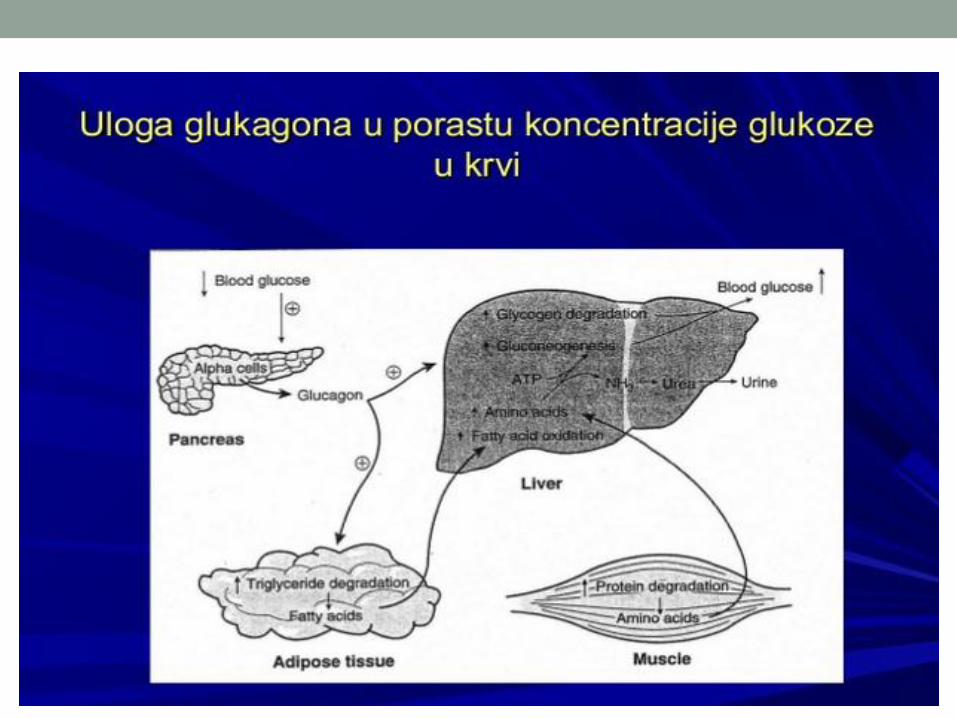
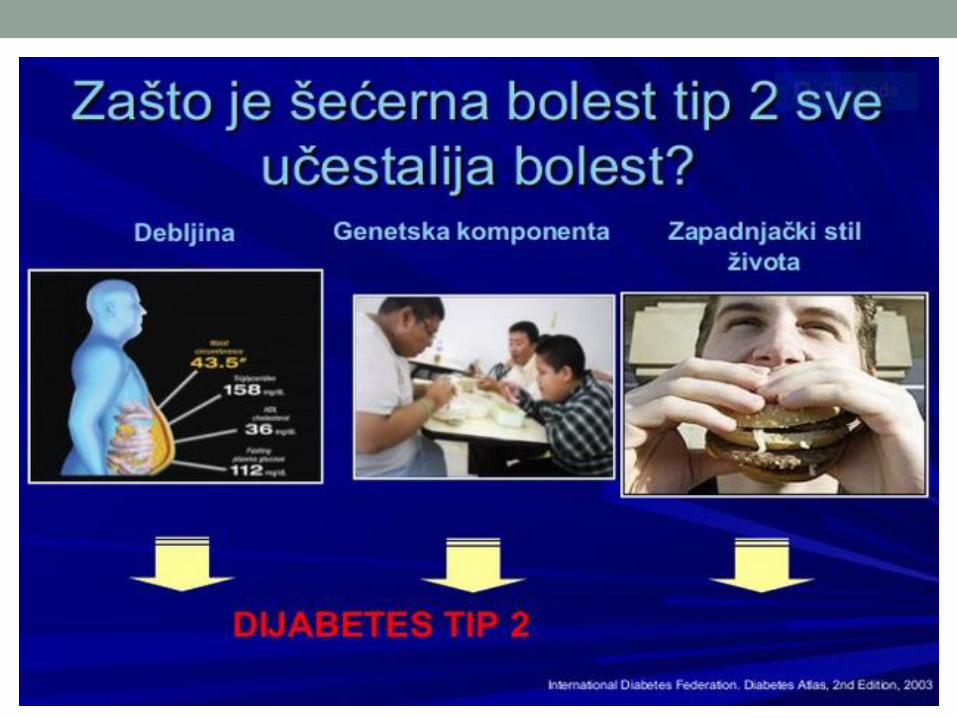
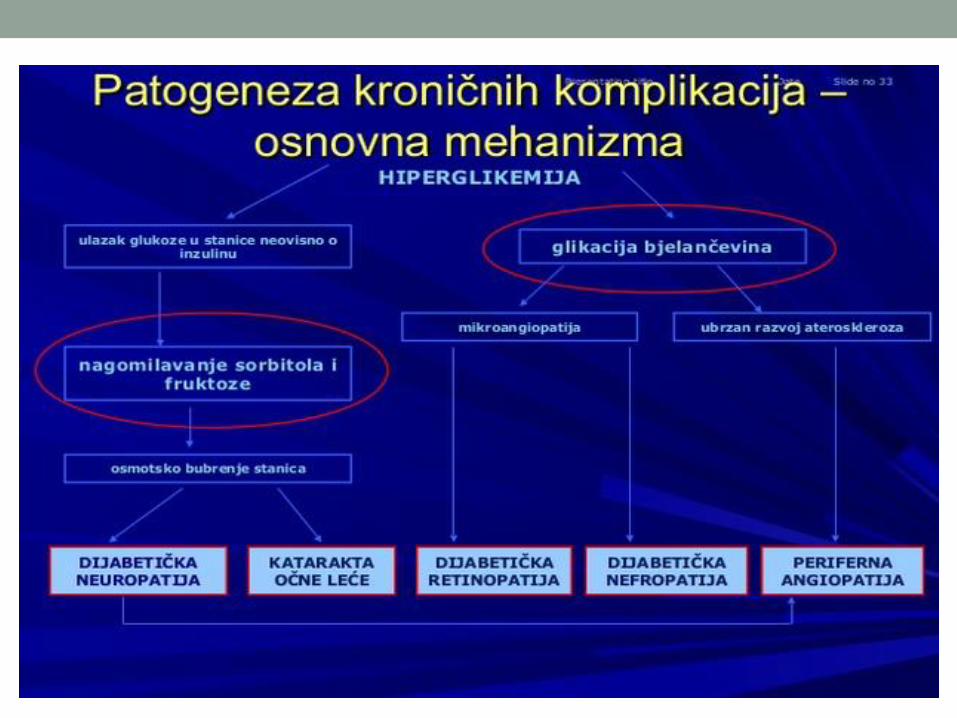
metabolizam ugljenih hidrata - ucg.ac.me uh... · metabolizam ugljenih hidrata 14.02.2018. zbirni...
TRANSCRIPT

METABOLIZAM UGLJENIH HIDRATA14.02.2018.








