morphological delineation and distribution patterns of four … · morphological delineation and...
TRANSCRIPT

Morphological delineation and distribution patterns of fournewly described species within the Synura petersenii speciescomplex (Chrysophyceae, Stramenopiles)
PAVEL ŠKALOUD, MAGDA ŠKALOUDOVÁ, ANNA PROCHÁZKOVÁ AND YVONNE NĚMCOVÁ
Department of Botany, Charles University in Prague, Benátská 2, CZ-12801, Prague, Czech Republic
(Received 18 October 2013; revised 22 January 2014; accepted 28 January 2014)
The Synura petersenii species complex represents a common, cosmopolitan and highly diverse taxon of autotrophic freshwaterflagellates. In this paper, we describe and characterize four new species (S. borealis, S. heteropora, S. hibernica and S. laticarina)that have been identified during our extensive sampling of freshwater habitats in 15 European countries. Morphometric analysesof siliceous scales led to the significant phenotypic differentiation of all four newly described species, and their separation fromother related species of the S. petersenii complex. Two of these newly described species (S. hibernica and S. borealis) can beclearly distinguished by characteristic large colonies consisting of elongated, lanceolate-shaped cells. Development of stronglyelongated, narrow cells in S. hibernica could be explained by the adaptation of this species to oligotrophic conditions. Thoughmorphologically distinct, S. borealis possesses an exceptionally high degree of genetic diversity, possibly indicating recentspeciation and evolutionary diversification within this taxon. Three of the four newly described species exhibit restrictedbiogeographic distribution. The evolutionarily related S. borealis and S. laticarina occur only in Northern Europe, and seem to beadapted to colder areas. The most remarkable distribution pattern was observed for S. hibernica, which has a geographicdistribution that is restricted to western Ireland.
Key words: biogeography, cox1, cryptic species, ITS rDNA, morphology, phylogeny, rbcL, speciation, Synura, taxonomy
Introduction
The order Synurales (Chrysophyceae, Stramenopiles)contains several genera of scale-bearing flagellates thatare important components of phytoplankton commu-nities of various freshwater bodies (Kristiansen &Preisig, 2007; Škaloud et al., 2013a). The most con-spicuous genus of the order, Synura, is a colonialorganism formed by a variable number of cells joinedtogether at their posterior ends. Each cell is covered byimbricate silica scales (Wee, 1997), consisting of aperforated basal plate with various upturned or bentparts, and a secondary ornamentation, which is used todelimit the different species (Kristiansen, 1986).According to the recently published phylogeny of theSynurales, the genus Synura should be split into fivesections: Echinulatae, Peterseniae, Spinosae,Splendidae and Synura (Škaloud et al., 2013a).Taxabelonging to the section Peterseniae are well character-ized by body scales with a central keel that may end in aspine-like projection. The most widely recognized spe-cies of this section, Synura petersenii sensu lato, isconsidered to be one of the most widely distributedfreshwater chrysophytes (Kristiansen, 1975), even
causing taste and odour problems in water supplies(Nicholls & Gerrath, 1985). This species has beensubjected to various ecophysiological experiments(e.g. Saxby-Rouen et al., 1997; Kim et al., 2008;Pichrtová & Němcová, 2011).
Due to the species-specific morphology of silicascales, Synura could be supposed to have one ofthe best morphological species concepts withinprotists. Synura scales are often well preserved inbenthic sediments and used in palaeoecologicalstudies. Because many taxa are distributed withina narrow range of ecological variables (e.g. pH,temperature and nutrient concentration), investiga-tion of fossilized Synura scales is used as a tool toassess eutrophication, acidification and shifts inclimate (Smol, 1995; Smol & Cumming, 2000).In addition, the records of fossilized Synura scalesdiscovered in Eocene sediments are used to datethe diversification and evolution of the genus (Booet al., 2010; Siver, 2013).
Recent molecular phylogenetic investigationshave revealed a conflict between the traditionalmorphological species concept based on ultrastruc-ture of silica structures and the phylogenetic data.Synura petersenii sensu lato represents the most
Correspondence to: Pavel Škaloud. E-mail: [email protected]
Eur. J. Phycol. (2014), 49(2): 213–229
ISSN 0967-0262 (print)/ISSN 1469-4433 (online)/14/020213-229 © 2014 British Phycological Societyhttp://dx.doi.org/10.1080/09670262.2014.905710
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

investigated species in this respect. First, Wee et al.(2001) demonstrated the existence of two well-sup-ported S. petersenii clades with different distributionpatterns. Later, Kynčlová et al. (2010) investigated anumber of European isolates, and revealed the exis-tence of six cryptic species. Almost concurrently,Boo et al. (2010) published a multigene phylogenyof approximately 100 S. petersenii isolates, confirm-ing the high degree of hidden, species-level diversitywithin this species. A taxonomic assessment ofobserved cryptic diversity redefined the species con-cept within the S. petersenii morphotype, and recog-nized six cryptic lineages as separate species(Škaloud et al., 2012).
Phylogenetic analyses performed by Boo et al.(2010) and Škaloud et al. (2012) clearly indicate thatthere are more than six hidden species within theS. petersenii complex. To further investigate the reallevel of species diversity within this complex, weperformed extensive sampling in 15 European coun-tries, including Greenland. According to the analysesof internal transcribed spacer (ITS) rDNA sequences,58 of more than 800 investigated Synura isolatesbelonged to four novel, undescribed species. Theprincipal aim of this study was to expand our knowl-edge and understanding of the diversity in theS. petersenii complex, and to describe and character-ize all these new taxa. In addition, we performed adetailed morphological investigation of all crypticspecies, and investigated whether they could be deli-neated by the morphology of silica scales.
Materials and methods
Origin and cultivation of the investigated strains
The strains used in this study were obtained during aninvestigation of silica-scaled chrysophytes in Europe andGreenland during the period 2008–2013 (Table 1). Thematerial was usually sampled from plankton and meta-phyton of various water bodies, using a plankton net with20-μm mesh. To ensure the highest viability of Synurastrains, individual colonies were isolated on the same daythat they were sampled. The colonies were removed fromthe natural sample by micropipetting, and transferred intoseparate wells of a 96-well plate containing 400 μl ofeither MES- or HEPES-buffered DY IV liquid medium(Andersen et al., 1997). The wells were kept at 15°C(fridge bag TK 51, Ardes SpA, Ponte Nossa, Italy), underconstant illumination of 50–200 μmol m−2 s−1 providedby 6 W LED diodes (LB115A-6W-X, Yuyao LianliangElectric Appliance Co Ltd, Ningbo, China). After return-ing them to the laboratory, the strains were cultivated at15°C (cooling box C5G, Helkama Oy, Helsinki, Finland),under illumination of 40 μmol m−2 s−1 and a 16-h light :8-h dark cycle (TLD 18W/33 fluorescent lamps, Philips,Amsterdam, the Netherlands). In addition, isolated colo-nies were separately transferred to 200 μl PCR tubes andsubsequently frozen at −24°C.
DNA extraction, PCR and DNA sequencing
For DNA isolation, 100 µl of the growing culture was cen-trifuged in PCR tubes (6000 rpm for 3 min), and 30 µl ofInstaGene matrix (Bio-Rad Laboratories, Hercules, CA,USA) was added to the pellet. The solution was vortexedfor 10 s, incubated at 56°C for 30 min, and heated at 99°C for8 min. After vortexing a second time, the tubes were centri-fuged at 12000 rpm for 2 min, and the supernatant wasdirectly used as a PCR template. Three molecular markerswere amplified by PCR: nuclear ITS rDNA, chloroplast rbcLand mitochondrial coxI. The amplification of ITS rDNAwasperformed as described in Kynčlová et al. (2010), using theprimers Kn1.1 (5ʹ-CAAGGT TTC CGTAGG TGAACC-3ʹ;Wee et al., 2001) and ITS4 (5ʹ-TCC TCC GCT TAT TGATAT GC-3ʹ; White et al., 1990). Amplification of the rbcLmarker was performed according to Jo et al. (2011), using theprimers rbcL_2F (5ʹ-AAAAGTGACCGT TATGAATC-3ʹ;Daugbjerg & Andersen, 1997) and rbcL_R3 (5ʹ-GTA ATATCT TTC TTC CATAAATCTAA-3ʹ; Jo et al., 2011). Thecox1 marker was amplified according to Boo et al. (2010),using the primers F692 (5ʹ-TTG TDT GGT CAG TTT TAATTA C-3ʹ) and R1433 (5ʹ-GGC ATA CCT GCWARA CCTAA-3ʹ; Boo et al., 2010). The PCR products were purifiedand sequenced at Macrogen Inc. in Seoul, Korea.
Phylogenetic analyses
A multiple alignment of the newly determined ITS rDNA,rbcL and coxI gene sequences and other sequences selectedfrom the GenBank/EMBL/DDBJ databases was built usingMAFFT, version 6, applying the Q-INS-i strategy (Katohet al., 2002). The sequences were selected to encompass allknown lineages within the Synura petersenii species complex(Table S1). ITS rDNA sequences were then aligned on thebasis of their rRNA secondary structure information(Kynčlová et al., 2010) with MEGA 4 (Kumar et al., 2008).The three loci were concatenated, yielding an alignment of2308 bases. The final matrix contained 51 ITS rDNA,36 rbcL and 33 coxI sequences. A suitable partitioning strat-egy and partition-specific substitution models were selectedin a multistep process (Verbruggen et al., 2010). Initially, aguide tree was obtained by carrying out a second-level max-imum likelihood (ML) search on the unpartitioned datasetwith a HKY + Γ8 model using TreeFinder (Jobb et al., 2004).Then, the dataset was divided by 17 different partitioningstrategies (combining different levels of locus segmentation).Subsequently, Bayesian information criterion (BIC) calcula-tions were performed for all 17 potential partitioning strate-gies, assuming the guide tree and HKY + Γ8 model for eachpartition. The five best-scoring partitioning strategies (lowestBIC scores) were retained for further analysis. In the nextstep, models of sequence evolution were selected for indivi-dual partitions using BIC. For each partition present in thefive retained partitioning strategies, 12 different nucleotidesubstitution models were evaluated (F81, HKY, GTR andtheir combinations with Γ, I and Γ + I). Finally, the partition-ing strategies were re-evaluated using the selected models forthe particular partitions. This BIC-based model selectionprocedure selected the following model with eight partitions:(1) internal transcribed spacers ITS1 and ITS2 –GTR + Γ; (2)5.8 S ribosomal locus – F81; (3), first codon position of therbcL gene – GTR + Γ; (4) second codon position of the rbcL
P. Škaloud et al. 214
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

