microbial communities performing anaerobic oxidation of...
TRANSCRIPT

Microbial communities performing
anaerobic oxidation of methane:
diversity of lipid signatures and habitats
Dissertation
zur Erlangung des Doktorgrades
der Naturwissenschaften
- Dr. rer. Nat. -
Am Fachbereich Geowissenschaften
der Universität Bremen
vorgelegt von
Pamela E. Rossel Cartes
Bremen
Februar 2009

1. Gutachter: Prof. Dr. Kai-Uwe Hinrichs, University of Bremen, Germany
2. Gutachter: Prof. Dr. Antje Boetius, Max Planck Institute for Marine Microbiology,
Bremen, Germany

No viniste de lejos, ni siquiera has llegado. Estabas desde siempre, como un lenguaje
escrito en el fondo de mí…
Para Xavi con mucho amor


TABLE OF CONTENTS
Abstract Thesis abstract……………………………………………………..I
Kurzfassung……………………………………………………...III
Acknowledgements………………………………………………………………………V
List of Figures……………………………………………………….............................VII
List of Tables………………………………………………………................................IX
List of Abbreviations………………………………………………………....................X
Chapter I: Introduction…………………………………………………….................1
General introduction………………………………………………………2
I.1. Properties and importance of methane………………………………..2
I.2. Production and consumption of methane……………………………..4
I.3. Microbial communities performing AOM…………………..............11
I.4. Distribution/Habitats of AOM communities………………………...13
I.5. Lipid signatures of communities performing AOM…………………18
I.6. Intact polar membrane lipids (IPLs)…..……………………………..21
I.7. Methods……………………………………………………………...28
I.8. Hypothesis and objectives…………………………………………...29
I.9. Contribution to publications…………………………………………30
I.10. References………………………………………………………….33
Chapter II: Intact polar lipids of anaerobic methanotrophic archaea and……………45
associated bacteria
II.1. Printed manuscript…………………………………………………..46
II.2. Supplementary online material……………………………………...61

Chapter III: Factors controlling the distribution of anaerobic………………………...63
methanotrophic communities in marine environments:
evidence from intact polar membrane lipids
III.1. Manuscript…………………………………………………………64
III.2. Supplementary material………..…………………...…………….106
Chapter IV: Experimental approach to evaluate stability and reactivity…………….111
of intact polar membrane lipids of archaea and bacteria in
marine sediments
Chapter V: Diversity of intact polar membrane lipids in marine…………………...125
seep environments
Chapter VI: Concluding remarks and perspectives………………………………….149
VI.1. Conclusions……………………………………………………….150
VI.2. Future perspectives……………………………………………….155
VI.3. Presentations and other activities…………………………………159

Thesis abstract ________________________________________________________________________
I
THESIS ABSTRACT
The main aim of this thesis was to study different microbial communities
involved in the process of anaerobic oxidation of methane (AOM) using lipid analysis.
During this work a variety of globally distributed methane-bearing systems characterized
by different environmental factors and anaerobic methanotrophic consortia were analyzed
for intact polar lipid (IPL) and apolar lipid composition. Moreover, an experiment was
designed in order to evaluate the stability of archaeal and bacterial IPLs in marine
sediments.
The three phylogenetically distinct clusters of Euryarchaeota called ANME-1, -2
and -3, which have been observed in association with sulfate-reducing bacteria of the
Desulfosarcina/Desulfococcus group (‘‘ANME-1/DSS and -2/DSS aggregates”) or
Desulfobulbus spp (‘‘ANME-3/DBB aggregates”) could be clearly distinguished by IPL
composition but not by apolar lipids. ANME-1/DSS was characterized by
glyceroldialkylglyceroltetraethers (GDGTs) with glycosidic, phospho, as well as mixed
of both , whereas diagnostic IPLs of ANME-2/DSS were archaeols with both glycosidic
and phospho headgroups. Distinctly, ANME-3/DBB contained neither glycosidic-
archaeols nor GDGT-based IPLs, but the phospho-archaeol composition was very similar
to ANME-2/DSS. The main and distinguishing feature of ANME-3/DBB was the high
contribution of the bacterial IPLs phosphatidyl-(N)-methylethanolamine (PME) and
phosphatidyl-(N,N)-dimethylethanolamine (PDME). Other bacterial IPLs that were
mainly found in ANME-2/DSS-dominated carbonate mats were IPLs with non-phospho
headgroups such as ornithine lipids, surfactins and betaine lipids, the latter with odd fatty
acid chains. In contrast, IPLs with phospho headgroups were generally more abundant in
sediment environments. The high contribution of glycosidic archaeal IPLs and the
presence of bacterial IPLs with non-phospho headgroups in carbonate mats can be
explained by adsorption of phosphate onto calcium carbonate.
In addition to the general differences in IPL composition of each of three AOM-
community types, the IPL distribution was also associated with several environmental
factors, allowing the characterization of their different habitats. ANME-1/DSS dominates

Thesis abstract ________________________________________________________________________
II
habitats with high temperature and low oxygen content in bottom waters. For ANME-
2/DSS systems, it was possible to differentiate between carbonate reef habitats and
sediment settings, with the former characterized by low temperature, high oxygen content
in bottom waters and high methane and sulfate concentrations, whereas the latter was
associated with higher sulfate reduction rates. ANME-3/DBB presented similar
environmental characteristics to ANME-2/DSS.
Furthermore, degradation of archaeal and bacterial IPLs was evaluated in marine
sediments, showing a loss of 80% for the archaeal and ~50% for the bacterial IPL at 5°C
after 465 days of incubation under sterile conditions. However, in non-sterile conditions
at 5°C, an increase in concentration of both IPLs at the end of the experiment was
observed. Therefore, biotic degradation of IPLs could not be proved because the pools of
produced and degraded IPLs in the non-sterile conditions were indistinguishable.
The results obtained during this thesis support the distinction of microbial
communities performing AOM based on IPL diversity and address the role of
environmental factors in the distribution of three major AOM-community types. This
work contributes substantially to the understanding of the distribution of AOM systems
on a global scale.

Kurzfassung ________________________________________________________________________
III
KURZFASSUNG
Der Schwerpunkt dieser Doktorarbeit liegt auf der Untersuchung von
unterschiedlichen Mikrobengemeinschaften, die an der anaeroben Oxidation von Methan
(AOM) beteiligt sind mit Hilfe von Lipidanalysen. Die Zusammensetzung von apolaren
und intakten polaren Lipiden (IPLs) wurde an einer breitgefächerten Auswahl von
methangeladenen Systemen analysiert, die durch verschiedene Umweltfaktoren und
anaerobische methanotrophische Konsortien charakterisiert sind. Außerdem wurde ein
Experiment konzipiert, um die Stabilität von bakteriellen und von Archaeen stammenden
IPLs in marinen Sedimenten zu untersuchen.
Die drei phylogenetisch unterschiedlichen Cluster von Euryarchaeen namens
ANME-1, -2 und -3, die oft zusammen mit sulfatreduzierenden Bakterien der Gruppe
Desulfosarcina/Desulfococcus (‘‘ANME-1/DSS und -2/DSS Aggregate”) oder
Desulfobulbus spp (‘‘ANME-3/DBB Aggregate”) beobachtet worden sind, konnten
eindeutig anhand der Zusammensetzung ihrer IPLs unterschieden werden, aber nicht
durch ihre apolaren Lipide. Charakteristisch für ANME-1/DSS sind
Glyceroldialkylglyceroltetraether (GDGT) mit sowohl glykosidischen, phospho und
gemischten Kopfgruppen, wohingegen diagnostische IPLs für ANME-2/DSS Archaeole
mit sowohl glycosidischen als auch phospho Kopfgruppen waren. Im Gegensatz dazu
zeigten ANME-3/DBB weder glykosidische Archaeole noch GDGT-basierte IPLs, aber
dafür eine zu ANME-2/DSS sehr ähnliche Zusammensetzung der Phosphoarchaeole. Der
größte Unterschied von ANME-3/DBB waren die bakteriellen IPLs phosphatidyl-(N)-
methylethanolamine (PME) und phosphatidyl-(N,N)-dimethylethanolamine (PDME).
Andere bakterielle IPLs, die hauptsächlich in ANME-2/DSS dominierten Karbonatmatten
gefunden wurden waren IPLs ohne phosphatbasierende Kopfgruppe wie Ornithinlipide,
Surfactin und Betainlipide, letztere mit ungeraden Fettsäureketten. Im Gegensatz dazu
hatten Lipide mit phosphatbasierenden Kopfgruppen einen höheren Anteil in
sedimentären Umgebungen. Der hohe Anteil von glykosidischen Archaeenlipiden und
bakteriellen IPLs ohne phosphatbasierende Kopfgruppen in Karbonatmatten kann durch
die Adsorption von Phosphat an Kalziumcarbonat erklärt werden.

Kurzfassung ________________________________________________________________________
IV
Zusätzlich zu den allgemeinen Unterschieden der IPL Zusammensetzung der drei
AOM-Gemeinschaften, war die Verteilung der IPLs auch mit verschiedenen
Umweltfaktoren verknüpft, was die Charakterisierung deren unterschiedlichen
Lebensräume ermöglicht. ANME-1/DSS dominiert Umgebungen mit hoher Temperatur
und niedrigem Sauerstoffgehalt im Bodenwasser. Für ANME-2/DSS Systeme war es
möglich zwischen Karbonatriffen und Sedimenten zu unterscheiden, wobei Erstere durch
niedrige Temperaturen, hohen Sauerstoffgehalt im Bodenwasser und hohe Methan- und
Sulfatkonzentrationen charakterisiert sind, während Letztere mit hohen
Sulfatreduktionraten verbunden waren. ANME-3/DBB zeigte ähnliche
Umweltcharakteristika wie ANME-2/DSS.
Zusätzlich wurde die Degradation von bakteriellen und von Archaeen
stammenden IPLs in marinen Sedimenten untersucht. Nach Inkubation für 465 Tage
unter sterilen Bedingungen bei 5°C wurde ein Abbau von 80% des Archaeen- und ~50%
des Bakterienlipids beobachtet. Unter nicht sterilen Bedingungen bei 5°C hingegen
wurde ein Anstieg der Konzentration von beiden IPLs am Ende des Experiments
festgestellt. Deshalb konnte der biologische Abbau von IPLs nicht belegt werden, da die
Pools von produzierten und abgebauten IPLs unter nicht-sterilen Bedingungen
ununterscheidbar waren.
Die Ergebnisse dieser Doktorarbeit zeigen, dass es möglich ist die verschiedenen
Mikrobengemeinschaften die an AOM beteiligt sind anhand ihrer IPL Zusammensetzung
zu unterscheiden und deuten auf die Rolle von Umweltfaktoren bei der Verteilung der
drei Typen von AOM Gemeinschaften hin. Diese Studie trägt wesentlich zum
Verständnis der Verteilung von AOM Systemen im globalen Maßstab bei.

Acknowledgements ________________________________________________________________________
V
ACKNOWLEDGEMENTS
I started my scientific career as a marine biologist, followed by a master in
oceanography, period during which I acquired the first knowledge about organic
geochemistry. This small background was widely extended during the realization of my
PhD under the supervision of Prof. Kai-Uwe Hinrichs, who gave me the opportunity to
join his working group. Thanks Kai for providing me support and inspiration during these
over three and half years. I would also like to thank the co-supervision of Marcus Elvert,
who contributed to my knowledge in GC and GC-MS and for the interesting and helpful
discussions. I am also grateful to Julius Lipp and Helen Fredricks for guiding my first
steps with HPLC-MS and in the analysis of IPLs. I would also like to thank the thesis
committee members for their review of my dissertation.
Additionally, I would like to thank all the colleges from the MPI in Bremen
involved in the MUMM project especially Antje Boetius, Tina Treude, Katrin Knittel,
Julia Arnds, Helge Niemann, Gunter Wegener, Janine Felden and Thomas Holler, for
supplying samples and for the useful discussions. I am also indebted to Julia Arnds,
Katrin Knittel, Antje Boetius and Alban Ramette for contributing in great part to the
work included in this thesis. Moreover, I would like to thank my friend Beth! Orcutt for
providing me samples from the Gulf of Mexico, together with some unpublished data
from this setting. Thanks also to Helge Niemann, Tina Treude and Janine Felden for
providing me some unpublished data. Thanks also to Florence Schubotz who helped me
with her expertise in bacterial IPLs and also for sharing unpublished data from the Black
Sea.
Thanks to Birgit Schmincke for being always so helpful with the administrative
paper work.
A special thank to all my colleges and friends from the Organic Geochemistry and
Geobiology groups in Bremen for providing a nice and pleasant working atmosphere.
Thanks for the interesting collaboration work with our lab guests John Pohlman and
Maria Pachiadaki. Thanks to Marcus and Xavi for technical support in the lab. I would
like to thanks also my friends Marcos Yoshinaga, Julius Lipp and Julio Sepulveda for
reading and reviewing part of my work.

Acknowledgements ________________________________________________________________________
VI
Thanks to Julio to be my brother all these years, to share so many histories and
experiences that I will never forget (gracias peladito espero que nuestros caminos se
junten nuevamente). Thanks also to Annette and Amaya; you have been my family in
Bremen, thanks for always being there in the good and bad moments, I will miss all of
you very much.
Thanks to my German teacher and good friend Ursula, who made me enjoy so
much the two hours of German lessons every Friday. I am very glad that I decided to stay
in Bremen, so I will be able to continue with that.
Thanks to my family in Barcelona, Montserrat, Julià and Jordi, for receiving me
as my own family, for taking care of me and giving me support during this PhD.
Thanks to my friends from South America, which despite the distance are always
so close to me: Lilian Nuñez, Andrea Elgueta, Jaime Letelier, Klaudia Hernandez,
Pamela Vaccari, Carlos Tapia and Marcelo Ayala. Thanks to my friends in Bremen for
giving me many great moments and to make me feel at home: Claudia & Sven, Petra,
Luisa, Elvan & Jerome, Cécile & Rick, Flo & Julius, Mathias & Susanne, Barbara &
Marius, Xavier & Gulnaz, Catalina, Ilham and Jeroen. To my former advisors and
friends Silvio Pantoja and Carina Lange, thanks for being always there.
A word of thanks to my family in Chile, Margarita, Gabriel, Soledad, Camila,
Aylin and Gabriel son, thanks for believe in me and give me your support during these
years. Especially to you mother for being a great friend and inspiring woman so strong
and perseverant, despite all the things you have being through, without you I wouldn’t be
this person.
Finalmente a Xavi, gracias por quererme tanto y por ser tan paciente en especial
este ultimo año. Gracias por tu compañía y atenciones. Por tu risa, tus miradas y caricias.
Espero seguir siendo tu compañera de viaje siempre en el polvo del tiempo. Este trabajo
te lo dedico a ti.

List of Figures ________________________________________________________________________
VII
LIST OF FIGURES
Figure I.1. Three-dimensional structure of the methane molecule………………..2
Figure I.2. Gas hydrate stability zone in the marine environment...………………3
Figure I.3. Model of methane hydrate structure…...……...………………………3
Figure I.4. Methane, temperature and past climate changes…...……….………....4
Figure I.5. Sources of atmospheric methane…………………………….……......5
Figure I.6. Classification of natural methane sources……………...………….......6
Figure I.7. Redox sequence in marine sediments………….……………………...7
Figure I.8. Phylogeny of archaea……………………………….………………....8
Figures I.9. Enzymatic pathway of CO2 reduction……………….………...............9
Figure I.10. Production and consumption of methane in marine sediments...........10
Figure I.11. Phylogenetic tree of Euryarchaeota including
anaerobic methanotrophic archaea (ANME)…………………...……12
Figure I.12. Methane-dependent sulfate reduction in ANME-1 and
ANME-2 in response to temperature variability..................................13
Figure I.13. Community distribution in relation to fluid flow……….….………...14
Figure I.14. Global distribution of ANMEs based on phylogenetic data..………..15
Figure I.15. Apolar lipids derived from ANME-1 and ANME-2............................20
Figure I.16. Phospholipid membrane bilayer.………….…………....……………22
Figure I.17. General features of archaeal and bacterial membranes…………........23
Figure I.18. HPLC-MS chromatogram from an IPL mixture………...…………...25
Figure I.19. Diversity of IPLs..……………………..……...……………………...26
Figure I.20. Characteristic mass spectra of PE in positive and
negative ion modes…………………………………..………………27
Figure II.1. Composite mass chromatograms of samples dominated by
different ANME communities…………………………………….…51
Figure II.2. Distribution of IPLs in AOM communities………………………….54
Figure II.3. Structure of IPLs…………………………..…………………………61
Figure III.1. Grouping of samples according to the dominance of
GDGT- and AR-based IPLs………………………………………….78

List of Figures ________________________________________________________________________
VIII
Figure III.2. Principal Component Analysis showing the distribution
of IPLs among the analyzed samples………………...………………81
Figure III.3. Redundancy Analysis in function of environmental data……………89
Figure III.4. Location of the samples included in the global survey……………..106
Figure III.5. Principal Component Analysis showing the distribution
of bacterial IPLs………………………………...…..………………107
Figure III.6. Principal Component Analysis showing the distribution
of apolar lipids among the samples…………………………………108
Figure IV.1. Experimental design of the degradation study……………………...115
Figure IV.2. Degradation of archaeal and bacterial IPLs at 5°C and 40°C
in sterile sediments………….…………………………….………...117
Figure IV.3. Degradation of archaeal and bacterial IPLs at 5°C and
40°C in active sediments………………………………….………...119
Figure IV.24 Variability of GDGT cores in sediments incubated at 5°C
in active sediments …………..………………………..….………...120
Figure V.1. MS2 positive ion spectra of glycosidic archaeols...………………...130
Figure V.2. MS2 positive ion spectra of glycosidic GDGTs.....………………...132
Figure V.3. MS2 positive ion spectra of phospholipid archaeols…………....….134
Figure V.4. MS2 positive ion spectra of phospholipid GDGTs…...…………….135
Figure V.5. MS2 positive ion spectra of the phospholipids PE
and its methyl derivates...…………………………………………..136
Figure V.6. MS2 positive ion spectra of ornithine lipids………………….....….137
Figure V.7. MS2 positive ion spectra of betaine lipids……………………....….138
Figure V.8. MS2 positive ion spectra of surfactins…...……………………...….139
Figure V.9. MS2 positive ion spectra of unknown IPLS a and b…….……...….141

List of Tables ________________________________________________________________________
IX
LIST OF TABLES
Table I.1. General guidelines to distinguish phospholipids…………………….27
Table II.1. Overview of analyzed samples and IPLs…………………………….50
Table III.1. Overview of analyzed samples, with sample location
and AOM-phylotypes……………………………………………..….68
Table III.2. Environmental data selected for redundancy analysis……………….72
Table III.3. Lipid code and source assignment of detected IPLs…………………75
Table III.4. Relative abundance of IPLs in percentage……………….………....109
Table III.5. Concentration of apolar lipids……………………….……………...110
Table IV.1. Frequency of analysis in experiments performed to test IPLs
stability…………….……………………………………………......116 Table V.1. IPL diversity in seep environments………………………….……...142

List of Abbreviations ________________________________________________________________________
X
LIST OF ABBREVIATIONS
16S Rrna Small ribosomal ribonucleic acid unit with a sedimentary unit of 16
ANME Anaerobic methanotrophic archaea
AOM Anaerobic oxidation of methane
APCI Atmospheric pressure chemical ionization
APT Phosphoaminopentatetrol
AR Archaeol
AS Arabian Sea
Beg Beggiatoa
BL Betaine lipids
BS Black Sea
Calyp Calyptogena
CARD-FISH Catalyzed reporter deposition fluorescent in situ hybridization
CH4 Methane concentration
Da Dalton
DAG Diacylglycerol
DAGEs sn-1,2-di-O-alkyl glycerol ethers
DCM Dichloromethane
DEG Dietherglycerol
DNA Desoxyribonucleic acid
EMS Eastern Mediterranean Sea
ER Eel River Basin
ESI Electrospray ionization
FA Fatty acid
FAME Fatty acid methyl esters
FISH Fluorescent in situ hybridization
GB Guaymas Basin
GC-MS Gas chromatography-mass spectrometry
GDGT Glyceroldialkylglyceroltetraether
GF Gullfaks oil field

List of Abbreviations ________________________________________________________________________
XI
Gly Glycosyl
GOM Gulf of Mexico
HMMV Håkon Mosby Mud Volcano
HPLC-MS High performance liquid chromatography mass spectrometry
HR Hydrate Ridge
IPL Intact polar membrane lipid
m/z mass to charge ratio
MAGE sn-1, mono-O-alkyl glycerol ether
MAPT Phosphomethylaminopentatrol
MAR Macrocyclic archaeol
MeOH Methanol
MS1 Primary order mass spectrometry stage
MS2 Secondary order daughter ion mass spectra
MSn Higher order daughter ion mass spectra
MUMM Methane in the Geo/Bio-System-turnover, metabolism and microbes
O2 Oxygen concentration in bottom waters
OH-AR Hydroxyarchaeol
OL Ornithine lipids
OM Organic matter
PAF Platelet activation factor (1-O-hexadecyl-2-acetoyl-sn-glycero-3-
-phosphatidylcholine)
PC Phosphatidylcholine
PCA Principal component analysis
PDME Phosphatidyl-(N,N)-dimethylethanolamine
PE Phosphatidylethanolamine
PG Phosphatidylglycerol
PI Phosphatidylinositol
PME Phosphatidyl-(N)-methylethanolamine
PMI 2,6,15,19-pentamethylicosane
PS Phosphatidylserine
RDA Redundancy analysis

List of Abbreviations ________________________________________________________________________
XII
rDNA Ribosomal ribonucleic acid
SMTZ Sulfate methane transition zone
SO42- Sulfate concentration
SOB Sulfide oxidizing bacteria
SR Sulfate reduction
SRB Sulfate reducing bacteria
SRR Sulfate reduction rate
Thio Thioploca
TLE Total lipid extract
TOC Total organic carbon
TOF-SIMS Time of flight mass spectrometry
VFA Volatile fatty acids

Chapter I ________________________________________________________________________
1
CHAPTER I
Introduction

Chapter I ________________________________________________________________________
2
GENERAL INTRODUCTION
The first chapter provides an overview about the significance of methane in the
global carbon cycle and a description of different processes during methane production
and consumption. Furthermore, this section will give an introduction to the role of the
oceans and the microorganism inhabiting marine sediments in the global methane budget.
A dominant part is dedicated to the identification of diverse microbial communities
involved in the anaerobic oxidation of methane (AOM) from widely distributed
hydrocarbon rich sediments. Finally, the last part of this section includes the main
objectives of this work.
I.1. Properties and importance of methane
Fig I.1. Three-dimensional tetrahedron of the methane molecule.
Methane is the simplest organic
molecule and the most reduced form of
carbon. Methane represents the main
component of natural gas, although this
can occur with other hydrocarbons such
as ethane, propane and butane. Methane
has a molecular weight of 16.04 and
consists of a central carbon atom
covalently bonded to four hydrogen
atoms (tetrahedron, Fig. I.1).
Methane solubility in water is rather low (~2,5 mM at 0°C and 1 atm of pressure)
and it is negatively affected by temperature (Duan et al., 1992) and salinity (Yamamoto et
al., 1976). Contrary to salinity and temperature, pressure has a positive effect on methane
solubility according to Henry’s law. However, in the marine environment, the
combination of low temperature and high pressure conditions enables the mixture of

Chapter I ________________________________________________________________________
3
methane and water molecules resulting in hydrate formation (Fig. I.2), which is a
crystalline, ice-like structure known as methane clathrate (Fig. I.3). Three different
methane clathrate structures have been described (I, II and H) and among these, structure
I is based on pure methane, while the other ones also include ethane, propane or butane
(Buffett, 2000). The stability of methane hydrates is also affected by the inclusion of
various ions and additional gases such as hydrogen sulfide or carbon dioxide (Fig. I.2).
Fig. I.2. Gas hydrate stability zone in the marine environment in relation to pressure and temperature (after Kvenvolden, 1998).
Fig. I.3. Model of methane hydrate structure I. Gas and water molecules are displayed in green and blue, respectively (Rehder and, Suess, 2004).
Methane is an important greenhouse gas due to its ability to absorb and re-emit
radiation, trapping the heat 25 times more efficiently than carbon dioxide (Lelieveld et
al., 1998). Thus, several studies focused on the relation between methane inventory, i.e.
fluctuations in atmospheric methane concentration, and temperature during glacial-
interglacial cycles (Petit et al 1999, Wuebbles and Hayhoe 2002, Kasting, 2004). These
studies provided strong evidence for the positive correlation of the greenhouse gas
content in the atmosphere (CO2 and CH4) and the temperature record of Antarctica during
the past four glacial-interglacial cycles (Fig. I.4).

Chapter I ________________________________________________________________________
4
Fig. I.4. Variations of methane, CO2 and temperature recorded in the Vostok ice core (Petit, 1999).
Past global warming events have been related to an increase in the emissions of
methane gas to the atmosphere. Among the responsible sources for these releases,
methane hydrate dissociation has been discussed. Dickens (2004) suggests that the
depleted �13C values from several sediment cores from north and central Atlantic Ocean
during the warming period of the initial Eocene maximum (IETM), at about 55 million
years ago, can be explained by a methane release from gas hydrate source. Similarly,
Kennett et al. (2002), based on the light �13C values of benthic and planktonic
foraminifera recorded in a core from the Santa Barbara basin, proposed that the end of the
last glacial maximum was caused by a big methane release due to a destabilization of gas
hydrates, idea which is know as the clathrate gun hypothesis.
I.2. Production and consumption of methane According to the Intergovernmental Panel on Climate Change (IPCC), methane
concentration in the atmosphere has increased by ~150% since pre-industrial times
(IPCC, 2001). Several sources have been identified which contribute to the release of
methane to the atmosphere (Fig. I.5, Reeburgh, 2007). Among these, human-related
sources such as rice cultivation contribute with 20%, production of coal with 7%, and

Chapter I ________________________________________________________________________
5
ruminant animals with 16%. Additionally, incomplete combustion of organic matter and
degradation of organic carbon in landfills contribute with 11% and 8%, respectively.
Fig. I.5. Sources of atmospheric methane in Tg (1012g) and relative contribution presented in percentages (in parentheses) of the total (Reeburgh, 2007).
Natural sources of methane
include wetlands, termites, oceanic and
geological sources. Wetlands contribute
with 23%, while termites contribute only
with 4% (based on cellulose utilization
by methanogens living in their guts).
Ocean and freshwater contributes with
2%, while geological sources, like
hydrates and gas production (including
seeps) contribute with 1% and 8% to the
atmosphere methane budget,
respectively. However, the real
contribution of hydrates is still not very
well constrained.
Several of the identified sources of methane release are not affected by microbial
consumption such as animal production, biomass burning, coal production and venting or
methane flaring. Contrary to these sources, the oceans play an effective role in
controlling methane emissions to the atmosphere with only 2% of contribution in the
methane global budget, although they cover 70% of the Earth surface (Reeburgh, 2007).
The use of stable isotopes to distinguish natural methane sources is a very
common approach. The isotopic value of methane in nature can be affected by the
contribution of the different isotopomers (12C, 13C and 1H, 2H). During the utilization of
carbon by living organisms a discrimination against the heavier isotope (13C) results in
products enriched in 12C (lower or more negative �13C value, Eq. 1). However, different
metabolic pathways can discriminate differently against 13C. The �13C value is expressed
as per mil (‰) deviation from VPDB (Vienna Pee Dee Belemnite standard) according to
equation 1.

Chapter I ________________________________________________________________________
6
� �� �
31213
121313 101
Standard/Sample /
��
���
�
CCCCC� Eq. 1
Fig. I.6. Bernard-diagram used for the classification of natural methane sources (Whiticar, 1999).
Sources of methane can be
classified as thermogenic or
biogenic/bacterial (Fig. I.6, Whiticar,
1999 and references therein).
Thermogenic methane is formed during
thermocatalytic degradation of kerogen
at temperatures above ~120°C (Tissot
and Welte, 1984) and it is generally
more enriched in 13C (�13C > -50‰) than
the methane from biogenic sources (�13C
< -50‰; Whiticar, 1999).
Methane derived from bacterial sources is restricted to lower temperatures (< 60°C,
Ziebis and Haese, 2005) and shows carbon isotopic compositions which are dependent on
the environment (freshwater and marine or saline sediments). Bacterial methane from
marine environments is generally more depleted in 13C compared to freshwater
ecosystems, resulting from the dominance of CO2-reduction as opposed to acetoclastic
methanogenesis. Furthermore, the relation between �13C values and the occurrence of
longer chain hydrocarbons relative to methane expressed by the ratio C1/(C2+C3) also
provides information about the methane source, with values of less than 50 and more than
100 for thermogenic and microbial origin, respectively (Whiticar, 1999).
During the microbial degradation of organic matter in sediments, macromolecular
organic compounds are broken down into smaller molecules in a sequence of redox
reactions (Fig. I.7, Jørgensen, 2001). This redox sequence ends with the generation of
methane by methanogenic archaea, which either use carbon dioxide or other low
molecular weight compounds (formate, acetate, methanol and methylated amines) as
substrates under anaerobic conditions. Among the metabolic pathways used to produce
methane (Eq. 2a-e), the production of methane by CO2 reduction (Eq. 2a) and acetoclastic
metanogenesis (Eq. 2d) are the most important.

Chapter I ________________________________________________________________________
7
Fig. I.7. Redox sequence during the degradation of organic matter in marine sediments (Jørgensen, 2001).
Methanogenic reactions:
CO2 reduction:
OHCHHCO 2422 24 � , �G0= -135.6 Eq.2a
Methanol reduction:
OHCHHOHCH 2423 � , �G0= -112.5 Eq.2b
Disproportionation of formate:
OHCOCHHHCOO 224 2344 � , �G0= -130.1 Eq.2c
Acetoclastic methanogenesis:
243 COCHHCOOCH � , �G0= -31.0 Eq.2d
Disproportionation of methylamines: � 424233 4324 NHCOCHOHNHCH , �G0= -75.0 Eq.2e
Methanogens are strictly anaerobic microorganisms, due to instability of the
hydrogenase enzyme complex F420 in the presence of oxygen, nitrate and nitrite
(Schönheit et al., 1981). This coenzyme works as electron donor during the reduction of
different one-carbon intermediates involved in CO2 and methanol reduction (Hedderich

Chapter I ________________________________________________________________________
8
and Whitman, 2006). Methanogens are represented by five orders of the Euryarchaeota:
Methanobacteriales, Methanococcales, Methanomicrobiales, Methanosarcinales and
Methanopyrus (Fig. I.8). Among these groups, different metabolic pathways have been
described. The utilization of CO2, formate or methanol (Methanobacteriacea), CO2 or
formate (Methanococcacea), CO2, formate or alcohols (Methanomicrobiacea), as
substrate has been observed (Blotevogel and Fisher, 1985; Jones et al., 1987; Hedderich
and Whitman., 2006). Additionally, Methanosarcinales can also disproportionate
methanol, use acetate, methylamines and other methylated compounds to produce
methane (Eq.2b, d and e) (Ferguson and Mah, 1983; Jones et al., 1987; Hedderich and
Whitman., 2006).
Fig. I.8. Phylogeny of archaea. Euryarchaeotal methanogens are displayed in red (Kasting, 2004).

Chapter I ________________________________________________________________________
9
Fig. I.9. Enzymatic pathway of CO2 reduction (Hedderich and Whitman, 2006). Abbreviations: MFR, methanofuran; H4MPT, tetrahydromethanopterin, S-CoM, coenzyme M and B, CoM-S-S-CoB; reduced coenzyme F420H2
During methanogenic
reactions a complex series of
enzymes are involved (e.g., CO2
reduction, Fig. I.9). However,
besides the different carbon sources
used during methanogenesis, all
methanogens share the same final
step in which the methyl-coenzyme
M reductase (mcr) catalyzes the
reaction between the methyl-
coenzyme M and the coenzyme B
promoting the reduction of the
methyl group into methane.
Methane oxidation in the troposphere and stratosphere is caused by the production
of hydroxyl radicals during UV degradation of ozone (Lelieveld et al., 1998). In the
biosphere, methane consumption is microbially-mediated under both aerobic and
anaerobic conditions (Eq. 3a and b), thus reducing the escape of methane to the
atmosphere.
10
2224 842,22 �� � molkJGOHCOOCH Eq. 3a
1023
244 25, �� � molkJGOHHSHCOSOCH Eq. 3b
Aerobic methanotrophy is performed by bacteria utilizing the methane
monooxygenase enzyme. Aerobic methanotrophs are members of the �, � and �
subdivision of the Proteobacteria (Hanson and Hanson, 1996). These bacteria are
ubiquitously occurring in soils, sediments, water and also as endosymbionts of mussels.
Based on different metabolic pathways used during the oxidation of methane and
assimilation of formaldehyde, aerobic methanotrophs are classified as type I, II or X
(Hanson and Hanson, 1996). Type I methanotrophs use the ribulose monophosphate
(RuMP) pathway, whereas type II methanotrophs use the serine pathway. Methanotrophs

Chapter I ________________________________________________________________________
10
of the type X can use both pathways. The utilization of other carbon sources besides
methane, such as chlorinated hydrocarbons, has also been observed in methanotrophs.
The utilization of chlorinated hydrocarbons by this group of bacteria makes these
microbes commercially interesting (e.g., Hanson and Hanson, 1996).
The recognition of anaerobic oxidation of methane (AOM) was reported for the
first time in the mid 70’s in anoxic marine sediments (Martens and Berner, 1974; Barnes
and Goldberg, 1976; Reeburgh, 1976). For a long time, oxidation of methane was
assumed to take place only under oxic conditions. However, due to the rapid utilization of
oxygen during the organic matter degradation, aerobic oxidation of methane is very
limited in marine sediments.
The diffusion of methane from deep sediments and its disappearance before
reaching the oxygen layer pointed to the utilization of methane in the presence of another
electron acceptor.
Fig. I.10. Scheme showing production and consumption of methane in marine sediments (figure obtained from ifm-geomar.de after Whiticar, 1999 and DeLong, 2000).
Barnes and Goldberg
(1976) proposed sulfate as most
possible electron acceptor in this
process due to the simultaneous
consumption of both methane and
sulfate in the sulfate methane
transition zone (SMTZ) of marine
sediments (Fig. I. 10). The
utilization of sulfate as electron
acceptor during AOM was later
confirmed by the detection of
radioactively labeled products (i.e.,
sulfide and CO2) formed during
turnover of artificially labeled
substrates (i.e., 14CH4 and 35SO42-)
in sediments from the SMTZ (Devol, 1983; Iversen and Jørgensen, 1985).
The process of AOM, contrary to aerobic methanotrophy, results in increased
alkalinity (Eq. 3b, Barnes and Goldberg 1976), which favors the precipitation of

Chapter I ________________________________________________________________________
11
carbonate. The precipitates formed during AOM are mainly aragonites and Mg-rich
calcites, which can vary in shape and size ranging from small crystals (Aloisi et al., 2000)
to carbonate chimneys (Michaelis et al., 2002) and are preserved in time back to the
Carboniferous (~300 My; Birgel et al., 2008).
OHHSCaCOCaSOCH 2322
44 � Eq. 4
After the first reports of AOM three decades ago, subsequent investigations have
provided detailed evidence of Archaea and Bacteria involved in AOM. Based on field
and laboratory studies, Hoehler et al. (1994) proposed for the first time the presence of a
consortium of methanogenic archaea and sulfate reducing bacteria (SRB) in sediments of
Cape Lookout Bight, North Carolina. These authors suggested that AOM is
thermodynamically favorable at hydrogen concentrations below 0.3 nM. Because the
energy yield produced during AOM is approximately half of the energy necessary to
produce an ATP molecule (Eq. 3b), the growth rates of methanotrophic communities in
natural environments has been of controversial debate. However, the discovery of large
amounts of AOM biomass from different methane-rich environments has provided
indisputable evidence for the feasibility of this process (Boetius et al., 2000; Michaelis et
al., 2002).
I.3. Microbial communities performing AOM During the last ten years subsequent studies have reported different microbial
groups responsible for AOM in marine sediments. Because ANaerobic MEthanotrophs
(ANME) have not been successfully isolated so far, information has been dominantly
obtained from cultivation-independent techniques. Among these, the analysis of 16S
rRNA and lipid biomarkers have been mostly applied, providing evidence for the
occurrence of three main clusters in the Euryarchaeota named ANME-1, ANME-2 and
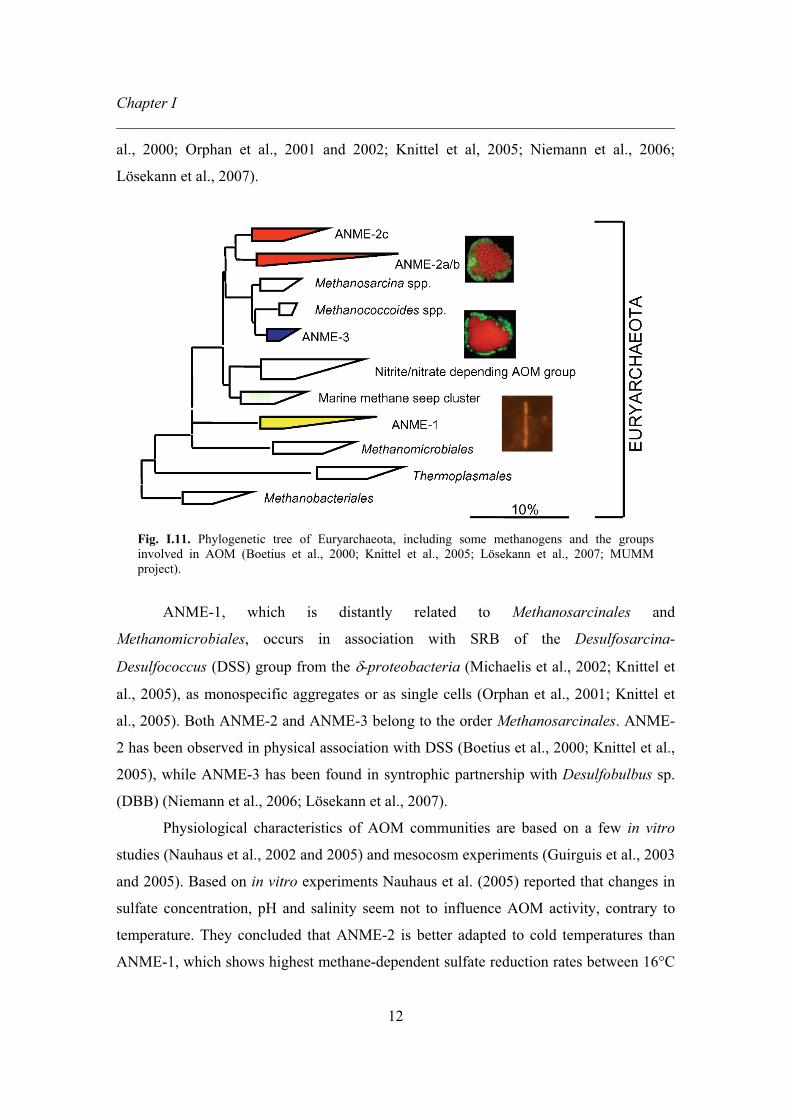
ANME-3 (Fig. I.11). These cluster were found in close association with two dominant
groups of SRB (SEEP-SRB1 and 4) involved in AOM (Hinrichs et al., 1999; Boetius et
Chapter I ________________________________________________________________________
12
al., 2000; Orphan et al., 2001 and 2002; Knittel et al, 2005; Niemann et al., 2006;
Lösekann et al., 2007).
Fig. I.11. Phylogenetic tree of Euryarchaeota, including some methanogens and the groups involved in AOM (Boetius et al., 2000; Knittel et al., 2005; Lösekann et al., 2007; MUMM project).
ANME-1, which is distantly related to Methanosarcinales and
Methanomicrobiales, occurs in association with SRB of the Desulfosarcina-
Desulfococcus (DSS) group from the �-proteobacteria (Michaelis et al., 2002; Knittel et
al., 2005), as monospecific aggregates or as single cells (Orphan et al., 2001; Knittel et
al., 2005). Both ANME-2 and ANME-3 belong to the order Methanosarcinales. ANME-
2 has been observed in physical association with DSS (Boetius et al., 2000; Knittel et al.,
2005), while ANME-3 has been found in syntrophic partnership with Desulfobulbus sp.
(DBB) (Niemann et al., 2006; Lösekann et al., 2007).
Physiological characteristics of AOM communities are based on a few in vitro
studies (Nauhaus et al., 2002 and 2005) and mesocosm experiments (Guirguis et al., 2003
and 2005). Based on in vitro experiments Nauhaus et al. (2005) reported that changes in
sulfate concentration, pH and salinity seem not to influence AOM activity, contrary to
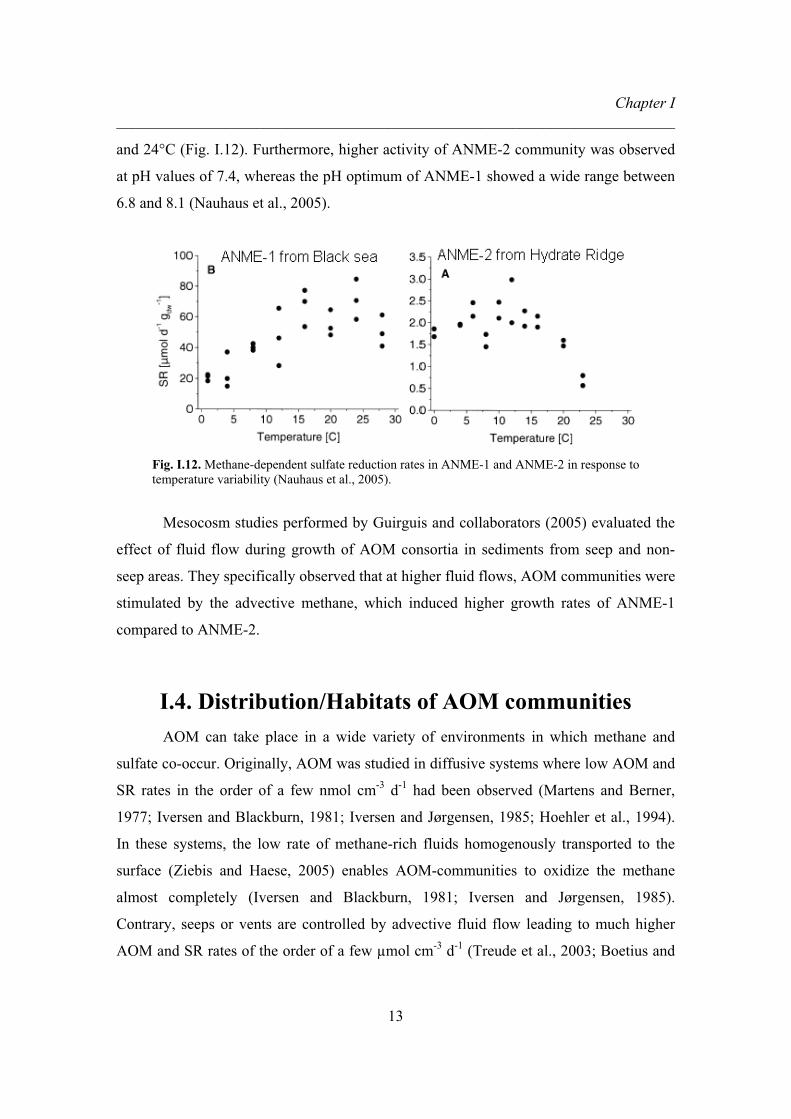
temperature. They concluded that ANME-2 is better adapted to cold temperatures than
ANME-1, which shows highest methane-dependent sulfate reduction rates between 16°C
Chapter I ________________________________________________________________________
13
and 24°C (Fig. I.12). Furthermore, higher activity of ANME-2 community was observed
at pH values of 7.4, whereas the pH optimum of ANME-1 showed a wide range between
6.8 and 8.1 (Nauhaus et al., 2005).
Fig. I.12. Methane-dependent sulfate reduction rates in ANME-1 and ANME-2 in response to temperature variability (Nauhaus et al., 2005).
Mesocosm studies performed by Guirguis and collaborators (2005) evaluated the
effect of fluid flow during growth of AOM consortia in sediments from seep and non-
seep areas. They specifically observed that at higher fluid flows, AOM communities were
stimulated by the advective methane, which induced higher growth rates of ANME-1
compared to ANME-2.
I.4. Distribution/Habitats of AOM communities AOM can take place in a wide variety of environments in which methane and
sulfate co-occur. Originally, AOM was studied in diffusive systems where low AOM and
SR rates in the order of a few nmol cm-3 d-1 had been observed (Martens and Berner,
1977; Iversen and Blackburn, 1981; Iversen and Jørgensen, 1985; Hoehler et al., 1994).
In these systems, the low rate of methane-rich fluids homogenously transported to the
surface (Ziebis and Haese, 2005) enables AOM-communities to oxidize the methane
almost completely (Iversen and Blackburn, 1981; Iversen and Jørgensen, 1985).
Contrary, seeps or vents are controlled by advective fluid flow leading to much higher
AOM and SR rates of the order of a few μmol cm-3 d-1 (Treude et al., 2003; Boetius and

