meiosis in coprinus - journal of cell sciencejcs.biologists.org/content/joces/12/1/131.full.pdf ·...
TRANSCRIPT

J. Cell Sci. 12, 131-141 (1973) 131Printed in Great Britain
MEIOSIS IN COPRINUS
IV. MORPHOLOGY AND BEHAVIOUR OF SPINDLE
POLE BODIES
N. B. RAJU AND B. C. LUDepartment of Botany and GeneticsUniversity of Guelph, Guelph, Ontario, Canada
SUMMARY
Meiotic synchrony in the genus Coprinus has permitted the sequential study of spindle polebody (SPB) behaviour through the meiotic process. The SPBs are monoglobular in the youngbasidia immediately after the last premeiotic mitosis. From 10 to 15 h before karyogamy untilpachytene, spindle pole bodies are not found. They become conspicuous in diplotene andpersist until the completion of meiosis. During diplotene, metaphase, anaphase and telophasestages the spindle pole bodies are monoglobular but at late diplotene they duplicate and becomediglobular with an isthmus connecting the 2 globular elements. The spindle pole bodies remainin a diglobular state until diakinesis when the isthmus breaks separating the 2 daughter spindlepole bodies. The diglobular SPBs in late diplotene and prophase II are believed to representthe duplicated form of the monoglobular state. The spindle pole bodies in Coprinus contain nocentrioles. In thin sections the SPBs appear to be fibrillar amorphous structures with a denseinner core surrounded by a less-dense outer zone.
INTRODUCTION
In a comparative study with light and electron microscopy, Lu (1967 a) reportedthe fine structure of centrosomes in Coprinus lagopus. Since then a number ofultrastructural studies revealed different centrosomal forms in Ascomycetes andBasidiomycetes. In the latter, for example, 2 centrosomal types have been reported: amonoglobular or spherical form (Lu, 1967a; Motta, 1967, 1969) and a diglobular ordumbbell form (Girbardt, 1968, 1971; Lerbs & Thielke, 1969; McLaughlin, 1971).Both forms may be found in a single species at different times during the nuclearcycle. Whether the diglobular structure is a duplicated form of the monoglobularcentrosome is still not clear.
In Coprinus the close synchrony of meiotic divisions facilitated the sequential studyof meiosis (Raju & Lu, 1970) and made it possible to correlate the centrosome be-haviour observed with light and electron microscopy to the meiotic sequence. Sincethe meiotic process and the centrosome behaviour in 2 species of Coprinus (C. lagopusand C. comatus) are similar, the sequence of events is illustrated with the best evidenceobtained from both the species.
The centrosome is variously named in the literature (e.g. centriole: Lu, 1967ft;centriolar plaque: Robinow & Marak, 1966; Zickler, 1969; Wells, 1970; Centrioleahnlichekdrper: Lerbs & Thielke, 1969; centrosomal plaque: Zickler, 1970; spindleplaque: Moens & Rapport 1971a, b; Van Winkle, Biesele & Wagner, 1971; centro-
9-2

132 N. B. Raju and B. C. Lu
some: Lu, 1967a; Westergaard & von Wettestein, 1970; McLaughlin, 1971; kineto-chore equivalent: Girbardt, 1971; McCully & Robinow, 1971). All these names refer tothe electron-dense structures on the nuclear envelope which function as spindle polesduring nuclear division. In the light of a recent analysis of centrosomal terminology(Fulton, 1971), the term spindle pole body will be adopted in this paper.
MATERIALS AND METHODS
Basidiocarps of C. lagopus were obtained from cultures on horse dung or a syntheticmedium devised by Rao & Niederpruem (1969). The culture dishes, inoculated with a di-karyatic mycelium, were incubated at 35 °C for 5-6 days until the substrate was covered bymycelium. They were then incubated at 25 °C under a regime of 16 h light and 8 h dark wherethey subsequently produced basidiocarps in 4-5 days. The basidiocarps of C. comatus werecollected from Guelph gardens during the summer and fall months. They were brought intothe laboratory with a block of undisturbed soil and kept in a moist chamber at room temperaturewhere they developed and matured normally. A few gills from the developing basidiocarpswere taken at various time intervals until meiosis was completed and processed for light andelectron microscopy as described below.
