mapping of the combining sites of antibodies specific to polyalanine chains
TRANSCRIPT

MAPPING OF THE COMBINING SITES OF ANTIBODIES SPECIFIC TO POLYALANINE CHAINS
Israel Schechter*
Department of Chemical Immunology The Weizmann Institute of Science
Rehovot, Israel
INTRODUCTION
Information on the size and nature of the antibody combining site is ob- tained from studying the interaction of antibodies with haptens.' Since it was found to be rather large (e.g., it can accommodate up to 6 glucose units in anti- bodies to polysaccharides,'? or 3-7 amino acid residues in antibodies to poly- peptides 3-11, 2g), for the purpose of mapping the antibody combining site it is suggested that it be divided into several subsites.l2, l3 A subsite is defined as the region on the antibody molecule that interacts with a particular portion of the antigenic determinant (or hapten). Knowing the size of the binding site (or the number of subsites), one can proceed to map it out in terms of the particular interactions which the various subsites can exhibit.14, l5 These include binding forces as well as geometrical factors such as stereospecificity and space limitation. This approach was used with advantage in exploring binding proper- ties of chain-splitting enzymes such as Iysozyme,16, 17 papain,l8-21 carboxy- peptidase-A,?*P ?:{ and For example, it was found that papain can interact simultaneously with a stretch of seven amino acid residues in a peptide chain. In terms of the subsite concept, the active site of this enzyme is com- posed of seven subsites. Four subsites were located on one side of the catalytic point and three on the other. It was possible to evaluate the degree of stereo- specificity in the various subsites and to determine the binding energies of differ- ent amino acid side chains in two subsites.
Antibodies specific to poly-L(or D)-alanine chains carrying free &-amino groups are the subject of the present investigation. In this case, the combining site is visualized as being composed of several subsites, each accommodating one amino acid residue of the antigenic determinant. Subsite S, interacts with the N-terminal alanine residue (which is the most exposed one in the immuno- gen), whereas the adjacent residues in the antigenic determinant (or hapten) occupy subsites S, and so on (FIGURE 1 ) . The extent and properties of the binding site are deduced from the correlation between the structural features of series of haptens and their strength of interaction with the antibody. The types of structural changes applied here are: ( 1 ) elongation of the peptide chain of the hapten; (2) blocking of the amino function or the carboxyl function of the peptides; (3) replacement of an L-alanine residue in the peptide by its D antipode; (4) replacement of one 1.-alanine residue by a glycine, L-phenyl- alanine or L-valine residue. The influence of these changes on antibody-hapten
* Present address: National Institute of Child Health and Human Development, National Institutes of Health, Bethesda, Md. 20014.
394

Schechter: Mapping of Antibody Combining Sites
+ H,N. Alo- Alo- Alo- Ale- Ala-
ANTIBODY
395
FIGURE 1. Schematic representation of the complex formed between a polyalanine chain and its specific antibody. The combining site of the antibody is composed of three subsites (SI to S3), each accommodating one amino acid residue of the anti- genic determinant. The N-terminal alanine residue occupies Si, the penultimate residue SZ, the third residue S3, and the other alanine residues are outside the region of the combining site.
combination is taken as evidence of interaction of the modified portion of the peptide with the binding site.
Previous investigations on antibodies to poly-L-alanine and poly-Dalanine determinants have shown that the antibodies are strictly stereospecific, as ex- pected.”, s, From the capacity of various alanine peptide disastereoisomers t o inhibit the precipitin reaction it was concluded that, in both systems, the size of the combining site is such as to accommodate up to 3-4 alanine residues, and that the region of the antigenic determinant furthest removed from the protein carrier (the N-terminal alanine residue) is of paramount importance in determining the specificities of the antibodies formed.5! More recently, anti- bodies to poly-o-alanine chains were fractionated by adsorption on an im- munoadsorbent and stepwise elution with di-D-alanine and then with tetra-D- alanine. The main difference between the two fractions was that di-Dalanine reacted much better with the first antibody fraction as compared to the second fraction. Tetra and penta-o-alanine reacted with both fractions in a similar manner, as manifested by the capacity of the peptides to inhibit the precipitin reactions.?R
In the present report, detailed analysis of the binding sites of anti-poly-r. (or o)-alanine antibodies is described in terms of the subsite concept. A more refined fractionation of the antibodies was achieved by using for elution di, tri, and tetraalanine peptides. In each purified antibody fraction, the number of subsites was estimated and the association constants of series of alanine peptides of increasing size were measured by equilibrium dialysis. T h e binding energy of the ,8 methyl side chain of alanine, extent of stereospecificity and space limi- tation in the various subsites were also The results led to the conclusion that the subsites exhibit a high degree of heterogeneity with respect to interaction with an alanine residue. The same subsite (say, S,) in the three antibody fractions and the various subsites (Sl, S2, etc.) in the same fraction behaved differently. In this system antibodies with larger combining sites did not show increased binding capacity. The results obtained may provide a clue for some architectural features of the combining site. It is also shown that more precise information on the combining sites is obtained by comparing pairs of haptens of equal size that differ from one another in a particular feature rather than from studying the effect of elongation in a homologous series of haptens.

396 Annals New York Academy of Sciences
MATERIALS AND METHODS
Peptides
The synthesis and characterization of free alanine peptide diastereoisomers, of N-acetyl peptides and of the amides of L-alanine peptides used as inhibitors, was described before.l1? 30 Other peptides were prepared by the hydroxysuccini- mide method.31 The purity of these peptides was higher than 99%, as checked by high voltage paper electrophoresis at pH 1.4 and by chromatography in butanol-acetic acid-water. The labeled hapten, *4C-~Ala3-Gly, was prepared by coupling of benzyloxycarbonyl-tri-D-alanine with 2-l"C-glycine by the hydroxy- succinimide method, and subsequent removal of the benzyloxycarbonyl group by catalytic hydrogenation. The labeled free tetrapeptide had a specific activity of 1.5mclmmole and the radiochemical purity was higher than 99.5%.37
Polyalanyl Proteins
The N-carboxy-L (or D) -alanine anhydride was reacted with human serum albumin (HSA) or rabbit serum albumin (RSA) in a water-dioxane mixture as described b e f ~ r e . ~ Analytical data of the protein conjugates prepared are given in TABLE 1. In this polymerization procedure the attached polyalanine chains carry free N-terminal a-amino groups.43
Immunoadsorbents
The water insoluble poly-L(or D ) -AlaRSA-cellulose conjugates were pre- pared according to published procedure.32 About 150 q g of purified antibodies could be extracted from the appropriate antiserum by means of 1 g immuno- adsorbent.ll9 **
TmLe 1
CHARACTERIZATION OF POLYALANYL PROTEINS
Ncarboxy- Moles of Designation L(or D)-alanine Alanine and No. of Anhydride Attached per Derivative (g/g protein) Mole Protein
poly-L-AlaHSA (1330) 0.8 1 20 poly-L-AlaRSA (569) 0.8 222 poly-D-AlaHSA (1341) 0 . 8 128 poly-D-AlaRSA (570) 0 . 8 205
Moles of Peptide Chains
per Mole Derivative t
25 26 23 25
Average No. of Alanine
Residues per Chain
4 . 8 8.5 5 .4 8 . 2
Calculated from amino acid analyses of the protein before and after polyalanylation.6 t Calculated from the lysine content of the derivative after deamination with nitrous
acid.6, U

Schechter: Mapping of Antibody Combining Sites 397
Antisera
Randomly bred rabbits (2.5-3 kg) of both sexes were used. Four injections of poly-L(or D)-AlaHSA (10 mg per injection) in complete Freund's adjuvant were administered at fortnightly intervals. The first injection was divided among the four footpads, the other three were injected intramuscularly. Bleeding was started after the second injection and then at weekly intervals for another two months. Purified antibody fractions were prepared from pooled antisera (P-4 to P-8) obtained from groups of four to six animals and from antisera collected from one rabbit (1-1).
Purified Antibody Fractions
The immunospecific isolation of antibodies from the sera was achieved with the aid of immunoadsorbents as described.11, 28, 32 Antisera from rabbits im- munized with poly-L-AlaHSA (P-4 to P-7) were adsorbed on poly-L-AlaRSA- cellulose conjugate, sera from animals immunized with poly-D-AlaHSA (P-8 and 1-1) were adsorbed on poly-D-AlaRSA-cellulose conjugate. The adsorbed antibodies were eluted by stepwise extraction with 0.01 M phosphate buffer p H 7.3-0.9% sodium chloride (PBS) solutions containing L-alanine peptides (P-4 to P-7) or D-alanine peptides (P-8 and 1-1 ) , and finally with 0.1 M acetic acid. After dialysis against PBS and concentration by negative pressure ultrafiltration the antibody solution was ready for use. The antibodies of P-5, P-6, P-7, P-8, and 1-1 were dissociated from the immunoadsorbent by stepwise extraction with dialanine, trialanine, tetraalanine, and acetic acid to yield Ab-2, Ab-3, Ab-4, and Ab-Ac fractions, respectively. The P-4 and portions of P-5 and P-8 were extracted by tetraalanine (without prior elutions with di and trialanine) and then by acetic acid to yield Ab-4, and Ab-Act fractions, respectively. The amount of antibody eluted from the immunoadsorbent was calculated from the absorbance of the solution at 280 mp, using the value Ei;Yrnp = 1.4 for 1 mg/ml of IgG. Some fractions were run in the ultracentrifuge and one peak was observed with values of szow = 6.2-6.48, corresponding to that of IgG. The peptide concentrations employed for extraction, the amounts of antibodies ob- tained and their precipitability with antigens are given in TABLE 2. Since L-alanine peptides are readily degraded in serum the purified antibody frac- tions were kept with 0.01M tetra-L-alanine under the conditions of inhibition experiments (see below). The reaction mixtures were then analyzed by high voltage paper electrophoresis at pH 1.4 and by paper chromatography in butanol-acetic acid-water. It was found that all fractions were devoid of any proteolytic activity."
Quantitative Precipitin Reactions
To a constant volume of antiserum or of purified antibody fraction (0.2 to 0.5 ml) increasing amounts of the precipitant (poly-L(or D)-AlaRSA) dissolved in PBS were added, and the final volume was brought to 1.7 ml with PBS. The reaction mixture was kept at 36" for 30 min and then at 5' for 18 hours. Precipitates formed were separated by centrifugation, washed twice with 2.0 ml of PBS, dissolved in 1.1 ml of 0.1N sodium hydroxide, and the absorbancy at 280 mp was determined.

