manipulation, rearing and storage …ufdcimages.uflib.ufl.edu/uf/e0/04/54/82/00001/chen_x.pdf ·...
TRANSCRIPT

1
MANIPULATION, REARING AND STORAGE OFTAMARIXIA RADIATA (HYMENOPTERA: EULOPHIDAE) PARASITOID OF DIAPHORINA CITRI
(HEMIPTERA: PSYLLIDAE)
By
XULIN CHEN
A THESIS PRESENTED TO THE GRADUATE SCHOOL
OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
UNIVERSITY OF FLORIDA
2013

2
© 2013 Xulin Chen

3
To my parents

4
ACKNOWLEDGMENTS
Funding for this research was provided by Dr. Phil Stansly from Citrus Research
and Development Foundation.
I thank my major professor Dr. Phil Stansly for his patient guidance, support and
pertinent suggestions. I also thank my other two committee members, Dr. Howard Frank
and Dr. Eric Rohrig, for their continuous support and advice. I thank Dr. Jawwad
Qureshi, for providing suggestions in my research.

5
TABLE OF CONTENTS page
ACKNOWLEDGMENTS .................................................................................................. 4
LIST OF TABLES ............................................................................................................ 8
LIST OF FIGURES .......................................................................................................... 9
ABSTRACT ................................................................................................................... 10
CHAPTER
1 LITERATURE REVIEW .......................................................................................... 12
Huanglongbing ........................................................................................................ 12 Distribution ....................................................................................................... 12
Symptoms ........................................................................................................ 12 Pathogen .......................................................................................................... 13
Diaphorina Citri ....................................................................................................... 13
Distribution ....................................................................................................... 14 Transmission Mechanism between Pathogen and Vector ................................ 14
Host Plants ....................................................................................................... 15 Biology of D. Citri .............................................................................................. 16
Life cycle .................................................................................................... 16
Seasonal history ........................................................................................ 18 Dispersion of D. citri ................................................................................... 19
Damage by D.citri ...................................................................................... 19
Influence of temperature and humidity ....................................................... 20
Influence of light ......................................................................................... 21 Natural Enemies ............................................................................................... 21
Predators ................................................................................................... 21
Parasitoids ................................................................................................. 22 Tamarixia Radiata ................................................................................................... 22
Distribution ....................................................................................................... 22 Taxonomy and Identification ............................................................................. 23 Host Specificity ................................................................................................. 23 Biology of T. Radiata ........................................................................................ 24
Life cycle .................................................................................................... 24 Mating ........................................................................................................ 25
Oviposition ................................................................................................. 25
Host preference ......................................................................................... 26 Host-feeding ............................................................................................... 26 Sex ratio ..................................................................................................... 27 Reproduction .............................................................................................. 27 Superparasitism ......................................................................................... 28

6
Host density effects .................................................................................... 28
Influence of food ........................................................................................ 29 Development time ...................................................................................... 29
Temperature and humidity effects .............................................................. 30
2 CARBON DIOXIDE ANESTHESIA OF TAMARIXIA RADIATA (WATERSTON) (HYMENOPTERA: EULOPHIDAE) PARASITOID OF DIAPHORINA CITRI (HEMIPTERA: PSYLLIDAE) ................................................................................... 33
Introduction ............................................................................................................. 33
Materials and Methods............................................................................................ 34 Colonies ........................................................................................................... 34 Gas Chamber ................................................................................................... 34 Recovery Time ................................................................................................. 35
Survival Rate .................................................................................................... 35 Percent Parasitism ........................................................................................... 36
Results .................................................................................................................... 36 Discussion .............................................................................................................. 37
3 FUNCTIONAL RESPONSE OF TAMARIXIA RADIATA (HYMENOPTERA: EULOPHIDAE) TO DENSITIES OF ITS HOST, DIAPHORINA CITRI (HEMIPTERA: PSYLLIDAE) ................................................................................... 40
Introduction ............................................................................................................. 40 Materials and Methods............................................................................................ 42
Colonies ........................................................................................................... 42 Arenas .............................................................................................................. 43
Statistical Analysis .................................................................................................. 43 Results .................................................................................................................... 44
Fecundity, Percent Parasitism and Percent Superparasitism ........................... 44
Functional Response ........................................................................................ 44 Discussion .............................................................................................................. 45
4 THE INFLUENCE OF DIET ON EGG FORMATION IN TAMARIXIA RADIATA (HYMENOPTERA: EULOPHIDAE), A PARASITOID OF DIAPHORINA CITRI (HEMIPTERA: PSYLLIDAE) ................................................................................... 50
Introduction ............................................................................................................. 50 Materials and Methods............................................................................................ 51
Colonies ........................................................................................................... 51
Diets ................................................................................................................. 52
Egg Load .......................................................................................................... 53 Statistical Analysis .................................................................................................. 53 Results .................................................................................................................... 54 Discussion .............................................................................................................. 54
5 DISCUSSION AND CONCLUSIONS ...................................................................... 61

7
LIST OF REFERENCES ............................................................................................... 64
BIOGRAPHICAL SKETCH ............................................................................................ 73

8
LIST OF TABLES
Table page 1-1 Demographic parameters T (generation time) per day, R0 (net reproductive
rate) nymphs per female, and r (intrinsic rate of increase) per day. .................... 32
3-1 Number of parasitized hosts (Mean ± SEM) at different host densities .............. 47
3-2 Percent parasitism (Mean ± SEM) at different host densities ............................. 47
3-3 Percent superparasitism (Mean ± SEM) in different host densities, mean number ............................................................................................................... 47
4-1 Mean ± SEM number of eggs by treatment from dissections after 5, 10, 15,
and 20 days at 17 C. .......................................................................................... 56

