microenvironmental vegf distribution is critical for stable and
TRANSCRIPT

The FASEB Journal • FJ Express Full-Length Article
Microenvironmental VEGF distribution is critical forstable and functional vessel growth in ischemia
Georges von Degenfeld,*,‡ Andrea Banfi,*,§ Matthew L. Springer,*,� Roger A. Wagner,†
Johannes Jacobi,† Clare R. Ozawa,* Milton J. Merchant,* John P. Cooke,†
and Helen M. Blau*,1
*Baxter Laboratory in Genetic Pharmacology, Departments of Molecular Pharmacology and ofMicrobiology and Immunology; †Division of Cardiovascular Medicine, Stanford University School ofMedicine, Stanford, California, USA; ‡Cardiovascular Research, Bayer Healthcare, Wuppertal,Germany; §Institute for Surgical Research and Hospital Management, Department of Research andDepartment of Surgery, University Hospital, Basel, Switzerland; and �Division of Cardiology,University of California, San Francisco, California, USA
ABSTRACT The critical role of vascular endothelialgrowth factor (VEGF) expression levels in developmen-tal angiogenesis is well established. Nonetheless, theeffects of different local (microenvironmental) VEGFconcentrations in ischemia have not been studied in theadult organism, and VEGF delivery to patients has beendisappointing. Here, we demonstrate the existence ofboth lower and upper threshold levels of microenviron-mental VEGF concentrations for the induction of ther-apeutic vessel growth in ischemia. In the ischemic hindlimb, implantation of myoblasts transduced to expressVEGF164 at different levels per cell increased bloodflow only moderately, and vascular leakage and aber-rant preangiomatous vessels were always induced.When the same total dose was uniformly distributed byimplanting a monoclonal population derived from asingle VEGF-expressing myoblast, blood flow was fullyrestored to nonischemic levels, collateral growth wasinduced, and ischemic damage was prevented. Heman-giomas were avoided and only normal, pericyte-coveredvessels were induced persisting over 15 mo. Surpris-ingly, clones uniformly expressing either lower orhigher VEGF levels failed to provide any functionalbenefit. A biphasic effect of VEGF dose on vesselnumber and diameter was found. Blood flow was onlyimproved if vessels were increased both in size and innumber. Microenvironmental VEGF concentrations de-termine efficacy and safety in a therapeutic setting.—von Degenfeld, G., Banfi, A., Springer, M. L., Wagner,R. A., Jacobi, J., Ozawa, C. R., Merchant, M. J., Cooke,J. P., Blau, H. M. Microenvironmental VEGF distribu-tion is critical for stable and functional vessel growth inischemia. FASEB J. 20, E2277–E2287 (2006)
Key Words: angiogenesis � gene therapy � cell transplantation
Delivery of vascular endothelial growth factor-A(VEGF) in patients with ischemic heart disease orperipheral artery disease is a promising approach buthas not yet provided clear evidence of therapeutic
efficacy (1). The mixed clinical results have beencorroborated by recent experimental studies in animalmodels of coronary and peripheral ischemia that havefailed to observe sustained improvements in blood flowfollowing VEGF delivery (2–7). Increasing evidencesuggests that the lack of robust and lasting effects is dueto difficulties in achieving an appropriate dosage ofVEGF, a growth factor with an apparently narrowtherapeutic window (8, 9).
Indeed, VEGF expression requires exceedingly tightregulation during development, as both reduced ex-pression following the targeted deletion of just oneallele (10, 11) and modest overexpression (12) lead toearly embryonic lethality. In adult animals, VEGF hasbeen found to induce pathological, glomeruloid vascu-lar proliferation using diverse gene delivery vectors indifferent tissues (7, 9, 13, 14). When VEGF is constitu-tively delivered long term, vascular proliferation isunabated and results in hemangiomas (15). Moreover,VEGF-induced vascular leakage can cause severe edema(16, 17), which has been linked to limb loss (3, 18) andeven death (19) in different animal models. Appropri-ate VEGF dosage may be critical to inducing therapeu-tic improvement while avoiding deleterious effects.
We recently showed that in normal, nonischemicmuscle, the “local” (microenvironmental), concentra-tion, rather than the “total” dose, of VEGF164 deter-mined whether its delivery induced the formation ofhemangiomas or capillaries of normal appearance (9).However, the functional consequences were not inves-tigated, and it is not known whether microenvironmen-tal VEGF concentrations are critical for the achieve-ment of therapeutic effects in the clinically relevantpathology of peripheral ischemia. In this study, weinvestigated the dose-dependent mechanism of VEGF-induced functional vessel growth in ischemia by ex-
1 Correspondence: Baxter Laboratory in Genetic Pharma-cology, Stanford University School of Medicine, Stanford, CA94305-5175, USA. E-mail: [email protected]
doi: 10.1096/fj.06-6568fje
E22770892-6638/06/0020-2277 © FASEB

ploiting the unique advantages of a cell-based deliverysystem that provides uniform VEGF levels in vivo:homogeneous monoclonal populations of geneticallyengineered myoblasts derived from a single cell thatexpress equal amounts of VEGF per cell. We found thatthe parental polyclonal population of VEGF-expressingmyoblasts induced a mixture of normal capillaries andglomeruloid vascular structures that increased bloodflow only modestly. In contrast, the same total VEGFdose delivered in a microenvironmentally controlledmanner by monoclonal populations derived from asingle cell achieved full recovery of blood flow tononischemic levels, without increasing vascular leakageand while avoiding completely the appearance of aber-rant vasculature or hemangioma growth, even after 15mo of constitutive expression. Our approach revealed abiphasic dose-dependent effect of VEGF: whereas thelowest concentration already induced the maximal in-crease in new vessel number, vessel diameter enlargedcontinuously with higher VEGF doses without apparentupper limit. Vessel size and shape, rather than vesselnumber, correlated with therapeutic improvement inblood flow and collateral growth.
In conclusion, this study revealed a pathophysiologi-cal mechanism by which microenvironmental levels ofVEGF improve functional benefit long term with re-duced toxicity, findings that have implications in de-signing clinical trials of therapeutic angiogenesis.
MATERIALS AND METHODS
Myoblast culture
Primary myoblasts expressing LacZ were infected using MFG-vascular endothelial growth factor164 retrovirus, and culturedas described previously (20). Single cells isolated by flowcytometry were expanded into monoclonal populations,and VEGF secretion was monitored periodically by ELISA(R&D Systems, Minneapolis, MN, USA) (9). The four clonalpopulations secrete �10%, 100%, 180%, and 325% of theaverage VEGF levels of the parental polyclonal populationper cell in vitro (9).
Surgical induction of hind limb ischemia
Male C.B-17-SCID mice (16–20 wk old) were obtained fromthe Stanford University Department of Comparative Medi-cine and treated according to the guidelines of the StanfordAdministrative Panel on Laboratory Animal Care. Anesthesiawas induced with 0.01 mg Avertin and maintained by Metoxi-fane inhalation. Unilateral hind limb ischemia was surgicallyinduced by ligation and transection of the medial portion ofthe right superficial femoral artery distal to the deep femoralartery origin. Muscle blood velocimetry was assessed over 2min before and after surgery using a calibrated laser Doppler-probe (Perimed-PF3/Perisoft-software, Perimed, Jarfalla, Swe-den) positioned on the distal adductor muscle using athree-dimensional micromanipulator stage, as described byJacobi et al. (21). In preliminary studies, laser Dopplervelocimetry manifested greater variability than microspheremeasurements but reliably identified severe hypoperfusion.Specifically, postoperative blood flow values that were gener-ally less than 40% of the contralateral leg correlated well
when determined by laser Doppler or measured by micro-spheres in all 20 animals studied with both techniques.Accordingly, laser Doppler was used to document postopera-tive ischemia after surgery. Mice were randomized aftersurgery to receive 8 � 106 myoblasts suspended in 0.5%BSA/PBS (108cells/ml) or vehicle injected into the distalthigh muscles (adductor and quadriceps femoris groups) (8injections/106 cells each).
Microsphere measurement of blood flow andmicroangiography
Fourteen days after surgery, blood flow was measured usingfluorescent microspheres as described previously (21).After median thoracotomy, 2 � 105 15-�m diameter red-fluorescent microspheres (Molecular Probes, Eugene, OR,USA) were continuously injected over 60 s into the beatingleft ventricle. Perfusion fixation was performed in fourmice/group: the heart was cannulated and perfused at 110mmHg with Tris-HCl buffer containing Na�, Ca2�, Mg2�,and 0.1% adenosine (2 min), followed by 1.5% formalde-hyde (2 min) (22). The muscle group of the thigh wasexcised, cut in midthigh, weighed, embedded in OCT-compound, and snap frozen. Kidneys were reference or-gans for equilateral microsphere distribution. Micro-spheres were individually counted by direct fluorescencemicroscopy on 100-�m cryosections from the entire sam-ples, a method that avoids difficulties associated withfluorescence recovery and background fluorescence intissue homogenates (23, 24). An additional advantage isthat neighboring thin cryosections can be obtained forhistology and immunofluorescence analysis matching mi-crosphere counts from the same muscle (21). Microspherecounts, normalized for muscle weight, from the right,ischemic leg were normalized to the contralateral, nonisch-emic leg. Microangiography was performed in some ani-mals as described (21). Following cannulation of theabdominal aorta, barium-sulfate solution (0.5 mg/ml) wasinjected and images acquired using a Faxitron radiographysystem (Faxitron, Wheeling, IL, USA). To determine cap-illary perfusion in vivo, fluorescein-isothiocyanate-labeledtomato lectin (Vector Laboratories, Burlingame, CA, USA)was injected intravenously (i.v.) and allowed to circulatefor 2 h before perfusion fixation (n�2/group) using amodified published protocol (25).