Chapter I ________________________________________________________________________
14
Suess, 2004). AOM and SR rates are usually coupled in a 1:1 ratio (Hinrichs and Boetius,
2002; Nauhaus et al., 2002 and 2005). However, due to the fact that SR can as well be
fueled by other carbon substrates, a decoupling of both processes has been observed in
places where seepage of oil and higher hydrocarbon gases, such as ethane and propane,
are detected (e.g., Gulf of Mexico, Joye et al., 2004).
Methane-rich fluids in advective systems are transported along permeable
pathways (faults, cracks, scarps) induced by pressure gradients (Ziebis and Haese, 2005),
which result in varying fluid flow regimes. This affects the small scale heterogeneity of
seep communities which are dependent on hydrogen sulfide produced during AOM (Fig.
I.13).
Fig. I.13. Community distribution in relation to fluid flow in sediments from Hydrate Ridge (Sahling et al., 2002; Torres et al., 2002).
The input of methane, together with the sulfide rich fluids advected as a result of
AOM, is the basis for the abundant communities of organism living in seeps such as
sulfide oxidizing microbial communities and diverse benthic macrofauna with
methanotrophic symbionts (Sahling et al., 2002; Levin, 2005).
Cumulative molecular data provide evidence of a global distribution of AOM
communities (Fig. I.14). The occurrence of different AOM communities is observed in a
wide range of natural habitats, which are dominated by one of the consortia described
above. Hot spots of AOM communities are cold seep environments from globally-

Chapter I ________________________________________________________________________
15
distributed habitats including anoxic water bodies, mud volcanoes and oil fields, all of
which are often found in conjunction with methane gas hydrates. Moreover, AOM has
been observed at hydrothermal vent systems. A description of these environments is
provided below.
Fig. I.14. Global distribution of AOM communities based on fluorescence in situ hybridization (FISH) microscopy obtained during the projects MUMM I and II.
Cold seeps. Cold seeps are habitats where seepage of gases and methane-rich
fluids are transported by advective forces without a considerable increase in temperature.
In contrast to hydrothermal vents, the fluid rates and temperatures at hydrocarbon seeps
are dependent on the accumulation and burial of organic matter (Campbell, 2006). Since
the first report of cold seeps 20 years ago (Paull et al., 1984), several new cold seeps have
been found in passive (e.g., Suess et al., 1985, 1998; Yun et al., 1999) and active
continental margins (e.g., Paull et al., 1995). In this environment, the supply of methane
enables growth of diverse microbial communities such as methanotrophic archaea and
SRB.
Hydrothermal vents. Hydrothermal vents are observed at mid-ocean ridges,
where abiotic methane is produced by serpentinization of iron and manganese minerals
during the contact of basaltic material with sea water (Eq. 5a and b, Reeburgh et al.,
Black Sea
Haakon Mosby Mud Volcano
Eel River Basin
Hydrate Ridge
Wadden Sea
Eckernförder Bight
Congo Basin
Gulf of Mexico
Guaymas Basin

Chapter I ________________________________________________________________________
16
2007). Once the sulfide- and sulfate-rich vent fluids get in contact with the cold seawater
the precipitation of minerals produce the characteristic black smokers observed in
hydrothermal systems (Haymon, 1983). Characteristic features of hydrothermal vent
fluids are high temperatures (Lutz et al., 1994) and typically acidic pH values, although
higher pH values have also been reported (pH >10, von Damm et al., 1985). Due to the
presence of chemical and thermal energy produced in hydrothermal systems, this habitat
is a major focus of interest because it represents an analog for the origin of life.
� �� � � �� �)(magnetite e)(serpentin (olivine) HOFeOHOSiMg30H7SiOFeMg6 2434523245.05.1 � Eq. 5a
O2H4 2422 HCHCO � Eq.5b
Hydrothermal vent fluids sustain diverse communities including tube worms,
shrimps, clams and chemosynthetic microorganisms (Levin et al., 2005). Moreover,
AOM has also been reported in the Guaymas Basin hydrothermal field where ANME-1
and ANME-2 communities occur (Teske et al., 2002).
Anoxic water bodies. The largest anoxic marine basin is the Black Sea
(Reeburgh et al., 1991). Concentration of methane in the anoxic water column are in the
micromolar range (Reeburgh et al., 1991), which seems to facilitate the build-up of
chimney-like structures that harbors carbonate-rich microbial mats of AOM communities
(Michaelis et al., 2002; Treude et al., 2005). Both, lipid biomarkers strongly depleted in 13C and FISH data confirm the presence of ANME-1/DSS and AMME-2/DSS utilizing
methane as a carbon source (Michaelis et al., 2002; Blumenberg et al., 2004). Besides
these structures, the occurrence of pockmarks, mud volcanoes and gassy sediments is also
observed in the Black sea. Similarly, the occurrence of AOM in sediments and water
column of Cariaco Basin has been documented (Reeburgh, 1976; Ward et al., 1987),
although no evidence of chimney-like structures has been provided.
Mud volcanoes. Mud volcanoes are another important habitat, with high, but
episodic gas escape (Reeburgh et al., 2007). Most mud volcanoes are found as submarine
structures close to subduction zones and orogenic belts, in which high sedimentation rates
and the formation of hydrocarbons and fluids occur (Dimitrov et al., 2002; Milkov et al.,

Chapter I ________________________________________________________________________
17
2003). Methane release from these structures is estimated in the order of 13 Tg and 15 Tg
during inactive and eruptive periods, respectively (Milkov et al., 2003). At distinct mud
volcanoes, such as the Haakon Mosby Mud Volcano (HMMV), up to 40% of the released
methane is oxidized by aerobic and anaerobic methonotrophs (Niemann et al., 2006).
Distinctive from other seep environments is the dominance of ANME-3/DBB
communities at HMMV (Lösekann et al., 2007). A relative higher abundance of ANME-
3, although accompanied by other ANME groups, has been also reported at the mud
volcano from the Nile deep sea fan at the eastern Mediterranean Sea (Omoregie et al.,
2008).
Oil fields. Shallow and deep oil fields have been observed at Gullfaks and in the
Gulf of Mexico, respectively. Gullfaks is a big Norwegian oil and gas field located in the
northern North Sea at 140 m water depth (Hovland, 2007). This area is covered by sand,
which was deposited during the last glacial maximum (Hovland and Judd, 1988).
Microbial mats of sulfide oxidizing bacteria provide evidence of the occurrence of AOM
just a few centimeters below the seafloor, in which ANME-2a and -2c dominated
communities inhabit (Wegener et al., 2008). The northern Gulf of Mexico is a
hydrocarbon gas reservoir positioned over salt deposits of Jurassic age (Roberts et al.,
1999). The tectonic characteristics of this location produce conduits that allow the
transport of gas through seeps, brine pools and mud volcanoes, as well as the formation
of methane hydrates (Sassen et al., 1994). Large amounts of sulfide oxidizing bacteria,
inhabiting surface of sediments, together with a high abundance of ANME-1/DSS have
been observed at Gulf of Mexico seeps (Orcutt et al., 2005).
Gas hydrate environments. The occurrence of methane hydrates in cold seeps is
very well documented from several locations such as the Gulf of Mexico (Sassen et al.,
1994), the Eel River Basin (Kvenvolden and Field, 1981) and the Cascadia continental
margin (Suess et al., 1999). Among these locations, one of the most studied is Hydrate
Ridge, a geological feature discovered at the Cascadia Margin in the mid ‘80s (Suess et
al., 1985). Hydrate Ridge is characterized by high fluid flow and shallow deposits of gas
hydrates (Suess et al., 1999; Torres et al., 2002). In this habitat, the consortium of
ANMEs and SRB responsible of AOM was visually observed for the first time (Boetius
et al., 2000) in agreement with previous findings of huge amounts of AOM-derived

Chapter I ________________________________________________________________________
18
carbonate structures (Ritger t al., 1987) and 13C-depleted lipid biomarkers (Elvert et al.,
1999).
Besides the fact that AOM communities are widely distributed in various habitats
in which methane and sulfate co-occur, the dominance of single communities has been
reported. For example, ANME-1/DSS seems to dominate in subsurface sediments
(Knittel et al., 2005) and microbial mat structures (Michaelis et al., 2002), ANME-2/DSS
occurs in surface sediments related to methane hydrates (Knittel et al., 2005), and
ANME-3/DBB in mud volcanoes (Niemann et al., 2006, Lösekann et al., 2007). This
indicates that the selection of the respective groups depends on a yet unknown
environmental conditions found at the sites.
I.5. Lipid signatures of communities performing AOM The first description of a biomarker related to AOM came from the irregular tail-
to-tail isoprenoid crocetane (2,6,11,15-tetramethylhexadecane), which was observed in
the SMTZ of sediments in the Kattegat (Bian, 1994; Bian et al., 2001). Moreover,
crocetane was reported from recent and ancient cold seep environments associated with
marine gas hydrates (Elvert et al., 1999) and limestone formation (Peckmann et al., 1999;
Thiel et al., 1999), respectively. In all of these studies, crocetane was suggested to be a
biomarker of anaerobic methanotrophic archaea due to its structural characteristic and
strong depletion in 13C relative to the assimilated methane. Together with the occurrence
of crocetane in AOM environments, subsequent studies have provided a series of other
biomarkers characterized by very low �13C values as a consequence of methane
utilization. The first unambiguous evidence of archaea mediating AOM was the presence
of archaeol and sn-2-hydroxyarchaeol with �13C values < -100‰, which were found in
concert with ANME-1 sequences in methane rich sediments from the Eel River Basin
(Hinrichs et al., 1999). In a following study, Hinrichs et al. (2000) provided evidence for
not only archaeol and sn-2-hydroxyarchaeol as indicators of ANMEs but also bacterial-
derived fatty acids as well as straight-chain monoalkyl and dialkyl glycerol ethers
(MAGEs and DAGEs, respectively), which were less depleted in 13C compared to the
archaeal lipids. The presence of these non-isoprenoidal lipid biomarkers was attributed to

Chapter I ________________________________________________________________________
19
the SRB partners associated with the ANMEs (Hinrichs et al., 2000). The occurrence of
these and other biomarkers in various cold seep systems, including methane-hydrate
environments (Elvert et al., 1999, 2003 and 2005; Boetius et al., 2000), hydrothermal
vents (Teske et al., 2002), mud volcanoes (Pancost et al., 2000 and 2001; Niemann et al.,
2006), carbonate reefs (Thiel et al., 2001; Michaelis et al., 2002; Blumenberg et al., 2004)
and oil fields (Wegener et al., 2008), support the extensive distribution of these
communities performing AOM.
Several diagnostic biomarkers have been related to the dominance of the different
AOM communities in the marine environment. ANME-1 microbial mats from the Black
Sea were characterized by a high abundance of GDGT-derived biphytanes and higher
amounts of archaeol as opposed to hydroxyarchaeol (Fig. I.15A). In contrast, ANME-2
dominated mats were found to contain crocetane and crocetenes, and a higher abundance
of hydroxyarchaeol relative to archaeol (Fig. I.15B). Similar conclusions were drawn by
Elvert et al. (2005) who reported the diversity of biomarkers occurring in sediments from
Hydrate Ridge off the coast of Oregon. Biomarker patterns observed were specifically
related to different fluid flow regimes causing the development of distinct seep provinces,
namely Beggiatoa mats, Calyptogena fields and Acharax fields (Fig. I.13). Besides
archaeal biomarkers, high amounts of DSS-specific fatty acids (i.e., C16:1�5c and
cyC17:0�5,6) were detected at the Beggiatoa site (Fig. I.15C), where also high numbers of
ANME-2a/DSS aggregates were observed, whereas ANME-1 in deeper horizons of the
Calyptogena site showed higher contents of the fatty acid ai-C15:0 (Fig. I.15D). Generally,
sediments from the Calyptogena site were dominated by ANME-2c and characterized by
the additional occurrence of GDGTs containing 1 and 2 cyclopentyl rings, which have
been frequently detected in AOM environments (e.g., Pancost et al., 2001; Wakeham et
al., 2003). Carbon isotopic values of the biomarkers from ANME-2 were usually 20‰
more negative than the ones from ANME-1 dominated sediment horizons (Elvert et al.,
2005). This carbon isotopic difference between the two communities was previously
indicated in other studies (Hinrichs et al., 2000; Orphan et al., 2001; Blumenberg et al.,
2004).

Chapter I ________________________________________________________________________
20
Fig. I.15. Characteristic apolar lipids derived from ANME-1 and ANME-2 dominated chimney-like structures in the Black Sea (A and B, Blumenberg et al., 2004) and sediments underneath a Beggiatoa mat from Hydrate Ridge (C and D, Elvert et al., 2005).
The differentiation of ANME-3 from ANME-1 and -2 is less obvious and was
characterized by the sole presence of highly unsaturated 2,6,10,15,19-
pentamethylicosanes (PMI:4 and PMI:5) together with archaeol and hydroxyarchaeol, but
the absence of both crocetane and GDGTs (Niemann et al., 2006). The bacterial partner
of the Desulfobulbus group, however, was indicated by the high abundance of the
specific fatty acid C17:1�6c.

Chapter I ________________________________________________________________________
21
In summary, the occurrence of strongly 13C-depleted archaeal biomarkers in
AOM studies is accompanied by the presence of slightly 13C-enriched bacterial lipid
biomarkers. Among these bacterial lipids, the occurrence of complex fatty acids with 14-
18 carbon atoms, with and without double bonds, methyl-branches and cyclopropyl
isomers has been observed (Hinrichs et al., 2000; Elvert et al., 2003 and 2005). Also the
presence of MAGEs and DAGEs with similar patterns to the ones detected in the fatty
acids has been reported (Hinrichs et al., 2000; Elvert et al., 2005). However, all of these
previous biomarker studies targeted GC-amenable lipids, which are assumed to represent
only a minor fraction in living cells and may have only been found as a relict of deceased
microbial communities. To reduce the obstacles associated with apolar lipids, we
therefore targeted intact polar lipids (IPLs) which are the building blocks of the
cyctoplasmic membrane of all living cells and which can be directly related to
microbiological investigations using FISH or other techniques.
I.6. Intact polar membrane lipids (IPLs) The cytoplasmic cell membrane acts as a semi-permeable barrier and protects the
cell from the external environment. The membrane is composed of proteins and a lipid
bilayer (Fig. I.16).
Proteins can play different roles in the cell membrane such as recognizing
substrates, performing enzymatic activity and transporting substances (nutrients, ions and
waste) between the cytoplasm and the exterior of the cell (Madigan et al., 2003). On the
other hand, lipids are indispensable for the membrane structure due to their chemical
properties (hydrophobicity and hydrophilicity), which directly involve these molecules in
membrane permeability (Madigan et al., 2003). Because the cell membrane regulates the
transport between the exterior and interior of the cell, it is also important in the
conservation of cell energy (Madigan et al., 2003).
According with the fluid mosaic model, the cell membrane is composed of a
double layer or bilayer of lipids. The bilayer formed by phospholipids contains a fatty
acid tail (hydrophobic side) and a phosphate group in the polar part of the molecule
(hydrophilic side). The hydrophobic side is oriented inwards, while the hydrophilic side

Chapter I ________________________________________________________________________
22
or head group is facing outwards (i.e. the aqueous cytosol of the cell or the environment)
(Fig. I.16).
Fig. I.16. The phospholipid membrane bilayer (Tortora et al., 2004).
Lipids in the cell membrane of prokaryotes are represented by phospholipids,
glycolipids and sometimes hopanoids (e.g., in methanotrophic bacteria, Madigan et al.,
2003). In total, they represent up to 6% of the cell dry weight (Langworthy et al., 1983).
Membrane lipids are good candidates to distinguish Bacteria and Archaea. Bacteria
generally contain a phospholipid bilayer composed of fatty acids linked to a glycerol
backbone via ester bonds in sn-1 and sn-2 position (ester-bond acyl chains, Fig. I.17). In
sulfate reducers, these fatty acids may include methyl branching, double bonds and
cyclopropyl isomers (Taylor and Parkes, 1983; Dowling et al., 1986). Archaeal
membranes can occur both as a bilayer or monolayer (Fig. I.17). The bilayer of archaeal
cells contains isoprenoidal chains linked to the glycerol backbone in sn-2 and sn-3
position via an ether bond (i. e., isoprenoidal alkyl chains) and is generally formed by two
C20 hydrocarbon chains (phytanyl ethers) (Langworthy and Pond, 1986). Archaeal
monolayer membranes are composed of glycerol tetraethers, in which two glycerol
molecules are linked via two C40 hydrocarbon chains (biphytanyl ethers) (Langworthy

Chapter I ________________________________________________________________________
23
and Pond, 1986). Generally, ether bonds from archaeal membranes are more resistant to
higher temperature, pressure and pH (De Rosa et al., 1989) than the ester bonds present in
bacteria.
Fig. I.17. General features of archaeal and bacterial lipid membranes (Valentine, 2007).
Because the cell membrane is affected by external conditions such as temperature,
pH, pressure or salinity, several adaptations in prokaryotic cell membranes are related to
cell evolution, physiology, biogeochemistry and ecology (Langworthy, 1982). Among
these adaptations, changes in fatty acid compositions have been observed depending of
the habitat temperatures. In contrast to shorter saturated and unsaturated fatty acids in
psychrophilic bacteria, evidence of longer and saturated fatty acids, predominantly iso-
branched, is found in thermophilic bacteria (Langworthy, 1982). Additionally, the effects
of pH and temperature in a thermoacidophile were evaluated (De Rosa et al., 1974). At
lower pH and increasing temperature, the proportion of iso- and anteiso-fatty acids

Chapter I ________________________________________________________________________
24
increases, whereas at higher pH and increasing temperature cyclohexyl fatty acids
increase (De Rosa et al., 1974). Furthermore, the effect of temperature on polar head
group compositions of a thermophilic organism (i.e., Bacillus caldotenax) has been
investigated by Hasegawa et al. (1980). These authors reported a decrease in the amount
of PE (from 57% to 37%) and increase of PG (from 27% to 46%) in the total
phospholipid content induced by a temperature decrease from 65°C to 45°C.
Modifications observed in the hydrocarbon chains of archaeal-based tetraether
lipids include the increase in membrane stability at higher growth temperatures by the
formation of cyclopentane rings (Langworthy and Pond, 1986).
All the modifications in the membrane described above intent to protect the cell
from the environment. In general, archaeal membranes are less permeable, thus they may
be better adapted to hostile environments than bacterial ones (Valentine, 2007). Due to
this characteristic of Archaea, these microorganisms were assumed to live in extreme
environments in which low pH and high temperatures occur (Rothschild and Mancinelli,
2001). However, cumulative evidence shows that Archaea are not only prevalent in the
deep biosphere (Biddle et al., 2006; Lipp et al., 2008), hydrothermal vents (Teske et al.,
2002; Reysenbach et al., 2000; Schouten et al., 2003) and cold seeps (Boetius et al.,
2000; Knittel et al., 2005), but are also widely distributed in ocean waters (Karner et al.,
2001; DeLong, 2003).
The investigation on the diversity of intact polar membrane lipids (IPLs) from
both Bacteria and Archaea was extended by the utilization of high-performance liquid
chromatography mass spectrometry (HPLC-MS). Contrary to the other techniques (e.g.,
gas chromatography), the advantage of HPLC-MS is the possibility to study the intact
membrane lipid molecules instead of core or side chain products. During the analysis, the
chromatographic separation of IPLs is based on their polarity, which is mainly related to
the molecule’s head groups (Fig. I. 18).

Chapter I ________________________________________________________________________
25
Fig. I.18. HPLC-MS chromatogram (A) and density map (B) of an IPL mixture of commercially available standards mixed with an extract of microbial mat from the Black Sea. IPLs elution depends on their polarity, with less polar compound eluting at early retention times. Density map is a representation of the IPL peaks in relation to the retention time and the mass to charge ratio (range scanned from 500 to 2000 m/z). In it, the intensity of the black lines is correlated to the concentration of the IPL in the sample mixture. Bacterial-derived IPLs (PE, PG and PDME) in the density map are displayed in series due to the presence of different fatty acid chain lengths. Abbreviations of IPLs according to Fig. I.19
Diversity of polar head groups in IPLs has been described from cultures and
environmental samples based on HPLC-ESI-MS (Fig. I.19A), providing taxonomic
information that allows the distinction of different microorganisms (e.g., De Rosa et al.,
1986; Koga et al., 1998; Sturt et al., 2004; Koga and Morii, 2005; Van Mooy et al., 2006;
Koga and Nakano, 2008). HPLC-ESI-MS is equipped with an electrospray ionization
source (ESI) that produces a soft ionization of the analytes, which is particularly
appropriate for polar molecules like IPLs. Using this technique, the diversity of IPLs
characteristic of archaea from marine systems has been reported, including archaeol- and
GDGT-based IPLs with glycosidic head groups (Fig. I.19B, Sturt et al., 2004; Biddle et
al., 2006; Lipp et al., 2008). Furthermore, a variety of phospholipids from Bacteria has
been documented, including ether and ester phospholipids (Fig. I.19C) with diverse types
of head groups (Rütters et al., 2002; Sturt et al., 2004; Van Mooy et al., 2006).

Chapter I ________________________________________________________________________
26
OOPOH
O OO
OOPOH
O OO
OOPOH
O OO
R'
R''
R''
R''O
R'O
R'O
Diacylglycerophospholipid DAG
Acyl/ether glycerophospholipid AEG
Dietherglycerophospholipid DEG
OO
O
X=H, Diglycosyl archaeolX=OH, Diglycosyl hydroxyarchaeol
X
O
OO O
OHO
Diglycosyl glyceroldialkylglyceroltetraether GDGT with 0 cyclopentyl rings
OHOOH O
POH
O
OHONH2
O
Phosphatidylserine PS
Phosphatidylglycerol PG
OPOH
O
OOHHO
HOHO OH
Phosphatidylethanolamine PE
OPOH
OON
Phosphatidylcholine PC Phosphatidylinositol PI
OPO
O
OH2N
OPOH
O
ON
OPOH
O
ONH
OPOH
O
Phosphatidyl-(N)-methylethanolamine PME
Phosphatidyl-(N,N)-dimethylethanolamine PDME
O
HO OHHOO
OHOHO
HO OH
O
HO OHHOO
OHOHO
HO OH
OHO
HO OH
O OHO
HOHO
HO OH
OO O
OHO
Diglycosyl glyceroldialkylnonitoltetraether GDNT with 0 cyclopentyl rings
HOOPOH
O
Phosphatiddic acid PA
OOPOH
OHO
OH
NH2OH
Phosphoaminopentatetrol APT
OOPOH
OH2N
O
OHHOHO O
OPOH
OH
Glyco-phosphoethanolamine GPE
HOO
OHHOHO O O
O
O
O
n=1 Monogalactosyldiacylglycerol MGDG n=2 Digalactosyldiacylglycerol DGDG
n
HO3SO
OHHOHO O O
O
O
O
Sulfoquinovosyldiacylglycerol SQDG
A C
B
Fig. I.19. Diversity of IPL-head groups present in Bacteria and Archaea (A), glycolipids commonly observed in Archaea (B), and ester and ether linkages observed in phospholipids (C).
Structural information of IPLs can be obtained by ion-trap mass spectrometry (IT-
MS) configured to trap ions of interest which are later fragmented producing daughter ion
mass spectra (MSn). Identification of IPLs is based on fragmentation patterns obtained
from MSn experiments in positive and negative modes, and by comparison with
previously reported mass spectral data (Table I.1) (Sturt et al., 2004) and molecular
structures (Koga and Nakano, 2008 and references therein). Most of the structural
characteristics of IPLs can be obtained in MS2 (Fig. I.20). However, additional
information is obtained by analyzing the sample under positive and negative ionization
modes. IPLs positively ionized frequently loose the head groups providing information of
the lipid class (Fig. I.20A), whereas IPLs negatively ionized loose the fatty acid chain
located in the sn-2 position (Fig. I.20B). Structural information of diverse IPLs from
Archaea and Bacteria observed in this study are provided in the Chapter V of this work.

Chapter I ________________________________________________________________________
27
Positive ion mode [M +H]+ Negative ion mode [M -H]- Headgroup AEG, DAG DEG AEG, DAG DEG
PE 141 Da loss (phosphoethanolamine)
43 Da loss (ethanolamine) 43 Da loss
(ethanolamine)
APT 231 Da loss (phospho-APT) 133 Da loss (APT)
AEG-P; loss of sn-2 fatty
acid 133 Da loss (APT)
PG 189 Da loss
(phosphoglycerol + NH4
+ adduct) 75 Da loss (glycerol)
DAG-P; loss of head
group+ sn-2 fatty acid
75 Da loss (glycerol)
PI 162 Da loss hexose Major ion m/z 241 (phosphoglycosyl –
H2O)
PS 185 Da loss (phosphoserine) 87 Da loss (serine) 87 Da loss (serine)
PC All give a major ion m/z 184 (phosphocholine) All show 60 Da loss (CH3+ HCOO- adduct)
Table I.1. Characteristic headgroup losses of common phospholipids under HPLC-ESI-MS conditions in positive and negative ion modes (Sturt et al., 2004).
Fig. I.20. Mass spectra of phosphatylethanolamine (PE) diacylglycerol (DAG). Difference of mass between the positive (A) and negative ion mode (B) are explained by the addition and lost of one proton in the molecule, respectively. MS2 data in positive ion mode indicate the lost of 141 Da (PE) from the glycerol and fatty acid core with C31:2 (sum of both fatty acids). Negative ion mode indicates the lost of C15:2 from sn-2 position of the glycerol first (lyso fragment 434 Da) and the presence of the fatty acid C16:0 in the sn-1 position of the glycerol (fragment 255 Da).

Chapter I ________________________________________________________________________
28
I.7. Methods Most samples analyzed in this study were freeze-dried and extracted according to
a modified Bligh and Dyer protocol (Sturt et al., 2004) by microwave-assisted extraction
system (MARS-X, CEM, USA) for 15 min at a temperature of 70°C, while a few others
were extracted by ultrasonication. A mixture of standards covering different lipid classes
was added to the samples. The standards included cholestane (hydrocarbons), behenic
acid methyl ester (ketones), C-19 alcohol (alcohols) and C19-fatty acid (fatty acids) for
GC-amenable lipids, and C16-PAF for IPL analysis. The solvent mixture used during the
extractions was methanol:dichloromethane:buffer in a proportion of 2:1:0.8. The volume
of the solvent mixture used was 40 mL per every 10 g of dry sediment and 1 g of dry mat.
The first two extraction steps were performed with phosphate buffer, whereas the last two
were performed with trichloroacetic acid buffer (TCA). After collection of all
supernatants, the organic phase was separated from the aqueous one by multiple additions
of dichloromethane (DCM) and milli-Q water. This liquid-liquid extraction was
performed by using the same amount of water and DCM than the total solvent mixture
added during the extractions, starting with DCM (3 times) and then with water (3 times).
The organic phase or total lipid extract (TLE) was evaporated to dryness under a stream
of nitrogen and re-dissolved in a mixture of DCM:methanol (1:1), which was finally
injected into the HPLC-ESI-MS.
Due to the nature of the sample (e.g., oily etc.), additional clean-up steps were
performed on Eel river Basin, Guaymas Basin and two sediment samples from Gulf of
Mexico. Here, separation of the TLE into apolar, glyco- and phospholipids was carried
out on activated silica column (2 g of silica for 50-200 mg of extract) by elution with 20
mL of DCM, 40 mL of acetone and 40 mL of methanol, respectively. Acetone and
methanol eluted fractions were combined and evaporated under a nitrogen stream and re-
dissolved in DCM:methanol (1:1) prior to analysis. This procedure allows the detection
of IPLs previously not observed in the TLE probably due to matrix problems and ion
suppression. It is well documented that ESI signal can be affected by the sample matrix,
which, if contain endogenous material (in this case hydrocarbons), could interfere in the
ionization of the analytes of interest (Mallet et al., 2004). This problem can be solved to
some degree by additional clean-up steps (Mallet et al., 2004).

Chapter I ________________________________________________________________________
29
Parallel analyses of apolar lipid biomarkers were performed in order to compare
both intact (IPLs) and non-intact lipids (GC-amenable lipids). For the analysis of apolar
lipids, a fraction of the TLE was added to a Pasteur pipette with glass wool and separated
into maltene and asphaltene fraction, eluting the first of them with 2.5 mL hexane and the
second with 4 mL of DCM. The maltene fraction was further separated into four fractions
of increasing polarity on Supelco LC-NH2 glass cartridges (500 mg sorbent) using 4 mL
of hexane (hydrocarbons), 6 mL hexane/DCM (3:1; ketones/esters), 7 mL DCM/acetone
(9:1; alcohols) and 8 mL of 2% formic acid in DCM (free fatty acids). Each fraction was
evaporated to dryness under a stream of nitrogen and re-dissolved in hexane prior to
analysis. Previously alcohols were derivatized into trimethylsilylesters (TMS-derivatives)
by addition of N,O-bis(trimethylsilyl) fluoracetamide (BSTFA) and pyridine. Similarly,
fatty acids were transformed to methylesters (FAME) before analysis, using 20% Boron
trifluoride (BF3) in methanol. Both reactions were performed at 70°C for 1h. All
fractions were analyzed via gas chromatography-mass spectrometry (GC-MS) and GC-
flame ionization detection (GC-FID). Identification of GC-amenable lipids was based on
the comparison of retention times, mass spectra of commercial standards and from
literature.
I.8. Hypothesis and objectives The aim of this PhD work is the elucidation of the microbial community
structures in different marine methane-rich environments based on the diversity of lipid
signatures. This work is part of the MUMM II (Methane in the Geo/bio-System-
Turnover, Metabolism and Microbes) project, a multidisciplinary BMBF project which
started in a first phase already in January 2001.
AOM is, based on current knowledge, associated with the presence of three
phylogenetic clusters of methanotrophic archaea (ANME) and two groups of SRB (DSS
and DBB) in various marine environments (gas hydrate, mud volcanoes, hydrothermal
sediments and coastal subsurface environments). Different biogeographical patterns of
these clusters are probably related to varying environmental conditions found in a wide
range of settings (e.g., Arabian Sea, Black Sea, Eastern Mediterranean Sea, Eel River

Chapter I ________________________________________________________________________
30
Basin, Guaymas Basin, Gulf of Mexico, Gullfaks oil field, Häkon Mosby Mud Vulcano
and Hydrate Ridge). In order to evaluate the global distribution of these AOM
communities, this study reviews the diversity of lipids, known so far from the analysis of
characteristic apolar lipids, and extends the knowledge to intact polar membrane lipids
that provide valuable information of both Archaea and Bacteria. Additionally, the
combination of lipid biomarkers, available microbiological data, together with the
environmental characterization of each setting should improve our understanding of the
distribution of AOM communities and the factors controlling them.
Specifically, the present study addresses the following questions regarding AOM:
� What is the diversity of IPLs present in AOM environments?
� Is it possible, using IPL diversity to distinguish between ANME-1/-2/-3,
their SRB partners?
� Is it possible to assign the dominant ANME group in a sample without
molecular information?
� Is the IPL composition of an AOM community inhabiting carbonate
chimneys the same as the one found in the corresponding AOM
community in sediments?
� Is it possible to identify the most important environmental variables that
define the ecological niches of AOM communities?
� Do classical apolar lipid biomarkers provide the same information as
IPLs?
� What is the relation between IPLs and apolar lipids?
I.9. Contribution to publications
This thesis includes the complete version of two manuscripts. Chapter II is a
published manuscript and Chapter III is a manuscript version close to submission.
Chapter IV is a draft of a degradation experiment and Chapter V is a draft in which a
deeper insight into the diversity of intact polar membrane lipids observed during this
study is provided.

Chapter I ________________________________________________________________________
31
CHAPTER II - full manuscript Intact polar lipids of anaerobic methanotrophic archaea and associated bacteria
Pamela E. Rossel, Julius S. Lipp, Helen F. Fredricks, Julia Arnds, Antje Boetius, Marcus
Elvert, Kai-Uwe Hinrichs
Pamela E. Rossel extracted membrane lipids from three samples and identified
diverse archaeal and bacterial lipids with the support of Helen F. Fredricks and Julius S.
Lipp. Helen F. Fredricks extract membrane lipids from Hydrate Ridge sediments. Julia
Arnds and Antje Boetius provided phylogenetic data of the two microbial mats from the
Black Sea. Pamela Rossel, Marcus Elvert and Kai-Uwe Hinrichs wrote the paper jointly
with editorial input from all co-authors.
Published in Organic Geochemistry vol. 39, page 992-999,
doi:10.1016/j.orggeoche.2008.02.021.
CHAPTER III - full manuscript Factors controlling the distribution of anaerobic methanotrophic communities in
marine environments: evidence from intact polar membrane lipids
Pamela E. Rossel, Marcus Elvert, Alban Ramette, Antje Boetius and Kai-Uwe Hinrichs
Pamela E. Rossel extracted sediment and microbial mat samples, identified
diverse archaeal and bacterial polar and apolar lipids and compiled diverse environmental
data from literature to characterize the environments analyzed. Alban Ramette provided
expertise in multivariate analyses. Antje Boetius provided phylogenetic data and supplied
several samples analyzed in this study. Pamela Rossel, Marcus Elvert and Kai-Uwe
Hinrichs wrote the paper jointly with editorial input from all co-authors.
The manuscript is prepared for submission.

Chapter I ________________________________________________________________________
32
CHAPTER IV - draft Experimental approach to evaluate stability and reactivity of intact polar
membrane lipids of archaea and bacteria in marine sediments
Pamela E. Rossel, Julius S. Lipp, Verena Heuer and Kai-Uwe Hinrichs
Pamela E. Rossel prepared the experiment, extracted sediment samples and
quantified both archaeal and bacterial membrane lipids and
glyceroldialkylglyceroltetraether cores over the time of the experiment. Verena Heuer
performed acetate analysis. Julius S. Lipp gave support in the lab and with the membrane
lipids quantification. Pamela Rossel and all co-authors participated in the experimental
design. Unfortunately, due to several uncertainties in the results of this work, a new
experiment is indispensable, in which several problems related to the actual experimental
design should be overcome. Therefore this draft is just a guideline for further
experiments.
CHAPTER V - draft Diversity of intact polar membrane lipids in marine seep environments
Pamela E. Rossel, Marcus Elvert and Kai-Uwe Hinrichs
Pamela E. Rossel extracted sediment and microbial samples, identified diverse
archaeal and bacterial polar lipids and provided the molecular structures identified in seep
environments based on the mass spectral interpretation. All co-authors provided expertise
on lipid identification. Pamela Rossel wrote the paper jointly with editorial input from all
co-authors.

Chapter I ________________________________________________________________________
33
I.10. References Aloisi, G., Pierre, C., Rouchy, M. J., Foucher, J. P., Woodside, J., MEDINAUT scientific
party., 2000. Methane-related authigenic carbonates of eastern Mediterranean Sea
mud volcanoes and their possible relation to gas hydrate destabilization. Earth and
Planetary Science Letters 184, 321-338.
Barnes, R., Goldberg, E., 1976. Methane production and consumption in anoxic marine
sediments. Geology 4, 297-300.
Bian, L., Hinrichs, K. -U., Xie, T., Brassell, S. S., Iversen, N., Fossing, H., Jørgensen, B.
B., Hayes, J. M., 2001. Algal and archaeal polyisoprenoids in a recent marine
sediment: molecular isotopic evidence for anaerobic oxidation of methane.
Geochemistry Geophysics Geosystems 2, 2000GC000112.
Bian, L., 1994. Isotopic biogeochemistry of individual compounds in a modern coastal
marine sediment (kattegat, Denmark and Sweden). MSc thesis, Indiana University.
Biddle, J. F., Lipp, J. S., Lever, M. A., Lloyd, K. G., Sörensen, K. B., Anderson, R.,
Fredricks, H. F., Elvert, M., Kelly, T. J., Schrag, D. P., Sogin, M. L., Brenchley, J.
E., Teske, A. House, C. H., Hinrichs, K. -U., 2006. Heterotrophic archaea dominate
sedimentary subsurface ecosystems off Peru. Proceedings of the National Academy
of Science U.S.A. 103, 3846-3851.
Birgel, D., Himmler, T., Freiwald, A., Peckmann, J., 2008. Anew constrain on the
antiquity of anaerobic oxidation of methane: Late Pennsylvanian seep limestones
from southern Namibia. Geology 36, 543-546.
Blotevogel, K. -H., Fisher, U., 1985. Isolation and characterization of a new thermophilic
and autotrophic methane producing bacterium: Methanobacterium
thermoaggregans spec.nov. Archives of Microbiology 142, 218-222.
Blumenberg, M., Seifert, R., Reitner, J., Pape, T., Michaelis, W., 2004. Membrane lipid
patterns typify distinct anaerobic methanotrophic consortia. Proceedings of the
National Academy of Science U.S.A. 101, 11111-11116.
Boetius, A., Suess, E., 2004. Hydrate Ridge: a natural laboratory for the study of
microbial life fueled by methane from near-surface gas hydrates. Chemical Geology
205: 291-310.

Chapter I ________________________________________________________________________
34
Boetius, A., Ravenschlag, K., Schubert, C. J., Rickert, D., Widdel, F., Gieseke, A.,
Amann, R., Jørgensen, B. B., Witte, U., Pfannkuche, O., 2000. A marine microbial
consortium apparently mediating anaerobic oxidation of methane. Nature 407, 623-
626.
Buffett, B. A., 2000. Clathrate Hydrates. Annual Reviews of Earth and Planetary Science.
28, 477-507.
Campbell, K. A., 2006. Hydrocarbon seep and hydrothermal vent paleoenvironments and
paleontology: past developments and future research directions. Palaeogeography,
Palaeoclimatology, Palaeoecology 232, 362-407.
De Rosa, M., Gambacorta, A., Bu’Lock, J. D., 1974. Effects of pH and temperature on
the fatty acid composition of Bacillus acidocaldarius. Journal Bacteriology 117,
212-214.
De Rosa, M., Gambacorta, A., Gliozzi, A., 1986. Structure, biosynthesis, and
physicochemical Properties of Archaeabacterial Lipids. Microbiological Reviews
50, 70-80.
De Rosa, M., Gambacorta, A., Huber, R., Lanzotti, V., Nicolaus, B., Stetter, K. O.,
Trincone, A., 1989. Lipid structures in Thermophaga maritima. In: Microbiology of
Extreme Environments and its Potential for Biotechnology (eds. da Costa, M. S.,
Duarte, J. C., Williams, R. A. D.), 167-173, Elsevier Applied Science, London and
New York.
DeLong, E., 2003. Oceans of Archaea. American Society for Microbiology News 69, 503-
511.
Devol, A. H., 1983. Methane oxidation rates in the anaerobic sediments of Saanich Inlet.
Limnology and Oceanography 28, 738-742.
Dickens, G., 2004. Hydrocarbon-driven warming. Nature 429, 513-515.
Dimitrov, L. I., 2002. Mud volcanoes - the most important pathway for degassing deeply
buried sediments. Earth-Science Reviews 59, 49-76.
Dowling, N. J. E., Widdel, F., White, D. C., 1986. Phospholipid ester-linked fatty acid
biomarkers of acetate-oxidizing sulphate-reducers and other sulphide-forming
bacteria. Journal of General Microbiology 129, 3303-3309.

Chapter I ________________________________________________________________________
35
Duan, Z., Möller, N., Greenberg, J., Weare, J. H., 1992. The prediction of methane
solubility in natural waters to high ionic strengths from 0° to 250°C and from 0 to
1600 bar. Geochimica et Cosmochimica Acta 56, 1451-1460.
Elvert, M., Boetius, A., Knittel, K., Jørgensen, B. B., 2003. Characterization of specific
membrane fatty acids as chemotaxonomic markers for sulfate-reducing bacteria
involved in anaerobic oxidation of methane. Geomicrobiology Journal 20, 403-419.
Elvert, M., Hopmans, E. C., Treude, T., Boetius, A., Suess E., 2005. Spatial variations of
methanotrophic consortia at cold methane seeps: implications from a high-
resolution molecular and isotopic approach. Geobiology 3, 195–209.
Elvert, M., Suess E., Whiticar, M. J., 1999. Anaerobic methane oxidation associated with
marine gas hydrates: superlight C-isotopes from saturated and unsaturated C20 and
C25 irregular isoprenoids. Naturwissenschaften 86, 295–300.
Ferguson, T. J., Mah, R. A., 1983. Effect of H2-CO2 on Methanogenesis from acetate or
Methanol in Methanosarcina spp. Applied and Environmental Microbiology 46,
348-355.
Guirguis, P. R., Cozen, A. E., DeLong, E. F., 2005. Growth and population dynamics of
anaerobic methane oxidizing archaea and sulfate.reducing bacteria in a continuous-
flow bioreactor. Applied and environmental microbiology 71, 3725-3733.
Guirguis, P. R., Orphan, V. J., Hallam, S. J., DeLong, E. F., 2003. Growth and methane
oxidation o anaerobic methanotrophic archaea in a continuous-flow bioreactor.
Applied and environmental microbiology 69, 5472-5482.
Hanson, R. S., Hanson, T. E., 1996. Methanotrophic bacteria. Microbiological Reviews
60, 439-471.
Harvey, R. H., Fallon, R. D., Patton, J. S., 1986. The effect of organic matter and oxygen
on the degradation of bacterial membrane lipids in marine sediments. Geochimica
et Cosmochimica Acta 50, 795-804.
Hasegawa, Y., Kawada, N., Nosoh, Y., 1980. Change in Chemicals composition of
membrane of Bacilus caldotenax after shifting the growth temperature. Archives of
Microbiology 126, 103-108.
Haymon R. M., 1983. Growth history of hydrothermal black smoker chimneys. Nature
301, 695-698.

Chapter I ________________________________________________________________________
36
Hedderich, R., Whitman, W. B., 2006. Physiology and biochemistry of the methane-
producing archaea. Prokaryotes 2, 1050-1079.
Hinrichs, K. -U, Summons, R. E, Orphan, V., Sylva, S. P., Hayes, J. M., 2000. Molecular
and isotopic analyses of anaerobic methane-oxidizing communities in marine
sediments. Organic Geochemistry 31,1685-1701.
Hinrichs, K. -U., Boetius, A., 2002. The anaerobic oxidation of methane: New insights in
microbial ecology and biogeochemistry. In: Ocean Margin Systems (eds. Wefer G.,
Billett D., Hebbeln D., Jørgensen B. B., Schlueter M., van Weering T. C. E.), 457-
477. Springer-Verlag.
Hinrichs, K. -U., Hayes, J. S., Sylva, S. P., Brewer, P. G., DeLong, E. F., 1999. Methane-
consuming archaebacteria in marine sediments. Nature 398, 802-805.
Hoehler, T. M., Alperin, M. J., Albert, D. B., Martens, C. S., 1994. Field and laboratory
studies of methane oxidation in an anoxic marine sediment - evidence for a
methanogen-sulfate reducer consortium. Global Biogeochemical Cycles 8, 451-463.
Hovland, H., Judd, A. G., 1988. Seabed pockmarks and seepages: impact in geology,
biology and marine environment. Graham and Trotman, London, 1-293.
Hovland. M., 2007. Discovery of prolific natural methane at Gullfaks, northern North
Sea. Geo-Marine Letters 27, 197-201.
Iversen, N., Blackburn, H. T., 1981. Seasonal rates of methane oxidation in anaerobic
marine sediments. Applied and Environmental Microbiology 41, 1295-1300.
Iversen, N., Jørgensen, B. B., 1985. Anaerobic methane oxidation rates at the sulfate-
methane transition in marine sediments from Kattegat and Skagerrak (Denmark).
Limnology and Oceanography 30, 944-955.
Jones, W. J., Nagle Jr, D. P., Whitman, W. B., 1987. Methanogens and the diversity of
archaebacteria. Microbial Reviews 51, 135-177.
Jørgensen, B., 2001. Bacteria and marine biogeochemistry. In: Marine Geochemistry
(eds. Schulz H. D., Zabel M.), 169-206, Springer Verlag.
Joye, S. B., Orcutt, B. N., Boetius, A., Montoya, J. P., Schulz, H., Erickson, M. J., Lugo,
S. K., 2004. The anaerobic oxidation of methane and sulfate reduction in sediments
from at Gulf of Mexico cold seeps. Chemical Geology 205, 219-238.

Chapter I ________________________________________________________________________
37
Karner. M. B., DeLong, E. F., Karl, D. M., 2001. Archaeal dominance in the mesopelagic
zone of the Pacific Ocean. Nature 409, 507-510.
Kasting, J. K., 2004. When methane made climate. In: Scientific American, 78-85.
Kennett, J. P., Cannariato, K. G., Hendy, I. L., Behl, R. J., 2002. Methane Hydrates in
Quaternary Climate Change: The Clathrate Gun Hypothesis. American Geophysical
Union, Washington, USA.
Knittel, K., Lösekann, T., Boetius, A., Kort, R., Amann, R., 2005. Diversity and
Distribution of Methanotrophic Archaea at Cold Seeps. Applied and Environmental
Microbiology 71, 467-479.
Koga, Y., Morii, H., 2005. Recent Advances in Structural Research on Ether Lipids from
Archaea Including Comparative and Physiological Aspects. Bioscience
Biotechnology Biochemistry 69, 2019-2034.
Koga, Y., Morii, H., Akagawa-Matsushita, M., Ohga, M., 1998. Correlation of polar lipid
composition with 16S rRNA phylogeny in methanogens. Further analysis of lipid
component parts. Bioscience Biotechnology Biochemistry 62, 230-236.
Koga, Y., Nakano, M., 2008. A dendrogram of archaea based on lipid component parts
composition and its relationship to rRNA phylogeny. Systematic and Applied
Microbiology 31, 169-182.
Kvenvolden, K. A., 1998. A primer on the geological occurrence of gas hydrate. In: Gas
hydrates: Relevance to World Margin Stability and Climate Change 137 (eds.
Henriet, J. P., Mienert, J.), 9-30. Especial Publications Geological society, London.
Kvenvolden, K. A., Field, M. E., 1981. Thermogenic hydrocarbons in unconsolidated
sediment of Eel River, offshore northern California. American Association of
Petroleum Geologist Bulletin 65, 1642-1646.
Langworthy, T. A., 1982. Lipids of bacteria in extreme environments. In: Current Topics
in Membranes and Transport, Membrane lipids of prokaryotes 17 (eds. Razin, S.,
Rottem, S.), 45-77. Academic press.
Langworthy, T. A., Holzer, G., Zeikus, J., Tornabene, T. G., 1983. Iso- and anteiso-
branched glycerol diethers of the thermophilic anaerobe Thermodesulfotobacterium
commune. Systematic and Applied Microbiology 4, 1-17.