9
LIST OF FIGURES
Figure page 2-1 Carbon dioxide gas chamber .............................................................................. 38
2-2 Tamarixia radiata female and male recovery time frequency (%) distribution .... 38
2-3 Mean (SEM) survivorship of CO2 treated and untreated Tamarixia radiata adults. ................................................................................................................. 39
3-1 Number of parasitized host at different host densities, error bar stands for SEM .................................................................................................................... 48
3-2 Percent parasitism at different host densities, error bar stands for SEM ............ 48
3-3 Percent superparasitism in different host densities ............................................ 49
3-4 Number of D. citri parasitized by T. radiata and the functional response curve (Type II) .............................................................................................................. 49
4-1 Newly emerged (unfed) T. radiata female digestive system ............................... 57
4-2 Newly emerged (unfed) T. radiata female .......................................................... 57
4-3 Paired T. radiata ovaries after feeding on honey for 20 days (egg resorption) ... 58
4-4 T. radiata ovary after feeding on Nu-Lure+honey for 10 days ............................ 58
4-5 Paired T. radiata ovaries after feeding on nymphs for 5 days ............................ 59
4-6 Paired T. radiata ovaries after feeding on honey +nymphs for 10 days .............. 59
4-7 Paired T. radiata ovaries after feeding on Nu-Lure+ nymphs for 10 days........... 60
4-8 Paired T. radiata ovaries after feeding on Nu-Lure+honey+nymphs for 5 days .. 60

10
Abstract of Thesis Presented to the Graduate School of the University of Florida in Partial Fulfillment of the
Requirements for the Master of Science
MANIPULATION, REARING AND STORAGE OF TAMARIXIA RADIATA (HYMENOPTERA: EULOPHIDAE) PARASITOID OF DIAPHORINA CITRI
(HEMIPTERA: PSYLLIDAE)
By
Xulin Chen
May 2013
Chair: Phil. Stansly Major: Entomology and Nematology
Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae), is an arrhenotokous
ectoparasite of the Asian citrus psyllid (ACP) Diaphorina citri (Kuwayama) (Hemiptera:
Psyllidae), vector of citrus greening disease or huanglongbing (HLB). Tamarixia radiata
is being tested as an augmentive biological control agent, since the number present in
the field is low following winter when few hosts are available. Any program aimed at
augmentation of a natural enemy would require an efficient system of mass production.
Key components would include methods of manipulating, rearing and storing large
populations of insects. This research investigated the use of carbon dioxide (CO2) to
anesthetize T. radiata, determined optimal host densities for rearing the parasitoid, and
evaluated diets to maintain fecundity during storage periods.
Tamarixia radiata adults held in an atmosphere of 100% CO2 for 5 min were
immobilized for about 4 min. However, survivorship and fecundity were reduced
significantly, although sex ratio of progeny from treated adults was not affected.
Consequently, lighter doses of CO2 or other methods of anesthesia are needed.

11
One pair of three-day-old T. radiata was released in a centrifuge tube for five
days with different 4th instar D. citri nymphs densities: 10, 20, 30, 40, 50, and 60. At a
density 40, 4th instar nymphs per female fecundity, incidence of parasitism and
superparasitism were all optimal. The pattern of parasitism for the first five days
conformed to a Type II functional response. Estimated searching efficiency was 0.442 ±
0.036 per day and estimated handing time was 0.045 ± 0.008 days. Therefore, 40 hosts
per female T. radiata should be a target for mass rearing.
Females feeding on eight different diets were dissected every 5, 10, 15 and 20
days to check the number of eggs in ovaries. The result showed that T. radiata formed
more eggs feeding on mixed diets (Nu-Lure+ honey+ nymphs or Nu-Lure+ nymphs)
compared to nymphs alone. Thus, it is recommended that protein and carbohydrate
supplements along with host nymphs be provided to T. radiata before release in the
field.

12
CHAPTER 1 LITERATURE REVIEW
This chapter reviews (1) distribution, symptoms and pathogen of Huanglongbing,
(2) biology, distribution of Diaphorina citri Kuyawama, (Hemiptera: Psyllidae), vector of
Huanglongbing, the transmission mechanism between pathogen and vector, and natural
enemies of D. citri, (3), biology of Tamarixia radiata (Waterston) (Hymenoptera:
Eulophidae), a major biological control agent for D. citri.
Huanglongbing
Distribution
Huanglongbing (HLB) is one of the most destructive diseases of citrus in the
world (Halbert and Manjunath 2004, Teixeira et al. 2005, Bové 2006, Wang et al. 2006,
Batool et al. 2007, Manjunath et al. 2008). It has been reported from Asia: eastern
Japan, southern China, Indian subcontinent and Pakistan, and also in the Arabian
peninsula for some time. An African form is found in eastern, central, and southern
Africa (Gottwald 2007). HLB was first discovered in the western hemisphere in Brazil in
July 2004 (Teixeira et al. 2005). The first discovery of HLB in North America occurred in
south Florida in August 2005 (Halbert 2005). By December 2007, it had spread through
30 counties which were all located south of Marion County (Hall 2008a).
Symptoms
Huanglongbing means "yellow dragon disease" or “yellow shoot disease” in
Chinese because of the yellow shoots which are early symptoms of the disease. In
addition to yellow shoots, typical symptoms also include blotchy (asymmetric) mottling,
chlorosis resembling zinc deficiency which is followed by leaf drop and twig dieback
(Halbert 2004). Symptomatic fruit are small, lopsided with small, dark aborted seed and