Light microscopy
Gills were fixed in modified Lu's fixative (Lu, 1962) containing 9 parts ethanol, 6 parts pro-pionic acid and 2 parts 10% aqueous chromic acid. Since fungal cells are highly basophillic,acid hydrolysis was important for proper staining of chromosomes and spindle pole bodies.A simplified iron haematoxylin method (Henderson & Lu, 1968) was used with slight modifica-tions (Lu & Raju, 1970).
Electron microscopy
Samples were fixed for 2-4 h in 5 % glutaraldehyde in either o-1 M cacodylate buffer atpH 72 or 0067 M phosphate buffer at pH 60. They were washed overnight with the samebuffer and postfixed in 2 % OsO4 for 1-2 h, dehydrated in an ethanol series and embedded ina low viscosity resin mixture (Spur, 1969). Figs. 3 and 7 were taken earlier using OsO4 fixationand Araldite embedding. Sections were cut with a Porter-Blum MT-2 ultramicrotome usinga glass knife and examined with a Philips EM 200 or a Zeiss EM 9 at either 60 or 80 kV.
RESULTS
Light-microscope observations revealed spindle pole bodies at the last premeioticmitosis as well as during meiosis in the basidia of Coprinus. The spindle pole bodiesare well stained and appear to be spherical at the mitotic metaphase (Fig. 1). Thesespherical structures were also seen in the young basidia immediately following mitosis(Fig. 2). However, during the period from 10 to 15 h before karyogamy to pachytene,extensive studies with light and electron microscope failed to reveal any structureanalogous to a spindle pole body. This organelle can be seen again from late pachy-tene/diplotene to the completion of meiosis. At late pachytene/early diplotene aspherical spindle pole body was seen attached to an indented portion of the outermembrane of the nuclear envelope (Fig. 3). Identification of this stage as late pachytene/diplotene was confirmed by the fact that on the same sections synaptonemal complexeswere found. The spindle pole bodies become quite conspicuous during diplotene and

Spindle pole bodies in Coprinus 133
were readily seen by light or electron microscopy (Fig. 4.) In some cells the spindlepole bodies may be found at a distance from the nuclear material since acid hydrolysisin the staining procedure destroys the nuclear envelope. At late diplotene the spindlepole bodies are usually diglobular with 2 globular elements (daughter SPB) connectedby SPB isthmus (Figs. 4, 9). The isthmus stretches and breaks at diakinesis separatingthe 2 daughter spindle pole bodies (Fig. 9), which are spherical during the subsequentmetaphase, anaphase and telophase stages (Figs. 5, 6, 8, 10). At metaphase I, thespindle pole bodies occupy opposite poles at right angles to the basidium. Figs 5 and 6show the ultrastructure of the organelle in the lateral view and the polar view respec-tively. The spindle pole bodies are fibrillar amorphous structures. In the polar view,however, a dark band was seen around the inner dense sphere which is surroundedby a less-dense outer region (Fig. 6). In Coprinus the nuclear envelope is disrupted atmetaphase I (Fig. 5) and reformed in telophase. The spindle pole bodies are sphericalfor a short time at the end of the first meiotic division but soon become diglobularin interkinesis or in prophase II (Figs. 7, 10, 11). The size of spindle pole body atmetaphase I and at the end of the first division is approximately the same (Figs. 8, 10).In prophase II, however, the diglobular spindle pole body is almost twice as large asthe monoglobular one of metaphase I and telophase I suggesting that its duplicationinvolves the synthesis of some SPB material. During the second metaphase and ana-phase stages the spindle pole bodies are monoglobular (Fig. 12). At the end of seconddivision, there is one spindle pole body associated with each nucleus. At this stage, itappears to be monoglobular although the photomicrograph does not clearly illustratethis (Fig. 13). Electron microscopy at this stage revealed only the monoglobular form.
DISCUSSION
Since the spindle pole bodies exist either in monoglobular or diglobular formdepending upon the state of the nucleus (Girbardt, 1971) it is important to under-stand its behaviour in relation to the meiotic sequence. In Coprinus the diglobularspindle pole bodies observed in late diplotene and prophase II are believed to representthe duplicated form.
