the cells were washed with PBS and used for immuno-cytochemistry Three biological replicates of each treatmentwere used for analysis
23 Animals Tg2576mice expressing the APP Swedishmut-ation (APPSWE2576Kha) aged 5ndash7 months (3 females (F)3 males (M)) and 6ndash9 months (17 F 13M) were obtained bybackcrossing B6SJL (F1) females (Taconic) at the KarolinskaInstitutet animal care facility as previously described [15]Age-matched wild type littermates (3 F 3M) were used ascontrol animals in the pilot study assessing memory inTg2576mice with theMorris water maze (MWM) navigationtask All mice were housed in enriched cages with a 12-hourlight-dark cycle and access to food and water ad libitum Allexperimental procedures complied with the guidelines andregulations of the Swedish National Board for LaboratoryAnimals and the Regional Ethics and Animal Research Com-mittee at Karolinska Institutet approved the study protocol
24 Drug Treatment Tg2576 mice aged 6ndash9 months weredivided into three treatment groups and given intraperitonealinjections of 03mgkg JN403 (119899 = 5) or 25mgkg (+)-phenserine (119899 = 7) solubilized in physiological saline solutionor vehicle (physiological saline solution) (119899 = 18) once dailyfor 1 week To monitor potential adverse drug reactionsJN403 was initially administered at dosages of 001mgkg(days 1-2) and 01mgkg (days 3-4) before reaching the fulldose from day 5
25 hNSC Transplantation Cells were triturated countedand diluted with cell medium (vehicle) Tg2576 mice wereanesthetized using a constant flow of 4 isoflurane and keptwarm under a heating lamp throughout the transplantationprocedure The head of each mouse was fixed using ear andtooth bars before a skin incision into the skull bone wasmadeusing a 07mm steel burr (Meisinger Neuss Germany) with
Neural Plasticity 3
the following coordinates relative to the bregma AP minus206ML plusmn175 and DV minus175mm Using a 26-gauge microsyringe(ILS Microsyringes Stutzerbach Germany) approximately25000 cells per hemisphere were injected in a total volumeof 1 120583L into the hippocampal DG of each Tg2576 mouseand the needle was thereafter kept in the injection site forapproximately 30 seconds to allow the volume to diffuseThe wound was then sutured with metal clips Lidocaine wasused for local anesthesia during the procedure and 032Msucrose solution was injected subcutaneously after surgeryThe animalsweremonitored daily for bodyweight and properhealing of the incision site after surgery No immunosuppre-ssants were used and no symptoms indicative of a reactionto the transplant were observed within the time frame inves-tigated
Drug treatment was interrupted on the day of transplan-tation and the mice were allowed a 3-day recovery periodbefore drug treatment was continued for an additional 4weeks Demographic information of all the mice included inthe transplantation is provided in Supplementary Table 1 (seeSupplementary Material available online at httpdxdoiorg1011552015370432) and the study outline is illustrated inFigure 2(a)
26 Morris Water Maze First a pilot study was conductedin which 5ndash7-month-old Tg2576 mice and their age-matchedwild type littermates underwent testing for spatial learningand memory in the MWM task The escape latency (time toreach the hidden platform) for Tg2576mice and age-matchedwild type littermates was compared during acquisition trialsof 60 secondsrsquo duration which were performed 6 times a dayat 10-minute intervals for 4 days Both the latency to the cen-ter of the platform (time to reach the former platform loca-tion) and the number of platform crossings were measured
Later in the follow-up acquisition study the Tg2576mice(vehicle- or hNSC-transplanted and saline- or drug-treated)now aged 6ndash9 months underwent 60-second trials 4 times aday at 15-second intervals for 5 days To assess retention ofspatial memory 24 hours after the last acquisition trial theplatform was removed and the mice were tracked in the poolfor 60 seconds in a probe trial Both the latency to the centerof the platform and the time spent in the target quadrant weremeasured The mice were randomly placed in one of fourfixed positions around thewall of a 1mdiameter round swim-ming pool During the acquisition period the mice learnedthe location of a hidden platform (9 times 9 cm) which wasplaced 1 cm under the water surface and 25 cm from the wallThe location of the platform was learned through visual cues(wall posters) on the walls around the pool The water tem-perature was maintained at 21∘C plusmn 2∘C and the water wasmade opaque (white) to contrast the dark color of the micefrom the black pool The mice were habituated a day beforethe first acquisition day by letting them swim in the pool for60 seconds without the platform The platform location wasfixed during the whole acquisition phase Mice that did notfind the platform within 60 seconds were aided to stay onthe platform for 10 seconds to assist the learning processThebehavior of the mice in the MWM task was recorded by anautomated video-tracking system (Ethovision) To evaluate
the differences in memory retention between the groups inthe probe trial the baseline values for latency were subtractedfrom the follow-up latency values (Δ latency)
Two mice (one in the hNSC-transplanted and saline-treated group as well as one in the hNSC-transplanted and(+)-phenserine treated group) could not be detected by theautomated video-tracking system because of their color andthese mice were therefore omitted from the MWM tests
27 Tissue Processing for Biochemical Analyses Within 12ndash16hours of completing the final behavioral assessments the ani-mals were anesthetized with a 1 1 ketaminexylazine mixture(100mgkg ketamine and 20mgkg xylazine) and sacrificedby transcardial perfusion with PBS buffer (pH 74) Thebrain from each animal was separated into hemispheres theleft hemisphere was snap frozen on dry-ice and the righthemisphere was postfixed in buffered 4 paraformaldehydeand then transferred to a 032M sucrose cryoprotectant solu-tion for 24 h at 4∘C The brain hemispheres were stored atminus80∘C until further analysisThe left-brain hemispheres weresubsequently used in the biochemical assays and the righthemispheres were cryostat sectioned and used in immuno-histochemistry studies
28 Immunohistochemistry For staining of differentiatedhNSCs in culture cells were incubated with rabbit anti-human glial fibrillary acidic protein (GFAP Dako GlostrupDenmark) mouse anti-120573III-tubulin (Sigma St Louis MOUSA) or mouse polyclonal anti-microtubule-associated pro-tein 2 (MAP2 Sigma St Louis MO USA) All primary anti-bodies were diluted 1 250 Thereafter cells were incubatedwith secondary Alexa Fluor conjugated antibodies (1 500donkey anti-mouse Alexa Fluor 546 or donkey anti-rabbitAlexa Fluor 488 Molecular Probes Eugene OR USA) Cellsimmunopositive for DAPI and GFAP or 120573III-tubulin or thenumber of dendritic branches perDAPI andMAP2 immuno-positive cell was counted in three random fields per cover slip(six fields per treatment under a Nikon E800 microscope)
In order to obtain comparable results between the differ-ent animals for each individual animal three coronal sectionsfrom the same hippocampal regions were selected for immu-nostaining Newly generated neurons in the hippocampuswere labeled with an antibody for themicrotubule-associatedprotein Doublecortin (DCX) (goat anti-DCX 1 200 SantaCruz Heidelberg Germany) according to the method des-cribed previously by Lilja et al [10] DCX immunoreactivitywas analyzed in three serial sections for each animal AllDCX+ cells in the DG were quantified blindly in which thetotal number of DCX+ cells the proportion of DCX+ cellswith dendrites and the number of branch points on theDCX+ cells with more pronounced dendritic branching werecounted
To investigate graft survival and the fate of transplantedhNSCs in the DG fluorescent immunohistochemical anal-ysis was carried out on serial sections from each animalusing primary antibodies polyclonal rabbit antiglial fibril-lary acidic protein (GFAP) (1 500 Dako Glostrup Den-mark) polyclonal rabbit anti-microtubule-associated protein2 (MAP2) (1 500 Millipore Temecula CA USA) and
4 Neural Plasticity
monoclonal mouse anti-human nuclei (hNuclei) (1 200Millipore Temecula CA USA) Secondary antibodies wereconjugated with Alexa Fluor (AF AF 488 donkey anti-rabbitand AF 546 donkey anti-mouse 1 500 Molecular ProbesEugene OR USA) In control experiments the primaryantibodywas omitted and a blocking bufferwas substituted toverify the specificity of the secondary antibodies The speci-ficity of hNuclei antibody staining was also assessed in micethat had not received any hNSCs implants Immunoreactivecells within the DG (including the subgranular zone andgranular and polymorph layers) were counted in three serialsections from each animal
The number of GFAP1205727 nAChR immunopositive cellswas evaluated on three serial coronal sections per animalafter consecutive incubations with primary antibodies (1 500rabbit anti-GFAP Dako 1 100 rat anti-1205727 nAChR Abcam)and secondary antibodies (1 200 goat anti-rabbit IgG-Fabhorseradish peroxidase- (HRP-) conjugated goat anti-ratIgG-Fab HRP-conjugated Invitrogen La Jolla CA USA)Immunoreactivity was visualized using peroxidase substratekits (DAB and SG Vector resp) The results were quanti-fied at 20x magnification in a prespecified area (325 120583m times435 120583m= 141375 120583m2) using software ImageJ (NIH BethesdaMD USA)
Nonfluorescent staining was evaluated under a light mic-roscope (Leica Wetzlar Germany) with an attached ProgResSpeedXT core5 video camera and an image analysis system(ProgRes Capture Pro288) Fluorescent staining was evalu-ated under a Nikon E800 microscope at 20x magnificationusing Adobe Photoshop CS5 software (San Jose CA USA)
29 A12057340 and A12057342 ELISA Frontal cortices and hippocampiwere extracted from the left hemisphere immersed in PBSbuffer (pH 74) and homogenized Samples were diluted 1 25in BSAT-DPBS (Dulbeccorsquos PBS with 5 BSA 003 Tween-20 and 1x protease inhibitor cocktail) A120573
40
and A12057342
levelswere measured using a sandwich ELISA assay (InvitrogenCamarillo CA USA) according to the manufacturersrsquo ins-tructions and the absorbance was read at 450 nm (TecanInfinite M1000)
210 Statistical Analysis GraphPad Prism 50 or 6 (GraphPadSoftware Inc) was used for all statistical analyses Datawere analyzed using the Mann-Whitney test for comparisonbetween two groups and theKruskal-Wallis one-wayANOVAtest by rank followed by Dunnrsquos post hoc test for compar-ison of gt2 groups for both in vitro and in vivo analysesRepeated measures ANOVA was applied for comparison ofwild type and Tg2576 mice in the acquisition tests in Morriswater maze Spearmanrsquos rank correlation was used in thecorrelation analysis which was visualized graphically usingsimple regression analysisThe data are presented as means plusmnstandard error of the mean (SEM) 119901 values lt 005 wereconsidered to be significant
3 Results
31 JN403 Exerts Neuroprotective Effects on hNSC-DerivedNeurons in Culture Neurotrophic actions of the 1205727 nAChR
partial agonist JN403 and the A120573 modulatory drug (+)-phenserine were assessed by examining the survival dif-ferentiation and maturation of progenitor cells in vitroJN403 protected hNSC-derived120573III-tubulinmarked neuronsagainst brain extract from a patient with AD containingvarious A120573 species (Figure S1A) On the other hand JN403 or(+)-phenserine did not significantly alter the number andorpattern of MAP2-positive Tg2576 cortical neurons in culture(Figure S1B)
32 Tg2576 Mice Demonstrate Impaired Spatial Memory Nav-igation Hippocampal-dependent spatial learning andmem-ory in Tg2576 mice (119899 = 6) and age-matched nontransgeniclittermates (wild type 119899 = 6) were tested in the MWM nav-igation task During the four days of acquisition training ofTg2576 mice resulted in an expected learning curve whereastraining of the wild type mice resulted in a very shallowlearning curve where escape latencies were similar over time(Figure 1(a)) The latency to the former platform location inthe probe trial 24 hours after the last day of training (day4) was also significantly longer for Tg2576 mice than forwild type mice (471 s compared to 119 s 119901 lt 005 Mann-Whitney test Figure 1(b))Wild typemice crossed the formerlocation of the platform more often than the Tg2576 mice(119901 lt 005 Mann-Whitney test Figure 1(c)) In contrast theswimming velocity and the total distance the mice swamduring the acquisition period (Figures S2A and S2B) did notdiffer significantly between the groups Differences in escapelatency between female andmalemice were also assessed andno significant differences appeared