13
discolored vascular bundles in the fruit axis. The stylar end of symptomatic fruit remains
green even as they mature (Gottwald 2007). Roistacher (1996) reported that HLB
damaged at least 10-15% of trees each year in Thailand, which could lead many citrus
areas to go out of business. Infected citrus trees do not die immediately but decline
within 8-9 years (Roistacher 1996).
Pathogen
The causative agent is a vector-borne α-proteobacterium (Jagoueix et al.1994)
which has not yet been isolated in pure culture (Garnier and Bové 1993). Three etiologic
agents of HLB have been implicated based on their 16S rRNA sequence: Candidatus
Liberibacter asiaticus (Asia, North America, and Brazil), Ca. Liberibacter americanus
(Brazil), and Ca. Liberibacter africanus (Africa) (Garnier et al. 1984, Jagoueix et al.
1996, Sagaram et al. 2009). Of the three agents, only Candidatus Liberibacter asiaticus
(Las) was discovered in Florida in August 2005 (Halbert 2005), which led to an
estimated 1.6% of orange trees in Florida being infected by 2008 (Morris et al. 2009a,b).
Candidatus Liberibacter asiaticus is transmitted by psyllid vectors, and can also be
transmitted by grafting, dodder, and possibly by seed, but not by contamination of
personnel and tools or by wind and rain (Halbert 2010).
Diaphorina Citri
To date, two vectors have been reported for HLB disease in the world, Asian
citrus psyllid, Diaphorina citri (Kuwayama) (Hemiptera: Sternorrhyncha: Psyllidae), and
African psyllid, Trioza erytreae (del Guercio) (Hemiptera: Sternorrhyncha: Triozidae). D.
citri is responsible for transmitting Ca. L asiaticus in North America, Brazil and Asia, and
Ca. L americanus in Brazil, while T. erytreae transmits Ca. L africanus in the Middle
East, Reunion and Africa (Halbert and Manjunath 2004). Of the two species, D. citri is

14
more heat tolerant, while T. erytreae is sensitive to high temperatures (Massonie et al.
1976). D. citri was first detected in Florida in 1998 (Halbert 1998) and later become
established throughout the state (Halbert 2010) and in other southeastern states.
Distribution
Diaphorina citri is thought to be indigenous to tropical and subtropical Asia and
has been reported in the following geographical areas: China, India, Burma, Taiwan,
Philippine Islands, Malaysia, Indonesia, Ceylon, Pakistan, Thailand, Nepal, Sikkim,
Hong Kong, Ryukyu Islands, Afghanistan, Saudi Arabia, Reunion and Mauritius (Mead
1977, Halbert and Manjunath 2004). D. citri was also known to occur in South America
in Brazil (Lima 1942, Mead 1977). During the 1990s, D. citri invaded the West Indies
(Guadeloupe), Abaco Island, Grand Bahama Island, and Cayman Islands (Halbert and
Núñez 2004). In June 1998, D. citri was detected on the east coast of Florida, from
Broward to St. Lucie counties (Halbert 1998), by September 2000, this pest had spread
to 31 counties in Florida (Halbert et al. 2001). During 2001, it was found in the
Dominican Republic, Cuba (Halbert and Núñez 2004), Puerto Rico (Pluke et al. 2008)
and Texas (French et al. 2001).
Transmission Mechanism between Pathogen and Vector
Information about interactions between D. citri and the pathogen Las is quite
limited. Nymphal stages of D. citri have the ability to acquire HLB pathogen during the
later instar development when growing on the pathogen-infected trees. Adults emerging
from these infected nymphs can transmit the pathogen immediately after emergence
(Capoor et al. 1974, Xu et al. 1988). Uninfected adults feeding on infected trees can
also acquire the pathogen (Xu et al. 1988). However, compared with the adults, later
instar nymph stages have a higher efficiency in acquiring the pathogen in nature (Xu et

15
al. 1988). In more recent studies, nymphs reared on Las-infected trees were also more
likely to acquire the pathogen than adults (Pelz-Stelinski et al. 2010). Acquisition access
period (AAP), latent period, and inoculation access period (IAP) all varied a lot in the
past studies. An AAP of 15min to 24h was reported for both D. citri and T. erytreae by
Capoor et al. (1974), and Buitendag and von Broembsen (1993), whereas Xu et al.
(1988) reported an AAP of between 30min to 5 h for D. citri.
The transmission mechanism between D. citri and the pathogen Las is persistent
and presumably propagative transmission. The pathogen apparently multiplies in the
vector, and the adult D. citri generally remain infective during their whole lifespan (Xu et
al. 1988, Hung et al. 2004); but Pelz-Stelinski et al. (2010) found that the ratio of Las
infected adults declined over time when reared on healthy trees, even though the
pathogen appears to multiply in the vector body.
Following acquisition, adults of T. erytreae may require 21 days of latent period
before they can transmit the pathogen (Moll and Van Vuuren 1977). Latent period for D.
citri was also reported to be 8 to 12 days by Cappor et al. (1974). IAP was reported from
15min to 7h for both psyllid species (Capoor et al. 1974, Buitendag and von Broembsen
1993), but a study with D. citri concluded that the disease could be transmitted after
adults had fed on healthy trees for 5hr to 7hr (Xu et al. 1988). Transovarial transmission
was not reported to occur between D. citri parents and offspring (Xu et al. 1988, Capoor
et al. 1974, and Hung et al. 2004). However, Pelz-Stelinski et al. (2010) reported that
transovarial transmission does occur at a rate from 2% to 6%.
Host Plants
Diaphorina citri has a restricted range of host plants including citrus, orange
jasmine (Murraya paniculata), orange boxwood (Severinia buxifolia), and some other