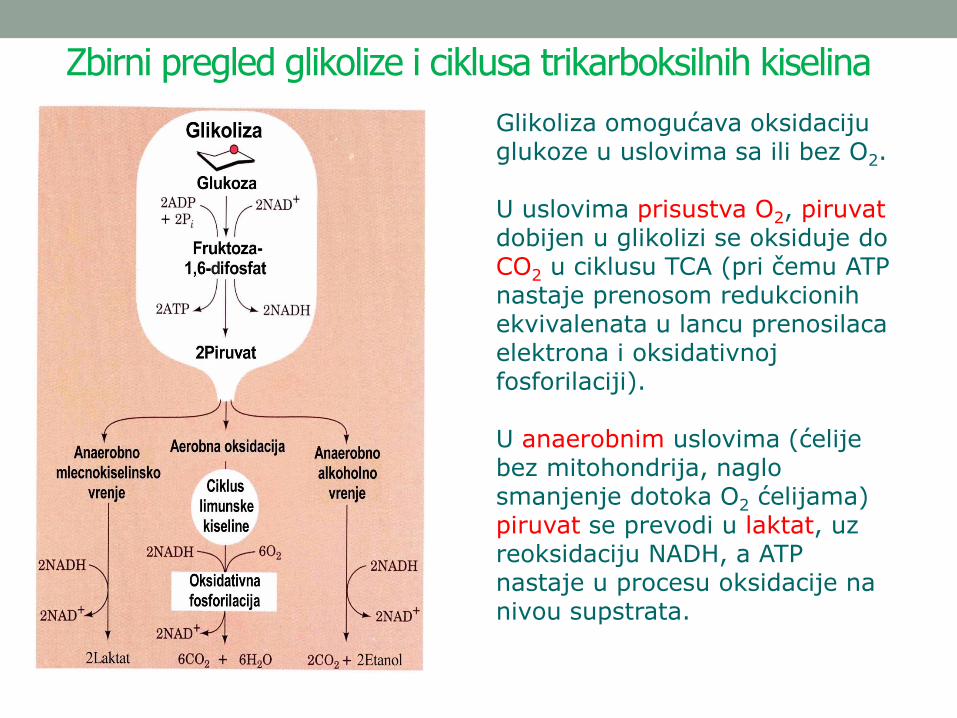
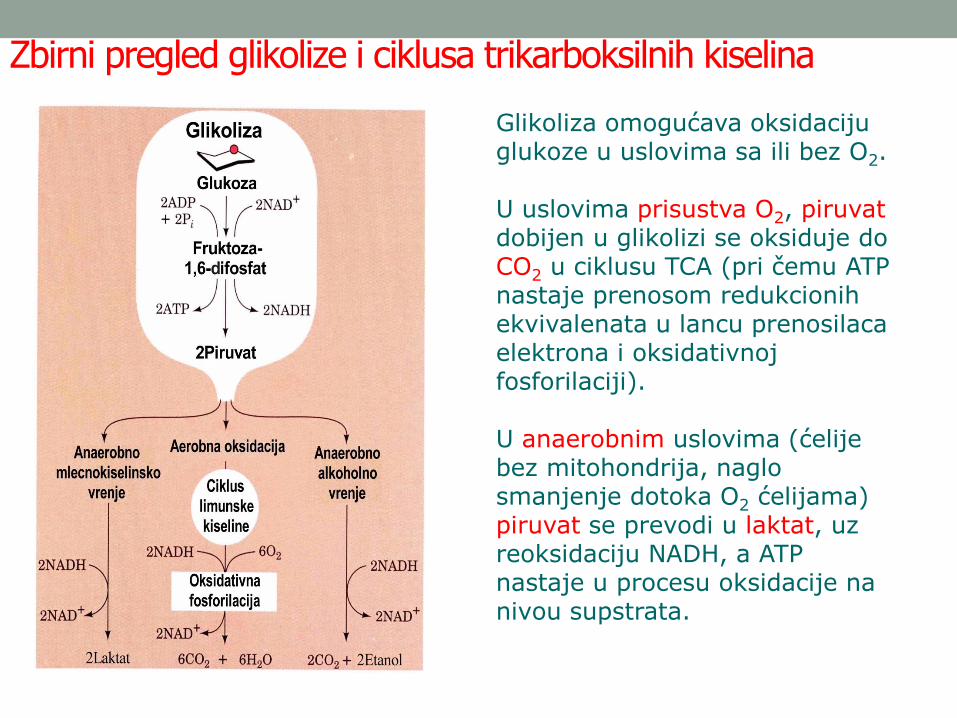
Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina
Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O2.
U uslovima prisustva O2, piruvatdobijen u glikolizi se oksiduje do CO2 u ciklusu TCA (pri čemu ATP nastaje prenosom redukcionih ekvivalenata u lancu prenosilaca elektrona i oksidativnoj fosforilaciji).
U anaerobnim uslovima (ćelije bez mitohondrija, naglo smanjenje dotoka O2 ćelijama) piruvat se prevodi u laktat, uz reoksidaciju NADH, a ATP nastaje u procesu oksidacije na nivou supstrata.


Transport glukoze u ćelije

GLIKOLIZA
Zbirni pregled glikolize i ciklusa trikarboksilnih kiselina
Glikoliza omogućava oksidaciju glukoze u uslovima sa ili bez O2.
U uslovima prisustva O2, piruvatdobijen u glikolizi se oksiduje do CO2 u ciklusu TCA (pri čemu ATP nastaje prenosom redukcionih ekvivalenata u lancu prenosilaca elektrona i oksidativnoj fosforilaciji).
U anaerobnim uslovima (ćelije bez mitohondrija, naglo smanjenje dotoka O2 ćelijama) piruvat se prevodi u laktat, uz reoksidaciju NADH, a ATP nastaje u procesu oksidacije na nivou supstrata.

•Glikolizu sačinjavaju 2 faze: pripremna ifaza u kojoj se dobija energija (SVAKAFAZA TRAJE PO 5 CIKLUSA).
•Prvih pet reakcija čine pripremnu fazu,u kojoj nastaju dva mola gliceraldehid-3-fosfata iz jednog mola glukoze, uzutrošak 2 mola ATP-a.

Pripremna faza

(1) Fosforilacija D-glukoze: heksokinaza
Utrošak prvog mola ATP
Ireverzibilna reakcija
Enzimi: heksokinaza, glukokinaza
O
CH2OH
HH
OH
H
OH
OH
H
OH
H
+ ATP
Heksokinaza
Mg2+
O
CH2OPO
3
HH
OH
H
OH
OH
H
OH
H
+ ADP H+
Glukoza
Glukoza-6-fosfat (G6P)
+
2-
O
CH2OH
HH
OH
H
OH
OH
H
OH
H
+ ATP
Heksokinaza
Mg2+
O
CH2OPO
3
HH
OH
H
OH
OH
H
OH
H
+ ADP H+
Glukoza
Glukoza-6-fosfat (G6P)
+
2-
HK
Mg2+

Sudbine glukozo-6-fosfata
Fosforilacijom se glukoza ”zarobljava” u ćeliji i reakcija je pod unutarćelijskim uslovima bespovratna.
I druge heksoze (D-galaktoza, D-fruktoza i D-manoza), nakon fosforilacije, mogu da uđu u pripremnu fazu glikolize.
Od glukozo-6-P se granaju metabolički putevi, jer jeovaj molekul prekursor praktično svakog puta
glukoze :glikoliza, pentozni put, sinteza glikogena, sinteza glukuronske kiseline i aminošećera. Takođe ovaj molekuli i nastaje u nekim metaboličkim putevima (razgradnja glikogena, glukoneogeneza).
Ćelijska membrana nepropusna za fosforne estrešećera, samo jetra i bubreg imaju glc-6-fosfatazu
koja otpušta slobodnu glc u krv.

Heksokinaza D (ili tip IV, glukokinaza), najzastupljeniji izoenzim u hepatocitima, ima znatno manji afinitet za glukozu od drugih heksokinaza i katališe fosforilaciju heksoza u heksozo-6-fosfat.
Ovakve karakteristike različitih izoenzima heksokinaze imaju za posledicu da:
(i) porast glikemije dovodi do povećanja brzine fosforilacije glukoze u jetri
(ii) u tkivima u kojima su prisutni izoenzimi heksokinaze sa velikim afinitetom za supstrat, fosforilacija glukoze postoji čak i u uslovima relativno niske koncentracije glukoze u krvi, što je od velikog značaja za ćelije koje u potpunosti zavise od glukoze kao izvora energije (mozak).
Postoji nekoliko izoenzima heksokinaze, koji katališu fosforilaciju glukoze. Većina ih ima visok afinitet za glukozu (Km<0,1 mmol/L), i inhibisani su proizvodom reakcije, glukozo-6-fosfatom