Vascular leakage
Vascular leakage was assessed 4 days after surgery and cellimplantation using a modified described protocol (n�3/group) (9). Evans Blue (J. T. Baker, Phillipsburg, NJ, USA)was injected i.v. (30 �g/g body wt). Four hours later, micewere perfused (1% paraformaldehyde/0.05 M citric acid, pH3.5), muscle samples from the distal thigh were harvested,Evans Blue was extracted in formamide at 55°C overnight,quantified with a spectrophotometer at 610 nm, and normal-ized to tissue wet wt.
Immunofluorescence and histomorphometry
For immunofluorescent staining, 10-�m sections werefixed in 1.6% formaldehyde, and blocked using 2% normalgoat serum, 0.5% casein, and 0.3% Triton-X-100 (1 h).Slides were incubated (1 h) with rat monoclonal antibody(mAb) against CD31/PECAM-1 (PharMingen, San Diego,CA, USA; 1:100 dilution), mouse mAb against �-SMA(ICN-Biomedicals, Aurora, OH, USA, 1:400), rabbit anti-body (Ab) against laminin (Chemicon, Temecula, CA,
E2278 Vol. 20 December 2006 VON DEGENFELD ET AL.The FASEB Journal

USA, 1:200) or rabbit Ab against �-galactosidase (Eppen-dorff-5-Prime, Westbury, NY, USA, 1:200). Secondary anti-bodies conjugated with Alexa-fluorophores (MolecularProbes) and Hoechst-33258 nuclear stain were used. Adja-cent sections were stained with H&E or X-gal. Slides wereanalyzed using a Zeiss Axioplan microscope.
Vessel growth was differentially quantified for length andsize. For vessel length density, three images of cryosections ofthe quadriceps femoris and adductor muscles stained forPECAM were randomly acquired (20� objective). Center-lines of vessels were manually drawn using a modified tech-nique, as previously described (9), quantified using calibratedOpenlab™ image analysis software (Improvision, Lexington,MA), and normalized to the number of muscle fibers. Vesselsizes were measured on 3 randomly chosen images of thequadriceps femoris muscle (20� objective) with a standard-ized grid overlay, using a modified protocol described previ-ously (9, 18, 26). The diameter of each vessel (if any) in thecenter of randomly selected squares was measured usingcalibrated software. Muscle regions expressing LacZ followingfusion with the injected myoblasts were compared with re-mote LacZ-negative regions. Collateral vessels �30 �m indiameter were identified on cross sections of the proximaladductor muscle by costaining for PECAM and �-SMA andquantified as described previously (21, 27).
Damaged muscle was defined as either inflammation(mononucleated cell infiltrates) or necrosis (“ghost fibers”lacking nuclei) (28). Areas were manually drawn on digitalimages (5� objective) and quantified using calibrated soft-ware (3 legs/group).
Implantation of myoblasts into the ear muscle andwhole-mount preparation
5 � 105 myoblasts in 5 �l 0.5% BSA/PBS were implanted intothe posterior auricular muscle (9). In vivo intravascularstaining was performed 4 wk after cell implantation by i.v.injection of biotinylated tomato lectin (Vector Laboratories),as described previously (9, 19). Ears were removed, bisectedin the cartilage plane, and stained with X-gal and avidin-biotin complex (ABC)/diaminobenzidine histochemistry(Vector Laboratories), dehydrated through an alcohol series,and cleared with toluene. Vessel diameters and length densi-ties around LacZ-positive muscle fibers were measured onimages of ear whole-mounts. The centerline of blood vessels(brown lectin stain) and LacZ-expressing muscle fibers weremanually traced and quantified on digital photomicrographs(20� objective) overlaid with a computer-generated squarelattice using calibrated software for vessel length density(vessel length/muscle fiber length) and branching (branch-ing points/100 �m muscle fiber length) on 3 fields/ear (3ears/group). The diameter of each vessel in the center ofrandomly selected squares was measured (30 measurements/ear; 3 ears/group).
Statistics
Data are presented as mean � sems. Differences betweengroups with respect to vessel length, diameter, blood flow,collateral score, VEGF quantity, and leakage were assessedusing ANOVA and Student’s t test; P 0.05 was consideredstatistically significant and P 0.01 highly significant [Statis-tical Packages for the Social Sciences-13.0, statistical Packagesfor the Social Sciences (SPSS) Inc., Chicago, IL, USA].
RESULTS
Implantation into the ischemic hind limb of clonalmyoblast populations uniformly expressing differentlevels of VEGF
To study the microenvironmental dose effects ofVEGF164 in ischemia, we generated monoclonal popu-lations of myoblasts that express equal amounts ofVEGF per cell. Primary murine myoblasts expressingLacZ were infected with MFG-vascular endothelialgrowth factor164 retrovirus. From the resulting poly-clonal population of VEGF-expressing myoblasts, singlecells were isolated by flow cytometry and expanded intomonoclonal populations as described (9). Comparedwith the parental polyclonal myoblast population pro-ducing on average 61 � 5 ng/106 cells/24 h VEGF invitro, each of the four derived clonal populations se-creted different VEGF amounts per cell: “10% clone”:6 � 1, “100% clone”: 55 � 5, “180% clone”: 104 � 8,and “325% clone”: 191 � 15.
To induce hind limb ischemia, the right superficialfemoral artery was surgically ligated and transected.Laser-Doppler velocimetry performed after surgeryshowed marked blood flow reduction below 40% ofpreoperative levels in each individual mouse withoutdifferences between treatment groups (not shown).Fourteen days later, regional blood flow was deter-mined by injection of fluorescent microspheres, awell-characterized method considered as the goldstandard, as it measures the effective perfusion ofthe microvasculature that correlates well with meta-bolic and functional measures of ischemia (16, 21,29 –31).
The area of myoblast engraftment (X-Gal staining)was distributed along the needle tracks and com-prised �20% of the cross-sectional muscle area (ex-ample shown in Fig. 6G). No difference was seen withregard to the extent and distribution of cell engraft-ment or fusion with endogenous muscle fibers alongthe needle tracks between the parental polyclonalpopulation and any of the monoclonal populations14 days after implantation. Furthermore, no tumori-genicity was seen with any myoblast population,including the clones. Finally, we quantified theamount of VEGF expressed following implantation ofthe parental polyclonal population and the 100%clone into the ischemic muscle in order to rule outVEGF-mediated toxicities to the myoblasts expressing“high” amounts of VEGF in vivo. As predicted fromthe in vitro expression levels, VEGF expression fol-lowing implantation of the 100% clone into themuscle was similar to that of the parental polyclonalpopulation (see below). In conclusion, there was noevidence for differences in cell survival, distributionor VEGF production between the monoclonal popu-lations, and compared with the parental polyclonalpopulation from which they had been derived.
E2279MICROENVIRONMENTAL VEGF DETERMINES FUNCTION

Control over microenvironmental distributionimproves the efficacy of VEGF delivery in ischemiawithout increasing leakage
To determine the effects of VEGF dosage, mice wererandomized after surgery to be injected either withvehicle only, LacZ-expressing control myoblasts, theparental polyclonal VEGF-expressing myoblast popu-lation, or one of the monoclonal VEGF-expressingpopulations (6 groups, n�9/group). Fourteen daysafter surgery and cell implantation, the controlgroup injected with vehicle showed markedly re-duced perfusion in the ischemic leg compared withthe contralateral leg (54.6�6.8%), showing that thesurgical procedure provided sustained blood flowreduction (Fig. 1). Injection of control myoblasts didnot improve blood flow (55.3�13.1%). Mice im-planted with the parental polyclonal VEGF popula-tion exhibited only moderately increased blood flow(74.1�8.6%, P0.05 vs. controls). Improvement wassubstantially greater in mice that received the 100%vascular endothelial growth factor clone, in whichblood flow was similar to the contralateral, nonisch-emic leg (96.8�10.5%, P0.01 vs. controls; P0.05vs. polyclonal VEGF population). By contrast, blood
flow was neither improved in mice having receivedlow (10% clone: 54.7�17.5%) nor high microenvi-ronmental VEGF doses (180% clone: 69.8�18.9%),although the latter showed a positive trend.
Differences in blood flow were accompanied byvarious degrees of ischemic muscle damage, quantifiedon H&E-stained muscle sections, according to previ-ously described criteria (28). Muscle damage wassmaller in mice treated with the 100% vascular endo-thelial growth factor clone compared with all othergroups (P0.05; Table 1). Furthermore, the incidenceof foot gangrene appeared lower in mice injected withthe 100% VEGF clone (Table 1). Thus, although the100% clone expresses the same total VEGF dose as theparental polyclonal population, they have much greaterpositive effects on perfusion and salvage of ischemictissue.