398 Annals New York Academy of Sciences
TABLE 2
PURIFIED ANTIBODY FRACTIONS
Antiserum Source
Antibody Fraction P-4 P-5 P-6 P-7 P-8 1-1
Ab-2 dialanine (mM) * Percent of total t Percent precipitated $
Ab-3 trialanine (mM) Percent of total Percent precipitated
Ab-4 tetraalanine (mM) Percent of total Percent precipitated
Ab-Ac percent of total Percent precipitated
Ab-4t tetraalanine (mM) Percent of total Percent precipitated
Ab-Act percent of total Percent precipitated
10 13.1 60
4 13.1 67
9 16.6 80
55.6 71
11 8 47 35.7 77 70
38 62.1 58 64
10 12.2 63
4 12.4 75
10 11.7 83
54 70
10 17.7 57
4 14.1 77
4 12.8 82
45.9 55
10 23 88
5 28 87
5 10 n.d.
17 n.d.
5 77 83
22 n.d.
8 31.6 n.d.
5 39.8 n.d.
5 12.6 n.d.
16 n.d.
Antisera were adsorbed on water insoluble immunoadsorbent containing poly-L-AlaRSA (in the case of P-4 to P-7) or poly-o-AlaRSA (P-8 and 1-1). Purified antibody fractions were obtained by stepwise extraction with L-alanine peptides (P-4 to P-7) or D-alanine peptides (P-8 and 1-1) and finally with 0.1 M acetic acid; n.d.: not determined.
* Peptide concentration in the PBS solution used for extracting the antibody from the immunoadsorbent.
t Total refers to the antibody content of the serum which is taken as 100%. 1 The percent of purified antibody precipitated by poly-L-AlaRSA (P-4 to P-7) or poly-
D-AlaRSA (P-8 and 1-1).
The percent of purified antibody fraction precipitated by antigen was cal- culated from the absorbancy of the dissolved precipitate and of the supernatant obtained at the optimal zone of the precipitin curve, and both values agreed within 5%.
Quantitative Inhibition of the Precipitin Reaction
Reaction mixtures contained purified antibodies, precipitating antigen and peptide. A number of concentrations of peptide were employed to determine the point of 50% inhibition. Further details were published e l ~ e w h e r e . ~
Equilibrium Dialysis Experiments
The direct and competitive binding experiments were carried out with the

Schechter : Mapping of Antibody Combining Sites 399
specially designed pyrex cells described by K a r u ~ h . ~ ~ Solutions were made u p in PBS. One ml of purified antibody solution (5 x lo-’ to 5 x M, based on a molecular weight of 140,000) was placed in one compartment of the cell, and 1 ml of 14C-DAla,-Gly was placed in the other. In the direct binding experi- ments, the initial concentrations of the labeled hapten were in the range of 1 0 - 7 - 1 0-4 M. In the competitive binding experiments, increasing concentrations of the inhibitor (cold peptide) were added to solutions containing a constant concentration of 14C-DAh,-Gly ( 1-2 x 1 O-c M, depending on the antibody frac- tion and inhibitor employed). The cells were rotated at a speed of 5 rpm a t + 4 O for 44 hr. The radioactivity of samples (0.5 ml) withdrawn from both compartments was measured in a Packard liquid scintillation counter. In each experimental run, controls in which a number of concentrations of labeled hapten were equilibrated against PBS were also included. These controls validated that equilibrium between the two compartments was achieved. In addition it was found that the amount of labeled hapten adsorbed on the cellu- lose membrane and glass walls was very small (2-3% ), and it was neglected in the calculation of free and bound hapten concentrations.
In direct binding experiments, the Scatchard plot 34 was used to determine the valency (n) of the antibody fractions by extrapolation of the curve t o infinite tC-DAh,-Gly concentration. The Sips distribution function 3 6 - 36 was used to calculate the average intrinsic association constant (KO) and the heterogeneity index ( a ) by plotting log r / ( n - r ) vs. log c, where r is the average number of labeled peptide molecules bound per antibody molecule a t the equilibrium concentration of the free labeled peptide (c).
The binding of cold alanine peptides was determined indirectly by measur- ing the capacity of these peptides to inhibit the binding of the labeled hapten. The inhibition constants ( K i ) of the peptides were calculated by making use of equations proposed by Karush.:l:’ K i value, which very closely represents the average association constant of the inhibitor in the absence of l4C-~Ala,-G1y, is obtained when equimolar amounts of cold and labeled peptide are bound per mole antibody.:lJ For this purpose, K , values determined at various in- hibitor concentrations were plotted against the average number of moles in- hibitor bound per mole antibody (ii). The average number of molecules of labeled peptide bound per antibody molecule in the presence of inhibitor ( r ’ ) at each point was also known. Thus, by interpolation, it was possible to deter- mine the K i value when ri = r‘ ( FIGURE 2 ) . In most experiments r, = r’ = 0.6, the range being from 0.52-0.78 for all antibody fractions. FIGURE 2 shows that the measurcd K i decreases with increasing values of T i , that is by in- creasing inhibitor concentration. This trend is expected, since the increased inhibitor concentration brings up interaction with antibody molecules of lower affinity, therefore the apparent K i decreases. There is good agreement between the K , and K, values for the binding of DAIa,,-Gly determined indirectly (using cold peptide, see TABLE 12) and by the direct method (using labeled peptide, see TABLE 11 ).
RESULTS AND DISCUSSION
Results obtained with purified antibodies prepared by extraction with alanine peptides dissolved in PBS will be discussed mainly. I t was previously shown that