(3) Fosforilacija F6P: fosfofruktokinaza
Fosfofruktokinaza – glavni regulatorni enzim glikolize
Fosfofruktokinaza (PFK)
O
CH2OPO
3
H
OH
H
H
OH
OH
CH2OH
2-
Fruktoza-6-fosfat (F6P)
Mg2+
+ ATP
O
CH2OPO
3
H
OH
H
H
OH
OH
CH2OPO
3
2-
2-
Fruktoza-1,6-difosfat (FDP)
+ ADP H+
+
Fosfofruktokinaza (PFK)
O
CH2OPO
3
H
OH
H
H
OH
OH
CH2OH
2-
Fruktoza-6-fosfat (F6P)
Mg2+
+ ATP
O
CH2OPO
3
H
OH
H
H
OH
OH
CH2OPO
3
2-
2-
Fruktoza-1,6-difosfat (FDP)
+ ADP H+
+
PFK1
Mg2+

(4) Cepanje FDP: aldolaza
CH2OPO
3
C
C
C
C
CH2OPO
3
O
HOH
OH
H
H
OH
CH2OPO
3
C
CH2
O
OH
C
OH
C OHH
CH2OPO
3
Dihidroksiaceton-fosfat (DHAP)
Gliceraldehid-3-fosfat (GAP)
Fruktoza-1,6-difosfat (FDP)
Aldolaza
2-
2-
2-
2-
1
2
3

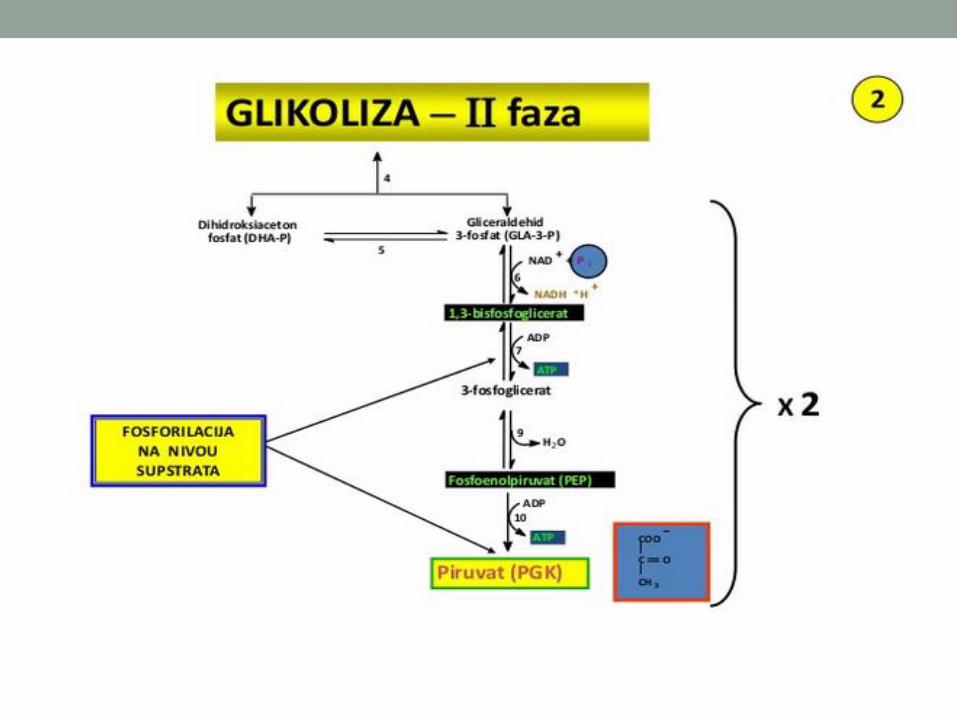
Stvaranje prvog visokoenergetskog jedinjenja, 1,3-bisfosfoglicerat, (mešoviti anhidrid karboksilne i fosfornekiseline) oksidacijom gliceraldehid-3-fosfata.
C
HO
C OHH
CH2OPO
3
Gliceraldehid- 3-fosfat (GAP)
2-
GAPDH+ NAD + P
+i
C
OPO3O
C OHH
CH2OPO
3
1,3-Difosfoglicerat (1,3-DPG)
2-
2-
+ NADH + H+
1
2
3
1
2
3
(6) Oksidacija gliceraldehid-3-fosfata:gliceraldehid-fosfat dehidrogenaza ENERGETSKO
JEDINJENJE

Faza dobijanja energije - oksidacija i fosforilacija na nivou supstrata
Regeneracija NAD+:
- u laktat dehidrogenaznoj reakciji,
- prenošenjem redukcionih ekvivalenata u mitohondrije mehanizmima povratnog transporta.


Sudbine piruvata
Piruvat se, u aerobnim uslovima, oksidativnom dekarboksilacijomprevodi u acetil-CoA, koji ulazi u ciklustrikarboksilnih kiselina (ciklus TCA). Potpuna oksidacija glukoze do CO2 i H2O daje 29,5-31 molekul ATP-a pomolekulu glukoze.
U anaerobnom uslovima, piruvat se redukuje u laktat,čime se obezbeđuje oksidacija NADH+H+ i dalje odvijanje glikolize

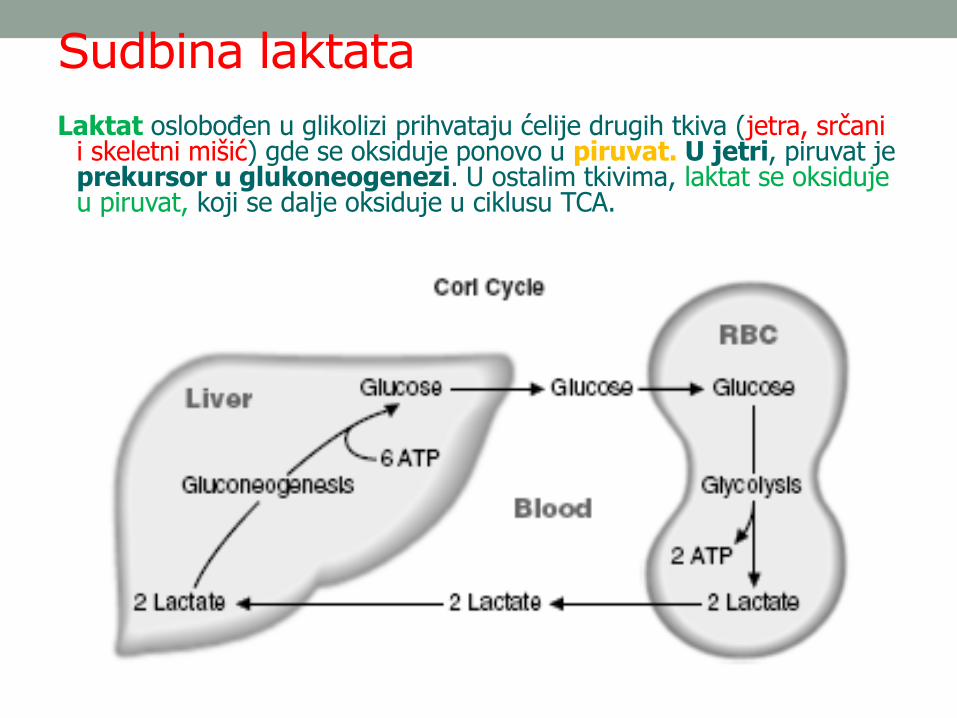
Sudbina laktata
Laktat oslobođen u glikolizi prihvataju ćelije drugih tkiva (jetra, srčani i skeletni mišić) gde se oksiduje ponovo u piruvat. U jetri, piruvat je prekursor u glukoneogenezi. U ostalim tkivima, laktat se oksiduje u piruvat, koji se dalje oksiduje u ciklusu TCA.