16
related species of Rutaceae (Halbert and Manjanath 2004). Murraya paniculata was
reported to be more tolerant than citrus of direct feeding damage from D. citri (Skelley
and Hoy 2004). Murraya paniculata has been widely used to rear D. citri, and it is also
an ornamental plant and a host for both psyllids and bacteria. However, Walter et al.
(2012) suggested that, M. paniculata is serving as a minor source of Candidatus
Liberibacter asiaticus inoculum, because they found less than 1% of psyllids and 1.8%
of plants were positive, using sensitive quantitative polymerase chain reaction (qPCR)
targeting at two prophage genes of Candidatus Liberibacter asiaticus. The
development, longevity and fecundity of D. citri varied on different host plants (Fung and
Chen 2006, Nava et al. 2007). Tsai and Liu (2000) reported that D. citri has a higher
rate of development on grapefruit compared with rough lemon, sour orange and orange
jasmine. Nymphal viability tested on Sunki mandarin was lower than that on ‘Rangpur’
lime and M. paniculata (Nava et al. 2007). Fecundity was higher on M. paniculata.
Diaphorina citri developed better on Rangpur lime and orange jasmine compared to
Sunki mandarin (Nava et al. 2007).
Biology of D. Citri
Life cycle
The life cycle of D. citri is incomplete metamorphosis, so there are only three life
stages: egg, nymph and adult.
The egg has been described as oval or almond-shaped, elongated, slightly
curved with ends in a blunt point (Husain and Nath 1927). A slender stalk-like process
appears from its rounded base which links the egg with the plant tissue (Husain and
Nath 1927, Tsai and Liu 2000). Egg length was reported to be 0.3 mm on average
without the stalk, or about 0.038 mm with the stalk. The greatest diameter was around

17
0.13 mm (Husain and Nath 1927). The color of the eggs is light yellow when newly
deposited, gradually turning bright orange with two distinct red spots, the eyes of the
nymph, just before hatching (Husain and Nath 1927, Hall 2008a).
D. citri development includes five nymphal instar stages. The general color of the
nymph is light yellow or dull orange (Husain and Nath 1927, Hall 2008a). As nymphs
mature, the abdominal color of some turn green while some others turn pale orange
(Tsai and Liu 2000). The first nymphal instar is 0.3mm long and 0.17 mm wide with two
visible red eyes (Tsai and Liu 2000). Wing-pads are not yet visible. Some setae arise
from the surface of the dorsum and two golf-club-shaped tarsal setae arise from of the
middle and hind legs with a single seta on each of the fore legs (Husain and Nath
1927). All three features differentiate the first instar from the other stages (Husain and
Nath 1927). The second instar is 0.45mm long and 0.25 wide (Tsai and Liu 2000), with
small triangular wing-pads distinct on the dorsum of the thorax (Tsai and Liu 2000,
Husain and Nath 1927). The third instar nymphs are 0.74mm long and 0.43 mm wide on
average, the wing pads are well developed and the antennal segmentation is visible
(Tsai and Liu 2000). There is a single lanceolate seta on each antenna (Husain and
Nath 1927). The fourth instar nymphs have two setae on each of the antennae (Husain
and Nath 1927). The fifth instar nymphs averaged 1.6mm long and 1.02mm wide (Tsai
and Liu 2000), with a distinctly marked thorax and well developed wing-pads (Husain
and Nath 1927). There is only one seta on each tarsus on the leg, and three setae on
each antenna (Husain and Nath 1927). Paiva and Parra (2012) found that, the egg to
adult survivorship was really low with only 7 adults emerging from 406 eggs under field

18
conditions. Of all the generations, egg to first nymphal instar showed the lowest survival
of 38.8% in field
D.citri adults are 2.7 to 3.3 mm in length, with mottled brown wings (Hall 2008a).
When feeding or resting, they always align their heads against the surface of the leaves,
such that the axis of the body is at 45°angle with the leaf surface (Hall 2008a). Adults
can copulate soon after emergence, and eggs are laid if young shoots are available
(Husain and Nath 1927). Both oviposition and development of immature D. citri are
confined to young, new growth (Hall and Albrigo 2007), and all the activities are
restricted to the daylight (Wenninger and Hall 2007). Sex ratio of adult D. citri was
reported around 1:1 by Aubert and Quilici (1988). New adults become physically mature
in 2 to 3 days, and female adults begin to lay eggs 1 or 2 days after mating (Wenninger
and Hall 2007). Females can lay eggs continuously throughout their lifetime if new and
young shoots are available (Hall 2008a). Adult females live approximately 40 to 48
days, and lay 500 to 800 eggs on average during lifetime, and 1900 at most (Husain
and Nath 1927, Nava et al. 2007, Tsai and Liu 2000). Adults usually lay eggs in the
folds of partially opened leaves, push them between the bud and stem or petioles of
leaves and axillary buds, and other similar situations (Husain and Nath 1927). Eggs are
attached to plant tissue by means of their stalks, which can protect them from bad
climatic conditions, such as washing off by heavy rain (Husain and Nath 1927).
Seasonal history
All three stages of D. citri are found on citrus plants throughout the year, and no
hibernation of a specific stage has been discovered (Husain and Nath 1927). Adults
have been observed throughout winter, but usually lay no eggs until spring because of
the lack of new growth. Large numbers of eggs and nymphs were found during March

