mismatch_cleavage_by_cel-1_1__final
TRANSCRIPT

Mismatch Cleavage by CEL-I Endonuclease: A Tool for Rapid Detection 1
of Homozygous and Heterozygous Mutants
Mismatch Cleavage by CEL-I Endonuclease:
A Tool for Rapid Detection of Homozygous
and Heterozygous Mutants
Sulabha Sharma1, Kamal Tyagi2, M Lakshmi Narasu3, Y Sreelakshmi4 and Rameshwar Sharma5
In recent years, mutants have served as useful starting material to unravel the mechanisms
governing many biological processes, including both plants and animals. Mainly, there are
three ways of inducing mutations; by using physical agents (radiations UV, X-rays, -rays),
chemical agents (Ethyl Methane Sulphonate (EMS), Nitrous Oxide (NO), acridine, etc.) and
biological agents (transposon and T-DNA). The mutation induced by these agents lead to
isolation of most commonly recessive or uncommonly dominant mutants. The isolation and
screening of a mutant are tedious and cumbersome job and even after isolation, ascertaining
the homozygous nature of the plant is equally difficult, especially for a dominant mutant.
The commonly known methods for screening of homo/heterozygosity of the mutant are
either highly expensive as sequencing of the gene or involve very tedious/time-consuming
genetic segregation analysis. Here, we present a rapid tool for identifying the homo/
heterozygous nature of the plants by simple digestion of PCR-amplified genomic DNA with
CEL-I endonuclease and resolving the cleaved product on conventional agarose gels.
This method is quite robust and provides a great help to researchers in developing countries
where instrumentation access is limited.
Keywords: Homozygosity, Heterozygosity, Homoduplex, Heteroduplex, CEL-I Endonuclease, Mutant,
Agarose
Introduction
Plant mutants have been used in genetic studies and breeding for decades, yet huge
number of mutants remains to be characterized at the molecular level. In recent years,
the information of genome sequence has become available for several plant species but
the functions of many of the genes are not known. The analysis of well-characterized
mutants using saturated mutant populations, coupled with recent methods for the detection
1 Sulabha Sharma (Researcher), Department of Plant Sciences, School of Life Sciences, Hyderabad Central University, Hyderabad
500046, India. E-mail: m a i l 2 s u l a b h a @ g m a i l . c o m
2 Kamal Tyagi, Department of Plant Sciences, School of Life Sciences, Hyderabad Central University, Hyderabad
500046, India. E-mail: [email protected]
3 Professor. M. Lakshmi Narasu ( Director), Institute of Science and Technology, Department of Biotechnology, Jahawarlal Nehru Technological
University, Hyderabad, India. E-mail: [email protected]
4 Dr. Y. Sreelakshmi (Associate Professor), Department of Plant Sciences, School of Life Sciences, Hyderabad Central University, Hyderabad
500046, India. E-mail: [email protected]
5 Professor. Rameshwar Sharma (Professor), Department of Plant Sciences, School of Life Sciences, Hyderabad Central University, Hyderabad
500046, India; and is the corresponding author. E-mail: [email protected]
© 2011 IUP. All Rights Reserved.

and generation of mutants are bridging the gap between plant genes and their functions.
There are basically two ways to analyze function of a mutated gene: reverse and forward
genetics (Caldwell et al., 2004; Jose and Joseph, 2006; and Christian et al., 2009). In reverse
genetic approaches, one starts with a (sequenced) gene of interest, selects a mutation in
that gene, and then tries to identify a phenotypic change associated with mutation.
In forward genetic approaches (Caldwell et al., 2004), one begins with a prediction of
specific effect of a mutation for physiological and morphological processes and then
isolates mutants with the predicted phenotype followed by mapping and isolation of the
genetic sequence that determines the above phenotype. Thus, contrary to reverse genetics
(Jansen et al., 1997; Sessions et al., 2002; Jose and Joseph, 2006; and Christian et al., 2009),
forward genetics starts with phenotyping of mutants and later identifies the gene
responsible for the altered phenotype and both the approaches are valuable and
complementary. Once corres ponding gene linked to a mutation is identified, the
introgression of the gene in a new cultivar for breeding requires at least two backcrosses
for removal of background mutations and elimination of genotype of donor plant. Moreover,
the essential confirmation that a phenotype of interest results from a given mutation
though can be achieved via complementation testing, which is also used for determining
allelism of recessive mutations, such confirmation is not possible in case of dominant
mutants. In addition, once the mutant is isolated the next requirement is to maintain the
purity of the mutant, i.e., homozygous nature, which is slightly difficult in case of dominant
mutant (Jack et al., 1997), where the mutant shows same phenotype in both homozygous
and heterozygous forms. Likewise, if the population of different mutants were grown
together, there are always chances of cross pollination from neighboring plants and
heterozygous progeny may result. Tomato crop grows well under warm climate, but
cannot be grown under temperate climate. As a consequence in temperate climate
tomatoes are grown densely in green houses (Liu et al., 2004; and Menda et al., 2004).