33 Intrahippocampal hNSC Transplantation Prevents Mem-ory Deterioration in Tg2576 Mice To ensure that there wereno differences in learning and memory among the Tg2576mice that could be attributed to the age range (6ndash9 months)the mice were subjected to a baseline MWM test beforethe onset of treatment (an overview of the study design isprovided in Figure 1(d)) No significant differences in testresults were observed among the different treatment groups(at baseline or at follow-up) during the 5 days of acquisitiontraining In general improvements in escape latencies wereminor and resulted in similar shallow learning curves as thoseobserved during training of wild type mice in the pilot studyIn the 24-hour probe trial therewere no differences in latencybaseline values between groups (Figure 1(e) 119901 = 026 Dunnrsquostest) The cohort of hNSC-transplanted Tg2576 mice had amemory performance comparable to their baseline valuesat the MWM follow-up test whereas the other treatmentsgroups deteriorated compared to baseline (Figure 1(e)) Theprogression of pathology as well as extensive neurosurgerycould have influenced the performance at follow-up ThehNSC-transplanted mice found the former platform locationsignificantly faster than SHAM-transplanted mice in theMWM follow-up test (Δ latency 110 s and 399 s resp119901 lt 005 Dunnrsquos test Figure 1(f)) The hNSC-transplantedTg2576 mice treated with either JN403 or (+)-phenserineshowed no memory improvements compared to SHAM-transplanted mice (Figure 1(f))
Neural Plasticity 5
60
40
20
0
1 2 3 4
Esca
pe la
tenc
y (s
)
Time (days)
WtTg2576
(a)
Wt Tg2576
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
lowast
(b)
Wt Tg2576
3
2
1
0
Num
ber o
f pla
tform
cros
ses
lowast
(c)
Drug treatment
Was
hout
per
iod
Mor
ris w
ater
maz
e
Mor
ris w
ater
maz
e
Tran
spla
ntat
ion
Treatment groups
minusfo
llow
-up
minusba
selin
e
1W 1W 1W4W 1W
Vehicle injected + saline (SHAM + SAL) n = 9
hNSC + saline (hNSC + SAL) n = 9
hNSC + JN403 (hNSC + JN) n = 5
hNSC + (+)-phenserine (hNSC + PHEN) n = 7
(d)
80
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
BaselineFollow-up
SHAM + SAL + SALhNSC + JNhNSC + PHENhNSC
(e)
SHAM + SAL
60
40
20
0
Δ la
tenc
y (s
)
lowast
+ SALhNSC + JNhNSC + PHENhNSC
(f)
Figure 1 Continued
6 Neural Plasticity
SHAM + SAL
50
40
30
20
10
0
Tim
e in
targ
et q
uadr
ant (
)
+ SALhNSC + JNhNSC + PHENhNSC
(g)
Figure 1 Intrahippocampal hNSCs transplantation affects spatial navigation memory in Tg2576 mice Learning and memory were assessedin 5- to 7-month-old wild type (wt 119899 = 6) and Tg2576 (APPswe 119899 = 6) mice in the pilot Morris water maze task (a) Escape latency during 4days of acquisition training (b) Latency to the former location of the platform or (c) the number of platform crosses in a probe test 24 hourslater lowast119901 lt 005 compared to wild typeThe data are expressed as means plusmn SEM (d)The experimental design of the study with 6- to 9-month-old Tg2576 mice (119899 = 30) comprised (i) Morris water maze tests with 5 days of acquisition followed 24 hours later by a probe memory test(ii) daily intraperitoneal injections of 03mgkg JN403 or 25mgkg (+)-phenserine for 5 weeks in total (iii) bilateral hippocampal injectionsof sim25000 hNSCshemisphere after 1 week of drug treatment (iv) repetition of the Morris water maze tasks 5 weeks after transplantationand (v) sacrifice by transcardial perfusion with PBS (e) Time taken to find the former platform location in the baseline and follow-up test(f) Differences in time taken to find the former platform location (Δ latency follow-up probe test values minus baseline probe values) (g)Percent time spent in the target quadrant where the platform was located lowast119901 lt 005 compared to SHAM + SAL The data are expressed asmeans plusmn SEM
The standardmeasurement ofmemory is the percent timespent in the target quadrant compared with the other quad-rants rather than the latency to reach the former location ofthe platform The different groups of mice spent an equallyshort time in the target quadrant thus showing a low persis-tence which is not unusual for mice (Figure 1(g)) Swimmingvelocity or the total distance swum did not differ amongTg2576 mice in the various groups (Figures S2C and S2D)
34 hNSCTransplantation Increases EndogenousNeurogenesisin the Dentate Gyrus The early neuronal marker DCX istransiently expressed during a period that extends fromapro-liferative progenitor cell stage to a postmitotic phase at whichDCX-positive cells differentiate into mature neurons withincreasing dendritic arborization in the DG as illustrated inFigure 2(a) We thus characterized neural stemprogenitorcells DCX+ cells as an index of neurogenesisThese cells mayderive from the resident pool of progenitor cells in the hippo-campal DG or from the exogenous source of transplantedhNSCs DCX+ cells were detected exclusively in the subgran-ular layer while cells positive for the marker human nuclei(hNuclei+ cells) as described in the following section wereobservedmainly in the polymorph layer of the DGThis indi-cates that the DCX+ cells detected were mainly derived fromthe endogenous stem cell pool Immunohistochemical analy-sis demonstrated that hNSC transplantation in Tg2576 miceaugmented endogenous neurogenesis as increases in boththe total number of DCX+ cells (56 higher) and the numberof maturing DCX+ cells (82 higher) were observed in theDG compared to SHAM transplantation (both 119901 lt 005Dunnrsquos test Figures 2(b) and 2(c))
Treatment with JN403 and (+)-phenserine howevercounteracted the beneficial effects of hNSC transplantationon endogenous neurogenesis Moreover JN403 reduced thetotal number of DCX+ cells (43 lower 119901 lt 001 Dunnrsquostest) as well as the number of DCX+ cells with dendrites (55lower 119901 lt 005 Dunnrsquos test) compared to vehicle-treatedhNSC-transplanted mice Treatment of hNSC-transplantedmice with (+)-phenserine did not significantly alter thenumber of DCX+ cells (Figures 2(b) and 2(c))There were nosignificant differences in the number of dendritic branchesper DCX+ cell among the different groups (Figure 2(d))
The number of DCX+ cells with dendrites inversely cor-related with measures of the Δ latency obtained in the probetrial (119901 lt 001 119903 = minus061 Spearmanrsquos rank Figure 3(e))indicating that the increasedmaturation of newborn neuronsin the DG following hNSC transplantation was related toan improvement in hippocampus-dependent memory Rep-resentative images of DCX+ cells in the DG of hNSC- andSHAM-transplanted Tg2576 mice are shown in Figures 2(f)and 2(g)
35 (+)-Phenserine Treatment Increases the Survival of hNSC-Derived Neurons in the Hippocampus of Tg2576 Mice Chro-nic treatment with (+)-phenserine increased the survival ofengrafted hNSCs in the DG of Tg2576 mice as demonstratedby human nuclei antibody labeling The number of hNuclei+cells was larger in Tg2576 mice treated with (+)-phenserinethan in those receiving saline (43 increase 119901 lt 005 Dunnrsquostest) while there were no differences in numbers of hNuclei+cells between JN403- and saline-treated mice (Figure 3(a))Costaining hNuclei+ cells with MAP2 or GFAP markers for
Neural Plasticity 7
Proliferation Maturation Maturation with dendritic arborization
(a)
50
40
30
20
10
0SHAM + SAL
lowast
Num
ber o
f DCX
+ce
lls in
den
tate
gyr
us
+ SALhNSC + JNhNSC + PHENhNSC
(b)
SHAM + SAL
lowast
10
15
20
25
0
5
Num
ber o
f DCX
+ce
lls w
ith d
endr
ites
+ SALhNSC + JNhNSC + PHENhNSC
(c)
SHAM + SAL0
5
1
2
3
4
Den
driti
c bra
nche
sD
CX+
cells
with
den
drite
s
+ SALhNSC + JNhNSC + PHENhNSC
(d)
0
20
0 10 20 30 40
40
60
80
minus20
Prob
eΔla
tenc
y
r = minus061lowastlowastp lt 001
Number of DCX+ cells with dendrites
(e)
SHAM + SAL + SALhNSC
(f)
SHAM + SAL + SALhNSC
(g)
Figure 2 hNSC transplantation increases the number and the maturation of new neurons in the dentate gyrus (a) Doublecortin- (DCX-)labeled cells in the dentate gyrus (DG) of Tg2576micewere characterized according to their numbers and the level ofmaturation and dendriticarborization (b)The total number of DCX+ cells (c) the number of DCX+ cells possessing dendrites and (d) the average number of dendriticbranches on DCX+ cells in the DG inmice receiving SHAM (vehicle only) transplantation and saline (SHAM+ SAL) or mice receiving hNSCtransplantation and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (e) Correlation between the number ofDCX+ cells with dendrites and the difference in time taken to find the position of the platform (Δ latency follow-up probe test values minusbaseline acquisition values) in theMorris watermaze task in groups of Tg2576mice treatedwith hNSC+ SAL hNSC+ JN and hNSC+PHEN(119903 = minus061 lowastlowast119901 lt 001 119899 = 18) Representative images of immunostaining with DCX+ cells in the DG of TG2576 mice at (f) 10x and (g) 20xmagnification lowast119901 lt 005 compared to SHAM + SAL 119901 lt 005 and
119901 lt 001 compared to hNSC + SALThe data are expressed as means plusmnSEM
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 3
the following coordinates relative to the bregma AP minus206ML plusmn175 and DV minus175mm Using a 26-gauge microsyringe(ILS Microsyringes Stutzerbach Germany) approximately25000 cells per hemisphere were injected in a total volumeof 1 120583L into the hippocampal DG of each Tg2576 mouseand the needle was thereafter kept in the injection site forapproximately 30 seconds to allow the volume to diffuseThe wound was then sutured with metal clips Lidocaine wasused for local anesthesia during the procedure and 032Msucrose solution was injected subcutaneously after surgeryThe animalsweremonitored daily for bodyweight and properhealing of the incision site after surgery No immunosuppre-ssants were used and no symptoms indicative of a reactionto the transplant were observed within the time frame inves-tigated
Drug treatment was interrupted on the day of transplan-tation and the mice were allowed a 3-day recovery periodbefore drug treatment was continued for an additional 4weeks Demographic information of all the mice included inthe transplantation is provided in Supplementary Table 1 (seeSupplementary Material available online at httpdxdoiorg1011552015370432) and the study outline is illustrated inFigure 2(a)
26 Morris Water Maze First a pilot study was conductedin which 5ndash7-month-old Tg2576 mice and their age-matchedwild type littermates underwent testing for spatial learningand memory in the MWM task The escape latency (time toreach the hidden platform) for Tg2576mice and age-matchedwild type littermates was compared during acquisition trialsof 60 secondsrsquo duration which were performed 6 times a dayat 10-minute intervals for 4 days Both the latency to the cen-ter of the platform (time to reach the former platform loca-tion) and the number of platform crossings were measured
Later in the follow-up acquisition study the Tg2576mice(vehicle- or hNSC-transplanted and saline- or drug-treated)now aged 6ndash9 months underwent 60-second trials 4 times aday at 15-second intervals for 5 days To assess retention ofspatial memory 24 hours after the last acquisition trial theplatform was removed and the mice were tracked in the poolfor 60 seconds in a probe trial Both the latency to the centerof the platform and the time spent in the target quadrant weremeasured The mice were randomly placed in one of fourfixed positions around thewall of a 1mdiameter round swim-ming pool During the acquisition period the mice learnedthe location of a hidden platform (9 times 9 cm) which wasplaced 1 cm under the water surface and 25 cm from the wallThe location of the platform was learned through visual cues(wall posters) on the walls around the pool The water tem-perature was maintained at 21∘C plusmn 2∘C and the water wasmade opaque (white) to contrast the dark color of the micefrom the black pool The mice were habituated a day beforethe first acquisition day by letting them swim in the pool for60 seconds without the platform The platform location wasfixed during the whole acquisition phase Mice that did notfind the platform within 60 seconds were aided to stay onthe platform for 10 seconds to assist the learning processThebehavior of the mice in the MWM task was recorded by anautomated video-tracking system (Ethovision) To evaluate
the differences in memory retention between the groups inthe probe trial the baseline values for latency were subtractedfrom the follow-up latency values (Δ latency)
Two mice (one in the hNSC-transplanted and saline-treated group as well as one in the hNSC-transplanted and(+)-phenserine treated group) could not be detected by theautomated video-tracking system because of their color andthese mice were therefore omitted from the MWM tests