400 Annals New York Academy of Sciences
some damage is induced by treatment of antibodies with acetic acid. Purified antibodies prepared by elution with peptides in PBS were subsequently kept in 0.1 M acetic acid for one hour a t 37". As a result, the capacity of peptide-
FIGURE 2. Inhibition constants (KI) of amides of o-alanine peptides determined by equilibrium dialysis (at +4"). A number of concentrations of the cold peptides were used to inhibit the binding of 14C-~Ala&ly to Ab-3 fraction of P-8. f i r average number of moles inhibitor bound per mole antibody. r', average number of moles of 14C-~Ala3-Gly bound per mole antibody in the presence of inhibitor. Full symbols, Ki values at a given ri. Empty symbols, r' values at a given KI. The inter- section point gives Ki when ri = r'. The symbols are: 0 , 0, HmAla.NHz (lower); H, 0, HeDAlazaNHz; A, A, He~Ala3aNH2; 0 , 0, H-oAla.NH2 (upper).
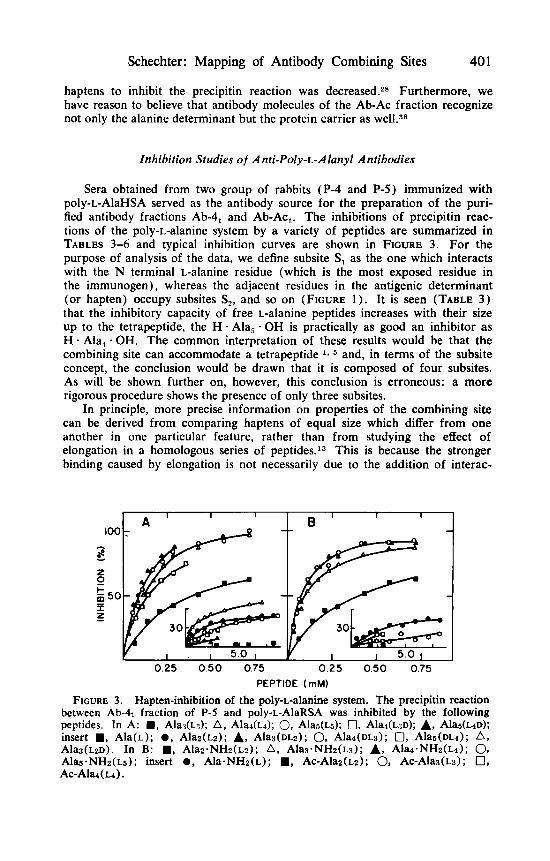
Schechter : Mapping of Antibody Combining Sites 40 1
haptens to inhibit the precipitin reaction was decreased.28 Furthermore, we have reason to believe that antibody molecules of the Ab-Ac fraction recognize not only the alanine determinant but the protein carrier as
Inhibition Studies of Anti-Poly-~-Alanyl Antibodies
Sera obtained from two group of rabbits (P-4 and P-5) immunized with poly-L-AlaHSA served as the antibody source for the preparation of the puri- fied antibody fractions Ab-4, and Ab-Act. The inhibitions of precipitin reac- tions of the poly-L-alanine system by a variety of peptides are summarized in TABLES 3-6 and typical inhibition curves are shown in FIGURE 3. For the purpose of analysis of the data, we define subsite S, as the one which interacts with the N terminal L-alanine residue (which is the most exposed residue in the immunogen), whereas the adjacent residues in the antigenic determinant (or hapten) occupy subsites S,, and so on (FIGURE 1) . It is seen (TABLE 3) that the inhibitory capacity of free L-alanine peptides increases with their size up to the tetrapeptide, the H . Ala, . OH is practically as good an inhibitor as H Ala, OH. The common interpretation of these results would be that the combining site can accommodate a tetrapeptide and, in terms of the subsite concept, the conclusion would be drawn that it is composed of four subsites. As will be shown further on, however, this conclusion is erroneous: a more rigorous procedure shows the presence of only three subsites.
In principle, more precise information on properties of the combining site can be derived from comparing haptens of equal size which differ from one another in one particular feature, rather than from studying the effect of elongation in a homologous series of peptides.13 This is because the stronger binding caused by elongation is not necessarily due to the addition of interac-
100 - e z 0 ' 50 z I
0.25 0.50 0.75 0.25 0.50 0.75 PEPTIDE (mM)
FIGURE 3. Hapten-inhibition of the poly-L-alanine system. The precipitin reaction between Ab-4t fraction of P-5 and poly-L-AlaRSA was inhibited by the following peptides. In A: ., AlaT(L3); A, Ala&); 0, Alaa(~5); 0, Alar(L:{D); A, A l a ( ~ 4 ~ ) ; insert W, Ala(L); 0 , Alaz(Lz); A, A l a s ( ~ ~ z ) ; 0, Ala4(DL3); 0, A l a d ~ h ) ; A , A~%(LZD). In B: ., Alaz.NHz(Lz); A, Alas.NHz(~3); A, Alaa.NHz(~a); 0, Ala .NHz(~6) ; insert 0 , Ala.NHz(L); W , Ac-Alaz(~2); 0, A~-Ala3(~3); 0, Ac-Ala(~4).

P 0
h,
TA
BL
E 3
INH
IBIT
ION
O
F THE P
REC
IPIT
IN
REA
CTIO
N OF
AN
TIB
OD
IES SP
ECIF
IC
TO
PO
LY-L
-ALA
NIN
E C
HA
INS BY
AL
AN
ME
PEW
IDES
Pept
ide
Con
cent
ratio
n (m
M) a
t 50%
Inhi
bitio
n > 3 3
Ant
ibod
y A
ntib
ody
Sour
ce
Frac
tion
Ala
n H
.L.O
H
H.L
-iD
.OH
H
. DL
-1 .O
H
H.L
-NH
2
Ac-
L.O
H
E P-
5 A
b-4t
A
lal
>8.
5 (6
%)
>8.5
(4
2%)
Z A
la2
8.5
>8.5
(2
5%)
0.32
>
3 (0%)
2 4
b
Ala
2 >
6 (4
3%)
0.26
>
1.8 (0%)
e,
>1.
8(4%
) E
Ala
, 0.
30
>0.
6 (2
0%)
>1.
8 (0%)
0.08
2 >
1.8 (0%)
Ala
, 0.
07
0.09
>
1.8
(15%
) 0.
053
Ala
, 0.
055
0.06
>
1.8
(20%
) 0.
053
Ala
3 0.
32
2.4
>3
(31%
) 0.
08
>3
(10%
) A
la.,
0.09
0.
11
>3
(31%
) 0.
07
>3
(22%
) A
h6
0.08
0.
07
>3
(30%
) 0.
07
% 0
P-4
Ab-
4t
Ah
>
6 (5
%)
>6
(42%
)
2.0
&
v1
0 3
c)
CD
P-4
Ab-
Act
A
la,
>6 (8%)
Q.
Ala
z >
6 (3
7%)
0.71
>
2.4 (0%)
Ala
, 1.
47
2.9
>1.
8 (0%)
0.41
>
2.4 (0%)
Ala
, 0.
40
0.71
>
1.8 (0%)
0.32
>1.8 (0%)
Ala
, 0.
35
0.38
>
1.8
(15%
) 0.
32
v1
Rea
ctio
n m
ixtu
res
cont
aine
d pu
rifie
d an
tibod
ies,
pol
y-L-
Ala
RSA
and
pep
tide.
A n
umbe
r of
con
cent
ratio
ns o
f pe
ptid
es w
ere
empl
oyed
to
dete
rmin
e th
e po
int
of 5
0% i
nhib
ition
. Whe
n SO
OJ, i
nhib
ition
was
not
ach
ieve
d, th
e pe
rcen
t inh
ibiti
on o
btai
ned
at th
e m
axim
al p
eptid
e co
ncen
- tr
atio
n us
ed i
s gi
ven
in p
aren
thes
es. H
.L.O
H,
free
L-a
lani
ne p
eptid
e; H
.L-,
D.O
H,
free
ala
nine
pep
tide
with
a D
res
idue
at t
he C
term
inus
; H
.DL
-I.O
H, f
ree
alan
ine
pept
ide
with
a D
resi
due
at th
e N
term
inus
; H.L
.NH
2, a
mid
e of
ala
nine
pep
tide;
Ac-
L,.O
H,
N-a
cety
l ala
nyl p
eptid
e.