Tab
le1.
Originandsamplingdetails
ofanalysed
strains.
Taxo
nStrain
Locality
Geographiccoordinates
Sam
plingdate
Tem
p.(°C)
pHCon
d.(µScm
−1)
S.am
erican
aS81
.C7
Záplavy
NR,C
zech
Repub
lic50
.146
43°N,1
4.01
389°
E19
.3.201
25
7.2
670
S.am
erican
aS10
4.C4
Schinkelbos,N
etherlands
52.299
27°N,4
.807
42°E
7.12
.201
24
7.9
1257
S.bo
realis
S58
.C7
Lillesjön,
Sweden
56.955
43°N,1
4.71
428°
E22
.4.2011
126.5
81S.
borealis
S62
.D7
unnamed
lake,D
isko
island
,Greenland
69.289
13°N,5
3.49
322°
W28
.7.2011
76.3
81S.
borealis
S90
.F3
Rutajärvi,H
arjulahti,Finland
61.906
36°N,2
6.02
054°
E5.5.20
128
5.8
–S.
borealis
S90
.G4
Harjujärvi,Finland
61.885
47°N,2
6.01
479°
E5.5.20
127
4.9
–S.
borealis
S90
.M34
Jämsänjarvi,F
inland
62.250
60°N,2
5.18
766°
E5.5.20
126
5.2
–S.
borealis
S110.E2/F3
asm
allp
ooln
earKallijõgi,E
ston
ia58
.335
39°N,2
7.26
385°
E10
.5.201
321
6.7
157
S.bo
realis
S110.B9
Apn
ajõgi,E
ston
ia58
.343
05°N,2
7.26
793°
E10
.5.201
319
6.7
110
S.bo
realis
S113.F4/F6
Emajõg
i,Eston
ia58
.396
36°N,2
6.31
094°
E10
.5.201
316
7.5
322
S.bo
realis
S114.B8/B9/C8
unnamed
lake
(A1),S
weden
68.348
05°N,1
9.03
784°
E14
.6.201
313
6.3
134
S.bo
realis
S114.F3
unnamed
lake
near
Paittasjärvi,S
weden
67.862
23°N,1
9.02
746°
E15
.6.201
316
6.5
23S.
borealis
S114.G6
Syv
äjärvi,S
weden
67.756
64°N,2
0.09
297°
E16
.6.201
312
6.6
49S.
borealis
S115.F4
apo
olnear
Arosnjarkajaute,S
weden
68.428
76°N,1
8.35
156°
E19
.6.201
34
6.8
88S.
borealis
S115.G2/G3
apo
olnear
Vassijaure,Sweden
68.433
05°N,1
8.23
966°
E19
.6.201
37
6.9
45S.
borealis
S115.G7
Vassijaure,Sweden
68.430
72°N,1
8.12
755°
E19
.6.201
311
6.9
24S.
borealis
S116.B6
Kjerringd
alsvatnet,Norway
68.660
19°N,1
5.54
052°
E19
.6.201
316
6.2
37S.
borealis
S117.D3
apo
olnear
Paittasjärvi,S
weden
67.858
03°N,1
9.02
454°
E15
.6.201
316
6.0
13S.
borealis
S118.C6
apo
olnear
Torneträsk,
Sweden
68.355
76°N,1
8.82
491°
E17
.6.201
314
7.4
173
S.bo
realis
SCFIN
13A
Ojala,F
inland
61.847
78°N,2
6.29
145°
E5.5.20
1210
6.5
–S.
borealis
SCFIN
22B
Heinäjärvi,Finland
62.88112
°N,2
5.49
724°
E5.5.20
127
5.9
–S.
borealis
SCGRL34
0Sanning
assupTasia,G
reenland
69.269
39°N,5
3.47
820°
W28
.7.2011
156.8
72S.
borealis
SCGRL36
4amorrainelake,D
isko
island
,Greenland
69.297
54°N,5
3.51
961°
W28
.7.2011
5–
–S.
borealis
SCSWE9
Helgassjön,
Sweden
56.956
11°N,1
4.71
630°
E22
.4.2011
116.6
72S.
cono
pea
S10
3.B3
apo
olnear
Smědava,Czech
Repub
lic50
.846
06°N,1
5.23
6750
°E
14.10.20
126
5.0
40S.
glab
raS89
.F5
Saarijärvi,Finland
62.701
73°N,2
5.26
388°
E5.5.20
126
6.5
–S.
heteropo
raS20
.1Lochan
Add
,Scotland
,UK
56.032
98°N,5
.534
510°
W4.6.20
08–
––
S.heteropo
raS20
.45
CrinanCanal,S
cotland
,UK
56.060
51°N,5
.477
146°
W4.6.20
08–
––
S.heteropo
raS40
.F11
Pod
hradskápo
ol,C
zech
Repub
lic50
.46112
°N,1
4.9117
6°E
3.4.20
1116
7.4
660
S.heteropo
raS54
.E11
Klejnarka
river,Czech
Repub
lic49
.969
56°N,1
5.32
123°
E19
.4.2011
––
–S.
heteropo
raS86
.F2
Zbý
šovpo
nd,C
zech
Repub
lic49
.81160
°N,1
5.35
409°
E20
.3.201
2–
––
S.heteropo
raS87
.C6
anephemeralpu
ddlenear
Kufstein,
Austria
47.597
91°N,1
2.15
115°
E22
.4.201
2–
––
S.heteropo
raS10
1.F7
apo
ndin
Nyg
årdsparken,B
ergen,
Norway
60.386
14°N,5
.324
47°E
21.10.20
12–
6.2
117
S.heteropo
raS112.E2
Saardepaisjärv,E
ston
ia58
.143
55°N,2
4.97
033°
E10
.5.201
319
8.7
313
S.heteropo
raS112.F5
Rahum
eri,Eston
ia58
.182
46°N,2
5.05
247°
E10
.5.201
317
7.5
213
S.heteropo
raS113.C8
KarulaJarv,E
ston
ia58
.398
58°N,2
5.59
201°
E10
.5.201
315
6.9
458
S.heteropo
raS117.G6
Torneträsk,
Sweden
68.355
73°N,1
8.82
172°
E17
.6.201
312
7.5
53S.
heteropo
raSCIRL8
GarlanLou
gh,Ireland
55.003
43°N,7
.905
883°
W29
.9.2011
166.1
121
S.hibernica
SIE
E4
The
Lon
gRange,Ireland
51.997
25°N,9
.550
89°W
17.7.200
916
8.0
23S.
hibernica
SIE
E8
unnamed
lake
near
Maam
Cross,Ireland
53.450
49°N,9
.544
07°W
19.7.200
916
6.2
56S.
hibernica
SIE
E11
Glend
ollagh
Lou
gh,Ireland
53.466
58°N,9
.738
63°W
18.7.200
917
7.9
53S.
hibernica
SIE
M38
The
Lon
gRange,Ireland
51.997
25°N,9
.550
89°W
19.9.200
8–
7.6
32S.
hibernica
SIE
103.C8/C9/C11
Gow
laun
Lou
gh,Ireland
51.783
16°N,9
.765
03°W
6.3.20
107
4.4
31
(con
tinued)
Four new species of Synura 215
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

gene – F81; (5) third codon position of the rbcL gene – GTR+ Γ; (6) first codon position of the coxI gene – GTR + Γ; (7)second codon position of the coxI gene – F81 + Γ; and (8)third codon position of the coxI gene – GTR + Γ.
The phylogenetic tree was inferred with Bayesian infer-ence (BI) using MrBayes version 3.1.2 (Ronquist &Huelsenbeck, 2003). Two parallel MCMC runs were carriedout for 10 million generations. The dataset was divided intoeight partitions, for which different substitution models wereselected according to the BIC-based model selection. Treesand parameters were sampled every 100 generations.Convergence of the two runs was assessed during the runby calculating the average standard deviation of split frequen-cies (SDSF). The SDSF value between simultaneous runswas 0.003908. The burn-in was determined using the ‘sump’command. Bootstrap analyses were performed with ML andweighted parsimony (wMP) criteria using GARLI version0.951 (Zwickl, 2006) and PAUP* version 4.0b10(Swofford, 2002), respectively. ML analyses consisted ofrapid heuristic searches (100 pseudoreplicates) using auto-matic termination (genthreshfortopoterm command set to100 000). The dataset was divided into eight partitions withdifferent substitution models. The wMP bootstrapping (1000replications) was performed using heuristic searches with100 random sequence addition replicates, tree bisectionreconnection swapping, random addition of sequences (thenumber limited to 10 000 for each replicate), and gap char-acters treated as a fifth character state. Character weightswere assigned using the rescaled consistency index on ascale of 0 to 1000. New weights were based on the mean fitvalues for each character over all trees in the memory.
Morphological investigations and statistical analyses
For morphological observations, the strains were cultivated in50-ml Erlenmeyer flasks for 1–2 months. To avoid depletionof nutrients, the strains were grown under lower light andtemperature regimes. Light microscopy (LM) observationswere performed using an Olympus BX51 microscope. Forscanning electron microscopy (SEM) observations, a drop ofglutaraldehyde (GA)-fixed (2%GA overnight) cell suspensionwas sedimented for 60 min on polyL-lysine-coated glass cov-erslips to ensure appropriate cell adhesion. The coverslipswere washed by repeated transfer into drops of deionizedwater dispensed onto the hydrophobic surface of a Parafilmstrip, and subsequently dehydrated via acetone series. Thecells were dried to a critical point with liquid carbon dioxide(Bal-Tec CPD 030), coated with platinum for 90 s with a Bal-Tec SCD 050 sputter coater, and observed with a JEOL JSM-740 1F FESEM scanning electron microscope. For transmis-sion electronmicroscopy (TEM) investigations of silica scales,a drop from the living cultures was placed onto formvar-coatedcopper grids and dried. After washing in a series of waterdroplets, the grids were examined in a TEM JEOL 1011electron microscope. At least three strains were randomlychosen from each of the clades identified by the molecularstudy, including the cultures deposited in the CultureCollection of Algae of Charles University in Prague (CAUP)and the National Center for Marine Algae and Microbiota inMaine (designated as CCMP). For each clade, the followingcharacters were measured in 30 randomly selected scales: (1)scale length; (2) scale width; (3) area of a base hole; (4)T
able1.
Con
tinued.
Taxo
nStrain
Locality
Geographiccoordinates
Sam
plingdate
Tem
p.(°C)
pHCon
d.(µScm
−1)
S.hibernica
SIE
104.D11
Glanm
oreLake,Ireland
51.739
01°N,9
.771
94°W
6.3.20
109
5.2
40S.
hibernica
SIE
105.F6
CahaLakes,Ireland
51.721
71°N,9
.659
94°W
6.3.20
107
5.4
31S.
hibernica
SCIRL20
aDerrynaherrivaLou
gh,Ireland
54.070
93°N,9
.512
78°W
30.9.2011
145.8
87S.
hibernica
SCIRL41
Easky
Lou
gh,Ireland
54.157
17°N,8
.844
04°W
2.10
.2011
146.1
51S.
hibernica
SCIRL60
Maumwee
Lou
gh,Ireland
53.474
38°N,9
.543
63°W
19.7.200
9–
6.9
39S.
laticarina
S89
.D5
Ojala,F
inland
61.847
78°N,2
6.29
145°
E5.5.20
1210
6.5
–S.
laticarina
S90
.C8
Tehriselkä,Finland
61.756
52°N,2
6.48
581°
E5.5.20
126
5.5
–S.
laticarina
S110.C9
Apn
ajõgi,E
ston
ia58
.343
05°N,2
7.26
793°
E10
.5.201
319
6.7
110
S.laticarina
S113.E5
apo
olnear
Emajõg
i,Eston
ia58
.396
37°N,2
6.31
094°
E10
.5.201
316
7.6
357
S.laticarina
S115.B2
Bardu
elva,N
orway
69.013
95°N,1
8.48
489°
E17
.6.201
310
7.0
80S.
laticarina
S115.D2
apo
olnear
Arosnjarkajaute,S
weden
68.428
76°N,1
8.35
156°
E19
.6.201
34
6.8
88S.
laticarina
S115.E5
apo
olnear
Vassijaure,Sweden
68.433
05°N,1
8.23
966°
E19
.6.201
37
6.9
45S.
macropo
raS71
.B4
Pod
hradskápo
ol,C
zech
Repub
lic50
.46112
°N,1
4.9117
6°E
4.3.20
125.5
6.5
230
S.macropo
raS10
4.D7
acanalinOosteinde,N
etherlands
52.288
16°N,4
.809
22°E
7.12
.201
23.3
7.9
850
S.macropo
raS10
4.F11
apo
olin
Aalsm
eer,Netherlands
52.271
48°N,4
.761
43°E
7.12
.201
24.1
7.9
975
S.petersenii
S89
.D6
Ojala,F
inland
61.847
78°N,2
6.29
145°
E5.5.20
129.5
6.5
–S.
petersenii
S89
.F9
Heinäjärvi,Finland
62.88112
°N,2
5.49
724°
E5.5.20
127.0
5.9
–S.
truttae
S62
.B5
apeatbo
gnear
Přebu
z,Czech
Repub
lic50
.383
08°N,1
2.59
668°
E24
.5.2011
12.6
4.7
107
P. Škaloud et al. 216
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4
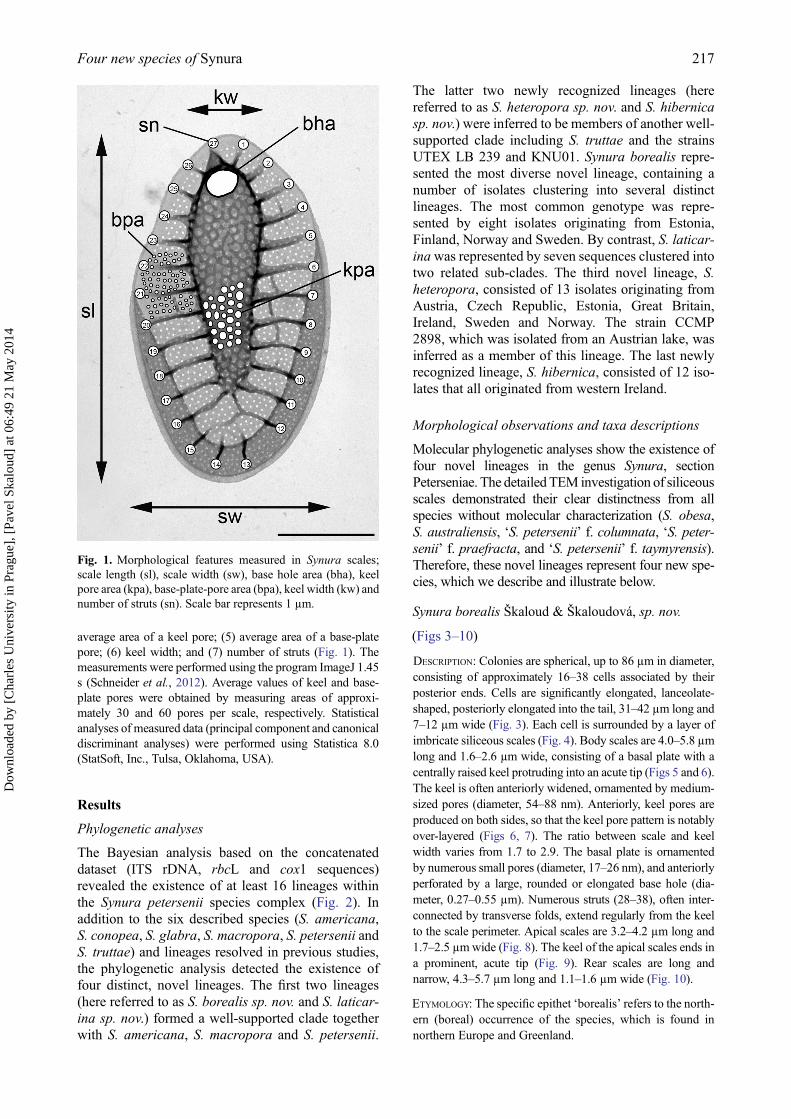
average area of a keel pore; (5) average area of a base-platepore; (6) keel width; and (7) number of struts (Fig. 1). Themeasurements were performed using the program ImageJ 1.45s (Schneider et al., 2012). Average values of keel and base-plate pores were obtained by measuring areas of approxi-mately 30 and 60 pores per scale, respectively. Statisticalanalyses of measured data (principal component and canonicaldiscriminant analyses) were performed using Statistica 8.0(StatSoft, Inc., Tulsa, Oklahoma, USA).
Results
Phylogenetic analyses
The Bayesian analysis based on the concatenateddataset (ITS rDNA, rbcL and cox1 sequences)revealed the existence of at least 16 lineages withinthe Synura petersenii species complex (Fig. 2). Inaddition to the six described species (S. americana,S. conopea, S. glabra, S. macropora, S. petersenii andS. truttae) and lineages resolved in previous studies,the phylogenetic analysis detected the existence offour distinct, novel lineages. The first two lineages(here referred to as S. borealis sp. nov. and S. laticar-ina sp. nov.) formed a well-supported clade togetherwith S. americana, S. macropora and S. petersenii.
The latter two newly recognized lineages (herereferred to as S. heteropora sp. nov. and S. hibernicasp. nov.) were inferred to be members of another well-supported clade including S. truttae and the strainsUTEX LB 239 and KNU01. Synura borealis repre-sented the most diverse novel lineage, containing anumber of isolates clustering into several distinctlineages. The most common genotype was repre-sented by eight isolates originating from Estonia,Finland, Norway and Sweden. By contrast, S. laticar-inawas represented by seven sequences clustered intotwo related sub-clades. The third novel lineage, S.heteropora, consisted of 13 isolates originating fromAustria, Czech Republic, Estonia, Great Britain,Ireland, Sweden and Norway. The strain CCMP2898, which was isolated from an Austrian lake, wasinferred as a member of this lineage. The last newlyrecognized lineage, S. hibernica, consisted of 12 iso-lates that all originated from western Ireland.
Morphological observations and taxa descriptions
Molecular phylogenetic analyses show the existence offour novel lineages in the genus Synura, sectionPeterseniae. The detailed TEM investigation of siliceousscales demonstrated their clear distinctness from allspecies without molecular characterization (S. obesa,S. australiensis, ‘S. petersenii’ f. columnata, ‘S. peter-senii’ f. praefracta, and ‘S. petersenii’ f. taymyrensis).Therefore, these novel lineages represent four new spe-cies, which we describe and illustrate below.
Synura borealis Škaloud & Škaloudová, sp. nov.
(Figs 3–10)
DESCRIPTION: Colonies are spherical, up to 86 µm in diameter,consisting of approximately 16–38 cells associated by theirposterior ends. Cells are significantly elongated, lanceolate-shaped, posteriorly elongated into the tail, 31–42 µm long and7–12 µm wide (Fig. 3). Each cell is surrounded by a layer ofimbricate siliceous scales (Fig. 4). Body scales are 4.0–5.8 µmlong and 1.6–2.6 µm wide, consisting of a basal plate with acentrally raised keel protruding into an acute tip (Figs 5 and 6).The keel is often anteriorly widened, ornamented by medium-sized pores (diameter, 54–88 nm). Anteriorly, keel pores areproduced on both sides, so that the keel pore pattern is notablyover-layered (Figs 6, 7). The ratio between scale and keelwidth varies from 1.7 to 2.9. The basal plate is ornamentedby numerous small pores (diameter, 17–26 nm), and anteriorlyperforated by a large, rounded or elongated base hole (dia-meter, 0.27–0.55 µm). Numerous struts (28–38), often inter-connected by transverse folds, extend regularly from the keelto the scale perimeter. Apical scales are 3.2–4.2 µm long and1.7–2.5 µm wide (Fig. 8). The keel of the apical scales ends ina prominent, acute tip (Fig. 9). Rear scales are long andnarrow, 4.3–5.7 µm long and 1.1–1.6 µm wide (Fig. 10).
ETYMOLOGY: The specific epithet ‘borealis’ refers to the north-ern (boreal) occurrence of the species, which is found innorthern Europe and Greenland.
Fig. 1. Morphological features measured in Synura scales;scale length (sl), scale width (sw), base hole area (bha), keelpore area (kpa), base-plate-pore area (bpa), keel width (kw) andnumber of struts (sn). Scale bar represents 1 µm.
Four new species of Synura 217
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