Although spindle pole bodies are observed in young basidia, their continuedpresence up to late pachytene/early diplotene could not be demonstrated in the presentstudy. Similarly, Zickler (1970) and McLaughlin (1971} failed to find any structurebetween karyogamy and pachytene that resembled a spindle pole body. During latepachytene and early diplotene, Lu (1967 a) showed a monoglobular amorphous struc-ture as the Coprinus spindle pole body. However, as cautioned by Girbardt (1971),unless serial sections are examined even a diglobular form may appear as mono-globular if the sections are cut perpendicular to the axis of the diglobular spindlepole body.
McLaughlin (1971) reported diglobular spindle pole bodies in the basidia of Boletusrubinellus at prekaryogamy, prophase I and interphase I. In our opinion McLaughlin'sobservation of a diglobular form before karyogamy is not conclusive. Meiotic divisionsin Boletus are not synchronous and the stage of basidial development was estimated by

134 N- B- RaJu and B- c- Lu
nuclear size and condition of the cytoplasm, both of which are unreliable criteria. Inelectron micrographs it is difficult to distinguish between late prekaryogamy basidiaand those at prophase II since in both cases 2 nuclei are found without much changein the size and shape of basidia.
Recent observations of Moens & Rapport (1971a) on Saccharomyces cerevisiaesupport our observations in Coprinus. Since meiotic stages could not be identified inthis yeast, time course studies were made after the cells had been transferred to thesporulation medium. Moens & Rapport noted that during the first 4-5 h period thecells have only one indistinct spindle pole body which they called a spindle plaque.After about 8 h on the sporulation medium many of the larger yeast cells have2 plaques, side by side, connected by an isthmus (plaque bridge). In a more recentpaper, they showed the presence of polycomplexes in the cells after 8 h on the sporula-tion medium - which suggested that the cells were in diplotene (Moens & Rapport,19716). Thus, in yeast, the spindle pole body is single in the early stages of meiosisbut becomes double at a later stage (diplotene?). In our opinion the diglobular spindlepole body reported in Basidiomycetes should be considered a duplicated one. Further-more, in Coprinus the high frequency of diglobular spindle pole bodies in late di-plotene suggests that there is considerable time between their duplication andseparation.
In the mitotic cell cycle of Polystictus versicolor, Girbardt (1971) showed that thekinetochore equivalent (spindle pole body) becomes diglobular in interphase as earlyas 30-40 min following the previous nuclear division. He suggested that the changeto the diglobular condition in interphase was an indication that DNA synthesis hadstarted. The apparent correlation of SPB duplication and DNA synthesis is fortuitoussince the SPB can duplicate in the absence of gross DNA synthesis in interkinesis orprophase II (Figs. 10, 11).
McLaughlin (1971) suggested that at karyogamy both the spindle pole bodiesdisappear and one reforms during prophase. He drew parallels between the fate ofthe centrosome at fertilization in animals and karyogamy in Boletus. Although thefailure to find spindle pole bodies at karyogamy and pachytene is believed to be dueto the disappearance of this organelle, which is synthesized de novo when needed assuggested by Fulton & Dingle (1971), other explanations are equally attractive. Thestructure may actually be present at this time in the monoglobular form but probablyin an indistinct state. Unless a thorough search for the SPB or its precursor was carriedout as Fulton & Dingle did in Naegleria, this latter possibility cannot be ruled out.
The spindle pole bodies in Ascomycetes and Basidiomycetes lack the 9 triplettubular structure that is characteristic of centrioles in animals, algae, and lowerfungi (see reviews by Went, 1966; Pickett-Heaps, 1969; Fulton, 1971). Nevertheless,in Ascobolus and Podospora, Zickler (1970) showed that at metaphase and anaphasethe spindle pole body consisted of 2 zones situated on each side of the nuclear envelope:an electron-opaque outer zone and a less-dense inner zone in which most of themicrotubules end. Some form of zone differentiation is evident, although to a lesserextent, in other Ascomycetes (Moens & Rapport, 1971a; McCully & Robinow, 1971).The presence of a darker band in the central core of the Coprinus spindle pole body

Spindle pole bodies in Coprinus 135
is noteworthy although its significance cannot be established at this time. Its similarityto the dark band of the spindle pole body in Ascomycetes may be of some phylogeneticsignificance.