In such growth conditions, close proximity of plants also increases the chances of cross
pollination leading to heterozygous population.
In mutation breeding experiments, it is essential that the plant materials be genetically
pure and uniform for the traits to be examined and that pollination be rigidly controlled
both prior to and during the experiment to prevent outcrossing. The identification of
homozygous wild type, heterozygous and homozygous mutant plants, can be done by
normal genetic crossing experiments. However, it involves considerable time and money
for genetically screening of mutant. Moreover, in crops, it is difficult to distinguish between
homozygous dominant or heterozygous dominant mutations. In procedures, that assist
in the identification of mutations include growing progeny of suspected mutants and
observing whether segregation occurs or back crossing the suspected mutant to the
parent followed by selfing or sibling of the progeny of a true recessive mutant.
Apart from genetic screening, another technique for the confirmation of mutation
identity is sequencing of the mutated gene. But the cost of sequencing is not affordable
to every laboratory or company, particularly, when a large plant population of at least 100
2 The IUP Journal of Genetics & Evolution, Vol. IV, No. 2, 2011

to 200 plants need to be analyzed for homozygosity or heterozygosity of mutation.
Therefore, there is an enormous potential for simple and fast methods, which can allow
one to distinguish homo/heterozygosity of mutants efficiently using in house facility.
Recently, use of mismatch specific endonuclease CEL-I (Oleykowski et al., 1998; Till et al.,
2004a; and Yeung et al., 2005) has been extensively applied for TILLING, a reverse
genetic method for mutant isolation. Currently, the use of above endonuclease enzymes
is being made in numerous approaches like TILLING ECOTILLING, SNP detection, etc.
(Peterson et al., 1997; McCallum et al., 2000; Colbert et al., 2001; Henikoff and Comai,
2003; Perry et al., 2003; Till et al., 2004b; Gilchrist and Haughn, 2005; Altmann et al., 2007;
and Raghavan et al., 2007). Here we describe the use of CEL-I endonuclease enzyme
(Oleykowski et al.1998; and Till et al., 2004a) for detection of homo/heterozygous
mutants in a pool of mixed population. This method can be easily carried out in a
breeding lab with simple PCR machine and conventional gel electrophoresis apparatus.
The above tool can detect both homozygous and heterozygous mutants in a cost-
effective manner. This method is also quite convenient for detecting dominant
mutations (Wilson et al., 1990) where it is difficult to differentiate between homozygous
and heterozygous mutants. Moreover, this method can be applied to any organism right
from bacteria to higher eukaryotes for any gene, whose gene sequence is known.
Materials and Methods
Plant Material and Growth Conditions
The tomato mutant population and wild type were grown in green houses, net houses,
growth chambers in confined areas to avoid adverse climatic conditions and even protect
crop from damage caused by birds and animals. The plants used for screening of
homozygous and heterozygous mutation from a pool of mixed population were Solanum
lycopersicon. Mill cv Ailsa Craig (wild type), Money maker, (wild type), fri (far red insensitive
mutant) (Galina et al., 1998) obtained from Prof. M Korneeff and Prof. R E Kendrick,
University of Wageningen, Netherlands in 1992 and multiplied in green houses at
University of Hyderabad.
Plant Genomic DNA Isolation
Extraction of DNA was carried out, as described earlier (Sreelakshmi et al., 2010) with
slight modifications. The leaf tissue (200-300 mg) was homogenized with three steel
balls in microcentrifuge tube in Mini-Bead Beater for 2 min in the presence of 1,000 L
preheated (65 °C) extraction buffer (0.1 M Tris-HCl, pH 7.5, 0.05 M EDTA, pH 8.0; 1.25%
( w / v ) SDS ) c o n t a in in g 0 .2 M - me rc a pt o et h a n o l a n d 3 0 mg o f i n s o l u b l e
polyvinylpolypyrrolidone (PVPP). The tubes were incubated at 65 °C for 30 min for cell
disruption. The residual RNA was degraded by adding 1 L from 10 mg/mL stock and
incubated at 37 °C in a water bath for 30 min. The protein precipitation was done by
the addition of 600 L of 6 M ammonium acetate and incubating the samples at 4 °C for
15 min. The precipitated proteins along with other cellular debris were pelleted by
centrifugation at 13,000 rpm for 10 min and the clear supernatant (800 L) containing
Mismatch Cleavage by CEL-I Endonuclease: A Tool for Rapid Detection 3
of Homozygous and Heterozygous Mutants

2
DNA in aqueous phase was transferred into a fresh microcentrifuge tube. The nucleic
acid was precipitated by adding chilled 700 L of isopropanol and precipitation was
maximized by incubating the sample at –20 °C for 30 min. The DNA was pelleted by
centrifugation at 13,000 rpm for 10 min, and tubes were dec anted and residual
isopropanol was replaced and salts were removed by washing the pellet with 70% (v/v)
ethanol and finally pellets were air-dried and dissolved in 200 L TE (10 mM Tris, pH
7.5, 1 mM EDTA pH 8.0). The estimation of the yield of isolated DNA samples was done
spectrophotometrically by Nanodrop-1,000 and by electrophoresis on 1% (w/v) agarose
gel by comparative fluorescence quantification of ethidium bromide stained bands using
quantified standard DNA.