Chapter I ________________________________________________________________________
38
Langworthy, T. A., Pond, J. L., 1986. Archaeal ether lipids and chemotaxonomy.
Systematic and Applied Microbiology 7, 253-257.
Lelieveld, J., Crutzen, P. J., Dentener, F. J., 1998. Changing concentration, lifetime and
climate forcing of atmospheric methane. Tellus 50B, 128-150.
Levin, L., 2005. Ecology of cold seep sediments: interactions of fauna with fluid flow,
chemistry and microbes. In: Oceanography and Marine Biology: An annual Review
43 (eds. Gibson, R. N., Atkinson, R. J. A., Gordon, J. D. M), 1-46. Taylor and
Francis.
Lipp, J. S., Morono, Y, Inagaki, F., Hinrichs, K. -U., 2008. Significant contribution of
Archaea to the extant biomass in marine subsurface sediments. Nature 454, 991-
994.
Lösekann, T., Knittel, K., Nadalig, T., Fuchs, B., Niemann, H., Boetius, A., Amann, R.,
2007. Diversity and abundance of aerobic and anaerobic methane oxidizers at the
Haakon Mosby Mud Volcano, Barents Sea. Applied and Environmental
Microbiology 73, 3348–3362.
Lutz, R. A., Kennis, M. J., Pooley, A. S., Fritz, L. W., 1994. Calcium carbonate
dissolution rates in Hydrothermal vent fields of the Guaymas Basin. Journal of
Marine Research 52, 969-982.
Madigan, M. T., Martinko, J. M., Parker, J., 2003. Brock - Biology of Microorganisms.
10th Edition, Pearson education Inc. Upper Saddle River, New Jersey.
Mallet, C. R., Lu, Z., Mazzeo, J. R., 2004. A study of ion suppression effects in
electrospray ionization from mobile phase additives and solid phase extracts. Rapid
Communications in Mass Spectrometry 18, 49-58.
Martens, C. S., Berner, R. A., 1974. Methane production in the interstitial waters of
sulfate-depleted marine sediments. Science 185, 1167-1169.
Martens, C. S., Berner, R. A., 1977. Interstitial water chemistry of Long Island Sound
sediments, 1: Dissolved gases. Limnology and Oceanography 22, 10-25.
Michaelis, W., Seifert, R., Nauhaus, K., Treude, T., Thiel, V., Blumenberg, M., Knittel,
K., Gieseke A., Peterknecht F., Pape T., Boetius A., Amann R., Jørgensen B. B.,
Widdel, F., Peckmann, J., Pimenkov, N., Gulin, M. B., 2002. Microbial reefs in the
Black Sea fueled by anaerobic methane oxidation. Science 297, 1013-1015.

Chapter I ________________________________________________________________________
39
Milkov, A. V., Sassen, R., Apanasovich, T. V., Dadashev, F. G., 2003. Global gas flux
from mud volcanoes: A significant source of fossil methane in the atmosphere and
the ocean. Geophysical Research Letters 30, doi:10.1029/2002GL016358.
Nauhaus, K., Boetius, A., Krüger, M., Widdel, F., 2002. In vitro demonstration of
anaerobic oxidation of methane coupled to sulfate reduction from a marine gas
hydrate area. Environmental Microbiology 4, 296-305.
Nauhaus, K., Treude, T., Boetius, A., Krüger, M., 2005. Environmental regulation of the
anaerobic oxidation of methane a comparison of ANME-1 and ANME-II
communities. Environmental Microbiology 7, 98-106.
Niemann, H., Lösekann T., de Beer, D., Elvert, M., Nadalig, T., Knittel, K., Amann, R.,
Sauter, E., Schlüter, M., Klages, M., Foucher, J. -P., Boetius, A., 2006. Novel
microbial communities of the Haakon Mosby mud volcano and their role as a
methane sink. Nature 443, 854-858.
Omoregie, E. O., Mastalerz, V., de Lange, G., Straub, K. L., Kappler, A., Røy, H.,
Stadniskaia, A., Faucher, J. -P., Boetius, A., 2008. Biogeochemistry and community
composition of iron-and sulfur-precipitating microbial mats at the Chefren Mud
Volcano (Nile deep sea fan, Eastern Mediterranean). Applied and Environmental
Microbiology 74, 3198-3215.
Orcutt, B. N., Boetius, A., Elvert, M., Samarkin, V. A., Joye, S. B., 2005. Molecular
biogeochemistry of sulfate reduction, methanogenesis and the anaerobic oxidation
of methane at the Gulf of Mexico cold seeps. Geochimica et Cosmochimica Acta
69, 4267-4281.
Orphan, V. J., House, C. H., Hinrichs, K. -U., McKeegan, K. D., DeLong, E. F., 2001.
Methane-consuming archaea revealed by directly coupled isotopic and phylogenetic
analysis. Science 293, 484-487.
Orphan, V. J., House, C. H., Hinrichs, K. -U., McKeegan, K. D., DeLong, E. F., 2002.
Multiple archaeal groups mediate methane oxidation in anoxic cold seep sediments.
Proceedings of the National Academy of Science U.S.A. 99, 7663-7668.
Pancost, R. D, Sinninghe Damsté, J. S, Lint, S. D., van der Maarel MJEC, Gottschal J. C.,
Shipboard Scientific Party., 2000. Biomarker evidence for widespread anaerobic

Chapter I ________________________________________________________________________
40
methane oxidation in Mediterranean sediments by a consortium of methanogenic
archaea and bacteria. Applied Environmental Microbiology 66, 1126-1132.
Pancost, R. D., Bouloubassi, I., Aloisi, G., Sinninghe Damste, J. S., Party M. S. S., 2001.
Three series of non-isoprenoidal dialkyl glycerol diethers in cold-seep carbonate
crusts. Organic Geochemistry 32, 695-707.
Paull, C. K., Hecker, B., Commeau, R., Freman-Lynde, R. P., Neumann, C., Corso, W.
P., Golubic, S., Hook, J. E., Sikes, E., Curray, J., 1984. Biological communities at
the Florida escarpment resemble hydrothermal vent taxa. Science 206, 965-967.
Paull, C. K., Ussler III, W., Browski, W. S., Spiess, F. N., 1995. Methane- Rich plumes
on the Carolina continental rise: associations with gas hydrates. Geology 23, 89-92.
Peckmann, J., Thiel, V., Michaelis, W., Clari, P., Gaillard, C., Martire, L., Reitner, J.,
1999. Cold seep deposits of Beauvoisin (Oxfordian; southeastern France) and
Marmorito (Miocene; northern Italy): microbially induced authigenic carbonates.
International Journal of Earth Sciences 88, 60-75.
Petit, J. R., Jousel, J., Raynaud, D., Barkov, N. I., Barnola, M. -J., Basile, I., Bender, M.,
Chappellaz, J., Davis, M., Delaygue, G., Delmotte, M., Kotlyakov, V. M., Legrand,
M., Lipenkov, V. Y., Lorius, C., Pépin, L., Ritz, C., Saltzman, E., Stievenard, M.,
1999. Climate and atmospheric history of the the past 420,000 years from the
Vostok ice core, Antarctica. Nature 399, 429-436.
Reeburgh, W. S., 1976. Methane consumption in Cariaco trench waters and sediments.
Earth Planetary Science Letters 28, 337-341.
Reeburgh, W. S., 2007. Oceanic methane biogeochemistry. Chemistry Reviews 107, 486-
513.
Reeburgh, W. S., Ward B. B., Whalen S. C., Sandbeck K. A., Kilpatrick K. A., Kerkhof
L. J., 1991. Black Sea methane geochemistry. Deep-Sea Research 38 (suppl. 2),
S1189-S1210.
Rehder, G., Suess, E., 2004. Marine gas hydrates. IFM-geomar report 2002-2004, 35-38.
Reysenbach, A. -L., Longnecker, K., Kirshtein, J., 2000. Novel bacterial and archaeal
lineages from an in situ growth chamber deployed at a Mid-Atlantic Ridge
hydrothermal vent. Applied and Environmental Microbiology 66, 3798-3806.

Chapter I ________________________________________________________________________
41
Ritger, S., Carson, B., Suess, E., 1987. Methane-derived authigenic carbonates formed by
subduction-induced pore-water expulsion along Oregon/Washington margin.
Geological Society of America Bulletin 98, 147-156.
Roberts, H. H., McBride, R. A., Coleman, J., 1999. Outer shelf and slope geology of the
Gulf of Mexico: an overview In: The Gulf of Mexico Large Marine Ecosystems:
Assesment Sustainability, and Management (eds. Kumpf, H., Steidinger, K.,
Sherman, K.), 93-112. Black Science Ltd.
Rothschild, L. J., Mancinelli, R. L., 2001. Life in extreme environments. Nature 409,
1092-1101.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2002. Phospholipid analysis as a tool
to study complex microbial communities in marine sediments. Journal of
Microbiological Methods 48, 149-160.
Sahling, H., Rickert, D., Lee, R. W., Linke, P., Suess, E., 2002. Macrofaunal community
structure and sulfide flux at gas hydrate deposits from the Cascadia convergent
margin, NE Pacific. Marine Ecological Progress Series 231, 121-138.
Sassen, R., MacDonald, I. R., Requejo, A. G., Guinasso, N. L., Kennicutt, M. C., Sweet,
S. T., Brooks, J. M., 1994. Organic geochemistry of sediments from chemosythetic
communities, Gulf of Mexico slope. Geo-Marine Letters 14, 110-119.
Schönheit, P., Keweloh, H., Thauer, R. K., 1981. Factor F420 degradation in
Methanobacterium thermoautotrophicum during exposure to oxygen. Federation of
European Microbiological Societies Microbiology Letters 12, 347-349.
Schouten, S., Wakeham, S. G., Hopmans, E. C., Sinninghe Damsté J. S., 2003.
Biogeochemical evidence that thermophilic archaea mediate the anaerobic
oxidation of methane. Applied and Environmental Microbiology 69, 1680-1686.
Sturt, H. F., Summons, R. E., Smith, K. J., Elvert, M., Hinrichs, K. -U., 2004. Intact polar
membrane lipids in prokaryotes and sediments deciphered by high-performance
liquid chromatography/electrospray ionization multistage mass spectrometry-new
biomarkers for biogeochemistry and microbial ecology. Rapid Communications in
Mass Spectrometry 18, 617-628.

Chapter I ________________________________________________________________________
42
Suess, E., Bohrmann, G., Huene, R., Linke, P., Wallmann, K., Lammers, S., Sahling, H.,
1998. Fluid venting in the eastern Aleutian subduction zone. Journal of
Geophysical research 103, 2597-2614.
Suess, E., Carson B., Ritger S. D., Moore, J. C., Jones, M. L., Kulm, L. D., Cochrane, G.
R., 1985. Biological communities at vent sites along the subduction zone off
Oregon. Biological Society of Washington Bulletin 6, 475-484.
Suess, E., Torres, M. E., Bohrmann, G., Collier, R. W., Greinert, J., Linke, P., Rehder,
G., Trehu, A., Wallmann, K., Winckler, G., Zuleger, E., 1999. Gas hydrate
destabilization: enhanced dewatering, benthic material turnover and large methane
plumes at the Cascadia convergent margin. Earth Planet Science Letters 170:1-15.
Taylor, J., Parkes, R. J., 1983. The cellular fatty acids of the sulphate-reducing bacteria,
Desulfobacter sp., Desulfobulbus sp. and Desulfovibrio desulfuricans. Journal of
General Microbiology 129, 3303-3309.
Teske, A., Hinrichs, K. -U., Edgecomb, V., de Vera Gomez, A., Kysela, D., Sylva, S. P.,
Sogin, M. L., Jannasch, H. W., 2002. Microbial diversity of hydrothermal
sediments in the Guaymas Basin: Evidence for anaerobic methanotrophic
communities. Applied and Environmental Microbiology 68, 1994-2007.
Thiel, V., Peckmann, J., Richnow, H. H., Luth, U., Reitner, J., Michaelis, W., 2001.
Molecular signals for anaerobic methane oxidation in Black Sea seep carbonates
and a microbial mat. Marine Chemistry 73,97-112.
Thiel, V., Peckmann, J., Seifert, R., Wehrung, P., Reitner, J., Michaelis, W., 1999. Highly
isotopically depleted isoprenoids: Molecular markers for ancient methane venting.
Geochemica et Cosmochemica Acta 63, 3959-3966.
Tissot, B. P., Welte, D. H., 1984. Petroleum formation and occurrence. pp. 527, Springer-
Verlag, Heidelberg, 1984.
Torres, M., McManus, J., Hammond, D. E., de Angelis, M. A., Heeschen, K., Colbert, S.
L., Tyron, M. D., Brown, K. M., Suess, E., 2002. Fluid and chemical fluxes in and
out of sediments hosting methane hydrate deposits on Hydrate Ridge, OR, I:
hydrological provinces. Earth and Planetary Science Letters 201, 525-540.
Tortora, G. J., Funke, B. R., Case, C. L., 2004. Microbiology: An introduction. 8th
Edition, Pearson education Inc. Benjamin/Cumminmgs, San Francisco, USA.

Chapter I ________________________________________________________________________
43
Treude, T., Boetius, A., Knittel, K., Wallmann, K., Jørgensen B. B., 2003. Anaerobic
oxidation of methane above gas hydrates at Hydrate Ridge, NE Pacific Ocean.
Marine Ecology Progress Series 264: 1-14.
Treude, T., Knittel, K., Blumenberg, M., Seifert, R., Boetius, A., 2005. Subsurface
microbial methanotrophic mats in the Black Sea. Applied and environmental
microbiology 71, 6375-6378.
Valentine, D. L., 2007. Adaptations to energy stress dictate the ecology and evolution of
the Archaea. Nature Reviews Microbiology 4, 316-323.
Van Mooy, B. A. S., Rocap, G., Fredricks, H. F., Evans, C. T., Devol, A. H., 2006.
Sulfolipids dramatically decrease phosphorus demand by picocyanobacteria in
oligotrophic marine environments. Proceedings of the National Academy of Science
U.S.A. 103, 8607-8612.
Von Damm, K. L., 1990. Seafloor hydrothermal activity: black smoker chemistry and
chimneys. Annual Reviews Earth Planetary Science 49, 2197-2220.
Von Damm, K. L., Edmond, J. M., Grant, B., Measures, C. I., 1985. Chemistry of
submarine hydrothermal solutions at 21°N, East Pacific Rise. Geochimica et
Cosmochimica Acta 49, 2197-2220.
Wakeham, S. G., Lewis, C. M., Hopmans, E. C., Schouten, S., Sinninghe Damsté J. S.,
2003. Archaea mediate anaerobic oxidation of methane in deep euxinic waters of
the Black Sea. Geochemica et Cosmochimica Acta 67, 1359-1374.
Ward, B. B., Kilpatrick, K.A., Novelli, P. C., Scranton, M. I., 1987. Methane oxidation
and methane fluxes in the ocean surface layer and deep anoxic waters. Nature 327,
226-229.
Wegener, G., Shovitri, M., Knittel, K., Niemann, H., Hovland, M., Boetius, A., 2008.
Biogeochemical processes and microbial diversity of the Gullfaks and Tommeliten
methane seeps (Northern North Sea). Biogeosciences 5, 1127-1144.
White, D. C., Davis, W. M., Nickels, J. S., Kind, J. D., Bobbie, R. J., 1979. Oecologica
40, 51-62.
Whiticar, M. J., 1999. Carbon and hydrogen isotope systematics of bacterial formation
and oxidation of methane. Chemical Geology 161, 291-314.

Chapter I ________________________________________________________________________
44
Wuebbles, D. J., Hayhoe, K., 2002. Atmospheric methane and global change. Earth
Science Reviews 57, 177-20.
Yamamoto, S., Alcauskas, J. B., Crozier, T. E., 1976. Solubility of methane in distilled
water and seawater. Journal of Chemical Engineering Data 21, 78-81.
Yun, J. W., Orange, D. L., Field, M. E., 1999. Subsurface gas offshore of Northern
Califorornia and its link to submarine geomorphology. Marine Geology 154, 357-
368.
Ziebis, W., Haese, R. R., 2005. Interactions between fluid flow, geochemistry, and
biogeochemical processes at methane seeps. In: Macro- and Microorganisms in
Marine Sediments (eds. Kristensen E., Kostka J., Haese R. R.), 267-298, AGU
Coastal and Estuarine Studies.

Chapter II ________________________________________________________________________
45
CHAPTER II
Intact polar lipids of anaerobic methanotrophic archaea
and associated bacteria
Pamela E. Rossela, Julius S. Lippa, Helen F. Fredricksb, Julia Arndsc, Antje Boetiusc,
Marcus Elverta, Kai-Uwe Hinrichsa
Published in Organic Geochemistry.
vol. 39, page 992-999, doi:10.1016/j.orggeoche.2008.02.021
aOrganic Geochemistry Group, Department of Geosciences, University of Bremen, 28334 Bremen,
Germany bWoods Hole Oceanographic Institution, Department of Marine Chemistry and Geochemistry, Woods Hole,
MA 02543, USA cMax-Planck-Institute for Marine Microbiology, 28359 Bremen, Germany

Chapter II ________________________________________________________________________
46
II.1. PRINTED MANUSCRIPT
ABSTRACT
Previous biomarker studies of microbes involved in anaerobic oxidation of
methane (AOM) have targeted non-polar lipids. We have extended the biomarker
approach to include intact polar lipids (IPLs) and show here that the major community
types involved in AOM at marine methane seeps can be clearly distinguished by these
compounds. The lipid profile of methanotrophic communities with dominant ANME-1
archaea mainly comprises diglycosidic GDGT derivatives. IPL distributions of microbial
communities dominated by ANME-2 or ANME-3 are consistent with their phylogenetic
affiliation with the euryarchaeal order Methanosarcinales, i.e., the lipids are dominated by
phosphate-based polar derivatives of archaeol and hydroxyarchaeol. IPLs of associated
bacteria strongly differed among the three community types analyzed here; these
differences testify to the diversity of bacteria in AOM environments. Generally, the
bacterial members of methanotrophic communities are dominated by
phosphatidylethanolamine and phosphatidyl-(N,N)-dimethylethanolamine species; polar
dialkylglycerolethers are dominant in the ANME-1 community while in ANME-2 and
ANME-3 communities mixed acyl/ether glycerol derivatives are most abundant. The
relative concentration of bacterial lipids associated with ANME-1 dominated
communities appears significantly lower than in ANME-2 and ANME-3 dominated
communities. Our results demonstrate that IPL analysis provides valuable molecular
fingerprints of biomass composition in natural microbial communities and enables
taxonomic differentiation at the rank of families to orders.
Abbreviations: ANME, anaerobic methanotrophic archaea; AR, archaeol; AOM, anaerobic oxidation of
methane; CARD–FISH, catalyzed reporter deposition–fluorescence in situ hybridization; GDGT,
glyceroldialkylglyceroltetraether; IPL, intact polar lipid; SRB, sulfate-reducing bacteria; OH-AR,
hydroxyarchaeol; PC, phosphatidylcholine; PDME, phosphatidyl-(N,N)-dimethylethanolamine; PE,
phosphatidylethanolamine; PG, phosphatidylglycerol; PI, phosphatidylinositol; PME, phosphatidyl-(N)-
methylethanolamine; PS, phosphatidylserine; 2-Gly, diglycosyl; 2OH-AR, dihydroxyarchaeol.

Chapter II ________________________________________________________________________
47
INTRODUCTION
Anaerobic oxidation of methane (AOM) in the marine environment is mediated
by three phylogenetically distinct clusters of Euryarchaeota called ANME-1, -2 or -3 (cf.
Hinrichs et al., 1999; Boetius et al., 2000; Hinrichs and Boetius, 2002; Niemann et al.,
2006) that form consortia with sulfate-reducing bacteria (SRB) (Boetius et al., 2000;
Orphan et al., 2001a, 2002; Lösekann et al., 2007). ANME-2 are phylogenetically
affiliated with the order Methanosarcinales and are typically observed in physical
association with SRB of the Desulfosarcina/Desulfococcus group (Boetius et al., 2000;
Orphan et al., 2001b, ‘‘ANME-2/DSS aggregates”). ANME-3 are closely related to the
genera Methanococcoides and Methanolobus and have been found in association with
SRB related to Desulfobulbus spp. (Lösekann et al., 2007, ‘‘ANME-3/DBB aggregates”).
ANME-1 are not directly affiliated with any of the major orders of methanogens
(Hinrichs et al., 1999; Orphan et al., 2001b; Knittel et al., 2005). These archaea have been
observed in physical association with SRB of the Desulfosarcina/Desulfococcus group in
microbial mats (Michaelis et al., 2002) but also frequently as monospecific aggregates or
as single cells without a clear bacterial partner (Orphan et al., 2002).
Previous biomarker studies of AOM communities have focused on non-polar
lipids, such as hydrocarbons of archaeal origin, bacterial fatty acids, and archaeal and
bacterial glycerol-based ether lipids (e.g., Elvert et al., 1999, 2005; Hinrichs et al., 1999,
2000; Pancost et al., 2000; Blumenberg et al., 2004; Niemann et al., 2006). However, an
interpretation of the lipid profiles with regard to the distribution and composition of
active methanotrophic communities is limited by their relatively low taxonomic
specificity and the likelihood of incorporating signals from the past. The latter point is
particularly crucial due to the temporally highly dynamic physical–chemical conditions
encountered in many of the intensely studied AOM environments. By contrast, intact
polar lipids (IPLs) offer a more detailed view of microbial communities due to their
higher taxonomic specificity and property to select for live biomass (Rütters et al., 2002;
Sturt et al., 2004; Biddle et al., 2006).
Here we report the composition of IPLs in environmental samples dominated by
either one of the three major ANME groups and associated bacteria. We show that IPL

Chapter II ________________________________________________________________________
48
profiles can serve as valuable community fingerprints and relative indicators of biomass
of ANME archaea and associated bacteria in natural systems.
MATERIAL AND METHODS
IPL analysis
Samples from four different seep environments were analyzed, each dominated by
one distinct ANME group (Table II.1 and Fig. II.1): two microbial mats from the
northwestern Black Sea, one sediment sample from Hydrate Ridge and one sediment
sample from Häkon Mosby Mud Volcano. Both surface sediment samples from Hydrate
Ridge and Häkon Mosby Mud Volcano were covered by Beggiatoa mats.
IPL analysis was performed with a HPLC–ESI–MSn system using protocols
described previously by Sturt et al. (2004) and Biddle et al. (2006). Total lipid extracts
from microbial mats from the Black Sea and the sediment from Häkon Mosby Mud
Volcano were obtained with an automated microwave-assisted extraction system
(MARS-X, CEM, USA) at a temperature of 70°C, while the sediment from Hydrate
Ridge was extracted via ultrasonication. The latter sample was analyzed after
chromatographic separation as glyco- and phospholipids fraction (Sturt et al., 2004),
while the former samples were analyzed as total lipid extracts. Structural assignments
were based on mass spectral interpretation (cf. Sturt et al., 2004) and by comparison with
IPL inventories of cultured archaea and bacteria (e.g., Koga et al., 1998; Koga and Morii,
2005; Hinrichs et al., unpublished data). Chain length assignment, degree of unsaturation,
and determination of ether and ester bond linkages of bacterial IPLs were based on
molecular masses and fragments according to Sturt et al. (2004). Due to the limited
availability of commercial standards, we did not use response factors for IPL
quantification. Based on calibration curves we observed response factors for various
commercially available IPLs that can differ up to a factor of three. Thus, reported relative
distributions are semi-quantitative. Only compounds with a signal-to-noise ratio higher
than 6 were reported. The least concentrated reported compounds amounted to 0.12%,
0.06%, 5.0% and 5.2% of the total quantified IPLs in samples from Black Sea (two
samples), Hydrate Ridge and Häkon Mosby Mud Volcano, respectively. Composite

Chapter II ________________________________________________________________________
49
chromatograms of extracted quasi-molecular ions of individual IPLs were obtained from
the full scan (m/z 500–2000) from each sample (Fig. 1). Structural details of hexoses
linked via glycosidic bonds to archaeal ether lipids are not resolved; hence hexoses are
designated as glycolipids.
Catalyzed reporter deposition-fluorescence in situ hybridization (CARD-FISH)
analyses
Microbial mats were fixed as described previously (Treude et al., 2007) and
homogenized. In situ hybridizations with horseradish peroxidase (HRP)-conjugated
probes followed by tyramide signal amplification were carried out as described by
Pernthaler et al. (2002) with slight modifications: endogenous peroxidases were inhibited
with methanol (30 min) and rigid archaeal cell walls were permeabilized with proteinase
K(15 μg ml-1, for 2 min at room temperature). Total cell counts were determined by 4’,6’-
diamidino-2-phenylindole (DAPI)-staining.
Hybridized and DAPI-stained samples were examined with an epifluorescence
microscope (Axiophot II microscope; Carl Zeiss, Jena, Germany). For each probe and
sample 700 DAPI-stained cells in 70 independent microscopic fields were counted. Probe
sequences and formamide concentrations required for specific hybridization were:
ARCH915 (35% formamide), EUB338 I–III (35% formamide), ANME-1–350 and
EelMS932 (40 and 60% formamide) (Amann et al., 1990; Daims et al., 1999; Boetius et
al., 2000).

Cha
pter
II
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
50
Tab
le II
.1:
Ove
rvie
w o
f AO
M sa
mpl
es a
naly
zed,
rela
tive
dist
ribut
ion
of b
oth
arch
aeal
and
bac
teria
l cel
ls a
nd IP
Ls, a
nd c
ore
lipid
dis
tribu
tion
of d
igly
cosy
l-G
DG
Ts.
Loca
tion
Sam
ple
Archaea
vs.
Bacteria
(%
of t
otal
cel
ls) a
AN
ME-
1/-2
/-3
(% o
f tot
al
arch
aeal
cel
ls)
Sour
ce
Arc
haea
l gly
co-
and
phos
phol
ipid
s
(% o
f tot
al a
rcha
eal I
PLs)
Bac
teria
l ph
osph
olip
ids
(% o
f to
tal b
acte
rial I
PLs)
b
Cor
e lip
id ri
ng
dist
ribut
ion
of
2-G
ly-G
DG
Ts
Nor
thw
este
rn
Bla
ck S
ea,
Dni
epr a
rea
Mat
795
, pin
k
(189
m w
ater
de
pth)
33
/16
100/
0/0
this
stu
dy
2-G
ly-G
DG
T (>
99),
2-G
ly-A
R (<
1)
DE
G-P
E C
30:0
(31)
, D
AG
/AE
G-P
E C
33:2
(19)
, D
EG
-PE
C31
:1 (1
8),
DA
G/A
EG
-PE
C35
:2 (1
1),
DA
G/A
EG
-PE
C32
:2 (1
1),
othe
r PE
s (1
0)
3>2>
>1>>
0>5
Nor
thw
este
rn
Bla
ck S
ea,
Dni
epr a
rea
Mat
822
, ree
f to
p (1
90 m
w
ater
dep
th)
31/4
3 35
/65/
0 th
is s
tudy
PG
-AR
(35)
, 2-G
ly-G
DG
T (3
3), t
enta
tive
phos
pho-
AR
(1
9), 2
-Gly
-AR
(5),
PS
-OH
-A
R (2
), 2-
Gly
-OH
-AR
(1),
PE
-OH
-AR
(1)
DA
G/A
EG
-PE
C31
:2 (3
2),
DE
G-P
E C
32:1
(11)
, D
AG
/AE
G-P
E C
31:1
(10)
, D
AG
/AE
G-P
E C
32:2
(9),
DE
G-P
E C
31:1
(8),
othe
r P
Es
and
PC
s (3
2)
3>2>
>1>>
0 c
Hyd
rate
R
idge
Sedi
men
t (S
tatio
n 19
-2,
2-3
cm)
31/6
7 <2
/97/
0 K
nitte
l et
al. (
2005
)
PG
-OH
-AR
(23)
, PE
-OH
-A
R (1
5), P
I-OH
-AR
(13)
, P
S-O
H-A
R (1
4), P
G-A
R
(12)
, 2-G
ly-A
R (1
1), 2
-Gly
-G
DG
T (6
), P
S-2
OH
-AR
(2
), te
ntat
ive
phos
pho-
AR
(2
), P
S-A
R (1
)
DA
G/A
EG
-PE
C32
:2 (1
6),
DA
G/A
EG
-PG
C34
:2
(15)
, DA
G/A
EG
-PE
C
34:2
(11)
, DA
G/A
EG
-PE
C
32:1
(7),
DA
G/A
EG
-PG
C
36:2
(7),
othe
r PE
s,
PC
s, a
nd P
Gs,
(44)
3>2>
1 d
Håk
on
Mos
by M
ud
Vol
cano
Sedi
men
t (S
tatio
n 19
, 1-
2 cm
) 77
/10
0/
0/99
Lö
seka
nn
et a
l. (2
007)
PG
-OH
-AR
(53)
, PS
-OH
-A
R (1
6), P
I-OH
-AR
(9),
PS
-2O
H-A
R (5
), P
S-A
R (2
)
DA
G/A
EG
-PD
ME
C32
:2
(21)
, DA
G/A
EG
-PE
C
32:2
(18)
, DA
G/A
EG
-P
DM
E C
34:2
(11)
, oth
er
PE
s, P
Cs,
PG
s, P
ME
s an
d P
DM
Es
(50)
No
GD
GTs
Rel
ativ
e am
ount
s of I
PLs a
re b
ased
on
peak
are
a in
mas
s chr
omat
ogra
ms o
f sel
ecte
d m
olec
ular
ions
. For
bac
teria
l IPL
s, bo
nd ty
pes b
etw
een
alky
l moi
etie
s and
gl
ycer
ol a
re d
istin
guis
hed
(DEG
= d
ieth
er, D
AG
= d
iacy
l, A
EG =
mix
ed),
follo
wed
by
head
gro
ups,
the
sum
of c
arbo
n at
oms i
n bo
th a
lkyl
cha
ins a
nd n
umbe
r of
unsa
tura
tions
. Bac
teria
l IPL
dat
a fro
m H
ydra
te R
idge
wer
e pr
evio
usly
repo
rted
by S
turt
et a
l. (2
004)
. a P
erce
ntag
e de
rived
from
CA
RD
–FIS
H, e
xpre
ssed
rela
tive
to D
API
cou
nts (
100%
). b D
istin
ctio
n be
twee
n D
AG
and
AEG
not
pos
sibl
e un
der H
PLC
–MS
cond
ition
s app
lied,
indi
cate
d al
kyl c
hain
s pro
vide
d fo
r DA
G.
C N
o G
DG
T w
ith 5
ring
s det
ecte
d.
D N
o G
DG
Ts w
ith 0
and
5 ri
ngs d
etec
ted.

Chapter II ________________________________________________________________________
51
RESULTS AND DISCUSSION
Fig. II.1. Composite mass chromatograms of molecular ions of IPLs in samples dominated by ANME-1 (A), a mixed ANME-1/ANME-2 community (B), both from microbial mats collected in the Black Sea (BS), and sediments dominated by ANME-2 (C) and ANME-3 (D) from Hydrate Ridge (HR) and Ha�kon Mosby Mud Volcano (HMMV), respectively. Extracted m/z of quasi-molecular ions for archaeal IPLs are 1632-1645, 994, 807, 956, 823, 792, 820, 836, 852, 911 for the identified 2-Gly-GDGT, 2-Gly-AR, PG-AR, tentative P-AR, PGOH-AR, PE-OH-AR, PS-AR, PS-OH-AR, PS-2OH-AR, PI-OH-AR, respectively. The major bacterial IPLs are represented by the following quasi-molecular ions: m/z 674 and 688 for DAG/AEG-PE, 662 and 660 for DEG-PE, 764 and 736 for DAG/AEG-PG, 760 for DAG/AEG PC, 716 for DAG/AEG-PDME and 702 for PME.
IPLs of ANME-1
In the microbial mat from the trunk of a microbial reef in the Black Sea, all
archaeal cells were affiliated with ANME-1 (Table II.1). In this sample, diglycosyl
glyceroldialkylglyceroltetraethers (2-Gly-GDGTs) were the most abundant IPLs (Fig.
II.1A). Only small amounts of 2-Gly-archaeol (2-Gly-AR) were detected (Fig. II.2A).
The main GDGT core lipids in 2-Gly-GDGT were the di- and tri-cyclopentyl derivatives

Chapter II ________________________________________________________________________
52
(Table II.1). No polar derivative of hydroxyarchaeol (OH-AR) was detected. In terms of
the IPL composition, ANME-1 are distinct from other methanogens (e.g., Koga and
Morii, 2005), i.e., all major families of methanogens produce significant amounts of AR
and multiple types of phosphate-based IPLs. In fact, ANME-1 is most similar to members
of the hyperthermophilic Archaeoglobales that largely produce 2-Gly-GDGT, combined
with lower amounts of both 1-Gly-GDGT and small quantities of Gly-AR (Hinrichs et
al., unpublished data). Our results are consistent with evidence provided by Thiel et al.
(2007) who applied molecular imaging techniques based on ToF-SIMS to mat sections
obtained from the same reef system and dominated by cells of the ANME-1 morphology;
these sections consisted mainly of free GDGTs and 2-Gly-GDGTs.
The absence or extremely low relative abundance of OH-AR in ANME-1 archaea
was not apparent in earlier studies that focused on its non-polar derivatives (Hinrichs et
al., 1999; Blumenberg et al., 2004). However, in the ANME-1 dominated mat, the
concentration of non-polar OH-AR of 15 μg/g mat was very low compared to the mat
dominated by ANME-2 (436 μg/g mat). Low ratios of OH-AR/AR have been used as
indicator signatures of active ANME-1 (Blumenberg et al., 2004; Niemann and Elvert, in
press), but probably have to be interpreted with caution. We suggest that in ANME-1-
dominated environments lacking polar OH-AR, non-polar OH-AR is a relict from the
past, when environmental conditions selected for ANME-2.
IPLs of ANME-2
The mat from the top part of a reef structures in the Black Sea was characterized
by a mixture of ANME-1 and ANME-2 (Table II.1, 35% and 65% of total archaeal cells,
respectively), while the surface sediment sample from Hydrate Ridge was dominated by
ANME-2 (Table II.1, 97% of total archaeal cells, Knittel et al., 2005). Archaeal IPLs of
ANME-2 were largely based on AR and OH-AR with either glycosidic or phosphate-
based headgroups. Specifically, these included 2-Gly-AR, 2-Gly-OH-AR,
phosphatidylglycerol- (PG-) OH-AR, PG-AR, phosphatidylinositol- (PI-) AR, PI-OH-
AR, phosphatidylserine- (PS-) AR, PS-OH-AR, PS-2-OH-AR and a tentatively identified
AR with a phosphate-based headgroup of unknown structure (Fig. II.1B and C). This
phospho-AR, present in both samples containing ANME-2, but not in the ANME-1 and

Chapter II ________________________________________________________________________
53
ANME-3 dominated communities, was tentatively assigned based on information
obtained in negative ionization mode, which yielded an intense fragment of 433.5 Da
(interpreted as dehydrated lyso fragment with one phytanyl chain and without head
group). The unknown compound is formed by two masses 956.0 and 939.3 that probably
correspond to the ammonium adduct and the protonated lipid, respectively. None of the
corresponding ions yielded intense, interpretable fragments during MS2 experiments in
positive ionization mode. 2-Gly-GDGT was also detected in the two ANME-2 dominated
communities from Black Sea and Hydrate Ridge; relative concentrations of this
compound are consistent with the relative amounts of ANME-1 cells in these samples
(Table II.1 and Fig. II.1).
IPLs of ANME-3
The sediments from Häkon Mosby Mud Volcano were dominated by ANME-3
(Table II.1, 99% total archaeal cells, Lösekann et al., 2007). The sample contained the
most diverse distribution of archaeal and bacterial IPLs (Fig. II.1D). The main archaeal
IPLs were various phospholipids of AR and OH-AR, similar to those observed in the
ANME-2 system of Hydrate Ridge. In contrast to the ANME-1 and ANME-2 dominated
communities, neither GDGT-based IPLs nor glycosidic archaeol derivatives were
present. Likewise, the tentatively identified phospho-AR from the ANME-2 community
(Fig. II.1B and C) was not detected.
Bacterial IPLs
Compositional differences of bacterial IPLs reflect differences in the phylogenetic
affiliation of the bacterial members of AOM communities such as SRB (Fig. II.1A–D).
Bacterial IPLs vary in both structural diversity and relative abundance; ANME-3 and
ANME-2 dominated samples displayed both the highest abundance and highest diversity
of bacterial IPLs (Table II.1 and Fig. II.2). The Black Sea ANME-1 system was
dominated by phosphatidylethanolamine (PE) derivatives of dietherglycerol (DEG) lipid
types (Table II.1), while the ANME-2 systems contained mainly PE of mixed acyl/ether
glycerol (AEG) lipids or diacyl glycerol (DAG) lipids, although the corresponding DEG
types were also present. The high relative amounts of DAG/AEG lipids in combination

Chapter II ________________________________________________________________________
54
with PE and PG is consistent with the IPL composition of Desulfosarcina variabilis
(Rütters et al., 2001), a close relative of the sulfate reducers in ANME-2 communities,
although the chain length distribution and degree of unsaturation differ (Table II.1; cf.
Rütters et al., 2001; Sturt et al., 2004). In ANME-2 dominated communities, we also
observed phosphatidylcholine (PC) and PG (the latter observed in the Hydrate Ridge
sample only). With respect to the bacterial IPLs, the ANME-3 community is
distinguished from the ANME-2 community by a higher abundance of phosphatidyl-(N)-
methylethanolamine (PME) and phosphatidyl-(N,N)-dimethylethanolamine (PDME).
Fig. II.2. Compositional variation of IPL groups in the four AOM communities. (A) Distribution of archaeal IPLs (Gly-GDGT, Gly-AR and Gly-OH-AR, P-AR and P-OH-AR [P = phospho]), (B) relative amounts of archaeal and bacterial IPLs.
In all samples, PE was a major bacterial IPL and contributes between ~1 and 15%
to total IPLs in Black Sea mat samples, and between 15% and 40% in the sediments from
Häkon Mosby Mud Volcano and Hydrate Ridge, respectively (cf. Sturt et al., 2004, for
detailed discussion of bacterial IPLs in Hydrate Ridge sample). The total number of
carbon atoms in glycerol-bound acyl and/or alkyl moieties further distinguished the three
ANME community types. In the ANME-1 dominated sample, the dominant bacterial IPL

Chapter II ________________________________________________________________________
55
was a C30:0 DEGPE; in samples from Hydrate Ridge and Häkon Mosby Mud Volcano,
C32:2 DAG/AEG-PE and C32:2 DAG/AEG-PDME, respectively, were more abundant
(Table II.1).
Lipid taxonomy of uncultured AOM archaea
The presence of AR and OH-AR based core lipids in ANME-2 and ANME-3
archaea is consistent with their affiliation with the methanogenic orders Methanococcales
and Methanosarcinales (cf. Kates, 1997). When considering the presence and/or absence
of polar headgroups in ANME-2 and ANME-3 communities, the taxonomic relationship
is narrowed down to the Methanosarcinales: PI and PG derivatives are abundant in the
Methanosarcinales and but are absent in the Methanococcales (Koga and Morii, 2005).
Notably, no clear chemotaxonomic relationship exists between the phylogenetically
distinctive ANME-1 and any of the cultured methanogens (cf. Koga et al., 1998; Koga
and Morii, 2005). Closest relatives of ANME-1 in terms of IPL composition are the
Archaeoglobales (Hinrichs et al., unpublished data). Environmental IPL fingerprints that
resemble those from ANME-1 are those related to uncultured marine sedimentary
archaea (Biddle et al., 2006).
Archaeal vs. bacterial biomass
Relative amounts of archaeal vs. bacterial IPLs strongly varied in the samples
dominated by a single ANME type (percentages of archaeal IPLs are ~99%, 85%, 31%
and 52% for samples from the Black Sea, Hydrate Ridge, and Häkon Mosby Mud
Volcano, respectively; Fig. II.2B). These pronounced differences probably reflect
similarly large differences in archaeal vs. bacterial biomass among active AOM
community members, which in turn probably relate to ecophysiological characteristics of
the three types of AOM communities sampled here. Notably, the IPLs partly provided a
different picture of the relative abundance of archaeal vs. bacterial biomass than cell
counts by CARD–FISH (Table II.1). For example, for the Black Sea ANME-1 sample,
IPL analysis suggests a lower bacterial contribution to the microbial community than
CARD–FISH, while in the Häkon Mosby Mud Volcano ANME-3 sample, CARD–FISH
detected more archaea than IPL analysis (Fig. II.2B and Table II.1). Possible causes

Chapter II ________________________________________________________________________
56
include varying cellular IPL contents, e.g., due to differences in cell size and
morphology, and/or differences in physiological status of the bulk community that in turn
may affect both cellular IPL abundance and the ability to bind to CARD–FISH probes.
CONCLUSIONS
This study provides an unprecedented view of the lipid diversity of the three
globally relevant anaerobic methanotrophic communities, that is, communities dominated
by ANME-1, ANME-2, ANME-3 populations, and associated bacteria. The diversity and
relative amounts of both archaeal and bacterial IPLs differ remarkably between the three
community types. While lipid analysis is unable to capture the entire microbial diversity,
our results demonstrate that quantitative differences in microbial community structure
can be effectively resolved. Specifically, IPL analysis enables the differentiation of the
major players in natural microbial communities at the rank of taxonomic orders or higher.
ACKNOWLEDGMENTS
We thank the crew and shipboard scientist of R/V SONNE SO 148-1, R/V
L’Atalante, and R/V Poseidon for support during sample collection. Tina Treude is
gratefully acknowledged for providing microbial mat samples from the Black Sea, Katrin
Knittel for helping with analysis of FISH data, and Philippe Schaeffer and Richard
Pancost for their constructive reviews. This study was part of the program MUMM II
(grant 03G0608C), funded by the Bundesministerium für Bildung und Forschung
(BMBF, Germany) and the Deutsche Forschungsgemeinschaft (DFG, Germany). Further
support was provided from the Center of Marine Environmental Sciences (MARUM) at
the University of Bremen funded by the DFG. This is publication GEOTECH-316 of the
R&D program GEOTECHNOLOGIEN and MARUM-publication 0573.