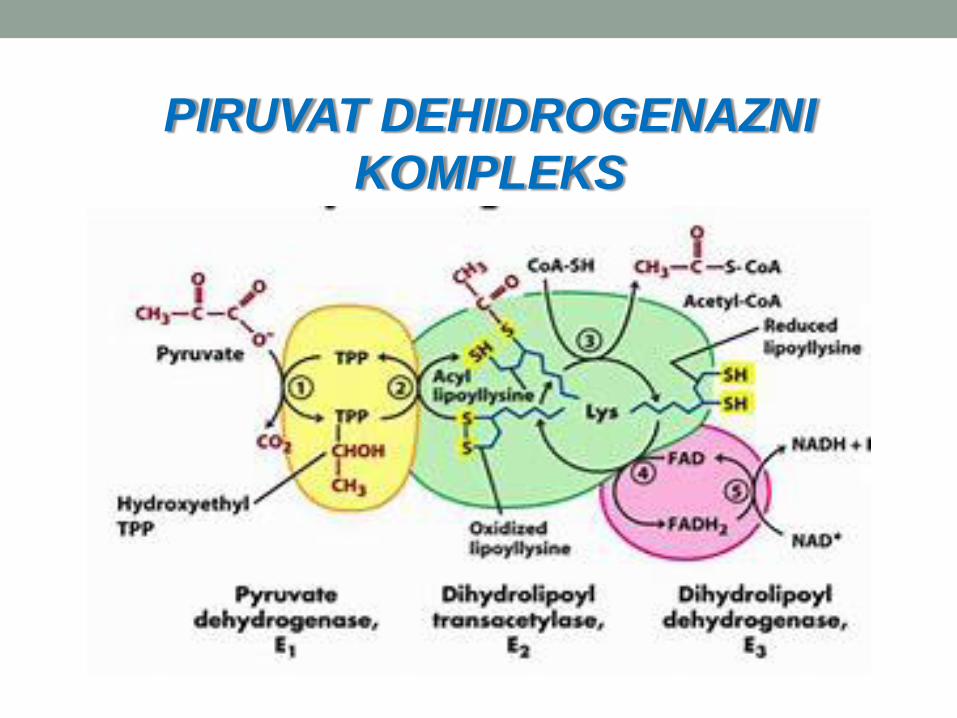
PIRUVAT DEHIDROGENAZNI
KOMPLEKS

glukoza
Glukozo-6-P
1,3-bisfosfoglicerat
3-fosfoglicerat
piruvat
Acetil-CoA
Glutamat i druge AK
MK
Glicerol-P
TAG
Šećer sa 5C
pentoze
2,3-bisfosfoglicerat
serin
alanin
OSTALE ULOGEGLIKOLIZE
TCAciklus
Glikoliza obezbeđuje i prekursore za neke biosintetske puteve (najviše u jetri):
Sinteza nukleotida
Ostali šećeri (npr. UDP-glukoza, manoza, sijalinska kiselina itd.
Aminokiseline
Triacilgliceroli
2,3 bisfosfoglicerat

Regulacija enzima glikolize
Enzimi heksokinaza, fosfofruktokinaza 1 i piruvat kinaza kontrolišubrzinu metaboličkog protoka u glikolizi, jer katališu reakcije koje su podfiziološkim uslovima nepovratne (DG<0), te se smatraju regulatornimenzimima glikolize.
Najvažniji regulatorni korak u glikolizi je reakcija pretvaranja fruktozo-6-fosfata u fruktozo-1,6-bisfosfat, koju katališe fosfofruktokinaza 1 (PFK1). Aktivnost ovog enzima uglavnom je kontrolisana alosteričkimefektorima.
Uopšteno uzevši, može se reći da brzina glikolize u ćeliji zavisi od:• energetskog statusa ćelije (ATP, AMP, Pi) — kada je nivo ATP-a
visok, glikoliza je inhibirana i obrnuto;• unutrašnje sredine ćelije (pH);• dostupnosti alternativnih izvora energije u vidu masnih kiselina ili
ketonskih tela (čijom razgradnjom se dobija acetil-CoA, koji u reakciji saoksalacetatom daje citrat) — dovoljna količina supstrata ulazi u ciklus TCAi glikoliza je inhibirana;
• odnosa insulin/glukagon u krvi (od koga zavisi koncentracijafruktozo-2,6-bisfosfata, kao i aktivnost piruvat kinaze);

KREBSOV CIKLUS



Krebsov ciklus je amfibolički metabolički put u celini smešten u mitohondrijama
Supstrat za ovaj ciklus je acetil CoA; najveći deo nastaje oksidativnom dekarboksilacijom piruvata nastalog u glikolizi pod dejstvom PDH kompleksa
Nastali redukcioni ekvivalenti najvažniji izvor elektrona za lanac prenosilaca elektrona
Potpunom oksidacijom 1 molekula glukoze u glikolizi, Krebsovom ciklusu i lancu prenosalaca elektrona dobija se 29,5-31 ATP, zavisno od toga kojim sistemom povratnog transporta se redukcioni ekvivalenti prebacuju iz citosola u mitohondrije.

Odvijanje Krebsovog ciklusa zavisi od:Odnosa NAD+/NADH unutar mitohondrijaParcijalnog pritiska O2
Energetskog statusa ćelije (alosterička aktivacija izocitrat dehidrogenaze ADP-om)Dostupnosti jedinjenja bogatih energijom (acetil-CoA, sukcinil-CoA)

Niz međuproizvoda ovog ciklusa su početna jedinjenja u sintezi aminokiselina, hema ili steroida.
Anaplerotski putevi obezbeđuju da, uprkos uključivanju u druge puteve, koncentracija međuproizvoda Krebsovog ciklusa unutar mitohondrija bude uglavnim konstantna.














OKSIDATIVNA FOSFORILACIJA

•U eukariota u mitohondrijama.
•Odigrava se redukcija O2 do H2O.
•Donori elektrona su NADH i FADH2.
Oksidativna fosforilacija

• Najveći deo energije za svoj rast, održavanje i rad ćelijadobija oksidacijom ugljenih hidrata i lipida; ovajedinjenja su u značajnoj meri redukovana te mogu dadaju veliku količinu redukcionih ekvivalenata koji seprenose do kiseonika; krajnji prozvodi oksidacije suvoda i ugljen dioksid.
• Pri oksidaciji se oslobađa energija koja se zahvata ipretvara u hemijsku energiju koja se čuva u oblikuvisokoenergetskih veza.

• Oksidacija je u ćeliji postepen proces. U prvoj fazi se redukcioniekvivalenti odvajaju od supstrata delovanjem dehidrogenaza.
• U sledećoj fazi elektroni i protoni se predaju lancu prenosilicaelektrona – respiratornom lancu koji je smešten na unutrašnjojmembrani mitohondrije.
• U trećoj fazi elektroni se prenose od jednog do drugog članarespiratornog lanca do kiseonika, krajnjeg primaoca elektrona.
Zahvatanje energije je ostvareno sprezanjem oslobađanjaenergije, koje se dešava pri prenosu elektrona sa višeg na nižipotencijal, sa reakcijama u kojima se formira visokoenergetskaveza u ATP-u.
• Celokupan proces je nazvan oksidativna fosforilacija.

Oksidativna fosforilacija
Peter Mitchell (1961)
- hemiosmotska teorija –
Transmembranska razlika u koncentraciji protonarezervoar za dobijanje energije iz biološkihoksidacija.

Dobijanje ATP-a u procesu oksidativne fosforilacije podrazumjevaučešće:• davaoca elektrona (NADH ili FADH2),• primaoca elektrona (O2), kao i• unutrašnju membranu mitohondrija koja je nepropusna zaprotone,• sve komponente lanca prenosilaca elektrona i• ATP sintazu.Oksidativna fosforilacija je regulisana stepenom iskorišćavanjaATP-a.