Primer Design and Amplification
Fo r wa rd ( 5'G A AG GATG ATG GC A GGA A A AT GC3 ') and rev ers e primers
(5'CACTCAGAAACACCAGCCA A ATTG3') were designed to amplif y a 982 bp region of
plant photoreceptor protein, phytochrome A defective mutant (far red insensitive) from
fri mutant of tomato. The mutant, fri has transition of A to T at the 3' end of the
in tron between exons 1 and 2, in the PhyA gen e of lengt h 6623 bp (AJ0 01913).
Upon digestion with CEL-I enzyme, it generated two fragments of 710 bp and 272 bp.
Amplification reaction was set up in a volume of 10 L with 25-30 ng of DNA having
fri mutant DNA in each of the samples. The reaction consisted of 5 L of template, IX
PCR buffer (10 mM Tris, 5 mM KCl, 1.5 mM MgCl , 0.1% (w/v) gelatin, 0.005% (v/v)
Tween-20, 0.005% (v/v) Np-40, pH 8.8), 0.2 mM dNTPs, 0.5 L Taq polymerase
(in-house isolated) and 5 pmoles of primers. The cycling conditions for amplification
were 94 °C-5 min, 35 cycles of 94 °C-20 s, 55 °C-30 s, 72 °C-30 s followed by elongation
at 72 °C for 8 min.
Heteroduplex Formation
The heteroduplex (Ruano and Kidd, 1992) was prepared by adding equal amount of WT
and fri PCR products and treating at 98 °C for 8 min; 80 °C for 20 s; 60 cycles of
80 °C for 7 s –0.3°/cycle (Colbert et al., 2001). The PCR product was digested as described
by Till et al. (2004a) using CEL-I endonuclease (Yeung et al., 2005). After mixing on ice,
the microcentrifuge tubes were incubated at 45 °C for 15 min and the digestion was
stopped by addition of 2 L of 75 mM EDTA.
Mismatch Cleavage and Mutant Detection
CEL-I was isolated from celery as previously described. The mismatch cleavage reaction
was performed in a total volume of 45 L containing 10 pi PCR product, IX CEL-I digestion
buffer (10 mM HEPES buffer pH 7.0, 10 mM KC1, 10 mM MgCl2, 0.002% (v/v) Triton X-100
and 10 g/mL BSA) and CEL-I enzyme at 1:300 dilution (1 L/300 L CEL-I digestion buffer).
The reaction was incubated at 45 °C for 15 min and then terminated by adding 2 L EDTA
(75 mM EDTA, pH 8.0). Later, the digestion mixture was run on 2% agarose gel, and the gel
was visually assessed for mutations.
4 The IUP Journal of Genetics & Evolution, Vol. IV, No. 2, 2011
of Homozygous and Heterozygous Mutants
Results and Discussion
Low Cost Genomic DNA Isolation
A cost-effective, efficient and high throughput method for genomic DNA isolation from
plants was used (Lin and Kuo, 1998; Kang and Yang, 2004; and Sreelakshmi et al., 2010)
that was specifically designed for high throughput DNA isolation from large number of
plants for TILLING. The basis of this methodology lies on good yield and quality of isolated
DNA along with use of inexpensive chemicals, and minimum time required for complete