27 Tissue Processing for Biochemical Analyses Within 12ndash16hours of completing the final behavioral assessments the ani-mals were anesthetized with a 1 1 ketaminexylazine mixture(100mgkg ketamine and 20mgkg xylazine) and sacrificedby transcardial perfusion with PBS buffer (pH 74) Thebrain from each animal was separated into hemispheres theleft hemisphere was snap frozen on dry-ice and the righthemisphere was postfixed in buffered 4 paraformaldehydeand then transferred to a 032M sucrose cryoprotectant solu-tion for 24 h at 4∘C The brain hemispheres were stored atminus80∘C until further analysisThe left-brain hemispheres weresubsequently used in the biochemical assays and the righthemispheres were cryostat sectioned and used in immuno-histochemistry studies
28 Immunohistochemistry For staining of differentiatedhNSCs in culture cells were incubated with rabbit anti-human glial fibrillary acidic protein (GFAP Dako GlostrupDenmark) mouse anti-120573III-tubulin (Sigma St Louis MOUSA) or mouse polyclonal anti-microtubule-associated pro-tein 2 (MAP2 Sigma St Louis MO USA) All primary anti-bodies were diluted 1 250 Thereafter cells were incubatedwith secondary Alexa Fluor conjugated antibodies (1 500donkey anti-mouse Alexa Fluor 546 or donkey anti-rabbitAlexa Fluor 488 Molecular Probes Eugene OR USA) Cellsimmunopositive for DAPI and GFAP or 120573III-tubulin or thenumber of dendritic branches perDAPI andMAP2 immuno-positive cell was counted in three random fields per cover slip(six fields per treatment under a Nikon E800 microscope)
In order to obtain comparable results between the differ-ent animals for each individual animal three coronal sectionsfrom the same hippocampal regions were selected for immu-nostaining Newly generated neurons in the hippocampuswere labeled with an antibody for themicrotubule-associatedprotein Doublecortin (DCX) (goat anti-DCX 1 200 SantaCruz Heidelberg Germany) according to the method des-cribed previously by Lilja et al [10] DCX immunoreactivitywas analyzed in three serial sections for each animal AllDCX+ cells in the DG were quantified blindly in which thetotal number of DCX+ cells the proportion of DCX+ cellswith dendrites and the number of branch points on theDCX+ cells with more pronounced dendritic branching werecounted
To investigate graft survival and the fate of transplantedhNSCs in the DG fluorescent immunohistochemical anal-ysis was carried out on serial sections from each animalusing primary antibodies polyclonal rabbit antiglial fibril-lary acidic protein (GFAP) (1 500 Dako Glostrup Den-mark) polyclonal rabbit anti-microtubule-associated protein2 (MAP2) (1 500 Millipore Temecula CA USA) and
4 Neural Plasticity
monoclonal mouse anti-human nuclei (hNuclei) (1 200Millipore Temecula CA USA) Secondary antibodies wereconjugated with Alexa Fluor (AF AF 488 donkey anti-rabbitand AF 546 donkey anti-mouse 1 500 Molecular ProbesEugene OR USA) In control experiments the primaryantibodywas omitted and a blocking bufferwas substituted toverify the specificity of the secondary antibodies The speci-ficity of hNuclei antibody staining was also assessed in micethat had not received any hNSCs implants Immunoreactivecells within the DG (including the subgranular zone andgranular and polymorph layers) were counted in three serialsections from each animal
The number of GFAP1205727 nAChR immunopositive cellswas evaluated on three serial coronal sections per animalafter consecutive incubations with primary antibodies (1 500rabbit anti-GFAP Dako 1 100 rat anti-1205727 nAChR Abcam)and secondary antibodies (1 200 goat anti-rabbit IgG-Fabhorseradish peroxidase- (HRP-) conjugated goat anti-ratIgG-Fab HRP-conjugated Invitrogen La Jolla CA USA)Immunoreactivity was visualized using peroxidase substratekits (DAB and SG Vector resp) The results were quanti-fied at 20x magnification in a prespecified area (325 120583m times435 120583m= 141375 120583m2) using software ImageJ (NIH BethesdaMD USA)
Nonfluorescent staining was evaluated under a light mic-roscope (Leica Wetzlar Germany) with an attached ProgResSpeedXT core5 video camera and an image analysis system(ProgRes Capture Pro288) Fluorescent staining was evalu-ated under a Nikon E800 microscope at 20x magnificationusing Adobe Photoshop CS5 software (San Jose CA USA)
29 A12057340 and A12057342 ELISA Frontal cortices and hippocampiwere extracted from the left hemisphere immersed in PBSbuffer (pH 74) and homogenized Samples were diluted 1 25in BSAT-DPBS (Dulbeccorsquos PBS with 5 BSA 003 Tween-20 and 1x protease inhibitor cocktail) A120573
40
and A12057342
levelswere measured using a sandwich ELISA assay (InvitrogenCamarillo CA USA) according to the manufacturersrsquo ins-tructions and the absorbance was read at 450 nm (TecanInfinite M1000)
210 Statistical Analysis GraphPad Prism 50 or 6 (GraphPadSoftware Inc) was used for all statistical analyses Datawere analyzed using the Mann-Whitney test for comparisonbetween two groups and theKruskal-Wallis one-wayANOVAtest by rank followed by Dunnrsquos post hoc test for compar-ison of gt2 groups for both in vitro and in vivo analysesRepeated measures ANOVA was applied for comparison ofwild type and Tg2576 mice in the acquisition tests in Morriswater maze Spearmanrsquos rank correlation was used in thecorrelation analysis which was visualized graphically usingsimple regression analysisThe data are presented as means plusmnstandard error of the mean (SEM) 119901 values lt 005 wereconsidered to be significant
3 Results
31 JN403 Exerts Neuroprotective Effects on hNSC-DerivedNeurons in Culture Neurotrophic actions of the 1205727 nAChR
partial agonist JN403 and the A120573 modulatory drug (+)-phenserine were assessed by examining the survival dif-ferentiation and maturation of progenitor cells in vitroJN403 protected hNSC-derived120573III-tubulinmarked neuronsagainst brain extract from a patient with AD containingvarious A120573 species (Figure S1A) On the other hand JN403 or(+)-phenserine did not significantly alter the number andorpattern of MAP2-positive Tg2576 cortical neurons in culture(Figure S1B)
32 Tg2576 Mice Demonstrate Impaired Spatial Memory Nav-igation Hippocampal-dependent spatial learning andmem-ory in Tg2576 mice (119899 = 6) and age-matched nontransgeniclittermates (wild type 119899 = 6) were tested in the MWM nav-igation task During the four days of acquisition training ofTg2576 mice resulted in an expected learning curve whereastraining of the wild type mice resulted in a very shallowlearning curve where escape latencies were similar over time(Figure 1(a)) The latency to the former platform location inthe probe trial 24 hours after the last day of training (day4) was also significantly longer for Tg2576 mice than forwild type mice (471 s compared to 119 s 119901 lt 005 Mann-Whitney test Figure 1(b))Wild typemice crossed the formerlocation of the platform more often than the Tg2576 mice(119901 lt 005 Mann-Whitney test Figure 1(c)) In contrast theswimming velocity and the total distance the mice swamduring the acquisition period (Figures S2A and S2B) did notdiffer significantly between the groups Differences in escapelatency between female andmalemice were also assessed andno significant differences appeared
33 Intrahippocampal hNSC Transplantation Prevents Mem-ory Deterioration in Tg2576 Mice To ensure that there wereno differences in learning and memory among the Tg2576mice that could be attributed to the age range (6ndash9 months)the mice were subjected to a baseline MWM test beforethe onset of treatment (an overview of the study design isprovided in Figure 1(d)) No significant differences in testresults were observed among the different treatment groups(at baseline or at follow-up) during the 5 days of acquisitiontraining In general improvements in escape latencies wereminor and resulted in similar shallow learning curves as thoseobserved during training of wild type mice in the pilot studyIn the 24-hour probe trial therewere no differences in latencybaseline values between groups (Figure 1(e) 119901 = 026 Dunnrsquostest) The cohort of hNSC-transplanted Tg2576 mice had amemory performance comparable to their baseline valuesat the MWM follow-up test whereas the other treatmentsgroups deteriorated compared to baseline (Figure 1(e)) Theprogression of pathology as well as extensive neurosurgerycould have influenced the performance at follow-up ThehNSC-transplanted mice found the former platform locationsignificantly faster than SHAM-transplanted mice in theMWM follow-up test (Δ latency 110 s and 399 s resp119901 lt 005 Dunnrsquos test Figure 1(f)) The hNSC-transplantedTg2576 mice treated with either JN403 or (+)-phenserineshowed no memory improvements compared to SHAM-transplanted mice (Figure 1(f))
Neural Plasticity 5
60
40
20
0
1 2 3 4
Esca
pe la
tenc
y (s
)
Time (days)
WtTg2576
(a)
Wt Tg2576
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
lowast
(b)
Wt Tg2576
3
2
1
0
Num
ber o
f pla
tform
cros
ses
lowast
(c)
Drug treatment
Was
hout
per
iod
Mor
ris w
ater
maz
e
Mor
ris w
ater
maz
e
Tran
spla
ntat
ion
Treatment groups
minusfo
llow
-up
minusba
selin
e
1W 1W 1W4W 1W
Vehicle injected + saline (SHAM + SAL) n = 9
hNSC + saline (hNSC + SAL) n = 9
hNSC + JN403 (hNSC + JN) n = 5
hNSC + (+)-phenserine (hNSC + PHEN) n = 7
(d)
80
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
BaselineFollow-up
SHAM + SAL + SALhNSC + JNhNSC + PHENhNSC
(e)
SHAM + SAL
60
40
20
0
Δ la
tenc
y (s
)
lowast
+ SALhNSC + JNhNSC + PHENhNSC
(f)
Figure 1 Continued
6 Neural Plasticity
SHAM + SAL
50
40
30
20
10
0
Tim
e in
targ
et q
uadr
ant (
)
+ SALhNSC + JNhNSC + PHENhNSC
(g)
Figure 1 Intrahippocampal hNSCs transplantation affects spatial navigation memory in Tg2576 mice Learning and memory were assessedin 5- to 7-month-old wild type (wt 119899 = 6) and Tg2576 (APPswe 119899 = 6) mice in the pilot Morris water maze task (a) Escape latency during 4days of acquisition training (b) Latency to the former location of the platform or (c) the number of platform crosses in a probe test 24 hourslater lowast119901 lt 005 compared to wild typeThe data are expressed as means plusmn SEM (d)The experimental design of the study with 6- to 9-month-old Tg2576 mice (119899 = 30) comprised (i) Morris water maze tests with 5 days of acquisition followed 24 hours later by a probe memory test(ii) daily intraperitoneal injections of 03mgkg JN403 or 25mgkg (+)-phenserine for 5 weeks in total (iii) bilateral hippocampal injectionsof sim25000 hNSCshemisphere after 1 week of drug treatment (iv) repetition of the Morris water maze tasks 5 weeks after transplantationand (v) sacrifice by transcardial perfusion with PBS (e) Time taken to find the former platform location in the baseline and follow-up test(f) Differences in time taken to find the former platform location (Δ latency follow-up probe test values minus baseline probe values) (g)Percent time spent in the target quadrant where the platform was located lowast119901 lt 005 compared to SHAM + SAL The data are expressed asmeans plusmn SEM
The standardmeasurement ofmemory is the percent timespent in the target quadrant compared with the other quad-rants rather than the latency to reach the former location ofthe platform The different groups of mice spent an equallyshort time in the target quadrant thus showing a low persis-tence which is not unusual for mice (Figure 1(g)) Swimmingvelocity or the total distance swum did not differ amongTg2576 mice in the various groups (Figures S2C and S2D)
34 hNSCTransplantation Increases EndogenousNeurogenesisin the Dentate Gyrus The early neuronal marker DCX istransiently expressed during a period that extends fromapro-liferative progenitor cell stage to a postmitotic phase at whichDCX-positive cells differentiate into mature neurons withincreasing dendritic arborization in the DG as illustrated inFigure 2(a) We thus characterized neural stemprogenitorcells DCX+ cells as an index of neurogenesisThese cells mayderive from the resident pool of progenitor cells in the hippo-campal DG or from the exogenous source of transplantedhNSCs DCX+ cells were detected exclusively in the subgran-ular layer while cells positive for the marker human nuclei(hNuclei+ cells) as described in the following section wereobservedmainly in the polymorph layer of the DGThis indi-cates that the DCX+ cells detected were mainly derived fromthe endogenous stem cell pool Immunohistochemical analy-sis demonstrated that hNSC transplantation in Tg2576 miceaugmented endogenous neurogenesis as increases in boththe total number of DCX+ cells (56 higher) and the numberof maturing DCX+ cells (82 higher) were observed in theDG compared to SHAM transplantation (both 119901 lt 005Dunnrsquos test Figures 2(b) and 2(c))