OKSIDATIVNA FOSFORILACIJAReakcije transfera elektrona u mitohondrijama
RESPIRATORNI
LANAC
NADH+H FADHNADPH+
H
Katabolički
putevi
ProteiniUgljeni hidrati
Lipidi
Dehidrogenaze sakupljaju elektrone iz kataboličkih
puteva i prenose do univerzalnih akceptora
elektrona NAD+, NADP+, FMN ili FAD.
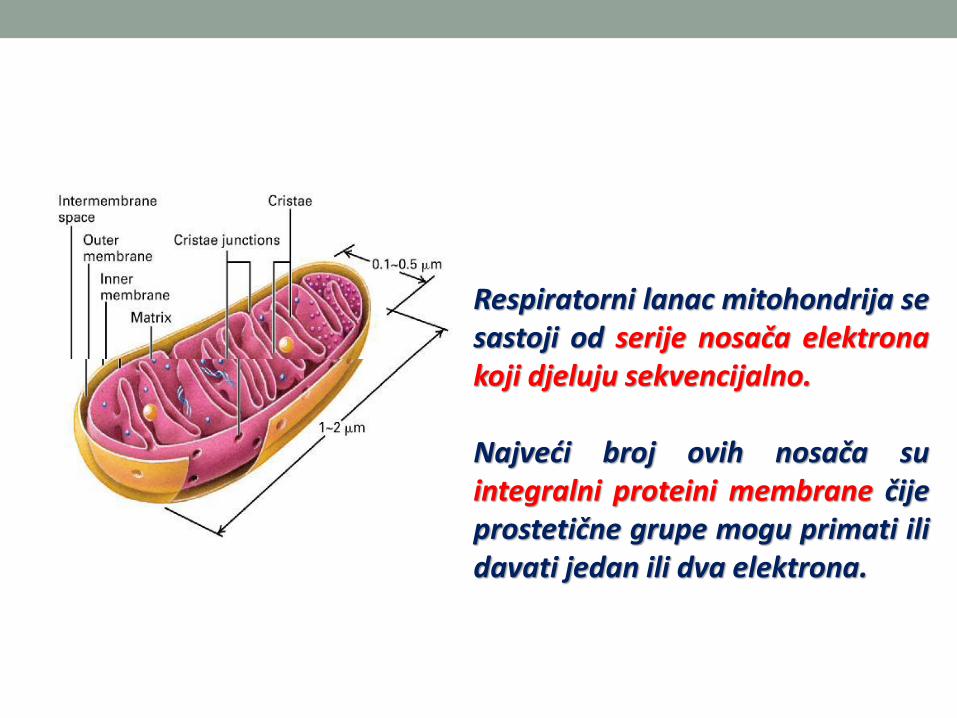
Oksidativna fosforilacija se odvija umitohondrijama
Respiratorni lanac mitohondrija sesastoji od serije nosača elektronakoji djeluju sekvencijalno.
Najveći broj ovih nosača suintegralni proteini membrane čijeprostetične grupe mogu primati ilidavati jedan ili dva elektrona.

• Respiratorni lanac je skup katalitičkih proteina lokalizovanih uunutrašnoj membrani mitohondrija, a čije funkcionalne grupe mogu daprimaju i predaju elektrone. Ti proteini su NADH dehidrogenaza,citohrom b, citohrom c1, citohrom c, citohrom a i citohrom a3.
• Uz ove, respiratorni lanac čine i proteini koji sprežu oslobađanje energijekoje se dešava pri promeni elektrohemijskog potencijala sa sintezomvisokoenergetske veze u ATP-u.
• Koenzim Q, koji prenosi elektrone od flavoproteina do citohroma b takođepripada ovom sistemu. U samom lancu elektronski nosači su poredani porastućem elektropozitivnom potencijalu; ukupna potencijalna razlikaizmeđu prvog i poslednjeg člana lanca je 1,12 V. Ova razlika uelektropozitivnosti omogućava protok elektrona uz oslobađanje energije.
Respiratorni lanac čine prenosioci elektronačiji je redosled određen rastućimelektropozitivnim potencijalom

• Lanac prenosilaca elektrona je organizovan sistem, i prostorno i funkcionalno.
• Članovi lanca prenosilaca elektrona su poredani u definisanom redosledu,prevashodno određenim njihovim redoks potencijalom – od najnegativnijeg ,NADH dehidrogenaze, do najpozitivnijeg, citohroma aa3.
• Prostorno, članovi respiratornog lanca su organizovani u funkcionalne koplekse,smeštene u unutrašnjoj membrani mitohondrija:
Kompleks I – NADH dehidrogenaza i Fe-S centri enzima
Kompleks II – sukcinat dehidrogenaza sa svojim Fe-S centrima i citohrom b
Kompleks III – citohromi b i c1 i specifični Fe-S centar
Kompleks IV – citohromi a i a3 (citohrom oksidaza)
Kompleks V – F0F1 ATP sintaza
Ubihinon spaja kompleksa I, II i III, dok je citohrom c veza između kompleksa III ikompleksa IV.
Respiratorni lanac je organizovan ukomplekse
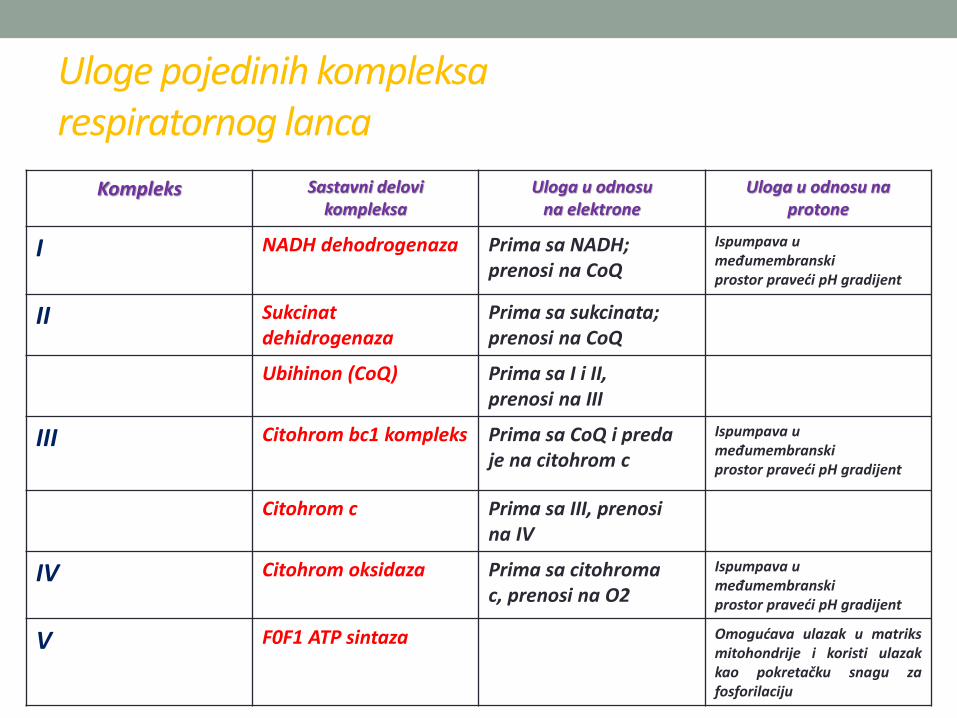
Kompleks Sastavni delovikompleksa
Uloga u odnosuna elektrone
Uloga u odnosu naprotone
I NADH dehodrogenaza Prima sa NADH;prenosi na CoQ
Ispumpava u međumembranskiprostor praveći pH gradijent
II Sukcinatdehidrogenaza
Prima sa sukcinata;prenosi na CoQ
Ubihinon (CoQ) Prima sa I i II,prenosi na III
III Citohrom bc1 kompleks Prima sa CoQ i predaje na citohrom c
Ispumpava u međumembranskiprostor praveći pH gradijent
Citohrom c Prima sa III, prenosina IV
IV Citohrom oksidaza Prima sa citohromac, prenosi na O2
Ispumpava u međumembranskiprostor praveći pH gradijent
V F0F1 ATP sintaza Omogućava ulazak u matriksmitohondrije i koristi ulazakkao pokretačku snagu zafosforilaciju
Uloge pojedinih kompleksarespiratornog lanca