The authors wish to thank the National Research Council of Canada, Ottawa, for financialsupport.
REFERENCES
FULTON, C. (1971). Centriole. In Origin and Continuity of Cell Organelles (ed. J. Reinert &H. Ursprung), pp. 170—213. Berlin: Springer-Verlag.
FULTON, C. & DINGLE, A. D. (1971) Basal bodies, but not centrioles in Naegleria. J. Cell Biol.51, 826-836.
GIRBARDT, M. (1968). Ultrastructure and dynamics of the moving nucleus. In Aspects of CellMotility, 22nd Symp. Soc. exp. Biol. (ed. P. L. Miller), pp. 249-259. Cambridge: UniversityPress.
GIRBARDT, M. (1971). Ultrastructure of the fungal nucleus. II . The kinetochore equivalent(KCE). J. Cell Sci. 9, 453-473-
HENDERSON, S. A. & Lu, B. C. (1968). The use of haematoxylin for squash preparations ofchromosomes. Stain Technol. 43, 233—236.
LERBS, V. & THIELKE, C. (1969). Die Entstehung der Spindel wahrend der Meiose von Coprinusradiatus. Arch. Mikrobiol. 68, 95-98.
Lu, B. C. (1962). A new fixative and improved propionocarmine squash technique for stainingfungus nuclei. Can. J. Bot. 40, 843-847.
Lu, B. C. (1967a). Meiosis in Coprinus lagopus: a comparative study with light and electronmicroscopy. J. Cell Sci. 2, 529-536.
Lu, B. C. (19676). The course of meiosis and centriole behaviour during the ascus developmentof the Ascomycete Gelasinospora calospora. Chromosoma 22, 210-226.
Lu, B. C. & RAJU, N. B. (1970). Meiosis in Coprinus. II. Chromosome pairing and the lamp-brush diplotene stage of meiotic prophase. Chromosoma 29, 305—316.
MCCULLY, E. K. & ROBINOW, C. F. (1971). Mitosis in the fission yeast Schizosaccharomycespombe: A comparative study with light and electron microscopy. J. Cell Sci. 9, 475-507.
MCLAUGHLIN, D. J. (1971). Centrosomes and microtubules during meiosis in the mushroomBoletus rubinellus. J. Cell Biol. 50, 737-745.
MOENS, P. B. & RAPPORT, E. (1971a). Spindles, spindle plaques, and meiosis in the yeastSaccharomyces cerevisiae (Hansen). J. Cell Biol. 50, 344-361.
MOENS, P. B. & RAPPORT, E. (19716). Synaptic structures in the nuclei of sporulating yeast,Saccharomyces cerevisiae. J. Cell Sci. 9, 665-677.
MOTTA, J. J. (1967). A note on the mitotic apparatus in the rhizomorph meristem of Armillariamellea. Mycologia 59, 370-375.
MOTTA, J. J. (1969). Somatic nuclear division in Armillaria mellea. Mycologia 61, 873-886.PICKETT-HEAPS, J. D. (1969). The evolution of the mitotic apparatus: an attempt at com-
parative ultrastructural cytology in dividing plant cells. Cytobios 3, 257-280.RAJU, N. B. & Lu, B. C. (1970). Meiosis in Coprinus. I II . Timing of meiotic events in C. lagopus
(sensu Buller). Can. J. Bot. 48, 2183-2186.RAO, P. S. & NIEDERPRUEM, D. J. (1969). Carbohydrate metabolism during morphogenesis of
Coprinus lagopus (sensu Buller). J. Bact. 100, 1222-1228.ROBINOW, C. F. & MARAK, J. (1966). A fiber apparatus in the nucleus of the yeast cell. J. Cell
Biol. 29, 129-151.SPUR, A. R. (1969). A low-viscosity epoxy resin embedding medium for electron microscopy.