We compared the amounts of VEGF expressed bythe 100% clone and the parental polyclonal popula-tion in vivo. Both populations express similaramounts of VEGF in vitro. The aim of the in vivoquantification was to rule out increased cell survivalor proliferation of the 100% clone, which might haveoccurred as a result of cell transformation in theprocess of clonal expansion from a single cell. Aver-age VEGF levels were measured in muscles harvested3 days after ischemia induction and cell injection(ELISA). Endogenous VEGF was up-regulated inischemia (control myoblasts: 25�2 ng/mg protein;vehicle: 26�2) compared with nonischemic muscle(7�1) (Fig. 2A). The VEGF myoblasts strongly in-creased total VEGF content, but levels were notdifferent for the parental polyclonal population andthe 100% clone (polyclonal: 129�22 ng/mg protein;100% clone: 128�47). Hence, although microenvi-ronmental distribution of VEGF concentrations inmuscle cannot be directly measured, the finding thatboth VEGF populations expressed similar totalamounts of VEGF implies that the greater antiisch-emic effects provided by the 100% clone was due toa more favorable, uniform local distribution and notto increase survival or proliferation of the clonallyexpanded population.
We determined whether functional improvementsinduced by the 100% clone came at the expense ofadverse effects. Vascular leakage is a hallmark ofVEGF effects that leads to potentially harmful tissueedema (3, 19). Leakage peaks 4 days after intramus-cular (i.m.) implantation of polyclonal VEGF myo-blasts before decreasing to baseline (9) (D. M. Mc-Donald, UCSF, personal communication). Leakagewas quantified 4 days after surgery and myoblastinjection by i.v. injection of Evans Blue. Both VEGFpopulations induced similar amounts of leakage inischemic muscle (Fig. 2B) that were higher than thatseen in the ischemic control legs, resulting fromendogenous VEGF up-regulation. Hence, increasedtherapeutic effects provided by the 100% VEGF clone
Figure 1. Microenvironmental concentrations of vascularendothelial growth factor (VEGF) determine blood flowimprovement in ischemia. Regional blood flow was mea-sured in the ischemic leg 14 days after surgical induction ofhind limb ischemia using microspheres (n�9/group).Blood flow in ischemic legs (expressed as percentage of thenonischemic contralateral leg) was severely reduced inmice injected with vehicle or control myoblasts. Implanta-tion of the parental polyclonal population of VEGF-ex-pressing myoblasts improved blood flow only modestly(*P0.05 vs. controls). Diverse effects were induced by theclonal populations uniformly expressing distinct levels ofVEGF “per cell”. In particular, the 100% clone markedlyimproved blood flow in the ischemic leg to a level compa-rable with the contralateral nonischemic leg (**P0.01 vs.controls; P0.05 vs. polyclonal VEGF myoblasts). In con-trast, neither the 10% nor the 180% clone improved bloodflow significantly, although the latter showed a positivetrend
E2280 Vol. 20 December 2006 VON DEGENFELD ET AL.The FASEB Journal

compared with the parental polyclonal populationdid not come at the expense of increased leakage.
Vessel number increase peaks at the lowest VEGFdose
To determine the structural features underlying thedistinct functional effects elicited by different microen-
vironmental VEGF doses, we examined the morphol-ogy, length, and diameter of vessel growth 14 days afterischemia induction and cell injection. Cryosectionsfrom ischemic muscle were stained for endothelialcells (platelet endothelial cell adhesion molecule-1PECAM/CD31). Vessel growth was seen exclusivelytightly localized around the muscle fibers that ex-pressed VEGF. Vessel length density was assessed bydrawing a mask in randomly selected fields and nor-malized to muscle fiber counts to avoid distortionscaused by ischemia-induced muscle shrinkage. Vessellength density was increased in the ischemic muscles ofall groups compared with the nonischemic contralat-eral leg, reflecting endogenous angiogenesis. Implan-tation of control myoblasts had no further effect (Fig.3A, G). In contrast, vessel length density was increasedin all groups injected with VEGF-myoblasts (Fig. 3B–G).Surprisingly, the effect was not dose-dependent: vessellength density peaked already in ischemic musclesimplanted with the 10% clone and was not furtherincreased by even the highest VEGF levels.
Although vessel density can be reliably quantified ontwo-dimensional cryosections, it is impossible to distin-guish between longitudinal growth or tortuousity ofpreexisting vessels and truly increased number of ves-sels resulting from sprouting or intussusception. Theclones were therefore studied in the ear muscle, whichis amenable to three-dimensional vessel analysis. Sam-ples were analyzed 4 wk after cell implantation becausemyoblast fusion and angiogenesis are delayed in the earcompared with leg muscles (9). In vivo blood vesselstaining was performed by i.v. injection of biotinylatedlectin (9, 19). Following ABC/diaminobenzidine stain-ing, branching patterns and segment length betweenbranches were analyzed in detail by quantifying amanually drawn mask overlay on whole-mount imagesof the vasculature around LacZ-expressing muscle fi-bers. The 10% clone induced a peak increase in thenumber of branching points and vessel length (Fig. 4B,E, F) and no further increase was seen with the 100%and 180% clones (Fig. 4C–F). VEGF-induced vesselswere tortuous, indicating longitudinal growth, but thiseffect was balanced by an equivalent increase in branchpoints. Thus, the calculated average length of vesselsegments between branch points was unchanged be-tween the controls and all three VEGF clones. There-fore, the data from the ear muscle whole-mountsconfirmed the findings from the ischemic leg musclesand showed that the VEGF clones truly increased vesselnumber.
TABLE 1. Blood flow improvement made with VEGF
Controls No VEGF VEGF-Poly 10% Clone 100% Clone 180% Clone
Gangrene foot/n 3/11 3/11 2/9 2/11 1/12 3/11Muscle damage %� 15.5 � 4.8 17.5 � 2.5 12.4 � 4.8 12.0 � 5.8 3.5 � 3.1* 12.5 � 4.4
In accordance with blood flow improvement, legs implanted with the 100%-vascular endothelial growth factor (VEGF) clone showedreduced muscle damage and a lower incidence of foot gangrene compared with all other groups, indicating functional benefit of optimizedmicroenvironmental VEGF levels (*P0.05).
To
tal V
EG
F e
xpre
ssio
nin
mu
scle
[pg
/mg
pro
tein
]
Vas
cula
r le
akag
e[E
van
’s b
lue
extr
avas
atio
nn
g/m
g m
usc
le w
et w
eig
ht]
A
B
Control m
yobla
sts
Polycl
onal
VEGF myo
blast
s
100%
VEGF c
lone
Vehic
le
No isch
emia
0102030405060708090100
# #
* *
020406080100120140160180
# #
* *
Figure 2. Despite improved blood flow by uniform VEGFdelivery, vascular leakage is not increased. A) “Total” VEGFprotein concentration in ischemic muscle was increased 3days after ischemia induction, reflecting endogenous up-regulation (ELISA). VEGF levels were increased after implan-tation of VEGF-expressing myoblasts; however, there was nodifference between the parental polyclonal population andthe 100% clone confirming in vitro levels. B) Vascular leakagemeasured by Evans Blue extravasation was increased 4 daysafter ischemia induction. Implantation of VEGF-expressingmyoblasts further increased leakage, but no difference wasfound between the parental polyclonal population and the100% clone. #P 0.01 vs. nonischemic muscle; *P 0.01 vs.ischemic control groups (n�3/group).
E2281MICROENVIRONMENTAL VEGF DETERMINES FUNCTION
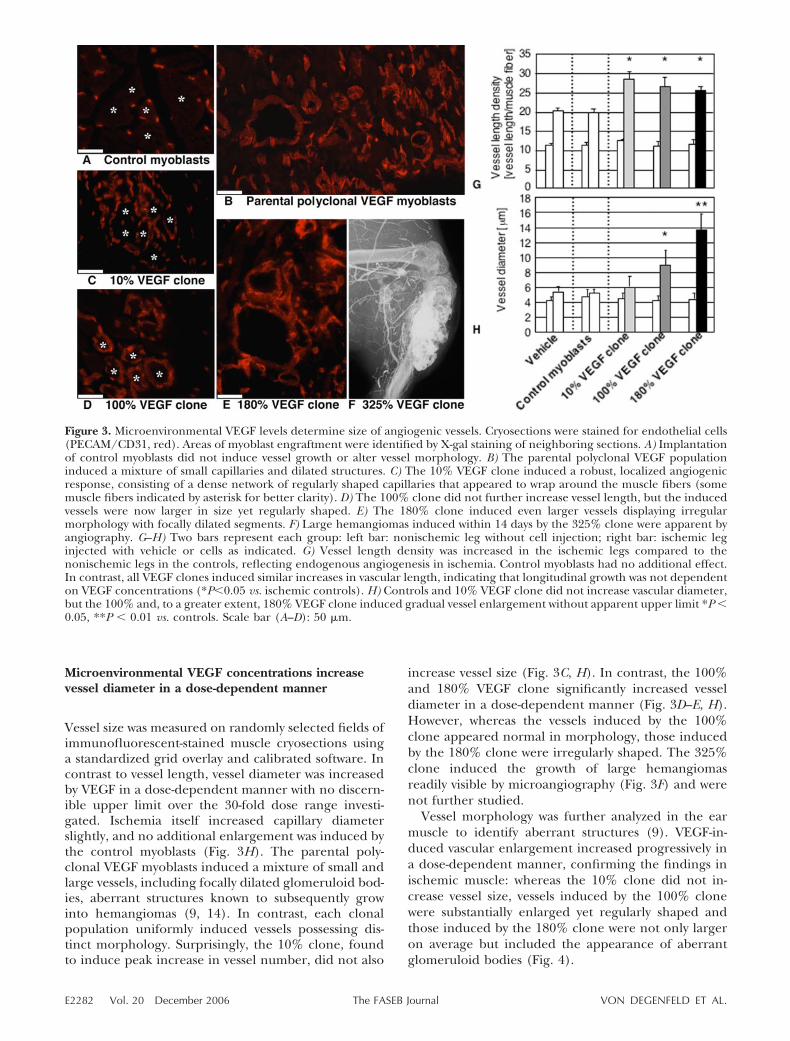
Microenvironmental VEGF concentrations increasevessel diameter in a dose-dependent manner
Vessel size was measured on randomly selected fields ofimmunofluorescent-stained muscle cryosections usinga standardized grid overlay and calibrated software. Incontrast to vessel length, vessel diameter was increasedby VEGF in a dose-dependent manner with no discern-ible upper limit over the 30-fold dose range investi-gated. Ischemia itself increased capillary diameterslightly, and no additional enlargement was induced bythe control myoblasts (Fig. 3H). The parental poly-clonal VEGF myoblasts induced a mixture of small andlarge vessels, including focally dilated glomeruloid bod-ies, aberrant structures known to subsequently growinto hemangiomas (9, 14). In contrast, each clonalpopulation uniformly induced vessels possessing dis-tinct morphology. Surprisingly, the 10% clone, foundto induce peak increase in vessel number, did not also
increase vessel size (Fig. 3C, H). In contrast, the 100%and 180% VEGF clone significantly increased vesseldiameter in a dose-dependent manner (Fig. 3D–E, H).However, whereas the vessels induced by the 100%clone appeared normal in morphology, those inducedby the 180% clone were irregularly shaped. The 325%clone induced the growth of large hemangiomasreadily visible by microangiography (Fig. 3F) and werenot further studied.