OKSIDATIVNA FOSFORILACIJARespiratorni lanac
Prenosioci elektrona funkcionišu u okviru multienzimskihkompleksa• Kompleksi I i II katalizuju prenos elektrona do koenzima Q• Kompleks III sa redukovanog koenzima Q do citohroma c• Kompleks IV sa citohroma c do O2
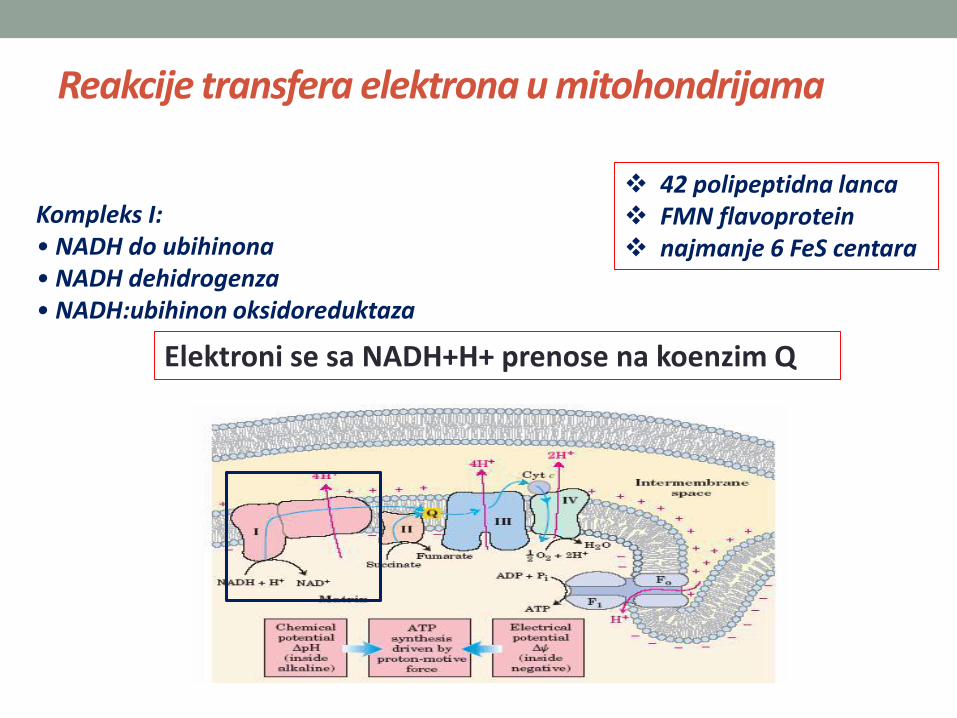
Reakcije transfera elektrona u mitohondrijama
Kompleks I:• NADH do ubihinona• NADH dehidrogenza• NADH:ubihinon oksidoreduktaza
42 polipeptidna lanca FMN flavoprotein najmanje 6 FeS centara
Elektroni se sa NADH+H+ prenose na koenzim Q

Reakcije transfera elektrona u mitohondrijama
FeS 2Fe-2S 4Fe-4S
Gvožđe-sumpor proteiniGvožđe je u asocijaciji sa neorganskim sumporom ili sa atomima sumporacisteinskih rezidua.
• Svi gvožđe sumpor proteini učestvuju u jedno-elektronskomtransferu u kome se jedan atom gvožđa oksiduje ili redukuje.
• Najmanje osam FeS proteina funkcioniše u mitohondrijalnomtransferu.

Reakcije transfera elektrona u mitohondrijama
Kompleks IKatalizuje dva simultana ipovezana procesa.
To je dakle protonska pumpakoja energiju dobija od prenosaelektrona.
Egzergoni prenos hibridnog jona(:H-) sa NADH i protona iz matriksa.
Endergoni prenos 4 protona iz matriksa umeđumembnski prostor.
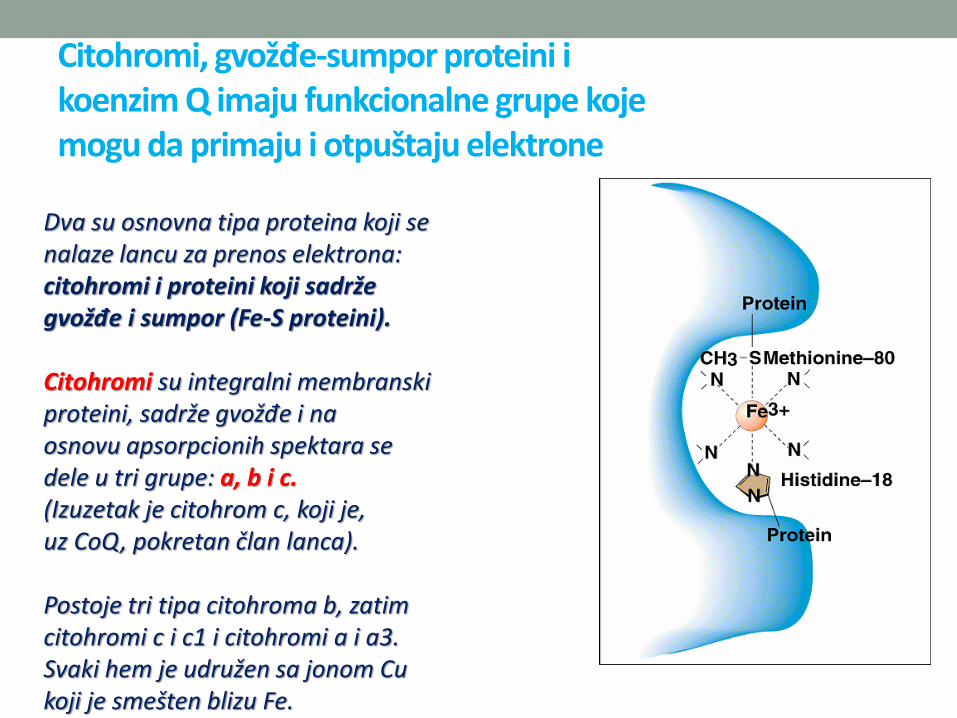
Citohromi, gvožđe-sumpor proteini ikoenzim Q imaju funkcionalne grupe kojemogu da primaju i otpuštaju elektrone
Dva su osnovna tipa proteina koji senalaze lancu za prenos elektrona:citohromi i proteini koji sadržegvožđe i sumpor (Fe-S proteini).
Citohromi su integralni membranskiproteini, sadrže gvožđe i naosnovu apsorpcionih spektara sedele u tri grupe: a, b i c.(Izuzetak je citohrom c, koji je,uz CoQ, pokretan član lanca).
Postoje tri tipa citohroma b, zatimcitohromi c i c1 i citohromi a i a3.Svaki hem je udružen sa jonom Cukoji je smešten blizu Fe.

• Citohromi su jednoelektronski prenosioci – gvožđeprelazi iz +3 u +2, a bakar iz +2 u +1 stanje.
• Uobičajeno je svih 6 koordinativnih veza Fepopunjeno, te se O2 ne može direktno vezati za njih.Izuzetak je citohrom aa3, citohrom oksidaza, jediničlan respiratornog lanca koji može da reaguje sakiseonikom, za koji ima veoma veliki afinitet; reakcijaprenosa redukcionih ekvivalenata na kiseonik je jedinanepovratna reakcija u lancu. Ove dve osobineomogućavaju neprekidni jednosmjerni prenoselektrona, čak i kada je parcijalni pritisak kiseonikamali.
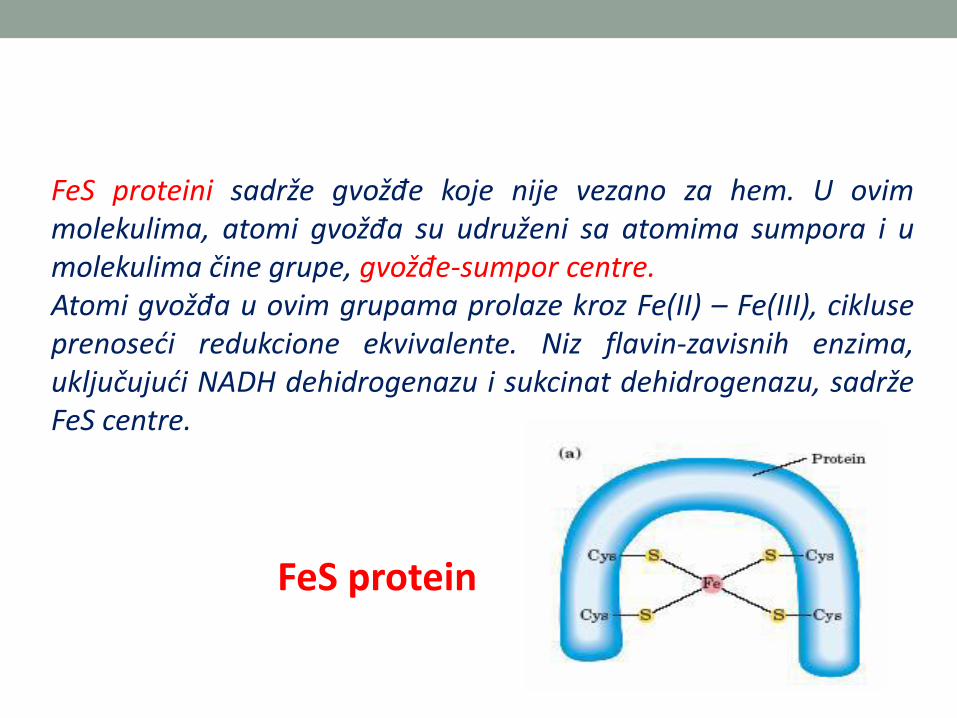
FeS proteini sadrže gvožđe koje nije vezano za hem. U ovimmolekulima, atomi gvožđa su udruženi sa atomima sumpora i umolekulima čine grupe, gvožđe-sumpor centre.Atomi gvožđa u ovim grupama prolaze kroz Fe(II) – Fe(III), cikluseprenoseći redukcione ekvivalente. Niz flavin-zavisnih enzima,uključujući NADH dehidrogenazu i sukcinat dehidrogenazu, sadržeFeS centre.
FeS protein