19
and April, but in the northern Punjab, the most frequent stage found in the dry May and
June period is the adult stage (Husain and Nath 1927). If rains start in July, there will be
a second peak of the nymphal population coinciding with the burst of new growth.
Populations begin to decline in the middle of October, and always remain low until
January the next year (Husain and Nath 1927).
Dispersion of D. citri
Adults are very active; they can move a short distance through jumping and flying
(Husain and Nath 1927, Hall 2008a). Aubert and Hua (1990) reported that adults fly to
disperse all day long, but are most apt to move during warm, sunny afternoons between
4 to 6 pm. Hall (2008a) speculated that flying psyllids could be carried by wind over a
0.5 to 1.0 km distance according to wind speed and the duration of flight. Adults
congregate on young and fresh leaves, and move quickly when searching for a place to
oviposit (Husain and Nath 1927). Husain and Nath (1927) stated that adult D. citri are
positively phototropic and negatively geotropic (Husain and Nath 1927). Nymphs
concentrate on the young leaves close to the emergence site. Nymphs do not move
much, but will crawl down the stem to larger leaves when the population density is high
(Husain and Nath 1927).
Damage by D.citri
Besides being the vector of the HLB pathogen, D. citri can also cause damage
through feeding on plants. D. citri is a sucking insect that inserts its mouthparts into the
plant tissue to suck the sap (Husain and Nath 1927, Hall 2008). High adult populations
cause defoliation of shoots attacked and dieback of branches (Husain and Nath 1927).
Nymphs suck the cell sap and exclude thick sugary honeydew from the anus encased in
a waxy secretion secreted by the circumanal glands (Husain and Nath 1927). Nymphs

20
move their abdomen to dislodge the wax-covered honeydew, and some fungi will also
grow on it, which makes the leaves look to be covered by a black sooty deposit (Husain
and Nath 1927).
Influence of temperature and humidity
Tsai and Liu (2000) recorded that eggs hatch in 4 days at 25 C. At this
temperature, the nymphal stage lasts over 13 d, leading to a total 17 days from egg to
adult. The life cycle varies significantly with different temperatures, 24 to 28 C being the
optimal temperature range. Diaphorina citri totally fails to develop at temperatures
above 33 C or below 10 C (Liu and Tsai 2000). Nava et al. (2007) reported that, with
temperature varying from 18 to 32℃, the duration of the egg and nymphal stages is 2.6
to 7.7 d and 9.4 to 35.8 d, respectively. Lower temperature development threshold (TT)
and thermal constant (K) for egg, nymph and whole life cycle were 12.0 C - 52.6
Degree-Day (DD), 13.9 C - 156.9 DD and 13.5 C - 210.9 DD respectively (Nava et al.
2007). Longevity differs between females and males even at the same temperature;
adult males live 21 to 25 d, while females live 31 to 32 d on average (Nava et al. 2007).
Different temperatures also influence adult longevity which ranges from 117 days at 15
C to 51 days at 30 C (Liu and Tsai 2000). Skelley and Hoy (2004) showed that D. citri
stopped laying eggs at 34 ℃ or above. However, fecundity returned if temperature was
later reduced. Fecundity is reduced at RH below 40% (Hall 2008a).
The survival of D. citri may increase with increasing relative humidity (McFarland
and Hoy 2001). Yang (1989) reported that nymphal mortality is low from RH 43% to
75% but increases at RH 85% to 92%. The combined influence of temperature and
humidity can reinforce the effect. Nymphal mortality was high at high temperature (34

21
℃) and a high humidity (82-92%) and low at moderate temperature (20-30 ℃) and a low
humidity level (43-75%) (Yang 1989). However, in general, temperature plays a more
important role in influencing nymphal survival than humidity (Yang 1989).
Influence of light
Yang (1989) found that the number of eggs laid per female increased with
increasing light intensity and duration under constant conditions below 11000lux and
light duration less than 18 h per day. Both light duration and light intensity influenced
preoviposition period, fecundity, and mortality of females. However, light duration had a
slightly greater effect (Yang 1989).
Natural Enemies
Predators
Diaphorina citri is commonly attacked by ladybeetles (Coleoptera: Coccinellidae),
syrphid flies (Diptera: Syrphidae), lacewings (Neuroptera: Chrysopidae, Hemerobiidae),
and spiders (Araneae) (Michaud 2002, Michaud 2004, Gonzalez et al.2003, Qureshi
and Stansly 2009). Adults are not very vulnerable to these natural enemies (Husain and
Nath 1927). Coccinellid predators, such as Harmonia axyridis Pallas and Olla V-nigrum
Mulsant have been considered the most important predators in Florida (Michaud 2002,
2004). Exochomus childreni Mulsant, Cycloneda sanguinea L. (Michaud 2004), and
Curinus coeruleus Mulsant (Michaud and Olsen 2004) have also been observed to
attack D.citri in Florida. Two lacewings, Ceraeochrysa sp. and Chrysoperla rufilabris
Burmeister, were reported to contribute to psyllid mortality in Florida (Michaud 2004).
The spider Hibana velox (Becker) was reported to be of great importance in Florida
(Michaud, 2004). Ants can just prey on immature D. citri in Florida (Michaud 2002).