Fig. 2. Phylogeny of the genus Synura, section Peterseniae, obtained by Bayesian inference of the concatenated ITS rDNA, rbcL andcox1 dataset. The analysis was performed under a partitioned model, using different substitution models for each partition. Values atthe nodes indicate statistical support estimated by three methods; MrBayes posterior node probability (left), maximum likelihoodbootstrap (middle), and maximum parsimony bootstrap (right). Only statistical supports higher than 0.90/50/50 are shown. Thickbranches highlight nodes receiving the highest posterior probability (PP) support (1.00). Newly sequenced strains are marked in bold.Strains used for the statistical analyses of morphological features are marked by asterisks. Scale bar represents the expected number ofsubstitutions per site.
P. Škaloud et al. 218
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

TYPE LOCALITY: Unnamed lake, Disko Island, Greenland(69.28913°N, –53.49322°W).
HOLOTYPE: Synura borealis strain S 62.D7, frozen materialdeposited in the Culture Collection of Algae of CharlesUniversity in Prague (CAUP). Figure 6 presents an illustra-tion of the holotype.
HOLOTYPE DNA BARCODE: GenBank Accession no.HG514176.
DISTRIBUTION (Table 1): Estonia, Finland, Greenland, Norwayand Sweden.
Synura heteropora Škaloud, Škaloudová &Procházková, sp. nov.
(Figs 11–19)
DESCRIPTION: Colonies are spherical, up to 50 µm in diameter,consisting of approximately 16–36 cells associated by theirposterior ends (Fig. 11). Cells are pyriform, posteriorly elon-gated into the tail, 20–25 µm long and 7–11 µm wide. Eachcell is surrounded by a layer of imbricate siliceous scales(Fig. 12). Body scales are 2.5–3.8 µm long and 1.1–1.9 µmwide, consisting of a basal plate with a centrally raised keelprotruding into an acute tip (Fig. 13). The keel is cylindrical,occasionally slightly widened anteriorly, and ornamented by
larger pores (diameter, 49–100 nm) (Fig. 14). The ratiobetween scale and keel width varies from 1.7 to 3.9. Thebasal plate is ornamented by numerous small pores (diameter,20–31 nm), and anteriorly perforated by a rounded base hole(diameter, 0.19–0.42 µm). Numerous struts (22–28), ofteninterconnected by transverse folds, extend regularly from thekeel to the scale perimeter (Fig. 15). Apical scales are 2.6–3.0µm long and 1.3–1.6 µm wide (Fig. 16). The keel of theapical scales ends in a rounded tip (Fig. 17). Rear scales are1.8–3.2 µm long and 0.6–1.0 µm wide (Figs 18, 19).
ETYMOLOGY: Named for the different sizes of the keel- andbase-plate pores.
TYPE LOCALITY: Crinan Canal, Scotland, UK (56.060461° N,5.481813° W).
HOLOTYPE: Synura heteropora strain S 20.45, frozen materialdeposited in the Culture Collection of Algae of CharlesUniversity in Prague (CAUP). Figure 14 presents an illustra-tion of the holotype.
HOLOTYPE DNA BARCODE: GenBank Accession no.HG514198.
DISTRIBUTION (Table 1): Austria, Czech Republic, Estonia,Ireland, Norway, Sweden and the UK.
Synura hibernica Škaloud & Škaloudová, sp. nov.
Figs. 3–10. Scale morphology of Synura borealis sp. nov. (Fig. 3: LM; Figs 4, 5: SEM; Figs 6–10: TEM). Scale bars represent 10 µm(Fig. 3) and 1 μm (Figs 4–10). Fig. 3. Colony consisting of elongated, lanceolate-shaped cells (strain S 90.G4). Fig. 4. Single cellsurrounded by a layer of siliceous scales (S 90.G4). Fig. 5. Body scale (S 90.G4). Fig. 6. Body scale with transverse foldsinterconnecting the struts. Note the over-layered pore pattern at the scale keel (S 62.D7). Fig. 7. Body scale with obviously anteriorlywidened keel (S 62.D7). Fig. 8.Apical scale (S 62.D7). Fig. 9.Apical scales with prominently protruding keel tips (S 58.C7). Fig. 10.Rear scale (S 62.D7).
Four new species of Synura 219
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

(Figs 20–28)
DESCRIPTION: Colonies are spherical, up to 94 µm in diameter,consisting of approximately 12–52 cells associated by theirposterior ends (Fig. 20). Cells are significantly elongated,anteriorly cylindrical, posteriorly tapering into the tail, 26–47 µm long and 6–12 µm wide. Each cell is surrounded by alayer of imbricate siliceous scales (Fig. 21). Body scales are3.4–5.6 µm long and 1.2–2.0 µm wide, consisting of a basalplate with a centrally raised keel protruding into an acute tip(Fig. 22). The keel is cylindrical, ornamented by medium-sized pores (diameter, 49–89 nm) (Figs 23, 24). The ratiobetween scale and keel width varies from 1.9 to 3.9. The basalplate is ornamented by numerous small pores (diameter, 18–27 nm), and anteriorly perforated by a rounded base hole(diameter, 0.14–0.38 µm). A large number of struts (30–47),often interconnected by transverse folds, extend regularlyfrom the keel to the scale perimeter (Figs 22, 23). Apicalscales are 2.9–3.3 µm long and 1.6–1.9 µm wide (Fig. 25).The keel of the apical scales ends in a prominent, acute tip(Fig. 26), sometimes terminated by two short teeth. Rearscales are long and narrow, 3.0–4.9 µm long and 0.9–1.0µm wide (Figs 27, 28).
ETYMOLOGY: The specific epithet ‘hibernica’ refers to thecommon occurrence of the species in Ireland.
TYPE LOCALITY: Glanmore Lake, Ireland (51.732724° N,9.767121° W).
HOLOTYPE: Synura hibernica strain S IE 104.D11, frozenmaterial deposited in the Culture Collection of Algae ofCharles University in Prague (CAUP). Figure 22 presentsan illustration of the holotype.
HOLOTYPE DNA BARCODE: GenBank Accession no.HG514216.
DISTRIBUTION (Table 1): Currently known only in Ireland.
Synura laticarina Škaloud & Škaloudová, sp. nov.
(Figs 29–36)
DESCRIPTION: Colonies are spherical, up to 64 µm in diameter,consisting of approximately 12–28 cells associated by theirposterior ends (Fig. 29). Cells are pyriform, posteriorly elon-gated into the tail, 21–32 µm long and 7–13 µmwide. Each cellis surrounded by a layer of imbricate siliceous scales (Fig. 30).Body scales are 3.1–4.3 µm long and 1.6–2.1 µm wide, con-sisting of a basal plate with a centrally raised keel protrudinginto an acute tip (Fig. 31). The keel is often anteriorly widened,ornamented bymedium-sized pores (diameter, 52–76 nm) (Figs32, 33). Anteriorly, keel pores are produced on both sides, sothat the keel pore pattern is notably over-layered (Fig. 33). Theratio between scale and keel width varies from 1.9 to 2.9. Thebasal plate is ornamented by numerous small pores (diameter,19–25 nm), and anteriorly perforated by a rounded base hole(diameter, 0.20–0.33 µm). Numerous struts (24–32), ofteninterconnected by transverse folds, extend regularly from the
Figs. 11–19. Scale morphology of Synura heteropora sp. nov. (Fig. 11: LM; Figs 12, 13, 16: SEM; Figs 14, 15, 17–19: TEM). Scalebars represent 10 μm (Fig. 11) and 1 μm (Figs 12–19). Fig. 11. Colony consisting of densely grouped, pyriform cells (strain S 20.45).Fig. 12. Single cell surrounded by a layer of siliceous scales (S 54.E11). Fig. 13.Body scale (S 54.E11). Fig. 14.Body scale (S 20.45).Fig. 15. Body scale with transverse folds interconnecting the struts (S 87.C6). Fig. 16. Apical scale (S 54.E11). Fig. 17. Apical scalewith a pronounced, rounded keel tip (S 101.F7). Figs 18, 19. Rear scales (S 101.F7).
P. Škaloud et al. 220
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4
keel to the scale perimeter (Fig. 34). Apical scales are 2.5–3.0µm long and 1.7–2.0 µm wide (Fig. 35). The keel of the apicalscales ends in a prominent, sometimes rounded tip. Rear scalesare 2.7–4.3 µm long and 0.7–1.3 µm wide (Fig. 36).
ETYMOLOGY: The specific epithet is derived from the Latin‘latus’ (broad) and ‘carina’ (keel), referring to the remarkableanterior widening of the keel.
TYPE LOCALITY: Ojala Lake, Finland (61.847778° N,26.291447° E).
HOLOTYPE: Synura laticarina strain S 89.D5, frozen materialdeposited in the Culture Collection of Algae of CharlesUniversity in Prague (CAUP). Figure 33 presents an illustra-tion of the holotype.
HOLOTYPE DNA BARCODE: GenBank Accession no.HG514221.
DISTRIBUTION (Table 1): Estonia, Finland, Norway andSweden.
Morphological analyses
To investigate morphological differences among the10 recognized, closely related Synura species in
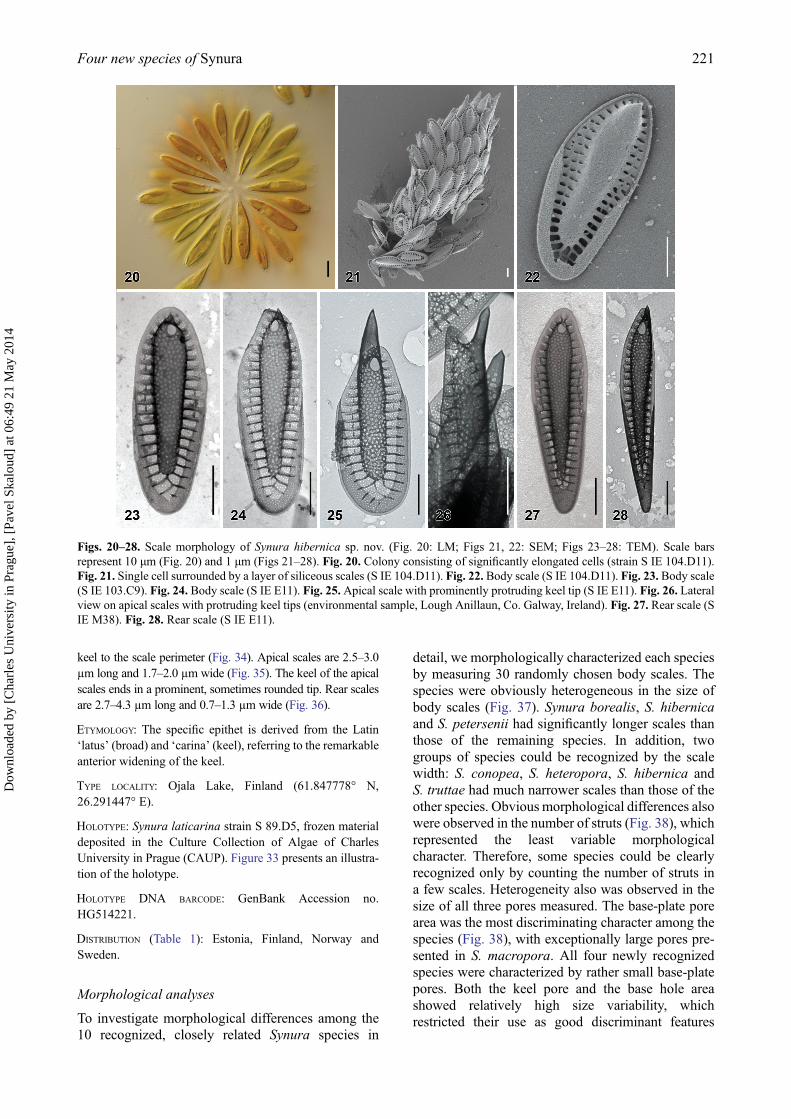
detail, we morphologically characterized each speciesby measuring 30 randomly chosen body scales. Thespecies were obviously heterogeneous in the size ofbody scales (Fig. 37). Synura borealis, S. hibernicaand S. petersenii had significantly longer scales thanthose of the remaining species. In addition, twogroups of species could be recognized by the scalewidth: S. conopea, S. heteropora, S. hibernica andS. truttae had much narrower scales than those of theother species. Obvious morphological differences alsowere observed in the number of struts (Fig. 38), whichrepresented the least variable morphologicalcharacter. Therefore, some species could be clearlyrecognized only by counting the number of struts ina few scales. Heterogeneity also was observed in thesize of all three pores measured. The base-plate porearea was the most discriminating character among thespecies (Fig. 38), with exceptionally large pores pre-sented in S. macropora. All four newly recognizedspecies were characterized by rather small base-platepores. Both the keel pore and the base hole areashowed relatively high size variability, whichrestricted their use as good discriminant features
Figs. 20–28. Scale morphology of Synura hibernica sp. nov. (Fig. 20: LM; Figs 21, 22: SEM; Figs 23–28: TEM). Scale barsrepresent 10 μm (Fig. 20) and 1 μm (Figs 21–28). Fig. 20. Colony consisting of significantly elongated cells (strain S IE 104.D11).Fig. 21. Single cell surrounded by a layer of siliceous scales (S IE 104.D11). Fig. 22. Body scale (S IE 104.D11). Fig. 23. Body scale(S IE 103.C9). Fig. 24. Body scale (S IE E11). Fig. 25.Apical scale with prominently protruding keel tip (S IE E11). Fig. 26. Lateralview on apical scales with protruding keel tips (environmental sample, Lough Anillaun, Co. Galway, Ireland). Fig. 27. Rear scale (SIE M38). Fig. 28. Rear scale (S IE E11).
Four new species of Synura 221
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