J. Ultrastruct. Res. 26, 31-43.VAN WINKLE, W. B., BIESELE, J. J. & WAGNER, R. P. (1971). The mitotic spindle apparatus of
Neurospora crassa. Can. J. Genet. Cytol. 13, 873-887.WELLS, K. (1970). Light and electron microscope studies of Ascobolus stercorarius. I. Nuclear
divisions in the ascus. Mycologia 62, 761-790.WENT, H. A. (1966). The behavior of centrioles and the structure and formation of the
achromatic figure. Protoplasmatologia 6, 1-109.

136 TV. B. Raju and B. C. Lu
WESTERGAARD, M. & VON WETTESTEIN, D. (1970). The nucleolar cycle in an Ascomycete.C. r. Trav. Lab. Carlsberg 37, 195-237.
ZICKLER, D. (1969). Sur l'appareil cinetique de quelques Ascomycetes. C. r. hebd. Seanc. Acad.Sci., Paris 268 D, 3040-3042.
ZICKLER, D. (1970). Division spindle and centrosomal plaques during mitosis and meiosis insome Ascomycetes. Chromosoma 30, 287-304.
{Received 8 May 1972)
ABBREVIATIONS ON PLATES
cchn
spindle pole bodychromosomenucleus
nenu
nuclear envelopenucleolus
Figs. 1-4. Spindle pole body behaviour during meiosis in C. lagopus.
Fig. 1. Metaphase of premeiotic mitosis. Spindle pole bodies are spherical (mono-globular), x 2500.
Fig. 2. Young basidia 15 h before karyogamy. One spindle pole body is visible ineach basidium. x 2500.
Fig. 3. Electron micrograph of a basidium at early diplotene. Spindle pole bodyis attached to an indented portion of the nuclear envelope. OsO4 fixation, x 60000.
Fig. 4. Electron micrograph at late diplotene. Spindle pole body is diglobular.Glutaraldehyde-OsO4 fixation. x 46000. The inset is a photomicrograph of abasidium at late diplotene. x 2500.
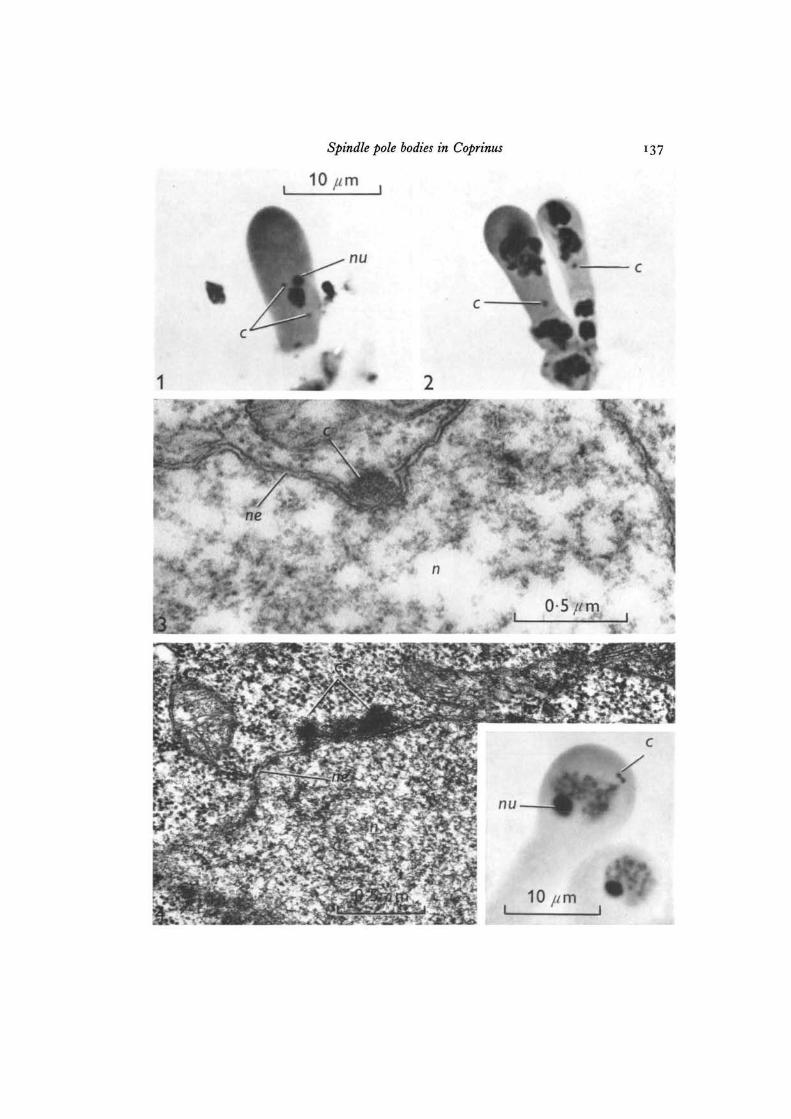
Spindle pole bodies in Coprinus
10//m .