DNA isolation (Sreelakshmi et al., 2010). In the present protocol, we modified few steps
as DNA extraction was carried out in microcentrifuge tubes/Eppendorf tubes leading to
the higher yield. One of the advantages of higher yield was that the same DNA after
confirming the homozygous nature of the mutant plant can also be used for other
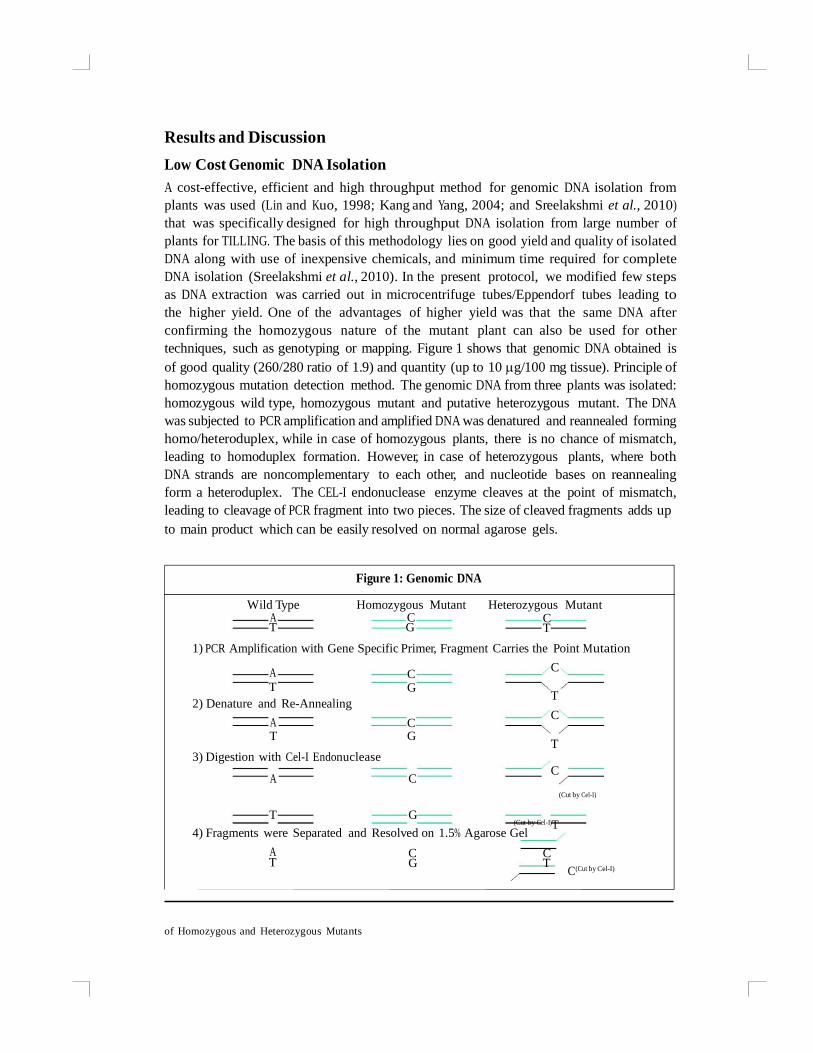
techniques, such as genotyping or mapping. Figure 1 shows that genomic DNA obtained is
of good quality (260/280 ratio of 1.9) and quantity (up to 10 g/100 mg tissue). Principle of
homozygous mutation detection method. The genomic DNA from three plants was isolated:
homozygous wild type, homozygous mutant and putative heterozygous mutant. The DNA
was subjected to PCR amplification and amplified DNA was denatured and reannealed forming
homo/heteroduplex, while in case of homozygous plants, there is no chance of mismatch,
leading to homoduplex formation. However, in case of heterozygous plants, where both
DNA strands are noncomplementary to each other, and nucleotide bases on reannealing
form a heteroduplex. The CEL-I endonuclease enzyme cleaves at the point of mismatch,
leading to cleavage of PCR fragment into two pieces. The size of cleaved fragments adds up
to main product which can be easily resolved on normal agarose gels.
Figure 1: Genomic DNA
Wild Type Homozygous Mutant Heterozygous Mutant A C C T G T
1) PCR Amplification with Gene Specific Primer, Fragment Carries the Point Mutation
A C C
T G T
2) Denature and Re-Annealing
A C C
T G T
3) Digestion with Cel-I Endonuclease
A C C
(Cut by Cel-I)
T G (Cut by Cel-I)T
4) Fragments were Separated and Resolved on 1.5% Agarose Gel
A C C T G T
C(Cut by Cel-I)

PCR Quality and Quantity
To take advantage of agarose gel detection, special attention should be given towards
the quality and quantity of PCR products. Initially, it is of great importance to amplify
specific and single PCR product from a PCR reaction since the presence of nonspecific
products may lead to heteroduplex formation. The nonspecific amplification would easily
result in the detection of varied CEL-I cleaved products (Ruano and Kidd, 1992; and
Oleykowski et al., 1998). Though here we are aware about the site of mutation in the
gene and the size of expected cleaved products, still complications can easily be avoided
by the single and specific PCR product. Secondly, the yield of PCR product should be
higher since the sensitivity of detection on agarose gel is little low. The PCR products
less than 10 ng/L are not normally detected on conventional agarose gel. A good yield of
PCR product is required to obtain better resolution of both the cleaved products on the
regular agarose gels.