(Fig. 39). Synura borealis and S. truttae were wellcharacterized by a notably large base hole. Strongheterogeneity was observed in the base-plate area tokeel area ratio (Fig. 40). The keel pores of S. macro-pora and S. glabrawere approximately comparable insize to the base-plate pores; however, those of S.borealis, S. heteropora, S. hibernica and S. laticarinawere usually distinctively larger. Synura americana,S. hibernica and S. laticarina were well characterizedby an anteriorly widened keel (Fig. 40). The centralmorphological characteristics of these 10 lineages,and other members of the genus Synura, sectionPeterseniae, are presented in Table 2. A key to thesespecies is given in Table S2.
The principal component analysis (PCA) of theentire dataset resulted in a relatively well-definedgrouping of scales belonging to the particular species(Fig. 41). Scales of S. macropora and S. borealisformed two distinct clusters with negative values onthe first PCR axis. By contrast, scales of S. conopeawere mostly intermixed with other species. The cano-nical discriminant analysis (CDA) yielded much bet-ter grouping of scales based on their morphologicaldata (Fig. 42). With the exception of S. conopea andS. americana, all species formed distinct, separateclusters. The discriminant analysis (DA) indicated
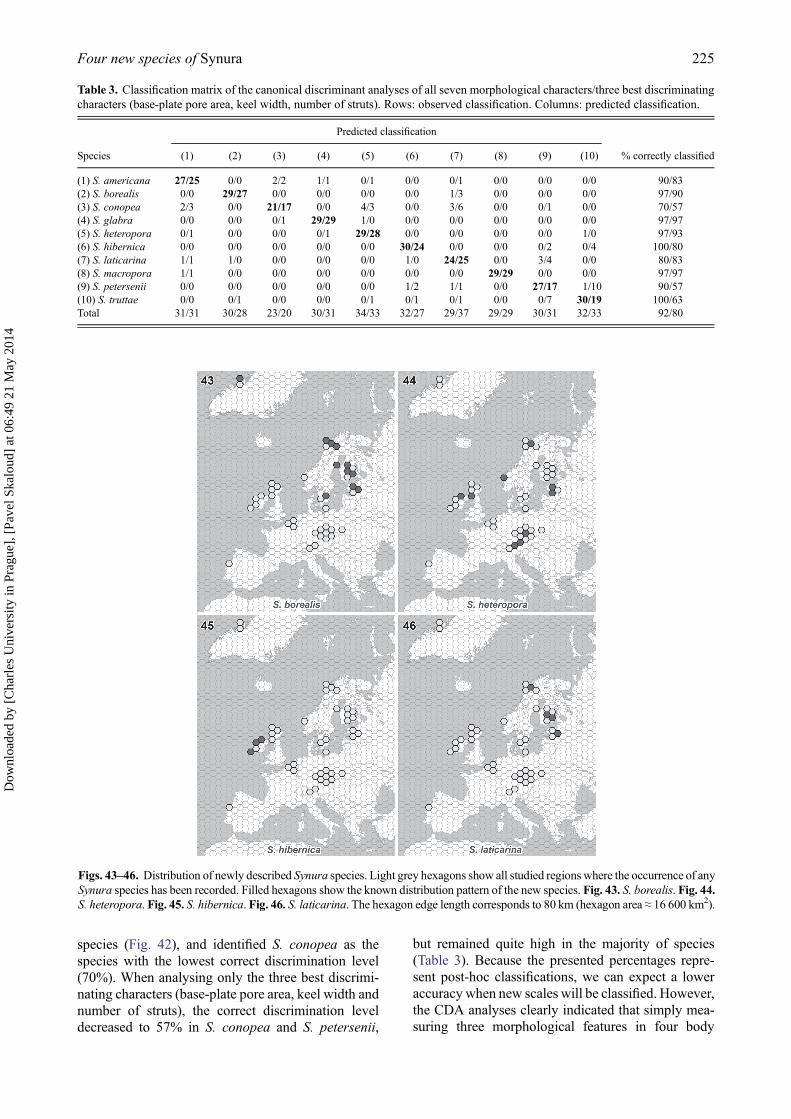
strongly significant differentiation among the 10 spe-cies (Wilk’s λ = 0.0008; P < 0.00001). The forwardstepwise analysis indicated that all tested morpholo-gical characters were significant for species recogni-tion (P < 0.00001), and selected the base-plate porearea (Partial Wilk’s λ = 0.20), the keel width (PartialWilk’s λ = 0.41) and the number of struts (PartialWilk’s λ = 0.46) as the three best discriminating char-acters. The discrimination of species was highly sig-nificant even when only these three characters wereanalysed (Wilk’s λ = 0.0058; P < 0.00001).Congruently, whereas the first CDA axis was princi-pally correlated with the base-plate pore area and thenumber of struts (correlation coefficients −0.82 and0.63, respectively), the second CDA axis correlatedwith keel and scale widths (correlation coefficients0.68 and 0.64, respectively). The average correct dis-crimination of individual scales on the basis of theirmorphology reached 92% (Table 3). The lowest cor-rect discrimination levels were recovered in S. con-opea (70.0%) and S. laticarina (80.0%).
Biogeography
A 6-year sampling of Synura isolates performed in 15different countries enabled us to determine their
Figs. 29–36. Scale morphology of Synura laticarina sp. nov. (Fig. 29: LM; Figs 30 and 31: SEM; Figs 32–36: TEM). Scale barsrepresent 10 μm (Fig. 29) and 1 μm (Figs 30–36). Fig. 29. Colony consisting of pyriform cells (strain S 90.C8). Fig. 30. Single cellsurrounded by a layer of siliceous scales (S 90.C8). Fig. 31.Body scale (S 90.C8). Fig. 32.Body scale with anteriorly widened keel (S89.D5). Fig. 33. Body scale. Note over-layered pore pattern at the scale keel (S 89.D5). Fig. 34. Body scale with transverse foldsinterconnecting the struts (S 90.C8). Fig. 35. Apical scale (S 89.D5). Fig. 36. Rear scale (S 89.D5).
P. Škaloud et al. 222
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4
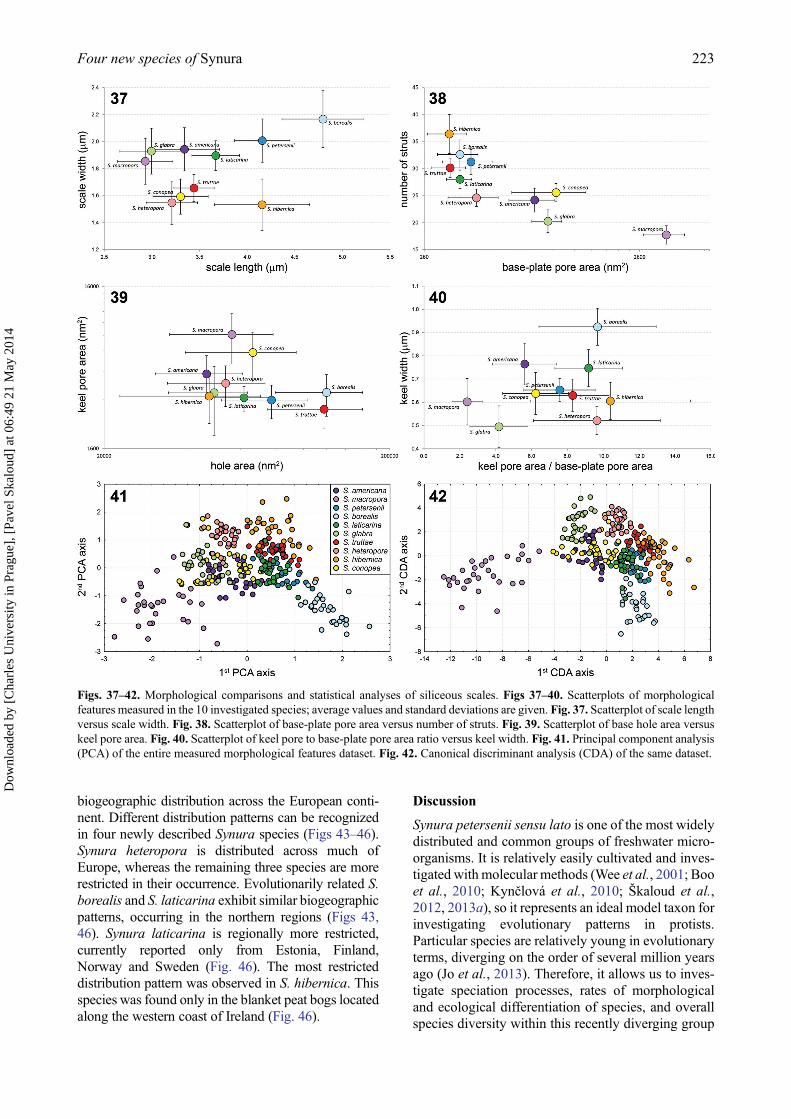
biogeographic distribution across the European conti-nent. Different distribution patterns can be recognizedin four newly described Synura species (Figs 43–46).Synura heteropora is distributed across much ofEurope, whereas the remaining three species are morerestricted in their occurrence. Evolutionarily related S.borealis and S. laticarina exhibit similar biogeographicpatterns, occurring in the northern regions (Figs 43,46). Synura laticarina is regionally more restricted,currently reported only from Estonia, Finland,Norway and Sweden (Fig. 46). The most restricteddistribution pattern was observed in S. hibernica. Thisspecies was found only in the blanket peat bogs locatedalong the western coast of Ireland (Fig. 46).
Discussion
Synura petersenii sensu lato is one of the most widelydistributed and common groups of freshwater micro-organisms. It is relatively easily cultivated and inves-tigated with molecular methods (Wee et al., 2001; Booet al., 2010; Kynčlová et al., 2010; Škaloud et al.,2012, 2013a), so it represents an ideal model taxon forinvestigating evolutionary patterns in protists.Particular species are relatively young in evolutionaryterms, diverging on the order of several million yearsago (Jo et al., 2013). Therefore, it allows us to inves-tigate speciation processes, rates of morphologicaland ecological differentiation of species, and overallspecies diversity within this recently diverging group
Figs. 37–42. Morphological comparisons and statistical analyses of siliceous scales. Figs 37–40. Scatterplots of morphologicalfeatures measured in the 10 investigated species; average values and standard deviations are given. Fig. 37. Scatterplot of scale lengthversus scale width. Fig. 38. Scatterplot of base-plate pore area versus number of struts. Fig. 39. Scatterplot of base hole area versuskeel pore area. Fig. 40. Scatterplot of keel pore to base-plate pore area ratio versus keel width. Fig. 41. Principal component analysis(PCA) of the entire measured morphological features dataset. Fig. 42. Canonical discriminant analysis (CDA) of the same dataset.
Four new species of Synura 223
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