Chapter II ________________________________________________________________________
57
REFERENCES
Amann, R.I., Binder, B.J., Olson, R.J., Chisholm, S.W., Devereux, R., Stahl, D.A., 1990.
Combination of 16S rRNA targeted oligonucleotide probes with flow cytometry
for analyzing mixed microbial populations. Applied and Environmental
Microbiology 56, 1919–1925.
Biddle, J.F., Lipp, J.S., Lever, M.A., Lloyd, K.G., Sörensen, K.B., Anderson, R.,
Fredricks, H.F., Elvert, M., Kelly, T.J., Schrag, D.P., Sogin, M.L., Brenchley,
J.E., Teske, A., House, C.H., Hinrichs, K.-U., 2006. Heterotrophic archaea
dominate sedimentary subsurface ecosystems off Peru. Proceedings of the
National Academy of Sciences of the USA 103, 3846–3851.
Blumenberg, M., Seifert, R., Reitner, J., Pape, T., Michaelis, W., 2004. Membrane lipid
patterns typify distinct anaerobic methanotrophic consortia. Proceedings of the
National Academy of Sciences of the USA 101, 11111–11116.
Boetius, A., Ravenschlag, K., Schubert, C.J., Rickert, D., Widdel, F., Gieseke, A.,
Amann, R., Jørgensen, B.B., Witte, U., Pfannkuche, K., 2000. A marine microbial
consortium apparently mediating anaerobic oxidation of methane. Nature 407,
623–626.
Daims, H., Bruhl, A., Amann, R., Schleifer, K.H., Wagner, M., 1999. The domain-
specific probe EUB338 is insufficient for the detection of all Bacteria:
development and evaluation of a more comprehensive probe set. Systematic and
Applied Microbiology 22, 434–444.
Elvert, M., Suess, E., Whiticar, M.J., 1999. Anaerobic methane oxidation associated with
marine gas hydrates: superlight Cisotopes from saturated and unsaturated C20 and
C25 irregular isoprenoids. Naturwissenschaften 86, 295–300.
Elvert, M., Hopmans, E.C., Treude, T., Boetius, A., Suess, E., 2005. Spatial variations of
methanotrophic consortia at cold methane seeps: implications from a high-
resolution molecular and isotopic approach. Geobiology 3, 195–209.
Hinrichs, K.-U., Boetius, A., 2002. The anaerobic oxidation of methane: new insights in
microbial ecology and biogeochemistry. In: Ocean Margin Systems (eds. Wefer,

Chapter II ________________________________________________________________________
58
G., Billett, D., Hebbeln, D., Jørgensen, B.B., Schlüter, M., van Weering, T.C.E.,
57–477, Springer-Verlag
Hinrichs, K.-U., Hayes, J.M., Sylva, S.P., Brewer, P.G., Delong, E.F., 1999. Methane-
consuming archaebacteria in marine sediments. Nature 398, 802–805.
Hinrichs, K.-U., Summons, R.E., Orphan, V., Sylva, S.P., Hayes, J.M., 2000. Molecular
and isotopic analysis of anaerobic methane-oxidizing communities in marine
sediments. Organic Geochemistry 31, 1685–1701.
Kates, M., 1997. Diether and tetraether phospholipids and glycolipids as molecular
markers for archaeabacteria (archaea). In: Molecular Markers in Environmental
Geochemistry (ed. Eganhause, R.P.), 35–48, Oxford University Press.
Knittel, K., Lösekann, T., Boetius, A., Kort, R., Amann, R., 2005. Diversity and
distribution of methanotrophic archaea at cold seeps. Applied and Environmental
Microbiology 71, 467–479.
Koga, Y., Morii, H., 2005. Recent advances in structural research on ether lipids from
archaea including comparative and physiological aspects. Bioscience
Biotechnology and Biochemistry 69, 2019–2034.
Koga, Y., Morii, H., Akagawa-Matsushita, M., Ohga, I., 1998. Correlation of polar lipid
composition with 16S rRNA phylogeny in methanogens. Further analysis of lipid
component parts. Bioscience Biotechnology and Biochemistry 62, 230–236.
Lösekann, T., Knittel, K., Nadalig, T., Fuchs, B., Niemann, H., Boetius, A., Amann, R.,
2007. Diversity and abundance of aerobic and anaerobic methane oxidizers at the
Haakon Mosby Mud Volcano, Barents Sea. Applied and Environmental
Microbiology 73, 3348–3362.
Michaelis, W., Seifert, R., Nauhaus, K., Treude, T., Thiel, V., Blumenberg, M., Knittel,
K., Gieseke, A., Peterknecht, K., Pape, T., Boetius, A., Amann, R., Jorgensen,
B.B., Widdel, F., Peckmann, J.R., Pimenov, N.V., Gulin, M.B., 2002. Microbial
reefs in the Black Sea fueled by anaerobic oxidation of methane. Science 297,
1013–1015.
Niemann, H., Elvert, M., 2008. Diagnostic lipid biomarker and stable carbon isotope
signatures of microbial communities mediating the anaerobic oxidation of
methane with sulphate. Organic Geochemistry 39, 1668-1677.

Chapter II ________________________________________________________________________
59
Niemann, H., Lösekann, T., Beer, D., Elvert, M., Nadalig, T., Knittel, K., Amann, R.,
Sauter, E.J., Schlüter, M., Klages, M., Foucher, J.P., Boetius, A., 2006. Novel
microbial communities of the Häkon Mosby mud volcano and their role as a
methane sink. Nature 443, 854–858.
Orphan, V.J., Hinrichs, K.U., Ussler, W., Paull, C.K., Taylor, L.T., Sylva, S.P., Hayes,
J.M., Delong, E.F., 2001a. Comparative analysis of methane-oxidizing archaea
and sulfatereducing bacteria in anoxic marine sediments. Applied and
Environmental Microbiology 67, 1922–1934.
Orphan, V.J., House, C.H., Hinrichs, K.-U., McKeegan, K.D., Delong, E.F., 2001b.
Methane-consuming archaea revealed by directly coupled isotopic and
phylogenetic analysis. Science 293, 484–487.
Orphan, V., House, C.H., Hinrichs, K.-U., McKeegan, K.D., Delong, E.F., 2002.
Multiple archaeal groups mediate methane oxidation in anoxic cold seep
sediments. Proceedings of the National Academy of Sciences of the USA 99,
7663–7668.
Pancost, R.D., Sinninghe Damsté, J.S., de Lint, S., van der Maarel, M.J.E.C., Gottschal,
J.C.and the Medinaut Shipboard Scientific Party, 2000. Biomarker evidence for
widespread anaerobic methane oxidation on Mediterranean sediments by a
consortium of methanogenic archaea and bacteria. Applied and Environmental
Microbiology 66, 1126–1132.
Pernthaler, A., Pernthaler, J., Amann, R., 2002. Fluorescence in situ hybridization and
catalyzed reporter deposition for the identification of marine bacteria. Applied and
Environmental Microbiology 68, 3094–3101.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2001. Monoalkylether phospholipids in
the sulfate-reducing bacteria Desulfosarcina variabilis and Desulforhabdus
amnigenus. Archives of Microbiology 176, 435–442.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2002. Phospholipid analysis as a tool
to study complex microbial communities in marine sediments. Journal of
Microbiological Methods 48, 149–160.
Sturt, H.F., Summons, R.E., Smith, K., Elvert, M., Hinrichs, K-U., 2004. Intact polar
membrane lipids in prokaryotes and sediments deciphered by high-performance

Chapter II ________________________________________________________________________
60
liquid chromatography/electrospray ionization multistage mass spectrometry –
new biomarkers for biogeochemistry and microbial ecology. Rapid
Communications in Mass Spectrometry 18, 617–628.
Thiel, V., Heim, C., Arp, G., Hahmann, U., Sjövall, P., Lausmaa, J., 2007. Biomarkers at
the microscopic range: ToF-SIMS molecular imaging of Archaea-derived lipids in
a microbial mat. Geobiology 5, 413–421.
Treude, T., Orphan, V., Knittel, K., Gieseke, A., House, C.H., Boetius, A., 2007.
Consumption of methane and CO2 by methanotrophic microbial mats from gas
seeps of the anoxic Black Sea. Applied and Environmental Microbiology 73,
2271–2283.

Chapter II ________________________________________________________________________
61
II.2. SUPPLEMENTARY ONLINE MATERIAL
Supplementary Figure
Supplementary Fig. II.3. Structure of IPLs identified in this study

Chapter II ________________________________________________________________________
62

Chapter III ________________________________________________________________________
63
CHAPTER III
Factors controlling the distribution of anaerobic
methanotrophic communities in marine environments:
evidence from intact polar membrane lipids
Pamela E. Rossela, Marcus Elverta, Alban Rametteb,
Antje Boetiusb and Kai-Uwe Hinrichsa
Prepared for submission
aOrganic Geochemistry Group, Department of Geosciences, University of Bremen, 28334 Bremen,
Germany cMax-Planck-Institute for Marine Microbiology, 28359 Bremen, Germany

Chapter III ________________________________________________________________________
64
III.1. MANUSCRIPT
ABSTRACT
Three distinct types of anaerobic methanotrophic microbial consortia are globally
distributed in marine sediments. These communities are dominated by archaea of the
ANME-1, ANME-2 and ANME-3 clades and their bacterial partners. All three ANME
groups co-occur with sulfate reducing bacteria either of the Desulfosarcina-
Desulfococcus branch (ANME- 1/DSS and-2/DSS) or with Desulfobulbus spp (ANME-
3/DBB). Frequently one ANME group dominates, but the factors controlling their
distribution and abundance are not well constrained. We used a lipid-based approach to
investigate linkages between the composition of anaerobic methanotrophic communities
and environmental factors in a geographically diverse set of seep systems. Intact polar
lipids (IPLs) provided a better distinction of the composition of living communities than
their apolar (fossil) derivatives, probably due to the preservation of the apolar lipids
beyond the lifetime of the cells. Based on the analysis of a substantial set of different
microbial communities, assignments of IPLs to certain ANME community types were
found to be robust and taxonomically useful. In ANME-1/DSS communities glycosidic-
and phospho- glyceroldialkylglyceroltetraethers were abundant, while ANME-2/DSS and
ANME-3/DBB communities were dominated by a diverse range of glycosidic- and
phospho- archaeols in combination with bacterial phospholipids from sulfate reducing
bacteria. Beside these main IPL signatures, additional differences were related to the
habitat characteristics of these communities (e.g., lower amount of phosphorus-
containing IPLs were observed in communities inhabiting carbonate reefs compared to
sediments). Moreover, the habitats of ANME-1/DSS communities were characterized by
higher temperatures and lower oxygen content (or even anoxia) compared to ANME-
2/DSS and ANME-3/DBB habitats. In ANME-2 dominated environments, higher
oxygen availability from bottom waters and efficient supply of methane and sulfate were
the controlling factors.

Chapter III ________________________________________________________________________
65
Abbreviations: AR = archaeol, BL = betaine lipids, Crocetane = 2,6,11,15-tetramethylhexadecane,
2Gly = diglycosyl, DAG=diacylglycerol, DEG=dietherglycerol, GDGT = glyceroldialkylglyceroltetraether,
OH-AR = hydroxyarchaeol, OL = ornithine lipids, PC = phosphatidylcholine, PDME = phosphatidyl-
(N,N)-dimethylethanolamine, PE = phosphatidylethanolamine, PG = phosphatidylglycerol, PI =
phosphatidylinositol, PME = phosphatidyl-(N)-methylethanolamine, PMI =2,6,11,15,19-
pentamethylicosane, PS = phosphatidylserine, SRB = sulfate reducing bacteria, SRR = sulfate reduction
rate.
INTRODUCTION
Anaerobic oxidation of methane (AOM) is an important process in the carbon
cycle of marine environments and represents a major sink of the greenhouse gas methane
(Reeburgh, 1996). AOM has been documented in diffusive sedimentary environments
(Martens and Berner, 1974; Reeburgh, 1980; Iversen and Jørgensen, 1985) and at
advection-dominated cold seeps (Elvert et al., 1999; Hinrichs et al., 1999; Boetius et al.,
2000; Pancost et al., 2000; Michaelis et al, 2002). Cold seeps are broadly distributed
along active (Suess et al., 1985, 1998; Yun et al., 1999) and passive margins (Paull et al.,
1995) and are characterized by fluids expelled from deeper reservoirs which have high
contents of methane or other hydrocarbon gases. Furthermore, some hydrothermal vents
with high methane fluxes, such as the ones found in the Guaymas Basin, are also
supporting methanotrophic communities (Teske et al., 2002).
AOM is mediated by a syntrophic consortium of archaea and sulfate reducing
bacteria (SRB) (Boetius et al., 2000; Hinrichs et al., 2000; Orphan et al., 2001a) and
subsequent studies have shown evidence of at least three phylogenetically distinct
clusters of Euryarchaeota involved in the process. Two clusters of ANerobic
MEthanotrophs (i.e., ANME-1 and ANME-2) have been observed in close association
with SRB from the Desulfosarcina-Desulfococcus branch (DSS) (Hinrichs et al., 1999;
Boetius et al., 2000; Michaelis et al., 2002; Knittel et al., 2003), although ANME-1 has
also been frequently observed as monospecific aggregates or even single cells (Orphan et
al., 2002). Finally, members of the third cluster (i.e., ANME-3) occur together with
Desulfobulbus spp (DBB) (Niemann et al., 2006; Lösekann et al., 2007).

Chapter III ________________________________________________________________________
66
Several studies have provided evidence of diagnostic lipid biomarkers associated
with AOM-communities at hydrocarbon seeps (e.g., Elvert et al., 1999; Hinrichs et al.,
1999; Hinrichs et al., 2000; Pancost et al., 2000; Thiel et al., 2001; Michaelis et al., 2002;
Blumenberg et al., 2004; Elvert et al., 2005; Niemann et al., 2006; Rossel et al., 2008).
Distinct biomarker patterns have been attributed to ANME-1 and ANME-2 communities
(Blumenberg et al., 2004) with ANME-1 being characterized by a high abundance of
glyceroldialkylglyceroltetraethers (GDGTs), the absence of crocetane and a low
concentration of hydroxyarchaeol (OH-AR). In contrast, the main features of ANME-2
dominated communities are high amounts of crocetane and OH-AR, whereas GDGTs are
absent. ANME-3, on the other hand, are characterized by the presence of OH-AR,
polyunsaturated 2,6,10,15,19-pentamethylicosanes (PMI:4 and PMI:5), and the absence
of crocetane and GDGTs (Niemann et al., 2006).
Recently, the analysis of intact polar membrane lipids (IPLs) by high-performance
liquid chromatography/electrospray ionization mass spectrometry (HPLC-ESI-MS) has
largely extended the analytic window of lipid diversity in biogeochemistry and microbial
ecology (Rütters et al., 2002; Sturt et al., 2004). The IPL approach is based on ESI – ion
trap - MSn analysis that allows the simultaneous detection of characteristic IPLs from all
domains of life in a single analysis (Sturt et al., 2004; Ertefai et al., 2008; Rossel et al.,
2008). IPLs are reactive biomarkers considered to be indicative of living biomass (White
et al., 1979; Sturt et al., 2004; Biddle et al., 2006; Lipp et al., 2008) and sufficiently
specific for taxonomic distinction of the various ANME groups and their associated
bacterial partners (Rossel et al., 2008). ANME-1 is characterized by a high abundance of
diglycosyl-GDGT (2Gly-GDGT), while ANME-2 and ANME-3 produce mainly
archaeol-based IPLs, either with glycosidic and phospho headgroups or only phospho
headgroups, respectively. SRB members of AOM communities are characterized by
phosphatidylethanolamine (PE) and phosphatidyl-(N,N)-dimethylethanolamine (PDME)
headgroups, with the former occurring as dietherglycerol (DEG) phospholipids in
ANME-1/DSS dominated communities and as diacyl (DAG) or mixed acyl/ether (AEG)
phospholipids in ANME-2/DSS and ANME-3/DBB environments. The occurrence of
PDME is one distinctive feature of the bacteria associated with ANME-3. Relative
amounts of archaeal vs. bacterial IPLs differ systematically among the three major

Chapter III ________________________________________________________________________
67
community types, with the bacterial IPL contribution in ANME-1/DSS communities
being notably small (Rossel et al., 2008).
Most AOM studies focused on the diversity of community types but neglected the
importance of environmental factors selecting for each of these communities. Based on
field observations, it has been suggested that ANME-1/DSS dominate subsurface
sediments (Knittel et al., 2005) and microbial reef structures (Michaelis et al., 2002),
whereas ANME-2/DSS occurs in surface sediments above dissociating methane hydrates
(Elvert et al., 2005; Knittel et al., 2005) or in settings with high methane flux regimes
(Blumenberg et al., 2004) and ANME-3/DBB at mud volcanoes (Niemann et al., 2006;
Lösekann et al., 2007; Omoregie et al., 2008). Furthermore, based on results from in vitro
experiments, it has been suggested that ANME-2/DSS, contrary to ANME-1/DSS, is
better adapted to cold temperatures (Nauhaus et al., 2005). The influence of salinity and
pH on AOM activity has been evaluated but these factors seem not to be important
(Nauhaus et al., 2005). To better constrain the environmental factors influencing this
distribution we targeted AOM communities in a geographically diverse range of
hydrocarbon seeps. We performed the first comprehensive study of IPLs of AOM
communities, supplemented by a framework of community-related data (molecular
ecology and apolar lipid biomarkers) and information on geochemical conditions (e.g.,
concentrations of methane and sulfate, pH, salinity, etc.). IPL analysis provides a holistic
molecular view that integrates signals of all major microbial community members that
contribute substantially to the bulk living biomass. By contrast, other commonly used
culture-independent techniques such as fluorescence in situ hybridization (FISH) and
catalyzed reporter deposition-FISH (CARD-FISH) are highly selective and provide
information only on targeted organisms. Additionally, FISH techniques do not provide
information on the physiological status of microbes in relation to the environment
(Wagner et al., 2003). IPLs, on the other hand, may reflect environmental characteristics
because the structural composition of lipid membranes is influenced by growth
temperature (Khuller and Goldfine, 1974; Oliver and Colwell, 1973; Shimada et al.,
2008), pH (Minnikin and Abdolrahhimzadeh, 1974) and nutrient limitation (Van Mooy et
al., 2006). Therefore, our IPL analyses from globally distributed hydrocarbon seeps
Chapter III ________________________________________________________________________
68
represent a unique opportunity to evaluate both the distribution and the composition of
AOM communities and their relationship to the environmental conditions.
MATERIAL AND METHODS
Sample description
Table III.1. Samples analyzed in this study. Sampling location and the dominant AOM-phylotypes are indicated. Sample codes are based on the location, type of sample (sediment or mat) and field characteristics. Multiple samples of the same type are numbered sequentially.
Location Sample name Dominant
AOM community Lat. Long.
Water depth
(m) Station Research
Cruise
Arabian Sea (AS): AS-S-SOB orange unknown 24°54'N 63°01'E 551 GeoB12320 PC45, 2-3 cm,
Makran subduction zone
M74-3, 2007
AS-S-Thio unknown 24°51'N 63°01'E 1038 GeoB12313 PC4, 2-3 cm Makran subduction zone
M74-3, 2007
AS-S-Calyp unknown 24°51'N 63°01'E 1038 GeoB12313 PC15, 2-3 cm Makran subduction zone
M74-3, 2007
Black Sea (BS): BS-M-trunk-1a ANME-1/DSS [1] 44°47'N 31°59'E 189 P795, Dniepr area PO 317/3,
2004 BS-M-nodule-1a ANME-1/-2 mixed [1] 44°47'N 31°59'E 190 P822, Dniepr area PO 317/3,
2004 BS-M-interior ANME-1/DSS [2] 44°47'N 31°59'E 190 P822, Dniepr area PO 317/3,
2004 BS-M-trunk-2 ANME-1/DSS [2] 44°47'N 31°59'E 190 P822, Dniepr area PO 317/3,
2004 BS-M-trunk-3 unknown 44°01'N 36°41'E 2004 346, Shatsky Ridge R/V
Logachev TTR-15,
2005 BS-M-nodule-2 ANME-2a/DSS [2] 44°51'N 30°28'E 370 P780, Danube area PO 317/3,
2004 BS-M-nodule-3 ANME-2a/DSS [2] 43°57'N 30°17'E 295 P784, Danube area PO 317/3,
2004 BS-S unknown 44°48'N 31°55'E 235 Station 112, 0-2cm,
Crimean area R/V
Logachev TTR-11,
2001 Eastern
Mediterranean Sea (EMS):
EMS-S-SOB unknown 32°32'N 030°21’E 1698 770, PC 44, 2-4 cm Nile Delta
M70-2, 2006
Eel River Basin (ER):
ER-S-SOB ANME-1 [3] 40°48'N 124°36'W 500-520
PC 45, 3-6 cm Northern California continental slope
R/V Melville,
1998 Guaymas Basin
(GB):
GB-S-SOB orange ANME-1 [4] 27°1'N 111°24'W 2000 Core A, 0-2 cm Gulf of California
R/V Atlantis,
1998

Chapter III ________________________________________________________________________
69
Gullfaks oil field (GF):
GF-S-SOB white ANME-2a/2c mixed/DSS [5]
61°10'N 02°14’E 150 766, 0-10 cm North Sea
HE208, 2004
Gulf of Mexico (GOM):
GOM-S-SOB white ANME-1/DSS [6] 27°33'N 90°59'W 950 161, 0-10 cm Northern Gulf of Mexico
SO174, 2003
GOM-S-Campeche knolls
ANME-1/DSS [6] 21°54'N 93°26'W 2902 140, 8-10 cm Southern Gulf of Mexico
SO174, 2003
Håkon Mosby Mud Volcano
(HMMV):
HMMV-S-Beg-1a ANME-3/DBB [7] 72°00’N 14°44'E 1250 ATL 19, 1-2 cm South West Barents Sea
shelf
ATL, 2003
HMMV-S-Beg-2 ANME-3/DBB [7] 72°00’N 14°44'E 1250 Station 322, 0-2 cm South West Barents Sea
shelf
PS64, 2003
HMMV-S-Beg-3 ANME-3/DBB [7] 72°00’N 14°44'E 1250 Station 317, 0-1 cm South West Barents Sea
shelf
PS64, 2003
HMMV-S-Beg-4 ANME-3/DBB [7] 72°00’N 14°44'E 1250 Station 317, 2-3 cm South West Barents Sea
shelf
PS64, 2003
Hydrate Ridge (HR):
HR-S-Beg-1a ANME-2a/DSS [8, 9] 44°34'N 125°09'W 777 Station 19-2, 2-3 cm
Cascadia Margin SO148-1,
2000 HR-S-Beg-2 unknown 44°34'N 125°09'W 777 Station 19-2, 8-10 cm
Cascadia Margin SO148-1,
2000 HR-S-Beg-3 unknown 44°34'N 125°09'W 777 Station 165, 0-3 cm
Cascadia Margin SO165-2,
2002 HR-S-Calyp-1b ANME-2c/DSS [8, 9] 44°34'N 125°09'W 787 Station 38, 2-6 cm
Cascadia Margin SO148-1,
2000 HR-S-Calyp-2b ANME-2c/DSS [10] 44°34'N 125°09'W 787 Station 44D, 4-6 cm
Cascadia Margin SO148-1,
2000 HR-S-Calyp-3 unknown 44°34'N 125°09'W 787 Station 44D, 16-19 cm
Cascadia Margin SO148-1,
2000 a Samples previously reported by Rossel et al. (2008): BS-M-trunk-1(Black Sea mat 795), BS-M-nodule-1(Black Sea mat 822 reef top), HR-S-Beg-1(HR sediment station 19-2, 2-3 cm) and HMMV-S-Beg-1(HMMV sediment station 19, 1-2cm). b Phylogenetic information of HR-S-Calyp-2 indicates that this sample contains 80% of ANME-2c/DSS aggregates (2E10 of total cells from which 7E9 are archaeal cells) and an average of 15% of single ANME-1 cells (6.5E9 total cells), with cell diameters of 0.5nm and 0.6nm for ANME-2 and ANME-1, respectively (Knittel et al., 2003, 2005). Based on the cell shapes (coccus vs. rods) we calculate 1.7 and 10.3 fg of lipid for ANME-2 and ANME-1 cells, respectively (Lipp et al., 2008), which suggest that 60% of the lipids in this sample are associated with ANME-2c and 40% to ANME-1. The same estimate is probably valid for HR-S-Calyp-2, a sample collected in parallel to HR-S-Calyp-1 (Elvert et al., 2005). Abbreviations: Beg = Beggiatoa; Calyp = Calyptogena, SOB = sulfide oxidizing bacteria, Thio = Thioploca observed at the surface sediment, M = mat, S = sediment. References: [1] Rossel et al., 2008; [2] Arnds et al., unpublished data; [3] Orphan et al., 2002; [4] Teske et al., 2002; [5] Wegener et al., 2008; [6] Orcutt PhD thesis 2007; [7] Lösekann et al., 2007; [8] Knittel et al., 2003; [9] Knittel et al., 2005 and [10] Elvert et al., 2005.
The survey of IPLs associated with AOM communities included a broad range of
methane-rich sediments from nine major hydrocarbon seep settings: Arabian Sea, Black
Sea, Eastern Mediterranean Sea, Eel River Basin, Guaymas Basin, Gulf of Mexico,
Gullfaks oil field, Håkon Mosby Mud Volcano, and Hydrate Ridge (Fig. III.4,

Chapter III ________________________________________________________________________
70
supplementary material). IPL analyses were performed at locations at which AOM and
the community members had been previously reported as well as at sites for which no
prior taxonomic characterization was available (see ref.s in Table III.1).
AOM community composition
The taxonomic identification of the different community types studied here
(ANME-1/DSS, ANME-2/DSS and ANME-3/DBB) was based on culture independent
techniques such as 16S ribosomal RNA clone libraries, FISH and CARD-FISH analyses
as described elsewhere (see ref.s in Table III.1).
Lipid analysis
Samples were extracted using an automated microwave assisted extraction system
(MARS-X, CEM, USA) following a modified Bligh and Dyer protocol (Sturt et al., 2004)
except for HR-S-Beg-1, GB-S-SOB orange and ER-S-SOB samples, which were
extracted via ultrasonication at Woods Hole Oceanographic Institution (Teske et al.,
2002; Orphan et al., 2002; Sturt et al., 2004). IPLs were analyzed in total lipid extracts
(TLEs) for the majority of samples, except samples HR-S-Beg-1, GOM-S-SOB white,
GOM-S-Campeche knolls, GB-S-SOB orange and ER-S-SOB, from which the polar
fractions were analyzed after liquid chromatographic separation of the TLE on silica
(White et al., 1998). The utilization of different extraction and separation methods were
tested in some of the samples providing no difference in the lipid distribution (data not
shown). IPL analyses were performed on a HPLC–ESI–MSn system with the instrumental
parameters described previously (Sturt et al., 2004; Biddle et al., 2006). IPL
identification was based on mass spectral interpretation (cf. Sturt et al., 2004; Ertefai et
al., 2008; Rossel et al., 2008; Schubotz et al., submitted) and by comparison with IPL
inventories of cultures of different archaea and bacteria (e.g., Koga et al., 1998; Koga and
Morii, 2005; Hinrichs et al., unpublished data). Due to the limited availability of
commercial standards for the accurate determination of absolute concentrations, IPL
diversity was evaluated based on their relative abundances (cf. Rossel et al., 2008) under
the assumption of uniform response factors. While this procedure is inadequate to
accurately reflect the “real” relative IPL distribution, it is suitable for the differentiation

Chapter III ________________________________________________________________________
71
of a large set of samples. Additionally, unidentified IPLs that were present in at least
three samples and with relative concentrations of more than 2% in at least one sample
were included in the data set. These criteria were used to avoid the inclusion of rare IPLs
signatures potentially not related to methanotrophic habitats and those with unclear
molecular structure. The IPL inventories of the samples HR-S-Beg-1, HMMV-S-Beg-1,
BS-M-trunk-1, and BS-M-nodule-1 were previously reported (Rossel et al., 2008) and
have now been complemented by a few additional compounds such as
phosphatidylglycerol-GDGTs, ornithine (OL) and betaine (BL) lipids due to recent
progress in IPL identification.
In this study we integrated data from apolar lipid biomarkers associated with
AOM, that is, degradation products of IPLs formed either in the sediment or during
sample manipulation and analysis (e.g., free fatty acids, the archaeal core lipids archaeol
(AR) and OH-AR and bacterial glycerol-ether lipids) as well as hydrocarbons (crocetane,
PMI, and their unsaturated derivatives). We presumed that most compounds in this pool
have longer turnover times than IPLs and are thus likely to integrate longer episodes in
the evolution of the respective seep ecosystem. Therefore, the inclusion of these data
may provide additional clues on intrinsic properties of a seep system.
Some of the respective data were acquired in previous studies focusing on the
distribution of apolar lipids (Elvert et al., 2005; Niemann et al., 2006; Wegener et al.,
2008). For microbial mats from the Black Sea, sediments from the Arabian Sea, and
sample HMMV-S-Beg-2, apolar lipids and IPLs of the same TLEs were analyzed.
Chromatographic separation, identification, and quantification were performed according
to previously reported methods (Hinrichs et al., 2000; Elvert et al., 2005).
Environmental data
Biogeochemical parameters from all locations are summarized in Table III.2.
Environmental data were mainly represented by variables associated with regional scale
characteristics (fluid flow, temperature, salinity as well as bottom water oxygen and
phosphate concentration), while other variables focused on small-scale variations at the
respective sampling location (total organic carbon, sulfate reduction rate,

Chapter III ______________________________________________________________________
72
Table III.2. Environmental data selected for redundancy analysis (RDA).
Oxygen
(μM) SRR
(μmol cm-3 d-1) TOC
(wt%) Methane
(mM) Sulfate (mM) Temp (°C) pH
Arabian Sea (AS):
AS-S-SOB orange 15 [1] <0.1 [2] 2.0 [3, 4] 1.25 [5] 33 [2] 12.5 [1] 7.4 [2]
AS-S-SOB Thio 15 [1] <0.1 [2] 2.0.[3, 4] 0.08 [5] 32 [2] 8.0 [1] 7.4 [2]
AS-S-SOB Calyp 15 [1] <0.1 [2] 2.0 [3, 4] 0.08 [5] 32 [2] 8.0 [1] 7.4 [2]
Black Sea (BS):
BS-M-trunk-1 <10 [6] 39.6 [7]* 35 [8] 3.8 [7, 9] 3.0 [9, 10] 8.5 [11] 7.7 [12]
BS-M-nodule-1 <10 [6] 113 [7]* 35 [8] 3.8 [7, 9] 3.0 [9, 10] 8.5 [11] 7.7 [12]
BS-M-interior <10 [6] 36 [7]* 35 [8] 3.8 [7, 9] 3.0 [9, 10] 8.5 [11] 7.7 [12]
BS-M-trunk-2 <10 [6] 39.6 [7]* 35 [8] 3.8 [7, 9] 3.0 [9, 10] 8.5 [11] 7.7 [12]
BS-M-trunk-3 <10 [6] 39.6 [7]* 35 [8] 3.8 [7, 9] 3.0 [9, 10] 8.5 [11] 7.7 [12]
BS-M-nodule-2 <10 [6] 113 [7]* 15 [8] 3.8 [7, 9] 3.0 [9, 10] 8.5 [11] 7.7 [12]
BS-M-nodule-3 <10 [6] 113 [7]* 15 [8] 3.8 [7, 9] 3.0 [9, 10] 8.5 [11] 7.7 [12]
BS-S <10 [6] <0.1 [13] 3.2 [14] 0.1 [15] 18 [13, 15] 8.5 [11] 7.5 [9]
Eastern Mediterranean Sea (EMS):
EMS-S-SOB 200 [16] 0.2 [17, 18] 0.6 [17] 0.3 [17, 19] 24 [18] 14 [17] 8.0 [19]
Eel River Basin (ER):
ER-S-SOB 90 [20] <0.1 [21] 1.0 [20] 0.16 [22] 12 [22] 5.5 [20] 8.4 [23]
Guaymas Basin (GB):
GB-S-SOB orange 28 [24] 0.3 [24] 3.3 [25] 14 [26] 26 [24] 15 [27] 7.5 [28]
Gullfaks oil field (GF):
GF-S-SOB white 275 [29] 0.5 [30]* 0.2 [30] 25 [30] 28 [30] 8.0 [31] 7.4 [32]
Gulf of Mexico (GOM):
GOM-S-SOB-white 200 [33] 0.6 [34] 11 [34] 38 [34] 14 [34] 8.0 [34] 8.0 [35]
GOM-S-Campeche knolls 200 [33] <0.1 [34] 13 [34] 1.5 [34] 3.0 [34] 8.0 [34] 8.0 [35] Haakon Mosby Mud Volcano
(HMMV):
HMMV-S-Beg-1 300 [36] 0.6 [37] 1.0 [38] 2.5 [37] 14 [39] -1.0 [36,39] 8.0 [36]
HMMV-S-Beg-2 300 [36] 1.5 [37] 1.0 [38] 2.5 [37] 19 [37] -1.0 [36,39] 8.0 [36]
HMMV-S-Beg-3 300 [36] 0.5 [37] 1.0 [38] 2.5 [37] 15 [37] -1.0 [36,39] 8.0 [36]
HMMV-S-Beg-4 300 [36] 0.2 [37] 1.0 [38] 2.5 [37] 11 [37] -1.0 [36, 39] 8.0 [36]
Hydrate Ridge (HR):
HR-S-Beg-1 70 [40] 1.0 [41, 42] 2.6 [41] 50 [43, 44] 16 [42, 43] 3.0 [43] 8.3 [11]
HR-S-Beg-2 70 [40] 0.4 [41, 42] 1.6 [41] 50 [43, 44] 2.0 [42] 3.0[43] 8.3 [11]
HR-S-Beg-3 70 [40] 1.0 [42] 2.2 [41] 50 [43, 44] 16 [42, 43] 3.0 [43] 8.3 [11]
HR-S-Calyp-1 70 [40] 1.2 [41] 1.9 [41] 5.5 [43, 44] 18 [42] 3.0 [43] 8.3 [11]
HR-S-Calyp-2 70 [40] 1.2 [41] 1.8 [41] 5.5 [43, 44] 18 [42] 3.0 [43] 8.3 [11]
HR-S-Calyp-3 70 [40] 0.3 [45] 1.6 [41] 5.5 [43, 44] 2.0 [42] 3.0 [43] 8.3 [11] *SRR from the Black Sea mats and Gullfaks sediments were transformed from μmol gdw-3 d-1 to μmol cm-3 d-1 considering 0.12 gdw and 1.2 gdw for 1cm3 mat and sediment, respectively. References: [1] Bohrmann and cruise participants, 2008; [2] Schmaljohann et al., 2001; [3] Cowie et al., 1999; [4] Grandel et al., 2000; [5] Yoshinaga unpublished data; [6] Shaffer, 1986; [7] Arnds et al., unpublished data; [8] Roberts et al., 2008; [9] Krüger et al., 2008; [10] Treude et al., 2005; [11] Nauhaus et al., 2004; [12] Lichtschlag, Wenzhöfer, DeBeer unpublished data [13] Jørgensen et al., 2001; [14] Wakeham et al., 1995; [15] Knab PhD thesis 2007; [16] Yilmaz and Tugrul 1998; [17] Omoregie et al, 2008; [18] Felden unpublished data; [19] Heijs et al., 2007; [20] Levin et al., 2003; [21] Ziebis and Haese, 2005; [22] Orphan et al., 2004; [23] Day MSc dissertation 2003; [24] Weber and Jørgensen 2002; [25] Schouten et al., 2003; [26] Teske et al., 2002; [27] Jorgensen et al., 1992; [28] Gieskes et al., 1982; [29] Lohse et al., 1996; [30] Wegener et al., 2008; [31] Shovitri MSc dissertation 2007; [32] Dando et al., 1994; [33] Yan et al., 2006; [34] Orcutt PhD thesis 2007; [35] Aharon and Fu, 2000; [36] De Beer et al., 2006; [37] Niemann PhD thesis 2005; [38] Milkov et al., 2004; [39] Niemann et al., 2006; [40] Suess et al.,1999; [41] Elvert et al., 2005; [42] Treude et al., 2003; [43] Knittel et al., 2005 [44] Torres et al., 2002; [45] Boetius and Suess 2004.

Chapter III ______________________________________________________________________
73
pH, and concentrations of methane, sulfate and sulfide). Only the environmental
variables that explained the IPL variability according to the redundancy analysis (RDA)
are presented in Table III.2.
Statistical analyses
IPL patterns were subjected to a Hellinger transformation prior to applying
linear multivariate methods (Legendre and Gallagher, 2001). Principal component
analysis (PCA) was performed with a focus on inter-species distances and principal
axes were calculated for samples with available molecular characterization (Table
III.1). The remaining samples, whose ANME community types were unknown, were
then projected as passive samples in the ordination plot by using their IPL patterns. In
order to relate variation in IPL patterns to variation in contextual parameters, RDA was
performed on quantitative variables that were standardized to unit variance and zero
mean, and qualitative variables (i.e., fluid flow, sample type [sediment vs. mat]) were
converted to dummy variables (Ramette, 2007). A forward selection procedure was
performed to retain only the spatial terms that significantly explained variation in the
lipid data. The selected terms were then analyzed in concert with the other contextual
parameters. Significances in the RDA models were assessed by 1000 data permutations
using CANOCO (version 4.5. Microcomputer Power, Ithaca, NY).
The overall distribution and total variability of lipids in the different settings
was evaluated first by the relative abundance of different IPL types (Fig. III.1)
combined with three PCA. In the first PCA (Fig. III.2), archaeal IPLs were represented
as full molecules, whereas bacterial IPLs were distinguished by the headgroup and
bond type between the alkyl moieties and the glycerol backbone (DEG or DAG; AEG
was not possible to distinguish, therefore alkyl chains are given as DAG). This
approach used for bacterial IPLs intends to avoid underestimation of bacterial IPLs,
which would occur from the separation of each IPL depending on the variability in the
fatty acid chains (e.g., PE-DAG has 25 different fatty acid combinations represented by
diverse chain lengths and degrees of unsaturation). Furthermore, betaine lipids (BL)
were separated in two groups according to the presence of odd (BL-odd) and even (BL-

Chapter III ______________________________________________________________________
74
even) fatty acid chains in order to evaluate the possible contribution of specific sources
distinct from algae (Schubotz et al., submitted).
In the second PCA (Fig. III.5, supplementary material), possible differences in
the side chain distribution of fatty acids in bacterial IPLs were evaluated. Four samples
were excluded in this analysis because no bacterial IPLs were detected: HR-S-Calyp-1
to -3 and GOM-S-Campeche knolls. Similarly, the overall distribution of apolar lipids
was evaluated by a third PCA (Fig. III.6, supplementary material).
RESULTS AND DISCUSSION
Diversity of IPLs at hydrocarbon seeps
A total of 46 different IPLs (35 known, nine with tentative names and two
unknowns) were evaluated in detail (Table III.3). 34 IPLs (25 known and nine
tentatively identified) were assigned to archaeal sources (Arabian numbers) and 10 to
bacterial sources (Roman numbers, except VIII which is derived from aquatic algae).
Additionally, two unknown IPLs (a and b) were likely derived from bacteria and
archaea inhabiting carbonate reefs and sediments, respectively. These assignments
were based on characteristics in the mass spectra, which indicate the presence of a
series of acyl moieties in compound a and a lipid structure analogical to a glycosidic-
AR in compound b (Table III.3). Archaeal IPL diversity included several glycosidic-
GDGTs (IPLs # 1 to 7) and glycosidic-ARs (IPLs # 18 to 22) as well as phospho-
GDGTs (IPLs # 8 to 17) and phospho-ARs (IPLs # 23 to 34) (Table III.3).

Chapter III ______________________________________________________________________
75
Table III.3. Lipid code and source assignment of IPLs detected in this study. Lipid Code Lipid name Source assignement
1 2Gly-GDGT Archaea, ANME-1 (Rossel et al., 2008) and deep subsurface (Biddle et al., 2006; Lipp et al., 2008; Sturt et al., 2004), Sulfolobus shibatae (Sturt et al., 2004); Methanobacterium thermoautotrophicum (Koga et al., 1993).
2 3Gly-GDGT Archaea, ANME-1 (this study).
3 4Gly-GDGT Archaea, ANME-1 (this study).
4 2Gly-GDGT+14 Archaea, ANME-1 (this study) and in deep subsurface (Fredricks and Hinrichs, 2007).
5 2Gly-GDGT+18 Archaea, ANME-1(this study) and in deep biosphere sediments (Lipp and Hinrichs, unpublished data) and Nitrosopumilus maritimus (Schouten et al., 2008).
6 2Gly-GDGT+28 Archaea, ANME-1 (this study).
7 2Gly-GDGT+145 Archaea, ANME-1 (this study).
8 Tentative 2Gly-GDGT-PE
Archaea, ANME-1 (this study), Methanobacterium thermoautotrophicum (Koga et al., 1993).
9 MAPT-GDGT-PG Archaea, ANME-1 (this study), aminopentatetrol -GDGTs with two and three methyl groups on the amino group have been describe in Methanomicrobiales (Koga and Morii, 2005; Koga and Nakano, 2008).
10 Gly-GDGT-PG Archaea, ANME-1 (this study).
11 2Gly-GDGT-PG Archaea, ANME-1 (this study), Methanospirillum hungatei (Koga et al., 1993).
12 PG-GDGT Archaea, ANME-1 (this study).
13 2PG-GDGT Archaea, ANME-1 (this study).
14 Tentative PE-GDGT-PG Archaea, ANME-1 (this study).
15 Tentative APT-GDGT-PG
Archaea, ANME-1 (this study), aminopentatetrol-GDGTs without methyl group on the amino group have been describe in Methanomicrobiales (Koga and Morii, 2005; Koga and Nakano, 2008).
16 Tentative APT-GDGT-238
Archaea, ANME-1 (this study), aminopentatetrol-GDGTs without methyl group on the amino group have been describe in Methanomicrobiales (Koga and Morii, 2005; Koga and Nakano, 2008).
17 Tentative 2P-GDGT+155 Archaea, ANME-1 (this study).
18 Gly-MAR Archaea, ANME-2 (this study), Methanocaldococcus jannaschii (Sturt et al., 2004).
19 2Gly-AR Archaea, ANME-2 (Rossel et al., 2008; this study), Methanocaldococcus jannaschii (Sturt et al., 2004), deep subsurface (Biddle et al., 2006, Fredricks and Hinrichs, 2007, Lipp et al., 2008).
20 Gly-OH-AR Archaea, ANME-2 (this study); Methanothrix soehngenii (koga et al., 1993).
21 2Gly-OH-AR Archaea, ANME-2 (Rossel et al., 2008; this study), Methanothrix soehngenii (koga et al., 1993).
22 2Gly-MAR Archaea, ANME-2 (this study), Methanocaldococcus jannaschii (Koga et al., 1993; Sturt et al., 2004).

Chapter III ______________________________________________________________________
76
23 PE-OH-AR Archaea, ANME-2 (Rossel et al., 2008; this study), Methanothrix soehngenii (koga et al., 1993), Methanosarcina barkeri (Koga and Morii et al., 2005).
24 PG-AR Archaea, ANME-2 (Rossel et al., 2008; this study).
25 PG-OH-AR Archaea, ANME-2 (Rossel et al., 2008; this study), Methanosarcina barkeri (Koga and Morii et al., 2005), Halophiles (Kates, 1978).
26 Tentative APT-OH-AR Archaea, ANME-2 (this study).
27 PI-OH-AR Archaea, ANME-2 and ANME-3 (Rossel et al., 2008; this study).
28 PS-AR Archaea, ANME-2 and ANME-3 (Rossel et al., 2008; this study), Methanobacterium thermoautotrophicum (Koga et al., 1993), Methanocaldococcus jannaschii (Sturt et al., 2004).
29 PS-OH-AR Archaea, ANME-2 and ANME-3 (Rossel et al., 2008; this study), Methanosarcina barkeri (Koga et al., 1993).
30 PS-2OH-AR Archaea, ANME-2 and ANME-3 (Rossel et al., 2008; this study).
31 Tentative P-AR+223 Archaea, ANME-2 (Rossel et al., 2008; this study).
32 Gly-PG-AR Archaea, ANME-1 (this study).
33 Tentative Gly-PS-AR Archaea, possibly ANME-2 (this study).
34 Tentative Gly-P-OH-AR, extended
Archaea, possibly ANME-2 (this study), archaeols with C25 chain have been previously reported in extreme Halophiles (Koga et al., 1993; 2008) and in cold seep sediments from Eastern Mediterranean Sea (Stadnitskaia et al., 2008).
I PC-DAG* Methanotrophic bacteria (Makula, 1978; Goldfine, 1984; Fang et al., 2000), photosynthethic bacteria and green algae (Imhoff and Bias-Imhoff, 1995; Thompson, 1996).
II PG-DAG*
Methanotrophic bacteria (Makula, 1978; Goldfine, 1984; Fang et al., 2000), Desulfosarcina variabilis (Rütters et al., 2001; Sturt et al., 2004), photosynthethic bacteria and green algae (Imhoff and Bias-Imhoff, 1995; Thompson, 1996).
III PE -DAG* Methanotrophic bacteria (Makula, 1978; Goldfine, 1984; Fang et al., 2000), Desulfosarcina variabilis (Rütters et al., 2001; Sturt et al., 2004).
IV PE-DEG Methanotrophic bacteria (Makula, 1978; Goldfine, 1984; Fang et al., 2000).
V PME-DAG* Methanotrophic bacteria (Makula, 1978; Goldfine, 1984; Fang et al., 2000), sulfide oxidizer (Barridge and Shively, 1968).
VI PDME-DAG* Methanotrophic bacteria (Makula, 1978; Goldfine, 1984; Fang et al., 2000), sulfide oxidizer (Barridge and Shively, 1968).
VII OL Bacteria gram-negative performing SR, sulfur and iron oxidation (Makula and Finerty, 1975; Knoche and Shively, 1972; Imhoff and Bias-Imhoff, 1995).
VIII BL-even Photosynthetic eukaryote (Sato, 1992; Dembitsky, 1996; Kato, 1996).
IX BL-Odd Bacteria from anoxic waters (Schubotz et al., submitted; this study).
X Surfactin Bacillus sp (Vater, 1986).