Treatment with JN403 and (+)-phenserine howevercounteracted the beneficial effects of hNSC transplantationon endogenous neurogenesis Moreover JN403 reduced thetotal number of DCX+ cells (43 lower 119901 lt 001 Dunnrsquostest) as well as the number of DCX+ cells with dendrites (55lower 119901 lt 005 Dunnrsquos test) compared to vehicle-treatedhNSC-transplanted mice Treatment of hNSC-transplantedmice with (+)-phenserine did not significantly alter thenumber of DCX+ cells (Figures 2(b) and 2(c))There were nosignificant differences in the number of dendritic branchesper DCX+ cell among the different groups (Figure 2(d))
The number of DCX+ cells with dendrites inversely cor-related with measures of the Δ latency obtained in the probetrial (119901 lt 001 119903 = minus061 Spearmanrsquos rank Figure 3(e))indicating that the increasedmaturation of newborn neuronsin the DG following hNSC transplantation was related toan improvement in hippocampus-dependent memory Rep-resentative images of DCX+ cells in the DG of hNSC- andSHAM-transplanted Tg2576 mice are shown in Figures 2(f)and 2(g)
35 (+)-Phenserine Treatment Increases the Survival of hNSC-Derived Neurons in the Hippocampus of Tg2576 Mice Chro-nic treatment with (+)-phenserine increased the survival ofengrafted hNSCs in the DG of Tg2576 mice as demonstratedby human nuclei antibody labeling The number of hNuclei+cells was larger in Tg2576 mice treated with (+)-phenserinethan in those receiving saline (43 increase 119901 lt 005 Dunnrsquostest) while there were no differences in numbers of hNuclei+cells between JN403- and saline-treated mice (Figure 3(a))Costaining hNuclei+ cells with MAP2 or GFAP markers for
Neural Plasticity 7
Proliferation Maturation Maturation with dendritic arborization
(a)
50
40
30
20
10
0SHAM + SAL
lowast
Num
ber o
f DCX
+ce
lls in
den
tate
gyr
us
+ SALhNSC + JNhNSC + PHENhNSC
(b)
SHAM + SAL
lowast
10
15
20
25
0
5
Num
ber o
f DCX
+ce
lls w
ith d
endr
ites
+ SALhNSC + JNhNSC + PHENhNSC
(c)
SHAM + SAL0
5
1
2
3
4
Den
driti
c bra
nche
sD
CX+
cells
with
den
drite
s
+ SALhNSC + JNhNSC + PHENhNSC
(d)
0
20
0 10 20 30 40
40
60
80
minus20
Prob
eΔla
tenc
y
r = minus061lowastlowastp lt 001
Number of DCX+ cells with dendrites
(e)
SHAM + SAL + SALhNSC
(f)
SHAM + SAL + SALhNSC
(g)
Figure 2 hNSC transplantation increases the number and the maturation of new neurons in the dentate gyrus (a) Doublecortin- (DCX-)labeled cells in the dentate gyrus (DG) of Tg2576micewere characterized according to their numbers and the level ofmaturation and dendriticarborization (b)The total number of DCX+ cells (c) the number of DCX+ cells possessing dendrites and (d) the average number of dendriticbranches on DCX+ cells in the DG inmice receiving SHAM (vehicle only) transplantation and saline (SHAM+ SAL) or mice receiving hNSCtransplantation and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (e) Correlation between the number ofDCX+ cells with dendrites and the difference in time taken to find the position of the platform (Δ latency follow-up probe test values minusbaseline acquisition values) in theMorris watermaze task in groups of Tg2576mice treatedwith hNSC+ SAL hNSC+ JN and hNSC+PHEN(119903 = minus061 lowastlowast119901 lt 001 119899 = 18) Representative images of immunostaining with DCX+ cells in the DG of TG2576 mice at (f) 10x and (g) 20xmagnification lowast119901 lt 005 compared to SHAM + SAL 119901 lt 005 and
119901 lt 001 compared to hNSC + SALThe data are expressed as means plusmnSEM
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

4 Neural Plasticity
monoclonal mouse anti-human nuclei (hNuclei) (1 200Millipore Temecula CA USA) Secondary antibodies wereconjugated with Alexa Fluor (AF AF 488 donkey anti-rabbitand AF 546 donkey anti-mouse 1 500 Molecular ProbesEugene OR USA) In control experiments the primaryantibodywas omitted and a blocking bufferwas substituted toverify the specificity of the secondary antibodies The speci-ficity of hNuclei antibody staining was also assessed in micethat had not received any hNSCs implants Immunoreactivecells within the DG (including the subgranular zone andgranular and polymorph layers) were counted in three serialsections from each animal
The number of GFAP1205727 nAChR immunopositive cellswas evaluated on three serial coronal sections per animalafter consecutive incubations with primary antibodies (1 500rabbit anti-GFAP Dako 1 100 rat anti-1205727 nAChR Abcam)and secondary antibodies (1 200 goat anti-rabbit IgG-Fabhorseradish peroxidase- (HRP-) conjugated goat anti-ratIgG-Fab HRP-conjugated Invitrogen La Jolla CA USA)Immunoreactivity was visualized using peroxidase substratekits (DAB and SG Vector resp) The results were quanti-fied at 20x magnification in a prespecified area (325 120583m times435 120583m= 141375 120583m2) using software ImageJ (NIH BethesdaMD USA)
Nonfluorescent staining was evaluated under a light mic-roscope (Leica Wetzlar Germany) with an attached ProgResSpeedXT core5 video camera and an image analysis system(ProgRes Capture Pro288) Fluorescent staining was evalu-ated under a Nikon E800 microscope at 20x magnificationusing Adobe Photoshop CS5 software (San Jose CA USA)
29 A12057340 and A12057342 ELISA Frontal cortices and hippocampiwere extracted from the left hemisphere immersed in PBSbuffer (pH 74) and homogenized Samples were diluted 1 25in BSAT-DPBS (Dulbeccorsquos PBS with 5 BSA 003 Tween-20 and 1x protease inhibitor cocktail) A120573
40
and A12057342
levelswere measured using a sandwich ELISA assay (InvitrogenCamarillo CA USA) according to the manufacturersrsquo ins-tructions and the absorbance was read at 450 nm (TecanInfinite M1000)
210 Statistical Analysis GraphPad Prism 50 or 6 (GraphPadSoftware Inc) was used for all statistical analyses Datawere analyzed using the Mann-Whitney test for comparisonbetween two groups and theKruskal-Wallis one-wayANOVAtest by rank followed by Dunnrsquos post hoc test for compar-ison of gt2 groups for both in vitro and in vivo analysesRepeated measures ANOVA was applied for comparison ofwild type and Tg2576 mice in the acquisition tests in Morriswater maze Spearmanrsquos rank correlation was used in thecorrelation analysis which was visualized graphically usingsimple regression analysisThe data are presented as means plusmnstandard error of the mean (SEM) 119901 values lt 005 wereconsidered to be significant
3 Results
31 JN403 Exerts Neuroprotective Effects on hNSC-DerivedNeurons in Culture Neurotrophic actions of the 1205727 nAChR
partial agonist JN403 and the A120573 modulatory drug (+)-phenserine were assessed by examining the survival dif-ferentiation and maturation of progenitor cells in vitroJN403 protected hNSC-derived120573III-tubulinmarked neuronsagainst brain extract from a patient with AD containingvarious A120573 species (Figure S1A) On the other hand JN403 or(+)-phenserine did not significantly alter the number andorpattern of MAP2-positive Tg2576 cortical neurons in culture(Figure S1B)
32 Tg2576 Mice Demonstrate Impaired Spatial Memory Nav-igation Hippocampal-dependent spatial learning andmem-ory in Tg2576 mice (119899 = 6) and age-matched nontransgeniclittermates (wild type 119899 = 6) were tested in the MWM nav-igation task During the four days of acquisition training ofTg2576 mice resulted in an expected learning curve whereastraining of the wild type mice resulted in a very shallowlearning curve where escape latencies were similar over time(Figure 1(a)) The latency to the former platform location inthe probe trial 24 hours after the last day of training (day4) was also significantly longer for Tg2576 mice than forwild type mice (471 s compared to 119 s 119901 lt 005 Mann-Whitney test Figure 1(b))Wild typemice crossed the formerlocation of the platform more often than the Tg2576 mice(119901 lt 005 Mann-Whitney test Figure 1(c)) In contrast theswimming velocity and the total distance the mice swamduring the acquisition period (Figures S2A and S2B) did notdiffer significantly between the groups Differences in escapelatency between female andmalemice were also assessed andno significant differences appeared
33 Intrahippocampal hNSC Transplantation Prevents Mem-ory Deterioration in Tg2576 Mice To ensure that there wereno differences in learning and memory among the Tg2576mice that could be attributed to the age range (6ndash9 months)the mice were subjected to a baseline MWM test beforethe onset of treatment (an overview of the study design isprovided in Figure 1(d)) No significant differences in testresults were observed among the different treatment groups(at baseline or at follow-up) during the 5 days of acquisitiontraining In general improvements in escape latencies wereminor and resulted in similar shallow learning curves as thoseobserved during training of wild type mice in the pilot studyIn the 24-hour probe trial therewere no differences in latencybaseline values between groups (Figure 1(e) 119901 = 026 Dunnrsquostest) The cohort of hNSC-transplanted Tg2576 mice had amemory performance comparable to their baseline valuesat the MWM follow-up test whereas the other treatmentsgroups deteriorated compared to baseline (Figure 1(e)) Theprogression of pathology as well as extensive neurosurgerycould have influenced the performance at follow-up ThehNSC-transplanted mice found the former platform locationsignificantly faster than SHAM-transplanted mice in theMWM follow-up test (Δ latency 110 s and 399 s resp119901 lt 005 Dunnrsquos test Figure 1(f)) The hNSC-transplantedTg2576 mice treated with either JN403 or (+)-phenserineshowed no memory improvements compared to SHAM-transplanted mice (Figure 1(f))
Neural Plasticity 5
60
40
20
0
1 2 3 4
Esca
pe la
tenc
y (s
)
Time (days)
WtTg2576
(a)
Wt Tg2576
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
lowast
(b)
Wt Tg2576
3
2
1
0
Num
ber o
f pla
tform
cros
ses
lowast
(c)
Drug treatment
Was
hout
per
iod
Mor
ris w
ater
maz
e
Mor
ris w
ater
maz
e
Tran
spla
ntat
ion
Treatment groups
minusfo
llow
-up
minusba
selin
e
1W 1W 1W4W 1W
Vehicle injected + saline (SHAM + SAL) n = 9
hNSC + saline (hNSC + SAL) n = 9
hNSC + JN403 (hNSC + JN) n = 5
hNSC + (+)-phenserine (hNSC + PHEN) n = 7
(d)
80
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
BaselineFollow-up
SHAM + SAL + SALhNSC + JNhNSC + PHENhNSC
(e)
SHAM + SAL
60
40
20
0
Δ la
tenc
y (s
)
lowast
+ SALhNSC + JNhNSC + PHENhNSC
(f)
Figure 1 Continued
6 Neural Plasticity
SHAM + SAL
50
40
30
20
10
0
Tim
e in
targ
et q
uadr
ant (
)
+ SALhNSC + JNhNSC + PHENhNSC
(g)
Figure 1 Intrahippocampal hNSCs transplantation affects spatial navigation memory in Tg2576 mice Learning and memory were assessedin 5- to 7-month-old wild type (wt 119899 = 6) and Tg2576 (APPswe 119899 = 6) mice in the pilot Morris water maze task (a) Escape latency during 4days of acquisition training (b) Latency to the former location of the platform or (c) the number of platform crosses in a probe test 24 hourslater lowast119901 lt 005 compared to wild typeThe data are expressed as means plusmn SEM (d)The experimental design of the study with 6- to 9-month-old Tg2576 mice (119899 = 30) comprised (i) Morris water maze tests with 5 days of acquisition followed 24 hours later by a probe memory test(ii) daily intraperitoneal injections of 03mgkg JN403 or 25mgkg (+)-phenserine for 5 weeks in total (iii) bilateral hippocampal injectionsof sim25000 hNSCshemisphere after 1 week of drug treatment (iv) repetition of the Morris water maze tasks 5 weeks after transplantationand (v) sacrifice by transcardial perfusion with PBS (e) Time taken to find the former platform location in the baseline and follow-up test(f) Differences in time taken to find the former platform location (Δ latency follow-up probe test values minus baseline probe values) (g)Percent time spent in the target quadrant where the platform was located lowast119901 lt 005 compared to SHAM + SAL The data are expressed asmeans plusmn SEM
The standardmeasurement ofmemory is the percent timespent in the target quadrant compared with the other quad-rants rather than the latency to reach the former location ofthe platform The different groups of mice spent an equallyshort time in the target quadrant thus showing a low persis-tence which is not unusual for mice (Figure 1(g)) Swimmingvelocity or the total distance swum did not differ amongTg2576 mice in the various groups (Figures S2C and S2D)