Large-Scale Population Screening for Homozygous and Heterozygous
Mutants in Mixed Crop
In the current era of genomics, plant biologists are striving to understand the function
of all the plant genes. The function of a particular gene can be best determined when
the gene of interest is mutated and the resulting mutant is analyzed for differences
compared to its wild type. The isolation of mutants either by forward genetics or
reverse genetics (Caldwell et al., 2004; and Christian et al., 2009) is equally tedious and
time-consuming. However, maintain ing homozygo sit y of mutants in subsequen t
generations is equally difficult, more so for dominant mutants (Wilson et al., 1990) and
also for those crops where cross pollination is feasible. Therefore, having a tool to
identif y homo/heterozygosity is of great advantage. Here, we adopted a use of CEL-I
endonuclease enzyme (Oleykowski et al., 1998; and Till et al., 2004) for detection of
homozygous and heterozygous mutants in a pool of mixed population. This method is
cost-effective and institutes or small companies with just a PCR facility can deploy this
method (Figure 2). The basic methodo logy is demo nstrated using a phytochro meA
receptor defective mutant in tomato (fri, far red insensitive) (Galina et al., 1998). The
cartoon depicts the methodology in Figure 3. The DNA from wild type and fri mutant
was PCR amplified using the phytochrome A gene specific primers and subjected to
mutation detection using the standard protocol of TILLING (Till et al., 2004). The
mismatch cleavage by CEL-I cuts the heteroduplex DNA fragment of 982 bp size at the
mismatch site, releasing two fragments of 272 bp and 710 bp (Figure 4). The specificity
of the reaction was confirmed since the cut bands were present only in samples where
genomic DNA from wild type and fri were combined to make heteroduplex. In contrast,
samples with either wild type or fri mutant DNA alone, no complementary fragments
were released, on CEL-I digestion. The resultant PCR product of homozygous mutant
releases a band on CEL-I digestion, only when it was mixed with WT PCR products to
make heteroduplex. In all other samples, cleavage of CEL-I endonuclease shows no cut
fragments, as there was no heteroduplex formation (Figure 5).
6 The IUP Journal of Genetics & Evolution, Vol. IV, No. 2, 2011

of Homozygous and Heterozygous Mutants
Hom
ozy
gous
WT
Hom
ozy
gous
frt
Dig
este
d W
T/f
ri
Un
dig
este
d W
T/f
ri
M
Figure 2: Plant Genomic DNA Isolation from a Mixed Population of Homozygous
and Heterozygous Plants
A
B
Note: The protocol used for tomatoplant genomic DNA isolation is highly cost-effective and adapted from
our previous publication.
Source: Sreelakshmi et al. (2010)
Figure 3: Gene-Specific PCR Amplification Using in House Isolated Taq DNA Polymerase
Note: The genomic DNA isolated from homo/heterozygous mixed population was used for PCR amplification.
The PCR amplified bands were resolved on 1% agarose gel. Amplification of the PCR product without
any nonspecific amplification is important for this technique.
Figure 4: Detection of Mutation in PHYA Gene of Fri mutant
by Mismatch Cleavage Using CEL-I
Nonspecific Band
0.98 Kb 0.71 Kb
0.27 Kb
1.0 Kb 0.3 Kb
Note: Homoduplex PCR product is ca. 0.98 kb in both wild type and fri mutant, while digestion of heteroduplex
shows presence of cleaved bands of 0.71 kb and 0.27 kb (adding uptothe size of full length product
0.98 kb); and * indicates nonspecific band.
Mismatch Cleavage by CEL-I Endonuclease: A Tool for Rapid Detection 7

Figure 5: Methodology for Screening of Homozygous Mutation
from a Pool of Homo/Heterozygous Population
1 Kb 0.9 Kb 0.8 Kb 0.7 Kb 0.6 Kb 0.5 Kb 0.4 Kb
0.3 Kb
0.2 Kb
M 1 2 3 4 5 6 7 8 9 1 0
Note: Lanes 1 and 3 show undigested PCR amplified DNA fragment, homozygous wild type and mutant, lanes
2 and 4 show CEL-I digested homozygous wild type and mutant DNA fragment. Lane 5 shows undigested
heteroduplex of Wild type DNA fragment mixed with mutant DNA fragment and lane 6 shows
heteroduplex made of WT + mutant DNA fragment digested with CEL-I endonuclease. The two extra
bands indicated by arrow shows the cleaved bands after digestion of CEL-I. Lanes 7 and 9 show one
putative homozygous and one heterozygous mutant plants respectively, the genomic DNA of putative
plants was PCR amplified using gene specific primer and then subjected to CEL-I endonuclease treatment,
the presence of two extra bands indicated by an arrow shows that the plant in lane 10 is heterozygous
carrying one copy of wild type and another copy of mutant DNA while lane 8 appears to be a
homozygous mutant with no other extra cleaved bands even after CEL-I treatment. The star* shows
some nonspecific cleavage observed in all the samples.