Schechter : Mapping of Antibody Combining Sites 403
tion by the extra piece, but can also be attributed to the abolishment of un- favorable interactions in the smaller hapten. Thus, the finding that H . Ala, . OH is a better inhibitor, by a factor of 4, than H . Ala, . OH does not prove that the additional alanine residue interacts with the binding site, i.e., it does not prove the existence of subsite S,. It can also be due to the lack of interaction, or even to interference between the carboxyl group of H * Ala, . OH and S,. Indeed, the inhibitory capacity of H * Ala, . NH2, which is equal in size to H . Ala:, . OH but without the carboxylate ion, is better than that of H * Ala, * OH and similar to that of tetra- and penta-L-alanine (TABLE 3). Other erroneous conclusions could be drawn from the use of the free peptide series alone. For example, the contribution of the third residue is overestimated by using data obtained with free peptides, since elongation from di- to tripeptide increased the inhibitory efficiency by factors of 23 and 27. The data obtained with the cor- responding amides give factors of 3 and 4 (TABLE 3), which seems to be a better estimate for this elongation (see below).
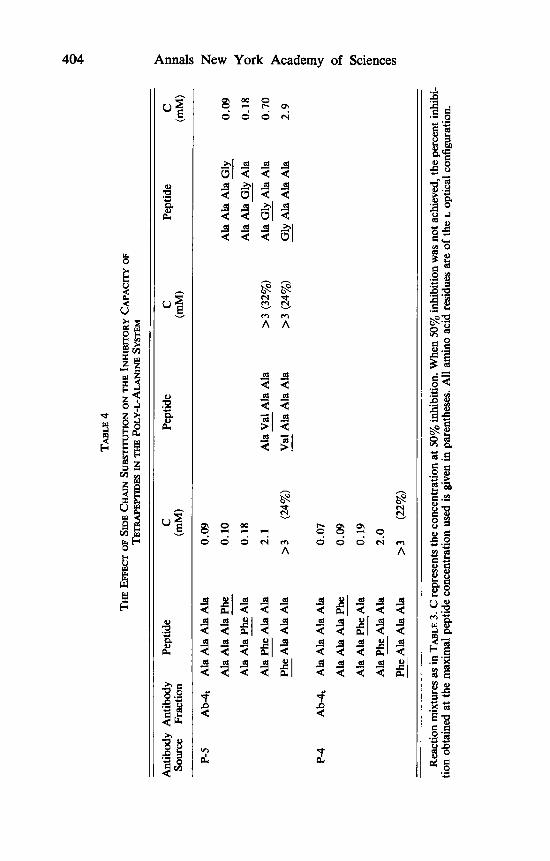
The effects of systematic variations in the structure of tetra-L-alanine are shown in TABLES 4 and 5. These involve an increase or decrease in the bulki- ness of the amino acid side chain, as well as diastereoisomeric replacements. It is seen that substitution of an L-alanine residue by an L-phenylalanine (or L-valine) residue in the N-terminal position reduced binding remarkably. This effect drops gradualy with distance from the N-terminal end, and disappears in the fourth position (counting from the N-terminal end). A similar pattern is observed by substituting a glycine residue for an L-alanine residue (TABLE 4). Replacement of an L-alanine residue by a D-alanine residue reduces binding when occurring in the first three positions, but it is not effective in the fourth position (the C-terminal end). The effect of diastereoisomeric replacement is stronger the closer it is to the N-terminal end (TABLE 5). In agreement with the above are the findings that in tripeptides the L, isomer is much better as an inhibitor than the L,D isomer, whereas the corresponding isomers of tetrapep- tides (L, and L,D) and pentapeptides (L, and L,D) have similar inhibitory potency (TABLE 3).
Data given in TABLE 3 show that in the free L-alanine peptide series the in- crease in inhibitory capacity levels off at the tetrapeptide ( H * Ala, . OH is practically as good in inhibition as H . Ala, . OH), whereas in the L-alanine amide series leveling off occurs with the tripeptide (H . Ala, . NH, is practically as good as H . Ala, . NH,). More relevant information on the binding site is derived from the use of alanine amides because these peptides bear closer struc- tural resemblance of the antigenic determinant. The poly-L-alanine chain in the antigen is composed of the charged N-terminal ammonium group and of re- peating sequences of polar amide groups and non-polar (Y and p carbons of alanine. The alanine chains coupled to the protein do not contain any carboxyl- ate group which is present in free alanine peptides (FIGURE 4). A comparison between the amides and the free peptides shows that the binding of H * Alan . NH, is almost equal to that of H . Ala,,,, . OH (TABLE 3). This means that in free peptides either the carbon portion of the C-terminal residue does not interact with the combining site, or that interference caused by the carboxylate of H . Ala,, . OH is counterbalanced by the interaction of the carbon portion of the C-terminal alanine in H . Ala,,,, . OH. The latter conclusion probably is the correct one, since Ala, (L, ) is a better inhibitor than Ala, (L,D). The finding that amidation improved the inhibitory capacity of alanine, Ala, and Ala, (but
TA
BL
E
4
TE
TR
AP
E~
ES
IN
THE P
OLY
-L-A
LAN
INE SY
STEM
TH
E EFFECT OF SIDE C
HA
IN sU
L3S
7llV
TIO
N O
N T
HE
IN
HIB
ITO
RY
CA
PAC
ITY
OF
9 E
Ant
ibod
y A
ntib
ody
Pept
ide
C
Pept
ide
C
Pept
ide
C
Sour
ce
Frac
tion
(mM
) (m
M)
(mM
) E
P-5
Ab
4
Ala
Ala
Ala
Ala
0.
09
Z CD
Ala
Ala
Ala
Phe
0.
10
Ala
Ala
Phe
Ala
0.
18
-
-
Ala
Ala
Ala
Gly
0.
09
8 A
la A
la G
ly A
la
0.18
2
_.
4
-
P?
9
Gly
Ala
Ala
Ala
2.
9 0 E
P-4
Ab
4
Ala
Ala
Ala
Ala
0.
07
r
-
Ala
Gly
Ala
Ala
0.
70
Ala
Val
Ala
Ala
A
la P
he A
la A
la
2.1
Val
Ala
Ala
Ala
Ph
e A
la A
la A
la
>3
(32%
) -
-
>3
(24%
) -
>3
(24%
) -
-
4 A
la A
la A
la P
he
0.09
Ala
Ala
Phe
Ala
0.
19
Ala
Phe
Ala
Ala
2.
0 -
-
-
Phe
Ala
Ah
Ala
>3
(2
2%)
Rea
ctio
n m
ixtu
res a
s in
TA
BL
E
3. C
repr
esen
ts th
e co
ncen
trat
ion
at 5
0% i
nhib
ition
. Whe
n 50
% i
nhib
ition
was
not
ach
ieve
d, th
e pe
rcen
t inh
ibi-
tion
obta
ined
at t
he m
axim
al p
eptid
e co
ncen
trat
ion
used
is
give
n in
par
enth
eses
. All
amin
o ac
id r
esid
ues
are
of t
he L
opt
ical
con
figur
atio
n.

Schechter : Mapping of Antibody Combining Sites 405
TABLE 5
THE EFFECX OF DIASTEREOWMERIC REPLACEMENT ON THE INHIBITORY CAPACITY OF THE TETRAALANINE PEPTIDES IN Ab4t FRACTION OF P-5
H . Ala, . OH Configuration at 50% Inhibition
Concentration (mM)
LLLL 0.09 LLLD 0.11 LLDL 1.8 LDLL > 3 (28%) DLLL > 3 (31%)
Reaction mixtures as in TABLE 3. When 50% inhibition was not achieved, the percent inhibition obtained at the maximal peptide concentration used is given in parentheses.
not of Ala, and Ala,) shows that the carboxylate ion interferes with binding up to the third residue.
To sum up, it was found that diastereoisomeric replacements, the presence of carboxylate ion and changes in bulkiness of the side chain (an increase Ala -+ Phe, or decrease Ala + Gly) are felt over three residues from the N- terminal end of an L-alanine peptide. This means that the size of the combining site is such as to accommodate three L-alanine residues, namely, it is composed of three subsites (Sl, S , and S,, FIGURE 1) . Stereospecificity is very strong in S, and S,, and somewhat less in S , (TABLE 5 ) . The structural requirements for the interaction of a ligand with S , are very stringent, less so in S,, and still less in S,. The strict requirements of S , are deduced from the findings that N- acetyl-Ala, (L,) and Ala, ( D L ~ ) (which contain uninterrupted sequences of 4 L-alanine residues) are very poor inhibitors as compared with Ala, (see TABLE 3). This means that a non-N-terminal L-alanine residue does not bind efficiently in S,. Also, the highest degree of space limitation toward L-phenylalanine and L-valine residues is exhibited by subsite S, (TABLE 4).
Since a glycine residue occupies less space than an alanine residue the in- hibition data can be used, by a first approximation, to calculate the binding energy of the methyl side chain of L-alanine ( - AZF) in the various subsites. For this purpose, the inhibitory capacity of pairs of peptides of equal size are compared, using the relation - AZF = RTln (Cpep/Cala).39 For example, the free energy of binding of the /3 methyl in S, is calculated by using the con- centrations, at 50% inhibition, of AlaGlyAla, (Cpep) and Ala, (Gala). The - A2F values thus obtained are summarized in TABLE 6. From the data given in this Table, it is also seen that despite the fact that the inhibitory potencies of H - Ala, * OH and H Ala, . OH are quite different, the same - A2F values for the binding of a methyl group in S , are obtained from the pairs GlyAla, and Ala, as well as GlyAla, and Ala,. These findings strongly support our basic assumption that the peptides are lined up on the combining site in such a way that the free N-terminal residue always occupies the same subsite, i.e., S, . In all cases examined, the binding energies of the methyl groups in the various subsites were found to be independent and thus additive. The above example shows that the interaction of the methyl group in S , is independent of the presence of a charged carboxylate in S,. In connection with this it can be

406 Annals New York Academy of Sciences + NH3 I
I
+ I
I NH3 I
I I
I I
I I
I I
I I
I I
I I
I I
I H3C-CH
I H3C-CH
I CO
I CO I I NH NH I I
H3C-CH
CO
NH
H3C-CH H3C-CH
CO CO
NH NH
H3C4H
CO co NH NH
H3C-CH
H3C-CH H3C-CH
CO co NH NH
FIGURE 4. Schematic representation of poly-r. (or D)-alanyl protein prepared by polymerization of N-carboxy-L(or D)-alanine anhydride. The attached alanine chains are of somewhat variable size and carry free a-amino groups.43 Boldface -NH stands for &-amino groups of lysine residues of the protein.
mentioned that binding of a methyl in S , is independent of the presence of /?-methyl in the fourth position (compare Ala?GlyAla and Ala,Gly, with Ala,) . The additivity is shown by the fact that in peptides in which two alanine residues were replaced by glycine residues the - A2F values calculated were equal to the sum of the - A2F values of the appropriate single replacements. For example, the change in binding energy between Gly,Ala, and Ala, was found to be 3.38 Kcal mole-'. The latter value is the same as the sum of the - AzF values determined separately for S, and S, (2.1 + 1.25 = 3.35 Kcal mole-1; see TABLE 6 for other examples).