34 hNSCTransplantation Increases EndogenousNeurogenesisin the Dentate Gyrus The early neuronal marker DCX istransiently expressed during a period that extends fromapro-liferative progenitor cell stage to a postmitotic phase at whichDCX-positive cells differentiate into mature neurons withincreasing dendritic arborization in the DG as illustrated inFigure 2(a) We thus characterized neural stemprogenitorcells DCX+ cells as an index of neurogenesisThese cells mayderive from the resident pool of progenitor cells in the hippo-campal DG or from the exogenous source of transplantedhNSCs DCX+ cells were detected exclusively in the subgran-ular layer while cells positive for the marker human nuclei(hNuclei+ cells) as described in the following section wereobservedmainly in the polymorph layer of the DGThis indi-cates that the DCX+ cells detected were mainly derived fromthe endogenous stem cell pool Immunohistochemical analy-sis demonstrated that hNSC transplantation in Tg2576 miceaugmented endogenous neurogenesis as increases in boththe total number of DCX+ cells (56 higher) and the numberof maturing DCX+ cells (82 higher) were observed in theDG compared to SHAM transplantation (both 119901 lt 005Dunnrsquos test Figures 2(b) and 2(c))
Treatment with JN403 and (+)-phenserine howevercounteracted the beneficial effects of hNSC transplantationon endogenous neurogenesis Moreover JN403 reduced thetotal number of DCX+ cells (43 lower 119901 lt 001 Dunnrsquostest) as well as the number of DCX+ cells with dendrites (55lower 119901 lt 005 Dunnrsquos test) compared to vehicle-treatedhNSC-transplanted mice Treatment of hNSC-transplantedmice with (+)-phenserine did not significantly alter thenumber of DCX+ cells (Figures 2(b) and 2(c))There were nosignificant differences in the number of dendritic branchesper DCX+ cell among the different groups (Figure 2(d))
The number of DCX+ cells with dendrites inversely cor-related with measures of the Δ latency obtained in the probetrial (119901 lt 001 119903 = minus061 Spearmanrsquos rank Figure 3(e))indicating that the increasedmaturation of newborn neuronsin the DG following hNSC transplantation was related toan improvement in hippocampus-dependent memory Rep-resentative images of DCX+ cells in the DG of hNSC- andSHAM-transplanted Tg2576 mice are shown in Figures 2(f)and 2(g)
35 (+)-Phenserine Treatment Increases the Survival of hNSC-Derived Neurons in the Hippocampus of Tg2576 Mice Chro-nic treatment with (+)-phenserine increased the survival ofengrafted hNSCs in the DG of Tg2576 mice as demonstratedby human nuclei antibody labeling The number of hNuclei+cells was larger in Tg2576 mice treated with (+)-phenserinethan in those receiving saline (43 increase 119901 lt 005 Dunnrsquostest) while there were no differences in numbers of hNuclei+cells between JN403- and saline-treated mice (Figure 3(a))Costaining hNuclei+ cells with MAP2 or GFAP markers for
Neural Plasticity 7
Proliferation Maturation Maturation with dendritic arborization
(a)
50
40
30
20
10
0SHAM + SAL
lowast
Num
ber o
f DCX
+ce
lls in
den
tate
gyr
us
+ SALhNSC + JNhNSC + PHENhNSC
(b)
SHAM + SAL
lowast
10
15
20
25
0
5
Num
ber o
f DCX
+ce
lls w
ith d
endr
ites
+ SALhNSC + JNhNSC + PHENhNSC
(c)
SHAM + SAL0
5
1
2
3
4
Den
driti
c bra
nche
sD
CX+
cells
with
den
drite
s
+ SALhNSC + JNhNSC + PHENhNSC
(d)
0
20
0 10 20 30 40
40
60
80
minus20
Prob
eΔla
tenc
y
r = minus061lowastlowastp lt 001
Number of DCX+ cells with dendrites
(e)
SHAM + SAL + SALhNSC
(f)
SHAM + SAL + SALhNSC
(g)
Figure 2 hNSC transplantation increases the number and the maturation of new neurons in the dentate gyrus (a) Doublecortin- (DCX-)labeled cells in the dentate gyrus (DG) of Tg2576micewere characterized according to their numbers and the level ofmaturation and dendriticarborization (b)The total number of DCX+ cells (c) the number of DCX+ cells possessing dendrites and (d) the average number of dendriticbranches on DCX+ cells in the DG inmice receiving SHAM (vehicle only) transplantation and saline (SHAM+ SAL) or mice receiving hNSCtransplantation and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (e) Correlation between the number ofDCX+ cells with dendrites and the difference in time taken to find the position of the platform (Δ latency follow-up probe test values minusbaseline acquisition values) in theMorris watermaze task in groups of Tg2576mice treatedwith hNSC+ SAL hNSC+ JN and hNSC+PHEN(119903 = minus061 lowastlowast119901 lt 001 119899 = 18) Representative images of immunostaining with DCX+ cells in the DG of TG2576 mice at (f) 10x and (g) 20xmagnification lowast119901 lt 005 compared to SHAM + SAL 119901 lt 005 and
119901 lt 001 compared to hNSC + SALThe data are expressed as means plusmnSEM
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 5
60
40
20
0
1 2 3 4
Esca
pe la
tenc
y (s
)
Time (days)
WtTg2576
(a)
Wt Tg2576
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
lowast
(b)
Wt Tg2576
3
2
1
0
Num
ber o
f pla
tform
cros
ses
lowast
(c)
Drug treatment
Was
hout
per
iod
Mor
ris w
ater
maz
e
Mor
ris w
ater
maz
e
Tran
spla
ntat
ion
Treatment groups
minusfo
llow
-up
minusba
selin
e
1W 1W 1W4W 1W
Vehicle injected + saline (SHAM + SAL) n = 9
hNSC + saline (hNSC + SAL) n = 9
hNSC + JN403 (hNSC + JN) n = 5
hNSC + (+)-phenserine (hNSC + PHEN) n = 7
(d)
80
60
40
20
0
Late
ncy
to ce
nter
of p
latfo
rm (s
)
BaselineFollow-up
SHAM + SAL + SALhNSC + JNhNSC + PHENhNSC
(e)
SHAM + SAL
60
40
20
0
Δ la
tenc
y (s
)
lowast
+ SALhNSC + JNhNSC + PHENhNSC
(f)
Figure 1 Continued
6 Neural Plasticity
SHAM + SAL
50
40
30
20
10
0
Tim
e in
targ
et q
uadr
ant (
)
+ SALhNSC + JNhNSC + PHENhNSC
(g)
Figure 1 Intrahippocampal hNSCs transplantation affects spatial navigation memory in Tg2576 mice Learning and memory were assessedin 5- to 7-month-old wild type (wt 119899 = 6) and Tg2576 (APPswe 119899 = 6) mice in the pilot Morris water maze task (a) Escape latency during 4days of acquisition training (b) Latency to the former location of the platform or (c) the number of platform crosses in a probe test 24 hourslater lowast119901 lt 005 compared to wild typeThe data are expressed as means plusmn SEM (d)The experimental design of the study with 6- to 9-month-old Tg2576 mice (119899 = 30) comprised (i) Morris water maze tests with 5 days of acquisition followed 24 hours later by a probe memory test(ii) daily intraperitoneal injections of 03mgkg JN403 or 25mgkg (+)-phenserine for 5 weeks in total (iii) bilateral hippocampal injectionsof sim25000 hNSCshemisphere after 1 week of drug treatment (iv) repetition of the Morris water maze tasks 5 weeks after transplantationand (v) sacrifice by transcardial perfusion with PBS (e) Time taken to find the former platform location in the baseline and follow-up test(f) Differences in time taken to find the former platform location (Δ latency follow-up probe test values minus baseline probe values) (g)Percent time spent in the target quadrant where the platform was located lowast119901 lt 005 compared to SHAM + SAL The data are expressed asmeans plusmn SEM
The standardmeasurement ofmemory is the percent timespent in the target quadrant compared with the other quad-rants rather than the latency to reach the former location ofthe platform The different groups of mice spent an equallyshort time in the target quadrant thus showing a low persis-tence which is not unusual for mice (Figure 1(g)) Swimmingvelocity or the total distance swum did not differ amongTg2576 mice in the various groups (Figures S2C and S2D)
34 hNSCTransplantation Increases EndogenousNeurogenesisin the Dentate Gyrus The early neuronal marker DCX istransiently expressed during a period that extends fromapro-liferative progenitor cell stage to a postmitotic phase at whichDCX-positive cells differentiate into mature neurons withincreasing dendritic arborization in the DG as illustrated inFigure 2(a) We thus characterized neural stemprogenitorcells DCX+ cells as an index of neurogenesisThese cells mayderive from the resident pool of progenitor cells in the hippo-campal DG or from the exogenous source of transplantedhNSCs DCX+ cells were detected exclusively in the subgran-ular layer while cells positive for the marker human nuclei(hNuclei+ cells) as described in the following section wereobservedmainly in the polymorph layer of the DGThis indi-cates that the DCX+ cells detected were mainly derived fromthe endogenous stem cell pool Immunohistochemical analy-sis demonstrated that hNSC transplantation in Tg2576 miceaugmented endogenous neurogenesis as increases in boththe total number of DCX+ cells (56 higher) and the numberof maturing DCX+ cells (82 higher) were observed in theDG compared to SHAM transplantation (both 119901 lt 005Dunnrsquos test Figures 2(b) and 2(c))
Treatment with JN403 and (+)-phenserine howevercounteracted the beneficial effects of hNSC transplantationon endogenous neurogenesis Moreover JN403 reduced thetotal number of DCX+ cells (43 lower 119901 lt 001 Dunnrsquostest) as well as the number of DCX+ cells with dendrites (55lower 119901 lt 005 Dunnrsquos test) compared to vehicle-treatedhNSC-transplanted mice Treatment of hNSC-transplantedmice with (+)-phenserine did not significantly alter thenumber of DCX+ cells (Figures 2(b) and 2(c))There were nosignificant differences in the number of dendritic branchesper DCX+ cell among the different groups (Figure 2(d))
The number of DCX+ cells with dendrites inversely cor-related with measures of the Δ latency obtained in the probetrial (119901 lt 001 119903 = minus061 Spearmanrsquos rank Figure 3(e))indicating that the increasedmaturation of newborn neuronsin the DG following hNSC transplantation was related toan improvement in hippocampus-dependent memory Rep-resentative images of DCX+ cells in the DG of hNSC- andSHAM-transplanted Tg2576 mice are shown in Figures 2(f)and 2(g)
35 (+)-Phenserine Treatment Increases the Survival of hNSC-Derived Neurons in the Hippocampus of Tg2576 Mice Chro-nic treatment with (+)-phenserine increased the survival ofengrafted hNSCs in the DG of Tg2576 mice as demonstratedby human nuclei antibody labeling The number of hNuclei+cells was larger in Tg2576 mice treated with (+)-phenserinethan in those receiving saline (43 increase 119901 lt 005 Dunnrsquostest) while there were no differences in numbers of hNuclei+cells between JN403- and saline-treated mice (Figure 3(a))Costaining hNuclei+ cells with MAP2 or GFAP markers for
Neural Plasticity 7
Proliferation Maturation Maturation with dendritic arborization
(a)
50
40
30
20
10
0SHAM + SAL
lowast
Num
ber o
f DCX
+ce
lls in
den
tate
gyr
us
+ SALhNSC + JNhNSC + PHENhNSC
(b)
SHAM + SAL
lowast
10
15
20
25
0
5
Num
ber o
f DCX
+ce
lls w
ith d
endr
ites
+ SALhNSC + JNhNSC + PHENhNSC
(c)
SHAM + SAL0
5
1
2
3
4
Den
driti
c bra
nche
sD
CX+
cells
with
den
drite
s
+ SALhNSC + JNhNSC + PHENhNSC
(d)
0
20
0 10 20 30 40
40
60
80
minus20
Prob
eΔla
tenc
y
r = minus061lowastlowastp lt 001
Number of DCX+ cells with dendrites
(e)
SHAM + SAL + SALhNSC
(f)
SHAM + SAL + SALhNSC
(g)
Figure 2 hNSC transplantation increases the number and the maturation of new neurons in the dentate gyrus (a) Doublecortin- (DCX-)labeled cells in the dentate gyrus (DG) of Tg2576micewere characterized according to their numbers and the level ofmaturation and dendriticarborization (b)The total number of DCX+ cells (c) the number of DCX+ cells possessing dendrites and (d) the average number of dendriticbranches on DCX+ cells in the DG inmice receiving SHAM (vehicle only) transplantation and saline (SHAM+ SAL) or mice receiving hNSCtransplantation and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (e) Correlation between the number ofDCX+ cells with dendrites and the difference in time taken to find the position of the platform (Δ latency follow-up probe test values minusbaseline acquisition values) in theMorris watermaze task in groups of Tg2576mice treatedwith hNSC+ SAL hNSC+ JN and hNSC+PHEN(119903 = minus061 lowastlowast119901 lt 001 119899 = 18) Representative images of immunostaining with DCX+ cells in the DG of TG2576 mice at (f) 10x and (g) 20xmagnification lowast119901 lt 005 compared to SHAM + SAL 119901 lt 005 and
119901 lt 001 compared to hNSC + SALThe data are expressed as means plusmnSEM
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

6 Neural Plasticity
SHAM + SAL
50
40
30
20
10
0
Tim
e in
targ
et q
uadr
ant (
)
+ SALhNSC + JNhNSC + PHENhNSC
(g)
Figure 1 Intrahippocampal hNSCs transplantation affects spatial navigation memory in Tg2576 mice Learning and memory were assessedin 5- to 7-month-old wild type (wt 119899 = 6) and Tg2576 (APPswe 119899 = 6) mice in the pilot Morris water maze task (a) Escape latency during 4days of acquisition training (b) Latency to the former location of the platform or (c) the number of platform crosses in a probe test 24 hourslater lowast119901 lt 005 compared to wild typeThe data are expressed as means plusmn SEM (d)The experimental design of the study with 6- to 9-month-old Tg2576 mice (119899 = 30) comprised (i) Morris water maze tests with 5 days of acquisition followed 24 hours later by a probe memory test(ii) daily intraperitoneal injections of 03mgkg JN403 or 25mgkg (+)-phenserine for 5 weeks in total (iii) bilateral hippocampal injectionsof sim25000 hNSCshemisphere after 1 week of drug treatment (iv) repetition of the Morris water maze tasks 5 weeks after transplantationand (v) sacrifice by transcardial perfusion with PBS (e) Time taken to find the former platform location in the baseline and follow-up test(f) Differences in time taken to find the former platform location (Δ latency follow-up probe test values minus baseline probe values) (g)Percent time spent in the target quadrant where the platform was located lowast119901 lt 005 compared to SHAM + SAL The data are expressed asmeans plusmn SEM