This robust technique can be even applied to screen for homozygous plants from a
population exhibiting doubtful phenotypes. Some times in tomato, wild type or any mutant
plant grown in green houses or open field shows unusual phenotypes, which can be due
to adverse climatic stress, pests, or cross pollination from neighboring plants. So it is
not advisable to use these plants for any analysis before confirming the identity of the
mutant. The only available option is to wait for few generations before we can screen
them genetically by analyzing segregation pattern or obtaining expensive gene sequence
of these plants. Here we used CEL-I endonuclease mismatch cleavage for identifying the
mutant gene identity either homozygous or heterozygous in nature. From our mutant
plant growth facility, we have identified few plants with unusual phenotypes and to
determine the homo/heterozygous nature of the plants the plant genomic DNA was isolated
and used for gene specific amplification and the PCR products were subjected to CEL-I
digestion, and the digested products were resolved on the 1% agarose gel, all the putative
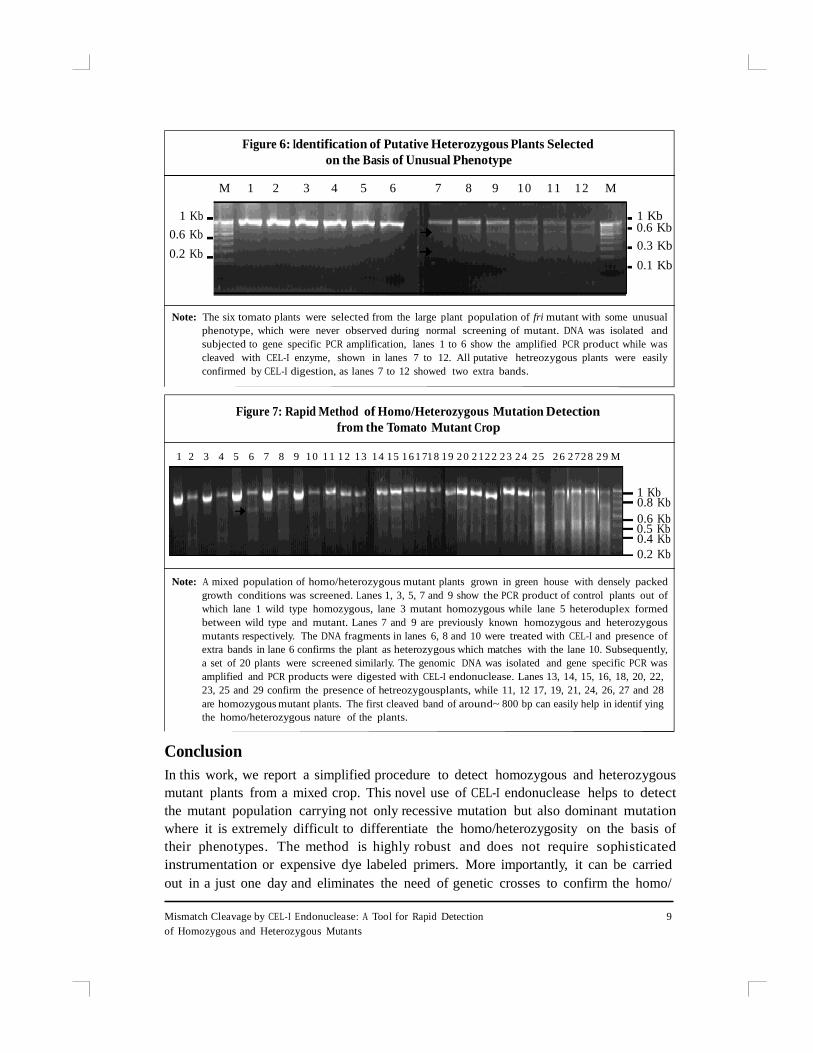
mutants turned out to be heterozygous in nature (Figure 6).
The important application of this method is large-scale screening of plants for detection
of homozygous and heterozygous mutations within a population. A set of 24 to 48 plants
can be screened in one batch. Once the genomic DNA is isolated 48 to 96 samples can be
amplified in a single PCR block in one set. A normal water bath or even PCR block can also
accommodate 48 to 96 samples for CEL-I digestion. The time taken for resolving the
cleaved fragments is important since this step cannot be done in a 96 well agarose gel
plates. In order to obtain better resolution, samples should be run at a voltage less than
50 V/min and a broad well gel casts should be used to cast agarose gel. Still 28 to 30
samples can be resolved in one batch (Figure 7). A population of about 150 to 200 plants
can easily be screened in less than three days.