Chapter III ______________________________________________________________________
77
Unknown IPLs Distinctive features a Unknown a+ Retention time: 27 min (-0.73 Retention index relative to C-16 PAF)
m/z of quasi-molecular ion: 706.3, 734.3 neutral loss in ms2 positive mode: 194 and then 18
b Unknown b+ Retention time: 43-45 min(+1.3 Retention index relative to C-16 PAF) m/z of quasi-molecular ion: 1148.0 ms2 in positive mode yield the m/z fragments: 873, 993
* Distinction of AEG and DAG not possible under HPLC-MS conditions applied, alkyl chain in Fig. III.3 of supplementary material provided for DAG. + Unknown IPL a was solely detected in microbial mats from the Black Sea. The occurrence of this IPL was specifically observed in ANME-2a/DSS and in the mat displayed between ANME-1 and ANME-2a grouping (BS-M-interior). This unknown was characterized by two major quasi-molecular ions (706.3 and 734.3, Table III.3), both with daughter fragments ions indicative of a loss of 193. Negative ion mode information showed the presence of fatty acids C16:1 and C17:1, which suggest that these lipids are bacterial-derived. The unknown IPL b was displayed, although with a small arrow, between ANME-2a and ANME-2c dominated sediments. The occurrence of this lipid was higher in sediments dominated by ANME-2a/DSS (except in GOM-S-SOB white where it makes up to 21%). This IPL was characterized by one quasi-molecular ion (1148.1, Table III.3) which shows a loss of a 155 (which could indicate the presence of PME) followed by a loss 120 Da in MS2. Negative ion mode information was rather noisy and did not allow a clear identification of the molecule. However, the occurrence of the 993.4 fragment during MS2, caused by the loss of 155, is a possible indication of the presence of 2Gly-MAR. Abbreviations: APT = phosphoaminopentatetrol, AR = archaeol, BL = betaine lipids, 2Gly = diglycosyl, 3Gly = triglycosyl, 4Gly = tetraglycosyl, OH-AR = hydroxyarchaeol, 2OH-AR = dihydroxyarchaeol, GDGT = glyceroldialkylglyceroltetraether, MAPT = phospho methylaminopentatetrol, MAR = macrocyclic archaeol, OL = ornithine lipids, P = phospho headgroup, PC = phosphatidylcholine, PDME = phosphatidyl-(N,N)-dimethylethanolamine, PE = phosphatidylethanolamine, PG = phosphatidylglycerol, PI = phosphatidylinositol, PME = phosphatidyl-(N)-methylethanolamine, PS = phosphatidylserine.
In order to evaluate the general trend of IPLs, the samples were separated in two
groups using the criteria of a contribution to the total archaeal IPLs higher than 50% of
GDGT-based IPLs (Fig. III.1a) or AR-based IPLs (Fig. III.1b), with the former
considered to be diagnostic of ANME-1 and the latter of ANME-2 and ANME-3
dominated communities (Rossel et al., 2008). In the first group the glycosidic-GDGTs
(IPLs # 1 to 7, Table III.4, supplementary material), with 2Gly-GDGT being the
dominant IPL, contributed over 75% in the microbial mats from the Black Sea, whereas
in sediments the contribution varied between 57 and 100%. However, this general
trend of a high glycosidic-GDGTs content was not followed by three samples (HR-S-
Calyp-1 and HR-S-Calyp-2 and AS-S-SOB orange, the first two taxonomically
affiliated with ANME-2c, Table III.1), which contained less than 40% of glycosidic-
GDGTs but more than 56% of phospho-GDGTs, with 2Gly-GDGT-PG and 2PG-
GDGT being the most abundant IPLs. Glycosidic-GDGTs have been previously
reported in isolates of Sulfolobus shibatae (Sturt et al., 2004), whereas the presence of
GDGTs with only phospho or mixed phospho and glycosidic headgroups has been
documented in Methanobacterium thermoautotrophicum (Koga et al., 1993).

Chapter III ______________________________________________________________________
78
Fig. III.1. Grouping of samples according to the dominance of GDGT- (a) and AR-based IPLs (b), considering the former as diagnostic for ANME-1 and the latter for ANME-2 and ANME-3 communities. Major lipids (BL-even and the unknowns a and b were not included) were grouped according to IPL classes and normalized to archaeal (a and b) and bacterial (c and d) IPLs. Contribution of archaeal IPLs relative to the total are shown (e). Abbreviations: Gly = glycosidic headgroups, P = phospho headgroups (including mixed glycosidic and phospho headgroup), GDGT = glyceroldialkylglyceroltetraether, AR = archaeol-based IPLs (archaeol and hydroxyarchaeol), PE = phosphoethanolamine, DAG = diacyl, DEG = diether. Notice that some of the samples in the first group (c) did not contain any bacterial IPLs.
The second group of samples was strongly dominated by AR-based IPLs with
both phospho and glycosidic headgroups (Fig. III.1b), compounds that have been found
in cultures of methanogens such as Methanocaldococcus jannaschii (Koga et al., 1993;
Sturt et al., 2004). All samples from Håkon Mosby Mud Volcano and AS-S-Calyp
contained exclusively phospho-ARs, with PG-OH-AR and PS-OH-AR being the major
IPLs in the first setting and PI-OH-AR the dominant in the second one. The microbial
mats from the Black Sea, which contained low amounts of glycosidic-GDGTs (< 34%;

Chapter III ______________________________________________________________________
79
Table III.1), presented high abundance of glycosidic-ARs (up to 57%), with 2Gly-AR
and 2Gly-MAR (macrocyclic) being the major IPLs. Besides the generally high
contribution of phospho-ARs in the sediments (> 56%), in the samples HR-S-Beg-3
and EMS-S-SOB also high amounts of GDGTs (~40% in each sample) were observed.
The bacterial IPL distribution in the two groups of samples was variable. In the
samples dominated by GDGT-based IPLs the bacterial IPLs presented a low
contribution or even absence (less than 28%), whereas in the group of samples
dominated by AR-based IPLs the abundance was as high as 66% (Figs. III.1c, d and e,
Table III.4, supplementary material). The most abundant bacterial IPL was PE-DAG
with contributions between 18 and 100% (Figs. III.1c and d) but revealed no clear
pattern in relation to ANME community types or sample characteristics. In contrast,
PE-DEG was present in relatively higher amounts (up to 38%) in the microbial mats
from the Black Sea dominated by glycosidic-GDGTs (Fig. III.1c). Additionally, high
contribution of PME and PDME in samples from the Håkon Mosby Mud Volcano was
a distinctive feature of these sediments dominated by ANME-3/DBB. PE has been
previously reported to be the major phospholipid in SRB such as Desulfosarcina
variabilis (Rütters et al., 2001; Sturt et al., 2004) and its occurrence together with PME
and PDME in the anoxic water column and surface sediments from the Black Sea has
also been suggested to derive from SRB (Schubotz et al., submitted). Nevertheless,
PME and PDME are also produced by methanotrophic bacteria such as Methylosinas
trichosporium and Methylobacterium organophilum (Makula, 1978; Goldfine, 1984;
Fang et al., 2000), as well as by sulfide oxidizers (Barridge and Shively, 1968). It
should be noted that two of these samples from the Håkon Mosby Mud Volcano, which
included the uppermost sediment surface (HMMV-S-Beg-2 and -3, Table III.1),
contained very low amounts of archaeal IPLs (7 to 9%, Fig. III.1e). This suggest an
additional contribution to the bacterial IPLs from either aerobic methanotrophic
bacteria, which have been shown to be present in surface sediments in this habitat
(Lösekann et al., 2007; Elvert and Niemann, 2008) or from sulfide oxidizers, both of
which contain PME and PDME in their membranes (Barridge and Shively, 1968;
Makula, 1978; Fang et al., 2000). PC and PG, which have been observed in
methanotrophic bacteria (Makula, 1978; Golfine, 1984; Fang et al., 2000) as well as in

Chapter III ______________________________________________________________________
80
green algae (Thompson, 1996), were highly variable but generally higher in sediment
samples (up to 85%) dominated by AR-based IPLs. The bacterial-derived BL-odd and
OL with non-phospho headgroups were mainly present in the mat samples from the
Black Sea but were also found (contribution up to 37%) in AS-S-SOB orange sample
dominated by GDGTs (Fig III.1c). BL-odd is probably derived from bacteria as
suggested by the exclusive presence in deep anoxic waters of the Black Sea (Schubotz
et al., submitted), whereas OL have been previously observed in SRB (Makula and
Finerty, 1975), sulfur-oxidizing and iron-oxidizing bacteria (Knoche and Shively,
1972). It has been suggested that OL play a functional role in the iron oxidation
metabolism in Thiobacillus ferrooxidans (Ghosh and Misha, 1987). Interestingly,
Reitner et al., (2005) observed that DSS in ANME-2 dominated mats from the Black
Sea presented intracellular iron sulfide precipitates, which suggest an active iron cycle
in these mats. Surfactin, a common lipopeptide previously found in Bacillus sp. (Vater,
1986), was only present in three of the mat samples from the Black Sea (BS-M-trunk-3,
BS-M-nodule-1 and -2), two of them dominated by AR-based IPLs in which surfactin
was as high as 55%. Because previous clone libraries from microbial mats do not
provide evidence for the presence of Bacillus sp (Knittel et al., 2005), we suggest that
other unidentified bacteria are the main producers of this compound.
Patterns of IPL – sample associations indicated by PCA
In order to examine systematic relationships between lipid distributions, AOM
community type and sample characteristics, we examined the data set with three PCAs:
(1) all IPLs, (2) all bacterial IPLs with individual acyl and alkyl moieties distinguished,
and (3) apolar lipids. In the first PCA (Fig. III.2) the total number of IPLs was reduced
from 46 to 41 due to the removal of some IPLs with low variation and/or frequency
among the samples (IPLs # 2, 3, 8 were found in Black Sea at the Shatsky Ridge, IPL
#33 was present at Eastern Mediterranean Sea and IPLs # 2 and 26 were observed in
Arabian Sea at the orange and Thioploca sites, respectively). The remaining 41 IPLs
explained most of the data variability (59% based on two principal components).

Chapter III ______________________________________________________________________
81
Fig.III.2. PCA plot displaying the overall distribution of IPLs among the various samples analyzed. Samples are shown in color codes according to the phylogenetic affiliation. The mixture of ANME-2a and -2c is displayed with both orange and brown circles combined (GF-S-SOB white). Eigenvectors of archaeal, bacterial, algal and unknown IPLs are displayed in red, blue, green and black, respectively. Their direction and length represents the main behavior of the lipid and the rate of change in two dimensional space, respectively (Ramette, 2007). IPL names are given according to Table III.3. As an example, mats dominated by ANME-2a/DSS (dots with white cross) were characterized by the archaeal IPLs # 22, 21, 24 and bacterial IPLs IV, X, VII. Notice that for the construction of this plot archaeal IPLs were represented as full molecules, whereas bacterial IPLs were distinguished by the headgroup and bond type between the acyl/alkyl moieties and the glycerol backbone (DEG or DAG; AEG was not possible to be distinguished). This approach was used to avoid underestimation of bacterial IPLs. Additionally, betaine lipids (BL) were separated in two groups according to the presence of odd (BL-odd) and even (BL-even) fatty acid chains.
IPLs of ANME-1/DSS dominated systems
A distinct group was formed by sediments and microbial mats samples
dominated ANME-1/DSS (BS-M-trunk-1, BS-M-interior, BS-M-trunk-2, GOM-S-
Campeche knolls, ER-S-SOB, GB-S-SOB-orange, and GOM-S-SOB white; Fig. III.2,
black circles) and taxonomically uncharacterized samples (BS-M-trunk-3, HR-S-Calyp-

Chapter III ______________________________________________________________________
82
3, BS-S, and AS-S-SOB orange; Fig. III.2, grey circles). The main feature of the
samples from this group was the high contribution of 2Gly-GDGT, which corroborates
our earlier findings (Rossel et al., 2008). In addition to glycosidic-GDGTs, diverse
types of GDGTs with mixed glycosidic and phospho headgroups (Gly-GDGT-PG,
2Gly-GDGT-PG) and phospho headgroups (PG-GDGT, 2PG-GDGT) were observed.
Together with 2Gly-GDGTs, 2Gly-GDGT-PG, 2PG-GDGT, other GDGTs with
unknown headgroups were observed (Table III.3). Only two AR-based IPLs were
observed in the ANME-1 grouping, however, with less than 1% contribution: Gly-PG-
AR and a tentatively identified extended Gly-P-OH-AR. In the ANME-1 group, which
includes sediments and microbial mats, the contribution of bacterial IPLs was very low
between 0 and 7% (Table III.4, supplementary material), in agreement with our
previous observations in ANME-1/DSS dominated mats (Rossel et al., 2008).
However, we observed two sediment samples with higher contributions of bacterial
IPLs: one from the Eel River Basin (ER-S-SOB) and one from the Black Sea (BS-S)
which contained 28 and 14% of the total IPLs, respectively. The low contribution or
even absence of bacterial IPLs in ANME-1/DSS dominated sediments and microbial
mats is in agreement with the observation of Orphan et al. (2002), who reported that
ANME-1 frequently occurs as a monospecific aggregates or as single cells.
IPLs of ANME-2/DSS dominated systems
Samples dominated by ANME-2 were separated into three main groups (Fig.
III.2; brown and orange circles). The first group was represented by microbial mat
nodules observed on the outside of carbonate reefs from the Black Sea (BS-M-nodule-1
to -3, brown circles with a white cross), which are dominated by ANME-2a/DSS
(Arnds et al., unpublished data). Characteristic features of these nodules were IPLs
based on AR and OH-AR with both glycosidic and phospho headgroups (Gly-OH-AR,
Gly-MAR, 2Gly-AR, 2Gly-OH-AR, 2Gly-MAR, 2-Gly-OH-AR, tentative-P-AR, PG-
AR and PE-OH-AR). These archaeal IPLs were accompanied by the presence of
bacterial-derived PE-DAG, PE-DEG, OL, BL-odd and surfactin. The general presence
of PE-DEG in microbial mats, independent of ANME type, suggests that it may not be

Chapter III ______________________________________________________________________
83
derived from a bacterial partner that is exclusively associated with one particular
ANME community type.
The three mat nodules dominated by the ANME-2a/DSS group differed by the
higher contribution of phospho-ARs in the first nodule (42%, BS-M-nodule-1), whereas
in the samples BS-M-nodule-2 to -3 both phospho and glycosidic headgroups were
similarly abundant. Additionally, the high contribution of surfactin of up to 27% in
these two nodules (Table III.4, supplementary material) suggests that this compound
has a functional role in ANME-2a/DSS mats. Surfactin is a lipopeptide composed of a
hydrophilic part (seven amino acids) and hydrophobic tail (hydroxylated fatty acids
with 13, 14 or 15 carbon atoms), and it is the most efficient microbial biosurfactant
known (Vater, 1986). Surfactin is mainly produced during the maximum growth phase
of the bacterial cell cycle, and so far has been mainly observed in several Bacillus
subtilis strains (Vater, 1986). It has surface-, interface- and membrane-active
properties and has been shown to improve mechanisms of cell adhesion (Ahimou et al.,
2000). This suggests that surfactin may facilitate the formation of zones of ANME-
2a/DSS aggregates in the methanotrophic mats.
The second cluster linked to the ANME-2 group comprised sediment samples
dominated by the ANME-2a subgroup. Some of the samples have been taxonomically
characterized (HR-S-Beg-1, Knittel et al., 2005; GF-S-SOB white, Wegener et al 2008)
whereas others are uncharacterized (AS-S-Thio, AS-S-Calyp, HR-S-Beg-3, HR-S-Beg-
2 and EMS-S-SOB; grey circles). The main feature of this group was the high
abundance of phospho-ARs (PS-2OH-AR, PS-AR, PS-OH-AR, PG-OH-AR and PI-
OH-AR) and bacterial IPLs, (PE-DAG, PG-DAG, PC-DAG) and the presence of BL-
even, considered to originate from aquatic plants (cf. Ertefai et al., 2008; Table III.4,
supplementary material). The archaeal IPL pattern was strongly dominated by OH-AR,
while glycosidic-AR was not abundant in this group (< 3%). The IPL composition of
sample EMS-S-SOB from the Eastern Mediterranean Sea differed from the rest due to a
high abundance of phospho-GDGTs and the tentatively identified Gly-P-OH-AR
extended (22 and 16%, respectively; Table III.4, supplementary material), with the
latter being the intact counterpart of the apolar OH-AR previously reported in this
setting by Stadniskaia et al. (2008). The occurrence of phospho-GDGTs and Gly-P-

Chapter III ______________________________________________________________________
84
OH-AR suggest the presence of a mixed ANME-1 and ANME-2a community in this
setting.
The third group of samples linked to ANME-2 was represented by two
Calyptogena-influenced sediment samples dominated by ANME-2c (Knittel et al.,
2005), HR-S-Calyp-1 and HR-S-Calyp-2 (Fig. III.2, orange circles). This group is
distinguished from the previous ANME-2 groups due to the high contribution of
GDGTs with phospho and mixed phospho and glycosidic headgroups. The main
GDGTs were 2Gly-GDGT-PG and 2PG-GDGT, followed by the tentatively identified
IPLs PE-GDGT-PG, APT-GDGT-PG and MAPT-GDGT-PG and additional unknown
intact GDGTs (# 16 and 17, Table III.4, supplementary material). By contrast, the IPLs
typically associated with ANME-2 (PG-OH-AR, PI-OH-AR, PS-OH-AR) were less
abundant (Table III.4, supplementary material). The presence of GDGTs in ANME-2c
was previously suggested by Elvert et al. (2005), who reported a maximum of free
GDGTs with one and two cyclopentane rings in sample HR-S-Calyp-2, which was
characterized by maximum rates of sulfate reduction and high numbers of ANME-
2c/DSS aggregates. However, relative cellular abundance is not necessarily a reliable
predictor for the corresponding IPL ratios since cellular size and surface area of
ANME-2c and ANME-1 cells differ significantly. Even though the ANME-1 cell
concentration is relatively small, due to their significantly larger total surface size
compared to ANME-2c cells, the expected IPL concentration account for up to 40% of
total archaeal IPL (Table III.1). And notably, in the sample from Gullfaks (GF-S-SOB
white), which is a mixture of ANME-2a and ANME-2c cells (Wegener et al., 2008), we
did not detect any GDGT-based IPLs. This suggests that the GDGTs observed in the
samples HR-S-Calyp-1 and -2 could be associated with the presence of ANME-1 cells.
Nevertheless, the GDGT composition of the sample is significantly different to those
from other ANME-1 dominated systems, which is specifically expressed in the low
abundance of glycosidic-GDGTs (Fig. III.1a, Table III.4, supplementary material).
Therefore, we suggest that the GDGTs in the samples HR-S-Calyp-1 and -2 originate
from ANME-1 that, unlike at other settings, produce phospholipids, although we cannot
exclude that ANME-2c, unlike ANME-2a, has the capability to produce GDGTs.
However, the absence of GDGTs at Gullfaks combined with the dominance of ANME-

Chapter III ______________________________________________________________________
85
1b genes in the clone library of this sample (Knittel et al., 2005) rather supports the
former alternative. Furthermore, we found no indications of bacterial phospho-IPLs,
which is contrary to the finding of SRBs by Elvert et al. (2005) and Knittel et al.
(2005), moving this group into closer relation to ANME-1 rather than to ANME-2.
IPLs of ANME-3/DBB dominated systems
ANME-3 dominated samples (HMMV-S-Beg-1 to -4; green circles) were
closely clustered with the group of sediments characterized by ANME-2a. The
respective ANME-3 samples were characterized by very similar phospho-ARs as those
commonly associated with ANME-2a and by the absence of GDGTs and glycosidic-
ARs, consistent with previous observations (Rossel et al., 2008). However, ANME-3
dominated samples were distinguished by high contributions of bacterial PMEs and
PDMEs (Table III.4, supplementary material), thus providing a possible indication of
DBB species (Rossel et al., 2008). Nevertheless, due to lack of information about IPLs
from DBB isolates and the previously reported production of similar lipids by aerobic
methanotrophs and sulfide oxidizers (Barridge and Shively, 1968; Makula, 1978; Fang
et al., 2000), it is possible, especially in surface sediments, that a fraction of the lipid
contribution may derive from these bacteria. The low contribution of bacterial IPLs in
sediments and microbial mats dominated by ANME-1/DSS compared to ANME-
2a/DSS and -3/DBB suggests that the latter two communities inhabit environments
suitable for a wide variety of microbes.
Diversity of bacterial IPLs in each ANME system
The bacterial communities inhabiting AOM environments were evaluated in
more detail by a second PCA including not only the characteristic headgroups of
bacterial IPLs (PE, PME, PDME, PC, PG, OL and BL), but also variations in the acyl
and alkyl chains (chain length and saturation degree) (Fig. III.5, supplementary
material). The most striking feature of this PCA was the strong separation of the
ANME-3/DBB group from Håkon Mosby Mud Volcano, whereas the other
methanotrophic communities were dominantly separated due to sample characteristics
into mats vs. sediments. The ANME-3/DBB group was characterized by phospho-IPLs

Chapter III ______________________________________________________________________
86
with the DAG bond type (PE C32:3, PME C32:2, PDME C34:2, PDME C32:2, PDME C32:1,
PC C32:2) as well as OL and BL (OL C32:1, OL C32:2, BL C32:2 and BL C34:2). All of
these were positively correlated. The association of PDME with combined C32:2 and
C34:2 acyl moieties with ANME-3/DBB is in agreement with previous observations
(Rossel et al., 2008). These acyl moieties are consistent with combinations of the fatty
acids such as C16:1�5c and C17:1�6c associated with ANME-3/DBB, but also with the
fatty acid C16:1�8 attributed to aerobic methanotrophs observed in surface sediments
from the same location (Niemann et al., 2006).
Distribution of apolar lipids and their taxonomic significance
Strongly 13C-depleted apolar lipids such as crocetane, PMI of archaeal origin
(e.g., Elvert et al., 1999, Thiel et al., 1999), various fatty acids and mono- and di-O-
alkyl glycerol ethers, putatively produced by SRB and the derivatives of polar DEG and
AEG lipids (Hinrichs et al., 2000; Pancost et al., 2000; Elvert et al., 2003), AR and sn-
2-OH-AR (e.g., Hinrichs et al., 1999, 2000) as well as biphytanes obtained by ether-
cleavage reactions of free GDGTs (e.g., Pancost et al., 2001; Schouten et al., 2001;
Thiel et al., 2001) have been routinely used to identify AOM. These and other apolar
lipids were analyzed by PCA in order to identify taxonomic relationships with
individual ANME groups (Fig. III.6, Table III.5, supplementary material). The sample
set had to be reduced from 27 down to 17 due to the lack of available contextual data.
As evident in supplementary Fig. III.6, apolar lipids alone did not separate the
three dominating AOM communities. Interestingly, most of the selected lipids (i.e.,
2OH-AR, DAGE C30:0, C23:1, AR, Crocetane, PMI, PMI:4, Crocetane:1, FA ai-C15:0, FA
cyC17: 0�5,6, C31:x and DAGE C32:2a) were mainly associated with the samples previously
grouped with ANME-1/DSS and ANME-2c/DSS based on IPLs. In opposite direction
to the majority of the lipids, were sn-2-OH-AR and MAGE C16:1�5c, (frequently
observed in ANME-2/DSS dominated systems, e.g., Blumenberg et al., 2004; Elvert et
al., 2005), here related to microbial mats containing both ANME-1 and ANME-2
populations, thus not providing a clear separation. Similarly, no clear relationship was
identified between ANME-3/DBB dominated samples and apolar lipids.

Chapter III ______________________________________________________________________
87
Possible explanations for the poor taxonomic differentiation in this data set are
the lack of appropriate data on GDGT abundances, the major ANME-1 core lipid, and,
importantly, the longer turnover times of apolar lipids compared to IPLs. As a
corollary, the mismatch between these two lipid-based lines suggests that community
compositions are probably not uniform through the time interval integrated by a typical
sample (largely on the order of 100 to 1000 yrs), suggestive of community changes in
the course of the geological evolution of highly dynamic seep systems.
Discrepancies between FISH and IPL data
We observed a number of discrepancies between the data sets of IPLs and FISH.
For example, we interpret the IPL distributions of two Calyptogena-influenced
sediment samples (HR-S-Calyp-1 and -2) as evidence of a substantial contribution of
ANME-1, while FISH analysis suggest a strong predominance of ANME-2c (Knittel et
al., 2003, 2005) (Table III.1). This discrepancy can partly be explained by the large
difference of cell surface areas of ANME-1 and ANME-2 (cf. Table III.1). If we take
these differences into account and calculate the surface area of ANME-1 vs. ANME-2
cells, we would predict that ANME-1 lipids are almost equally abundant as ANME-2
lipids. However, ANME-1 derived IPLs are much higher concentrated than their
ANME-2 counterparts. Potential explanations for these discrepancies can be sought in
both methodologies. For example, the large predominance of ANME-1 lipids in
samples HR-S-Calyp-1 and -2 could also be due to a fossil component in the IPL signal
although this would require that the system has evolved from an ANME-1 to an
ANME-2 dominated community. One explanation could be a period of starvation
which has been proposed to induce a dramatic decrease in phospholipid content (Oliver
and Stringer, 1984).
On the other hand, FISH counts may underestimate certain members of the
archaeal community due to the low permeability of their cell membranes (Wagner et
al., 2003). Based on the membrane lipid structure, we can argue that ANME-1 cells are
probably more rigid than ANME-2 cells (cf. Valentine, 2007), which may negatively
impact their detectability by FISH. Moreover, the selective FISH approach will not
detect archaea outside of the window of interest that may contribute IPLs similar to

Chapter III ______________________________________________________________________
88
those of ANME-1, i.e., the ubiquitous Marine Benthic Group B, which has been
detected previously in clone libraries of the Black Sea (Knittel et al., 2005) and is
presumed to produce 2Gly-GDGT as the major lipid (cf. Biddle et al., 2006; Lipp et al.,
2008). Additionally, there are several samples for which FISH suggests a relatively
high proportion of bacteria while IPL analysis did not detect bacterial IPLs at all (HR-
S-Calyp-1, -2 and GOM-S-Campeche knolls). This could be a result of generally lower
detectability of archaeal cells via FISH, thus resulting in an overestimation of bacteria.
Other factors such as current and past physiological state of a cell can likewise
influence the RNA content and therefore the detection by FISH techniques (Oda et al.,
2000), but probably also the lipid content of cells.
Environmental factors controlling the distribution and composition of AOM
communities
The linkage of environmental conditions and AOM community type was
evaluated by RDA based on the distribution of IPLs (Fig. III.3). Environmental
variables which were previously inferred to influence AOM community distribution are
fluid flow (Girguis et al., 2005), temperature (Nauhaus et al., 2005), oxygen, sulfate
and methane availability (Knittel et al., 2005). Furthermore, the influence of salinity
and pH over AOM activity has been investigated, although both do not appear to be
important (Nauhaus et al., 2005). From a total of twelve variables, 7 were shown to be
related to the IPL distribution (in priority order based on forward selection): oxygen
concentrations in the bottom water (O2), sulfate (SO42-) and methane concentrations
(CH4), sulfate reduction rate (SRR), total organic carbon concentration (TOC),
temperature and pH (Table III.2). From these variables, SRR and TOC were positively
correlated with each other and negatively correlated with SO42-. Variables additionally
included in the statistical analysis, which finally did not contribute to the variability of
the IPLs, were fluid flow (included as qualitative data), water depth, salinity, sulfide
and phosphate concentrations.

Chapter III ______________________________________________________________________
89
Fig. III.3. RDA plot showing the distribution of samples and IPLs in function of environmental variables that explain most of the variability. Environmental variables are shown in red arrows. TOC = total organic carbon, SRR= sulfate reduction rate, O2 = oxygen concentration in the bottom water, CH4 = methane concentration, SO4
2- = sulfate concentration. Color code is as in Figure III.2, except that molecularly uncharacterized samples (grey circles in Figure III.2) are colored according to the phylogenetic grouping in which they were displayed based on the IPL distribution. IPL names are given according to the abbreviation in Table III.3. As an example, mats dominated by ANME-2a/DSS (dots with white cross) were characterized by high sulfate reduction rates and diagnostic IPLs # 21 and 22.
The RDA separated all microbial mat samples from the Black Sea, independent
of the dominant AOM community type, from all of the sediment samples (Fig. III.3).
The main variables associated with this separation were temperature, TOC and SRR.
Microbial mats dominated by ANME-1/DSS (BS-M-interior, BS-M-trunk-1 to -3) were
characterized by higher TOC content and lower SRR compared to the ANME-2a/DSS
dominated mats (BS-M-nodule-1 to -3). Furthermore, temperature affected ANME-
1/DSS dominated AOM communities (mats and sediments), as illustrated by a very

Chapter III ______________________________________________________________________
90
similar direction of its vector in relation to that of 2Gly-GDGT, the IPL diagnostic of
ANME-1. A relationship between ANME-1/DSS and temperature has been previously
suggested by Nauhaus et al. (2005) based on results from in vitro experiments that
indicated higher AOM activity of ANME-1/DSS from microbial mats between 16°C
and 24°C compared to ANME-2a/DSS from sediments, for which a temperature
optimum between 10°C and 15°C was observed. Sediment samples of the ANME-
1/DSS type, by contrast, were more widely distributed during RDA and just weakly
affected by temperature and pH. The respective plot region was characterized by 2Gly-
GDGT-PG and 2PG-GDGT (# 11 and 13 in Fig. III.3, respectively), which were IPLs
also displayed in the Calyptogena-influenced sediments (HR-S-Calyp-1 and -2) and in
uncharacterized sediments from the Eastern Mediterranean Sea (EMS-S-SOB). The
wide distribution of ANME-1/DSS dominated samples relative to pH is in agreement
with previous observations that were not suggestive of a direct relationship (Nauhaus et
al., 2005).
O2 influenced the data distribution in an opposite direction as TOC and
contributed to the separation of mats and sediments. Macrofauna is less abundant in
areas where oxygen is scarce (Levin et al., 2002). Thus, grazing on microbial
communities by macrofauna in anoxic water bodies is absent, allowing the increase of
biomass and therefore TOC. Another environment with low oxygen concentrations
was represented by a sample from the oxygen minimum zone of the Arabian Sea (AS-
S-SOB orange - grouped with ANME-1/DSS dominated samples). However here, TOC
was not as high as in the carbonate reefs from the Black Sea, where typical values of
~25mg of TOC mL-1 of mat were reported (Michaelis et al., 2002).
Displayed opposite to SRR, pH varied similarly in both sediments and in the
Black Sea mats (between 7.4 and 8.3) and was expressed with a rather short vector.
Broader pH values between 6.8 and 8.1 are suggested to be optimum for ANME-1/DSS
activity, whereas for ANME-2/DSS communities the reported optimum is at 7.4
(Nauhaus et al., 2005). The overlapping of the pH values from ANME-1/DSS and
ANME-2/DSS dominated habitats suggested that the communities are not strongly
influenced by pH. Additionally, the metabolic activities of sulfide oxidizing bacterial
communities contribute to an effective supply of SO42-, a variable influencing the group

Chapter III ______________________________________________________________________
91
of sediment samples, particularly those dominated by ANME-2a/DSS. The occurrence
of both SO42-and CH4 (although expressed in a shorter vector compared to most of the
other variables) in the sediment samples dominated by ANME-2a/DSS suggests that a
high supply of these two reactants is an important criterion selective for ANME-
2a/DSS.
ANME-2a/DSS and ANME-3/DBB dominated sediments and some of the
molecularly uncharacterized samples (AS-S-Calyp, AS-S-Thio and HR-S-Beg-1) were
related to O2, SO42- and CH4. The most prominent IPLs found in this grouping were
PG-OH-AR and PI-OH-AR (# 25 and 27 in Fig. III.3, respectively), whereas for the
ANME-3 group, PME, PE-DAG, PDME and BL-even were observed. Among these,
the bacterial IPLs were inversely related to temperature. This relationship is explained
by the fact the ANME-3 type communities were only observed at Håkon Mosby Mud
Volcano, at bottom temperatures of around –1°C.
Based on IPL distribution, it was possible to distinguish microbial mats at
carbonate reefs from sediment samples (Figs. III.1 and III.2). This distinction was
corroborated by the inclusion of environmental variables for the purpose of the RDA
(Fig. III.3). With respect to IPL distribution, the major distinctive feature of these two
habitats is the importance of glycosidic vs. phospho-IPLs. Microbial mats in the Black
Sea affiliated with both ANME-1/DSS and ANME-2/DSS groups contained more than
75% glycosidic IPLs derived from archaea compared to the same consortia inhabiting
sediments which showed lower relative amounts (Figs. III.1a and b). This trend was
also accompanied by higher contributions of bacterial IPLs with non-phospho
headgroups such as OL, BL and surfactin in mats compared to sediments (Fig. III.1d).
The low abundance of phospho-IPLs in samples from reef-like structures in the Black
Sea could be related to phosphate availability (cf. van Mooy et al., 2006). Dissolved
phosphate in sediment pore water has been shown to be strongly adsorbed on calcium
carbonate (Cole et al., 1953; de Kanel and Morse, 1978). During AOM, precipitation
of calcium carbonate is highly stimulated by the increase in alkalinity (e.g., Barnes and
Goldberg, 1976; Ritger et al., 1987; Michaelis et al., 2002). In case of the chimney-like
structures of the Black Sea, Mg-calcite minerals rich in iron sulfide precipitates co-
occur with aragonite phases producing the characteristic highly cavernous stable fabric

Chapter III ______________________________________________________________________
92
(Reitner et al., 2005). Peckmann et al. (2001) suggested that, contrary to Mg-calcite
precipitation, aragonite precipitation in these chimney systems occurs under low
phosphate and high sulfate concentration. Hence, it is conceivable that the chimneys
act as a sink for dissolved phosphate, thus limiting phosphate availability for the
microbial communities inhabiting these carbonate structures. In analogy to marine
planktonic communities and cyanobacteria (van Mooy et al., 2006), AOM communities
may adapt their lipid membrane composition towards IPLs with higher proportions of
glycosidic lipids in case of archaeal, and BL and OL in case of bacterial community
members. For example, Pseudomonas fluorescens has been shown to substitute
phospholipids with OL in response to phosphate limitation (Minnikin and
Abdolrahimzadeh, 1974). Hence, the difference in IPL distribution of mats and
sediments may result from phosphate limitation rather than taxonomic control.
CONCLUSIONS
We distinguished the major microbial communities involved in AOM based on
the distribution of IPLs. IPL distribution allowed the identification of the major ANME
groups with and without phylogenetic information. In line with previous observations
(Rossel et al., 2008), one key feature of ANME-1/DSS dominated systems was the
higher abundance of intact GDGTs compared to ANME-2a/DSS and ANME-3/DBB in
which higher abundance of AR-based IPLs and bacterial lipids were characteristic.
Furthermore, within the main IPL types present in each community, additional
differences related to the habitat characteristics were also influencing the IPL
composition. For example, limitation of dissolved phosphate in AOM mats in
carbonate reef environments of the Black Sea is likely responsible for the generally low
amount of phospho-IPLs in both ANME-1/DSS and ANME-2a/DSS dominated mats in
the Black Sea when compared to sediments inhabited by the same communities.
We constrained several factors selecting for one of the three major ANME
community types. The dominance of ANME-1/DSS was associated with higher
temperatures and anoxia. In sediments dominated by ANME-2a/DSS, higher
concentrations of oxygen in the bottom water, methane, and, most importantly, sulfate

Chapter III ______________________________________________________________________
93
were key environmental parameters involved in selection of this community.
Effective supply of sulfate in sediments in which ANME-2a/DSS inhabits are possibly
facilitated by the production of sulfate coupled to removal of sulfide by sulfide
oxidizing bacterial mats.
The diversity of bacterial IPLs was high and strongly differed among the
settings analyzed. These differences reflect the diversity of bacteria in AOM
environments. Bacterial IPLs were generally less abundant and diverse in ANME-
1/DSS dominated systems compared to ANME-2a/DSS and ANME-3/DBB. The
taxonomic resolution of apolar lipids, i.e., compounds commonly targeted in lipid-
based studies of AOM environments, was insufficient for a distinction of the major
ANME community types.
ACKNOWLEDGMENTS
We thank the captain, crew, and shipboard scientist from the R/V SONNE SO
148-1, SO 165-2, SO 174, R/V L’Atalante 2003, R/V Polarsten PS64, R/V Poseidon
PO 317/3, Meteor M74-3, M70-2, R/V Atlantis, 1998, R/V Melville, 1998, R/V
Heincke HE208, R/V Logachev TTR-15 and TTR-11 for the support during sample
collection. Helge Niemann, Beth Orcutt, Victoria Orphan, Andreas Teske, Tina Treude,
and Gunter Wegener are gratefully acknowledged for providing several of the samples
analyzed here. We also thank Julius Lipp and Xavier Prieto for technical support on the
LC-ESI-MS and the GC-MS. We thank Julia Arnds and Katrin Knittel for phylogenetic
information and also Janine Felden, Helge Niemann, Florence Schubotz, Beth Orcutt,
Ana Lichtschlag, Frank Wenzhöfer and Dirk DeBeer for the unpublished data supplied.
This study was part of the program MUMM II (grant 03G0608C), funded by the
Bundesministerium für Bildung und Forschung (BMBF, Germany) and the Deutsche
Forschungsgemeinschaft (DFG, Germany). Further support was provided by the Center
for Marine Environmental Sciences (MARUM) at the University of Bremen funded by
the DFG-Research Center/Excellent Cluster “The Ocean in the Earth System.

Chapter III ______________________________________________________________________
94
REFERENCES
Aharon, P., Fu, B., 2000. Microbial sulfate reduction and sulfur and oxygen isotope
fractionations at oil and gas seeps in deepwater Gulf of Mexico. Geochimica et
Cosmochimica Acta 64, 233-246.
Ahimou, F., Jacques, P., Deleu, M., 2000. Surfactin and iturin A effect on Bacillus
subtilis surface hydrophobicity. Enzyme and Microbial Technology 27, 749-754.
Barnes, R., Goldberg, E., 1976. Methane production and consumption in anoxic marine
sediments. Geology 4, 297-300.
Barridge, J. K., Shively, J. M., 1968. Phospholipids of the Thiobacilli. Journal of
Bacteriology 95, 2182-2185.
Benning, C., Huang, Z. -H., Gage, D. A., 1995. Accumulation of a novel glycolipid and
a betaine lipid in cells of Rhodobacter sphaeroides grown under phosphate
limitation. Archives of Biochemistry and Biophysics 317, 103-111.
Biddle, J. F., Lipp, J. S., Lever, M. A., Lloyd, K. G., Sörensen, K. B., Anderson, R.,
Fredricks, H. F., Elvert, M., Kelly, T. J., Schrag, D. P., Sogin, M. L., Brenchley, J.
E., Teske, A. House, C. H., Hinrichs, K. –U., 2006. Heterotrophic archaea
dominate sedimentary subsurface ecosystems off Peru. Proceedings of the
National Academy of Science U.S.A. 103, 3846-3851.
Blumenberg, M., Seifert, R., Reitner, J., Pape, T., Michaelis, W., 2004. Membrane lipid
patterns typify distinct anaerobic methanotrophic consortia. Proceedings of the
National Academy of Science U.S.A. 101, 11111-11116.
Boetius, A., Ravenschlag, K., Schubert, C. J., Rickert, D., Widdel, F., Gieseke, A.,
Amann, R., Jørgensen, B. B., Witte, U., Pfannkuche, O., 2000. A marine
microbial consortium apparently mediating anaerobic oxidation of methane.
Nature 407, 623-626.
Boetius, A., Suess, E., 2004. Hydrate Ridge: a natural laboratory for the study of
microbial life fueled by methane from near-surface gas hydrates. Chemical
Geology 205, 291-310.
Bohrman, G., cruise participants., 2008. Report and preliminary results of R/V Meteor
cruise M74/3, Fujairah-Male, 30 October-28 November, 2007. Cold seeps of

Chapter III ______________________________________________________________________
95
Makran subduction zone (continental margin of Pakistan). Berichte, Fachbereich
Geowissenschaften, Universität Bremen, No. 266, 161 pages. Bremen.
Cole, C. V., Olsen, S. R., Scott, C. O., 1953. The nature of phosphate sorption by
calcium carbonate. Soil Science Society of America Journal 17, 352-356.
Cowie, G. L., Calvert, S. E., Pedersen, T. F., Schultz, H., Von Rad, U., 1999. Organic
content and preservational controls in surficial shelf and slope sediments from the
Arabian Sea (Pakistan margin). Marine Geology 161, 23-38.
Curatolo, E., 1987. The physical properties of glycolipids. Biochimica et Biophysica
Acta 906, 111-136.
Dando, P. R., Bussmann, I., Niven, S. J., O´Hara, S. C. M., Schmaljohann, R., Taylor,
L. J., 1994. A methane seep area in the Skagerrak, the habitat of the pogonophore
Siboglinum poseidoni and the bivalve mollusk Thyasira sarci. Marine Ecology
Progress Series 107, 157-167.
Day, A. S., 2003. Documenting modern and ancient methane release from cold seeps
using deep-sea benthic foraminifera, M.Sc. Thesis. University of Florida.
De Beer, D., Sauter, E., Niemann, H., Kaul, N., Foucher, J. -P. Witte, U., Schlüter, M.,
Boetius, A., 2006. In situ fluxes and zonation of microbial activity in surface of
the Håkon Mosby Mud Volcano. Limnology and Oceanography 51, 1315-1331.
De Kanel, J., Morse, J. W., 1978. The chemistry of orthophosphate uptake from
seawater on to calcite and aragonite. Geochimica et Cosmochimica Acta 42, 1335-
1340.
DeLong, E. F., Wickham, G. S., Pace, N. R., 1989. Phylogenetic stains: Ribosomal-
based probes for the identification of single cells. Science 243, 1360-1363.
Dembitsky, V. M., 1996. Betaine ether-linked glycerolipids: chemistry and biology.
Progress in Lipid Research 35, 1-51.
Elvert, M., Suess E., Whiticar, M. J., 1999. Anaerobic methane oxidation associated
with marine gas hydrates: superlight C-isotopes from saturated and unsaturated
C20 and C25 irregular isoprenoids. Naturwissenschaften 86, 295-300.
Elvert, M., Hopmans, E. C., Treude, T., Boetius, A., Suess E., 2005. Spatial variations
of methanotrophic consortia at cold methane seeps: implications from a high-
resolution molecular and isotopic approach. Geobiology 3, 195-209.

Chapter III ______________________________________________________________________
96
Elvert, M., Niemann, H., 2008. Occurrence of unusual steroids and hopanoids derived
from aerobic methanotrophs at an active marine mud volcano. Organic
Geochemistry 39, 167-177.
Ertefai, T. F., Fisher, M. C., Fredricks, H. F., Lipp, J. S., Pearson, A., Birgel, D., Udert,
K. M., Cavanaugh, C. M., Gschwend, P. M., Hinrichs, K. -U., 2008. Vertical
distribution of microbial lipids and functional genes in chemically distinct layers
of a highly polluted meromictic lake. Organic Geochemistry 39, 1572-1588.
Fang, J. S., Barcelona, M. J., Semrau, J. D., 2000. Characterization of methanotrophic
bacteria on the basis of intact phospholipid profiles. FEMS Microbiology Letters
189, 67-72.
Fredricks, H. F., Hinrichs, K. -U. 2007. Data report: intact membrane lipids as
indicators of subsurface life in Cretaceous and Paleogene sediments from site
1257 and 1258. Proceedings of the Ocean Drilling Program, Scientific Results
207.
Ghosh, M., Misha, A. K., 1987. Occurrence, identification and possible significance of
ornithine lipid in Thiobacillus ferrooxidans. Biochemical and Biophysical
Research Communications 142, 925-931.
Gieskes, J. M., Elderfield, H., Lawrence, J. R., Johnson, J., Meyers, B., Campbell, A.,
1982. Geochemistry of interstitial waters and sediments, Leg 64, Gulf of
California. Initial Reports, Deep Sea Drilling Project 64, 675-694. Washington,
DC: US Government Printing Office.
Goldfine, H., 1984. Bacterial membrane and lipid packing theory. Journal of Lipid
Research 25, 1501-1507.
Grandel, S., Rickert, D., Schlüter, M., Wallmann, K., 2000. Pore-water distribution and
quantification of diffusive benthic fluxes of silicic acid, nitrate and phosphate in
surface sediments of the deep Arabian Sea. Deep-Sea Research II 47, 2707-2734.
Guirguis, P. R., Cozen, A. E., DeLong, E. F., 2005. Growth and population dynamics of
anaerobic methane oxidizing archaea and sulfate.reducing bacteria in a
continuous-flow bioreactor. Applied and Environmental Microbiology 71, 3725-
3733.

Chapter III ______________________________________________________________________
97
Heijs, S. H., Haese, R. R., Van der Wielen, P. W. J. J., Forney, L. J., Van Elsas, J. D.,
2007. Use of 16S rRNA gene based clone libraries to assess microbial
communities involved in anaerobic methane oxidation in a Mediterranean cold
seep. Microbial Ecology 53, 384-398.
Hinrichs, K. -U., Hayes, J. S., Sylva, S. P., Brewer, P. G., DeLong, E. F., 1999.
Methane-consuming archaebacteria in marine sediments. Nature 398, 802-805.
Hinrichs, K. -U, Summons, R. E, Orphan, V., Sylva, S. P., Hayes, J. M., 2000.
Molecular and isotopic analyses of anaerobic methane-oxidizing communities in
marine sediments. Organic Geochemistry 31, 1685-1701.
Hofman, M., Eichenberger, W., 1996. Biosynthesis of diacylglyceryl-N,N,N-
trimethylhomoserine in Rhodobacter sphaeroides and evidence for lipid-linked N
methylation. Journal of Bacteriology 178, 6140-6144.
Imhoff, J. F., Bias-Imhoff, U., 1995. Lipids, Quinones and Fatty Acids of Anoxygenic
Phototrophic Bacteria. In Anoxygenic Photosynthetic Bacteria (eds. Blankenship,
R. E., Madigan, M. T., Bauer, C. E.), 179-205. Kluwer Academic Publishers,
Netherlands.
Ishikawa, M., Ichikuni, M., 1981. Coprecipitation of phosphate with calcite.
Geochemical Journal 15, 283-288.
Iversen, N., Jørgensen, B. B., 1985. Anaerobic methane oxidation rates at the sulfate-
methane transition in marine sediments from Kattegat and Skagerrak (Denmark).
Limnology and Oceanography 30, 944-955.
Jørgensen, B. B., Isaksen, M. F., Jannasch, H. W., 1992. Bacterial sulfate reduction
above 100°C in deep-sea hydrothermal vent sediments. Science 258, 1756-1757.
Jørgensen, B. B., Weber, A., Zopfi, J., 2001. Sulfate reduction and anaerobic methane
oxidation in Black Sea sediments. Deep-Sea Research I 48, 2097-2120.
Kato, M., Sakai, M., Adachi, K., Ikemoto, H., Sano, H., 1996. Distribution of betaine
lipids in marine algae. Phytochemistry 42, 1341-1345.
Khuller, G. K., Goldfine, H., 1974. Phospholipids of Clostridium butyricum. V. effects
of growth temperature on fatty acid, and alk-1-enyl ether group, and phospholipid
composition. Journal of Lipid Research 15, 500-507.