22
(Juan-Blasco et al. 2012) found that Amblyseius swirskii (Acari: Phytoseiidae) showed
significant predation on D. citri eggs, and also suck body fluids of first instar nymphs.
Parasitoids
Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae) and Diaphorencyrtus
aligarhensis (Shafee, Alam and Agarwal) (Hymenoptera: Encyrtidae) are two well know
parasitoids of D. citri (Hall 2008b, Grafton-Cardwell et al.2013). The ectoparasitoid T.
radiata and endoparasitoid D. aligarhensis were recorded as primary parasites (Tang
1990). Marietta leopardina was found attacking immature T. radiata and D. aligarhensis
(Hoddle 2012). D. aligarhensis was released in Florida but has not yet been reported to
have established (Rohrig et al. 2011).
Tamarixia Radiata
Distribution
Tamarixia radiata, the arrhenotokous ectoparasite of D. citri, was first recorded In
Pakistan (Waterston 1922). In 1978, T. radiata was introduced to Reunion Island from
the Punjab of India and was credited with achieving control of D. citri (Etienne and
Aubert 1980). It was also reported to have been successfully introduced in Mauritius in
1984 (Aubert 1987). It is well known to occur in Jiangxi and Fujian provinces in China
(Tang 1998), but was not released there on purpose (Yang et al. 2006). It was
introduced to Taiwan in 1983 for control of D. citri, and is very effective there (Chien and
Chu 1996). Distributions of T. radiata in Asia include India (Husain and Nath 1927),
Japan (Kohon et al. 2002), Nepal (Lama et al. 1988), Thailand (Markote and Nanta
1995), and Vietnam (Hoy et al. 1999). The parasitoid was imported from Taiwan and
Vietnam and released in Florida in 1999-2001 (Hoy and Nguyen 2001). Tamarixia
radiata has been detected in Texas (French et al. 2001), Brazil (Gomez et al. 2006),

23
and Puerto Rico (Pluke at al.2008) although no releases were ever recorded for those
locations. Tamarixia radiata is established in Florida now, although the parasitism rate is
relatively low except for late in the growing season (Tsai at al. 2002, Qureshi et al.
2009).
Taxonomy and Identification
Tamarixia radiata was originally described as Tetrastichus radiatus by Waterston
(1922) from specimens collected at Lyallpur, India (now Faisalabad, Pakistan). The
genus Tamarixia was split off as a separate genus from Tetrastichus by Mercet (1924).
All are parasitoids of Psylloidea (LaSalle 1994). Tang and Aubert (1990) described
some distinguishing characteristics of T. radiata: about 1 mm long including head; eyes
red in fresh specimens; head and body blackish and shining but without a metallic
sheen; underside of gaster pale with a large whitish basal patch on the 5th tergite of
dorsum; legs totally pale-white and wings transparent. Male and female are the same in
color and body structure, except for antennae. The female antenna has eight segments,
both funicle and club with three segments covered with fine, short setae. The funicle is
slender with the 1st segment longer than the 2nd and the 2nd segment longer than the 3rd.
The length of the 3rd segment is almost equal to the width. The male antenna is more
slender, and nine-segmented. The four-segmented funicle is covered with long, slightly
curved hairs, and the ventral scapal sensorium is near to the base of the scape (Tang
and Aubert 1990).
Host Specificity
T. radiata is not known to attack any psyllid other than D. citri (Aubert and Quilici
1984). (Mann et al. 2010) reported that T. radiata can be attracted by volatiles
emanating from D. citri nymphs.

24
Biology of T. Radiata
Life cycle
Structure of the 4 life stages of T. radiata was described by Chen et al. (1991a).
The egg is translucent, ivory, and reniform, with one end adhesive to the host. There
are four larval instars, each distinguished by head length (Chen et al. 1991a).
Development of the immature stages was reported by Xu and Tang (1993) and Chien et
al. (1991a). The newly eclosed larva sucks fluids externally from the site where it is
closely attached to the nymph’s integument (Husain and Nath 1923, Tang and Huang
1991, Hall 2008b). The third instar crawls to the ventral side of the host thorax to feed
(Chien et al. 1991a). The parasitized nymph continues to live and secrete honeydew for
some time (Husain and Nath 1923). By the time the parasitoid molts to the fourth instar,
all contents of the nymph have been consumed and the nymph turns to a dark-brown
mummy (Husain and Nath 1923, Chien et al. 1991a). The mature larva ceases to feed
as it progresses to the prepupal stage which secures the mummy to the plant surface by
means of silken threads (Chien et al. 1991a). After expelling the meconium, it molts to
the pupal stage which turns yellow, with red ommatidia and ocelli (Chen et al. 1991a).
Emergence
As soon as the adult hardens, it makes its way out of the mummy by chewing a
round hole of approximately 0.5 mm diameter in the region of thorax (Husain and Nath
1923, Chien et al. 1991a, Aubert 1987). Over 80% of adults emerge between 5 a.m. to
10 a.m., with the peak between 7 a.m. to 8 a.m. Male emergence occurs 1.5 hours
earlier than the female on average (Chien et al. 1991a).