The standardmeasurement ofmemory is the percent timespent in the target quadrant compared with the other quad-rants rather than the latency to reach the former location ofthe platform The different groups of mice spent an equallyshort time in the target quadrant thus showing a low persis-tence which is not unusual for mice (Figure 1(g)) Swimmingvelocity or the total distance swum did not differ amongTg2576 mice in the various groups (Figures S2C and S2D)
34 hNSCTransplantation Increases EndogenousNeurogenesisin the Dentate Gyrus The early neuronal marker DCX istransiently expressed during a period that extends fromapro-liferative progenitor cell stage to a postmitotic phase at whichDCX-positive cells differentiate into mature neurons withincreasing dendritic arborization in the DG as illustrated inFigure 2(a) We thus characterized neural stemprogenitorcells DCX+ cells as an index of neurogenesisThese cells mayderive from the resident pool of progenitor cells in the hippo-campal DG or from the exogenous source of transplantedhNSCs DCX+ cells were detected exclusively in the subgran-ular layer while cells positive for the marker human nuclei(hNuclei+ cells) as described in the following section wereobservedmainly in the polymorph layer of the DGThis indi-cates that the DCX+ cells detected were mainly derived fromthe endogenous stem cell pool Immunohistochemical analy-sis demonstrated that hNSC transplantation in Tg2576 miceaugmented endogenous neurogenesis as increases in boththe total number of DCX+ cells (56 higher) and the numberof maturing DCX+ cells (82 higher) were observed in theDG compared to SHAM transplantation (both 119901 lt 005Dunnrsquos test Figures 2(b) and 2(c))
Treatment with JN403 and (+)-phenserine howevercounteracted the beneficial effects of hNSC transplantationon endogenous neurogenesis Moreover JN403 reduced thetotal number of DCX+ cells (43 lower 119901 lt 001 Dunnrsquostest) as well as the number of DCX+ cells with dendrites (55lower 119901 lt 005 Dunnrsquos test) compared to vehicle-treatedhNSC-transplanted mice Treatment of hNSC-transplantedmice with (+)-phenserine did not significantly alter thenumber of DCX+ cells (Figures 2(b) and 2(c))There were nosignificant differences in the number of dendritic branchesper DCX+ cell among the different groups (Figure 2(d))
The number of DCX+ cells with dendrites inversely cor-related with measures of the Δ latency obtained in the probetrial (119901 lt 001 119903 = minus061 Spearmanrsquos rank Figure 3(e))indicating that the increasedmaturation of newborn neuronsin the DG following hNSC transplantation was related toan improvement in hippocampus-dependent memory Rep-resentative images of DCX+ cells in the DG of hNSC- andSHAM-transplanted Tg2576 mice are shown in Figures 2(f)and 2(g)
35 (+)-Phenserine Treatment Increases the Survival of hNSC-Derived Neurons in the Hippocampus of Tg2576 Mice Chro-nic treatment with (+)-phenserine increased the survival ofengrafted hNSCs in the DG of Tg2576 mice as demonstratedby human nuclei antibody labeling The number of hNuclei+cells was larger in Tg2576 mice treated with (+)-phenserinethan in those receiving saline (43 increase 119901 lt 005 Dunnrsquostest) while there were no differences in numbers of hNuclei+cells between JN403- and saline-treated mice (Figure 3(a))Costaining hNuclei+ cells with MAP2 or GFAP markers for
Neural Plasticity 7
Proliferation Maturation Maturation with dendritic arborization
(a)
50
40
30
20
10
0SHAM + SAL
lowast
Num
ber o
f DCX
+ce
lls in
den
tate
gyr
us
+ SALhNSC + JNhNSC + PHENhNSC
(b)
SHAM + SAL
lowast
10
15
20
25
0
5
Num
ber o
f DCX
+ce
lls w
ith d
endr
ites
+ SALhNSC + JNhNSC + PHENhNSC
(c)
SHAM + SAL0
5
1
2
3
4
Den
driti
c bra
nche
sD
CX+
cells
with
den
drite
s
+ SALhNSC + JNhNSC + PHENhNSC
(d)
0
20
0 10 20 30 40
40
60
80
minus20
Prob
eΔla
tenc
y
r = minus061lowastlowastp lt 001
Number of DCX+ cells with dendrites
(e)
SHAM + SAL + SALhNSC
(f)
SHAM + SAL + SALhNSC
(g)
Figure 2 hNSC transplantation increases the number and the maturation of new neurons in the dentate gyrus (a) Doublecortin- (DCX-)labeled cells in the dentate gyrus (DG) of Tg2576micewere characterized according to their numbers and the level ofmaturation and dendriticarborization (b)The total number of DCX+ cells (c) the number of DCX+ cells possessing dendrites and (d) the average number of dendriticbranches on DCX+ cells in the DG inmice receiving SHAM (vehicle only) transplantation and saline (SHAM+ SAL) or mice receiving hNSCtransplantation and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (e) Correlation between the number ofDCX+ cells with dendrites and the difference in time taken to find the position of the platform (Δ latency follow-up probe test values minusbaseline acquisition values) in theMorris watermaze task in groups of Tg2576mice treatedwith hNSC+ SAL hNSC+ JN and hNSC+PHEN(119903 = minus061 lowastlowast119901 lt 001 119899 = 18) Representative images of immunostaining with DCX+ cells in the DG of TG2576 mice at (f) 10x and (g) 20xmagnification lowast119901 lt 005 compared to SHAM + SAL 119901 lt 005 and
119901 lt 001 compared to hNSC + SALThe data are expressed as means plusmnSEM
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 7
Proliferation Maturation Maturation with dendritic arborization
(a)
50
40
30
20
10
0SHAM + SAL
lowast
Num
ber o
f DCX
+ce
lls in
den
tate
gyr
us
+ SALhNSC + JNhNSC + PHENhNSC
(b)
SHAM + SAL
lowast
10
15
20
25
0
5
Num
ber o
f DCX
+ce
lls w
ith d
endr
ites
+ SALhNSC + JNhNSC + PHENhNSC
(c)
SHAM + SAL0
5
1
2
3
4
Den
driti
c bra
nche
sD
CX+
cells
with
den
drite
s
+ SALhNSC + JNhNSC + PHENhNSC
(d)
0
20
0 10 20 30 40
40
60
80
minus20
Prob
eΔla
tenc
y
r = minus061lowastlowastp lt 001
Number of DCX+ cells with dendrites
(e)
SHAM + SAL + SALhNSC
(f)
SHAM + SAL + SALhNSC
(g)
Figure 2 hNSC transplantation increases the number and the maturation of new neurons in the dentate gyrus (a) Doublecortin- (DCX-)labeled cells in the dentate gyrus (DG) of Tg2576micewere characterized according to their numbers and the level ofmaturation and dendriticarborization (b)The total number of DCX+ cells (c) the number of DCX+ cells possessing dendrites and (d) the average number of dendriticbranches on DCX+ cells in the DG inmice receiving SHAM (vehicle only) transplantation and saline (SHAM+ SAL) or mice receiving hNSCtransplantation and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (e) Correlation between the number ofDCX+ cells with dendrites and the difference in time taken to find the position of the platform (Δ latency follow-up probe test values minusbaseline acquisition values) in theMorris watermaze task in groups of Tg2576mice treatedwith hNSC+ SAL hNSC+ JN and hNSC+PHEN(119903 = minus061 lowastlowast119901 lt 001 119899 = 18) Representative images of immunostaining with DCX+ cells in the DG of TG2576 mice at (f) 10x and (g) 20xmagnification lowast119901 lt 005 compared to SHAM + SAL 119901 lt 005 and
119901 lt 001 compared to hNSC + SALThe data are expressed as means plusmnSEM
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
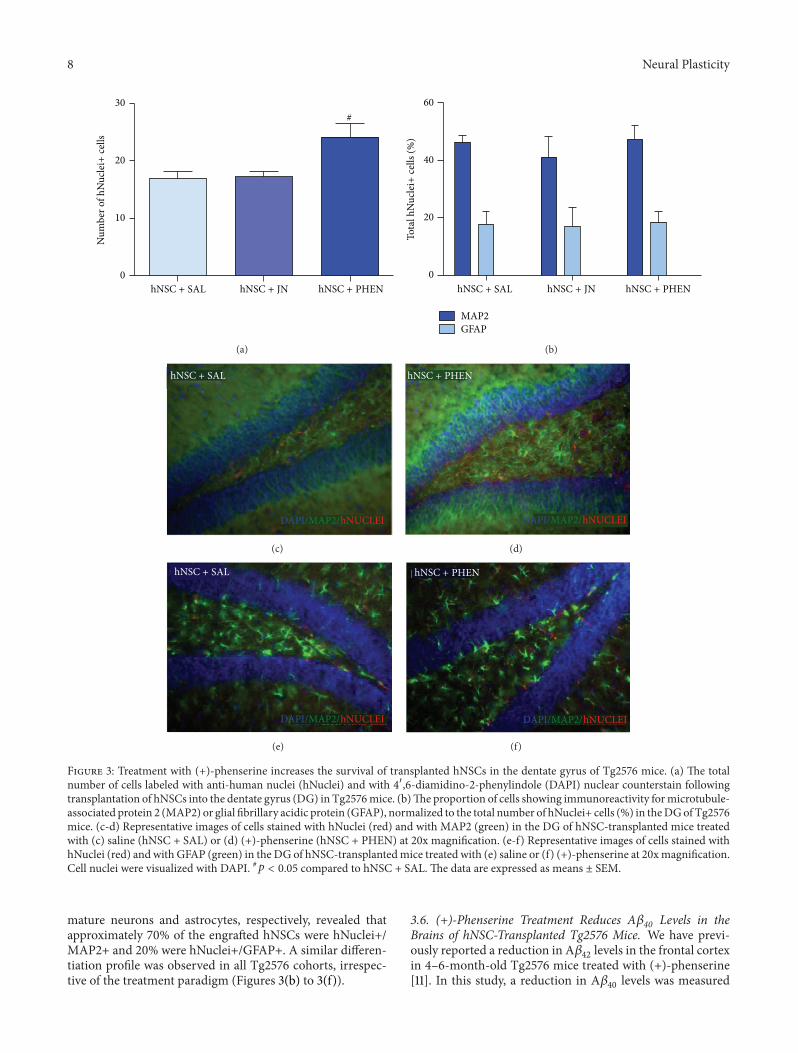
8 Neural Plasticity
10
20
30
0
Num
ber o
f hN
ucle
i+ce
lls
+ SALhNSC + JNhNSC + PHENhNSC
(a)
20
40
60
0
MAP2GFAP
Tota
l hN
ucle
i+ce
lls (
)
+ SALhNSC + JNhNSC + PHENhNSC
(b)
DAPIMAP2hNUCLEI
+ SALhNSC
(c)
DAPIMAP2hNUCLEI
+ PHENhNSC
(d)
DAPIMAP2hNUCLEI
+ SALhNSC
(e)
DAPIMAP2hNUCLEI
+ PHENhNSC
(f)
Figure 3 Treatment with (+)-phenserine increases the survival of transplanted hNSCs in the dentate gyrus of Tg2576 mice (a) The totalnumber of cells labeled with anti-human nuclei (hNuclei) and with 410158406-diamidino-2-phenylindole (DAPI) nuclear counterstain followingtransplantation of hNSCs into the dentate gyrus (DG) in Tg2576mice (b)The proportion of cells showing immunoreactivity formicrotubule-associated protein 2 (MAP2) or glial fibrillary acidic protein (GFAP) normalized to the total number of hNuclei+ cells () in theDGofTg2576mice (c-d) Representative images of cells stained with hNuclei (red) and with MAP2 (green) in the DG of hNSC-transplanted mice treatedwith (c) saline (hNSC + SAL) or (d) (+)-phenserine (hNSC + PHEN) at 20x magnification (e-f) Representative images of cells stained withhNuclei (red) and with GFAP (green) in the DG of hNSC-transplantedmice treated with (e) saline or (f) (+)-phenserine at 20xmagnificationCell nuclei were visualized with DAPI 119901 lt 005 compared to hNSC + SAL The data are expressed as means plusmn SEM
mature neurons and astrocytes respectively revealed thatapproximately 70 of the engrafted hNSCs were hNuclei+MAP2+ and 20 were hNuclei+GFAP+ A similar differen-tiation profile was observed in all Tg2576 cohorts irrespec-tive of the treatment paradigm (Figures 3(b) to 3(f))
36 (+)-Phenserine Treatment Reduces A12057340 Levels in theBrains of hNSC-Transplanted Tg2576 Mice We have previ-ously reported a reduction in A120573
42
levels in the frontal cortexin 4ndash6-month-old Tg2576 mice treated with (+)-phenserine[11] In this study a reduction in A120573
40
levels was measured
Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 9
SHAM + SAL
lowast
200
150
100
50
0
1205727 nAChR positive1205727 nAChR negative
Num
ber o
f GFA
P+ce
lls p
er fi
eld
+ SALhNSC + JNhNSC + PHENhNSC
(a)
+ SALhNSC + JN403hNSC
(b) (c)
Figure 4 Treatment with the cholinergic 1205727 nicotinic receptor agonist JN403 decreases the number of 1205727 nicotinic receptor-expressingastrocytes in the dentate gyrus (a) The number of 1205727 nicotinic receptorglial fibrillary acidic protein-positive (nAChR+GFAP+) cells andthe number of 1205727 nAChR-negativeGFAP+ cells in the dentate gyrus of Tg2576 mice that received SHAM (vehicle) transplants and saline(SHAM + SAL) or hNSC transplants and saline (hNSC + SAL) JN403 (hNSC + JN) or (+)-phenserine (hNSC + PHEN) (b) Representativeimages of 1205727 nAChR+GFAP+ cells (black) and 1205727 nAChR-negativeGFAP+ cells (brown) in hNSC + SAL and hNSC + JN mice (c)Representative image of 1205727 nAChR+GFAP+ cells (black) in the needle track after hippocampal injection lowast119901 lt 005 compared to SHAM +SAL The data are expressed as means plusmn SEM
in the frontal cortices and hippocampi of hNSC-transplan-ted Tg2576 mice treated with (+)-phenserine but not in thecohort of mice treated with JN403 or saline In comparison toSHAM-transplanted saline-treatedmice a significant reduc-tion in A120573
40
levels but not in A12057342
levels was measured inthe frontal cortex (29 reduction 119901 lt 005 Dunnrsquos testFigure S3A) and hippocampus (42 reduction 119901 lt 005Dunnrsquos test Figure S3B) of hNSC-transplanted mice trea-ted with (+)-phenserine No significant changes in A120573
40
and in A12057342
levels were observed in saline- or JN403-treated hNSC-transplanted Tg2576 mice versus SHAM-tra-nsplanted saline-treated mice
37 Association between Endogenous Neurogenesis and thePresence of 1205727 nAChR-Expressing Astrocytes in the DentateGyrus of hNSC-Transplanted Tg2576 Mice Reactive astrogli-osis and numerous 1205727 nAChR-expressing GFAP+ astrocyteswere detected in the hippocampi of hNSC-transplanted
Tg2576 mice Significant reductions were observed inthe total numbers of GFAP+ astrocytes (30 reduction 119901 lt005) and even greater decrease was seen in the number ofastrocytes expressing 1205727 nAChRs (39-40 reduction 119901 lt005 Dunnrsquos test) in the DG of hNSC-transplanted Tg2576mice treated with JN403 compared with mice receivingSHAM or hNSC transplants and saline (Figures 4(a) and4(b)) Treatment with (+)-phenserine did not alter the num-ber of 1205727 nAChR-expressing astrocytes in the hNSC-transplanted DG compared to saline treatment in SHAM- orhNSC-transplanted mice (Figure 4(a)) A large number of 1205727nAChR-expressing astrocyteswere observed in the area of theDG surrounding the injection site (Figure 4(c)) suggestingthe involvement of 1205727 nAChR-expressing astrocytes in repairand regenerative processes in the brain In support of thisa significant positive correlation was found between 1205727nAChR-expressing astrocytes and number of DCX+ cellswith dendrites (119903 = 070 119901 lt 001 Dunnrsquos test Figure 5) inhNSC-transplanted Tg2576 mice