of microorganisms. This study was the next stepnecessary for uncovering the real species diversityand to delimit and characterize particular specieswithin this complex.
Overall species diversity
Biological variation within Synura petersenii is verycomplex. Before information was available aboutgenetic differentiation, S. petersenii was generallyconsidered as a single species, with recognition of afew forms or varieties (Kristiansen & Preisig, 2007).In fact, this taxon contains a number of morphologi-cally similar, yet genetically distinct species. In addi-tion to the four newly proposed species, S. peterseniis.l. now includes 10 well-defined, genetically charac-terized species. Phylogenetic analysis clearly recog-nizes six lineages comprised of several strainsdeposited in various culture collections, which veryprobably represent new, yet undescribed species (Fig.2). Several taxa belonging to the section Peterseniaeare currently uncharacterized with molecular markers,and their phylogenetic position is still unknown. Allthese taxa, including S. australiensis (Playfair, 1915),‘S. petersenii’ f. columnata (Siver, 1988), ‘S. peterse-nii’ f. praefracta (Asmund, 1968), ‘S. petersenii’ f.taymyrensis (Kristiansen et al., 1997), and S. obesa(Němcová et al., 2008), could form additionallineages within S. petersenii s.l. Therefore, it is verydifficult to estimate the total number of species in thistaxon. In an extensive molecular investigation of morethan 100 S. petersenii s.l. strains, Boo et al. (2010)identified a large number of genotypes clustered intoseven groups. The strains originated from differentareas located in four continents. Of the four newspecies proposed in the present study, only S. hetero-pora was identified with a previously recognized gen-otype, the strain CCMP 2898 belonging to clade G IBoo et al. (2010). Obviously, there is still a largedegree of hidden diversity that cannot be fullyresolved without additional molecular and morpholo-gical data. However, even if we can predict that sev-eral tens of currently undiscovered taxa exist, it isprobable that the most frequently occurring speciesare already known and characterized.
Species delineation and morphological evolution
The six species delineated in previous studies(Kynčlová et al., 2010; Škaloud et al., 2012) wereshown to be distinguishable by the siliceous scalemorphology. However, the present study includesdescriptions of four additional, morphologicallyhighly similar taxa. Morphometric analyses of silic-eous scales enabled the significant phenotypic differ-entiation of all species of the S. petersenii complex,including the newly described species. The CDA ana-lysis significantly recognized all 10 investigatedT
able2.
Scalecharacteristicsof
selected
Synu
raspecies(sectio
nPeterseniae)w
ithsimilarscalemorph
olog
ies.New
lydescribedspeciesaregiveninbo
ld.For
term
inolog
yof
measuredcharacteristics,
seeFig.1
.
Taxo
nCelld
imension
s(µm)
Scaledimension
s(µm)
Num
berof
struts
Rearscales
long
erthan
body
scales?
Keelw
idth
Baseho
lediam
eter
(nm)
Base-platepo
rediam
eter
(nm)
Keelp
ore
diam
eter
(nm)
Bod
yscaleleng
thto
width
ratio
Con
nection
ofstruts
S.am
erican
a22
–28×8–
122.8–
4.1×1.6–2.2
20–2
8yes
0.6–
0.9
167–
336
27–43
54–9
41.4–
2.3
very
rare
S.cono
pea
20–28×8–
122.6–
3.7×1.3–1.9
22–2
9no
0.5–
0.8
189–
438
25–51
70–125
1.5–
2.5
rare
S.glab
ra19
–28×10
–14
2.2–
3.6×1.6–2.2
17–2
5no
0.4–
0.7
144–
327
29–41
44–100
1.3–
1.9
none
S.macropo
ra17
–25×8–
122.2–
3.4×1.5–2.2
15–2
2yes
0.4–
0.8
156–
391
50–78
69–137
1.3–
2.1
none
S.petersenii
20–31×8–
123.4–
4.7×1.7–2.3
26–3
7no
0.6–
0.8
244–
461
18–30
45–8
21.7–
2.5
frequent
S.truttae
22–31×11–13
3.0–
3.9×1.5–1.9
27–3
4no
0.5–
0.8
270–
557
18–25
47–7
01.8–
2.4
frequent
S.b
orealis
31–42×7–
124.0–
5.8×1.6–2.6
28–3
8no
0.8–
1.3
266–
554
17–26
54–8
81.7–
2.9
frequent
S.h
eterop
ora
20–2
5×7–
112.5–
3.8×1.1–1.9
22–2
8no
0.4–
0.7
187–
415
20–31
49–100
1.8–
2.6
frequent
S.h
ibernica
26–47×6–
123.4–
5.6×1.2–2.0
30–4
7no
0.5–
0.8
138–
378
18–27
49–8
92.2–
3.5
frequent
S.laticarina
21–32×7–
133.1–
4.3×1.6–2.1
24–3
2no
0.6–
1.0
200–
334
19–25
52–7
61.7–
2.4
frequent
‘S.p
etersenii’
f.columna
ta15
–23×8–
112.7–
3.5×1.5–2.2
22–2
7–
2.6–
3.1
––
–1.6–
1.8
none
‘S.p
etersenii’f.praefracta
–3.2–
3.4×1.8–2.0
24–2
7–
3.0–
3.5
––
–1.7–
1.8
frequent
‘S.p
etersenii’f.taym
yrensis
–2.7–
2.8×1.2
23–2
5no
1.7
––
–2.3
none
P. Škaloud et al. 224
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4
species (Fig. 42), and identified S. conopea as thespecies with the lowest correct discrimination level(70%). When analysing only the three best discrimi-nating characters (base-plate pore area, keel width andnumber of struts), the correct discrimination leveldecreased to 57% in S. conopea and S. petersenii,
but remained quite high in the majority of species(Table 3). Because the presented percentages repre-sent post-hoc classifications, we can expect a loweraccuracy when new scales will be classified. However,the CDA analyses clearly indicated that simply mea-suring three morphological features in four body
Table 3. Classification matrix of the canonical discriminant analyses of all seven morphological characters/three best discriminatingcharacters (base-plate pore area, keel width, number of struts). Rows: observed classification. Columns: predicted classification.
Species
Predicted classification
% correctly classified(1) (2) (3) (4) (5) (6) (7) (8) (9) (10)
(1) S. americana 27/25 0/0 2/2 1/1 0/1 0/0 0/1 0/0 0/0 0/0 90/83(2) S. borealis 0/0 29/27 0/0 0/0 0/0 0/0 1/3 0/0 0/0 0/0 97/90(3) S. conopea 2/3 0/0 21/17 0/0 4/3 0/0 3/6 0/0 0/1 0/0 70/57(4) S. glabra 0/0 0/0 0/1 29/29 1/0 0/0 0/0 0/0 0/0 0/0 97/97(5) S. heteropora 0/1 0/0 0/0 0/1 29/28 0/0 0/0 0/0 0/0 1/0 97/93(6) S. hibernica 0/0 0/0 0/0 0/0 0/0 30/24 0/0 0/0 0/2 0/4 100/80(7) S. laticarina 1/1 1/0 0/0 0/0 0/0 1/0 24/25 0/0 3/4 0/0 80/83(8) S. macropora 1/1 0/0 0/0 0/0 0/0 0/0 0/0 29/29 0/0 0/0 97/97(9) S. petersenii 0/0 0/0 0/0 0/0 0/0 1/2 1/1 0/0 27/17 1/10 90/57(10) S. truttae 0/0 0/1 0/0 0/0 0/1 0/1 0/1 0/0 0/7 30/19 100/63Total 31/31 30/28 23/20 30/31 34/33 32/27 29/37 29/29 30/31 32/33 92/80
Figs. 43–46. Distribution of newly described Synura species. Light grey hexagons show all studied regions where the occurrence of anySynura species has been recorded. Filled hexagons show the known distribution pattern of the new species. Fig. 43. S. borealis. Fig. 44.S. heteropora. Fig. 45. S. hibernica. Fig. 46. S. laticarina. The hexagon edge length corresponds to 80 km (hexagon area ≈ 16 600 km2).
Four new species of Synura 225
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

scales should be sufficient to correctly classify each ofthe species.
We detected a new lineage sister to S. americana,S. glabra and S. petersenii that we interpreted as twonovel species, the genetically diverse S. borealisand the more molecularly homogeneous S. laticarina(Fig. 2). Synura borealis possessed an exceptionallyhigh degree of genetic diversity, particularly in thecox1 gene, which could be interpreted as resultingfrom several independent species. However, therecognition of just these two species was stronglysupported by our morphological data. The silica scalesof all S. borealis strains shared characteristic morpho-logical features, including the anteriorly widened keelwith notably over-layered pore pattern, and large scaledimensions. Further, S. borealis and S. laticarina spe-cies could be undoubtedly recognized under the lightmicroscope. All investigated S. borealis strains werecharacterized by large colonies consisting of signifi-cantly elongated, lanceolate-shaped cells, whereasS. laticarina produced notably smaller colonies con-taining rounded, pyriform cells. Therefore, we inter-pret the high genetic diversity in S. borealis as anindication of recent speciation and ongoing evolution-ary differentiation. Further investigation of S. borealisgenotypes, including their ecophysiological differen-tiation, ecological preferences and distributional pat-terns could provide important insights into the driversof species diversification and mechanisms of popula-tion-level structuring in Synura species.
Particular species of the S. petersenii groupobviously underwent some degree of morphologicaldifferentiation, although they are relatively young inevolutionary terms. Some species were more signifi-cantly differentiated with respect to siliceous scalesthan others, and can be recognized at first glance. Forexample, S. macropora can be unambiguously identi-fied by its extremely large base-plate pores (Fig. 38).We can only speculate about the causes of this rapidmorphological evolution. Large pores could signifi-cantly decrease the density of the cell, and hencereduce the sinking rate. However, cells with largerpores could be more susceptible to viral infection(Losic et al., 2006). Synura macropora seems tooccur only in eutrophic habitats characterized byhigh conductivity values (Table 1; Škaloud et al.,2013b). It was shown that eutrophic conditionsdecreased the availability of silica, which often causeda shift from heavily silicified to less silicified diatomsin freshwater biotopes (Rabalais et al., 1996).Similarly, adaptation of S. macropora to eutrophicconditions could lead to the formation of less silicifiedscales characterized by large base-plate pores.
Synura hibernica represents another morphologi-cally well-defined species characterized by very nar-row, long siliceous scales bearing a large number ofstruts (Figs 37, 38). This species can even be distin-guished under the light microscope by the
characteristically large colonies consisting of stronglyelongated, narrow cells. The significantly differentmorphology of this Irish taxon has been discussedpreviously by Řezáčová & Škaloud (2005), whoemphasized its striking similarity with the tropicalspecies S. australiensis. Obviously, the characteristi-cally elongated shape of the siliceous scales is relatedto the elongated cell shape in both species. Synuraaustraliensis and S. hibernica represent another exam-ple of rapid morphological evolution within theS. petersenii group, leading to the speciation of organ-isms producing big colonies consisting of long andnarrow cells. Unfortunately, in the absence of mole-cular data for S. australiensis, it remains unclearwhether these two taxa are evolutionarily related toeach other. Significant morphological transformationmay have been caused by an ecological niche-shiftduring the speciation of both taxa, similarly to that ofS. macropora. In protists, a negative correlationbetween cell size and temperature has been proposedby Atkinson et al. (2003). However, S. hibernica iscurrently known only from Ireland, whereas the dis-tribution of S. australiensis is restricted to tropical andsubtropical regions (Kristiansen & Preisig, 2007).Therefore, we can rule out temperature-related mor-phological speciation of these two taxa.
The development of elongated cells in S. austra-liensis and S. hibernica could be explained by adapta-tion to oligotrophic conditions. It is known that mostoligotrophs achieve a high surface-to-volume ratio,which increases the organisms’ capacity to scavengeavailable nutrients (Reynolds, 2006). Elongation ofSynura cells would be the most effective way to sig-nificantly increase this ratio. All 12 isolated strains ofS. hibernica originated from nutrient-poor localities,with conductivity values ranging between 23–87 µScm−1 (Table 1). To our knowledge, all records ofS. australiensis have been associated with clear, oli-gotrophic water bodies (Croome & Tyler, 1985;Cronberg, 1989, 1996; Saha & Wujek, 1990; Weeet al., 1993; Hansen, 1996). The measured conductiv-ity of these water samples was 10–123 µS cm−1.
Biogeographic patterns
The biogeography of protists has become a highlycontroversial topic over the last 10 years (Martinyet al., 2006; Caron, 2009). It has been postulated thatthe small size, extremely large populations and highdispersal potential of protists result in the cosmopoli-tan distribution of the vast majority of species (Finlay,2002; Finlay & Fenchel, 2004). Conversely, limitedgeographical distributions have been implied byFoissner (1999), primarily based on the observedrestricted distribution of flagship species with easilyrecognizable morphologies and easily demonstrablepresence/absence (Foissner, 2006, 2008). Synurapetersenii s.l. represents a common group of
P. Škaloud et al. 226
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