25
Mating
Males use their antennae to locate receptive females. Once found, the male
crawls onto the dorsum of the female to make contact for an average of 68±7 seconds
before mating which lasts an average 33±3 seconds (Chien et al. 1991a). About 93% of
females mate once and only once during the first day of emergence. The remainder
mate twice during the first two days following emergence. Fecundity and longevity of
females were not affected by mating frequency. Males are capable of multiple matings
over a lifetime (Chien et al. 1991a).
Oviposition
Eggs can be laid immediately after emergence by mated or unmated females
(Chien et al. 1991a). From 5 am to 10 am is the most active time of day for oviposition
(Chu and Chien 1991). Host volatiles mediate host location (Mann et al. 2010). The
female moves actively among D. citri nymphs using her antennae to search for a
suitable host (Husain and Nath 1923). After an acceptable host is found, she deposits
an egg or occasionally two on the underside of the nymph, usually next to the mid or
posterior coxae (Husain and Nath 1923, Aubert 1987, Chien et al. 1991a, Hall 2008b,
Tang and Huang 1991). Oviposition took 3 to 4 min according to Husain and Nath
(1923), but only 61±8 s according to Chien et al. (1991a). Chien et al. (1991a) also
reported that the female T. radiata injects venom into the host nymph through the
ovipositor, immobilizing it for 4 to 8 min. If the egg was removed, the host nymph could
not molt, and died 8 days later at a temperature of 25 C. A first, second, or third stage
parasitoid larva placed on an unparasitized 5th instar nymph could not attach, and
dropped off when the nymph began crawling (Chien et al. 1991a).

26
Host preference
Studies on host selection of T. radiata females have given varying results. Chien
et al. (1991a) and Chu and Chien (1991) reported that 5th instar nymphs were preferred
for oviposition. Survival rate was 85%, compared with 33% and 71% from 3rd and 4th
instars, respectively (Chien et al. 1991a). Body length was also greater among offspring
from 5th instar hosts compared to 4th instar hosts: 1.12 mm compared with 0.91 mm
(female), 1.03 mm compared with 0.86 mm (male). The pattern was repeated with
fecundity: 215 compared to 120 eggs per female, and longevity: 18.0 d compared to
14.4 d (females), or 11.6 d compared to 7.2 d (males) (Chien et al. 1991). However, 4th
instar nymphs are parasitized significantly more than 3rd or 5th instars according to Tang
and Huang (1991).
Host-feeding
Both sexes feed on honeydew from D. citri nymphs, and females can suck
hemolymph after puncturing the nymph with the ovipositor (Chien et al.1991a, Skelley
and Hoy 2004). The T. radiata population oviposited and host-fed almost simultaneously
(Chu and Chien 1991). Host-feeding occurs in daytime and takes an average of 21±2 s
(Chien et al. 1991a). The host will die once fed upon and females avoid laying eggs and
feeding on the same host (Chien et al. 1991a, Tang and Huang 1991). Ratio of host-
feeding to oviposition correlates with host density and parasitoid age (Chien et al.
1991a). One egg was laid for an average 0.18 hosts fed upon by females between 4 to
18 d old. Younger or older females laid one egg for an average 0.29-0.38 hosts fed
upon respectively (Chien et al. 1991a). An average of 80% mortality resulted from
parasitism with an additional 20% from host-feeding (Chien et al. 1991a, 1994b). In this
way, a single female T. radiata could kill up to 500 nymphs during her lifetime (Chu and

27
Chien 1991). However, Chien et al. (1993) estimated host-killing capacity at 16, 25, 245,
196 nymphs per female at 15 C, 20 C, 25 C and 30 C respectively.
Sex ratio
T. radiata is arrhenotokous, meaning virgin females deposit eggs which produce
males, while eggs deposited by mated females can develop into either sex. The
average number of eggs deposited by virgin and mated females in one study was 209.2
and 215.4 respectively (Chu and Chien 1991). Female progeny ratio is highly correlated
with parasitoid age (Tang and Huang 1991, Chu and Chien 1991). The proportion of
female progeny increased as the mother aged, from 0.5 from a 1 d old female to 0.77
from a 22 d old female (Chu and Chien 1991). Sex ratio is also correlated with host
stage, although published results differ. Tang and Huang (1991) reported female ratios
of 0.88 from 5th instar hosts, 0.75 from 4th instars and 0.41 from 3rd instars. However,
Chu and Chien (1991) found female ratios of 0.67 from 5th instar nymphs, compared
with 0.16 from 4th instars. Skelley and Hoy (2004) reported female ratios of 0.64 and
0.67 with 4th instar nymphs for their Taiwanese and Vietnamese colonies of T. radiata.
Reproduction
Tamarixia radiata can be characterized as synovigenic autogenous, meaning that
some eggs become mature in the newly emerged wasp without feeding, but that host-
feeding is later required to mature more eggs. Most synovigenic parasitoids, including T.
radiata, can resorb eggs when hosts are absent or scarce (Chien et al. 1994a), thereby
maintaining reproductive resources and synchrony with the host population (Jervis et al.
2001). Egg resorption is thus a mechanism that aids adaption to host biology and
ecology. It can occur in T. radiata at either 15 C or 25 C when only honey is provided
and is positively related to host deprivation time (Chien et al. 1991a, 1994a).