Schechter : Mapping of Antibody Combining Sites 407
TABLE 6
VARIOUS SUBSITFS OF Ab4& FRACTION OF P-5 BINDING ENERGY OF THE METHYL SIDE CHAIN OF L-ALANINE IN THE
C &P - AzF Subsite Peptide (mM) Cnla (Kcal mole-') Explored
Ala Ala Ala Ala 0.09
Gly - Ala Ala Ala 2.9 32 2.1 S1
Ala - Gly Ala Ala 0.70 7.8 1.25 sz Ala Ala - Gly Ala 0.18 2.0 0.42 s3 Ala Ala Ala Gly - 0.09 1 .o 0 s4
_ _ Gly Gly Ala Ala 24 266 3.38 s1 + s2
Ala Ala Gly -- Gly 0.17 1.9 0.39 s3 + s4 Ala Ala Ala 0.32
Gly - Ala Ala 10 31 2.09 Sl
Ala Gly Gly 5.0 16 1.67 sz + s: --
Reaction mixtures as in TABLE 3. C represents the concentration at 50% inhibition. The free energy of binding of the methyl side chain (at 36") was calculated from the relation - A ~ F = RT In (C&,/CaI,,), using peptides of equal size. All alanine residues are of the L-optical configuration.
The finding that the methyl groups of an alanine peptide are bound in the three subsites with different energies (2.1 Kcal mole-' in S,, 1.2 Kcal mole-' in S2, and 0.4 Kcal mole-' in S,) may give an important clue to the architecture of the binding site. Since it is reasonable to assume that hydrophobic interaction is the main source of binding energy, the differences observed seem to reflect differences in the degree of contact. In other words, one may say that in S, the methyl group of the hapten fits into a depression which completely surrounds it, in S, the depression is either shallower or wider, producing less contact, whereas in S, contact is still weaker. This would also account for the fact that S , cannot contain a phenyl (or isopropyl) group, S , accommodates it with diffi- culty, and in S, the interference is rather slight. The varying degrees of stereo- specificity of the three subsites also fall into the same pattern.
A similar situation was found to exist in the binding of peptide inhibitors at the active site of papain which specifically binds aromatic residues in S2.10-21 Binding energies of various amino acid side chains in two subsites ( S , and S 2 ) were determined and correlated with preliminary x-ray diffraction data of the enzyme-inhibitor complex.21 The methyl side chain of alanine was found to be bound in S, with 0.85 Kcal mole-'. The model showed that only about half of it is in hydrophobic contact with the enzyme surface. The phenyl group of phenylalanine was bound with 0.85 Kcal mole-' in S, but with 3 Kcal mole-' in S,. The three-dimensional model showed that in S , contact is rather slight, whereas in S, hydrophobic contact is extensive. It is also of interest to mention

408 Annals New York Academy of Sciences
that in antibodies the phenyl group was found to be bound with 4 Kcal mole-1 in the anti-L(or D) -phenyl-(p-azobenzoylamino) -acetate and with 3.5 Kcal mole-' in the anti-DNP system.41 In terms of the lock-and-key theory, one may say that a given key (methyl or phenyl group) may fit one lock better than another.
Inhibition Studies of Subfractions of Anti-Poly-~-Alanyl Antibodies
Antibodies specific to poly-L-alanine chains from three groups of rabbits (P-5, P-6, and P-7) were fractionated by stepwise extraction from an immuno- adsorbant with di, tri, and tetra-L-alanine to yield respectively, Ab-2, Ab-3, and Ab-4 fractions. From the same P-5 antisera pool, another portion was adsorbed and extracted by tetra-L-alanine to yield Ab-4, fraction (the inhibition profile of this fraction is given in TABLES 3-6 and FIGURE 2). Thus, Ab-2, Ab-3, and Ab-4 fractions of P-5 are subfractions of Ab-4t.
The precipitin reactions of the various fractions were inhibited by free+ alanine peptides, the amides of L-alanine peptides, and by peptides composed of L-alanine and glycine residues. The results are summarized in TABLES 7- 10. It was found that the inhibitory efficacy of the alanine peptides increased with size up to a certain limit. In all fractions the upper limit was larger in the free alanine peptide series as compared to the alanine amide series (TABLES 7, 8) . For estimating the size of the combining site results obtained with the amides are more relevant, since these peptides bear closer structural resem- blance to the antigenic determinant (see previous section and FIGURE 4). A com- parison between the amides and the free peptides shows that in most cases the binding of H Alan NH, is almost equal to that of H . Alan+' OH. It is also seen that the interference caused by the C-terminal carboxyl ion in Ab-2 is strongly felt up to the dipeptide and to a lower degree in the tripeptide (H - Ala, NH, is 4.7-8.8 fold better than H - Ala, *OH, H Ala, - NH, is only 1.3-1.8 better than H Ala, * OH, and the inhibitory efficacy of the free and amide deriva- tives of tetra- and pentaalanine is practically identical), in Ab-3 up to the tripeptide (H * Ala, NH, is 2.4-3.8 better than H - Ala, * OH), and in Ab-4 up to the tetrapeptide (H .Ala, * NH, is 1.7 and 2.5 better than H Ala, OH). The binding energy of the methyl side chain of L-alanine in the various subsites (-AzF) was calculated as described in the previous sec- tion. Here again the -A2F values were found to be independent and there- fore additive, as seen in a detailed example given in TABLE 9. The -AzF for methyl-subsite interaction in all antibody fractions are summarized in TABLE 10. The data show that in all cases binding of the /3-methyl group is strongest in S, (1.9-2.4 Kcal mole-I), and it drops gradually towards S, and S,. There is no evident interaction in S,, even in the case of the Ab-4 fraction. The strength of interaction in the same subsite of the various fractions is different, particularly so in S, and S, (e.g., in P-6 the P-methyl group is bound to S, of the Ab-2, Ab-3, and Ab-4 with 0.68, 1.32, and 0.87 Kcal mole-', respectively).
The increment in inhibitory capacity caused by elongation of the peptide chain in the amide series leveled off at different peptide sizes (TABLE 8.) In the Ab-2 fractions the inhibitory capacity of penta, tetra, and trialanine amides are identical, the H Ala, * NH, is only slightly better than H . Ala, NH, (1.03-1.33 fold), whereas H * Ala, * NH, is much better than H * Ala NH,

TA
BL
E
7
POL
Y-L
-AL
AN
INE
C
HA
INS BY
FR
EE
L-A
LA
NIN
E
PE
~E
S
INHI
BITI
ON O
F T
HE
PRE
CIPm
N R
EAC
TIO
N
OF S
UBFR
ACTlO
N O
F A
NTI
BO
DIE
S SPEC
IFIC
TO
c( .. z z.
P-5
Ab-
2 >
7 (1
5%)
0.90
0.
20
0.14
7 0.
135
m
Ab-
3 >
6.6 (4%)
0.18
3 0.
098
0.090
0,
Pept
ide
Con
cent
ratio
n (m
M) a
t 50%
Inhi
bitio
n A
ntib
ody
Ant
ibod
y So
urce
Fr
actio
n H
. Ahl
. OH
H
. Ala
2. O
H
H .A
la,.O
H
H .A
la, .
OH
H
.Ala
S.O
H
a’ L! 2 A
b-4
>5.
9 (15%)
1.03
0.
176
0.11
7 A
b4
>8.5 (
6%)
8.5
0.
32
0.09
0.
08
P-6
Ab-
2 >1
4 (3
2%)
1.25
0.
14
Ab-
3 >1
4 (1
3%)
3.71
0.
14
Ab-
4 >1
4 (5
%)
>I4
(3
9%)
0.83
A
b-A
C >1
4 (4
%)
>14
(35%
) 14
0.11
7 0.
065
0.16
0 7.6
P-7
Ab-
2 >
7 (8
%)
0.70
5 0.
194
0.11
2 A
b-3
>4.
7 (2
0%)
0.23
0.
077
Ab-
4 >
5.9
(8%)
>0.4
7 (2
8%)
0.13
5
5.
5
0 a
r
0.11
7 0.
065
0.10
6 G
0.11
2 ki E
2.
0.07
7 3
0.07
7
Rea
ctio
n m
ixtu
res a
s in
TA
BL
E
3. W
hen 50%
inhi
bitio
n w
as n
ot a
chie
ved,
the
perc
ent i
nhib
ition
obt
aine
d at
the
max
imal
pep
tide
conc
entr
atio
n us
ed is
giv
en in
par
enth
eses
. r$ P
0
W

TA
BL
E
8
INH
IBIT
ION
O
F THE P
REC
IPIT
IN
RE
AC
~IO
N
OF
SUB
FRA
CTI
ON
S O
F A
NTI
BO
DIE
S SPEC
IFIC
TO
POLY
-L-A
LAN
INE
CH
AIN
S BY A
MID
ES
OF
L-A
LAN
INE ~E
P'I
IDE
S
E
Sour
ce
Frac
tion
H.A
lal.N
H2
H . A
la,.
NH
2 H
.Ala
, .N
H2
H .A
la,.
NH
2 H
.Ala
,.NZ
t 5
Ab-
3 4.
7 (3
2%)
0.31
2 0.
065
0.065
0.065
Ab-
4 n.
d.
n.d.
n.
d.
n.d.
n.
d.
Pept
ide
Con
cent
ratio
n (m
M) a
t 50%
Inhi
bitio
n m
Ant
ibod
y A
ntib
ody
P-5
Ab-
2 1.
88
0.12
6 0.
122
0.12
2 0.
122
4 %
Ab-
4t
>8.5
(42%
) 0.
32
0.08
0.
07
0.07
R
P-6
Ab-
2 1.
17
Ab-
3 7.
06
Ab-
4 13
.5
Ab-
Ac
14
0.14
1 0.
235
0.47
4.
1
0.10
6 0.059
0.16
7 3.
8
P-7
Ab-
2 2.
06
0.15
0.
117
Ab-
3 n.
d.
0.23
0.060
Ab-
4 >
4.7
(20%
) >0
.47
(41%
) 0.
145
0.10
6 0.059
0.09
7 4.
0
0.11
7 0.060
0.05
4
bj
0.11
5 0.059
0.10
5 -2 0
m
4.
3
E 0.
117
0.060
3
0.05
4 n
2 R
eact
ion
mix
ture
s as i
n T
mL
e 3.
Whe
n 50%
inhi
bitio
n w
as n
ot a
chie
ved,
the
perc
ent i
nhib
ition
obt
aine
d at
the
max
imal
pep
tide
conc
entr
atio
n used i
s gi
ven
in p
aren
thes
es, n
.d.:
not d
eter
min
ed.