Vessel morphology was further analyzed in the earmuscle to identify aberrant structures (9). VEGF-in-duced vascular enlargement increased progressively ina dose-dependent manner, confirming the findings inischemic muscle: whereas the 10% clone did not in-crease vessel size, vessels induced by the 100% clonewere substantially enlarged yet regularly shaped andthose induced by the 180% clone were not only largeron average but included the appearance of aberrantglomeruloid bodies (Fig. 4).
Figure 3. Microenvironmental VEGF levels determine size of angiogenic vessels. Cryosections were stained for endothelial cells(PECAM/CD31, red). Areas of myoblast engraftment were identified by X-gal staining of neighboring sections. A) Implantationof control myoblasts did not induce vessel growth or alter vessel morphology. B) The parental polyclonal VEGF populationinduced a mixture of small capillaries and dilated structures. C) The 10% VEGF clone induced a robust, localized angiogenicresponse, consisting of a dense network of regularly shaped capillaries that appeared to wrap around the muscle fibers (somemuscle fibers indicated by asterisk for better clarity). D) The 100% clone did not further increase vessel length, but the inducedvessels were now larger in size yet regularly shaped. E) The 180% clone induced even larger vessels displaying irregularmorphology with focally dilated segments. F) Large hemangiomas induced within 14 days by the 325% clone were apparent byangiography. G–H) Two bars represent each group: left bar: nonischemic leg without cell injection; right bar: ischemic leginjected with vehicle or cells as indicated. G) Vessel length density was increased in the ischemic legs compared to thenonischemic legs in the controls, reflecting endogenous angiogenesis in ischemia. Control myoblasts had no additional effect.In contrast, all VEGF clones induced similar increases in vascular length, indicating that longitudinal growth was not dependenton VEGF concentrations (*P0.05 vs. ischemic controls). H) Controls and 10% VEGF clone did not increase vascular diameter,but the 100% and, to a greater extent, 180% VEGF clone induced gradual vessel enlargement without apparent upper limit *P 0.05, **P 0.01 vs. controls. Scale bar (A–D): 50 �m.
E2282 Vol. 20 December 2006 VON DEGENFELD ET AL.The FASEB Journal

Optimized uniform VEGF delivery stimulatescollateral growth
Arteriogenesis, the growth of collaterals from preexist-ing arteries, is required for blood flow increase inischemia (1, 32). Collaterals were quantified on cryo-sections of the adductor muscles proximal of the siteof cell injection by costaining for endothelial cells(PECAM/CD31) and vascular smooth muscle cells (� -SMA). The number of collaterals � 30 �m in diameterwas increased in mice injected with 100% VEGF clonecompared with the controls, and a trend toward en-largement was seen with the 180% clone and theparental polyclonal population (Fig. 5A). Quantitativeanalyses were corroborated by microangiograms thatshowed more numerous arteries in the thigh area oflegs implanted with the 100% clone (Fig. 5B). Thus, theincrease in blood flow induced by the 100% clone wasaccompanied by proximal collateral growth.
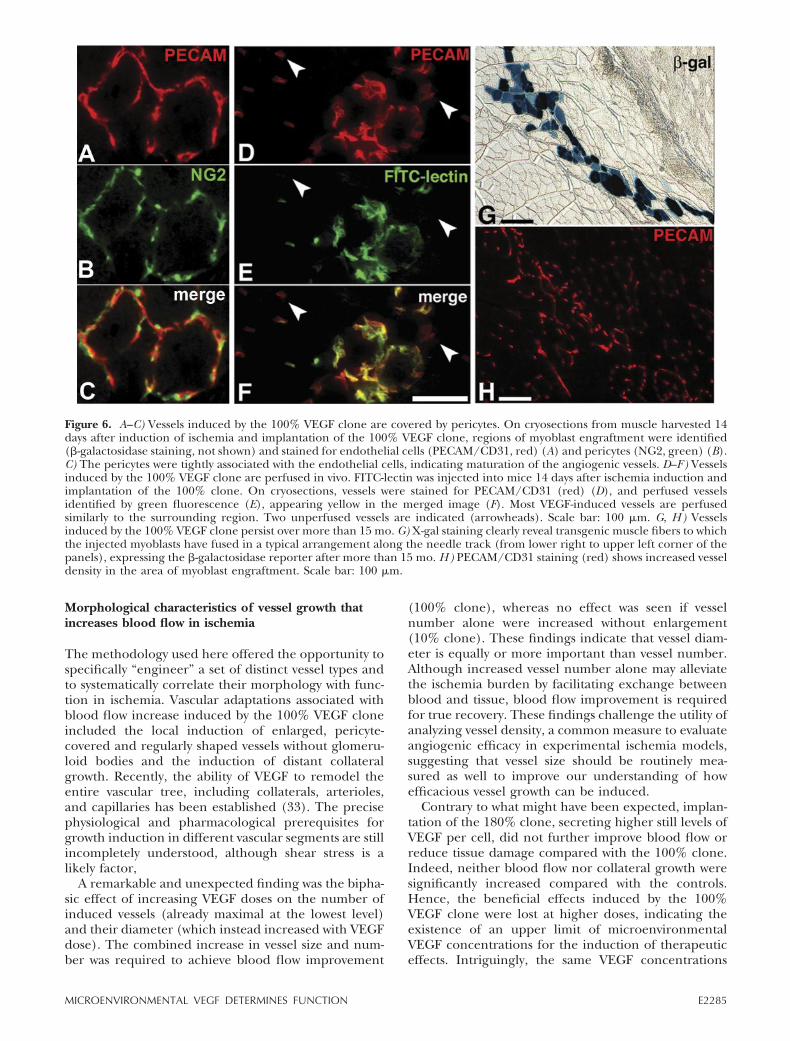
Vessels induced by optimized VEGF delivery areperfused in vivo and persist more than 15 mo
To confirm that vessels induced by the 100% VEGFclone were perfused in vivo, fluorescent lectin wasinjected i.v. Muscle sections costained for PECAMshowed that most vessels induced by the 100% clonewere perfused similarly to the surrounding vessels (Fig.6D–F). In accordance with blood flow improvement(microspheres), these findings demonstrate that the
vessels induced by the 100% clone were able to recruitblood flow.
Vessels induced by transient VEGF expression lackpericytes, are unstable, and are prone to regression (13,14). On the other hand, sustained VEGF expressioncan induce hemangioma growth, posing a challenge forsafe and therapeutic VEGF delivery. To address thisissue, we harvested muscles 66 wk after ischemia induc-tion and implantation of the 100% clone. Sustained�-galactosidase expression over more than 15 mo wasfound in muscle fibers to which the injected myoblastshad fused. In the area of cell engraftment, increasedvessel density was still readily visible (Fig. 6G–H). Re-markably, no evidence of glomeruloid bodies or hem-angioma growth was found despite long-term survival ofcells constitutively expressing VEGF. We examinedwhether long-term persistence of vessels induced by the100% clone correlated with pericyte coverage bycostaining muscle sections obtained 14 days after isch-emia induction and cell implantation for endothelialcells (PECAM/CD31) and pericytes (NG2). Vesselsinduced by the 100% clone displayed tightly associatedpericyte coverage (Fig. 6A–C), indicating that long-term stability was a result of adequate maturation of thenewly induced vessels. In contrast, the vessels inducedby the parental polyclonal VEGF population showedabnormal coverage with pericytes that appeared de-tached from the growing vessels and expressed smooth-muscle-actin (not shown), ultimately leading to thegrowth of glomeruloid bodies and hemangiomas. The
Figure 4. Vessel size, but not number, is dependent on microenvironmentalVEGF concentration. A–D) Whole-mount preparation of the ear 28 days aftermyoblast injection revealed clearly distinct morphologies of induced vessels.Perfused vascular lumen: brown lectin staining; engrafted myoblasts: bluishgreen X-gal staining. Scale bar: 50 �m. E–G) Morphometry shows that vesselnumber and branching were maximally increased by the 10% clone, withoutfurther increase with ascending VEGF doses. In contrast, vessels weregradually enlarged with increasing microenvironmental VEGF concentra-tions: unlike the 10% clone, the 100% clone induced the enlargement ofvessels that were, however, still normal in appearance; in contrast, the 180%clone induced vessels that were substantially enlarged and glomeruloidbodies. *P 0.05 vs. controls.