Chapter III ______________________________________________________________________
98
Knab, N. J., 2007. Controls of Anaerobic oxidation of methane in ocean margin
sediments. Ph.D. Thesis. University of Bremen.
Knittel, K., Boetius, A., Lemke, A., Eilers, H., Lochte, K., Pfannkuche, O., Linke, P.,
2003. Activity, distribution, and diversity of sulfate reducers and other bacteria in
sediments above gas hydrate (Cascadia Margin, Oregon). Geomicrobiology
Journal 20, 269-294.
Knittel, K., Lösekann, T., Boetius, A., Kort, R., Amann, R., 2005. Diversity and
Distribution of Methanotrophic Archaea at Cold Seeps. Applied and
Environmental Microbiology 71, 467-479.
Knoche, H. W., Shively, J. M., 1972. The structure of an ornithine –containing lipid
from Thiobacillus thioxidans. The Journal of Biological Chemistry 247, 170-178.
Koga, Y., Nishihara, M., Morii, H., Akagawa-Matsushita, M., 1993. Ether polar lipids
of methanogenic bacteria: structures, comparative aspects, and biosyntheses.
1996. Microbiological Reviews 57, 164-182.
Koga, Y., Morii, H., Akagawa-Matsushita, M., Ohga, I., 1998. Correlation of polar
lipid composition with 16S rRNA phylogeny in methanogens. Further analysis of
lipid component parts. Bioscience Biotechnology and Biochemistry 62, 230-236.
Koga, Y., Morii, H., 2005. Recent advances in structural research on ether lipids from
archaea including comparative and physiological aspects. Bioscience
Biotechnology and Biochemistry 69, 2019-2034.
Krüger, M., Wolters, H., Gehre, M., Joye, S. B., Richnow, H. -H., 2008. Tracing the
slow growth of anaerobic methane-oxidizing communities by 15N-labelling
techniques. FEMS Microbiology Ecology 63, 401-411.
Legendre, P., Gallagher, E. D., 2001. Ecologically meaningful transformations for
ordination of species data. Oecologia 129, 271-280.
Levin, L. A., 2002. Deep-ocean life where oxygen is scarce. American Scientist 90,
436-444.
Levin, L. A., Ziebis, W., Mendoza, G. F., Growney, V. A., Tryon, M. D., Brown, K.
M., Mahn, C., Gieskes, J. M., Rathburn, A. E., 2003. Spatial heterogeneity of
macrofauna at northern California methane seep: influence of sulfide
concentration and fluid flow. Marine Ecology Progress Series 265, 123-139.

Chapter III ______________________________________________________________________
99
Lipp, J. S., Morono, Y, Inagaki, F., Hinrichs, K. -U., 2008. Significant contribution of
Archaea to the extant biomass in marine subsurface sediments. Nature 454, 991-
994.
Lohse, L., Epping, E. H. G., Helder, W., Van Raaphorst, W., 1996. Oxygen pore water
profiles in continental shelf sediments of the North Sea: turbulent versus
molecular diffusion. Marine Ecology Progress Series 145, 63-75.
López -Lara, I. M., Sohlenkamp, C., Geiger, O., 2003. Membrane lipids in plant-
associated bacteria: their biosyntheses and possible functions. Molecular Plant-
Microbe Interactions Overview 16, 567-579.
Lösekann, T., Knittel, K., Nadalig, T., Fuchs, B., Niemann, H., Boetius, A., Amann, R.,
2007. Diversity and abundance of aerobic and anaerobic methane oxidizers at the
Haakon Mosby Mud Volcano, Barents Sea. Applied and Environmental
Microbiology 73, 3348–3362.
Makula, R. A., 1978. Phospholipid composition of methane-utilizing bacteria. Journal
of Bacteriology 134, 771-777.
Makula, R. A., Finnerty, W. R., 1975. Isolation and characterization of an ornithine-
containing lipid from Desulfovibrio gigas. Journal of Bacteriology 123, 523-529.
Martens, C. S., Berner, R. A., 1974. Methane production in the interstitial waters of
sulfate-depleted marine sediments. Science 185, 1167-1169.
Michaelis, W., Seifert, R., Nauhaus, K., Treude, T., Thiel, V., Blumenberg, M., Knittel,
K., Gieseke A., Peterknecht, F., Pape, T., Boetius A., Amann, R., Jørgensen, B.
B., Widdel, F., Peckmann, J., Pimenkov, N., Gulin, M. B., 2002. Microbial reefs
in the Black Sea fueled by anaerobic methane oxidation. Science 297, 1013-1015.
Milkov, A. V., Vogt, P. R., Crane, K., Lein, Y. A., Sassen, R., Cherkashev, G. A.,
2004. Geological, geochemical, and microbiological processes at the hydrate-
bearing Håkon Mosby Mud Volcano: a review. Chemical Geology 205, 347-366.
Mills, H. J., Martinez, R. J., Story, S., Sobecky, P. A., 2004. Identification of members
of the metabolically active microbial populations associated with Beggiatoa
species mat communities from Gulf of Mexico cold-seep sediments. Applied and
Environmental Microbiology 70, 5547-5458.

Chapter III ______________________________________________________________________
100
Minnikin, D. E., Abdolrahimzadeh, H., 1974. Effect of pH on the proportions of polar
lipids, in chemostat cultures of Bacillus subtilis. Journal of Bacteriology 120,
999-1003.
Mucci, A., 1986. Growth kinetics and composition of magnesian calcite overgrowths
precipitated from seawater: quantitative influence of orthophosphate ions.
Geochimica et Cosmochimica Acta 50, 2255-2265.
Nauhaus, K., Treude, T., Boetius, A., Krüger, M., 2005. Environmental regulation of
the anaerobic oxidation of methane a comparison of ANME-1 and ANME-II
communities. Environmental Microbiology 7, 98-106.
Niemann, H., 2005. Rates and signatures of methane turnover in sediments of
continental margins, Ph.D. Thesis. University of Bremen.
Niemann, H., Lösekann T., de Beer, D., Elvert, M., Nadalig, T., Knittel, K., Amann, R.,
Sauter, E., Schlüter, M., Klages, M., Foucher, J. -P., Boetius, A., 2006. Novel
microbial communities of the Haakon Mosby mud volcano and their role as a
methane sink. Nature 443, 854-858.
Oda, Y., Slagman, S. -J., Meijer, W. G., Forney, L. J., Gottschal, J. C., 2000. Influence
of growth rate and starvation on fluorescence in situ hybridization of
Rhodopseudomonas palustris. FEMS Microbiology Ecology 32, 205-213.
Oliver, J. D., Colwell, R. R., 1973. Extractable lipid of gram-negative marine bacteria:
phospholipid composition. Journal of Bacteriology 114, 897-908.
Oliver, J. D., Stringer, W. F., 1984. Lipid composition of a psychrophilic marine Vibrio
sp. during starvation-induced morphogenesis. Applied and Environmental
Microbiology 47, 461-466.
Omoregie, E. O., Mastalerz, V., De Lange, G., Straub, K. L., Kappler, A., Røy, H.,
Stadniskaia, A., Foucher, J. -P., Boetius, A., 2008. Biogeochemistry and
community composition of iron- and sulfur-precipitating microbial mats at the
Chefren Mud Volcano (Nile Deep Sea Fan, Eastern Mediterranean). Applied
Environmental Microbiology 74, 3198-3215.
Orcutt, B. N., 2007. Anaerobic oxidation of methane in cold seeps and gas hydrates:
responsible microorganisms, rates of activity, and interactions with other
processes, Ph.D. Thesis. University of Georgia, Athens.

Chapter III ______________________________________________________________________
101
Orphan, V. J., House, C. H., Hinrichs, K. -U., McKeegan, K. D., DeLong, E. F., 2001a.
Methane-consuming archaea revealed by directly coupled isotopic and
phylogenetic analysis. Science 20, 484-487.
Orphan, V. J., House, C. H., Hinrichs, K. -U., McKeegan, K. D., DeLong, E. F., 2001b.
Comparative analysis of methane-oxidizing archaea and sulfate-reducing bacteria
in anoxic marine sediments. Applied and Environmental Microbiology 67, 1922-
1934.
Orphan, V. J., House, C. H., Hinrichs, K. -U., McKeegan, K. D., DeLong, E. F., 2002.
Multiple archaeal groups mediate methane oxidation in anoxic cold seep
sediments. Proceedings of the National Academy of Science U.S.A. 99, 7663-
7668.
Orphan, V. J., Ussler, III. W., Naehr, T. H., House, C. H., Hinrichs, K. -U. Paull, C. K.,
2004. Geological, geochemical, and microbiological heterogeneity of the seafloor
around methane vents in the Eel River Basin, offshore California. Marine
Geology 205, 265-289.
Pancost, R. D., Sinninghe Damsté, J. S, Lint, S. D., van der Maarel, M. J. E. C.,
Gottschal J. C., Shipboard Scientific Party., 2000. Biomarker evidence for
widespread anaerobic methane oxidation in Mediterranean sediments by a
consortium of methanogenic archaea and bacteria. Applied Environmental
Microbiology 66, 1126-1132.
Pancost, R. D., Bouloubassi, I., Aloisi, G., Sinninghe Damsté, J. S., Party M. S. S.,
2001. Three series of non-isoprenoidal dialkyl glycerol diethers in cold-seep
carbonate crust. Organic Geochemistry 32,695-707.
Paull, C. K., Ussler III, W., Browski, W. S., Spiess, F. N., 1995. Methane- Rich plumes
on the Carolina continental rise: associations with gas hydrates. Geology 23, 89-
92.
Peckmann, J., Reimer, A., Luth, U., Hansen, B. T., Heinicke, C., Hoefs, J., Reitner. J.,
2001. Methane-derived carbonates and authigenic pyrite from northwestern Black
Sea. Marine Geology 177, 129-150.
Ramette, A., 2007. Multivariate analysis in microbial ecology. FEMS Microbiology
Ecology 62, 142-160.

Chapter III ______________________________________________________________________
102
Reeburgh, W. S., 1980. Anaerobic methane oxidation: rate depth distributions in Skan
Bay sediments. Earth and Planetary Science Letters 47, 345-352.
Reeburgh, W. S., 1996. “Soft Spots” in the global methane budget. In: Microbial
growth on C1 compounds (eds. Lidstrom, M. E., Tabita, F. R.), 334-342, Kluwer
Academic Publishers, Dordrecht.
Reitner, J., Peckmann, J., Reimer, A., Schumann, G., Thiel, V., 2005. Methane-derived
carbonate build-ups and associated microbial communities at cold seeps on the
lower Crimean shelf (Black Sea). Facies 51, 66-79.
Ritger, S., Carson, B., Suess, E., 1987. Methane-derived authigenic carbonates formed
by subduction-induced pore-water expulsion along Oregon/Washington margin.
Geological Society of America Bulletin 98, 147-156.
Roberts, Z. E., Meldrum, F. C., Pancost, R. D., 2008. The archaeal lipid composition of
partially lithified cold seep mats. Organic Geochemistry 39, 1000-1006.
Rossel, P. E., Lipp, J. S., Fredricks, H. F., Arnds, J., Boetius, A., Elvert, M., Hinrichs,
K. -U., 2008. Intact polar lipids of anaerobic methanotrophic archaea and
associated bacteria. Organic Geochemistry 39, 992-999.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2001. Monoalkylether phospholipids
in the sulfate-reducing bacteria Desulfosarcina variabilis and Desulforhabdus
amnigenus. Archives of Microbiology 176, 435-442.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2002. Phospholipid analysis as a tool
to study complex microbial communities in marine sediments. Journal of
Microbiological Methods 48, 149-160.
Sato, N., 1992. Betaine lipids. The botanical Magazine Tokyo 105, 185-197.
Schmaljohann, R., Drews, M., Walter, S., Linke, P., Von Rad, U., Imhoff, J, F., 2001.
Oxygen-minimum zone sediments in the northeastern Arabian Sea off Pakistan: a
habitat for the bacterium Thioploca. Marine Ecology Progress Series 211, 27-42.
Schouten, S., Wakeham, S. G., Sinninghe Damsté, J. S., 2001. Evidence for anaerobic
methane oxidation by archaea in euxinix waters of the Black Sea. Organic
Geochemistry 32,1277-1281.

Chapter III ______________________________________________________________________
103
Schouten, S., Wakeham, S. G., Hopmans, E. C., Sinninghe Damsté, J. S., 2003.
Biogeochemical evidence that thermophilic archaea mediate the anaerobic
oxidation of methane. Applied and Environmental Microbiology 69, 1680-1686.
Schouten, S., Hopmans, E. C., Bass, M., Boumann, H., Standfest, S., Könneke, M.,
Stahl, S. A., Sinninghe Damsté, J. S., 2008. Intact membrane lipids of
“Candidatus Nitrosopumilus maritimus”, a cultivated representative of the
cosmopolitan mesophilic group I Crenarchaeota. Applied and Environmental
Microbiology 74, 2433-2440.
Shaffer, G., 1986. Phosphate pumps and shuttles in the Black Sea. Nature 321, 515-
517.
Shimada, H., Nemoto, N., Shida, T., Yamagishi, A., 2008. Effects of pH and
temperature on the composition of polar lipids in Thermoplasma acidophilum
HO-62. Journal of Bacteriology 190, 5504-5411.
Shovitri, M., 2007. Biogeochemistry and molecular ecology of sandy sediment from
North Sea gas seep, M.Sc. Thesis. University of Bremen.
Stadnitskaia, A., Bouloubassi, I., Elvert, M., Hinrichs, K. -U., Sinninghe Damsté, J. S.,
2008. Extended hydroxyarchaeaol, a novel lipid biomarker for anaerobic
methanotrophy in cold seepage habitats. Organic Geochemistry 39, 1007-1014.
Sturt, H. F., Summons, R. E., Smith, K. J., Elvert, M., Hinrichs, K. -U., 2004. Intact
polar membrane lipids in prokaryotes and sediments deciphered by high-
performance liquid chromatography/electrospray ionization multistage mass
spectrometry-new biomarkers for biogeochemistry and microbial ecology. Rapid
Communications in Mass Spectrometry 18, 617-628.
Suess, E., Carson B., Ritger S. D., Moore, J. C., Jones, M. L., Kulm, L. D., Cochrane,
G. R., 1985. Biological communities at vent sites along the subduction zone off
Oregon. Biological Society of Washington Bulletin 6, 475-484.
Suess, E., Bohrmann, G., Huene, R., Linke, P., Wallmann, K., Lammers, S., Sahling,
H., 1998. Fluid venting in the eastern Aleutian subduction zone. Journal of
Geophysical Research 103, 2597-2614.
Suess, E., Torres, M. E., Bohrmann, G., Collier, R. W., Greinert, J., Linke, P., Rehder,
G., Trehu, A., Wallmann, K., Winckler, G., Zuleger, E., 1999. Gas Hydrate

Chapter III ______________________________________________________________________
104
destabilization: enhanced dewatering, benthic material turnover and öarge
methane plumes at the Cascadia convergent margin. Earth and Planetary Science
Letters 170, 1-15.
Teske, A., Hinrichs, K. -U., Edgecomb, V., de Vera Gomez, A., Kysela, D., Sylva, S.
P., Sogin, M. L., Jannasch, H. W., 2002. Microbial diversity of hydrothermal
sediments in the Guaymas Basin: Evidence for anaerobic methanotrophic
communities. Applied and Environmental Microbiology 68, 1994-2007.
Thiel, V., Peckmann, J., Siefert, R., Wehrung, P., Reitner, J., Michaelis, W., 1999.
Highly isotopically depleted isoprenoids: Molecular markers for ancient methane
venting. Geochimica et Cosmochimica Acta 73, 97-112.
Thiel, V., Peckmann, J., Richnow, H. H., Luth, U., Reitner, J., Michaelis, W., 2001.
Molecular signals for anaerobic methane oxidation in Black Sea seep carbonates
and a microbial mat. Marine Chemistry 73, 97-112.
Thompson, G. A., 1996. Lipids and membrane function in green algae. Biochimica et
Biophysica Acta 1302, 17-45.
Torres, M., McManus, J., Hammond, D. E., de Angelis, M. A., Heeschen, K., Colbert,
S. L., Tyron, M. D., Brown, K. M., Suess, E., 2002. Fluid and chemical fluxes in
and out of sediments hosting methane hydrate deposits on Hydrate Ridge, OR, I:
hydrological provinces. Earth and Planetary Science Letters 201, 525-540.
Treude, T., Boetius, A., Knittel, K., Wallmann, K., Jørgensen, B. B., 2003. Anaerobic
oxidation of methane above has hydrates at Hydrate Ridge, NE Pacific Ocean.
Marine Ecology Progress Series 264, 1-14.
Treude, T., Knittel, K., Blumenberg, M., Seifert, R., Boetius, A., 2005. Subsurface
microbial methanotrophic mats in the Black Sea. Applied and Environmental
Microbiology 71, 6375-6378.
Valentine, D. L., 2007. Adaptations to energy stress dictate the ecology and evolution
of the Archaea. Nature Reviews Microbiology 4, 316-323.
Van Mooy, B. A. S., Rocap, G., Fredricks H. F., Evans, C. T., Devol, A. H., 2006.
Sulfolipids dramatically decrease phosphorus demand by picocyanobacteria in
oligotrophic marine environments. Proceedings of the National Academy of
Science U.S.A. 103, 8607-8612.

Chapter III ______________________________________________________________________
105
Vater, J., 1986. Lipopeptides, an attractive class of microbial surfactants. Progress in
Colloid and Polymer Science 72, 12-18.
Wagner, M., Horn, M., Daims, H., 2003. Fluorescence in situ hybridization for the
identification and characterization of prokaryotes. Current opinion in
Microbiology 6, 302-309.
Wakeham, S. G., Sinninghe Damsté, J. S., Kohnen, M. E. L., de Leeuw, J. W., 1995.
Organic sulfur compounds formed during early diagenesis in Black Sea
sediments. Geochimica et Cosmochimica Acta 59, 521-533.
Weber, A., Jørgensen, B. B., 2002. Bacterial sulfate reduction in hydrothermal
sediments of the Guaymas Basin, Gulf of California, Mexico. Deep-Sea Research
I 49, 827-841.
Wegener, G., Shovitri, M., Knittel, K., Niemann, H., Hovland, M., Boetius, A., 2008.
Biogeochemical processes and microbial diversity of the Gullfaks and
Tommeliten methane seeps (Northern North Sea). Biogeosciences 5, 1127-1144.
White, D. C., Davis, W. M., Nickels, J. S., Kind, J. D., Bobbie, R. J., 1979. Oecologica
40, 51-62.
White, D. C., Ringelberg D. B., 1998. Signature lipid biomarker analysis. In:
Techniques in microbial ecology (eds. Burlage, R. S., Atlas, R, Stahl, D., Geesey,
G., Sayler, G.), 255-259, Oxford University Press, New York.
Yan, T., Ye, Q., Whou, J., Zhang, C. L., 2006. Diversity of functional genes for
methanotrophs in sediments associated with gas hydrates and hydrocarbon seeps
in the Gulf of Mexico. FEMS Microbiology Ecology 57, 251-259.
Yilmaz, A., Tugrul, S., 1998. The effect of cold-and warm-core eddied on the
distribution and stoichiometry of dissolved nutrients in the northeastern
Mediterranean. Journal of Marine Systems 16, 253-268.
Yun, J. W., Orange, D. L., Field, M. E., 1999. Subsurface gas offshore of Northern
California and its link to submarine geomorphology. Marine Geology 154, 357-
368.
Ziebis, W., Haese, R. R., 2005. Interactions between fluid flow, geochemistry, and
biogeochemical processes at methane seeps. In: Macro and microorganisms in

Chapter III ______________________________________________________________________
106
marine sediments (eds. Kristensen, E., Kostka, J., Haese, R. R.), 267-298, AGU
Coastal and Estuarine Studies (Vol 60 Coastal and Estuarine Studies).
III.2. SUPPLEMENTARY MATERIAL
Supplementary Figures and Tables
Supplementary Fig. III.4. Location of samples included in the global survey.

Chapter III ______________________________________________________________________
107
Supplementary Fig. III.5. PCA plot of the overall distribution of bacterial IPLs distinguishing their bond types (DEG = diether, DAG = diacyl), headgroups, sum of carbon atoms and number of unsaturations. Distinction between DAG and AEG was not possible, thus alkyl chains are provided for DAG. Bacterial IPLs with DEG are displayed with the names, all other are shown as DAG (AEG was not possible to distinguish, thus alkyl chains are provided for DAG). Color code of samples is according to Figure III.2 of the manuscript.

Chapter III ______________________________________________________________________
108
Supplementary Fig. III.6. PCA plot of the overall distribution of apolar lipids among the samples analyzed. Color code of samples is according to Figure III.2 of the manuscript. Abbreviations: ai = anteiso, AR = archaeol, Crocetane = 2,6,11,15-tetramethylhexadecane, Crocetene:1/2 = 2,6,11,15-tetramethylhexadecane with one or two double bond(s), DAGE = sn-1,2-di-O-alkyl glycerol ether, OH-AR = hydroxyarchaeol, 2OH-AR = dihydroxyarchaeol, FA = fatty acid, MAGE = sn-1 mono-O-alkyl glycerol ether, PMI =2,6,11,15, 19-pentamethylicosane, PMI:4 =2,6,11,15, 19-pentamethylicosene with four double bonds.

Cha
pter
III
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
109
Su
pple
men
tary
Tab
le II
I.4. R
elat
ive
abun
danc
e of
IPLs
in p
erce
ntag
e.
A
rabi
an S
ea
Bla
ck S
ea
Eas
tern
Med
iterr
anea
n Se
a E
el R
iver
B
asin
G
uaym
as
Bas
in
Gul
lfaks
G
ulf o
f Mex
ico
Håk
on M
osby
mud
V
olca
no
Hyd
rate
Rid
ge
IPL
AS-S-SOB orange
AS-S-thio
AS-S-calyp
BS-M-trunk-1
BS-M-nodule-1
BS-M-interior
BS-M-trunk-2
BS-M-trunk-3
BS-M-nodule-2
BS-M-nodule-3
BS-S
EMS-S-SOB
ER-S-SOB
GB-S-SOB orange
GF-S-SOB white
GOM-S-SOB white
GOM-S- Campeche
knolls
HMMV-S-Beg-1
HMMV-S-Beg-2
HMMV-S-Beg-3
HMMV-S-Beg-4
HR-S-Beg-1
HR-S-Beg-2
HR-S-Beg-3
HR-S-Calyp-1
HR-S-Calyp-2
HR-S-Calyp-3
2Gly
-GD
GT
(1)
29.7
74
.3
24.4
67
.2
82.1
79
.1
3.0
3.8
61.6
64.0
60
.9
41
.1
80.5
1.
4
5.6
18.1
7.
0 10
0.0
3Gly
-GD
GT
(2)
5.0
3.1
4G
ly-G
DG
T (3
)
1.6
2G
ly-G
DG
T+1
4 (4
)
14.5
2Gly
-GD
GT
+18
(5)
0.
6
1.2
1.1
2Gly
-GD
GT
+28
(6)
9.
1
2Gly
-GD
GT
+145
(7)
0.
5
1.
9 1.
5
0.6
4.
1
2Gly
-GD
GT
-PE
(8)
0.
3
MA
PT-G
DG
T-P
G (9
)
7.
9
G
ly-G
DG
T-P
G (1
0)
4.8
0.
2 1.
0
1.
3 3.
1
2Gly
-GD
GT
-PG
(11)
13
.6
0.3
1.1
3.7
14.2
8.
0 1.
2 8.
7
5.9
2.
7 14
.9
20.6
PG-G
DG
T (1
2)
20
.2
1.
7 6.
7 2.
5
2PG
-GD
GT
(13)
29
.5
0.3
0.
4 3.
7 0.
2
3.
7 14
.12
3.
3
3.3
20.2
48
.2
PE
-GD
GT
-PG
(14)
0.1
4.
8
APT
-GD
GT
-PG
(15)
13
.5
APT
-GD
GT
-238
(16)
4.
7
2P
-GD
GT
+155
(17)
14.3
GL
Y_M
AR
(18)
0.3
1.
4
2.
5 1.
1
2Gly
-AR
(19)
0.7
0.
3 3.
0 3.
2 0.
6 0.
9 14
.5
9.3
3.3
0.5
2.3
2.
4 1.
1 11
.2
2.8
2.2
1.6
G
ly-O
H-A
R (2
0)
0.4
0.
7
2G
ly-O
H-A
R (2
1)
0.9
0.7
0.1
3.
7 1.
9
2Gly
-MA
R (2
2)
0.
1 1.
7 1.
5 0.
4 0.
2 7.
9 4.
1
0.6
PE
-OH
-AR
(23)
0.
8 0.
2
0.
6 0.
6 3.
7
3.
7
0.6
PG
-AR
(24)
6.3
25.6
5.
9 0.
2
12.7
12
.5
4.
8
0.
6 2.
9
PG-O
H-A
R (2
5)
10
.2
4.5
0.2
0.6
16
.5
16.9
6.
7 19
.9
2.6
1.1
13.2
5.
8 29
.7
6.5
7.1
0.7
T
enta
tive
A
PT-O
H-A
R (2
6)
6.
5
PI-O
H-A
R (2
7)
2.5
11.9
18
.8
0.
1
7.
6 0.
4 0.
4 12
.5
1.4
1.6
2.8
0.3
0.5
1.3
5.1
13.0
2.
4 5.
9 0.
6
PS-A
R (2
8)
2.
2
0.
2
0.
6
0.5
0.8
0.3
1.0
0.8
PS
-OH
-AR
(29)
1.
0 29
.4
1.3
0.7
0.2
2.
2 0.
5
0.3
0.9
0.3
4.
9 0.
9 1.
1 4.
7 3.
4 25
.4
10.2
6.
0 2.
7
PS-2
OH
-AR
(30)
3.7
0.1
0.3
1.
6 0.
1 0.
3 0.
9 0.
6 3.
1 0.
6
Ten
t P-A
R+2
23 (3
1)
0.
9
14
.0
6.0
4.1
8.1
0.
5
0.4
G
LY
-PG
-AR
(32)
0.
1 0.
1
0.6
T
ent.G
ly-P
S-A
R (3
3)
6.
9
0.3
T
en. G
ly-P
-AR
ext
ende
d (3
4)
15
.7
0.5
0.4
0.
5
PC-D
AG
(I)
0.6
12.8
0.2
0.
1
1.
2
1.2
0.9
3.0
1.6
3.0
PG
-DA
G (I
I)
0.
1
23.2
21
.6
9.
7 3.
6
4.8
1.2
6.
2 20
.8
12
.5
PE
-DA
G (I
II)
2.9
10.8
27
.6
0.4
8.9
3.3
0.8
2.4
15.9
7.
8 8.
8 19
.2
6.6
2.9
47.6
1.
4
9.2
18.6
8.
0 23
.0
28.0
10
.2
36.5
PE-D
EG
(IV
) 1.
0 2.
7
0.4
3.9
0.9
0.3
2.2
4.1
2.5
4.7
1.9
0.5
3.7
3.0
10.6
PME
-DA
G- (
V)
0.8
6.
9
1.
4
8.
4
3.
0 5.
4 10
.2
3.7
0.3
PD
ME
.-DA
G (V
I)
11
.0
14.5
21
.4
14.5
O
L (V
II)
7.0
1.6
0.
1 4.
8 8.
0
1.4
2.
4 1.
4 1.
0 5.
6 0.
3
BL
-eve
n (V
III)
6.
3 3.
4 17
.5
1.4
5.1
0.7
2.4
3.9
33
.8
53.5
53
.4
23.8
13
.6
B
L-o
dd (I
X)
2.7
1.9
8.9
1.4
2.8
1.6
3.6
3.7
Su
rfac
tin (X
)
1.1
13.1
26
.5
U
nkno
wn
a
0.4
9.0
2.9
0.3
0.
4 0.
7 0.
4 2.
8 20
.7
3.
0 12
.2
0.8
1.
1
Unk
now
n b
1.
4
3.
6 5.
1

Cha
pter
III
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
____
110
Supp
lem
enta
ry T
able
III.5
. Con
cent
ratio
n of
apo
lar l
ipid
s.
A
rabi
an S
ea
Bla
ck S
ea
Gul
faks
H
åkon
Mos
by m
ud
Vol
cano
H
ydra
te R
idge
μg/g
AS-S-SOB orange
AS-S-Thio
AS-S-Calyp
BS-M-trunk-1
BS-M-nodule-1
BS-M-interior
BS-M-trunk-2
BS-M-trunk-3
BS-M-nodule-2
BS-M-nodule-3
GF-S-SOB white
HMMV-S-Beg-1
HMMV-S-Beg-2
HR-S-Beg-1
HR-S-Beg-2
HR-S-Calyp-2
HR-S-Calyp-3
Cro
ceta
ne
0.9
29.3
0.
7 0.
9 4.
8 12
.3
34.9
33
.6
8.4
120.
2 0.
0 0.
0 0.
0 1.
3 0.
8 0.
6 0.
3
CR
:1'''
0.
0 6.
0 0.
0 0.
0 0.
0 0.
4 0.
0 0.
0 0.
7 9.
1 0.
0 0.
0 0.
0 0.
5 0.
3 0.
1 0.
0
Cr:
2 0.
0 2.
1 0.
0 0.
0 0.
0 0.
0 0.
0 0.
0 0.
0 3.
9 0.
0 0.
0 0.
0 0.
2 0.
1 0.
0 0.
0
PMI
1.6
4.3
0.6
9.6
186.
8 3.
1 4.
6 7.
0 0.
0 18
8.8
0.0
0.0
0.0
0.3
0.2
0.5
0.3
PMI:
4 0.
3 2.
8 0.
0 7.
4 84
.9
1.2
4.4
11.8
19
.1
55.2
0.
0 0.
5 0.
5 1.
0 0.
6 0.
6 0.
2
C23
:1
0.2
0.5
0.0
7.4
0.0
1.6
4.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.0
0.1
C31
:x
0.0
0.0
0.0
0.0
6.4
0.0
0.0
9.9
4.3
1.0
0.0
0.7
0.0
1.7
0.6
0.5
0.0
FA a
i C15
:0
3.3
11.8
6.
0 16
3.3
3.5
0.0
0.0
442.
0 2.
3 46
9.6
0.6
3.0
3.1
3.4
1.4
1.8
0.7
FA C
16:1
w5c
3.
5 14
2.3
29.4
17
.3
2790
.9
0.0
0.0
109.
2 0.
0 51
33.4
3.
8 16
.3
5.3
17.3
9.
4 2.
0 0.
3
FA c
y C
17:0
w5.
6 1.
4 0.
0 0.
7 24
.7
0.0
0.0
0.0
62.9
0.
0 61
1.8
0.2
0.0
0.0
4.1
2.1
0.7
0.1
MA
GE
C16
:1w
5C
0.3
142.
3 6.
6 51
.7
2799
.7
5.1
174.
9 79
.8
445.
8 76
4.0
0.0
0.4
0.2
1.4
0.4
0.2
0.0
DA
GE
C30
:0
0.0
0.0
0.0
84.6
16
.8
6.3
30.6
12
7.2
64.9
21
.6
0.0
0.0
0.0
0.4
0.5
0.5
0.4
DA
GE
C32
:2a
0.0
182.
3 5.
4 7.
0 18
.1
0.0
0.0
38.7
19
1.8
35.3
0.
0 0.
0 0.
0 4.
4 1.
1 0.
7 0.
2
Arc
haeo
l 9.
1 39
.0
7.0
153.
3 22
6.5
44.4
38
0.3
252.
2 26
0.0
415.
4 0.
1 1.
5 9.
6 4.
5 2.
3 1.
4 0.
6
sn-2
-OH
-Arc
haeo
l 2.
0 11
0.3
18.3
15
.8
436.
2 19
0.7
964.
1 28
6.3
7001
.9
7599
9.5
0.2
3.7
14.5
13
.6
5.9
2.4
0.3
di-O
H-A
rcha
eol
0.5
5.9
1.3
0.0
0.0
22.4
92
.1
36.8
0.
0 0.
0 0.
0 0.
8 1.
3 1.
0 0.
4 0.
2 0.
0

Chapter IV ________________________________________________________________________
111
CHAPTER IV
Experimental approach to evaluate stability and reactivity of
intact polar membrane lipids of archaea and bacteria
in marine sediments
Pamela E. Rossela, Julius S. Lippa, Verena Heuera and Kai-Uwe Hinrichsa
aOrganic Geochemistry Group, Department of Geosciences, University of Bremen, 28334 Bremen,
Germany
Keywords: intact polar membrane lipids, biomarker, archaea, bacteria, sediment,
degradation

Chapter IV ________________________________________________________________________
112
IV.1. MANUSCRIPT
ABSTRACT
A 465-days-long incubation experiment was performed in order to asses the
stability and reactivity of archaeal and bacterial membrane lipids in anoxic marine
sediments. Subsurface sediments with low organic carbon content were spiked with both
archaeal (diglycosyl glycerodialkylglyceroltetraether, from a freeze dried and ground
microbial mat) and bacterial membrane lipid (C16-phosphatidylcholine, available as a
commercial standard), and incubated under oxygen-free conditions at 5 °C and 40 °C.
Incubations were performed using both “sterile” (previously autoclaved sediment) and
“alive or active” conditions to evaluate differences between biotic and abiotic
degradation. An overall decay for both membrane lipids, although at different rates, was
observed under sterile conditions at 5°C and 40°C, contrary to previous observations
suggesting only the occurrence of biotically mediated degradation. The degradation of
lipids under sterile conditions can be accounted by: 1) the presence of active microbial
enzymes likely derived from the microbial mat powder added to the sediments; 2) the
presence of active resistant spores even after sterilization of the sediment; 3) partial
decrease due to adsorption onto minerals, 4) partial degradation of the membrane lipids
takes place abiotically. Additionally, temperature appeared to be an important factor in
IPL degradation. In the incubations with active sediment the archaeal IPL increased at
5ºC and 40ºC, whereas the bacterial IPL only increased at 5ºC. This suggests that
microbes were growing during the experiments, although this could not be evaluated due
to the fact that both pools, degraded and newly produced IPLs, were indistinguishable.
Further improvements in future experiments are needed to better distinguish between
degraded and in situ produced IPLs as well as to evaluate the abundance of microbial
cells, the production of degradation products and the effect of adsorption processes over
time. However, our results provide an important baseline for guiding such experiments.

Chapter IV ________________________________________________________________________
113
INTRODUCTION
Intact polar lipids (IPLs) are ubiquitous in all cell membranes of living organisms.
Due to the instability of the bond between the head group and the glycerol backbone,
IPLs are assumed to be highly unstable after cell’s decay and are therefore used as
biomarkers for living biomass (White et al, 1979; Sturt et al 2004; Lipp et al., 2008).
During degradation, the cleavage of the polar head group from the intact molecule occurs,
leaving behind their apolar derivatives such as archaeol, hydroxyarchaeol or varying fatty
acid side chains. These derivatives have been commonly used in the study of modern
prokaryotic ecosystems such as those associated with anaerobic oxidation of methane
(AOM; e.g., Hinrichs et al., 2000; Blumenberg et al., 2004; Elvert et al., 2005). However,
the use of apolar lipids may be influenced by fossil biomass and therefore it does not
necessarily provide a direct evidence of active communities.
Even though IPLs are currently used as marker for living biomass, their stability
has not been studied systematically and the understanding of their reactivity is poorly
constrained. Therefore, the potential contribution of IPLs to a fossil sedimentary pool
remains unknown. In this study, we evaluated the stability and reactivity of archaeal and
bacterial IPLs, i.e., the hydrolytic cleavage of the glycosidic or phosphate-ester bond
between the polar head group and the core lipid, in a long-term experiment. A better
understanding of IPL degradation is essential because it severely affects the interpretation
of lipid biomarker signals in natural environments.
MATERIAL AND METHODS
Anoxic subsurface sediments with low organic carbon content (~0.7 wt%) were
obtained from IODP Leg 311 (Cascadia Margin; collected from 34 and 53 meter below
the sea floor). Sediments were mixed in a bottle with autoclaved artificial seawater (in
duplicate) in a 1:1 proportion to obtain 1 L of slurry per bottle, and were later incubated
at 5°C over a week to allow the formation of microbial films (Fig. IV.1). Artificial
seawater was prepared using sodium chloride (26.4 g L-1), magnesium chloride (5.7 g L-
1), potassium chloride (0.682 g L-1), potassium bromide (0.099 g L-1) and nutrients

Chapter IV ________________________________________________________________________
114
(ammonium chloride and potassium dihydrogen phosphate), the latter recommended for
culture media of sulfate reducing bacteria (SRB, Widdel and Bak, 1992). One slurry
bottle was kept at 5°C (active sediment incubation). The second bottle was autoclaved
twice (sterile conditions) with two days in between each autoclave cycle; afterwards,
water was removed and replaced for freshly autoclaved artificial seawater to avoid
contamination by potential microbe-derived spores growing in the sediment (Fig IV.1). In
parallel to the preparation of slurries, ~ 30 μg of bacterial C16-Phosphocoline (C16-PC)
and archaeal diglycosyl glyceroldialkylglyceroltetraether (2Gly-GDGTs) were introduced
in a series of Hungate tubes and stored at -80°C to avoid degradation (Fig IV.1). The
bacterial IPL correspond to a commercially available standard, whereas the archaeal IPL
is the dominant lipid in a microbial mat associated with an AOM system from the Black
Sea (R/V Logachev Cruise 2005). The occurrence of the ester lipid C16-PC has been
previously described in SRB such as Desulforhabdus amnigenus (Rütter et al., 2001),
methanotrophic bacteria (Makula, 1978) as well as photosynthetic eukaryotes
(Thompson, 1996). On the other hand, the ether lipid 2Gly-GDGT has been reported only
in methanogenic and thermogenic archaea (de Rosa et al., 1986) as well as in the
methanotrophic archaea ANME-1 (Rossel et al., 2008).
After both slurries were prepared, Hungate tubes with the IPL mixture were filled
up to completing a volume of 10 mL, and were sealed with butyl rubber stoppers
(previously sterilized) under anaerobic conditions using a glove box (Fig. IV.1). Only
anaerobic degradation was evaluated in this study because it is a most accurate
representation of the environmental conditions in which AOM take place.
Experiments were performed in order to monitor abiotic- and/or biologically-
mediated decay of IPLs. Incubations were performed under anaerobic conditions in
darkness for 465 days at 5°C and 40°C with irregular sampling intervals (Table IV.1).
Abiotic degradation of IPLs is known to occur under exposure to oxygen, light, or high
temperatures (Peterson and Cummings, 2006). Degradation of lipids and bulk AOM
biomass was monitored by analysis of IPLs and apolar GDGT cores concentrations, as
well as the concentrations and the carbon isotopic composition of the metabolite acetate
at irregular intervals (Table IV.1). For acetate analysis, an aliquot of ~1 mL was collected

Chapter IV ________________________________________________________________________
115
from the supernatant water and stored at -20°C, whereas for lipid, sediments were stored
at -80°C prior to extraction and analysis (Fig. IV.1).
Fig. IV.1. Diagram of the experimental design. 2Gly-GDGT = Diglycosyl dialkylglcerotetraether, C16-PC = C16-phosphocholine.
Acetate was analyzed by isotope ratio monitoring - liquid chromatography mass
spectrometry (irm-LC/MS) according to Heuer et al. (2006). Total lipid extracts (TLEs)
were obtained from freeze dried samples, previously stored at -80°C, with a microwave
assisted extraction systems (MARS-X, CEM, USA) for 15 min at 70°C using a modified
Bligh and Dyer method (Sturt et al., 2004). TLEs were evaporated to dryness under
nitrogen stream and stored at -80°C until IPL analysis was performed by high-
performance liquid chromatography/electrospray ionization mass spectrometry (HPLC-
ESI-MS) according to Sturt et al. (2004). For GDGT core analysis, selected TLE used for
IPL were analyzed by high-performance liquid chromatography/atmospheric pressure
chemical ionization-mass spectrometry (HPLC-APCI-MS) as described elsewhere
(Schouten et al., 2007; Lipp and Hinrichs, submitted). The quantification of the IPLs

Chapter IV ________________________________________________________________________
116
(using phosphatidyl-(N,N)-dimethylethanolamine as internal standard) and GDGTs are
expressed as the percentage relative to the initial amount (T0).
Table IV.1. Frequency of analysis performed in the experiments Days IPL at 5°C IPL at 40°C GDGT cores at 5°C Acetate at 40°C sterile active sterile active active active 0 + + + + + + 1 + + --- --- + --- 3 + + --- --- --- --- 6 + + --- --- --- --- 9 + + + + --- + 15 + + --- --- --- --- 21 + + --- --- --- --- 29 + + + + --- --- 40 --- + + + --- --- 95 + + + + + + 465 + + + + + +
RESULTS AND DISCUSSION
Sterile incubation
Generally, and despite the analytical error, a more rapid cleavage of the archaeal
IPL (glycosidic ether bond) compared to the bacterial IPL (phosphate ester bond) is
observed (Figs. IV.2 A and B). After 465 days of incubation, 14 and 16% of the initial
2Gly-GDGT was still present at 5°C and 40°C, respectively, whereas for C16-PC 46 and
~1% was present, respectively (Figs. IV.2 A and B). This result differs to previous
observations by Harvey et al. (1986) who reported a 20 fold higher degradation for the
bacterial phospholipid (phosphoethanolamine) than for a glycosidic-archaeol. However,
these authors performed incubations under oxic conditions, and suggested that the high
turnover of the phospholipid is expected due to its high solubility, making it more
accessible for enzymatic attack. Interestingly, by the end of the 40°C incubation the
decrease of bacterial IPLs was more dramatic than for the archaeal IPL. Although
stimulation of organic matter degradation rates due to higher temperatures is likely to
occur, a similar trend for both lipids is expected.
The abundance of polar lipids was highly variable through time, especially for the
bacterial IPL during the initial 50 days of the incubation. A possible mechanism
explaining such variability is adsorption of IPLs onto minerals, which seems to affect

Chapter IV ________________________________________________________________________
117
more strongly the bacterial IPL (larger error bars). The adsorption of organic matter onto
minerals facies may occur soon after deposition, providing physical protection which
decreases its availability for microbial degradation (Mayer, 1994; Keil, 1994; Hedges and
Keil, 1995). Experimental studies have reported that apparently recalcitrant organic
matter becomes more labile and it is rapidly degraded when separated from its mineral
matrix (Keil, 1994). Unfortunately, no controls to address the effect of adsorption on
IPLs were performed during our experiments. Therefore, we can not judge the
importance of this effect.
Time (days)0 50 100 150 450 500
0
10
20
30
40
50
60
70
80
90
100
110
120
130
140
150
Archaeal IPL (2Gly-GDGT) at 5°CArchaeal IPL (2Gly-GDGT) at 40°C
Time (days)0 50 100 150 450 500
0
10
20
30
40
50
60
70
80
90
100
110
120
130
140
150
Bacterial IPL (C16-PC) at 5°CBacterial IPL (C16-PC) at 40°C
A B
%IP
Ls re
lativ
e to
T0
Fig. IV.2. Degradation of archaeal 2Gly-GDGT (A) and bacterial C16-PC (B) at 5°C and 40°C in sterile sediments. Values are expressed as the percentage of the original IPL concentration at the beginning of the experiment.
The rapid decrease at the beginning of the experiment followed by a slow
turnover observed in the sterile experiment is in agreement with other observations of
IPLs degradation in natural sediments (White et al., 1979; Harvey et al., 1986). However,
contrary to previous observations by Harvey et al. (1986), who did not observed
abiotically-mediated degradation in the sterile sediments (previously autoclaved and
treated with formaldehyde), we observed a decrease of lipid abundance over time in the
sterile incubations. Degradation in sterile conditions may be caused by several reasons: 1)
it is possible that enzymes from the mat powder used as archaeal lipid standard were still
active; 2) that the used sterilization procedure (only autoclaved steps) did not efficiently
kill some resistant spores; 3) that the decrease of IPLs is due to adsorption; or 3) that an