Schechter : Mapping of Antibody Combining Sites 41 1
TABLE 9
BINDING ENERGY OF THE METHYL SIDE CHAIN OF L-ALANINE IN THE VARIOUS Sumsms OF Ab-3 FRACTION OF P-6
G P - A2F Subsite Peptide (mM) Gala (Kcal mole-1) Explored
- C
Ala Ala Ala Ala 0.065
Gly - Ala Ala Ala 2.46 37.8 2.24 SI
Ala Gly - Ala Ala 0.55 8.45 1.32 s2
Ala Ala Gly - Ala 0.13 2.0 0.43 SJ
Ala Ala Ala - Gly 0.059 0.91 -0.06 S,
Gly _ _ Gly Ala Ala 19.4 298 3.52 s , + sz Ala Ala Gly _ _ Gly 0.12 1.85 .38 s3 + s 4
Ala Ala Ala 0.14
Gly - Ala Ala 5.0 35.6 2.20 s1 Ala _- Gly Gly 1.72 12.3 1.55 sz + s3
Binding energies were calculated as described in TABLE 6.
(8.3-14.9 fold). In the Ab-3 fractions penta, tetra, and trialanine amides are identical, but H . Ala, * NH, is significantly better than H * Ala, . NH, (3.8-4.8 fold). In the Ab-4 fractions penta and tetraalanine amides are identical, and H . Ala, . NH, is somewhat better than H Ala, . NH, (1.7 and
TABLE 10
BINDING ENERGY OF THE METHYL SIDE CHAIN OF L-ALANINE I N THE VARIOUS SUBSITES OF ANTIBODY FRACTIONS SPECIFIC TO POLY-L-ALANINE CHAINS
-A2F Values (Kcal mole-I) Antibody Antibody Source Fraction SI sz S , S4
P-5 Ab-2 1.86 0.74 0.06 0 Ab-3 2.13 1.34 0.51 -0.20 Ab-4t 2.10 1.25 0.42 0
P-6 Ab-2 2.09 0.68 0.10 0 Ab-3 2.22 1.32 0.43 -0.06 Ab-4 2.38 0.87 0.35 0
Binding energies were calculated as described in TABLE 6. The -AZF values for Ab-4t of P-5 are from TABLE 6.

412
2.7 fold). Information on S, can be obtained from the inhibition data of alanine amide which reacts with this subsite. The alanine amide inhibits the Ab-2 fraction better than Ab-3, and Ab-3 better than Ab-4 (TABLE 8). This indicates that binding of H . Ala . NH, is strongest in S, of Ab-2, less so in S, of Ab-3, and even less in S, of Ab-4. Equilibrium dialysis experiments with the closely related anti-pOly-D-ahine system corroborate this suggestion (see next section),
To sum up, one may say that the binding sites of Ab-2, Ab-3, and Ab-4 fractions are composed essentially of three subsites, but the properties of the subsites differ considerably in the various antibody fractions. In Ab-2, most of the binding energy is brought up by interaction with S, and S2, whereas S, plays a minor role (S, shows a low sensitivity to the presence of the carboxyl- ate ion and, upon combination with H * Ala, NH, interaction with the third residue, is very low). Subsite S, in Ab-3 and Ab-4 exhibit significant interaction with the third residue of the ligand. The Ab-4 fraction contains an additional subsite, S,, but the contribution of S, to the total binding energy is quite small. The finding that there is no gain in free energy of binding by the methyl side chain of alanine in S, (TABLE 10) indicates that when an alanine residue occupies S, only the backbone of the residue (NH-CH-CO) interacts with the antibody molecule.
Annals New York Academy of Sciences
Equilibrium Dialysis Studies of Anti-Poly-D-A lanyl Antibodies
Antibodies specific to poly-D-alanine chains were obtained from rabbits immunized with poly-D-AlaHSA. Antisera collected from six animals (P-8) were pooled and adsorbed on poly-D-AlaRSA-cellulose conjugate. In one case, the adsorbed antibodies were fractionated by stepwise extraction with di-w alanine, tri-D-alanine, tetra-D-alanine, and acetic acid to yield Ab-2, Ab-3, Ab-4, and Ab-Ac fractions, respectively. In the other case, the adsorbed antibodies were extracted by tetra-D-alanine (without prior extraction with smaller pep- tides) and acetic acid to yield the Ab-4, and Ab-Ac, fractions. Antibodies ob- tained from an individual rabbit (1-1) were fractionated as described above (see TABLE 2). The equilibrium dialysis method was applied to determine interactions of the various purified antibody fractions with series of free and amides of Dalanine peptides of increasing size. For this purpose one labeled tetrapeptide (14C-~Ala,-Gly) served as a reference hapten, and the binding of cold peptides was determined by measuring their capacity to inhibit the binding of 14C-~Ala,-Gly.33
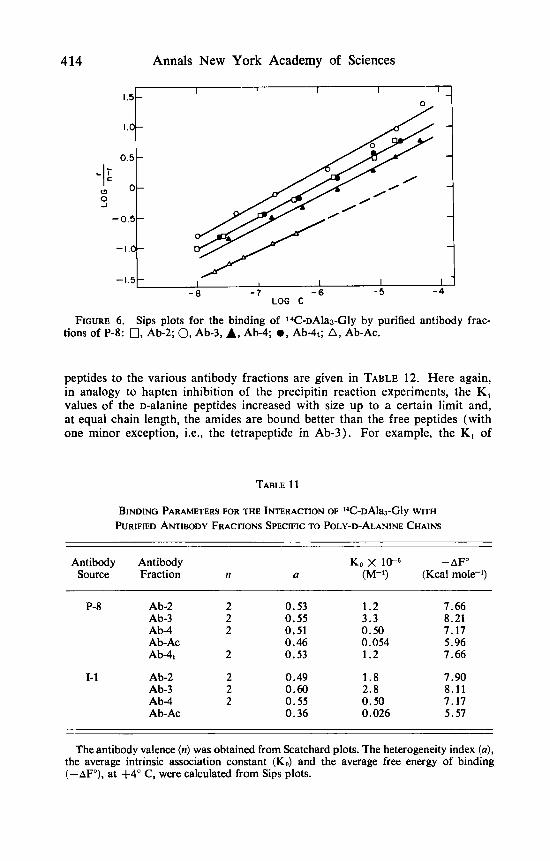
Representative binding curves of l4C-~A1a,-Gly are shown in FIGURES 5 and 6. By extrapolation of the Scatchard plot to the abscissa, it was found that all antibody fractions prepared by extraction with peptide solutions had a valence (n) of 2. The Ab-Ac fractions, which have quite poor binding, were saturated with the hapten up to r = 0.8 in 1-1 and r = 0.3 in P-8 ( r is the average number of peptide molecules bound per one antibody molecule). Values of the average intrinsic association constant (K.,), the average free energy of binding ( - AFO), and the heterogeneity index ( a ) for the inter- action of 14C-~Ala,-Gly with the various antibody fractions are given in TABLE 11. All antibody fractions exhibited a high degree of heterogeneity ( a values in the range of 0.36-0.6), irrespective of whether they were prepared from sera pooled from six animals (P-8) or from an individual rabbit (1-1). In the