E2283MICROENVIRONMENTAL VEGF DETERMINES FUNCTION

present findings show that prolonged, but uniformVEGF expression by the 100% clone in ischemic muscleyields mature, pericyte-covered vessels that persist formore than 15 mo, approximately half of the life span ofa mouse, without regressing or becoming aberrant.
DISCUSSION
Uniform VEGF distribution within amicroenvironmental dose window determines bloodflow improvement in ischemia
The most important and clinically relevant finding ofthis study is that that controlling the microenvironmen-tal concentration of VEGF, without changing totaldose, defines a dose window in which constitutive,long-term VEGF gene expression in ischemic musclepredictably induces stable and functional vessel growth,efficiently rescues blood flow to nonischemic levels,
while avoiding any aberrant angiogenesis and heman-gioma growth. These findings show clearly in a thera-peutic setting that, because of the pathophysiologicalmechanism of VEGF-induced vessel growth, averagetotal doses across muscle tissue are not the criticaldeterminant of efficacy and safety and that local con-centrations determine functional outcome. This studyis, to our best knowledge, the first to show that optimiz-ing the microenvironmental distribution is critical toimproving both efficacy and safety of VEGF in ischemia,without reducing or increasing total dose.
Because no available technique allows the directquantification of microenvironmental VEGF concentra-tions in tissue, we used a cell-based method to examinethe dose-dependent effects in ischemia (9). In thepolyclonal population, the amount of VEGF expressedper cell is variable, because the numbers of retroviralDNA copies and their integration sites into the hostgenome is unpredictable. By contrast, a monoclonalpopulation derived from a single cell expresses a givenamount of VEGF per cell. Remarkably, we find in theischemic hind limb that the uniformity of VEGF secre-tion critically influences both vascular structure andfunction. Blood flow was improved by the 100% VEGFclone compared with the parental polyclonal popula-tion, reaching a level similar to the nonischemic leg,and ischemic tissue damage was reduced. In contrast,neither low (10%) nor high (180%) microenvironmen-tal VEGF concentrations improved function, demon-strating the previously unrecognized existence of alower and an upper threshold VEGF concentration forthe induction of beneficial effects in ischemia.
Enhanced efficacy of the 100% clone compared withthe parental polyclonal population was not due to ahigher total VEGF dose, which is similar between bothpopulations in vitro and in vivo, showing that microen-vironmental factors are critical. Importantly, the vesselsinduced by the 100% clone were pericyte-covered andpersisted more than 15 mo, to our best knowledge, thelongest reported persistence of vessels induced byexogenous VEGF delivery, and possibly indefinitely, asa result of constitutive VEGF expression. On the otherhand, implantation of the 100% clone did not inducehemangioma formation, a form of aberrant vesselgrowth invariably induced by the polyclonal VEGFpopulation, regardless of how few cells were injected(9). The successful avoidance of hemangiomas is re-markable in that 8 million cells were injected in thepresent study, whereas as few as 35,000 cells of theparental polyclonal population had previously beenshown to induce aberrant vessel growth (9). Hence, ifmicroenvironmental VEGF concentrations are kept be-low a threshold level, the total VEGF dose in ischemiacan be dramatically increased without reintroducingthe risk of hemangioma growth. These findings showthat constitutive expression of VEGF has, indeed, thepotential to induce long-term stable vessels withoutgiving rise to aberrancies provided that microenviron-mental VEGF levels are controlled within a therapeuticdose window.
Figure 5. The 100%-vascular endothelial growth factor cloneinduces collateral growth. A) Collateral growth, determinedby costaining for PECAM/CD31 (endothelial cells) and�-SMA (vascular smooth muscle cells) of sections of theproximal adductor muscle, was significantly increased only bythe 100% VEGF clone, and showed a trend toward increase inthe parental polyclonal population and the 180% clone. B, C)Microangiograms corroborated the quantitative collateralscore, showing increased growth of collaterals originatingfrom large feeder vessels, crossing the thigh and connectingto the vasculature of the lower leg in mice implanted with the100% clone by comparison with control myoblasts. Numerouscollaterals display the typical, corkscrew-like morphology ofgrowing collaterals.
E2284 Vol. 20 December 2006 VON DEGENFELD ET AL.The FASEB Journal
Morphological characteristics of vessel growth thatincreases blood flow in ischemia
The methodology used here offered the opportunity tospecifically “engineer” a set of distinct vessel types andto systematically correlate their morphology with func-tion in ischemia. Vascular adaptations associated withblood flow increase induced by the 100% VEGF cloneincluded the local induction of enlarged, pericyte-covered and regularly shaped vessels without glomeru-loid bodies and the induction of distant collateralgrowth. Recently, the ability of VEGF to remodel theentire vascular tree, including collaterals, arterioles,and capillaries has been established (33). The precisephysiological and pharmacological prerequisites forgrowth induction in different vascular segments are stillincompletely understood, although shear stress is alikely factor,
A remarkable and unexpected finding was the bipha-sic effect of increasing VEGF doses on the number ofinduced vessels (already maximal at the lowest level)and their diameter (which instead increased with VEGFdose). The combined increase in vessel size and num-ber was required to achieve blood flow improvement
(100% clone), whereas no effect was seen if vesselnumber alone were increased without enlargement(10% clone). These findings indicate that vessel diam-eter is equally or more important than vessel number.Although increased vessel number alone may alleviatethe ischemia burden by facilitating exchange betweenblood and tissue, blood flow improvement is requiredfor true recovery. These findings challenge the utility ofanalyzing vessel density, a common measure to evaluateangiogenic efficacy in experimental ischemia models,suggesting that vessel size should be routinely mea-sured as well to improve our understanding of howefficacious vessel growth can be induced.
Contrary to what might have been expected, implan-tation of the 180% clone, secreting higher still levels ofVEGF per cell, did not further improve blood flow orreduce tissue damage compared with the 100% clone.Indeed, neither blood flow nor collateral growth weresignificantly increased compared with the controls.Hence, the beneficial effects induced by the 100%VEGF clone were lost at higher doses, indicating theexistence of an upper limit of microenvironmentalVEGF concentrations for the induction of therapeuticeffects. Intriguingly, the same VEGF concentrations
Figure 6. A–C) Vessels induced by the 100% VEGF clone are covered by pericytes. On cryosections from muscle harvested 14days after induction of ischemia and implantation of the 100% VEGF clone, regions of myoblast engraftment were identified(�-galactosidase staining, not shown) and stained for endothelial cells (PECAM/CD31, red) (A) and pericytes (NG2, green) (B).C) The pericytes were tightly associated with the endothelial cells, indicating maturation of the angiogenic vessels. D–F) Vesselsinduced by the 100% VEGF clone are perfused in vivo. FITC-lectin was injected into mice 14 days after ischemia induction andimplantation of the 100% clone. On cryosections, vessels were stained for PECAM/CD31 (red) (D), and perfused vesselsidentified by green fluorescence (E), appearing yellow in the merged image (F). Most VEGF-induced vessels are perfusedsimilarly to the surrounding region. Two unperfused vessels are indicated (arrowheads). Scale bar: 100 �m. G, H) Vesselsinduced by the 100% VEGF clone persist over more than 15 mo. G) X-gal staining clearly reveal transgenic muscle fibers to whichthe injected myoblasts have fused in a typical arrangement along the needle track (from lower right to upper left corner of thepanels), expressing the �-galactosidase reporter after more than 15 mo. H) PECAM/CD31 staining (red) shows increased vesseldensity in the area of myoblast engraftment. Scale bar: 100 �m.