137

138 N. B. Raju and B. C. Lu
Fig. 5. Lateral view of a section through the spindle at metaphase showing spindlepole bodies. Glutaraldehyde-OsO4 fixation, x 40000.Fig. 6. Polar view of a section through a spindle pole body at metaphase I. Notethe dark band (arrowed) around the inner dense sphere. Glutaraldehyde-OsO4fixation, x 60000.Fig. 7. Electron micrograph of a section through a diglobular spindle pole body atprophase II. OsO4 fixation, x 60000.
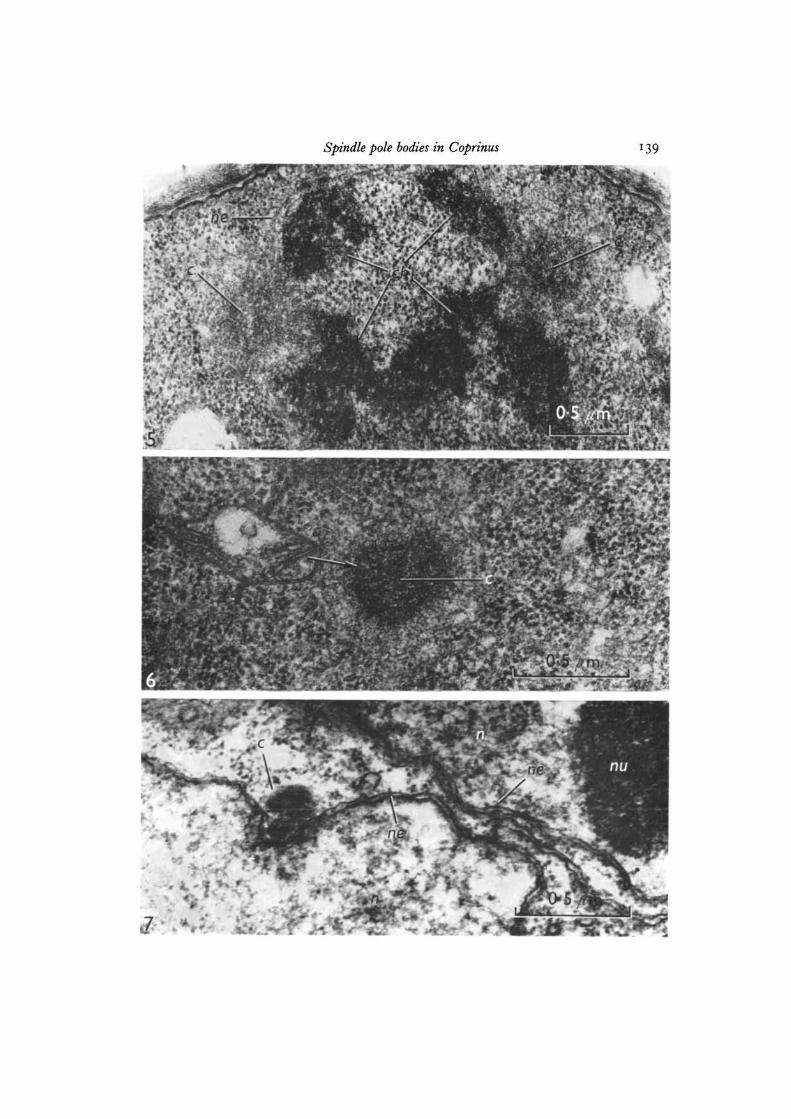
Spindle pole bodies in Coprinus

140 N. B. Raju and B. C. Lu
Figs. 8-13. Photomicrographs showing spindle pole body behaviour during meiosis inCoprinus comatus (Fig. 12, C. lagopus). All figures are x 2500.