10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

10 Neural Plasticity
0 10 20 30 40
r = 070lowastlowastp lt 001
250
200
150
100
50
0
1205727
nACh
RminusG
FAP+
cells
fiel
d
Number of DCX+ cells with dendrites
Figure 5 The number of newly generated neurons correlates withthe proportion of 1205727 nicotinic receptor-expressing astrocytes in thedentate gyrus of hNSC-transplanted Tg2576 mice The number ofmaturing Doublecortin- (DCX+-) labeled cells showed a significantpositive correlation with the number of 1205727 nicotinic receptorglialfibrillary acidic protein-positive (nAChR+GFAP+) cells in thedentate gyrus of Tg2576 mice (119903 = 070 lowastlowast119901 lt 001 119899 = 13) Thedata are expressed as the mean value for each parameter in thehNSC-transplanted saline-treated mice (hNSC + SAL) and thehNSC-transplanted JN403-treated mice (hNSC + JN)
4 Discussion
41 hNSCTransplantationEnhances EndogenousNeurogenesisand Prevents Memory Deterioration in Tg2576 Mice Trans-plantation of hNSCs increased endogenous neurogenesis inthe DG which correlated with improved hippocampal-depe-ndent memory performance The hNSC-transplanted micehad a memory performance comparable to baseline valueswhich might indicate stabilization of the disease whereas theother treatment groups continued to deteriorate It could beargued that extensive training (baseline and follow-up) mayimprove the performance However the memory deteriora-tion could be a consequence of the progression of the diseaseAlthough informative the behavioral paradigm employed inthis study has its limitations Reversal learning in the MWM(the ability of the mice to find a new location of the hiddenplatform) has recently gained preference for the assessment ofhippocampal-dependent memory in relation to neurogenesis[21] However subjecting the study mice to further assess-ments after they had undergone baseline follow-up andprobe trial sessions would have been too exhausting andmostlikely had an impact on their performance Another limita-tion in the behavioral testing is that we did not study visualdiscrimination learning A solution to this would have beento include an initial visual test to assess eyesight Despite theselimitations the findings presented here are in line with thoseof a previous study that showed that intrahippocampal trans-plantation of murine NSCs resulted in an increased secretionof brain-derived neurotrophic factor (BDNF) and synapticdensity as well as improved cognition in aged 3xTg-ADmiceharboring a heavy A120573 plaque burden and neurofibrillary
tangle pathology [22] Several studies have reported an imbal-ance between the levels of precursor and mature BDNF (theformer possessing proapoptotic features) as well as impairedBDNF signaling [23ndash27] Furthermore a decline in BDNFlevels in the brain occurs with disease progression and is cor-related with cognitive decline [25 28] Despite the promisingpotential for using BDNF as a biomarker to assess ther-apeutic effects quantitative measurements of BDNF levelswith ELISA-based techniques are difficult to perform asthe current available antibodies lack sufficient sensitivity todistinguish mature forms of BDNF from pro-BDNF [29 30]We hypothesize that impaired neurotrophic support in theAD brain may be one of the underlying mechanisms throughwhich endogenous neurogenesis is compromised prevent-ing maturation of newborn neurons Our current findingsindicate that engraftment of hNSCs in Tg2576 mouse brainsprovides neurotrophic support to existing neurons in theneural circuitry as well as to endogenous stem cells residingin the DG
42 (+)-Phenserine Treatment Decreases A120573 Levels and Enha-nces Graft Survival but Mitigates the hNSC-Mediated Bene-ficial Effects on Endogenous Neurogenesis and Cognition inTg2576 Mice (+)-phenserine treatment combined withhNSC transplantation in Tg2576 mice lowered both corticaland hippocampal A120573
40
levels and increased the survival oftransplanted hNSCs in the hippocampi However this treat-ment combination failed to improvememory in Tg2576miceSurprisingly we found that the administration of (+)-phen-serine prevented the hNSC-induced increase in endogenousneurogenesis in Tg2576 mice We have earlier demonstratedthat (+)-phenserine augments neuronal differentiation oftransplanted hNSCs in the brains of AD APP23 transgenicmice [4] We have also reported that (+)-phenserine in addi-tion to its A120573 lowering action exerts prosurvival effects onprogenitor cells [10] and increases the dendritic arborizationof hippocampal neurons [11] Hence the body of evidencethat (+)-phenserine possesses neurotrophic characteristicsrules out the possibility that (+)-phenserine per se inhibitedneurogenesis but rather antagonized the trophic effects ofhNSC transplantation
The mitogen-activated protein kinases (MAPK) and pro-tein kinase C signaling pathways have been identified aspotential mediators of the neurotrophic actions of (+)-phen-serine These pathways are involved in a diverse repertoire ofbiological events including proliferation differentiationmet-abolism motility survival and apoptosis [10] Both MAPKand the downstream transcription factorCREB are importantregulators of BDNF [31 32] and share common signalingpathways with other key regulators of adult neurogenesis andNSCs such as fibroblast growth factor-2 insulin-like growthfactor-1 and vascular endothelial growth factor [33] Hencea possible explanation for the unexpected and somewhatambiguous observations with (+)-phenserine in the presentstudy could be that the simultaneous combination of trans-plantation and chronic (+)-phenserine administration inter-feres with hNSC signaling cascades which could potentiallylead to an inhibition of the neurotrophic effects of these cells
Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 11
on the brain environment that normally supports neurogen-esis in Tg2576 mice Identifying the underlying mechanismswarrants future investigation yet our current findings raisethe question of whether the dosage regimen or interval of(+)-phenserine administration in combination with hNSCsneeds to be carefully considered in lieu of the different neu-rodegenerative stages in AD Although hNSC transplantationinduces beneficial effects impaired graft survival as a resultof the accumulation of pathological proteins in the AD brainshould be considered as has previously been demonstrated ingrafted neurons in Parkinson patients [34] Hence to sustainthe efficacy of this intervention we propose that combinationtreatment with drugs that target the different pathologicalprocesses without interfering with the stem cell-mediatedneurogenic effects should be administered at specific diseasestages
43 The 1205727 nAChR Agonist JN403 Impairs Neurogenesisand Cognition Which Correlates with Downregulation of 1205727nAChR-Expressing Astrocytes Neurotrophic effects for (+)-phenserine have been observed inmultiple studies [10 16 17]whereas similar effects of JN403 remained to be explored Tothis end we performed initial in vitro experiments to investi-gate the effects of JN403 on neuroprotection and on neu-ronal differentiation JN403 increased the survival of hNSC-derived neurons exposed to brain extract containing variousA120573 species in line with the characteristic neuroprotectiveeffects induced by treatment with 1205727 nAChR agonists [35ndash38]
Reactive gliosis is a complex and multistage process andplays an important role in the progression and resolution ofbrain pathology Astrocytes are a diverse group of cells withmany subclasses that carry out a wide range of functions inthe CNS including the homeostatic response limiting dam-age to the brain through formation of a glial scar and removalof neurotoxic pathogens and assist in postlesion regenerationof the neural networks [39] It has become amatter of growinginterest and ongoing investigation whether all astrocytes oronly certain types of astrocytes in the brain are involved in theregulation of cognitive processes Recent in vivo observationswith positron emission tomography (PET) imaging suggestthat astroglial reactivity correlates with the preservation ofcognitive function in patients with mild cognitive impair-ment and prodromal AD [40] The 1205727 nAChR-expressingastrocytes are reported to be associated with the pathophys-iological processes in AD as increased numbers have beenfound surrounding 120573-amyloid plaques in postmortem brainsof sporadic AD and APP Swedish mutation carriers [39] Inthe current study we detected a large number of 1205727 nAChR-expressing astrocytes surrounding the injection site in theDG of Tg2576 mice indicative of a tissue response to the celltransplantation In addition a significant positive correlationwas found between the numbers of 1205727 nAChR-expressingastrocytes and the numbers of newly generated neurons in theDGThese data suggest a role for this subclass of astrocytes inrepair and scarring processes in the brain
Inhibitory effects of chronic JN403 treatment on hNSC-induced neurogenesis and cognition were observed inTg2576 mice JN403 induced a downregulation in both
the total number of astrocytes and the number of 1205727 nAChR-expressing astrocytes in the DG It is possible that the dura-tion of treatment with JN403 led to a suppression of thenormal physiological and neuroprotective functions of 1205727nAChR-expressing astrocytes
Functional 1205727 nAChRs on astrocytes in hippocampal sli-ces from rat brain have previously been demonstrated [41]Considering the high Ca2+-permeability of 1205727 nAChRs acti-vation of these receptors would thus lead to increases in intra-cellular calcium levels in hippocampal 1205727 nAChR-express-ing astrocytes to an extent that may significantly impactsynaptic transmission and plasticity mechanisms Howeverthe 1205727 nAChRs also undergo rapid desensitization and thepharmacodynamic effects and impact of nAChR agonist sti-mulation versus desensitization on cognition are still poorlyunderstood The 1205727 nAChRs on neurons and astrocytes mayhave different roles and may be differentially affected duringthe various stages of disease Upregulation of neuronal 1205727nAChR expression with A120573 accumulation is reported in theearly stages of AD [42 43] as well as in Tg2576 mice [44]andmost likely reflects a compensatory response to maintainmemory function As the disease progresses neuronal 1205727nAChRsmay either decrease [45] or increase where hyperex-citable neuronal 1205727 nAChRs further exacerbate neurotoxicityand neurodegeneration [46] Reconciling the findings fromthese studies would therefore suggest that the modulation of1205727 nAChRs present on either neuronal or nonneuronal cellscould induce either protective or toxic effects depending onthe mode of agonist exposure and on the functional status ofthese receptors during the disease course The expression of1205727 nAChRs on neurons was not quantified in our studyand we cannot rule out the possibility that JN403 may haveexerted an antagonistic function on hippocampal-dependentmemory functions through mechanisms that also involvedneuronal 1205727 nAChRs The 1205727 nAChRs are present early dur-ing development and these receptors have also been detectedon human stem cells [47ndash49] where they likely mediateeffects of cholinergic signaling on stem cell survivalapopto-sis proliferation differentiation andmaturation An alterna-tive mechanism is that JN403 interacts with the 1205727 nAChRsexpressed on transplanted hNSCs with downstream conse-quences on 1205727 nAChR signaling pathways These changescould in turn result in a modulation of the proposed trophicactions of the grafted cells and consequently affect endoge-nous neurogenesis and cognition
The potential anti-inflammatory mechanisms and regu-lation of inflammatory processes in the brain caused by thestimulation 1205727 nAChRs with selective agonists should alsobe considered Although these mechanisms are complex andnot clearly understood it is conceivable that JN403 stimu-lation of the 1205727 nAChRs on astrocytes modulates the anti-inflammatory response where it is important that a certaininnate basal level of 1205727 nAChR-expressing GFAP+ astrocytesis maintained in the hippocampus In support of the lattera recent study using a rodent model of Parkinsonrsquos diseaseshowed that targeting 1205727 nAChRs induces anti-inflammatoryeffects via inhibition of astrocyte activation [38]
In conclusion the present study reveals that hNSC trans-plantation stimulates regenerative processes in the brain
12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

12 Neural Plasticity