E2285MICROENVIRONMENTAL VEGF DETERMINES FUNCTION

marked the transition from the induction of morpho-logically normal vasculature (100% clone) to that ofaberrant, glomeruloid bodies with insufficient and ab-normal pericyte coverage (180% clone) that later growinto hemangiomas. In a previous report, VEGF deliveryin a manner that induced hemangioma-like structuresalso failed to improve blood flow in ischemic myocar-dium (7). It is unclear whether the association betweenaberrant vessels and the loss of functional improvementin ischemia is a coincidence, or indicates a causalrelationship. However, striking parallels exist of theconcept of transient normalization of tumor vascula-ture that occurs after treatment with VEGF inhibitors(34). Tumor vasculature is frequently abnormally di-lated due to excessive proangiogenic stimulation. Ther-apeutic VEGF inhibition prunes such abnormalities,transiently resulting in a vasculature that is smaller indiameter and closer to normal, resulting in increasedtumor blood supply and oxygenation in a seeminglyparadoxical manner (34). The present study indicatesthat similar rules apply to angiogenesis in ischemia andthat excessive proangiogenic stimulation induces di-lated and glomeruloid vasculature (180% clone) thatabolishes the functional benefit seen at lower VEGFdoses. This imbalance is completely avoided if themicroenvironmental VEGF concentration is uniformlymaintained below a threshold level (100% clone). It iscurrently not fully understood, which biological mech-anisms underlie the “decision” of a vessel exposed toangiogenic stimuli to grow in length, form a newbranch or enlarge, and how these factors can be used toimprove therapeutic efficacy and safety. Factors down-stream of VEGF might act to stabilize (e.g., Angiopoi-etin-1) or further destabilize the angiogenic vessels(e.g., proteases). Spatial gradients of VEGF have beenproposed to induce angiogenic sprouting by guidingthe filopodia-mediated migration of endothelial tipcells (35). Furthermore, pericytes, which reside onmicrovessels and contact several endothelial cells withtheir long processes, could modulate control vesselsprouting and branching (36, 37).
The results of this study help explain the apparentlack of a manageable therapeutic window for VEGFdosage in reports from experimental and clinical stud-ies. In fact, gene vectors (e.g., adenovirus or plasmidDNA) or protein delivery cannot control the distribu-tion of microenvironmental VEGF levels in vivo. Forexample, adenoviral VEGF delivery results in a mixtureof different vessel types, most of which are unstable andregress (14). These results pose a challenge for thedevelopment of future therapeutic angiogenesis strate-gies. In conclusion, this study revealed that, althoughincreasing the total amount of VEGF in the ischemicmuscle is a prerequisite for the induction of angiogeniceffects, control over the microenvironmental levels ofVEGF delivery is required for the achievement oflong-term functional benefit without significant toxic-ity, suggesting that therapeutic strategies incorporatingthis biological concept have the potential to fullyharvest VEGF’s therapeutic power.
We thank Tim Kovachy and Chris Xu for technical assis-tance. This work was supported by a grant from the Deutsche-Forschungsgemeinschaft to G.v.D. (DE 740/1-1), and Amer-ican Heart Association Scientist Development Grant to A.B.(0430312N) and by NIH grants AG009521, HL065572,HD018179, AG020961, AG024987, and the Baxter Founda-tion to H.M.B.
REFERENCES
1. Simons, M. (2005) Angiogenesis: where do we stand now?Circulation 111, 1556–1566
2. Deindl, E., Buschmann, I., Hoefer, I. E., Podzuweit, T., Boen-gler, K., Vogel, S., van Royen, N., Fernandez, B., and Schaper,W. (2001) Role of ischemia and of hypoxia-inducible genes inarteriogenesis after femoral artery occlusion in the rabbit. Circ.Res. 89, 779–786
3. Masaki, I., Yonemitsu, Y., Yamashita, A., Sata, S., Tanii, M.,Komori, K., Nakagawa, K., Hou, X., Nagai, Y., and Hasegawa, M.,et al (2002) Angiogenic gene therapy for experimental criticallimb ischemia: acceleration of limb loss by overexpression ofvascular endothelial growth factor 165 but not of fibroblastgrowth factor-2. Circ. Res. 90, 966–973
4. Hershey, J. C., Baskin, E. P., Corcoran, H. A., Bett, A., Dough-erty, N. M., Gilberto, D. B., Mao, X., Thomas, K. A., and Cook,J. J. (2003) Vascular endothelial growth factor stimulates angio-genesis without improving collateral blood flow following hind-limb ischemia in rabbits. Heart Vessels 18, 142–149
5. Zhou, Y. F., Stabile, E., Walker, J., Shou, M., Baffour, R., Yu, Z.,Rott, D., Yancopoulos, G. D., Rudge, J. S., and Epstein, S. E.(2004) Effects of gene delivery on collateral development inchronic hypoperfusion: diverse effects of angiopoietin-1 versusvascular endothelial growth factor. J Am Coll Cardiol. 44, 897–903
6. Gounis, M. J., Spiga, M. G., Graham, R. M., Wilson, A., Haliko,S., Lieber, B. B., Wakhloo, A. K., and Webster, K. A. (2005)Angiogenesis is confined to the transient period of VEGFexpression that follows adenoviral gene delivery to ischemicmuscle. Gene Ther. 12, 762–771
7. Schwarz, E. R., Speakman, M. T., Patterson, M., Hale, S. S.,Isner, J. M., Kedes, L. H., and Kloner, R. A. (2000) Evaluation ofthe effects of intramyocardial injection of DNA expressingvascular endothelial growth factor (VEGF) in a myocardialinfarction model in the rat–angiogenesis and angioma forma-tion. J Am Coll Cardiol. 35, 1323–1330
8. Banfi, A., von Degenfeld, G., and Blau, H. M. (2005) Criticalrole of microenvironmental factors in angiogenesis. Curr Athero-scler Rep. 7, 227–234
9. Ozawa, C. R., Banfi, A., Glazer, N. L., Thurston, G., Springer,M. L., Kraft, P. E., McDonald, D. M., and Blau, H. M. (2004)Microenvironmental VEGF concentration, not total dose, deter-mines a threshold between normal and aberrant angiogenesis.J. Clin. Invest. 113, 516–527
10. Carmeliet, P., Ferreira, V., Breier, G., Pollefeyt, S., Kieckens, L.,Gertsenstein, M., Fahrig, M., Vandenhoeck, A., Harpal, K.,Eberhardt, C., et al. (1996) Abnormal blood vessel developmentand lethality in embryos lacking a single VEGF allele. Nature380, 435–439
11. Ferrara, N., Carver-Moore, K., Chen, H., Dowd, M., Lu, L.,O’Shea, K. S., Powell-Braxton, L., Hillan, K. J., and Moore,M. W. (1996) Heterozygous embryonic lethality induced bytargeted inactivation of the VEGF gene. Nature 380, 439–442
12. Miquerol, L., Langille, B. L., and Nagy, A. (2000) Embryonicdevelopment is disrupted by modest increases in vascular endo-thelial growth factor gene expression. Development 127, 3941–3946
13. Dor, Y., Djonov, V., Abramovitch, R., Itin, A., Fishman, G. I.,Carmeliet, P., Goelman, G., and Keshet, E. (2002) Conditionalswitching of VEGF provides new insights into adult neovascular-ization and pro-angiogenic therapy. EMBO J. 21, 1939–1947
14. Sundberg, C., Nagy, J. A., Brown, L. F., Feng, D., Eckelhoefer,I. A., Manseau, E. J., Dvorak, A. M., and Dvorak, H. F. (2001)Glomeruloid microvascular proliferation follows adenoviral vas-cular permeability factor/vascular endothelial growth factor-164gene delivery. Am. J. Pathol. 158, 1145–1160
E2286 Vol. 20 December 2006 VON DEGENFELD ET AL.The FASEB Journal

15. Springer, M. L., Chen, A. S., Kraft, P. E., Bednarski, M., andBlau, H. M. (1998) VEGF gene delivery to muscle: potential rolefor vasculogenesis in adults. Mol Cell. 2, 549–558
16. Luttun, A., Tjwa, M., Moons, L., Wu, Y., Angelillo-Scherrer, A.,Liao, F., Nagy, J. A., Hooper, A., Priller, J., De Klerck, B., et al.(2002) Revascularization of ischemic tissues by PlGF treatment,and inhibition of tumor angiogenesis, arthritis and atheroscle-rosis by anti-Flt1. Nat. Med. 8, 831–840
17. Vajanto, I., Rissanen, T. T., Rutanen, J., Hiltunen, M. O.,Tuomisto, T. T., Arve, K., Narvanen, O., Manninen, H.,Rasanen, H., Hippelainen, M., et al. (2002) Evaluation ofangiogenesis and side effects in ischemic rabbit hindlimbs afterintramuscular injection of adenoviral vectors encoding VEGFand LacZ. J. Gene Med. 4, 371–380
18. Rissanen, T. T., Markkanen, J. E., Arve, K., Rutanen, J., Ket-tunen, M. I., Vajanto, I., Jauhiainen, S., Cashion, L., Gruchala,M., Narvanen, O., et al. (2003) Fibroblast growth factor 4induces vascular permeability, angiogenesis and arteriogenesisin a rabbit hindlimb ischemia model. FASEB J. 17, 100–102
19. Thurston, G., Rudge, J. S., Ioffe, E., Zhou, H., Ross, L., Croll,S. D., Glazer, N., Holash, J., McDonald, D. M., and Yancopoulos,G. D. (2000) Angiopoietin-1 protects the adult vasculatureagainst plasma leakage. Nat. Med. 6, 460–463
20. Banfi, A., Springer, M. L., and Blau, H. M. (2002) Myoblast-mediated gene transfer for therapeutic angiogenesis. MethodsEnzymol. 346, 145–157
21. Jacobi, J., Sydow, S., von Degenfeld, G., Zhang, Y., Dayoub, H.,Wang, B., Patterson, A. J., Kimoto, M., Blau, H. M., and Cooke,J. P. (2005) Overexpression of dimethylarginine dimethylamino-hydrolase (DDAH) reduces tissue ADMA levels and enhancesangiogenesis. Circulation 111, 1431–1438