Chapter IV ________________________________________________________________________
118
important fraction of IPLs is really degraded abiotically, contrary to the observations of
Harvey et al. (1986).
Whereas Harvey et al. (1986) performed anoxic incubations in a glove box during
the curse of the experiment, we only prepared and sealed our samples under anoxic
conditions, but were not incubated under oxygen-free environment. This opens the
possibility that the samples were exposed to oxygen during part of the experiment and
thus oxic degradation may occur. Harvey et al. (1986) observed degradation rates 40%
lower under anoxic than under oxic conditions.
Active sediment incubations
Experiments with active sediments generally showed a decrease during the first
100 days of incubation, especially at 40°C (down to 11 and 1% for the archaeal and
bacterial IPLs, respectively; Figs. IV.3 A and B). After 100 days, a subsequent increase in
the abundance of 2Gly-GDGT up to 64 and 75% at 5°C and 40°C, respectively, was
observed, while C16-PC increased only during the 5°C incubation (Figs. IV.3 A and B).
The unexpected finding of high abundances of 2Gly-GDGT at higher temperatures can be
best explained by the growth of archaeal cells in the incubated sediments used for this
experiment. Subsurface sediments have been found to contain abundant archaea
producing 2Gly-GDGT (Biddle et al., 2006; Lipp et al., 2008). Moreover, due to the
extensive hydrogen bonding capacity, glycolipids-based membranes are more stable at
higher temperatures than phospholipid-based membranes (Curatolo, 1987). The growth of
2Gly-GDGT-producing archaea in the 40°C experiment is also in agreement with
previous observations by Nauhaus et al. (2005). Based on in vitro experiments, these
authors found evidence that ANME-1, the main producer of GDGTs in AOM
environments, showed higher activity than ANME-2 at higher temperatures.
The observed decrease of IPLs over the first 100 days in the active sediment
incubation at 40°C was accompanied by a rapid release of strongly 13C-depleted acetate
(Fig. IV.3B), whereas at 5°C acetate was below detection limit. The �13C value of the
acetate pool shifted from -26‰ to -73‰ in only nine days, strongly suggesting that fresh
AOM biomass was quickly turned over into acetate. After 465 days of incubation, acetate
concentrations were up to 590 μM, and exhibited a �13C value of -90‰. This strong
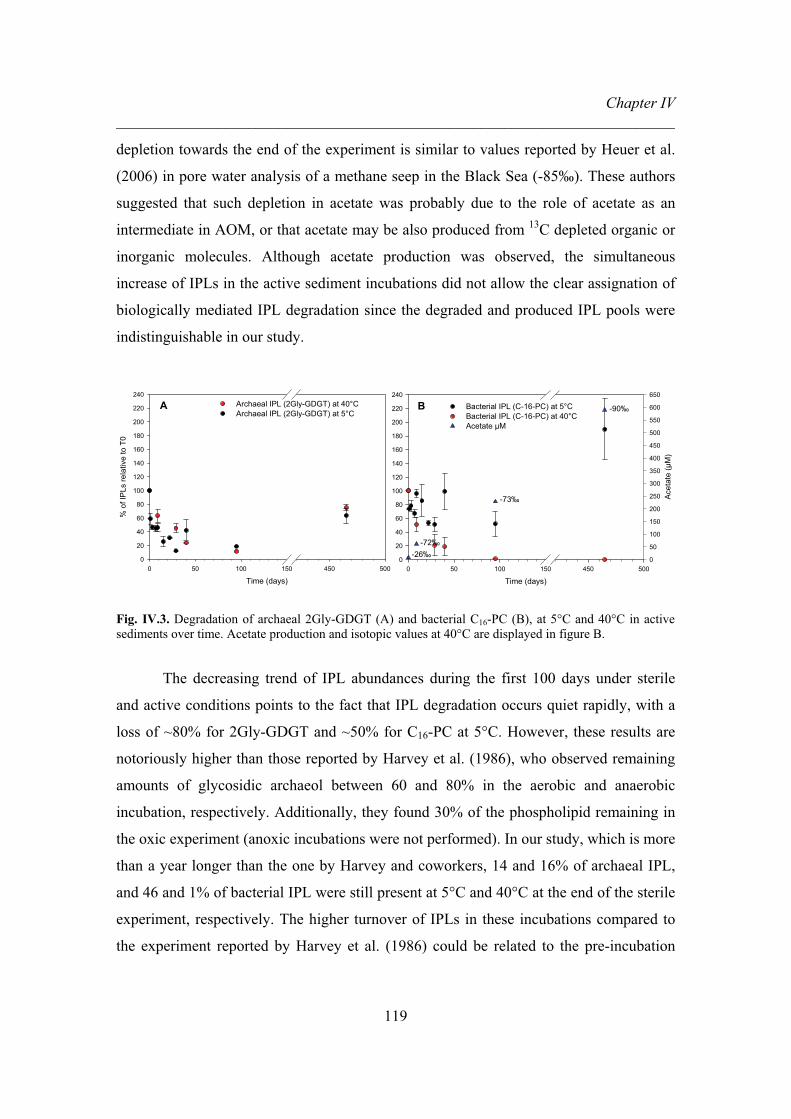
Chapter IV ________________________________________________________________________
119
depletion towards the end of the experiment is similar to values reported by Heuer et al.
(2006) in pore water analysis of a methane seep in the Black Sea (-85‰). These authors
suggested that such depletion in acetate was probably due to the role of acetate as an
intermediate in AOM, or that acetate may be also produced from 13C depleted organic or
inorganic molecules. Although acetate production was observed, the simultaneous
increase of IPLs in the active sediment incubations did not allow the clear assignation of
biologically mediated IPL degradation since the degraded and produced IPL pools were
indistinguishable in our study.
Time (days)0 50 100 150 450 500
% o
f IP
Ls re
lativ
e to
T0
0
20
40
60
80
100
120
140
160
180
200
220
240Archaeal IPL (2Gly-GDGT) at 40°CArchaeal IPL (2Gly-GDGT) at 5°C
A
D
Time (days)0 50 100 150 450 500
0
20
40
60
80
100
120
140
160
180
200
220
240
Acet
ate
(μM
)
0
50
100
150
200
250
300
350
400
450
500
550
600
650
Bacterial IPL (C-16-PC) at 5°CBacterial IPL (C-16-PC) at 40°CAcetate μM
B
-73‰
-72‰
-90‰
-26‰
Fig. IV.3. Degradation of archaeal 2Gly-GDGT (A) and bacterial C16-PC (B), at 5°C and 40°C in active sediments over time. Acetate production and isotopic values at 40°C are displayed in figure B.
The decreasing trend of IPL abundances during the first 100 days under sterile
and active conditions points to the fact that IPL degradation occurs quiet rapidly, with a
loss of ~80% for 2Gly-GDGT and ~50% for C16-PC at 5°C. However, these results are
notoriously higher than those reported by Harvey et al. (1986), who observed remaining
amounts of glycosidic archaeol between 60 and 80% in the aerobic and anaerobic
incubation, respectively. Additionally, they found 30% of the phospholipid remaining in
the oxic experiment (anoxic incubations were not performed). In our study, which is more
than a year longer than the one by Harvey and coworkers, 14 and 16% of archaeal IPL,
and 46 and 1% of bacterial IPL were still present at 5°C and 40°C at the end of the sterile
experiment, respectively. The higher turnover of IPLs in these incubations compared to
the experiment reported by Harvey et al. (1986) could be related to the pre-incubation
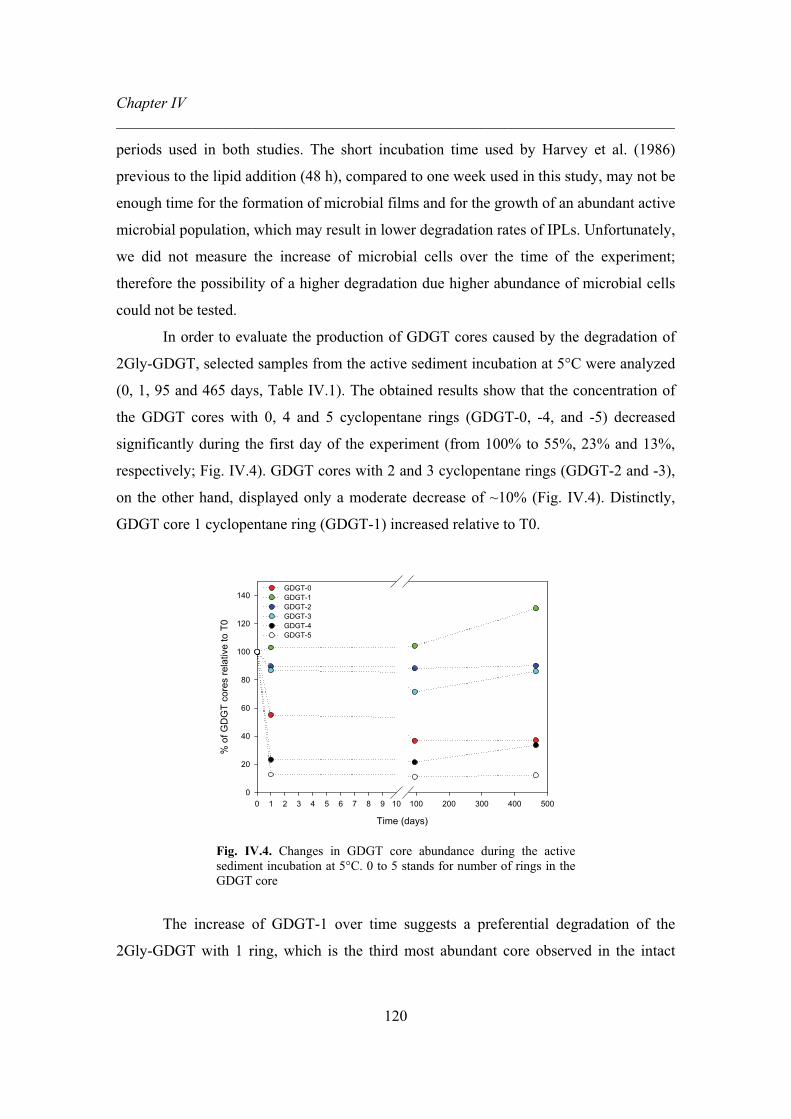
Chapter IV ________________________________________________________________________
120
periods used in both studies. The short incubation time used by Harvey et al. (1986)
previous to the lipid addition (48 h), compared to one week used in this study, may not be
enough time for the formation of microbial films and for the growth of an abundant active
microbial population, which may result in lower degradation rates of IPLs. Unfortunately,
we did not measure the increase of microbial cells over the time of the experiment;
therefore the possibility of a higher degradation due higher abundance of microbial cells
could not be tested.
In order to evaluate the production of GDGT cores caused by the degradation of
2Gly-GDGT, selected samples from the active sediment incubation at 5°C were analyzed
(0, 1, 95 and 465 days, Table IV.1). The obtained results show that the concentration of
the GDGT cores with 0, 4 and 5 cyclopentane rings (GDGT-0, -4, and -5) decreased
significantly during the first day of the experiment (from 100% to 55%, 23% and 13%,
respectively; Fig. IV.4). GDGT cores with 2 and 3 cyclopentane rings (GDGT-2 and -3),
on the other hand, displayed only a moderate decrease of ~10% (Fig. IV.4). Distinctly,
GDGT core 1 cyclopentane ring (GDGT-1) increased relative to T0.
Time (days)
0 1 2 3 4 5 6 7 8 9 10 100 200 300 400 500
% o
f GD
GT
core
s re
lativ
e to
T0
0
20
40
60
80
100
120
140GDGT-0 GDGT-1 GDGT-2 GDGT-3 GDGT-4 GDGT-5
Fig. IV.4. Changes in GDGT core abundance during the active sediment incubation at 5°C. 0 to 5 stands for number of rings in the GDGT core
The increase of GDGT-1 over time suggests a preferential degradation of the
2Gly-GDGT with 1 ring, which is the third most abundant core observed in the intact

Chapter IV ________________________________________________________________________
121
molecule from the mat used in this experiment. After a period of rather stable
concentrations during the first 100 days, relative abundances of GDGT-1, -3, and -4 also
increased, suggesting a higher degradation of intact GDGTs after this time of incubation.
However, IPL analyses showed the opposite trend, with degradation in the first 100 days
followed by a production after 100 days of incubation (Fig. IV.3A). Therefore, our results
from the active experiment, at 5°C incubation cannot be fully interpreted. Degradation of
intact GDGTs during the active experiments is not in accordance with the results from the
analyses of GDGT cores, calling for further and improved experiments which may allow
the distinction between the in situ produced and degraded IPL pools over time. A possible
solution for differentiating both pools could be the utilization of 13C labeled membrane
lipids. This approach would additionally improve the quantification of lipids over long-
time experiments, as well as the possibility to measure the production of gases and
degradation products specifically enriched in 13C.
CONCLUDING REMARKS
The results from our degradation experiments of IPLs suggest a rapid decrease of
membrane lipids (i.e., 2Gly-GDGT and C16-PC) under sterile conditions at 5°C and 40°C.
This decrease may be caused by several reasons: 1) the presence of active enzymes
derived from the added microbial mat powder with the archaeal lipid; 2) the loss of
anoxic conditions; 3) adsorption processes; 4) the presence of resistant spores even after
autoclaving the sediment; 4) the degradation of membrane lipids in marine sediments can
be partially abiotically mediated.
It was also observed that temperature, a factor not taken into account in previous
studies, affects degradation of both membrane lipids differently. During the incubation at
40°C, degradation of bacterial IPL was more dramatic than for the archaeal IPL.
Furthermore, an increase of membrane lipids after 465 days in the active sediment
incubations suggested that microbial communities were growing in situ.
Unfortunately, degraded and newly produced IPL pools were indistinguishable in
the active experiment; therefore the potential growth of microbes can not be proved.
Thus, an improved experimental design is required for future attempts. For these, not

Chapter IV ________________________________________________________________________
122
only the degraded pool and the fresh IPLs should be carefully determined, but also
problems such as possible loss of oxygen conditions, abundance of microbial cells,
degradation products and adsorption process should be evaluated over the curse of a long
term experiment.
ACKNOWLEDGMENTS
We thank the crew and shipboard scientist of IODP expedition 311 for support
during sample collection. Augusta Dibbel is gratefully acknowledged for laboratory
assistance and also Thomas Holler and Cristian Deusner from the Max Planck Institute
for Marine Microbiology in Bremen for assisting with the use of the glove box. This
study was part of the program MUMM II (grant 03G0608C), funded by the
Bundesministerium für Bildung und Forschung (BMBF, Germany) and the Deutsche
Forschungsgemeinschaft (DFG, Germany). Further support was provided from the Center
of Marine Environmental Sciences (MARUM) at the University of Bremen funded by the
DFG.
REFERENCES
Biddle, J. F., Lipp, J. S., Lever, M. A., Lloyd, K. G., Sörensen, K. B., Anderson, R.,
Fredricks, H. F., Elvert, M., Kelly, T. J., Schrag, D. P., Sogin, M. L., Brenchley, J.
E., Teske, A. House, C. H., Hinrichs, K. -U., 2006. Heterotrophic archaea dominate
sedimentary subsurface ecosystems off Peru. Proceedings of the National Academy
of Science U.S.A. 103, 3846-3851.
Blumenberg, M., Seifert, R., Reitner, J., Pape, T., Michaelis, W., 2004. Membrane lipid
patterns typify distinct anaerobic methanotrophic consortia. Proceedings of the
National Academy of Science U.S.A. 101, 11111-11116.
Curatolo, E., 1987. The physical properties of glycolipids. Biochimica et Biophysica
Acta 906, 111-136.

Chapter IV ________________________________________________________________________
123
De Rosa, M., Gambacorta, A., Gliozzi, A., 1986. Structure, biosíntesis, and
physicochemical properties of archaebacetrial lipids. Microbiological Reviews 50,
70-80.
Elvert, M., Hopmans, E.C., Treude, T., Boetius, A., Suess E., 2005. Spatial variations of
methanotrophic consortia at cold methane seeps: implications from a high-
resolution molecular and isotopic approach. Geobiology 3, 195–209.
Harvey, H. R., Fallon, R. D., Patton, J. S., 1986. The effect of organic matter and oxygen
on the degradation of bacterial membrane lipids in marine sediments. Geochimica et
Cosmochimica Acta 50, 795-804.
Hedges, J. I., Keil, R. G., 1995. Sedimentary organic matter preservation: an assesment
and speculative synthesis. Marine Chemistry 49, 81-115.
Heuer, V., Elvert, M., Tille, S., Krummen, M., Mollar, X. P., Hmelo, L. R., Hinrichs, K. -
U., 2006. Online �13C analysis of volatile fatty acids in sediment/porewater system
by liquid chromatography-isotope ratio mass spectrometry. Limnology and
Oceanography: Methods 4, 346-357.
Hinrichs, K. -U, Summons, R. E, Orphan, V., Sylva, S. P., Hayes, J. M., 2000. Molecular
and isotopic analyses of anaerobic methane-oxidizing communities in marine
sediments. Organic Geochemistry 31, 1685-1701.
Keil, R. G., Montlucon, D, B., Prahl, F. G., Hedges, J. I., 1994. Sorptive preservation of
labile organic matterin marine sediments. Nature 370, 549-552.
Lipp, J. S., Morono, Y, Inagaki, F., Hinrichs, K. -U., 2008. Significant contribution of
Archaea to the extant biomass in marine subsurface sediments. Nature 454, 991-
994.
Makula, R. A., 1978. Phospholipid composition of methane-utilizing bacteria. Journal of
Bacteriology 134, 771-777.
Mayer, L., 1994. Surface area controlof organic carbon accumulation in continental shelf
sediments. Geochimica et Cosmochimica Acta 58, 1271-1284.
Nauhaus, K., Treude, T., Boetius, A., Krüger, M., 2005. Environmental regulation of the
anaerobic oxidation of methane a comparison of ANME-1 and ANME-II
communities. Environmental Microbiology 7, 98-106.

Chapter IV ________________________________________________________________________
124
Peterson, B. L., Cummings, B. S., 2006. A review of chromatographic methods for the
assessment of phospholipids in biological samples. Biomedical Chromatography
20, 227-243.
Rossel, P. E., Lipp, J. S., Fredricks, H. F., Arnds, J., Boetius, A., Elvert, M., Hinrichs, K.
-U., 2008. Intact polar lipids of anaerobic methanotrophic archaea and associated
bacteria. Organic Geochemistry 39, 992-999.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2001. Monoalkylether phospholipids in
the sulfate-reducing bacteria Desulfosarcina variabilis and Desulforhabdus
amnigenus. Archives of Microbiology 176, 435-442.
Schouten, S., Hughet, C., Hopmans, E. C., Kienhuis, M. V. M., Sinninghe Damsté, J. S.,
2007. Analyticyl methodology for TEX86 paleothermometry by high-performance
liquid chromatography/atmospheric pressure chemical ionization-mass
spectrometry. Analytical Chemistry 79, 2940-2944.
Sturt, H. F., Summons, R. E., Smith, K. J., Elvert, M., Hinrichs, K. -U., 2004. Intact polar
membrane lipids in prokaryotes and sediments deciphered by high-performance
liquid chromatography/electrospray ionization multistage mass spectrometry-new
biomarkers for biogeochemistry and microbial ecology. Rapid Communications in
Mass Spectrometry 18, 617-628.
Thompson, G. A., 1996. Lipids and membrane function in green algae. Biochimica et
Biophysica Acta 1302, 17-45.
White, D. C., Davis, W. M., Nickels, J. S., Kind, J. D., Bobbie, R. J., 1979. Oecologica
40, 51-62.
Widdel, F., Bak, F., 1992. Gram-negative mesophilic sulfate-reducing bacteria. In: The
prokaryotes, a handbook on the biology of bacteria: ecophysiology, isolation,
identification, applications (eds. Balows A, Trüper, H. G., Dworkin, M., Harder,
W., Schleifer, K. H.), 3352-3378, 2nd edn. Springer, Berlin Heidelberg New York.

Chapter V ________________________________________________________________________
125
CHAPTER V
Diversity of intact polar membrane lipids
in marine seep environments
Pamela E. Rossela, Marcus Elvert and Kai-Uwe Hinrichsa
aOrganic Geochemistry Group, Department of Geosciences, University of Bremen, 28334 Bremen,
Germany
Keywords: intact polar membrane lipids, archaea, bacteria, seep, phospholipids,
glycolipids, non-phospholipids

Chapter V ________________________________________________________________________
126
ABSTRACT
Determination of the microbial community structure in natural habitats has been
the focus of many microbiological studies. However, most of the techniques applied are
inadequate because of their selectivity. Current approaches successfully applied to
characterize microbial communities include the analysis of intact polar membrane lipids
(IPLs). In this study structural information of IPLs from a variety of methane-bearing
environments is presented. This report provides a comprehensive spectral analysis of
IPLs from both archaea and bacteria occurring in seep environments. Analysis of lipid
extracts by high-performance liquid chromatography/electrospray ionization mass
spectrometry (HPLC-ESI-MS) provide information of the diversity in archaeal core lipids
including diphytanyl glyceroltetraether (GDGTs) and diphytanyl glyceroldiethers
(archaeols), with the latter presenting hydroxylation and cyclization in the lipid core.
Both core lipids were linked to a variety of glycosidic, phospho or mixed glycosidic and
phospho headgroups. Within the phospho headgroups, phosphatidylglycerol (PG) and
phosphatidylethanolamine (PE) were found attached to both GDGTs and archaeols,
whereas phosphatidylinositol (PI) and phosphatidylserine (PS) were only occurring with
archaeol. Additionally, bacteria produced both phospho and non-phospho lipids. Among
these, the major ones were PE and its methyl derivatives phosphatidyl-(N)-
methylethanolamine (PME) and phosphytidyl-(N,N)-dimethylethanolamine (PDME).
Bacteria derived non-phospho lipids included ornithine lipids, surfactin and betaine
lipids, with the latter characterized by odd fatty acid chains.
The results of membrane lipid analysis from a wide variety of seep environments
presented in this study confirmed the high diversity of microbes inhabiting these systems
and represent a base for further IPL studies from habitats in which anaerobic oxidation of
methane takes place.

Chapter V ________________________________________________________________________
127
INTRODUCTION
Determination of the microbial community structure in natural habitats has been
the focus of many microbiological studies. However, most of the applied methods,
including fluorescence in situ hybridization and cultures techniques, are inadequate for
several reasons: 1) they are selective methods (Wagner et al., 2003), 2) only a fraction of
the viable microbes is cultivable in laboratory conditions (MacCarthy and Murray, 1996
in fang 1998) and 3) limited information is gained about interaction between different
microbes (Findlay, 1996; Findlay et al., 1990). Current approaches to characterize
microbial communities include the analysis of intact polar membrane lipids (IPLs) by
high-performance liquid chromatography/electrospray ionization mass spectrometry
(HPLC-ESI-MS), technique that has been successfully applied in complex mixtures
(Sturt et al., 2004; Rütters et al., 2002; Biddle et al., 2006; Ertefai et al., 2008; Lipp et al.,
2008; Rossel et al., 2008; Schubotz et al., unpublished).
Methane-bearing environments represent a good opportunity to study microbial
communities using IPLs. These environments are considered as oases in which the
advection of fluids rich in methane and hydrogen sulfide support abundant
chemosynthetic life (Campbell, 2006). Communities inhabiting these systems include
sulfide oxidizing bacterial mats, diverse benthic macrofauna with methanotrophic
symbionts (Sahling et al., 2002; Levin, 2005) and the consortium of anaerobic
methanotrophic archaea (ANME) and sulfate reducing bacteria (SRB), which perform the
anaerobic oxidation methane (AOM, e.g., Hinrichs et al., 1999; Boetius et al., 2000;
Lösekann et al., 2007).
Among the diversity of IPLs observed in seep environments, several GDGTs and
archaeols with glycosidic and phospho headgroups from archaea and phospholipids from
bacteria have been observed (Rossel et al., 2008). Some of these lipids have already been
reported in other habitats such as anoxic water columns (ocean and lakes) (Schubotz et
al., unpublished; Ertefai et al., 2008) and in the deep subsurface (Biddle et al., 2006; Lipp
et al., 2008).

Chapter V ________________________________________________________________________
128
In order to evaluate the structural diversity of IPLs in seep systems using HPLC-
ESI-MS, a comprehensive spectral interpretation of both archaeal and bacterial IPLs from
these natural environments is provided.
MATERIAL AND METHODS
IPL analysis
Total lipid extracts of the samples were obtained with an automated microwave-
assisted extraction system (MARS-X, CEM, USA) for 15 minutes at 70°C or via
ultrasonication, using a modified Bligh and Dyer method (Sturt et al., 2004)
IPL analysis was performed with an HPLC system equipped with an ion-trap
mass spectrometer (ThermoFinnigan LCQ Deca XP) with an electrospray ionization
source (ESI) using protocols described previously by Sturt et al. (2004) and Biddle et al.
(2006). Briefly, a LiChospher Diol column (125mm x 2 mm, 5μm; Alltech Associates
INC., Deerfield, Il, USA) was used isothermally at 30°C in a ThermoFinnigan Surveyor
HPLC system. The following linear gradient was applied with a flow of 0.2 mL min-1:
100% A to 35% A: 65% B over 45 min, hold for 20min, then back to 100% A for 1 h to
equilibrate the system for the next injection, where A = 72:20:0.12:0.04 of hexane/2-
propanol/formic acid/14.8 M NH3aq and B = 88:10:0.12:0.04 of 2-propanol/water/formic
acid/14.8 M NH3aq..
Structural assignments were based on characteristic fragmentation patterns (cf.
Sturt et al., 2004) and by comparison with IPL inventories of cultured archaea and
bacteria (e.g., Koga et al., 1998; Koga and Morii, 2005; Hinrichs et al., unpublished
data). Individual IPLs were extracted by using the quasi-molecular ions obtained from the
full scan (m/z 500–2000), from which the MS2 daughter ion spectra information were
obtained. A general overview of the diversity of IPLs is given in Table V.1. The IPLs
identified are described according to their lipid class (i.e., glycolipids, phospholipids and
non-phospho lipids).

Chapter V ________________________________________________________________________
129
RESULTS AND DISCUSSION
IPL identification
1. Glycolipids
Carbohydrate-containing lipids are abundant molecules within thermophilic
archaea and bacteria (Langworthy, 1982). In some cases, these lipids are the major
components of the cytoplasmic membrane, especially in microbes without cell walls or
inhabiting hostile environments (Curatolo, 1987a). Most of the proposed functions for
glycolipids are based on their physical properties, which generally suggest that these
molecules participate in the stabilization, shape, extracellular recognition and ion bonding
in the membrane (Curatolo, 1987b).
Among the samples analyzed, the observed glycolipids were only associated with
archaea. Glycolipids have been suggested to be widely distributed among gram-positive
bacteria but rarely in gram-negative bacteria (López-Lara et al., 2003), which may
suggest low abundance of the former in seep environments. Within the archaeal
glycolipids, several archaeol-based IPLs (archaeol and hydroxyarchaeols) and GDGT-
based IPLs are described below.
1.1. Archaeol-based IPLs
Glycosidic archaeol-based IPLs ranged from archaeols (sn-2,3-diphytanyl
glycerol) containing 1 and 2 glycosidic headgroups (2Gly-AR, Fig. V.1a) to archaeols
with varying chain length, hydroxylation (extended archaeol, Fig. V.1b) and cyclization
(macrocyclic archaeol, Fig. V.1c). The occurrence of glycosidic archaeols has been
shown to be characteristic of the order Methanosarcinales (Koga et al., 1998).
Furthermore, the decrease in the proportion of archaeol relative to macrocyclic archaeol
as a response to the increase in the growth temperature has been observed in
Methanococcus jannaschii, a microbe isolated from hydrothermal vent systems (Sprott et
al., 1991).
During the fragmentation of archaeols, the MS2 daughter ion spectra exhibit a
main fragment indicative of the archaeol core (653 Da), together with another diagnostic
fragment (373 Da), which corresponds to one phytanyl chain with the glycerol positively

Chapter V ________________________________________________________________________
130
charged. Differently, the MS2 daughter ion spectra of hydroxylarchaeols show as major
fragment the loss of 296 Da, which represents the phytanyl chain with the hydroxyl
group. This pattern was consistently observed in all hydroxyarchaeols present in the
samples from this study.
Fig. V.1. MS2 positive ion spectra of several archaeol-based IPLs with glycosidic or mixed glycosidic and phospho headgroups. a) diglycosidic-archaeol (2Gly-AR), b) tentative glycosidic phospho-hydroxyarchaeol (Gly-P-OH-AR) and c) glycosidic-macrocyclic archaeol (Gly-MAR).
1.2. GDGT-based IPLs
Despite the occurrence of 1 to 8 cyclopentane rings depending of the growth
conditions (De Rosa et al., 1986), the C40 phytanyl chains of glycolipid tetraethers
(GDGTs) do not vary in length or saturation degree. However, GDGTs exhibit a wide
structural variety as is shown in this study. Glycosidic-GDGTs have been observed in
cultures of Sulfolobus schibatae and Methanobacterium thermoautotrophicum, the latter
also presenting GDGTs with mixed glycosidic and phospho headgroups (Koga et al.,
1993; Sturt et al., 2004).
The dominance of glycosidic headgroups in GDGTs has been reported in the deep
subsurface (Biddle et al., 2006; Lipp et al., 2008). Similarly, diglycosyl-GDGT (2Gly-

Chapter V ________________________________________________________________________
131
GDGT, Fig. V.2a) and 2Gly-GDGT with 18 Da more (Fig. V.2b) (H342-GDGT, Lipp et
al., 2008) have also been observed in seeps. The MS2 daughter ion spectrum of 2Gly-
GDGT shows the GDGT core as the most prominent fragment (Fig. V.2a). On the other
hand, the MS2 of 2Gly-GDGT with the additional 18 Da shows that this GDGT seems to
initially lose the diglycosyl headgroup with an ammonium adduct (1314 Da although
small fragment was observed), followed by a loss of 18 Da more (1314 Da to 1296 Da)
(Fig. V.2b). At this moment it is unclear if the additional 18 Da are contained in the
GDGT core or in the headgroup as previously suggested (GDGT core is the only
fragment observed in MS2, Lipp et al., 2008; Lipp and Hinrichs, submitted).
Nevertheless, the retention time and quasi-molecular ion information suggest that this
GDGT is the same as the one reported in deep subsurface environments. Furthermore,
although this lipid is present in seep environments, it is not the most abundant GDGT.
Other glycosidic GDGTs observed in seep environments included GDGTs with
up to 4 sugars (Fig. V.2c) and mixed glycosidic and phospho headgroups (Fig. V.2d and
e), as well as another 2Gly-GDGT with an additional unknown head group of 145 Da
(Fig. V.2e).
The MS2 daughter ion spectra of GDGTs show that GDGT containing only
glycosidic headgroups have the GDGT core as the most prominent fragment. However,
GDGTs containing both glycosidic and phospho headgroups lose first the glycosidic
headgroup and therefore the major fragment in MS2 is the GDGT core with the phospho
headgroup. Furthermore, during the fragmentation of some GDGTs with mixed
glycosidic and phospho headgroups it is also possible to observe two fragments in the
MS2: one indicative of the GDGT core with the phospho headgroup and another with the
GDGT core alone. Although we can not confirm the position of the glycosidic and
phospho headgroups, it has been previously suggested that in methanogens these two
headgroups are located in opposite ends of the GDGT core (Kates, 1997).

Chapter V ________________________________________________________________________
132
Fig. V.2. MS2 positive ion spectra of several glyceroldialkylglyceroltetraether (GDGT) based IPLs with glycosidic or mixed glycosidic and phospho headgroups. a) 2Gly-GDGT, b) 2Gly-GDGT+18, c) tetraglycosidic-GDGT (4Gly-GDGT), d) 2Gly-GDGT-phosphatidylglycerol (2Gly-GDGT-PG) and e) 2Gly-GDGT+ unknown head group of 145 Da (2Gly-GDGT+145).

Chapter V ________________________________________________________________________
133
2. Phospholipids
According to the fluid mosaic model, the primary function of phospholipids is to
define the permeability of the cell membrane (Madigan et al., 2003). Phospholipids are
also involved in solute transport, cell signaling as well as cell to cell recognition
(Madigan, et al., 2003). Additionally, phospholipids regulate the membrane structure by
modifying the headgroups, unsaturation degree and chain length of the acyl chains
(Hasegawa et al., 1980; Langworthy, 1982).
Phospholipids are ubiquitous in bacteria, however, the occurrence of archaeol and
GDGT with phospho headgroups among archaea is also widely distributed (Kates, 1997).
2.1 Phospholipids derived from archaea
Several phospholipids were observed in AOM environments. Among them,
archaeal phospholipids included phosphatidylglycerol (PG), phosphatidylinositol (PI),
phosphatidylserine (PS) and phosphatidylethanolamine (PE) linked to archaeol (Fig. V.3a
to 3e), whereas the common headgroups linked to GDGT were only PG and PE (Fig.
V.4a and b). The occurrence of PG, PI, PS and PE with GDGT and archaeol has been
frequently observed in methanogens (Kates, 1997; Sprott, 1992). Nevertheles, PG-
archaeol, which has been reported in Methanosarcina mazei (Sprott, 1992), is also a
characteristic IPL of Halophiles (Kates, 1997).
In agreement with the fragmentation pattern of hydroxyarchaeol described for
glycosidic-archaeol based IPL (section 1.1), the loss of the phytanyl chain with the
hydroxyl group (loss of 296 Da) was also observed in the MS2 daughter ion spectra of
the hydroxyarchaeols with phospho headgroups. Furthermore, the occurrence of 373 Da
fragment in MS2 previously described was also observed here. Nevertheless, the most
common fragment for phospho-hydroxyarchaeols in MS2 was 453 Da, which
corresponds to one phytanyl chain with the phosphate group. Different from glycosidic-
archaeols, the MS2 daughter ion spectra from archaeols with phospho headgroups show
the archaeol core with the phosphate group (733 Da). In addition to the previously
described phospho-archaeol based IPLs, the occurrence in ANME-2 dominated sediments
and carbonate mats of another archaeol with an unknown phospho headgroup of 223 Da
was observed.

Chapter V ________________________________________________________________________
134
Fig. V.3. MS2 positive ion spectra of diverse archaeol-based IPLs with phospho headgroups. a) PG-OH-AR, b) PI-OH-AR, c) PS-AR, c) PS-OH-AR, d) PE-OH-AR and e) archaeol with unknown phospho headgroup of 223 Da (P-AR+223).

Chapter V ________________________________________________________________________
135
Differently from the glycosidic GDGTs, the MS2 daughter ion spectra of the
GDGTs with phospho headgroups usually show two or three fragments: 1) fragment
indicative of the GDGT core with one of the two phospho headgroups, 2) the GDGT core
with only the phosphate group and 3) the GDGT core alone (Fig. V.4.).
Fig. V.4. MS2 positive ion spectra of diverse GDGT-based IPLs with phospho headgroups. a) PE-GDGT-PG and b) 2PG-GDGT.
2.2 Phospholipids derived from bacteria
The most common bacterial phospholipids observed were PE and its methyl
derivatives phosphatidyl-(N)-methylethanolamine (PME) and phosphytidyl-(N,N)-
dimethylethanolamine (PDME) (Fig. V.5a to V.5c). PE has been found to be the most
dominant IPL in SRB such as Desulfosarcina variabilis and Desulforhabdus amnigenus
(Rütter et al., 2001; Sturt et al., 2004). PME and PDME have been reported in
methanotrophic bacteria such as Methylosinas trichosporium and Methylobacterium
organophilum (Makula, 1978; Goldfine, 1984; Fang et al., 2000) as well as in sulfide
oxidizers (Barridge and Shively, 1968). The MS2 daughter ion spectra of the
diacylglycerol phospholipids PE, PME and PDME show the loss of their phospho
headgroups (141, 155 and 169 Da, respectively). This is the same fragmentation pattern

Chapter V ________________________________________________________________________
136
of phospholipids previously described in literature (e.g., Rütter et al., 2001; Sturt et al.,
2004).
Fig. V.5. MS2 positive ion spectra of the major bacterial phospholipids observed in seep environments. As examples a) PE-DAG C32:2, b) PME C32:2, c) PDME C34:2. DAG = diacylglycerol
3. Phosphorus-free membrane lipids
The production of phosphorus-free membrane lipids has been suggested to occur
in organism exposed to physiological stress conditions such as limitation of nitrogen or
phosphate (López-Lara et al., 2003). In our data set three main types of bacterial IPLs
which do not contain carbohydrates or phospho headgroups were identified. The
occurrence of some of them has been related to phosphate limitation conditions during
the growth (e.g., ornithine and betaine lipids), whereas others seem to provide surface
active properties to the membrane containing these molecules (e.g., surfactin, Vater,
1986).
3.1. Ornithine lipids
Ornithine lipids (OL) contain one amidified 3-hydroxy fatty acid to which another
fatty acid residue is attached (López-Lara et al., 2003). OL are widely spread among

Chapter V ________________________________________________________________________
137
gram-negative bacteria (Imhoff and Bias-Inhoff, 1995) involved in sulfate reduction
(Desulfovibrio gigas), sulphur oxidation (Thiobacillus thiooxidans) and iron metabolism
(Rhodomicrobium vannielii) (Makula and Finnerty, 1975; Knoche and Shively, 1972).
High abundance of OL seems to substitute phospholipids such as PE, PG and DPG, in
Pseudomonas fluorescens in response to change towards phospho-limited conditions
(Minnikin and Abdolrahimzadeh, 1974). Moreover, it has also been suggested that OL
partially control the iron oxidation metabolism in Thiobacillus ferrooxidans (Ghosh and
Misha, 1987).
The MS2 daughter ion spectra of OL usually show three fragments, with the first
indicative of the ornithine with one fatty acid, and the other two corresponding to the two
consecutive losses of 18 Da, indicative of loss of two molecules of water according to the
OL fragmentation pattern reported by Aygun-Sunar, et al. (2006) (Fig V.6.).
Fig. V.6. MS2 positive ion spectrum of C34:1 OL as an example for ornithine lipids.
3.2. Betaine lipids with odd fatty acid chains
Betaine ether linked glycerolipids (BL) are membrane components widely
distributed among higher plants, algae, protozoa and some fungi (Dembitsky, 1996).
Their structure in aquatic algae is frequently characterized by the presence of C14, C16,
C18, C20 and C22 fatty acids (Sato et al., 1992; Dembitsky, 1996). Their synthesis in
bacteria has been observed in the anoxygenic photosynthetic bacterium Rhodobacter
sphaeroides (Benning et al., 1995; Hoffman and Eichenberger, 1996) and in plant-
associated bacteria such as Sinorhizobium meliloti (Lopez-Lara et al., 2003). It has been
documented that phosphate-deprived cells of Rhodobacter s. growing in phosphate

Chapter V ________________________________________________________________________
138
concentrations <0.1mM can decrease their membrane phospholipid content from 90% to
22% (Benning et al., 1995).
Characteristic fatty acids of BL from bacteria have not been reported, but detailed
description of the BL from algae, which are composed by fatty acids with even carbon
numbers, suggest that the BL containing odd fatty acids chains (BL-odd) may be bacterial
derived. This has been previously suggested by Schubotz et al. (submitted), who
observed an increase of BL content with C15 and C17 fatty acids in the anoxic water from
the Black Sea.
In general the MS2 daughter ion spectra of BL show four fragments; the first and
second fragments correspond to the fatty acid chain in the sn1 position with and without
hydroxyl group, whereas the third and fourth fragments correspond the fatty acids in the
sn2 position with and without hydroxyl group (Fig V.7).
Fig. V.7. MS2 positive ion spectrum of C31:1 BL-odd as an example for betaine lipids.
3.3. Surfactins
Biosurfactants are molecules of interest in biotechnology and are grouped in five
different classes: glycolipids, phospholipids, lipopeptides/lipoproteins, polymeric
surfactants and particulate surfactants (Muthusamy et al., 2008). Among these classes,
surfactins, which are macrocyclic heptapeptides linked to a long-chain �-hydroxy fatty
acid (Hue et al., 2001), are considered as one of the most powerful biosurfactants (Vater,
1986). The cyclic form of surfactins results from the link between the hydroxyl group of
the fatty acid with the C-terminal carbonyl to form a lactone ring (Fig. V.8) (Hue et al.,
2001). Glutamic acid, leucine, valine and aspartic acid are the common amino acids

Chapter V ________________________________________________________________________
139
forming the ring. Surfactin structure has been shown to vary in both amino acid
composition and acyl chain length, the latter found with 12 and 15 carbon atoms (Hue et
al., 2001).
Several are the properties assigned to surfactins, including surface active (Vater et
al., 1986), antibiotics (Georgiou et al., 1992) and antifungal (Thimon et al., 1992), among
others. The surface active properties of surfactin have been suggested to increase
significantly when glutamic and aspartic acids are present (Georgiou et al., 1992).
The production of these molecules is affected by several factors. The carbon
source present in the system (usually hydrocarbons or carbohydrates) can influence not
only the surfactant production but also their structure, especially the hydrophobic tail
(Georgiou et al., 1992). Furthermore, temperature, pH and oxygen seem to affect
surfactant production as well (Kim et al., 1990; Gerson and Zajic, 1978). It has also been
reported that surfactin production is enhanced by the increase in iron and manganese
concentrations in the growth media (Cooper et al., 1981).
The MS2 daughter ion spectra of surfactins (Fig. V.8) were characterized by a
prominent 685 Da fragment, which corresponds to the loss of the protonated peptide (six
out of seven aminoacids with H+). The main quasi-molecular ion present is 1036.5, which
corresponds to a surfactin with glutamic acid, leucine, leucine, valine, aspartic acid,
leucine and leucine amino acids and a hydroxy fatty acid iso-C15. Other quasi-molecular
ions observed within the surfactin peak were 1008.5 and 1022.5, which indicate the
change of the hydroxy fatty acid from 15 to 13 and 14 carbon atoms, respectively.
Fig. V.8. MS2 positive ion spectrum of surfactin with glutamic acid, leucine, leucine, valine, aspartic acid, leucine and leucine amino acids and a hydroxyfatty acid iso-C15.

Chapter V ________________________________________________________________________
140
4. Unknown IPLs
Two unknown IPLs frequently observed in the analyzed samples were IPL a and
b, the first represented by the two quasi molecular ions 734.3 and 706.4 m/z, and the
second by the quasi molecular ion 1148.0 m/z. The MS2 daughter ion spectrum of IPL a
in positive mode (Fig. V.9a and b) shows three main fragments (losses of 193.5 Da, 18
Da and 46 Da). Information obtained from negative ion mode for 706.4 m/z indicate that
the molecule has 18 Da less when is negatively charged (687.5 m/z), which is also
indicated in the MS2 by a loss of 175.2 Da instead of 193.5 Da (Fig. V.9c). Additionally,
the occurrence of two other fragments in the MS2 of the negative ion mode, indicates the
presence of the fatty acids C17:1 and C16:1. The occurrence of these lipids in carbonate
mats from the Black Sea, together with presence of fatty acids in their lipid structure,
suggest that these lipids are bacterial derived.
The MS2 daughter ion spectrum of IPL b (Fig. V.9e) shows several fragments.
The first is 993.6 Da, which could be analogical to the diglycosyl archaeol core after loss
of 155 Da (possible analog to PME). However, the major fragment in the MS2 daughter
ion spectrum was 873.4 Da, which results from a consecutive loss of 120 Da.
Unfortunately, negative ion mode for this lipid was always very noisy and did not
provide additional information about its molecular structure. Unknown b was frequently
observed in ANME-2 dominated sediments.

Chapter V ________________________________________________________________________
141
Fig. V.9. MS2 positive ion spectra of two unknowns frequently observed in seep environments. a) unknown IPL a with quasi molecular ion 734.3 m/z b) unknown IPL a with quasi molecular ion 706.4 m/z c) MS2 daughter ion spectrum in negative mode for unknown IPL a 706.5, which is 687.5 m/z due to a loss of 18 Da and d) unknown IPL b with quasi molecular ion 1148.0 m/z.