Fig. 8. Diplotene to metaphase I: 1, monoglobular spindle pole body at diplotene;2, diglobular spindle pole body at diplotene; 3, monoglobular spindle pole body atmetaphase I. Note the size of the nucleolus in the 3 basidia.
Fig. 9. Daughter spindle pole bodies separating at diakinesis showing SPB isthmus(arrowed).
Fig. 10. The basidia (1 and2) are at the end of the firstmeiotic division. Spindle polebodies are monoglobular.
Fig. 11. Prophase II. Spindle pole bodies are diglobular.Fig. 12. The spindle pole bodies at anaphase II are monoglobular (C. lagopus).Fig. 13. Tetrad stage. One spindle pole body is associated with each of the 4 nuclei.
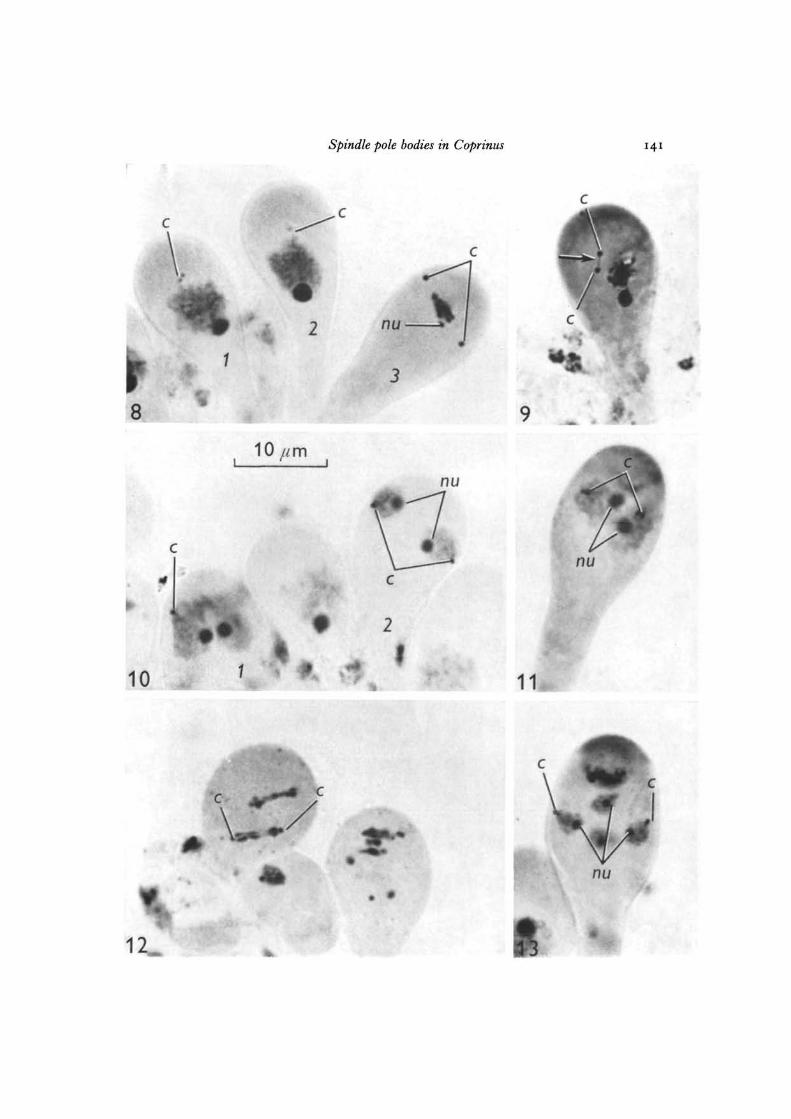
Spindle pole bodies in Coprinus
r «•
14I
c
nu
10
#-fc
2
9 ^
11
i nu