Furthermore the findings indicate that hNSC transplantationcan attenuate memory deterioration In contrast coadmin-istration with either drug (+)-phenserine or JN403 inhibitsthe beneficial effects of hNSC infusion The enhancement ofendogenous neurogenesis in mice following transplantationshows a positive correlation with 1205727 nAChR-expressing ast-rocytes in the DGTherapies that stimulate endogenous neu-rogenesis in AD brain could thus contribute to improvementof cognitive function
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Thework was supported by grants from the Swedish MedicalResearch Council (Project no 05817) the Stockholm CountyCouncil-Karolinska Institutet (ALF Grant) the KarolinskaInstitutet Strategic Neuroscience Program Swedish BrainPower the Brain Foundation the European Unionrsquos Sev-enth Framework Programme (FP72007ndash2013) under GrantAgreement no HEALTH-F2-2011-278850 (INMiND) Gunand Bertil Stohnersquos Foundation the Foundation for Old Ser-vantsMagnus Bergvallrsquos FoundationTheDementia Associa-tionTheLarsHiertaMemorial Foundation theOlle EngkvistByggmastare Foundation Karolinska Institutet Foundationsand the Alzheimer Association Sweden Anna M Lilja wassupported by funding for new doctoral students at KarolinskaInstitutet (KID) a grant from Erik and Edith FernstromrsquosFoundation and a stipend from Gun and Bertil StohnersquosFoundation The authors thank Dr Therese Pham (Karolin-ska Institutet) for assistance with the design of the behavioralstudies and Stephanie Shaia for quantification of DCX stain-ing JN403 was supplied as a kind gift fromNovartis and (+)-phenserine as a kind gift from Dr Nigel Greig (NIHNIAUSA)
References
[1] D Paterson and A Nordberg ldquoNeuronal nicotinic receptors inthe human brainrdquo Progress in Neurobiology vol 61 no 1 pp75ndash111 2000
[2] P Taupin ldquoNeurogenesis and Alzheimerrsquos diseaserdquo Drug TargetInsights vol 1 pp 1ndash4 2006
[3] K L Spalding O Bergmann K Alkass et al ldquoDynamics ofhippocampal neurogenesis in adult humansrdquo Cell vol 153 no6 pp 1219ndash1227 2013
[4] AMarutle M OhmitsuM Nilbratt N H Greig A NordbergandK Sugaya ldquoModulation of human neural stem cell differen-tiation in Alzheimer (APP23) transgenic mice by phenserinerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 104 no 30 pp 12506ndash12511 2007
[5] O Lazarov and R A Marr ldquoNeurogenesis and Alzheimerrsquosdisease at the crossroadsrdquo Experimental Neurology vol 223 no2 pp 267ndash281 2010
[6] Y Mu and F H Gage ldquoAdult hippocampal neurogenesis and itsrole in Alzheimerrsquos diseaserdquo Molecular Neurodegeneration vol6 no 1 article 85 2011
[7] J J Rodrıguez and A Verkhratsky ldquoNeurogenesis in Alz-heimerrsquos diseaserdquo Journal of Anatomy vol 219 no 1 pp 78ndash892011
[8] O Einstein and T Ben-Hur ldquoThe changing face of neural stemcell therapy in neurologic diseasesrdquo Archives of Neurology vol65 no 4 pp 452ndash456 2008
[9] K Jin L Xie X O Mao and D A Greenberg ldquoAlzheimerrsquosdisease drugs promote neurogenesisrdquo Brain Research vol 1085no 1 pp 183ndash188 2006
[10] A M Lilja Y Luo Q-S Yu et al ldquoNeurotrophic and neuropro-tective actions of (minus)- and (+)-phenserine candidate drugs forAlzheimerrsquos diseaserdquo PLoS ONE vol 8 no 1 Article ID e548872013
[11] A M Lilja J Rojdner T Mustafiz et al ldquoAge-dependent neu-roplasticity mechanisms in Alzheimer Tg2576 mice followingmodulation of brain amyloid-beta levelsrdquo PLoS ONE vol 8 no3 Article ID e58752 2013
[12] C U Lithner M M Hedberg and A Nordberg ldquoTransgenicmice as a model for Alzheimerrsquos diseaserdquo Current AlzheimerResearch vol 8 no 8 pp 818ndash831 2011
[13] T Mustafiz E Portelius M K Gustavsson et al ldquoCharacteri-zation of the brain 120573-amyloid isoform pattern at different agesof Tg2576 micerdquo Neurodegenerative Diseases vol 8 no 5 pp352ndash363 2011
[14] S Stewart F Cacucci and C Lever ldquoWhich memory task formymouseA systematic review of spatialmemory performancein the Tg2576 Alzheimerrsquos mouse modelrdquo Journal of AlzheimerrsquosDisease vol 26 no 1 pp 105ndash126 2011
[15] C Unger M M Svedberg W-F Yu M M Hedberg andA Nordberg ldquoEffect of subchronic treatment of memantinegalantamine and nicotine in the brain of Tg2576 (APPswe)transgenic micerdquo Journal of Pharmacology and ExperimentalTherapeutics vol 317 no 1 pp 30ndash36 2006
[16] D K Lahiri D Chen B Maloney et al ldquoThe experimentalAlzheimerrsquos disease drug posiphen [(+)-phenserine] lowersamyloid-beta peptide levels in cell culture and micerdquo Journal ofPharmacology and ExperimentalTherapeutics vol 320 no 1 pp386ndash396 2007
[17] M L Maccecchini M Y Chang C Pan V John H Zetterbergand N H Greig ldquoPosiphen as a candidate drug to lower CSFamyloid precursor protein amyloid-120573 peptide and 120591 levels tar-get engagement tolerabilityand pharmacokinetics in humansrdquoJournal of Neurology Neurosurgery and Psychiatry vol 83 no 9pp 894ndash902 2012
[18] D Feuerbach K Lingenhoehl H-R Olpe et al ldquoThe selectivenicotinic acetylcholine receptor 1205727 agonist JN403 is active inanimal models of cognition sensory gating epilepsy and painrdquoNeuropharmacology vol 56 no 1 pp 254ndash263 2009
[19] D Feuerbach J Nozulak K Lingenhoehl K McAllister andD Hoyer ldquoJN403 in vitro characterization of a novel nicotinicacetylcholine receptor 1205727 selective agonistrdquo Neuroscience Let-ters vol 416 no 1 pp 61ndash65 2007
[20] C L Brannen and K Sugaya ldquoIn vitro differentiation of mul-tipotent human neural progenitors in serum-free mediumrdquoNeuroReport vol 11 no 5 pp 1123ndash1128 2000
[21] A Garthe Z Huang L Kaczmarek R K Filipkowski and GKempermann ldquoNot all water mazes are created equal cyclin D2
Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Neural Plasticity 13
knockout mice with constitutively suppressed adult hippocam-pal neurogenesis do show specific spatial learning deficitsrdquoGenes Brain and Behavior vol 13 no 4 pp 357ndash364 2014
[22] M Blurton-Jones M Kitazawa HMartinez-Coria et al ldquoNeu-ral stem cells improve cognition via BDNF in a transgenicmodel of Alzheimer diseaserdquo Proceedings of the National Acad-emy of Sciences of the United States of America vol 106 no 32pp 13594ndash13599 2009
[23] M A Bruno W C Leon G Fragoso W E Mushynski GAlmazan and A C Cuello ldquoAmyloid 120573-induced nerve growthfactor dysmetabolism in Alzheimer diseaserdquo Journal of Neu-ropathology amp Experimental Neurology vol 68 no 8 pp 857ndash869 2009
[24] S Capsoni and A Cattaneo ldquoOn the molecular basis linkingNerve Growth Factor (NGF) to Alzheimerrsquos diseaserdquo Cellularand molecular neurobiology vol 26 no 4ndash6 pp 619ndash633 2006
[25] C Laske E Stransky T Leyhe et al ldquoStage-dependent BDNFserum concentrations in Alzheimerrsquos diseaserdquo Journal of NeuralTransmission vol 113 no 9 pp 1217ndash1224 2006
[26] B Michalski and M Fahnestock ldquoPro-brain-derived neu-rotrophic factor is decreased in parietal cortex in AlzheimerrsquosdiseaserdquoMolecular Brain Research vol 111 no 1-2 pp 148ndash1542003
[27] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[28] I M Ethell and D W Ethell ldquoMatrix metalloproteinases inbrain development and remodeling synaptic functions andtargetsrdquo Journal of Neuroscience Research vol 85 no 13 pp2813ndash2823 2007
[29] H Koshimizu K Kiyosue T Hara et al ldquoMultiple functions ofprecursor BDNF toCNSneurons negative regulation of neuritegrowth spine formation and cell survivalrdquoMolecular Brain vol2 no 1 article 27 2009
[30] R Lee P Kermani K K Teng and B L Hempstead ldquoRegula-tion of cell survival by secreted proneurotrophinsrdquo Science vol294 no 5548 pp 1945ndash1948 2001
[31] A E Autry and L M Monteggia ldquoBrain-derived neurotro-phic factor and neuropsychiatric disordersrdquo PharmacologicalReviews vol 64 no 2 pp 238ndash258 2012
[32] Y Lu K Christian and B Lu ldquoBDNF a key regulator forprotein synthesis-dependent LTP and long-term memoryrdquoNeurobiology of Learning and Memory vol 89 no 3 pp 312ndash323 2008
[33] R Faigle and H Song ldquoSignaling mechanisms regulating adultneural stem cells and neurogenesisrdquo Biochimica et BiophysicaActamdashGeneral Subjects vol 1830 no 2 pp 2435ndash2448 2013
[34] J-Y Li E Englund J L Holton et al ldquoLewy bodies in graftedneurons in subjects with Parkinsonrsquos disease suggest host-to-graft disease propagationrdquo Nature Medicine vol 14 no 5 pp501ndash503 2008
[35] T Kihara S Shimohama H Sawada et al ldquoNicotinic receptorstimulation protects neurons against beta-amyloid toxicityrdquoAnnals of Neurology vol 42 no 2 pp 159ndash163 1997
[36] A M Lilja O Porras E Storelli A Nordberg and A MarutleldquoFunctional interactions of fibrillar and oligomeric amyloid-120573with alpha7 nicotinic receptors in alzheimerrsquos diseaserdquo Journalof Alzheimerrsquos Disease vol 23 no 2 pp 335ndash347 2011
[37] Q Liu and B Zhao ldquoNicotine attenuates beta-amyloid peptide-induced neurotoxicity free radical and calcium accumulationin hippocampal neuronal culturesrdquo British Journal of Pharma-cology vol 141 no 4 pp 746ndash754 2004
[38] Y Liu J Hu J Wu et al ldquo1205727 nicotinic acetylcholine receptor-mediated neuroprotection against dopaminergic neuron loss inan MPTP mouse model via inhibition of astrocyte activationrdquoJournal of Neuroinflammation vol 9 article 98 2012
[39] A Verkhratsky M Olabarria H N Noristani C-Y Yeh and JJ Rodriguez ldquoAstrocytes in Alzheimerrsquos Diseaserdquo Neurothera-peutics vol 7 no 4 pp 399ndash412 2010
[40] S F Carter M Scholl O Almkvist et al ldquoEvidence forastrocytosis in prodromal alzheimer disease provided by 11C-deuterium-L-deprenyl a multitracer PET paradigm combining11C-Pittsburgh compound B and 18F-FDGrdquo Journal of NuclearMedicine vol 53 no 1 pp 37ndash46 2012
[41] J-X Shen and J L Yakel ldquoFunctional 1205727 nicotinic ACh recep-tors on astrocytes in rat hippocampal CA1 slicesrdquo Journal ofMolecular Neuroscience vol 48 no 1 pp 14ndash21 2012
[42] S E Counts B He S Che et al ldquo1205727 nicotinic receptor up-regulation in cholinergic basal forebrain neurons in Alzheimerdiseaserdquo Archives of Neurology vol 64 no 12 pp 1771ndash17762007
[43] MD Ikonomovic LWecker E E Abrahamson et al ldquoCortical1205727 nicotinic acetylcholine receptor and120573-amyloid levels in earlyAlzheimer diseaserdquo Archives of Neurology vol 66 no 5 pp646ndash651 2009
[44] I Bednar D Paterson A Marutle et al ldquoSelective nicotinicreceptor consequences in APP
119878119882119864
transgenic micerdquo Molecularand Cellular Neuroscience vol 20 no 2 pp 354ndash365 2002
[45] W-F Yu Z-Z Guan N Bogdanovic and A Nordberg ldquoHighselective expression of alpha7 nicotinic receptors on astrocytesin the brains of patients with sporadic Alzheimerrsquos disease andpatients carrying Swedish APP 670671 mutation a possibleassociation with neuritic plaquesrdquo Experimental Neurology vol192 no 1 pp 215ndash225 2005
[46] Q Liu X Xie R J Lukas P A StJohn and J Wu ldquoA novelnicotinic mechanism underlies 120573-amyloid-induced neuronalhyperexcitationrdquo Journal of Neuroscience vol 33 no 17 pp7253ndash7263 2013
[47] MNilbratt O Porras AMarutleOHovatta andANordbergldquoNeurotrophic factors promote cholinergic differentiation inhuman embryonic stem cell-derived neuronsrdquo Journal of Cellu-lar and Molecular Medicine vol 14 no 6 pp 1476ndash1484 2010
[48] L Wicklund R N Leao A-M Stromberg et al ldquoBeta-amyloid1-42 oligomers impair function of human embryonic stem cell-derived forebrain cholinergic neuronsrdquo PLoS ONE vol 5 no12 Article ID e15600 2010
[49] J A Court S Lloyd M Johnson et al ldquoNicotinic and mus-carinic cholinergic receptor binding in the human hippocam-pal formation during development and agingrdquo DevelopmentalBrain Research vol 101 no 1-2 pp 93ndash105 1997
Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014

Submit your manuscripts athttpwwwhindawicom
Neurology Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Alzheimerrsquos DiseaseHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentSchizophrenia
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neural Plasticity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAutism
Sleep DisordersHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neuroscience Journal
Epilepsy Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Psychiatry Journal
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
Depression Research and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Brain ScienceInternational Journal of
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Neurodegenerative Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Cardiovascular Psychiatry and NeurologyHindawi Publishing Corporationhttpwwwhindawicom Volume 2014