22. Scholz, D., Ziegelhoeffer, T., Helisch, A., Wagner, S., Friedrich,C., Podzuweit, T., and Schaper, W. (2002) Contribution ofarteriogenesis and angiogenesis to postocclusive hindlimb per-fusion in mice. J. Mol. Cell Cardiol. 34, 775–787
23. Chien, G. L., Anselone, C. G., Davis, R. F., and Van Winkle,D. M. (1995) Fluorescent vs. radioactive microsphere measure-ment of regional myocardial blood flow. Cardiovasc. Res. 30,405–412
24. Thein, E., Raab, S., Harris, A. G., Kleen, M., Habler, O.,Meisner, F., and Messmer, K. (2002) Comparison of regionalblood flow values measured by radioactive and fluorescentmicrospheres. Eur. Surg. Res. 34, 215–223
25. Inai, T., Mancuso, M., Hashizume, H., Baffert, F., Haskell, A.,Baluk, P., Hu-Lowe, D. D., Shalinsky, D. R., Thurston, G.,Yancopoulos, G. D., and McDonald, D. M. (2004) Inhibition ofvascular endothelial growth factor (VEGF) signaling in cancercauses loss of endothelial fenestrations, regression of tumorvessels, and appearance of basement membrane ghosts. Am. J.Pathol. 165, 35–52
26. Rissanen, T. T., Markkanen, J. E., Gruchala, M., Heikura, T.,Puranen, A., Kettunen, M. I., Kholova, I., Kauppinen, R. A.,Achen, M. G., Stacker, et al. (2003) VEGF-D is the strongest
angiogenic and lymphangiogenic effector among VEGFs deliv-ered into skeletal muscle via adenoviruses. Circ. Res. 92, 1098–1106
27. Heeschen, C., Jang, J. J., Weis, M., Pathak, A., Kaji, S., Hu, R. S.,Tsao, P. S., Johnson, F. L., and Cooke, J. P. (2001) Nicotinestimulates angiogenesis and promotes tumor growth and ath-erosclerosis. Nat. Med. 7, 833–839
28. Paek, R., Chang, D. S., Brevetti, L. S., Rollins, M. D., Brady, S.,Ursell, P. C., Hunt, T. K., Sarkar, R., and Messina, L. M. (2002)Correlation of a simple direct measurement of muscle pO(2) toa clinical ischemia index and histology in a rat model of chronicsevere hindlimb ischemia. J. Vasc. Surg. 36, 172–179
29. Baffour, R., Garb, J. L., Kaufman, J., Berman, J., Rhee, S. W.,Norris, M. A., and Friedmann, P. (2000) Angiogenic therapy forthe chronically ischemic lower limb in a rabbit model. J. Surg.Res. 93, 219–229
30. Grinberg, O. Y., Hou, H., Grinberg, S. A., Moodie, K. L.,Demidenko, E., Friedman, B. J., Post, M. J., and Swartz, H. M.(2004) pO2 and regional blood flow in a rabbit model of limbischemia. Physiol. Meas. 25, 659–670
31. von Degenfeld, G., Raake, P., Kupatt, C., Lebherz, C., Hinkel,R., Gildehaus, F. J., Munzing, W., Kranz, A., Waltenberger, J.,Simoes, M., et al. (2003) Selective pressure-regulated retro-infusion of fibroblast growth factor-2 into the coronary veinenhances regional myocardial blood flow and function inpigs with chronic myocardial ischemia. J. Am. Coll. Cardiol. 42,1120 –1128
32. Schaper, W., and Scholz, D. (2003) Factors regulating arterio-genesis. Arterioscler. Thromb. Vasc. Biol. 23, 1143–1151
33. Rissanen, T. T., Korpisalo, P., Markkanen, J. E., Liimatainen, T.,Orden, M. R., Kholova, I., de Goede, A., Heikura, T., Grohn,O. H., and Yla-Herttuala, S. (2005) Blood flow remodels grow-ing vasculature during vascular endothelial growth factor genetherapy and determines between capillary arterialization andsprouting angiogenesis. Circulation. 112, 3937–3946
34. Jain, R. K. (2005) Normalization of tumor vasculature: anemerging concept in antiangiogenic therapy. Science 307, 58–62
35. Ruhrberg, C., Gerhardt, H., Golding, M., Watson, R., Ioan-nidou, S., Fujisawa, H., Betsholtz, C., and Shima, D. T. (2002)Spatially restricted patterning cues provided by heparin-bindingVEGF-A control blood vessel branching morphogenesis. GenesDev. 16, 2684–2698
36. Nehls, V., Denzer, K., and Drenckhahn, D. (1992) Pericyteinvolvement in capillary sprouting during angiogenesis in situ.Cell Tissue Res. 270, 469–474
37. Morikawa, S., Baluk, P., Kaidoh, T., Haskell, A., Jain, R. K., andMcDonald, D. M. (2002) Abnormalities in pericytes on bloodvessels and endothelial sprouts in tumors. Am. J. Pathol. 160,985–1000
Received for publication May 29, 2006.Accepted for publication August 7, 2006.
E2287MICROENVIRONMENTAL VEGF DETERMINES FUNCTION

The FASEB Journal • FJ Express Summary
Microenvironmental VEGF distribution is critical forstable and functional vessel growth in ischemia
Georges von Degenfeld,*,‡ Andrea Banfi,*,§ Matthew L. Springer,*,� Roger A. Wagner,†
Johannes Jacobi,† Clare R. Ozawa,* Milton J. Merchant,* John P. Cooke,† andHelen M. Blau*,1
*Baxter Laboratory in Genetic Pharmacology, Departments of Molecular Pharmacology and ofMicrobiology and Immunology; †Division of Cardiovascular Medicine, Stanford University School ofMedicine, Stanford, California; ‡Cardiovascular Research, Bayer Healthcare, Wuppertal, Germany;§Institute for Surgical Research and Hospital Management, Department of Research and Departmentof Surgery, University Hospital, Basel, Switzerland; and �Division of Cardiology, University ofCalifornia, San Francisco, California
To read the full text of this article, go to http://www.fasebj.org/cgi/doi/10.1096/fj.06-6568fje
SPECIFIC AIMS
Vascular endothelial growth factor (VEGF) expressionrequires exceedingly tight regulation during develop-ment, as either reduced expression after the targeteddeletion of just one allele or modest overexpression leadto early embryonic lethality. Despite the great interest inharnessing the therapeutic potential of VEGF for thetreatment of patients with ischemic conditions, the effectsof different local (microenvironmental) VEGF concentra-tions in ischemia have not been studied in the adultorganism, and clinical trials of VEGF delivery have beendisappointing. This is apparently due to the limitations ofcurrent approaches to delivering the VEGF gene orprotein to the adult organisms which, unlike transgenicmodels, are inherently unable to provide uniform VEGFdistribution. We have developed a novel approach tosystematically study the effects of local (microenvironmen-tal) VEGF concentrations in ischemic muscle: monoclo-nal populations of VEGF-expressing myoblasts that, fol-lowing implantation, fuse with the muscle fibers toprovide uniform VEGF distribution in vivo. Using thistechnique, we investigated the effects of VEGF concentra-tion on functional recovery from ischemia, toxicities, andlong-term stability and morphology/architecture of in-duced vessel growth.
PRINCIPAL FINDINGS
1. Control over local (microenvironmental)distribution improves the efficacy of VEGF deliveryin ischemia without increasing total dose
We generated monoclonal populations of myoblaststhat express equal amounts of VEGF “per cell.” First, apolyclonal population expressing VEGF164 was gener-ated by retroviral transduction, from which single cellswere isolated and expanded to obtain a set of mono-clonal populations. Compared with the parental poly-
clonal myoblast population producing on average 61 �5 ng/106 cells/24 h VEGF in vitro, each of the fourderived clonal populations secreted different VEGFamounts per cell: “10% clone”: 6 � 1; “100% clone”:55 � 5; “180% clone”: 104 � 8, and “325% clone”:191 � 15. All populations also expressed the LacZreporter. Hind limb ischemia was induced in mice bysurgical transection of the superficial femoral artery,and 8 million myoblasts were implanted into the distalthigh muscles: either control myoblasts, the parentalpolyclonal VEGF-expressing population or one of themonoclonal VEGF-expressing populations or vehicle.
Fourteen days after surgery and cell implantation,the control groups injected with vehicle or controlmyoblasts showed strongly reduced blood flow (micro-sphere method) in the ischemic leg compared withthe contralateral nonischemic leg (54.6�6.8% and55.3�13.1%; Fig. 1). The parental polyclonal VEGFpopulation increased blood flow only moderately(74.1�8.6%, P�0.05 vs. controls). Improvement wassubstantially greater in mice that received the 100%VEGF clones, in which blood flow was similar to thecontralateral, nonischemic leg (96.8�10.5%, P�0.01 vs.controls; P�0.05 vs. polyclonal VEGF population) (Fig.1). Ischemia-induced muscle damage (histology) wasreduced only in mice injected with the 100% clone.Importantly, functional improvement was seen despitethe fact that the 100% clone produced similar totalamounts of VEGF to the parental polyclonal populationin vitro and in vivo. Furthermore, delivery of the 100%clone was less toxic, as hemangiomas were avoided andleakage was not increased. These findings show that, byproviding uniform VEGF distribution at the microenvi-ronmental level, therapeutic effects can be markedly
1 Correspondence: Baxter Laboratory in Genetic Pharma-cology, Stanford University School of Medicine, Stanford, CA94305-5175, USA. E-mail: [email protected]
doi: 10.1096/fj.06-6568fje
26570892-6638/06/0020-2657 © FASEB

improved and toxicities avoided compared with non-uniform distribution.