Cha
pter
V
_
142
Tab
le V
.1. I
ntac
t pol
ar m
embr
ane
lipid
div
ersi
ty in
seep
env
iron
men
ts
Lip
id n
ame
RT
R
ange
of q
uasi
m
olec
ular
ions
N
eutr
al lo
ssa
diag
nost
ic
ion
in M
S2
neut
ral l
oss o
r di
agno
stic
fr
agm
ent i
n M
S2 r
epre
sent
s O
bser
ved
in:
2Gly
-AR
-0
.68
994.
6 [M
+ N
H4]+1
34
1 65
3 Lo
ss
of
digl
ycos
yl
head
gr
oup
with
an
NH
4 add
uct
Arc
haea
, AN
ME-
2 (R
osse
l et
al.,
200
8), M
etha
noca
ldoc
occu
s ja
nnas
chii
(Stu
rt et
al.,
200
4), d
eep
subs
urfa
ce (B
iddl
e et
al.,
200
6; L
ipp
et a
l., 2
008)
Te
ntat
ive
Gly
-P-O
H-A
R
exte
nded
-0.9
5 98
1.7
[M+H
]+1
296
685
Loss
of
phyt
anyl
cha
in w
ith a
n hy
drox
yl g
roup
A
rcha
ea, p
ossi
bly
AN
ME-
2 (th
is s
tudy
), ar
chae
ols
with
C25
cha
in h
ave
been
pre
viou
sly
repo
rted
in
extre
me
Hal
ophi
les (
Kog
a et
al.,
199
3; 2
008)
and
in c
old
seep
sedi
men
ts fr
om E
aste
rn M
edite
rran
ean
Sea
(Sta
dnits
kaia
et a
l., 2
008)
G
ly-M
AR
-0
.75
831.
2 [M
+NH
4]+1
180
651
Loss
of
glyc
osyl
hea
d gr
oup
with
an
NH
4 add
uct
Arc
haea
, AN
ME-
2 (th
is st
udy)
, Met
hano
cald
ococ
cus j
anna
schi
i (St
urt e
t al.,
200
4)
PG-O
H-A
R
0.54
82
3.4
[M+H
]+1
296
527,
453
Lo
ss o
f ph
ytan
yl c
hain
with
an
hydr
oxyl
gro
up
Arc
haea
, A
NM
E-2
(Ros
sel
et a
l., 2
008)
, M
etha
nosa
rcin
a ba
rker
i (K
oga
and
Mor
ii et
al.,
200
5),
Hal
ophi
les (
Kat
es, 1
997)
PI
-OH
-AR
1.
00
911.
5 [M
+H]+1
29
6 61
5, 4
53
Loss
of
phyt
anyl
cha
in w
ith a
n hy
drox
yl g
roup
A
rcha
ea, A
NM
E-2
and
AN
ME-
3 (R
osse
l et a
l., 2
008)
PS-O
H-A
R
-0.9
0 83
6.4
[M+H
]+1
296
540,
453
Lo
ss o
f ph
ytan
yl c
hain
with
an
hydr
oxyl
gro
up
Arc
haea
, AN
ME-
2 an
d A
NM
E-3
(Ros
sel e
t al.,
200
8), M
etha
nosa
rcin
a ba
rker
i (K
oga
et a
l., 1
993)
PS-A
R
-0.8
7 82
0.4
[M+H
]+1
87
733,
453
Lo
ss o
f ser
ine
Arc
haea
, A
NM
E-2
and
AN
ME-
3 (R
osse
l et
al.,
200
8),
Met
hano
bact
eriu
m t
herm
oaut
otro
phic
um
(Kog
a et
al.,
199
3), M
etha
noca
ldoc
occu
s jan
nasc
hii (
Stur
t et a
l., 2
004)
PE
-OH
-AR
-0
.72
792.
4 [M
+H]+1
29
6 49
6, 4
53
Loss
of
phyt
anyl
cha
in w
ith a
n hy
drox
yl g
roup
A
rcha
ea,
AN
ME-
2 (R
osse
l et
al
., 20
08),
Met
hano
thri
x so
ehng
enii
(Kog
a et
al
., 19
93);
Met
hano
sarc
ina
bark
eri (
Kog
a an
d M
orii
et a
l., 2
005)
2G
ly-G
DG
T -0
.72
1632
.1-1
645.
1b [M
+NH
4]+1
341
GD
GT
core
Lo
ss
of
digl
ycos
yl
head
gr
oup
with
an
NH
4 add
uct
Arc
haea
, AN
ME-
1 (R
osse
l et a
l., 2
008)
and
dee
p su
bsur
face
(B
iddl
e et
al.,
200
6; L
ipp
et a
l., 2
008;
St
urt e
t al.,
200
4), S
ulfo
lobu
s sh
ibat
ae (
Stur
t et a
l., 2
004)
, Met
hano
bact
eriu
m th
erm
oaut
otro
phic
um
(Kog
a et
al.,
199
3)
2Gly
-GD
GT+
18
-0.7
7 16
50.1
-166
1.1b
[M+N
H4]+1
34
1 or
360
13
14,
GD
GT
core
Lo
ss
of
digl
ycos
yl
head
gr
oup
with
an
NH
4 ad
duct
or
unkn
own
head
gro
up o
f 34
2 D
a w
ith a
n N
H4 a
dduc
t (36
0 D
a)
Arc
haea
in
deep
bio
sphe
re s
edim
ents
(Li
pp a
nd H
inric
hs,
unp
ublis
hed
data
) an
d N
itros
opum
ilus
mar
itim
us (S
chou
ten
et a
l., 2
008)
4Gly
-GD
GT
+1.1
0 19
58.1
-196
9.1b
[M+N
H4]+1
66
7 G
DG
T co
re
Loss
of
tetra
glyc
osyl
hea
d gr
oup
with
an
NH
4 add
uct
Arc
haea
, AN
ME-
1 (th
is st
udy)
2Gly
-GD
GT-
PG
+1.0
6 17
87.1
-179
8.1b
[M+N
H4]+1
34
1 G
DG
T co
re+P
G
Loss
of
di
glyc
osyl
he
ad
grou
p w
ith a
n N
H4 a
dduc
t A
rcha
ea, A
NM
E-1
(this
stud
y), M
etha
nosp
irill
um h
unga
tei (
Kog
a et
al.,
199
3)
2Gly
-GD
GT+
145
-0.6
9 17
78.1
-178
9.7b
[M+N
H4]+1
34
1 G
DG
T co
re+1
45
Loss
of
di
glyc
osyl
he
ad
grou
p w
ith a
n N
H4 a
dduc
t A
rcha
ea, A
NM
E-1
(this
stud
y)
PE-G
DG
T-PG
+1
.06
1569
.1-1
579.
7b [M
+H]+1
15
4, 4
3 G
DG
T+PE
Lo
ss o
f PG
with
the
pho
spha
te
grou
p w
ithou
t on
e ox
ygen
, fo
llow
ed
by
the
lost
of
et
hano
lam
ine
Arc
haea
, AN
ME-
1(th
is st
udy)
2PG
-GD
GT
+1.0
9 16
00.1
-161
0.1b
[M+H
]+1
154,
74
GD
GT+
PG
Loss
of
PG w
ith t
he p
hosp
hate
gr
oup
with
out
one
oxyg
en,
follo
wed
by
the
lost
of g
lyce
rol
Arc
haea
, AN
ME-
1 (th
is st
udy)
PE (D
AG
) -0
.76
608.
6-74
4.5c
[M+H
]+1
141
Fatty
ac
ids
+gly
cero
l Lo
ss o
f PE
Met
hano
troph
ic b
acte
ria (M
akul
a, 1
978;
Gol
dfin
e, 1
984;
Fan
g et
al.,
200
0), D
esul
fosa
rcin
a va
riabi
lis
(Rüt
ters
et a
l., 2
001;
Stu
rt et
al.,
200
4)
PME
(DA
G)
-075
70
2.4-
802.
5c [M
+H]+1
15
5 Fa
tty
acid
s +g
lyce
rol
Loss
of P
ME
Met
hano
troph
ic b
acte
ria (
Mak
ula,
197
8; G
oldf
ine,
198
4; F
ang
et a
l., 2
000)
, su
lfide
oxi
dize
r (B
arrid
ge a
nd S
hive
ly, 1
968)
PD
ME
(DA
G)
-0.7
5 71
6.6-
746.
6c [M
+H]+1
16
9 Fa
tty
acid
s +g
lyce
rol
Loss
of P
DM
E M
etha
notro
phic
bac
teria
(M
akul
a, 1
978;
Gol
dfin
e, 1
984;
Fan
g et
al.,
200
0),
sulfi
de o
xidi
zer
(Bar
ridge
and
Shi
vely
, 196
8)
OL
-0.8
2 59
7.5-
721.
5c [M
+H]+1
--
----
----
--
Orn
ithin
e+
fatty
aci
d Lo
ss o
f on
e fa
tty a
cid
chai
n an
d re
mai
n th
e hy
drox
yl
fatty
ac
id
with
the
orni
thin
e he
adgr
oup
Bac
teria
gra
m-n
egat
ive
perfo
rmin
g su
lfur
redu
ctio
n, s
ulfu
r an
d iro
n ox
idat
ion
(Mak
ula
and
Fine
rty,
1975
; Kno
che
and
Shiv
ely,
197
2; Im
hoff
and
Bia
s-Im
hoff
, 199
5)
BL
-0.6
8 71
6.6-
746.
6c [M
+H]+1
--
----
----
--
236
Indi
cativ
e of
th
e be
tain
e he
adgr
oup
BL
with
C14
, C16
, C18
fat
ty a
cids
der
ived
fro
m a
quat
ic a
lgae
(D
embi
tsky
, 199
6), B
L w
ith o
dd f
atty
ac
ids s
uch
as C
15 a
nd, C
17 s
eem
to d
eriv
e fr
om b
acte
ria d
ue to
thei
r occ
urre
nce
in a
noxi
c w
ater
s fr
om
the
Bla
ck S
ea (S
chub
otz
et a
l., su
bmitt
ed; t
his s
tudy
) Su
rfac
tin
-0.5
9 10
08.5
-103
6.0d
[M+H
]+1
----
----
----
68
5 Lo
ss
of
the
prot
onat
ed
pept
ide
(six
out
of
seve
n am
inoa
cids
with
H
+ )
Baci
llus
sp (
Vat
er, 1
986)
, unk
now
n ba
cter
ia d
ue to
thei
r oc
curr
ence
in th
e bl
ack
nodu
les
from
the
Bla
ck S
ea m
ats (
this
stud
y)
Unk
now
n a
-0.7
3 70
6.3
and
734.
3 19
4 an
d th
en 1
8 --
----
----
--
----
----
----
O
bser
ved
in c
arbo
nate
mat
s fro
m th
e B
lack
Sea
U
nkno
wn
b +1
.3
1148
.0
993.
6 an
d 87
3 --
----
----
--
----
----
----
Fr
eque
ntly
obs
erve
d in
AN
ME-
2 do
min
ated
sedi
men
ts
RT=
rete
ntio
n in
dex
rela
tive
to C
16-P
AF
inte
rnal
sta
ndar
d, a
The
neut
ral l
oss
resu
lts fr
om th
e lo
ss o
f the
hea
dgro
up p
lus
the
nece
ssar
y [H
]+ to c
harg
e th
e co
re o
f the
IPL
in M
S2, b
Ran
ge o
f mas
ses
cons
ider
a G
DG
T co
re w
ith 0
to 5
cy
clop
enta
ne r
ings
; c Inc
lude
fat
ty a
cids
of
diff
eren
t len
gth
and
satu
ratio
ns; d I
nclu
de s
urfa
ctin
mol
ecul
es w
ith g
luta
mic
aci
d, le
ucin
e, le
ucin
e, v
alin
e, a
spar
tic a
cid,
leuc
ine
and
leuc
ine
and
the
hydr
oxyl
fat
ty a
cids
with
13,
14
and
15
carb
on a
tom
s. A
bbre
viat
ions
: AR
= a
rcha
eol,
BL
= be
tain
e lip
ids,
2Gly
= d
igly
cosy
l, D
AG
=dia
cylg
lyce
rol,
GD
GT
= gl
ycer
oldi
alky
lgly
cero
ltetra
ethe
r, O
H-A
R =
hyd
roxy
arch
aeol
,, O
L =
orni
thin
e lip
ids,
PDM
E =
phos
phat
idyl
-(N
,N)-
dim
ethy
leth
anol
amin
e, P
E =
phos
phat
idyl
etha
nola
min
e, P
G =
pho
spha
tidyl
gylc
erol
, PI =
pho
spha
tidyl
inos
itol,
PME
= ph
osph
atid
yl-(
N)-
met
hyle
than
olam
ine,
PS
= ph
osph
atid
ylse
rine.

Chapter V ________________________________________________________________________
143
CONCLUDING REMARKS
Using HPLC-ESI-MS a total of 25 different IPL structures observed in seep
environments were discussed. The interpretation of mass spectra provided useful
structural information of archaeal lipids including GDGTs and archaeols linked to a
variety of glycosidic and phospho headgroups such as diglycosyl, tetraglycosyl, PG, PE
PI and PS. Bacterial IPLs commonly observed included the phospholipids PE, PME and
PDME as well as non-phospho lipids such as ornithine lipids, surfactin and betaine lipids,
with the latter characterized by odd fatty acid chains.
These results show the potential of intact polar membrane lipid analysis in the
evaluation of microbial diversity in a variety of methane-bearing environments and
provide a base for further IPL studies in natural environments such as those in which
anaerobic oxidation of methane takes place.
REFERENCES
Aygun-Sunar, S., Mandaci, S., Koch, H. -G., Murray, I. V. J., Goldfine, H., Daldal, F.,
2006. Ornithine lipid required for optimal steady-state amounts of c-typer
cytochromes in Rhodobacter capsulatus. Molecular Microbiology 61, 418-435.
Barridge, J. K., Shively, J. M., 1968. Phospholipids of the Thiobacilli. Journal of
Bacteriology 95, 2182-2185.
Benning, C., Huang, Z. –H., Gage, D. A., 1995. Accumulation of a novel glycolipid and a
betaine lipid in cells of Rhodobacter sphaeroides grown under phosphate limitation.
Archives of Biochemistry and Biophysics 317, 103-111.
Biddle, J. F., Lipp, J. S., Lever, M. A., Lloyd, K. G., Sörensen, K. B., Anderson, R.,
Fredricks, H. F., Elvert, M., Kelly, T J., Schrag, D. P., Sogin, M. L., Brenchley, J.
E., Teske, A. House, C. H., Hinrichs, K. -U., 2006. Heterotrophic archaea dominate
sedimentary subsurface ecosystems off Peru. Proceedings of the National Academy
of Science U.S.A. 103, 3846-3851.
Boetius, A., Ravenschlag, K., Schubert, C. J., Rickert, D., Widdel, F., Gieseke, A.,
Amann, R., Jørgensen, B. B., Witte, U., Pfannkuche, O., 2000. A marine microbial

Chapter V ________________________________________________________________________
144
consortium apparently mediating anaerobic oxidation of methane. Nature 407, 623-
626.
Campbell, K. A., 2006. Hydrocarbon seep and hydrothermal vent paleoenvironments and
paleontology: past developments and future research directions. Palaeogeography,
Palaeoclimatology, Palaeoecology 232, 362-407.
Cooper, D. G., MacDonald, C. R., Duff, S. J. B., Kosaric, N., 1981. Enhanced production
of surfactin from Bacillus subtilis by continuos product removal and metal cation
additions. Applied and Environmental Microbiology 42, 408-412. 1981
Curatolo, E., 1987a. Glycolipid function. Biochimica et Biophysica Acta 906, 137-160.
Curatolo, E., 1987a. The physical properties of glycolipids. Biochimica et Biophysica
Acta 906, 111-136.
De Rosa, M., Gambacorta, A., Gliozzi, A., 1986. Structure, biosynthesis, and
physicochemical Properties of Archaeabacterial Lipids. Microbiological Reviews
50, 70-80.
Dembitsky, V. M., 1996. Betaine ether-linked glycerolipids: chemistry and biology.
Progress in Lipid Research 35: 1-51.
Ertefai, T. F., Fisher, M. C., Fredricks, H. F., Lipp, J. S., Pearson, A., Birgel, D., Udert,
K. M., Cavanaugh, C. M., Gschwend, P. M., Hinrichs, K. -U., 2008. Vertical
distribution of microbial lipids and functional genes in chemically distinct layers of
a highly polluted meromictic lake. Organic Geochemistry 39, 1572-1588.
Fang, J. S., Barcelona, M. J., Semrau, J. D., 2000. Characterization of methanotrophic
bacteria on the basis of intact phospholipid profiles. FEMS Microbial Letters 189,
67-72.
Findlay, R. H., 1996. The use of phospholipids fatty acids to determine microbial
community structure. Molecular Microbial Ecology Manual 4, 1-17.
Findlay, R. H., Trexler, M. B., White, D. C., 1990. Response of a benthic microbial
community to biotic disturbance. Marine Ecology Progress Series 62, 135-148.
Georgio, G., Lin, S. -C., Sharma, M., 1992. Surface-active compound from
microorganisms. Nature biotechnology 10, 60-64.

Chapter V ________________________________________________________________________
145
Gerson, D. F., Zajic, J. E., 1978. Surfactant production from hydrocarbons by
Corynebacterium lepus, sp nov. And Pseudomonas asphaltenicus, sp nov.
Developments in Industruial Microbiology 19, 597-599.
Ghosh, M., Misha, A. K., 1987. Occurrence, identification and possible significance of
ornithine lipid in Thiobacillus ferrooxidans. Biochemical and Biophysical Research
Communications 142, 925-931.
Goldfine, H., 1984. Bacterial membrane and lipid packing theory. Journal of Lipid
Research 25, 1501-1507.
Hasegawa, Y., Kawada, N., Nosoh, Y., 1980. Change in Chemicals composition of
membrane of Bacilus caldotenax after shifting the growth temperature. Archives of
Microbiology 126, 103-108.
Hinrichs, K. -U., Hayes, J. S., Sylva, S. P., Brewer, P. G., DeLong, E. F., 1999. Methane-
consuming archaebacteria in marine sediments. Nature 398, 802-805.
Hofman, M., Eichenberger, W., 1996. Biosynthesis of diacylglyceryl-N,N,N-
trimethylhomoserine in Rhodobacter sphaeroides and evidence for lipid-linked N
methylation. Journal of Bacteriology 178, 6140-6144.
Hue, N., Serni, L., Laprévote, O., 2001. Structural investigation of cyclic peptidolipids
from Bacillus subtilis by high-energy tandem mass spectrometry. Rapid
Communications in Mass Spectrometry 15, 203-209.
Imhoff, J. F., Bias-Imhoff, U., 1995. Lipids, Quinones and Fatty Acids of Anoxygenic
Phototrophic Bacteria. In Anoxygenic Photosynthetic Bacteria (eds. Blankenship, R.
E., Madigan, M. T., Bauer, C. E.), 179-205. 1995 Kluwer Academic Publishers.
Printed in The Netherlands.
Kates, M., 1997. Diether and tetraether phospholipids and glycolipids as molecular
markers for archaeabacteria (archaea). In: Molecular Markers in Environmental
Geochemistry (ed. Eganhause, R. P.), 35–48, Oxford University Press.
Kim, J. -S., Powalla, M., Lang, S., Wagner, F., Lundsdorf, H., Wray, F., 1990. Microbial
glycolipid production under nitrogen limitation and resting cell conditions. Journal
of Biotechnology 13, 257-266.
Knoche, H. W., Shively, J. M., 1972. The structure of an ornithine –containing lipid from
Thiobacillus thioxidans. The Journal of Biological Chemistry 247, 170-178.

Chapter V ________________________________________________________________________
146
Koga, Y., Morii, H., 2005. Recent advances in structural research on ether lipids from
archaea including comparative and physiological aspects. Bioscience Biotechnology
and Biochemistry 69, 2019-2034.
Koga, Y., Morii, H., Akagawa-Matsushita, M., Ohga, I., 1998. Correlation of polar lipid
composition with 16S rRNA phylogeny in methanogens. Further analysis of lipid
component parts. Bioscience Biotechnology and Biochemistry 62, 230-236.
Koga, Y., Nishihara, M., Morii, H., Akagawa-Matsushita, M., 1993. Ether polar lipids of
methanogenic bacteria: structures, comparative aspects, and biosyntheses. 1996.
Microbiological Reviews 57, 164-182.
Langworthy, T. A., 1982. Lipids of bacteria in extreme environments. In: Current Topics
in Membranes and Transport, Membrane lipids of prokaryotes 17 (eds. Razin, S.,
Rottem, S.), 45-77. Academic press.
Levin, L., 2005. Ecology of cold seep sediments: interactions of fauna with fluid flow,
chemistry and microbes. In: Oceanography and Marine Biology: An annual Review
43 (eds. Gibson, R. N., Atkinson, R. J. A., Gordon, J. D. M), 1-46. Taylor and
Francis.
Lipp, J. S., Morono, Y, Inagaki, F., Hinrichs, K.-U., 2008. Significant contribution of
Archaea to the extant biomass in marine subsurface sediments. Nature 454, 991-
994.
López –Lara, I. M., Sohlenkamp, C., Geiger, O., 2003. Membrane lipids in plant-
associated bacteria: their biosyntheses and possible functions. Molecular Plant-
Microbe Interactions Overview 16, 567-579.
Lösekann, T., Knittel, K., Nadalig, T., Fuchs, B., Niemann, H., Boetius, A., Amann, R.,
2007. Diversity and abundance of aerobic and anaerobic methane oxidizers at the
Haakon Mosby Mud Volcano, Barents Sea. Applied and Environmental
Microbiology 73, 3348–3362.
MacCarthy, C. M., Murray, L., 1996. Viability and metabolic features of bacteria
indigenous to a contaminated deep aquifer. Microbial Ecology 32, 305-321.
Madigan, M. T., Martinko, J. M., Parker, J., 2003. Brock - Biology of Microorganisms.
10th Edition, Pearson education Inc. Upper Saddle River, New Jersey.

Chapter V ________________________________________________________________________
147
Makula, R. A., 1978. Phospholipid composition of methane-utilizing bacteria. Journal of
Bacteriology 134, 771-777.
Makula, R. A., Finnerty, W. R., 1975. Isolation and characterization of an ornithine-
containing lipid from Desulfovibrio gigas. Journal of Bacteriology 123, 523-529.
Minnikin, D. E., Abdolrahimzadeh, H., 1974. Effect of pH on the proportions of polar
lipids, in chemostat cultures of Bacillus subtilis. Journal of Bacteriology 120, 999-
1003.
Muthusamy, K., Gopalakrishnan, S., Ravi, T. K., Sivachidambaram, P., 2008.
Biosurfactants: properties, commercial production and application. Current Science
94, 736-746.
Rossel, P. E., Lipp, J. S., Fredricks, H. F., Arnds, J., Boetius, A., Elvert, M., Hinrichs, K.
-U., 2008. Intact polar lipids of anaerobic methanotrophic archaea and associated
bacteria. Organic Geochemistry 39, 992-999.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2001. Monoalkylether phospholipids in
the sulfate-reducing bacteria Desulfosarcina variabilis and Desulforhabdus
amnigenus. Archives of Microbiology 176, 435-442.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2002. Phospholipid analysis as a tool
to study complex microbial communities in marine sediments. Journal of
Microbiological Methods 48, 149-160.
Sahling, H., Rickert, D., Lee, R. W., Linke, P., Suess, E., 2002. Macrofaunal community
structure and sulfide flux at gas hydrate deposits from the Cascadia convergent
margin, NE Pacific. Marine Ecological Progress Series 231, 121-138.
Sato, N., 1992. Betaine lipids. The botanical Magazine Tokyo 105, 185-197.
Sprott, D. G., 1992. Structures of archaebacterial membrane lipids. Journal of
Bioenergetics and Biomembranes 24, 555-565.
Sprott, D. G., Meloche, M., Richards, J. C., 1991. Proportions of diether, macrocyclic
diether and tetraether lipids in Methanococcus Jannaschii grown at different
temperatures. Journal of Bacteriology 173, 3907-3910.
Stadnitskaia, A., Bouloubassi, I., Elvert, M., Hinrichs, K. -U., Sinninghe Damsté, J. S.,
2008. Extended hydroxyarchaeaol, a novel lipid biomarker for anaerobic
methanotrophy in cold seepage habitats. Organic Geochemistry 39, 1007-1014.

Chapter V ________________________________________________________________________
148
Sturt, H. F., Summons, R. E., Smith, K. J., Elvert, M., Hinrichs, K. -U., 2004. Intact polar
membrane lipids in prokaryotes and sediments deciphered by high-performance
liquid chromatography/electrospray ionization multistage mass spectrometry-new
biomarkers for biogeochemistry and microbial ecology. Rapid Communications in
Mass Spectrometry 18, 617-628.
Thimon, L., Peypoux, F., Maget-Dana, R., Roux, B., Michel, G., 1992. Interactions of
surfactin, a biosurfactant from Bacillus subtilis, with inorganic cations.
Biotechnology Letters 14, 713-718.
Vater, J., 1986. Lipopeptides, an attractive class of microbial surfactants. Progress in
Colloid and Polymer Science 72, 12-18.
Wagner, M., Horn, M., Daims, H., 2003. Fluorescence in situ hybridization for the
identification and characterization of prokaryotes. Current opinion in Microbiology
6, 302-309.

Chapter VI ________________________________________________________________________
149
CHAPTER VI
Concluding remarks and perspectives

Chapter VI ________________________________________________________________________
150
VI.1. Conclusions
This dissertation focused on the study of different microbial communities
involved in the process of AOM. This work began with the identification of several intact
polar lipids (IPLs) from few samples phylogenetically dominated by each of three
anaerobic methanotrophic consortia (ANME-1, -2 and 3 and sulfate reducer bacterial
partners). After the identification of several diagnostic IPLs characteristic of the each
AOM-community, these lipids were analyzed in a variety of globally distributed cold
seep systems. Among these hot spots of AOM, different habitats were analyzed such as
anoxic water bodies, mud volcanoes, oil fields, gas hydrate environments and
hydrothermal vents. In the course of this work, it was possible to address several open
questions regarding AOM-research: (1) identification of communities involved in AOM
based on few diagnostic of IPLs, (2) microbial-derived IPL diversity in AOM hot spots
and (3) environmental factors influencing the dominance and distribution of AOM-
communities.
This work is the first to demonstrate that IPLs, which are biomarkers associated to
living biomass, enable not only the distinction of the three main groups of AOM-
mediating microbes from a wide variety of methane-bearing habitats (Chapter II and III)
but, more importantly, provides additional insights on the environmental factors
influencing the distribution of these communities (Chapter III).
The three phylogenetically distinct clusters of Euryarchaeota called ANME-1, -2
and -3 (e.g., Hinrichs et al., 1999; Boetius et al., 2000; Lösekann et al., 2007) which have
been observed in association with sulfate-reducing bacteria (SRB) of the
Desulfosarcina/Desulfococcus group (Boetius et al., 2000; Orphan et al., 2001; Michaelis
et al., 2002, ‘‘ANME-1/DSS and -2/DSS aggregates”) or Desulfobulbus spp (Lösekann et
al., 2007, ‘‘ANME-3/DBB aggregates”) exhibit a characteristic IPL composition.
ANME-1, which is not directly affiliated with any of the major orders of methanogens
(Hinrichs et al., 1999; Orphan et al., 2001; Knittel et al., 2005) is characterized by the
production of glyceroldialkylglyceroltetraether (GDGTs) with glycosidic and phospho as
well as mixed glycosidic and phospho headgroups. The main glycosidic-GDGT in

Chapter VI ________________________________________________________________________
151
ANME-1 system, is diglycosyl-GDGT (2Gly-GDGT, Rossel et al., 2008; Chapter II and
Chapter III), a lipid also frequently observed in deep subsurface (Biddle et al., 2006; Lipp
et al., 2008), as well among several species within the order Methanomicrobiales (Koga
et al., 1998). In addition to glycosidic-GDGTs, GDGTs with mixed glycosidic and
phospho or only phospho headgroups were dominated by 2Gly-GDGT-PG and 2PG-
GDGT (Chapter III), which have been also previously reported in Methanobacterium
thermoautotrophicum (Koga et al., 1993). Interestingly, contribution of 2Gly-GDGT,
2Gly-GDGT-PG and 2PG-GDGT varied depending of the ANME-1 habitat. Beside the
general dominance of 2Gly-GDGT, the contribution of 2Gly-GDGT-PG and 2PG-GDGT
was much higher in sediment than in carbonate reefs dominated by ANME-1.
Different from ANME-1, diagnostic IPLs of ANME-2 were archaeols with both
glycosidic and phospho headgroups, which also occur in Methanocaldococcus jannaschii,
Methanococcus voltae and Methanothirx soehngenii (Koga et al., 1993; Sturt et al.,
2004). Within the glycosidic archaeols the main IPLs were 2Gly-archaeol (2Gly-AR),
2Gly-MAR (2Gly-macrocyclic archaeol), 2Gly-hydroxyarchaeol (2Gly-OH-AR),
whereas the major phospho-archaeols were PG-OH-AR, phosphatidylserine-OH-AR (PS-
OH-AR) and phosphatidylinositol-OH-AR (PI-OH-AR) (Chapter III). Similar to ANME-
1 systems, archaeal IPLs containing phospho headgroups were more abundant in
sediments than in carbonate reefs.
ANME-3, contrary to ANME-2 and ANME-1 contained neither glycosidic-
archaeols nor GDGT-based IPLs. However, the phospho-archaeols composition was very
similar to ANME-2, although with a generally less contribution of PI-OH-AR (Chapter
III). The phylogenetic affiliation of ANME-2 and ANME-3 with the order
Methanosarcinales, was consistent with the dominance of archaeol and hydroxyarchaeol
with both glycosidic and phospho headgroups (Kates, 1997; Koga et al., 1998).
Among the major bacterial IPLs, relative high abundance of
phosphatidylethanolamine (PE), phosphatidyl-(N)-methylethanolamine (PME) and
phosphatidyl-(N,N)-dimethylethanolamine (PDME) with diacylglycerol (DAG) bond
type, were found in ANME-2/DSS and ANME-3/DBB dominated settings (Rossel et al.,
2008; chapter II and chapter III). PE is the major phospholipid type of SRB such as
Desulfosarcina variabilis (Rütters et al., 2001) and its occurrence together with PME and

Chapter VI ________________________________________________________________________
152
PDME in anoxic waters and surface sediments from the Black Sea has been also
suggested to derive from SRB (Schubotz et al., submitted). However PME and PDME
have been also described in some methanotrophic bacteria (Makula, 1978; Fang et al.,
2000) as well as sulfide oxidizers (Barridge and Shively, 1968). The presence of PME
and PDME seems to be a general feature of ANME-3/DBB dominated systems, although
it needs to be taken into account, that a fraction of these two IPLs may derived either
from aerobic methanotrophic bacteria or from sulfide oxidizers, both which contain
similar membrane lipids (Barridge and Shively, 1968; Makula, 1978; Fang et al., 2000).
Other bacterial IPLs, which contributed mainly to ANME-2/DSS dominated mats,
were the non-phospho IPLs ornithine lipids (OL), surfactin and betaine lipids (BL), with
the latter characterized by odd fatty acid chains (BL-odd) (Chapter III). OL have been
reported in SRB, and sulfur and iron oxidizing bacteria (Knoche and Shively, 1972;
Makula and Finerty, 1975), whereas surfactin is a lipopeptide with surface active
properties common of Bacillus sp. (Vater et al., 1986) that may also be produced by an
unknown bacteria in the mats from the Black Sea. On the other hand, BL-odd, contrary to
BL with even fatty acid chains, have been suggested to derive from bacteria, do to their
occurrence in deep anoxic water of the Black Sea (Schubotz et al., submitted).
Based on IPL distribution, it was possible to observe a clear separation within the
chimney-like structures and the sediment habitats. ANME-1 and ANME-2/DSS
inhabiting carbonate reefs contained high abundance of glycosidic-IPLs and IPL with
non-phospho headgroups. Both archaeal (2Gly-GDGT, 2Gly-AR, 2Gly-MAR, 2Gly-OH-
AR) and bacterial IPL (OL, surfactin and BL odd) composition point to the low
abundance of phospho-IPLs in carbonate mats compared to sediments. Dissolved
phosphate in sediment pore water has been shown to be strongly adsorbed on calcium
carbonate (Cole et al., 1953; de Kanel and Morse, 1978). Therefore, limitation of
dissolved phosphate in AOM carbonate mats from the Black Sea is likely responsible for
the generally low abundance of IPLs with phospho headgroups in both ANME-1/DSS
and ANME-2a/DSS dominated mats (Chapter III).
Beside the general differences in IPL composition of ANME-1, -2 and -3
communities, additional variations in the IPL pattern in relation to several environmental

Chapter VI ________________________________________________________________________
153
variables provided new insights into the ecological niches dominated by these
communities (Chapter III). ANME-1/DSS, in which the diagnostic IPL was 2Gly-GDGT,
dominates habitats with higher temperature and lower oxygen content in bottom waters
compared to the systems in which ANME-2/DSS and ANME-3/DBB inhabit. This
relationship between ANME-1/DSS and temperature is in agreement with the detected
higher AOM-activity of ANME-1/DSS at higher temperatures (up to 24°C) compared to
ANME-2/DSS (up to 15°C) based on in vitro experiments (Nauhaus et al., 2005).
Furthermore, the dominance of ANME-1/DSS in low oxygen bottom waters is in
agreement with previous field observations, which suggest that ANME-1/DSS may be
more sensitive to oxygen than ANME-2/DSS (Knittel et al., 2005). Based on IPL
diversity, ANME-2/DSS systems were separated in two groups: the carbonate reefs and
the sediments. ANME-2/DSS dominated sediments were characterized not only by lower
temperature and higher oxygen content in bottom waters, but also by higher methane and
sulfate concentrations. These environmental variables were accompanied by the presence
of PG-OH-AR and PI-OH-AR. On the other hand, ANME-2/DSS dominated carbonate
mats were associated with higher sulfate reduction rates (SRR) and to the occurrence of
2Gly-OH-AR and 2Gly-MAR. These differences between carbonate reefs and sediments
dominated by ANME-2/DSS could be explained by the presence of sulfide oxidizing
bacteria (SOB) in the sediments, which efficiently remove sulfide and produce sulfate.
The environmental characteristics, as well as the archaeal IPL composition of
ANME-3 and ANME-2 from sediments, suggest that these two communities dominate in
similar environments, although due to the fact that the lowest temperatures were observed
at ANME-3/DBB dominated sediments from Håkon Mosby Mud Volcano, it is possible
that temperature may also select for either ANME-2/DSS or ANME-3/DBB.
IPL data in general was in good agreement with the phylogenetic information
based on FISH methods. Nevertheless, in a few cases both methods have some
discrepancies due to several potential reasons. It was observed that in sediments
dominated by ANME-2c/DSS according to FISH counting, the contribution of ANME-1
derived GDGT-based IPLs was higher than the ANME-2/DSS IPL signal. The high
contribution of GDGT-based IPLs was probably due to the presence of extremely large
ANME-1 cells in this setting. Additionally, FISH methods could also underestimate

Chapter VI ________________________________________________________________________
154
archaeal abundance, especially ANME-1, due to the low permeability of their membranes
compared to the bacterial phospholipid (Wagner et al., 2003).
The evaluation of apolar lipids distribution provided a poor taxonomic separation
between the three AOM-communities (Chapter III). This was probably due to the lack of
GDGTs in our data set, which is the main core lipid of ANME-1, but also to the
presumed longer turnover times of apolar lipids than of IPLs. Apolar signals may
integrate longer periods in the geologic evolution of the studied seep systems, in which
community changes are likely to occur resulting in a mixed signal from current and past
microbial communities.
Furthermore, IPL behavior on marine sediment systems was evaluated using an
experimental approach (Chapter IV). Incubations were performed using slurries of
sediments with (sterile condition) and without sterilization (active condition), in which
membrane lipid of archaea (2Gly-GDGT) and bacteria (C16-PC) were spiked. Both sterile
and active conditions were incubated at 5°C and 40°C. According to our results both
archaeal and bacterial IPLs were degraded under sterile conditions. However, after 465
days of incubation under active conditions, an increase of both IPLs was observed
(although the bacterial IPL only increased at 5°C). This suggests that the microbial
communities were growing. Unfortunately, degradation of IPLs in the active conditions
could not be proved because the IPLs produced and degraded were indistinguishable.
Therefore, an improved experimental approach is necessary.
We demonstrated that few IPLs enable the distinction of AOM-communities,
although the diversity of IPLs identified in methane-bearing habitats is very high (chapter
III and V). Among the archaeal IPLs identified, GDGT-based and archaeol-based IPLs
with glycosidic, mixed phospho and glycosidic or pure phospho headgroups were
observed. Bacterial IPLs were also diverse having not only different phospho headgroups
but also containing non-phospho IPLs. Structural information and fragmentation patterns
of diverse IPL classes are provided in this thesis (Chapter V) as base for further IPL
identification in AOM systems.
The results obtained during this thesis provide a clear distinction between the
major microbial communities involved in AOM in marine sediments (ANME-1, -2 and -3

Chapter VI ________________________________________________________________________
155
and SRB partners) based on IPL distribution. Additionally these results demonstrate that
IPLs varied not only according to the community type but also in relation to the habitat
characteristics. Furthermore, IPL distribution was also related to several environmental
factors selecting for one of the three major AOM-community types. Thus, allowing to
define the ecological niches dominated by each of these groups.
VI.2. Future perspectives
This thesis contributes to a better understanding of the microbial communities
involved in the process of AOM and the environmental factors controlling their
dominance in a variety of seeps globally distributed. However, several open questions
regarding the process of AOM and the potential applications of IPLs for future research
can still be addressed:
� Few diagnostic IPLs enable the distinction between the three major communities
performing AOM. However the diversity of IPLs in hot spots of AOM is quite
high and includes some IPLs which are abundant in just a few settings. This
suggest that either the same microbial communities produce different IPLs
depending of the environment or other ANMEs, so far not identified, are present.
In this settings will be necessary to characterized in detail the microbial
community present.
� The high concentration of IPLs in AOM systems provides an excellent
opportunity to elucidate structural diversity of IPLs derived from both archaea and
bacteria living in marine systems. Some of these IPLs, still with tentative
structures, can be purified to confirm their structures.
� During this work, we have learned that IPLs are strongly influenced by the habitat
conditions in which microbial communities dominate. In this study the role of
phosphate limitation was discussed, although many other factors may influence
the composition of IPLs in microbial membranes. These effects can be studied
either by covering a variety of extreme environments or by culture experiments in

Chapter VI ________________________________________________________________________
156
which environmental factors such as nutrients, pH, pressure, temperature, carbon
source, starvation are controlled and properly monitored.
� Environmental factors selecting between ANME-1/DSS and ANME-2/DSS were
clearly defined. However ANME-2/DSS and ANME-3/DBB presented similar
IPL compositions as well as the habitat characteristics, which do not allow a good
separation between these two groups. It is necessary to study in more detail
ANME-3/DBB from other dominated settings, to confirm the presence of similar
diagnostic IPLs as well as the environmental factors influencing their distribution.
� An improvement of the experimental design used in this study to evaluate stability
of IPLs in sediments is needed. This information affects the interpretation and
validation of IPLs as biomarkers for currently active communities. In this new
experimental approach, the distinction between degraded and produced IPLs, the
effects of adsorption in IPLs behavior over time, the abundance of microbial cells
as well as degradation products should be considered.
REFERENCES
Barridge, J. K., Shively, J. M., 1968. Phospholipids of the Thiobacilli. Journal of
Bacteriology 95, 2182-2185.
Biddle, J. F., Lipp, J. S., Lever, M. A., Lloyd, K. G., Sörensen, K. B., Anderson, R.,
Fredricks, H. F., Elvert, M., Kelly, T J., Schrag, D. P., Sogin, M. L., Brenchley, J.
E., Teske, A. House, C. H., Hinrichs, K. –U., 2006. Heterotrophic archaea dominate
sedimentary subsurface ecosystems off Peru. Proceedings of the National Academy
of Science U.S.A. 103, 3846-3851.
Boetius, A., Ravenschlag, K., Schubert, C. J., Rickert, D., Widdel, F., Gieseke, A.,
Amann, R., Jørgensen, B. B., Witte, U., Pfannkuche, O., 2000. A marine microbial
consortium apparently mediating anaerobic oxidation of methane. Nature 407, 623-
626.
Cole, C.V., Olsen, S. R., Scott, C. O., 1953. The nature of phosphate sorption by calcium
carbonate. Soil Science Society of America Journal 17, 352-356.

Chapter VI ________________________________________________________________________
157
De Kanel, J., Morse, J. W., 1978. The chemistry of orthophosphate uptake from seawater
on to calcite and aragonite. Geochimica et Cosmochimica Acta 42, 1335-1340.
Elvert, M., Niemann, H., 2008. Occurrence of unusual steroids and hopanoids derived
from aerobic methanotrophs at an active marine mud volcano. Organic
Geochemistry 39, 167–177.
Fang, J. S., Barcelona, M. J., Semrau, J. D., 2000. Characterization of methanotrophic
bacteria on the basis of intact phospholipid profiles. FEMS Microbial Letters 189,
67-72.
Hinrichs, K.-U., Hayes, J. S., Sylva, S. P., Brewer, P. G., DeLong, E. F., 1999. Methane-
consuming archaebacteria in marine sediments. Nature 398, 802-805.
Kates, M., 1997. Diether and tetraether phospholipids and glycolipids as molecular
markers for archaeabacteria (archaea). In: Molecular Markers in Environmental
Geochemistry (ed. Eganhause, R.P.), pp. 35–48. Oxford University Press.
Knittel, K., Lösekann, T., Boetius, A., Kort, R., Amann, R., 2005. Diversity and
Distribution of Methanotrophic Archaea at Cold Seeps. Applied and Environmental
Microbiology 71, 467-479.
Knoche, H. W., Shively, J. M., 1972. The structure of an ornithine –containing lipid from
Thiobacillus thioxidans. The Journal of Biological Chemistry 247, 170-178.
Koga, Y., Nishihara, M., Morii, H., Akagawa-Matsushita, M., 1993. Ether polar lipids of
methanogenic bacteria: structures, comparative aspects, and biosyntheses. 1996.
Microbiological Reviews 57, 164-182.
Koga, Y., Morii, H., Akagawa-Matsushita, M., Ohga, I., 1998. Correlation of polar lipid
composition with 16S rRNA phylogeny in methanogens. Further analysis of lipid
component parts. Bioscience Biotechnology and Biochemistry 62, 230-236.
Lipp, J. S., Morono, Y, Inagaki, F., Hinrichs, K.-U., 2008. Significant contribution of
Archaea to the extant biomass in marine subsurface sediments. Nature 454, 991-
994.
Lösekann, T., Knittel, K., Nadalig, T., Fuchs, B., Niemann, H., Boetius, A., Amann, R.,
2007. Diversity and abundance of aerobic and anaerobic methane oxidizers at the
Haakon Mosby Mud Volcano, Barents Sea. Applied and Environmental
Microbiology 73, 3348–3362.

Chapter VI ________________________________________________________________________
158
Makula, R. A., Finnerty, W. R., 1975. Isolation and characterization of an ornithine-
containing lipid from Desulfovibrio gigas. Journal of Bacteriology 123, 523-529.
Makula, R. A., 1978. Phospholipid composition of methane-utilizing bacteria. Journal of
Bacteriology 134, 771-777.
Nauhaus, K., Treude, T., Boetius, A., Krüger, M., 2005. Environmental regulation of the
anaerobic oxidation of methane a comparison of ANME-1 and ANME-II
communities. Environmental Microbiology 7, 98-106.
Niemann, H., Lösekann T., de Beer, D., Elvert, M., Nadalig, T., Knittel, K., Amann, R.,
Sauter, E., Schlüter, M., Klages, M., Foucher, J. -P., Boetius, A., 2006. Novel
microbial communities of the Haakon Mosby mud volcano and their role as a
methane sink. Nature 443, 854-858.
Orphan, V. J., House, C. H., Hinrichs, K. –U., McKeegan, K. D., DeLong, E. F., 2001.
Comparative analysis of methane-oxidizing archaea and sulfate-reducing bacteria in
anoxic marine sediments. Applied and Environmental Microbiology 67, 1922-1934.
Rossel, P. E., Lipp, J. S., Fredricks, H. F., Arnds, J., Boetius, A., Elvert, M., Hinrichs, K.
–U., 2008. Intact polar lipids of anaerobic methanotrophic archaea and associated
bacteria. Organic Geochemistry 39, 992-999.
Rütters, H., Sass, H., Cypionka, H., Rullkötter, J., 2001. Monoalkylether phospholipids in
the sulfate-reducing bacteria Desulfosarcina variabilis and Desulforhabdus
amnigenus. Archives of Microbiology 176, 435-442.
Sturt, H. F., Summons, R. E., Smith, K. J., Elvert, M., Hinrichs, K.-U., 2004. Intact polar
membrane lipids in prokaryotes and sediments deciphered by high-performance
liquid chromatography/electrospray ionization multistage mass spectrometry-new
biomarkers for biogeochemistry and microbial ecology. Rapid Communications in
Mass Spectrometry 18, 617-628.
Wagner, M., Horn, M., Daims, H., 2003. Fluorescence in situ hybridization for the
identification and characterization of prokaryotes. Current opinion in Microbiology
6, 302-309.

Chapter VI ________________________________________________________________________
159
VI.3. Presentations and other activities
August 2008 Gordon Research Conference in Organic Geochemistry, Plymouth, USA. “Intact polar membrane lipids associated with microbial communities performing AOM from globally distributed hydrocarbon seeps. (Poster)
February 2008 Anaerobic Oxidation of Methane Exchange Meeting
together with groups from Universities of Wageningen and Nijmegen (The Netherlands), Aselage, Germany. “Intact polar membrane lipid analyses of anaerobic methanotrophic archaea and associated bacteria”. (Talk)
October 2007 International Conference and 97th Annual Meeting of the
Geologische Vereinigung e.V. University of Bremen, Bremen, Germany. “Diversity of polar lipids in anaerobic communities and their stability in marine sediments”. (Talk)
September 2007 International Meeting on Organic Geochemistry
Conference, Torquay, UK. “Polar and apolar lipids of anaerobic methanotrophic communities from marine seep environments and their relation to environmental conditions” (Poster)
August 2006 Gordon Research Conference in Organic Geochemistry,
Plymouth, USA. “Diversity of polar lipids in anaerobic methanotrophic communities”. (Poster)
November 2005 2nd Northern German Organic Geochemistry Meeting.
University of Oldenburg, Oldenburg, Germany. “Diversity of polar lipids in anaerobic methanotrophic communities”. (Talk)
Participation in Fieldtrips November 2007 M74/3 on board of R/V Meteor (Fujairah to Maldives,
Indian Ocean) in the framework of the “Methane seeps and sediment transport on the Makran accretionary prism and biogeochemical investigations of the oxygen minimum zone.
Academic Supervision July-August 2006 Supervision of summer student Augusta Dibbell,
Massachusetts Institute of Technology-USA.

Chapter VI ________________________________________________________________________
160
Courses September 2007 Organic Facies modelling. European Graduate College in
Marine Sciences (ECOLMAS) January 2007 Methane Biogeochemistry and Geophysics & Remote
Sensing and Ocean-Land Interaction. Austral Summer Institute (ASI-VII)
May-June 2006 Advanced Organic Biogeochemistry. European Graduate
College in Marine Sciences (ECOLMAS) March 2006 Signal and time series analysis. European Graduate College
in Marine Sciences (ECOLMAS)