freshwater microorganisms, known to occur in allcontinents except Antarctica (Kristiansen, 2008).Because its molecular diversity has been investigatedin three continents (Wee et al., 2001; Boo et al., 2010;Škaloud et al., 2012), it represents an ideal taxon tostudy the distribution patterns in protists.
The first evaluation of biogeographic patterns in theS. petersenii group was published by Boo et al.(2010), who described the presence of restricted bio-geographic distributions for many species. However,Škaloud et al. (2012) compared the genetic data withpublished morphological observations, and reportedmuch broader distributions for some of the investi-gated taxa. For example, putative North Americanendemic S. americana, although dominantly occur-ring in the USA and Canada, was also reported inSouth America and Europe (Cronberg, 1989;Škaloud et al., 2012). However, the restricted distri-bution of several lineages is evident (e.g. clades D andF sensu Boo et al. 2010). Our results support therestricted distribution of Synura species, includingthe strict regional endemism observed in S. hibernica.To the best of our knowledge, all four newly proposedspecies have been reported only in Europe andGreenland (Figs 43–46). A comparison of the mor-phological characteristics with previously publishedreports reveals that S. borealis and S. hibernica havebeen observed previously in Greenland and Ireland,respectively (Kristiansen, 1992;Řezáčová & Škaloud,2005); however, we found no other reports outsideEurope.
Three of four newly described species exhibitrestricted biogeographic distribution. Evolutionarilyrelated S. borealis and S. laticarina seem to be adaptedto colder areas, and their distributions appear to belimited by high summer temperatures (Figs 43, 46).The occurrence of S. borealis in Greenland indicatesno obvious dispersal limitations, so we expect itscircumboreal distribution. The most remarkable dis-tribution pattern, however, is that of S. hibernica. Thisspecies is restricted in its geographic distribution towestern Ireland (Fig. 45). Althoughwe found S. hiber-nica in nine localities located along the western coastof Ireland (Table 1), it was never detected in any of theother sampled localities. To check this highly unusualdistribution pattern, we undertook two expeditions in2008 and 2012 to investigate the diversity of Synuraspecies in neighbouring Scotland. Ecological and lim-nological conditions are very similar in westernIreland and Scotland: both areas are characterized bythe occurrence of extensive peat deposits restricted toa cool oceanic climate, known as blanket bogs(Moore, 1982). Moreover, the British Isles lie at theintersection of major bird migration corridors, andseveral bird species frequently pass through one partof the Isles to reach another (Newton, 2010).Therefore, the algal flora of the western British lake-lands, including western Ireland and northwestern
Scotland, are highly similar. The significant similarityof western British desmid flora was reported pre-viously (West & West, 1909). Although we sampleda total of 71 localities in northwestern Scotland, wedid not discover a single colony of S. hibernica. Thisdistribution pattern, which is restricted to an extre-mely small biogeographic area, is remarkable andentirely outstanding. Future work should study themechanisms underlying the obvious dispersal limita-tion of S. hibernica, including ecological and habitatrequirements, desiccation tolerance and mechanismsof cyst formation.
Acknowledgements
The authors would like to thank Fabio Rindi andMichael D. Guiry (Ryan Institute, National Universityof Ireland, Galway); Thomas Pröschold and John G. Day(Culture Collection of Algae and Protozoa, ScottishMarine Institute, Oban, UK); David John (NaturalHistory Museum, London, UK); Ole Stecher andFrantz Nielsen (Arctic Station, University ofCopenhagen, Greenland); Eugen Rott (Institute ofBotany, University of Innsbruck, Austria); HelenaBestová, Lucie Jelínková, Katarína Nemjová, PavlaSlámová, Pavel Svoboda and Vojtěch Scharfen(Department of Botany, Charles University in Prague,Czech Republic) for their kind support during sampling.This work was supported by the Czech ScienceFoundation [grant number P506/11/P056].
Supplementary information
The following supplementary material is accessible viathe Supplementary Content tab on the article’s onlinepage at http://dx.doi.org/10.1080/09670262.2014.905710
Table S1. Strain names, geographic origins, and thecorresponding ITS rDNA, rbcL, and cox1 GenBankaccession numbers for the taxa used in the phylogeneticanalyses. Newly obtained sequences are given in bold.
Table S2. Key to species of the genus Synura, sectionPeterseniae.
Alignment S1. The final, partitioned alignment ofconcatenated ITS rDNA, rbcL, and cox1 sequencesused for phylogenetic reconstructions.
References
ANDERSEN, R.A., MORTON, S.L. & SEXTON, J.P. (1997). Provasoli-Guillard National Center for Culture of Marine Phytoplankton1997 – list of strains. Journal of Phycology, 33 (suppl.): 1–75.
ASMUND, B. (1968). Studies on Chrysophyceae from some pondsand lakes in Alaska. VI. Occurrence of Synura species.Hydrobiologia, 31: 497–515.
ATKINSON, D., CIOTTI, B.J. & MONTAGNES, D.J.S. (2003). Protistsdecrease in size linearly with temperature: ca. 2.5% °C-l.Proceedings of the Royal Society of London Series B –Biological Sciences, 270: 2605–2611.
BOO, S.M., KIM, H.S., SHIN,W., BOO, G.H., CHO, S.M., JO, B.Y., KIM,J.-H., KIM, J.H., YANG, E.C., SIVER, P.A., WOLFE, A.P.,BHATTACHARYA, D., ANDERSEN, R.A. & YOON, H.S. (2010).Complex phylogeographic patterns in the freshwater alga Synura
Four new species of Synura 227
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

provide new insights into ubiquity vs. endemism in microbialeukaryotes. Molecular Ecology, 19: 4328–4338.
CARON, D.A. (2009). Past president’s address: protistan biogeogra-phy: why all the fuss? Journal of Eukaryotic Microbiology, 56:105–112.
CRONBERG, G. (1989). Scaled chrysophytes from the tropics. NovaHedwigia, Beiheft, 95: 191–232.
CRONBERG, G. (1996). Scaled chrysophytes from the OkavangoDelta, Botswana, Africa. Nova Hedwigia, Beiheft, 114: 91–108.
CROOME, R.L. & TYLER, P.A. (1985). Synura australiensis(Mallomonadaceae, Chrysophyceae), a light and electron micro-scopical investigation. Nordic Journal of Botany, 5: 399–401.
DAUGBJERG, N. & ANDERSEN, R.A. (1997). Phylogenetic analysis ofthe rbcL sequences from haptophytes and heterokont algae suggesttheir chloroplasts are unrelated.Molecular Biology and Evolution,14:1242–1251.
FINLAY, B.J. (2002). Global dispersal of free-living microbial eukar-yote species. Science, 296: 1061–1063.
FINLAY, B.J. & FENCHEL, T. (2004). The ubiquity of small species:patterns of local and global diversity. Bioscience, 54: 777–784.
FOISSNER, W. (1999). Protist diversity: estimates of the near-impon-derable. Protist, 150: 363–368.
FOISSNER, W. (2006). Biogeography and dispersal of micro-organ-isms: a review emphasizing protists. Acta Protozoologica, 45:111–136.
FOISSNER, W. (2008). Protist diversity and distribution: some basicconsiderations. Biodiversity and Conservation, 17: 235–242.
HANSEN, P. (1996). Silica-scaled Chrysophyceae and Synurophyceaefrom Madagascar. Archiv für Protistenkunde, 147: 145–172.
JO, B.Y., SHIN, W., BOO, S.M., KIM, H.S. & SIVER, P.A. (2011).Studies on ultrastructure and three-gene phylogeny of thegenus Mallomonas (Synurophyceae). Journal of Phycology, 47:415–425.
JO, B.Y., SHIN, W., KIM, H.S., SIVER, P.A. & ANDERSEN, R.A. (2013).Phylogeny of the genus Mallomonas (Synurophyceae) anddescriptions of five new species on the basis of morphologicalevidence. Phycologia, 52: 266–278.
JOBB, G., VON HAESELER, A. & STRIMMER, K. (2004). TREEFINDER:a powerful graphical analysis environment for molecular phyloge-netics. BMC Evolutionary Biology, 4: 18.
KATOH, K., MISAWA, K., KUMA, K. & MIYATA, T. (2002). MAFFT: anovel method for rapid multiple sequence alignment based on fastFourier transform. Nucleic Acid Research, 30: 3059–3066.
KIM, J.H., SHIN, M.O., LEE, K.L. & KIM, H.S. (2008). Effect ofenvironmental conditions on the growth of Synura petersenii(Synurophyceae) in vitro and two eutrophic water bodies inKorea. Nova Hedwigia, 86: 529–544.
KRISTIANSEN, J. (1975). On the occurrence of the species of Synura.Verhandlungen der Internationalen Vereinigung for Theoretischeund Angewandte Limnologie, 19: 2709–2715.
KRISTIANSEN, J. (1986). The ultrastructural bases of Chrysophytesystematics and phylogeny. CRC Critical Reviews in PlantScience, 4: 149–211.
KRISTIANSEN, J. (1992). Silica-scaled chrysophytes from WestGreenland: Disko Island and the Søndre Strømfjord region.Nordic Journal of Botany, 12: 525–536.
KRISTIANSEN, J. (2008). Dispersal and biogeography of silica-scaledchrysophytes. Biodiversity and Conservation, 17: 419–426.
KRISTIANSEN, J. & PREISIG, H.R. (2007). Chrysophyte and haptophytealgae, part 2: Synurophyceae. In Süsswasserflora vonMitteleuropa. Vol. 1 –2 (BÜDEL, B. et al., editors), 1–252.Spektrum Akademischer Verlag, Springer, Berlin.
KRISTIANSEN, J., DÜVEL, L. & WEGEBERG, S. (1997). Silica-scaledchrysophytes from the Taymyr Peninsula, Northern Siberia.Nova Hedwigia, 65: 337–351.
KUMAR, S., DUDLEY, J., NEI, M. & TAMURA, K. (2008). MEGA: abiologist-centric software for evolutionary analysis of DNA andprotein sequences. Briefings in Bioinformatics, 9: 299–306.
KYNČLOVÁ, A., ŠKALOUD, P. & ŠKALOUDOVÁ, M. (2010). Unveilinghidden diversity in the Synura petersenii species complex(Synurophyceae, Heterokontophyta). Nova Hedwigia, Beiheft,136: 283–298.
LOSIC, D., ROSENGARTEN, G., MITCHELL, J.G. & VOELCKER, N.H.(2006). Pore architecture of diatom frustules: potential nanostruc-tured membranes for molecular and particle separations. Journalof Nanoscience and Nanotechnology, 6: 1–8.
MARTINY, J.B., BOHANNAN, B.J., BROWN, J.H., COLWELL, R.K.,FUHRMAN, J.A., GREEN, J.L., HORNER-DEVINE, M.C., KANE, M.,KRUMINS, J.A., KUSKE, C.R., MORIN, P.J., NAEEM, S., OVREÅS, L.,REYSENBACH, A.L., SMITH, V.H. & STALEY, J.T. (2006). Microbialbiogeography: putting microorganisms on the map. NatureReviews, Microbiology, 4: 102–112.
MOORE, P.D. (1982). Beneath the blanket bogs of Britain. NaturalHistory, 91: 48–55.
NĚMCOVÁ, Y., NOVÁKOVÁ, S. & Řezáčová-Škaloudová, M. (2008).Synura obesa sp. nov. (Synurophyceae) and other silica-scaledchrysophytes from Abisko (Swedish Lapland). Nova Hedwigia,86: 243–254.
NEWTON, I. (2010). Bird Migration. Harper Collins, London.NICHOLLS, K.H. & GERRATH, J.F. (1985). The taxonomy of Synura(Chrysophyceae) in Ontario with special reference to tasteand odour in water supplies. Canadian Journal of Botany, 63:1482–1493.
PICHRTOVÁ, M. & NĚMCOVÁ, Y. (2011). Effect of temperature on sizeand shape of silica scales in Synura petersenii and Mallomonastonsurata (Stramenopiles). Hydrobiologia, 673: 1–11.
PLAYFAIR, G.I. (1915). Freshwater algae of the Lismore District: withan appendix on the algal fungi and Schizomycetes. Proceedings ofthe Linnean Society of New South Wales, 40: 310–362.
RABALAIS, N.N., TURNER, R.E., JUSTIC, D., DORTCH, Q., WISEMAN JR.,W.J. & SEN GUPTA, B.K. (1996). Nutrient changes in theMississippi River and system responses on the adjacent continen-tal shelf. Estuaries, 19: 386–407.
REYNOLDS, C.S. (2006). The Ecology of Phytoplankton. CambridgeUniversity Press, Cambridge.
ŘEZÁČOVÁ, M. & ŠKALOUD, P. (2005). Silica-scaled chrysophytes ofIreland. With an appendix: geographic variation of scale shape ofMallomonas caudata. Nova Hedwigia, Beiheft, 128: 101–124.
RONQUIST, F. & HUELSENBECK, J.P. (2003). MrBayes 3: Bayesianphylogenetic inference under mixed models. Bioinformatics, 19:1572–1574.
SAHA, L.C. & WUJEK, D.E. (1990). Scale-bearing chrysophytesfrom tropical Northeast India. Nordic Journal of Botany, 10:343–354.
SAXBY-ROUEN, K.J., LEADBEATER, B.S.C. & REYNOLDS, C.S. (1997).The growth response of Synura petersenii (Synurophyceae) tophoton flux density, temperature, and pH. Phycologia, 36: 233–243.
SCHNEIDER, C.A., RASBAND, W.S. & ELICEIRI, K.W. (2012). NIHImage to ImageJ: 25 years of image analysis. Nature Methods, 9:671–675.
SIVER, P.A. (1988). A new form of the common chrysophycean algaSynura petersenii. Transactions of the American MicroscopicalSociety, 107: 380–385.
SIVER, P.A. (2013): Synura cronbergiae sp. nov., a new speciesdescribed from two Paleogene maar lakes in northern Canada.Nova Hedwigia, 97: 179–187.
ŠKALOUD, P., KYNČLOVÁ, A., BENADA, O., KOFROŇOVÁ, O. &ŠKALOUDOVÁ, M. (2012). Toward a revision of the genus Synura,section Petersenianae (Synurophyceae, Heterokontophyta): mor-phological characterization of six pseudo-cryptic species.Phycologia, 51: 303–329.
ŠKALOUD, P., KRISTIANSEN, J. & ŠKALOUDOVÁ, M. (2013a).Developments in the taxonomy of silica-scaled chrysophytes –from morphological and ultrastructural to molecular approaches.Nordic Journal of Botany, 31: 385–402.
ŠKALOUD, P., ŠKALOUDOVÁ, M., PICHRTOVÁ, M., NĚMCOVÁ, Y.,KREIDLOVÁ, J. & PUSZTAI, M. (2013b). www. chrysophytes.eu – adatabase on distribution and ecology of silica-scaled chrysophytesin Europe. Nova Hedwigia, Beiheft, 142: 141–147.
SMOL, J.P. (1995). Application of chrysophytes to problems inpaleoecology. In Chrysophyte Algae. Ecology, Phylogeny andDevelopment (Sandgren, C.D., Smol, J.P. & Kristiansen, J., edi-tors), 303–330. Cambridge University Press, Cambridge.
P. Škaloud et al. 228
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