Schechter: Mapping of Antibody Combining Sites 413
5
4
3 0 I
0 1
0 \ L
2
I
FIGURE 5.
I I I I
0.4 0.8 I .2 I .6 2.0 r
Scatchard plots for the binding of 14C-~Ala&ly by purified antibody fractions of P-8: 0, Ab-2; 0, Ab-3; A, Ab-4; 0, Ab-4t; A, Ab-Ac.
case of P-8, Ab-2, Ab-3, and Ab-4 represent subfractions of Ab-4, (TABLE 2) . Since all four fractions had similar a values (0.51 to 0.55) , it may be concluded that stepwise extraction with peptides did not result in a more restricted heterogeneity, The KO values of the corresponding fractions of P-8 and 1-1 are quite similar. In both cases the highest average association constant was observed in Ab-3 (3.3 and 2.8 x lo6 M-l), it was reduced by a factor of about 2 in Ab-2, by a factor of about 6 in Ab-4, and a marked reduction in affinity (by two orders of magnitude) was found in the Ab-Ac fraction. An inspection of the K, values of Ab-4, and of its subfractions (Ab-2, Ab-3, and Ab-4) shows that some degree of fractionation with respect to affinity towards 14C-~Ala,-Gly was achieved.
The K, (which closely represents the average association constant, see Materials and Methods section) and - AFO values for binding of Dalanine
414 Annals New York Academy of Sciences
1.5-
1.0-
0.5
-1: a 0 J
-0.5-
-1.0-
-1.5-
I I I I I -
-
0-
- 8 - 7 - 6 - 5 - 4
FIGURE 6. Sips plots for the binding of 14C-~Ala3-Gly by purified antibody frac- tions of P-8: 0, Ab-2; 0, Ab-3, A, Ab-4; 0 , Ab-4t; A, Ab-Ac.
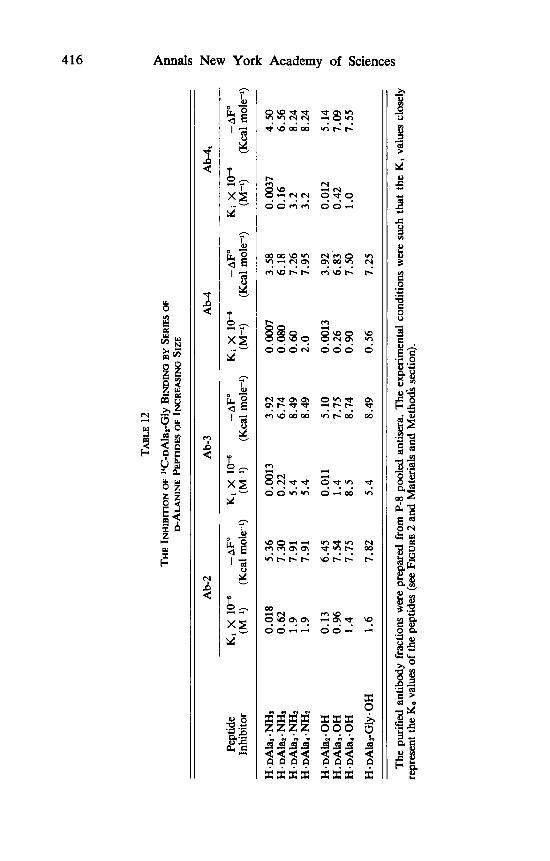
peptides to the various antibody fractions are given in TABLE 12. Here again, in analogy to hapten inhibition of the precipitin reaction experiments, the K i values of the D-alanine peptides increased with size up to a certain limit and, a t equal chain length, the amides are bound better than the free peptides (with one minor exception, i.e., the tetrapeptide in Ab-3). For example, the K i of
TABLE 11
BINDING PARAMETERS FOR THE INTERACTION OF 14C-~Alal-Gly WITH
PURIFIED ANTIBODY FRACTIONS SPECIFIC TO POLY-D-ALANINE CHAINS
Antibody Antibody KO X 10-' -AF" Source Fraction I 1 U (M-'1 (Kcal mole-')
P-8 Ab-2 2 Ab-3 2 Ab-4 2 Ab-Ac Ab-4t 2
1-1 Ab-2 2 Ab-3 2 Ab-4 2 Ab-Ac
0.53 0.55 0.51 0.46 0.53
0.49 0.60 0.55 0.36
1.2 3.3 0.50 0.054 1.2
1.8 2.8 0.50 0.026
7.66 8.21 7.17 5.96 7.66
7.90 8.11 7.17 5.57
The antibody valence (n) was obtained from Scatchard plots. The heterogeneity index (a), the average intrinsic association constant (KO) and the average free. energy of binding (-AF"), at +4" C, were calculated from sips plots.

Schechter : Mapping of Antibody Combining Sites 415
H . Ala, . NH, is larger than the K , of H * Ala, * OH by factors of 4.8, 20 and 61, respectively, in Ab-2, Ab-3 and Ab-4. These ratios reflect different degrees of sensitivity to the presence of carboxylate ion in S, of the three subfractions. Results obtained with the alanine amides will be discussed mainly because these peptides are very closely related to the structure of the antigenic determinant in the immunogen (see previous two sections). The upper value of - AFO ob- tained probably represents all possible interactions, since the increment in - AFO caused by elongation of the peptide chain leveled off. The maximal binding energy found for Ab-2, Ab-3, and Ab-4 was quite similar and cor- responded to 7.91, 8.49, and 7.85 Kcal mole-', respectively. There are, how- ever, big differences (absolute and relative) between the various antibody frac- tions when interactions in individual subsites are examined. Thus, subsite S, (which reacts with the N-terminal residue) in Ab-2 is the most efficient one: it binds H . DAla . NH, with 1.44 and 1.78 Kcal mole-' more than S, in Ab-3 and Ab-4, respectively. In terms of K , values, S, in Ab-2 is 16- and 24-fold better than S, in Ab-3 and Ab-4. In a first approximation, the binding energy of a D-alanine residue in all subsites ( - A2F) can be calculated by subtracting the - AFO values of appropriate pairs of peptides (e.g., the - A 2 F for S2 is given by the difference between the - AFO values of H DAla, . NH, and H . DAla. NH,). The data given in TABLE 13 show that the strongest inter- action exhibited by S, occurs in Ab-3 ( - A 2 F for binding a D-alanine residue by S, in Ab-2, Ab-3, and Ab-4 are 1.94, 2.82, and 2.60 Kcal mole-', respec- tively). Subsite S, is also strongest in Ab-3, whereas interaction in S, is ob- served only in Ab-4 fraction. The contribution ( in percent) of each subsite to the total binding energy can be calculated from the relation ( - A Z F / - AFO) x 100, where - A2F correspond to values given in TABLE 13, and - AFO is the value for binding H . DAla, . NH, (TABLE 12). The results are that S,, S,, S:, and S, contribute in Ab-2, 67.8, 24.5, 7.7, and 0%; respectively; in Ab-3 46.2, 33.2, 20.6, and 0%; in Ab-4 45., 32.8, 13.6, and 8.7% of the total binding energy.
The above results lead to the conclusion that the three antibody fractions are essentially composed of three subsites but they differ considerably with re- spect to the strength of interaction in individual subsites. In the Ab-4 fraction there is an additional subsite, S,, which contributes very little (8.7%) to the total binding energy.
Despite the fact that Ab-4 has a larger combining site than Ab-2, the pep- tide H .DAla, .NH, is equally bound to both fractions (K, , 2.0 and 1.9 x 106 M-I). Somewhat stronger binding of this peptide is observed in Ab-3 ( K , , 5.4 x lo6 M-l). Thus, in the poly-D-alanine system there is no in- crease in binding with larger combining sites. Similar results were obtained by Moreno and Kabat 4 2 who fractionated human antibodies to blood group A substance by stepwise extraction with monosaccharide (GalNAc) and penta- saccharide (AR,,0.52). They found that the KO values of the AR,0.52 eluates were higher (1.2-10-fold) than the K, values of the GalNAc eluates.
Some degree of fractionation of the molecules compromising the Ab-4, fraction was achieved by stepwise extraction with di, tri, and tetra-D-alanine peptides. This is apparent from the finding that the K , values of each peptide (TABLE 12) and the - A z F values for interaction in individual subsites (TABLE 13) in Ab-4, correspond to intermediate values of the K , and - A * F in Ab-2, Ab-3, and Ab-4.
TABLE
12
THE INHIBITION O
F 1
4hN
ai-G
ly BI
NDING B
Y S
ERIFS
OF
D-A
LAN
INE PE
FllD
ES OF
INCR
EAsM
G SIZE
E A
M
Ab-
4 5
Ab-
2 A
b-3
s Pe
ptid
e K
i X
lo-
' -A
Fo
Ki
X 1
o-B
-AFo
K
i X
1o-
B -A
F"
Ki
X 1
o-B
-AF
" In
hibi
tor
(M-1
) (K
cal m
ole-
') (M
-1)
(Kca
l mol
ecl)
(M-I
) (K
cal m
ole-
-1)
(Kca
l mol
e-')
2 H
. ~A
la,.
NH
2 0.
018
5.36
0.
0013
3.
92
0.00
07
3.58
0.
0037
4.
50
H . D
Ala
2. N
Ht
0.62
7.
30
0.22
6.
74
0.08
0 6.
18
0.16
6.
56
H. D
Ala
3. N
Ht
1.9
7.91
5.
4 8.
49
0.60
7.
26
3.2
8.24
H
. ~A
la,.
NH
2 1.
9 7.
91
5.4
8.49
2.
0 7.
95
3.2
H . ~
Ala
~.
OH
0.
13
6.45
0.
011
5.10
0.
0013
3.
92
0.01
2 5.
14
H . ~
Ala
,.0
H
1.4
7.75
8.
5 8.
74
0.90
7.
50
1 .o
7.55
Ha ~
Ala
,Gly
.0H
1.
6 7.
82
5.4
8.49
0.
56
7.25
% 0
8.24
r g Y
H. ~
Ala
,. OH
0.
96
7.54
1.
4 7.
75
0.26
6.
83.
0.42
7.
09
SL R 2
The
pur
ified
ant
ibod
y fr
actio
ns w
ere
prep
ared
fro
m P
-8 p
oole
d an
tiser
a. T
he e
xper
imen
tal c
ondi
tions
wer
e su
ch t
hat
the
Ki v
alue
s cl
osel
y re
pres
ent t
he K
O va
lues
of
the
pept
ides
(see F
IGU
RE
2
and
Mat
eria
ls a
nd M
eth
d se
ctio
n).