2. A lower and an upper threshold level ofmicroenvironmental VEGF concentrations determinea therapeutic window
The monoclonal populations of VEGF-expressing myo-blasts showed markedly different effects in the ischemicleg. Surprisingly, blood flow was not improved in micehaving received either low (10% clone: 54.7�17.5%) orhigh microenvironmental VEGF doses (180% clone:69.8�18.9%) (Fig. 1). These findings demonstrate theexistence of a therapeutic window determined by micro-environmental VEGF concentrations and indicate thatexpression levels need to be tightly controlled to befunctionally efficacious in the ischemic muscle of theadult organism.
3. VEGF has biphasic effects on vessel number anddiameter
To determine the structural features underlying thedistinct functional effects elicited by different microen-vironmental VEGF doses, we examined the morphol-ogy, length, and diameter of vessel growth by immuno-fluorescence. Myoblast engraftment (�-galactosidaseexpression) was distributed along the needle tracks andcomprised �20% of the cross-sectional muscle area
with no difference between the different populations.Vessel growth was tightly localized around the musclefibers that expressed VEGF. Vessel length density wasincreased in all groups injected with VEGF-expressingmyoblasts. Surprisingly, the effect was not dose depen-dent but peaked in muscles implanted with the 10%clones and was not further increased by even highestVEGF levels.
In contrast, vessel diameter was gradually in-creased by VEGF in a dose-dependent manner withno discernable upper limit over the 30-fold doserange investigated. The parental polyclonal VEGFmyoblasts induced a mixture of small and largevessels, including dilated glomeruloid bodies, aber-rant structures that subsequently grow into hemangi-omas. In contrast, each clonal population uniformlyinduced vessels of distinct morphology. Surprisingly,the 10% clone, found to induce a peak increase invessel number, did not also increase vessel size, butthe 100% and 180% VEGF clones significantly in-creased vessel diameter in a dose-dependent manner.However, whereas the vessels induced by the 100%clone were still normal in morphology, the 180%clone induced irregularly shaped, preangiomatousglomeruloid bodies. The 325% clone induced largehemangiomas visible by microangiography.
4. Control over microenvironmental distributionpromotes collateral growth
The growth of collaterals from preexisting arteries isrequired for blood flow increase in ischemia. Collat-erals were quantified on cryosections of the proximaladductor muscles by costaining for endothelial cells(PECAM) and vascular smooth muscle cells (�-SMA).The number of collaterals �30 �m diameter wasincreased in mice injected with 100% VEGF clonescompared with the controls (P � 0.05), whereas nosignificant collateral growth was seen with the paren-tal polyclonal population or with any of the otherclones. Thus, collateral growth was only seen in thegroup injected with the 100% clone, matching theimproved blood flow and reduced tissue damage.
5. Vessels induced by optimized VEGF delivery arepericyte-covered and persist long-term
Vessels induced by short-term VEGF expression lack peri-cytes, are unstable and prone to regression. On the otherhand, sustained VEGF expression can induce hemangio-mas, posing a challenge for safe and therapeutic VEGFdelivery. To address this issue, we harvested muscles 66 wkafter ischemia induction and implantation of the 100%clone. In the area of cell engraftment (�-galactosidase),vessel density was still clearly increased. Remarkably, noglomeruloid bodies or hemangiomas were found despitelong-term constitutive VEGF expression. We examinedwhether long-term persistence of vessels correlated withpericyte coverage by costaining for endothelial cells (PE-CAM) and pericytes (NG2). Vessels induced by the 100%clone displayed tightly associated pericyte coverage, indi-cating that long-term stability was a result of adequate
Figure 1. Microenvironmental concentrations of vascularendothelial growth factor (VEGF) determine blood flowimprovement in ischemia. Regional blood flow was measuredin the ischemic leg 14 days after surgical induction of hindlimb ischemia using microspheres (n�9/group). Blood flowin ischemic legs (expressed as percentage of the nonischemiccontralateral leg) was severely reduced in mice injected withvehicle or control myoblasts. Implantation of the parentalpolyclonal population of VEGF-expressing myoblasts im-proved blood flow only modestly (*P�0.05 vs. controls). The100% clone markedly improved blood flow in the ischemicleg to a level comparable with the contralateral nonischemicleg (**P�0.01 vs. controls; P�0.05 vs. polyclonal VEGFmyoblasts). In contrast, neither the 10% nor the 180% cloneimproved blood flow significantly.
2658 Vol. 20 December 2006 VON DEGENFELD ET AL.The FASEB Journal

maturation of the newly induced vessels. In contrast,vessels induced by the parental polyclonal VEGF popula-tion had abnormal coverage with pericytes that appeareddetached and expressed smooth-muscle-actin, ultimatelyleading to vascular aberrancies and hemangiomas. Thepresent findings show that VEGF expression that is pro-longed, but at a controlled and uniform level, by the100% clone in ischemic muscle yields mature, pericyte-covered vessels that persist presumably lifelong.
CONCLUSIONS AND SIGNIFICANCE
The most important and clinically relevant finding ofthis study is that controlling the microenvironmentalconcentration of VEGF, without changing total dose,defines a dose window in which constitutive, long-termVEGF gene expression in ischemic muscle predictablyinduces stable and functional vessel growth and effi-ciently rescues blood flow, while reliably avoiding aber-rant angiogenesis and hemangiomas. These findingsshow clearly in a therapeutic setting that, because of thepathophysiological mechanism of VEGF-induced vesselgrowth, average total doses across muscle tissue are notthe critical determinant of efficacy and that local con-centrations determine functional outcome. This studyis, to our best knowledge, the first to show that optimiz-ing the microenvironmental distribution is critical toimproving both efficacy and safety of VEGF in ischemia.
Importantly, vessels induced by the 100% clone werepericyte-covered and persisted more than 15 mo as aresult of constitutive VEGF expression. Remarkably,implantation of 8 million cells of the 100% clone didnot induce hemangiomas, whereas this form of aber-rant vessel growth was invariably induced by the poly-clonal VEGF population even if very few cells wereinjected. Hence, if microenvironmental VEGF concen-trations are kept below the upper threshold level, thetotal VEGF dose in ischemia can be dramatically in-creased without reintroducing the risk of hemangiomagrowth. These findings show that constitutive expres-sion of VEGF has the potential to induce long-term
stable vessels without aberrancies provided that micro-environmental VEGF levels are controlled.
The methodology used here offered the opportunityto specifically “engineer” a set of distinct vessel typesand to systematically correlate their morphology withfunction in ischemia. Blood flow increase and reducedischemic tissue damage provided by the 100% vascularendothelial growth factor clone correlated with specificvascular adaptations: growth of pericyte-covered vesselsthat were enlarged but regularly shaped vessels withoutglomeruloid bodies and collateral growth. A remark-able and unexpected finding was the biphasic effect ofincreasing VEGF doses on the number of inducedvessels (already maximal at the lowest level) and theirdiameter (which instead increased with VEGF dose).The combined increase in vessel size and number wasrequired to achieve blood flow improvement (100%clone), whereas no effect was seen if vessel numberalone was increased without enlargement (10% clone).These findings demonstrate the existence of a lowerthreshold level of VEGF concentration for the induc-tion of functional vessel growth. Contrary to what mighthave been expected, implantation of the 180% clone,secreting still higher levels of VEGF per cell, did notfurther improve blood flow or reduce tissue damagecompared with the 100% clone. Hence, the beneficialeffects seen with the 100% VEGF clone were lost athigher doses, marking an upper threshold of microen-vironmental VEGF concentrations (Fig. 2).
The results of this study help explain the apparentlack of a manageable therapeutic window for VEGF inreports from experimental and clinical studies, as genevectors or protein delivery cannot provide controlledmicroenvironmental VEGF distribution in vivo. In con-clusion, this study revealed that control over the micro-environmental levels of VEGF delivery in ischemiamakes the achievement of long-term functional benefitwithout significant toxicity possible and suggests thattherapeutic strategies incorporating this biological con-cept have the potential to fully reap VEGF’s therapeuticpower.
Figure 2. Uniform VEGF distribution inischemic muscle reveals a therapeuticwindow determined by microenviron-mental concentrations. Comparison ofthe parental polyclonal VEGF populationand the 100% clone shows marked differ-ences in the characteristics of inducedvessels and function, despite the fact thatboth populations express similar totalamounts of VEGF in vitro and in vivo.Blood flow recovery and collateral growthare much more pronounced with the100% clone, showing that microenviron-mental concentrations are critical. Com-parison between the 3 monoclonal pop-
ulations shows that neither the 10% nor the 180% clone induce collateral growth or improve blood flow, revealing a previouslyunrecognized therapeutic window determined by microenvironmental VEGF concentrations.
2659MICROENVIRONMENTAL VEGF DETERMINES FUNCTION