SMOL, J.P. & CUMMING, B.F. (2000). Tracking longterm changes inclimate using algal indicators in lake sediments. Journal ofPhycology, 36: 986–1011.
SWOFFORD, D.L. (2002). PAUP*. Phylogenetic Analysis UsingParsimony (*and Other Methods). Version 4. Sinauer Associates,Sunderland, MA.
VERBRUGGEN, H., MAGGS, C.A., SAUNDERS, G.W., LE GALL, L., YOON,H.S. & DE CLERCK, O. (2010). Data mining approach identifiesresearch priorities and data requirements for resolving the red algaltree of life. BMC Evolutionary Biology, 10: 16.
WEE, J. L. (1997). Scale biogenesis in Synurophycean protists:phylogenetic implications. CRC Critical Reviews in PlantSciences, 16: 497–534.
WEE, J.L., BOOTH, D.J. & BOSSIER, M.A. (1993). Synurophyceaefrom the southern Atlantic Coastal Plain of North America: apreliminary survey in Louisiana, USA. Nordic Journal ofBotany, 13: 95–106.
WEE, J.L., FASONE, L.D., SATTLER, A., STARKS, W.W. & HURLEY, D.L.(2001). ITS/5.8S DNA sequence variation in 15 isolates of Synurapetersenii Korshikov (Synurophyceae). Nova Hedwigia, Beiheft,122: 245–258.
WEST, W. & WEST, G.S. (1909). The British freshwater phytoplank-ton, with special reference to desmid-plankton and the distributionof British desmids. Proceedings of the Royal Society, London,81B: 165–206.
WHITE, T.J., BRUNS, T., LEE, S. & TAYLOR, J.W. (1990). Amplificationand direct sequencing of fungal ribosomal RNA genes for phylo-genetics. In PCR Protocols: A Guide to Methods and Applications(Innis, M.A., Gelfand, D.H., Sninsky, J.J. & White, T.J., editors),315–322. Academic Press, New York.
ZWICKL, D.J. (2006). Genetic algorithm approaches for the phylo-genetic analysis of large biological sequence datasets under themaximum likelihood criterion. PhD dissertation, University ofTexas at Austin, Austin.
Four new species of Synura 229
Dow
nloa
ded
by [
Cha
rles
Uni
vers
ity in
Pra
gue]
, [Pa
vel S
kalo
ud]
at 0
6:49
21
May
201
4

Key to species of the genus Synura, section Peterseniae
1. Base plate covered by struts extending from the keel to the scale perimeter ..................... 2
Struts missing, base plate with fine regular reticulation ............................... S. longisquama
2. Transverse struts connected in a dense net.......................................................................... 3
Transverse struts unconnected, or interconnected by single transverse struts .................... 4
3. Keel narrow, with a large base plate hole .................................................... S. macracantha
Keel extremely broad, with a very small base plate hole ......................................... S. obesa
4. Scales long and narrow (scale length to width ratio higher than 3.6) ........... S. australiensis
Scale length to width ratio less than 3.5 .............................................................................. 5
5. Scales with a very broad rim ............................................................................................... 6
Rim of scales narrower ........................................................................................................ 7
6. Struts of different length, irregularly spacered ................................................ S. asmundiae
Struts of similar length, regularly spacered ........................................................... S. bjoerkii
7. Inner portion of the rim ornamented by row of posts (in SEM) or dots (in TEM) ...............
.................................................................................................. “S. petersenii” f. columnata
Rim not ornamented ............................................................................................................ 8
8. Keel ornamented by a hexagonal pattern ............................... “S. petersenii” f. taymyrensis
Keel not ornamented ........................................................................................................... 9
9. Keel of apical scales terminates into a rounded tip, provided with a number of small teeth10
Keel of apical scales terminates into an acute tip ............................................................. 11
10. Body scales narrower, with 27-34 thickened and conspicuous struts ..................... S. truttae
Body scales broader, with 24-27 inconspicuous struts ............ “S. petersenii” f. praefracta
11. Body scales rounded to oval, struts never or very rarely connected, large base-plate pores
(diameter 25-78 nm) .......................................................................................................... 12

Body scales lanceolate, struts frequently connected by transverse ribs, small base-plate
pores (diameter 17-31 nm) ................................................................................................ 15
12. Rear scales longer than body scales .................................................................................. 13
Rear scales short ................................................................................................................ 14
13. Base-plate pores very large (diameter 50-78 nm) ............................................ S. macropora
Base-plate pores smaller (diameter 27-43 nm) ................................................ S. americana
14. Keel and ribs less developed, scales often almost rounded .................................... S. glabra
Keel and ribs well developed, scales oval ............................................................ S. conopea
15. Keel anteriorly greatly widened, with the over-layered pore pattern (in TEM) ............... 16
Keel more or less cylindrical, ornamented by a single layer of pores .............................. 17
16. Body scales short (length 3.1-4.3 m), cells shorter than 32 m ......................S. laticarina
Body scales long (length 4.0-5.8 m), cells longer than 32 m .......................... S. borealis
17. Keel anteriorly slightly widened, scale width up to 1.8(-2)m........................................ 18
Keel cylindrical, scale width greater than (1.7-)1.9m ..................................... S. petersenii
18. Body scales short (length 2.5-3.8 m), with 22-28 struts ................................ S. heteropora
Body scales long (length 3.4-5.6 m), with 30-47 struts ................................... S. hibernica

ITS
rbc
Lc
ox
1
Syn
ura
am
eric
an
a K
yn
člo
vá
& Š
ka
lou
dC
CM
P 8
62
Win
ter’s
Cre
ek, M
I, US
AG
U3
38
12
4G
U3
25
48
5G
U2
95
52
9
Syn
ura
am
eric
an
a K
yn
člo
vá
& Š
ka
lou
dC
CM
P 8
63
roa
d d
itch
ne
ar W
inte
r’s C
ree
k, M
I, US
AG
U3
38
12
5G
U3
25
48
6G
U2
95
53
0
Syn
ura
am
eric
an
a K
yn
člo
vá
& Š
ka
lou
dC
CM
P 8
66
New
fou
nd
lan
d, C
an
ad
aA
F3
08
84
0-
-
Syn
ura
am
eric
an
a K
yn
člo
vá
& Š
ka
lou
dU
TE
X L
B 2
40
4S
po
rtsm
an
’s L
ake
, WA
, US
AG
U3
38
13
2G
U3
25
49
5G
U2
95
53
8
Syn
ura
am
eric
an
a K
yn
člo
vá
& Š
ka
lou
dS
81
.C7
Zá
pla
vy N
R, C
ze
ch
Re
pu
blic
HG
51
41
66
--
Syn
ura
am
eric
an
a K
yn
člo
vá
& Š
ka
lou
dS
10
4.C
4S
ch
inke
lbo
s, N
eth
erla
nd
sH
G5
14
16
7-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 5
8.C
7L
illesjö
n, S
we
de
nH
G5
14
16
8H
G5
14
23
4H
G5
14
25
5
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 9
0.F
3R
uta
järv
i, Harju
lah
ti, Fin
lan
dH
G5
14
16
9-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 9
0.G4
Harju
järv
i, Fin
lan
dH
G5
14
17
0-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
10
.B9
Ap
na
jõg
i, Esto
nia
HG
51
41
71
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
10
.E2
a s
ma
ll po
ol n
ea
r Ka
lli jõg
i, Esto
nia
HG
51
41
72
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
16
.B6
Kje
rring
da
lsva
tne
t, No
rwa
yH
G5
14
17
3-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
SC
FIN
13
AO
jala
, Fin
lan
dH
G5
14
17
4-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
SC
SW
E 9
Helg
assjö
n, S
we
de
nH
G5
14
17
5-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 6
2.D
7a
sm
all la
ke
, Dis
ko
isla
nd
, Gre
en
lan
dH
G5
14
17
6H
G5
14
23
5H
G5
14
25
6
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
SC
GR
L 3
64
a m
orra
ine
lake
, Dis
ko
isla
nd
, Gre
en
lan
dH
G5
14
17
7-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 9
0.M
34
Jä
msä
nja
rvi, F
inla
nd
HG
51
41
78
HG
51
42
36
HG
51
42
57
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
10
.F3
a s
ma
ll po
ol n
ea
r Ka
lli jõg
i, Esto
nia
HG
51
41
79
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
13
.F6
Em
ajõ
gi riv
er, E
sto
nia
HG
51
41
80
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
15
.G2
a p
oo
l ne
ar V
assija
ure
, Sw
ed
en
HG
51
41
81
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
18
.C6
a p
oo
l ne
ar T
orn
eträ
sk, S
we
de
nH
G5
14
18
2-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
13
.F4
Em
ajõ
gi riv
er, E
sto
nia
HG
51
41
83
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
14
.B8
un
na
me
d la
ke
, Sw
ed
en
HG
51
41
84
HG
51
42
37
HG
51
42
58
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
14
.C8
un
na
me
d la
ke
, Sw
ed
en
HG
51
41
85
HG
51
42
38
HG
51
42
59
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
14
.B9
un
na
me
d la
ke
, Sw
ed
en
HG
51
41
86
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
14
.F3
un
na
me
d la
ke
ne
ar P
aitta
sjä
rvi, S
we
de
nH
G5
14
18
7-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
17
.D3
a p
oo
l ne
ar P
aitta
sjä
rvi, S
we
de
nH
G5
14
18
8H
G5
14
23
9H
G5
14
26
0
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
SC
GR
L 3
40
Sa
nn
ing
assu
p T
asia
, Gre
en
lan
dH
G5
14
18
9-
-
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
14
.G6
Syvä
järv
i, Sw
ed
en
HG
51
41
90
HG
51
42
40
HG
51
42
61
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
15
.F4
a p
oo
l ne
ar A
rosn
jark
aja
ute
, Sw
ed
en
HG
51
41
91
HG
51
42
41
HG
51
42
62
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
15
.G3
a p
oo
l ne
ar V
assija
ure
, Sw
ed
en
HG
51
41
92
--
Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
S 1
15
.G7
Va
ssija
ure
, Sw
ed
en
HG
51
41
93
HG
51
42
42
HG
51
42
63
Sy
nu
ra a
me
rica
na
Ta
xo
nS
train
nu
mb
er
Orig
inG
en
Ba
nk
ac
ce
ss
ion
Sy
nu
ra b
ore
alis