8 The IUP Journal of Genetics & Evolution, Vol. IV, No. 2, 2011
of Homozygous and Heterozygous Mutants
Figure 6: Identification of Putative Heterozygous Plants Selected
on the Basis of Unusual Phenotype
M 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 M
1 Kb
0.6 Kb
0.2 Kb
1 Kb 0.6 Kb
0.3 Kb
0.1 Kb
Note: The six tomato plants were selected from the large plant population of fri mutant with some unusual
phenotype, which were never observed during normal screening of mutant. DNA was isolated and
subjected to gene specific PCR amplification, lanes 1 to 6 show the amplified PCR product while was
cleaved with CEL-I enzyme, shown in lanes 7 to 12. All putative hetreozygous plants were easily
confirmed by CEL-I digestion, as lanes 7 to 12 showed two extra bands.
Figure 7: Rapid Method of Homo/Heterozygous Mutation Detection
from the Tomato Mutant Crop
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2 1 3 1 4 1 5 1 6 1 71 8 1 9 2 0 2 1 2 2 2 3 2 4 2 5 2 6 2 7 2 8 2 9 M
1 Kb 0.8 Kb
0.6 Kb 0.5 Kb 0.4 Kb
0.2 Kb
Note: A mixed population of homo/heterozygous mutant plants grown in green house with densely packed
growth conditions was screened. Lanes 1, 3, 5, 7 and 9 show the PCR product of control plants out of
which lane 1 wild type homozygous, lane 3 mutant homozygous while lane 5 heteroduplex formed
between wild type and mutant. Lanes 7 and 9 are previously known homozygous and heterozygous
mutants respectively. The DNA fragments in lanes 6, 8 and 10 were treated with CEL-I and presence of
extra bands in lane 6 confirms the plant as heterozygous which matches with the lane 10. Subsequently,
a set of 20 plants were screened similarly. The genomic DNA was isolated and gene specific PCR was
amplified and PCR products were digested with CEL-I endonuclease. Lanes 13, 14, 15, 16, 18, 20, 22,
23, 25 and 29 confirm the presence of hetreozygousplants, while 11, 12 17, 19, 21, 24, 26, 27 and 28
are homozygous mutant plants. The first cleaved band of around~ 800 bp can easily help in identif ying
the homo/heterozygous nature of the plants.
Conclusion
In this work, we report a simplified procedure to detect homozygous and heterozygous
mutant plants from a mixed crop. This novel use of CEL-I endonuclease helps to detect
the mutant population carrying not only recessive mutation but also dominant mutation
where it is extremely difficult to differentiate the homo/heterozygosity on the basis of
their phenotypes. The method is highly robust and does not require sophisticated
instrumentation or expensive dye labeled primers. More importantly, it can be carried
out in a just one day and eliminates the need of genetic crosses to confirm the homo/
Mismatch Cleavage by CEL-I Endonuclease: A Tool for Rapid Detection 9

heterozygosity of the population This technique will enable more laboratories, particularly
in developing countries to carry out work on mutants without worrying about costs and
facilities ^
References
1. Altmann T, Go ttwald S, Ho hmann U et al. (2007), ―Weis shaar SB: GABI-TILL:
Establishment of a Central Platform for Testing Lead Gene Function in Crops Based
on TILLING‖, GABI Progress Report, pp. 54-57.
2. Caldwell D G, McCallum N, Shaw P et al. (2004), ―Structured Mutant Population for Forward
and Reverse Genetics in Barley (Hordeum vulgare L.)‖, Plant J., Vol. 40, pp. 143-150.
3. Christian Rogers, Jiangqi Wen, Rujin Chen and Giles Oldroyd (2009), ―Deletion-Based
Reverse Genetics in Medicago Truncatula‖, Plant Physiol., Vol. 151, pp. 1077-1086.
4. Colbert T, Till B J, Tompa R et al. (2001), ―High-Throughput Screening for Induced
Point Mutations‖, Plant Physiol., Vol. 126, pp. 480-484.
5. Galina I, Lazarova L, Huub J et al. (1998), ―Molecular Analysis of PHYA in Wild-Type and
Phytochrome A—Deficient Mutants of Tomato‖, The Plant Journal, Vol. 14, No. 6,
pp. 653-662.
6. Gilchrist E J and Haughn G W (2005), ―TILLING Without a Plough: A New Method with
Applications for Reverse Genetics‖, Curr. Opin. Plant Biol., Vol. 8, pp. 211-215.
7. Henikoff S and Comai L (2003), ―Single-Nucleotide Mutations for Plant Functional
Genomics‖, Annu. Rev. Plant Biol., Vol. 54, pp. 375-401.
8. Henikoff S, Till B J and Comai L (2004), ―TILLING: Traditional Mutagenesis Meets
Functional Genomics‖, Plant Physiol., Vol. 135, pp. 630-636.
9. Jack Q Wilkinson, Michael B Lanahan, David G Clark et al. (1997), ―A Dominant Mutant
Receptor from Arabidopsis Confers Ethylene Insensitivity in Heterologous Plants‖,
Nature Biotechnology., Vol. 15, pp. 444-447.