Schechter : Mapping of Antibody Combining Sites 417
TABLE 13
BINDING ENERGY OF A D-ALANINE RFSDUE IN THE VARIOUS SUBSITES OF ANTIBODY FRACTIONS SPECIFIC TO POLY-D-ALANINE CHAINS
-A2F Values (Kcal mole@) Antibody Antibody
Source Fraction S , sz S3 S,
P-8 Ab-2 5.36 1.94 0.61 0 Ab-3 3.92 2.82 1.75 0 Ab-4 3.58 2.60 1.08 0.69 Ab-4t 4.50 2.06 1.68 0
The - A2F values for D-alanine-subsite interaction may reflect the extent of contact between the two interacting species. This is based on the analogy to papain where binding energies of amino acid side chains were correlated with the x-ray model of the enzyme-inhibitor complex (see first section and Refer- ence 21). The data given in TABLE 13 thus indicate that in the poly-D-alanine system the largest contact exists in S, and it drops gradually towards S,. The same geometrical relationships between hapten and binding site were also sug- gested in the poly-L-alanine system on the basis of the - A2F values for binding the P-methyl group of alanine (TABLES 6, 9, 10) and from the extent of space limitation and stereospecificity of the various subsites (TABLES 4, 5 ) .
It has been suggested that antibodies of high affinity should be capable of displaying a wider range of cross reactions than antibodies of low affinity." The above proposal cannot be taken as a generalization since our data contra- dict it. Subsites with high affinity (S,) exhibit a high degree of discrimination with respect to structural features of the ligand, whereas subsites with low affinity (S,) can accommodate unrelated ligands much more easily (TABLES
The antibody-hapten complex is a system of two molecules, a large and a small one which, by virtue of a special geometric relationship show a strong affinity for each other. The mapping procedure described here yields informa- tion on the number and spatial orientation of the subsites compromising the binding site, quantitative data on the free energies of binding in the various sub- sites, and one can easily determine the enthalpies and entropies of binding as well. It is also possible to break down the measured thermodynamic quantities into the sum of individual interactions of groups (e.g., the methyl side chain of alanine) . The state of x-ray crystallography today and the research in homo- geneous antibodies, obtained either from myelomas or as products of the normal immune response, make is possible that the three-dimensional structure of antibody-hapten complex will be resolved in the near future. Then one can attempt to explain the source of binding energy directly from the molecular model of the antibody-hapten complex. This will be done by correlating thermodynamic quantities with the extent of complementarity (in terms of geometry and in terms of the nature of the interacting groups) in the model. Correct interpretation of this model may lead to an understanding, on a molecular level, of biological recognition in general.
3-6).

418 Annals New York Academy of Sciences
1.
2. 3.
4. 5.
6.
7. 8. 9.
10. 11.
12. 13. 14. 15. 16. 17. 18. 19. 20. 21.
22.
23. 24. 25.
26. 27. 28. 29. 30. 31.
32. 33. 34. 35. 36. 37. 38. 39.
40. 41. 42. 43. 44.
REFERENCES
KABAT, E. A. 1968. Structural Concepts of Immunology and Immunochem-
KABAT, E. A. 1960. J. Immunol. 84: 82. SAGE, H. J., G. F. DEUTSCH, G. D. FASMAN & L. LEVINE.
ARNON, R., M. SELA, A. YARON & H. A. SOBER. SCHECHTER, I., B. SCHECHTER & M. SELA.
VAN VUNAKIS, H., J. KAPLAN, H. I. LEHRER & L. LEVINE.
GOODMAN, J. W. 1969. Immunochemistry 6: 139. HAIMOVICH, J., I. SCHECHTER & M. SELA. SCHECHTER, B., I. SCHECHTER & M. SELA. MURPHY, P. D. & H. J. SAGE. SCHECHTER, I., E. CLERICH & E. ZAZEPITZKI. 1971. European J . Biochem.
SCHECHTER, I. 1970. Israel J. Med. Sci. 6: 441. SCHECHTER, I. 1970. Nature 228: 639. SCHECHTER, I. 1971. Israel J. Med. Sci. 7: 614. LICHT, A. & I. SCHECHTER. PHILLIPS, D. C. 1966. Scient. Am. 215: 78. SHARON, N. & D. CHIPMAN. 1969. Science 165: 454. SCHECHTER, I. & A. BERGER. SCHECHTER, I. & A. BERGER. BERGER, A. & I. SCHECHTER. BERGER, A., I. SCHECHTER, H. BENDERLI & N. KURN.
ABRAMOWITZ, N., I. SCHECHTER & A. BERGER.
SCHECHTER, I. 1970. European J. Biochem. 14: 516. CUATRECASAS, P., M. WILCHEK & C. B. ANFINSEN. MORIHARA, K., T. OKA & H. TSUZUKI.
THOMA, J. A,, C. BROTHERS & J. SPRADLIN. ATLAS, D., S. LEVIT, I. SCHECHTER & A. BERGER. SCHECHTER, B., I. SCHECHTER & M. SELA. CLERICI, E., I. SCHECHTER & M. SELA. SCHECHTER, I. & A. BERGER. 1966. Biochemistry 5: 3362. ANDERSON, G. W., J. E. ZIMMERMAN & F. M. CALLAHAN.
ROBBINS, J. B., J. HAIMOVICH & M. SELA. KARUSH, F. 1956. J. Am. Chem. Soc. 78: 5519. SCATCHARD, G. 1949. Ann. N. Y. Acad. Sci. 51: 660. SIPS, R. 1948. J. Chem. Phys. 10: 490. KARUSH, F. 1962. Advan. Immunol. 2: 1. LICHT, A. & I. SCHECHTER. In preparation. SELA, M. This monograph. PRESSMAN, D. & A. L. GROSSBERG.
Specificity. Benjamin. New York. KARUSH, F. 1956. J. Am. Chem. SOC. 78: 5519. EISEN, H. N. & S. W. SISKIND. MORENO, C. & E. A. KABAT. KATCHALSKI, E. & M. SELA. ANFINSEN, C. B., M. SELA & J. P. COOKE.
istry. Holt, Rinehart and Winston. New York.
1964. Immuno-
1965. Biochemistry 4: 948. 1966. Biochim. Biophys. Acta 127:
1966. Immuno-
chemistry 1: 133.
438.
chemistry 3: 393.
1969. 1970.
1970. J. Immunol. 105: 460.
European J. Biochem. 7: 537. J. Biol. Chem. 245: 1438.
18: 561.
1971. Israel J. Med. Sci. 7: 616.
1967. Biochern. Biophys. Res. Commun. 27: 157. 1968. Biochem. Biophys. Res. Commun. 32: 898. 1970. Phil. Trans. Roy. SOC. Lond. (B) 257: 249.
1971. In Proc. Tenth
1967. Biochem. Biophys. Res. European Symp. on Peptides : 290-309. North-Holland. Amsterdam.
Commun. 29: 862.
1968. Science 162: 1491. 1969. Arch. Biochem. Biophys. 135:
1970. Biochemistry 9: 1768.
1970. Immunochemistry 7: 587.
311.
1970. FEBS Lett. 11: 281.
1970. Immunology 19: 267.
1964. J. Am. Chem. SOC. 86: 1839.
1967. Immunochemistry 4: 11.
1968. The Structural Basis of Antibody
1964. Biochemistry 3: 996. 1969. J. Exp. Med. 129: 871. 1958. Advan. Protein Chem. 13: 243.
1962. J. Biol. Chem. 237: 1825.

Schechter : Mapping of Antibody Combining Sites 419
DISCUSSION
DR. KABAT: When you elute your antibody fractions how much antibody might still be absorbed to the original column or antigen? In other words could you have some antibodies with a somewhat larger site? You really did have a fourth sub-site in some of these fractions evidentally.
DR. SCHECHTER: When you elute with a peptide you could elute up to 60 to 70% of the antibody. The 30% which are left I believe that they don’t have a larger combining site, actually you could elute it with acetic acid and I showed this fraction, and when you analyze this antibody you find two interest- ing things. First of all they probably have only two subsites rather than 4 or 5 as one would expect. Secondly, they bind peptides very poorly, by two orders of magnitude less than antibodies eluted by peptides. We have reasons to believe that these are antibodies which are directed to both the alanyl determinant and the protein carrier. So I really believe that the maximum number of subsites is four and, as you saw, subsite S, contributes very little.
DR. KABAT: There is really a striking parallelism in a contribution dropping off as you go up the sugar chain. Of course in each case in the sugar we could show that, since the reducing group is a mixture of alpha and beta, when you make the proper structure alpha as in dextran, you increase the binding energy even if you only put a methyl group on instead of a sugar.