Syn
ura
bo
rea
lis Š
ka
lou
d &
Ška
lou
do
vá
SC
FIN
22
BH
ein
äjä
rvi, F
inla
nd
HG
51
41
94
--
Syn
ura
co
no
pe
a K
yn
člo
vá
& Š
ka
lou
dC
CM
P 8
59
so
uth
ea
st b
ridg
e p
on
d, A
R, U
SA
GU
33
81
21
GU
32
54
82
GU
29
55
26
Syn
ura
co
no
pe
a K
yn
člo
vá
& Š
ka
lou
dS
10
.2H
uťs
ký p
on
d, C
ze
ch
Re
pu
blic
FM
17
85
07
--
Syn
ura
co
no
pe
a K
yn
člo
vá
& Š
ka
lou
dN
IES
10
07
To
ma
ko
ma
i, Ja
pa
nG
U3
38
11
9G
U3
25
47
9G
U2
95
52
4
Syn
ura
co
no
pe
a K
yn
člo
vá
& Š
ka
lou
dK
NU
13
Dals
eo
ng
gyo
, Ko
rea
GU
33
80
69
GU
32
54
30
GU
29
54
72
Syn
ura
co
no
pe
a K
yn
člo
vá
& Š
ka
lou
dC
AU
P B
70
7 (S
7.1
0)
Ba
bín
po
ol, C
ze
ch
Re
pu
blic
FM
17
85
06
--
Syn
ura
co
no
pe
a K
yn
člo
vá
& Š
ka
lou
dS
10
3.B
3a
po
ol n
ea
r Sm
ěd
ava
, Cze
ch
Re
pu
blic
HG
51
41
95
--
Syn
ura
gla
bra
Ko
rsh
iko
vN
IES
23
3H
iga
sh
iya
ta R
ive
r, Ja
pa
nG
U3
38
11
8G
U3
25
48
0G
U2
95
52
3
Syn
ura
gla
bra
Ko
rsh
iko
vC
NU
09
Jio
kji p
on
d, K
ore
aG
U3
38
07
7G
U3
25
43
8G
U2
95
48
0
Syn
ura
gla
bra
Ko
rsh
iko
vK
NU
10
Bo
mu
n re
se
rvo
ir, Ko
rea
GU
33
80
66
GU
32
54
27
GU
29
54
69
Syn
ura
gla
bra
Ko
rsh
iko
vS
9.2
co
nflu
en
ce
of th
e M
ora
va
an
d D
yje
rive
rs, C
ze
ch
Re
pu
blic
FM
17
85
13
--
Syn
ura
gla
bra
Ko
rsh
iko
vS
14
.1S
wa
mp
NR
, Cze
ch
Re
pu
blic
FM
17
85
14
--
Syn
ura
gla
bra
Ko
rsh
iko
vS
89
.F5
Sa
arijä
rvi, F
inla
nd
HG
51
41
96
--
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
CC
MP
28
98
Wa
llers
ee
, Au
stria
GU
33
81
36
GU
32
54
98
-
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 2
0.1
Lo
ch
an
Ad
d, S
co
tlan
d, U
KH
G5
14
19
7-
-
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 2
0.4
5C
rina
n C
an
al, S
co
tlan
d, U
KH
G5
14
19
8H
G5
14
24
3-
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 4
0.F
11
Po
dh
rad
ská
po
ol, C
ze
ch
Re
pu
blic
HG
51
41
99
--
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 5
4.E
11
Kle
jna
rka
rive
r, Cze
ch
Re
pu
blic
H
G5
14
20
0-
-
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 8
6.F
2Z
býšo
v p
on
d, C
ze
ch
Re
pu
blic
HG
51
42
01
--
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 1
12
.E2
Sa
ard
e p
ais
järv
, Esto
nia
HG
51
42
02
--
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 1
12
.F5
Rah
um
eri, E
sto
nia
HG
51
42
03
--
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 1
13
.C8
Ka
rula
Ja
rv, E
sto
nia
HG
51
42
04
--
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 1
17
.G6
To
rne
träsk, S
we
de
nH
G5
14
20
5-
-
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
SC
IRL
8G
arla
n L
ou
gh
, Irela
nd
HG
51
42
06
--
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 8
7.C
6a
n e
ph
em
era
l pu
dd
le n
ea
r Ku
fste
in, A
ustria
HG
51
42
07
HG
51
42
44
HG
51
42
64
Syn
ura
he
tero
po
ra Š
ka
lou
d, Š
ka
lou
do
vá
& K
yn
člo
vá
S 1
01
.F7
a p
on
d in
Nyg
ård
sp
ark
en
, Be
rge
n, N
orw
ay
HG
51
42
08
HG
51
42
45
HG
51
42
65
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
E4
Th
e L
on
g R
an
ge
, Irela
nd
HG
51
42
09
HG
51
42
46
HG
51
42
66
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
E1
1G
len
do
llag
h L
ou
gh
, Irela
nd
HG
51
42
10
--
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
10
3.C
11
Go
wla
un
Lo
ug
h, Ire
lan
dH
G5
14
21
1-
-
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
SC
IRL
41
Ea
sky L
ou
gh
, Irela
nd
HG
51
42
12
--
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
E8
un
na
me
d la
ke
ne
ar M
aa
m C
ross, Ire
lan
dH
G5
14
21
3H
G5
14
24
7H
G5
14
26
7
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
10
3.C
8G
ow
lau
n L
ou
gh
, Irela
nd
HG
51
42
14
--
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
10
3.C
9G
ow
lau
n L
ou
gh
, Irela
nd
HG
51
42
15
--
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
10
4.D
11
Gla
nm
ore
La
ke
, Irela
nd
HG
51
42
16
HG
51
42
48
HG
51
42
68
Sy
nu
ra g
lab
ra
Sy
nu
ra c
on
op
ea
Sy
nu
ra h
ibe
rnic
a
Sy
nu
ra h
ete
rop
ora

Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
M3
8T
he
Lo
ng
Ra
ng
e, Ire
lan
dH
G5
14
21
7-
-
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
SC
IRL
20
aD
erry
na
he
rriva
Lo
ug
h, Ire
lan
dH
G5
14
21
8-
-
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
S IE
10
5.F
6C
ah
a L
ake
s, Ire
lan
dH
G5
14
21
9H
G5
14
24
9H
G5
14
26
9
Syn
ura
hib
ern
ica
Ška
lou
d &
Ška
lou
do
vá
SC
IRL
60
Ma
um
we
e L
ou
gh
, Irela
nd
HG
51
42
20
--
Syn
ura
latic
arin
a Š
ka
lou
d &
Ška
lou
do
vá
S 8
9.D
5O
jala
, Fin
lan
dH
G5
14
22
1-
-
Syn
ura
latic
arin
a Š
ka
lou
d &
Ška
lou
do
vá
S 9
0.C
8T
eh
rise
lkä
, Fin
lan
dH
G5
14
22
2H
G5
14
25
0H
G5
14
27
0
Syn
ura
latic
arin
a Š
ka
lou
d &
Ška
lou
do
vá
S 1
10
.C9
Ap
na
jõg
i, Esto
nia
HG
51
42
23
--
Syn
ura
latic
arin
a Š
ka
lou
d &
Ška
lou
do
vá
S 1
13
.E5
a p
oo
l ne
ar E
ma
jõg
i, Esto
nia
HG
51
42
24
--
Syn
ura
latic
arin
a Š
ka
lou
d &
Ška
lou
do
vá
S 1
15
.B2
Ba
rdu
elv
a riv
er, N
orw
ay
HG
51
42
25
--
Syn
ura
latic
arin
a Š
ka
lou
d &
Ška
lou
do
vá
S 1
15
.D2
a p
oo
l ne
ar A
rosn
jark
aja
ute
, Sw
ed
en
HG
51
42
26
HG
51
42
51
HG
51
42
71
Syn
ura
latic
arin
a Š
ka
lou
d &
Ška
lou
do
vá
S 1
15
.E5
a p
oo
l ne
ar V
assija
ure
, Sw
ed
en
HG
51
42
27
--
Syn
ura
ma
cro
po
ra Š
ka
lou
d &
Kyn
člo
vá
S 5
.3a
lluvia
l po
ol, P
rag
ue
, Cze
ch
Re
pu
blic
FM
17
84
96
--
Syn
ura
ma
cro
po
ra Š
ka
lou
d &
Kyn
člo
vá
S 1
4.2
Sw
am
p N
R, C
ze
ch
Re
pu
blic
FM
17
84
97
--
Syn
ura
ma
cro
po
ra Š
ka
lou
d &
Kyn
člo
vá
S 7
1.B
4P
od
hra
dská
po
ol, C
ze
ch
Re
pu
blic
HG
51
42
28
HG
51
42
52
HG
51
42
72
Syn
ura
ma
cro
po
ra Š
ka
lou
d &
Kyn
člo
vá
S 1
04
.D7
a c
an
al in
Ooste
ind
e, N
eth
erla
nd
sH
G5
14
22
9-
-
Syn
ura
ma
cro
po
ra Š
ka
lou
d &
Kyn
člo
vá
S 1
04
.F1
1a
po
ol in
Aa
lsm
ee
r, Neth
erla
nd
sH
G5
14
23
0-
-
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vA
CO
I 17
07
Ab
ran
tes, P
ortu
ga
lG
U3
38
15
0G
U3
25
51
2G
U2
95
55
2
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vC
CA
P 9
60
/3P
riest P
ot, E
ng
lan
d, U
KG
U3
38
14
3G
U3
25
50
5G
U2
95
54
5
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vS
6.4
Horn
i Lu
zn
ice
NR
, Cze
ch
Re
pu
blic
FM
17
85
01
--
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vC
CM
P 8
60
Ced
ar L
ake
, IL, U
SA
GU
33
81
22
GU
32
54
83
GU
29
55
27
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vS
AG
12
0.7
9L
ün
eb
urg
er H
eid
e, M
ea
do
w p
oo
l, Ge
rma
ny
GU
33
81
44
GU
32
55
06
GU
29
55
46
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vC
AU
P B
70
3 (S
7.7
)B
ab
ín p
on
d, C
ze
ch
Re
pu
blic
FM
17
85
04
--
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vS
89
.D6
Oja
la, F
inla
nd
HG
51
42
31
--
Syn
ura
pe
ters
en
ii Ko
rsh
iko
vS
89
.F9
Hein
äjä
rvi, F
inla
nd
HG
51
42
32
--
Syn
ura
trutta
e (S
ive
r) Ška
lou
d &
Kyn
člo
vá
Nem
co
va
D5
Ka
ch
ní p
oto
k, C
ze
ch
Re
pu
blic
GU
33
81
40
GU
32
55
02
GU
29
55
42
Syn
ura
trutta
e (S
ive
r) Ška
lou
d &
Kyn
člo
vá
S 1
5.3
Úpské
raše
liniš
tě, C
ze
ch
Re
pu
blic
FM
17
85
08
--
Syn
ura
trutta
e (S
ive
r) Ška
lou
d &
Kyn
člo
vá
S 1
5.9
Úpské
raše
liniš
tě, C
ze
ch
Re
pu
blic
FM
17
85
10
--
Syn
ura
trutta
e (S
ive
r) Ška
lou
d &
Kyn
člo
vá
CA
UP
B7
05
(S 3
4.1
)fo
rest p
oo
l ne
ar M
ezn
í lou
ka
, Cze
ch
Re
pu
blic
FR
81
97
49
HG
51
42
53
-
Syn
ura
trutta
e (S
ive
r) Ška
lou
d &
Kyn
člo
vá
S 6
2.B
5a
pe
at b
og
ne
ar P
řeb
uz, C
ze
ch
Re
pu
blic
HG
51
42
33
HG
51
42
54
-
Syn
ura
sp
.C
NU
01
Sa
mh
wa
po
nd
, Ko
rea
GU
33
80
70
GU
32
54
31
GU
29
54
73
Syn
ura
sp
.C
NU
10
Bih
aksa
n, K
ore
aG
U3
38
07
8G
U3
25
43
9G
U2
95
48
1
Syn
ura
sp
.C
NU
12
Ge
om
uye
okri, K
ore
aG
U3
38
08
0G
U3
25
44
1G
U2
95
48
3
cla
de
D
cla
de
F
Sy
nu
ra p
ete
rse
nii
Sy
nu
ra la
tica
rina
Sy
nu
ra m
ac
rop
ora
Sy
nu
ra tru
ttae

Syn
ura
sp
.C
NU
47
Se
on
gd
on
g p
on
d, K
ore
aG
U3
38
11
4G
U3
25
47
5G
U2
95
51
8
Syn
ura
sp
.K
NU
01
Uksu
rese
rvo
ir, Ko
rea
GU
33
80
57
GU
32
54
18
GU
29
54
60
Syn
ura
sp
.U
TE
X L
B 2
39
un
kn
ow
nG
U3
38
13
5G
U3
25
49
3G
U2
95
53
6
Syn
ura
sp
.C
CA
C 0
05
2C
olo
gn
e, G
erm
an
yG
U3
38
14
6G
U3
25
50
8G
U2
95
54
8
Syn
ura
sp
.C
CM
P 8
69
To
ba
cco
Riv
er, M
I, US
AG
U3
38
12
8G
U3
25
48
9G
U2
95
53
3
oth
ers