Hinon rastvorljiv u mastima sa veoma dugim izoprenoidnim bočnim lancem (10izoprenoidnih jedinica - CoQ10). Količinski, postoji znatan stehiometrijski višakubihinona u odnosu na ostale članove respiratornog lanca – CoQ je pokretan isakuplja redukcione ekvivalente sa manje pokretnih flavoproteinskih kompleksa.
Uloga ubihinona je da prikuplja redukcione ekvivalente ne samo sa NADHdehidrogenaze već i sa drugih flavinzavisnih dehidrogenaza (npr. Sukcinatdehidrogenaze i acil–CoA dehidrogenaze masnih kiselina) i da ih predajecitohromima.
Kako ubihinon prima i predaje po jedan elektron u jednom trenutku, zahvaljujućipostojanju semihinonskog oblika, CoQ je tačka gdje dvoelektronski prenos postajejednoelektronski. Pri određenim uslovima u ćeliji moguće je nagomilavanjesemihinonskog oblika CoQ sa posledičnom pojavom slobodnih radikala i mogućimoštećenjima koja iz ovoga proizilaze.
Koenzim Q (ubihinon; CoQ)

• Ubihinon preuzima i redukcione ekvivalente sa flavin–zavisnihdehidrogenaza, kao što je npr. sukcinat dehidrogenaza, enzimciklusa trikarboksilnih kiselina (kompleks II). Direktan prenosredukcionih ekvivalenata na koenzim Q vrši se i sa a-glicerofosfat dehidrogenaze (enzim koji vrši povratni transportredukcionih ekvivalenata iz citosola u mitohondrije) i acil CoAdehidrogenaze iz β-oksidacije masnih kiselina.
• Ubihinon predaje elektrone citohromu b kompleksa III, acitohrom c preuzima elektrone sa ovog kompleksa i prenosi ihdo kompleksa IV, citohrom oksidaze, koja katališe prenoselektrona sa redukovane forme citohroma c na kiseonik, pričemu nastaje voda:
4 cit c (2+) + 4 H+ + O2 → 4 cit c (3+) + 2 H2OPotpunom redukcijom kiseonika nastaje bezopasno jedinjenje, voda.Međutim, ukoliko je kiseonik samo delimično redukovan nastajujedinjenja koja su veoma reaktivna i opasna za ćeliju.

Reakcije transfera elektrona u mitohondrijama
Kompleks II Sukcinat do ubihinona Sukcinat dehidrogenaza– enzim ciklusa trikarbonskih kiselina (CTK)– jedini enzim CTK vezan za membranuPrenosi elektrone sa sukcinata do koenzima Q
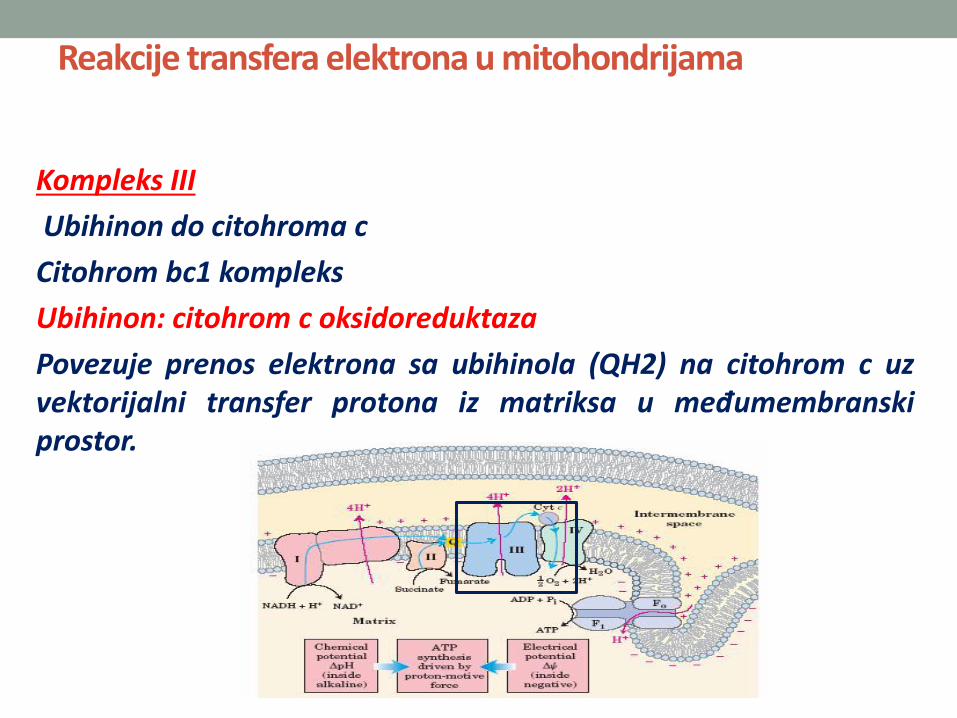
Kompleks III
Ubihinon do citohroma c
Citohrom bc1 kompleks
Ubihinon: citohrom c oksidoreduktaza
Povezuje prenos elektrona sa ubihinola (QH2) na citohrom c uzvektorijalni transfer protona iz matriksa u međumembranskiprostor.
Reakcije transfera elektrona u mitohondrijama

Reakcije transfera elektrona u mitohondrijama
Kompleks IIIDimer – sastavljen od dva identična monomera.Svaki monomer sadrži 11 različitih subjedinica.
Struktura monomeraFunkcionalno jezgro čine tri subjedinice.Citohrom b (zeleno) sa svoja dva hema (bh i b1).Rieske gvožđe-sumpor protein (ljubičast) sa 2Fe-2S centrom (žuto).Citohrom c1 (plavo) sa svojim hemom (c1, crveno).

Kompleks IVCitohrom c do O2Citohrom oksidazaPrenosi elektrone sa citohroma c do molekularnog kiseonikadajući vodu.

Kompleks IV
Redukcija kiseonika sa 4 elektronauključuje redoks centre kojinose po jedan elektron, i morase odvijati bez oslobađanjanekompletno redukovanihintermedijera (slobodni radikali(vodonik peroksid i hidroksilradikal).
Oni ostaju tesno vezani zakompleks do kompletneredukcije do vode.