10. Jansen G, Hazendonk E, Thijssen K L and Plasterk R H A (1997), ―Reverse Genetics by
Chemical Mutagenesis in Caenorhabditis Elegans‖, Nat. Genet., Vol. 17, pp. 119-121.
11. Jose M Alonso and Joseph R Ecker (2006), ―Moving Forward in Reverse: Genetic
Technologies to Enable Genome-Wide Phenomic Screens in Arabidopsis‖, Nature Reviews
Genetics., Vol. 7, pp. 524-536.
12. Kang T J and Yang M S (2004), ―Rapid and Reliable Extraction of Genomic DNA from
Various Wild-Type and Transgenic Plants‖, BMC Biotech., Vol. 4, p. 20.
13. Lin J J and Kuo J (1998), ―A New Reagent for Simple Isolation of Plant Genomic DNA‖,
Focus, Vol. 20, No. 2, pp. 46-48.
14. Liu Y, Roof S, Ye Z et al. (2004), ―Manipulation of Light Signal Transduction as a Means
of Modifying Fruit Nutritional Quality in Tomato‖, Proc. Natl. Acad. Sci. USA, Vol. 101,
pp. 9897-9902.
1 0 The IUP Journal of Genetics & Evolution, Vol. IV, No. 2, 2011

of Homozygous and Heterozygous Mutants
15. McCallum C M, Comai L, Greene E A and Henikoff S (2000), ―Targeting Induced Local
Lesions in Genomes (TILLING) for Pl ant Functional Genomics‖, Plant Physiol.,
Vol. 123, pp. 439-442.
16. Menda N, Semel Y, Peled D et al. (2004), ―In Silico Screening of a Saturated Mutation
Library of Tomato‖, Plant J., Vol. 38, pp. 861-872.
17. Oleykowski C A, Bronson Mullins C R, Godwin A K and Yeung A T (1998), ―Mutation
Detection Using a Novel Plant Endonuclease‖, Nucl. Acid. Res., Vol. 26, pp. 4597-4602.
18. Perry J A, Wang T L, Welham T J et al. (2003), ―TILLING Reverse Genetics Tool and a
Web-Accessible Collection of Mutants of the Legume Lotus japonicus‖, Plant Physiol.,
Vol. 131, pp. 866-871.
19. Peterson D G, Boehm K S and Stack S M (1997), ―Isolation of Milligram Quantities of
Nuclear DNA from Tomato (Lycopersicon esculentum), A Plant Containing High Levels of
Polyphenolic Compounds‖, Plant Mol. Bio. Rep., Vol. 15, pp. 148-153.
20. Raghavan C, Naredo E, Wang H et al. (2007), ―Rapid Method of Detecting SNPs and its
Application in Candidate Gene Mapping‖, Mol. Breed., Vol. 19, pp. 87-101.
21. Rogers S O and Bendich A J (1988), ―Extraction of DNA from Plant Tissues‖, in
S B Gelvin and R A Schilperoort (Eds.), Plant Molecular Biology Manual A6, pp. 1-10,
Kluwer Academic Publisher, Dordrecht, Belgium.
22. Ruano G and Kidd K K (1992), ―Modeling of Heteroduplex Formation During PCR
from Mixtures of DNA Templates‖, PCR Methods Appl., Vol. 2, pp. 112-116.
23. Sessions A, Burke E, Presting G et al. (2002), ―A High-Throughput Arabidopsis Reverse
Genetics System‖, Plant Cell, Vol. 14, pp. 2985-2994.
24. Sreelakshmi Y, Gupta S, Bodanapu R et al. (2010), ―NEATTILL: A Simplified Procedure
for Nucleic Acid Extractio n from Arrayed Tissue for TILLING and Other High-
Throughput Reverse Genetic Applications‖, Plant Methods, Vol. 6, p. 3.
25. Till B J, Burtner C, Comai L and Henikoff S (2004), ―Mismatch Cleavage by Single-
Strand Specific Nucleases‖, Nucleic. Acids Res., Vol. 32, pp. 2632-2641.
26. Till B J, Reynolds S H, Weil C et al. (2004), ―Discovery of Induced Point Mutations in
Maize Genes by TILLING‖, BMC Plant Biol., Vol. 4, p. 12.
27. Wilson A K, Pickett F B, Turner J C and Estelle M (1990), ―A Dominant Mutation in
Arabidopsis Confers Resistance to Auxin, Ethylene and Abscisic Acid‖, Mol. Gen. Genet.,
Vol. 222, Nos. 2 & 3, pp. 377-383.
28. Yeung A T, Hattangadi D, Blakesley L and Nicolas E (2005), ―Enzymatic Mutation
Detection Technologies‖, Biotechniques, Vol. 38, pp. 749-758.
Reference # 66J-2011-05-06-01
Mismatch Cleavage by CEL-I Endonuclease: A Tool for Rapid Detection 1 1