• Translokaciju protona vrše kompleksi respiratornog lanca, asintezu ATP-a ATP sintaza.
• FoF1 ATP sintaza je enzim koji sintetiše ATP.
• FoF1 kompleks, ATP sintaza, sačinjena je od pet vrsta proteina(a,b,g, d i e), koji čine čvor i peteljku; čvor i peteljka zajedno čineF1, čiji je sastav a3b3gde. Uz to se nalazi osnova, Fo, ugrađena uunutrašnju membranu mitohondrija.
• U osnovi ATP sintaze nalazi se kanal kroz koji prolaze protoni.Prolazak protona daje energiju za obrtanje osnove koje se prenosina peteljku i menja konformaciju proteina u čvoru.
Oksidativna fosforilacija –Sinteza ATP-a
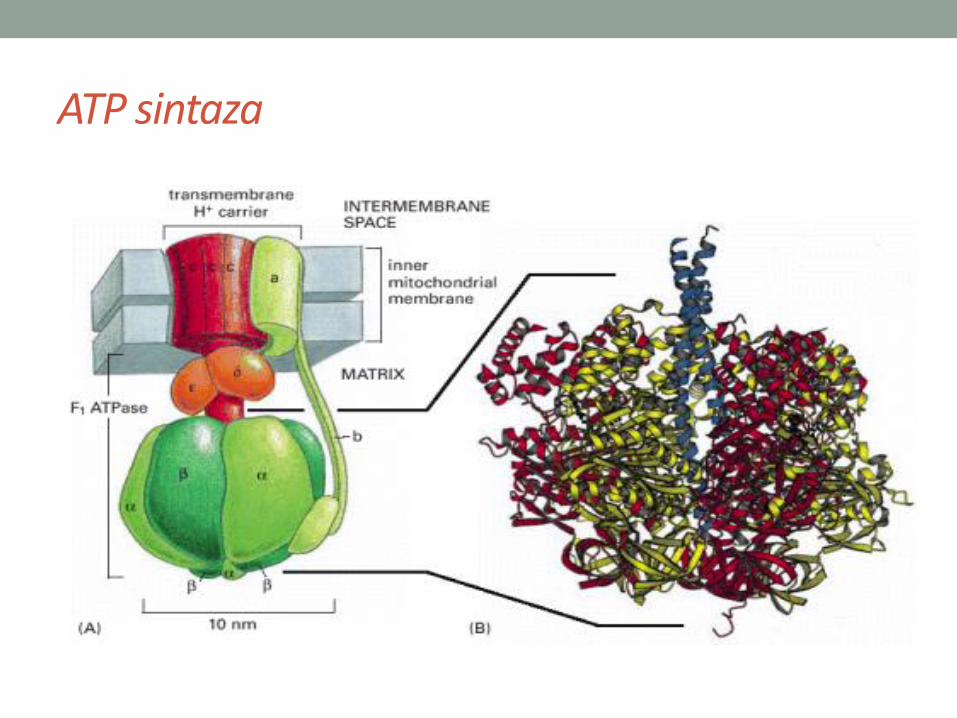
ATP sintaza

Translokacija protona se dešava na tri mesta urespiratornom lancu, u kompleksima I, III i IV
Smatra se da prolazak jednog para elektrona kroz cjelokupni respiratorni lanacispumpava ukupno 10 protona. To je dovoljno za sintezu 2,5 ATP-a (deoenergije se troši za izmjenu ADP:ATP između citosola i mitohondrije).

• Katalitičko mesto ATP sintaze se nalazi na spoju a i b subjedinica. b subjedinicamože da bude u tri konformaciona oblika, otvorenom, opuštenom i stegnutom,što je određeno asimetričnim interakcijama sa centralno postavljenom gsubjedinicom. ADP i Pi su vezani za opušteni oblik, dok se ATP nalazi vezan zastegnuti oblik b subjedinice.
Kako se energija protonskog gradijentapretvara u hemijsku vezu u ATP-u ?
Pri prolasku protona kroz kanal FO komponente, ceo a3b3 skup se okrene, te stegnutomesto postane otvoreno i ATP se otpušta; za mesto koje postaje opušteno vezuju se ADP iPi; na mestu koje prelazi u stegnuto, ADP i Pi daju ATP. Energija je potrebna za otpuštanjeATP-a, za konformacionu promenu koja pretvara stegnutu u otvorenu formu; naime,stvaranje ATP-a od ADP-a i Pi koji su već vezani za opuštenu formu zahteva malu promenuslobodne energije, sve dok je ATP vezan za katalitičko mesto – otpuštanje ATP-a zahtevautrošak energije.Makroskopski - energija je potrebna za pretvaranje ADP-a i Pi iz rastvora u ATP u rastvoru.

Oksidativna fosforilacija –Sinteza ATP-a
Pri prolasku para elektrona kroz respiratorni lanac dolazi do prebacivanja 10 H+iz matriksa u međumembranski prostor.Za sintezu jednog molekula ATP-a potrebno je da kroz protonski kanal prođe 4H+.NADH – 10 protona 2.5 ATPFADH2 – 6 protona 1.5 ATP

• Od svakog NADH+H+ u lanac prenosilaca elektrona uđu 2elektrona, što je ekvivalent redukciji ½O2.
• Ukoliko se 4 protona ispumpaju na nivou kompleksa I, 4 na nivoukompleksa III i 2 protona na nivou kompleksa IV, a procenjuje se daje za sintezu ATP-a potrebno da se 4 protona vrate kroz molekul ATPsintaze, oksidacijom svakog NADH+H+ se dobiju 3 ATP-a, a odFADH2 - 2 ATP. To znači da se samo 30% energije dostupneoksidacijom redukovanih koenzima iskoristi za sintezu ATP-a.
• Deo energije se iskoristi za transport jona, a ostatak se oslobađa uvidu toplote.
Energetski bilans lanca prenosilacaelektrona

• Ćelije mrkog masnog tkiva koriste transport elektrona zaproizvodnju toplote; životinje koje hiberniraju kao i mladunčad(uključujući i čoveka) imaju relativno veću proporciju ovogspecijalizovanog oblika masnog tkiva.
• Veća proizvodnja toplote je važna u održanju telesne temperatureu novorođenčadi jer je njihova telesna površina neproporcionalnovelika u odnosu na telesnu masu; u odraslih je količina mrkogmasnog tkiva veoma mala.
• Mrko masno tkivo (boja potiče od velikog broja mitohondrija iposledično velike količine citohroma) je efikasan proizvođač toplote– 400 W kg-1 u odnosu na 1 W kg-1 u tipičnom tkivu.
Uloga transporta elektrona u proizvodnjitoplote

Ćelije mrkog masnog tkiva dobijenih iz mišićnih
ćelija

U unutrašnjoj membranimitohondrija u mrkom masnomtkivu se nalazi proteintermogenin koji ima funkcijukanala za anjone: omogućavaOH- ili Cl- jonima da brzoprolaze kroz membranu.
Prenos OH- jona iz citosola u mitohondrijalni matriksima isti efekat kao prenos H+ u suprotnom pravcu –poništavanje protonskog gradijenta – što znači datermogenin deluje kao supstanca koja razdvaja prenoselektrona od sinteze ATP-a, sa posledičnimoslobađanjem velike količine toplote.

Brus Spigelman, ćelijski biolog sa Univerziteta Harvard,
načinio jako važan korak ka razumevanju razvića masnog
tkiva. 2009. godine Spigelman i saradnici su pokazali da se
partner protein nazvan C/EBP-beta vezuje za PRDM16
protein, i da se upravo povezivanjem ova dva faktora
mišićne ćelije i ćelije vezivnog tkiva pretvaraju u mrko
masno tkivo.

• Energija dobijena oksidacijom hranljivih materija se prevodi uvisoko-energetske veze adenozin trifosfata (ATP) u procesuoksidativne fosforilacije.
• Najveći deo energije dobijen u procesu oksidacije hranljivihmaterija u ciklusu trikarboksilnih kiselina i drugim kataboličkimputevima se čuva u obliku redukovanih koenzima, NADH iFAD(2H).
• U lancu prenosilaca elektrona NADH i FAD(2H) se oksiduju, pričemu predaju elektrone O2, koji se redukuje u H2O.
• Energija dobijena u redukciji O2 se koristi za fosforilacijuadenozin difosfata (ADP) u ATP dejstvom ATP sintaze (FoF1ATPaza).
• Neto prinos oksidativne fosforilacije je približno 2.5 mola ATP-apo molu oksidaovanog NADH, ili 1.5 mol ATP-a po moluoksidaovanog FAD(2H).