28
Once suitable hosts are fed upon, new mature eggs can be produced in the
ovary and oviposition can proceed normally (Chien et al. 1994a). Little or no effect on
total fecundity was observed after host deprivation for 10 days at 25 C with honey
provided as food (204 eggs), but there was a difference after host deprivation for 20 d
(156 eggs) (Chien et al. 1994a). Wasps thus stored for 10 to 20 d, wasps at 25 C laid
significantly more eggs (156 eggs) than wasps stored at 15 C (98 eggs). Fecundity
decreased greatly following host deprivation for 30 to 40 d at 15 C (25~59 eggs).
Superparasitism
Female T. radiata can discriminate between parasitized and unparasitized hosts
to avoid superparasitism (Chien et al. 1991a). Husain and Nath (1923) observed
superparasitism during December and January when hosts were scarce, but not when
hosts were abundant. Chien et al. (1991a, 1991b) observed superparasitism rates of up
to 5.6% when the host density was low and active space was limited.
Host density effects
Longevity, fecundity, sex ratio, and ratio of host-feeding to oviposition all
correlate with host density (Chu and Chien 1991, Chien et al. 1991a, 1995). The
relationship between host density and wasp longevity (both males and females) has
been described as following a domed parabolic response, meaning that female
longevity and fecundity ascend with host density to a peak and then decrease as host
density increases (Chien et al. 1995). Average female longevity increased from 15.9 to
18.6 to 20.3 d when host densities of 10, 20 and 30 were provided daily. However,
female longevity decreased from 23.6 to 17.2 to 11.2 over a range of 40, 60, 80 hosts
per day, respectively (Chien et al. 1995). Chu and Chien (1991) reported that females
lived an average 23.6 days and males 14.8 d when 40, 5th instars were provided at 25

29
C, 14:10 (L: D) photoperiod and 100% RH. Both daily and lifetime fecundity showed
similar parabolic responses to host density, with the peak at 40 hosts per day (Chien et
al. 1995).
Influence of food
Adults deprived of food or water survived 1.0 to 1.7 d (Chien et al. 1994b). These
authors found no difference in longevity among female adults deprived of hosts and fed
either honey alone (22.5 d), honey and pollen (23.0 d), or honey and yeast extract (23.4
d). However, all these food supplements increased fecundity and progeny survivorship
compared with adults held without food or water. Adults fed on a diet of honey and
yeast extract significantly decreased host-feeding while maintaining or improving
intrinsic rate of increase (0.2976 to 0.3014 per day) and the net reproductive rate (140
to 187 female eggs per female), respectively (Chien et al. 1994b).
Development time
Chien et al. (1991a) found the duration of one generation for T. radiata on orange
jasmine (egg to adult emergence) to be around 11.4 d at 25 C, 14:10 (L: D)
photoperiod, and 100% RH. This would include 45 h for the egg, 24, 24, 22, 26 h for the
1st through 4th instars, respectively, 14.4 h for the prepupal, and a 117.6 h pupal stage.
Xu and Tang (1993) reported a 12.6 d generation time at 25 ± 1 ℃, 14:10 (L:D)
photoperiod, and 75% to 85% RH: 40 h for the egg, 119 h = 25, 28, 32 and 34 h for 1st
to 4th larval instar respectively, 24 h for the prepupa and 120 h for the pupa. These
results could indicate that humidity may play an important role in development rate
especially in the prepupal and pupal stages.

30
Temperature and humidity effects
Tamarixia radiata completes development at 15 C to 32 C with an optimum
temperature of 25 ℃ (Chien et al. 1993). Gomez-Torres et al. (2012) found parasitism
rates to be highest at 25 and 30 °C (85.5 and 72.8%, respectively) compared to 23.1
and 40.2% at 15 and 35 °C respectively. They also found emergence rates to be
highest, 86.7 and 88.3%, at 25 and 30 °C respectively, compared to about 50% in the
15 to 20 °C range. At 70 ± 10% RH, and L: D= 14:10 they estimated maximum
parasitism rates of 77.2% at 26.3 ℃, whereas emergence was greatest (89.9%) at 30.8
℃. Pre-imaginal development was longer for females, varying from 489.6 h at 15° C to
247.2 h at 35 °C compared to males at 343.2 to 146.4 h over this same range.
Longevity with access to pure honey negatively correlated with temperature
between 8 C to 30 C (Quilici and Fauvergue 1990). These authors found that adult
longevity decreased from 34 d at 20 C to 22 d at 22 C, 10 d at 30 C and 8 d at 35 C.
Chien et al. (1993) found longevity to increase from 45.5 to 59.5 between 8 and 15 C
but then decrease to 22.5 and 9.6 days at 25 and 30 C respectively. Ten percent of T.
radiata adults survived for 50 d when stored at 25 C with access to honey and yeast
extract (Skelley and Hoy 2004).
McFarland and Hoy (2001) reported that T. radiata adults from Vietnam survived
longer without food and water compared to wasps from Taiwan over a range of RH from
7% to 97% at 25 °C and especially 30 °C. They attributed this difference to greater
moisture requirements of wasps from Taiwan.
Chien et al. (1993) estimated host-killing capacity at 16, 25, 245, 196 nymphs per
female at 15 C, 20 C, 25 C and 30 C. Estimates of intrinsic rate of increase (r), net

31
reproductive rate (R0), and mean generation time (T) for pairs of T. radiata provided 20
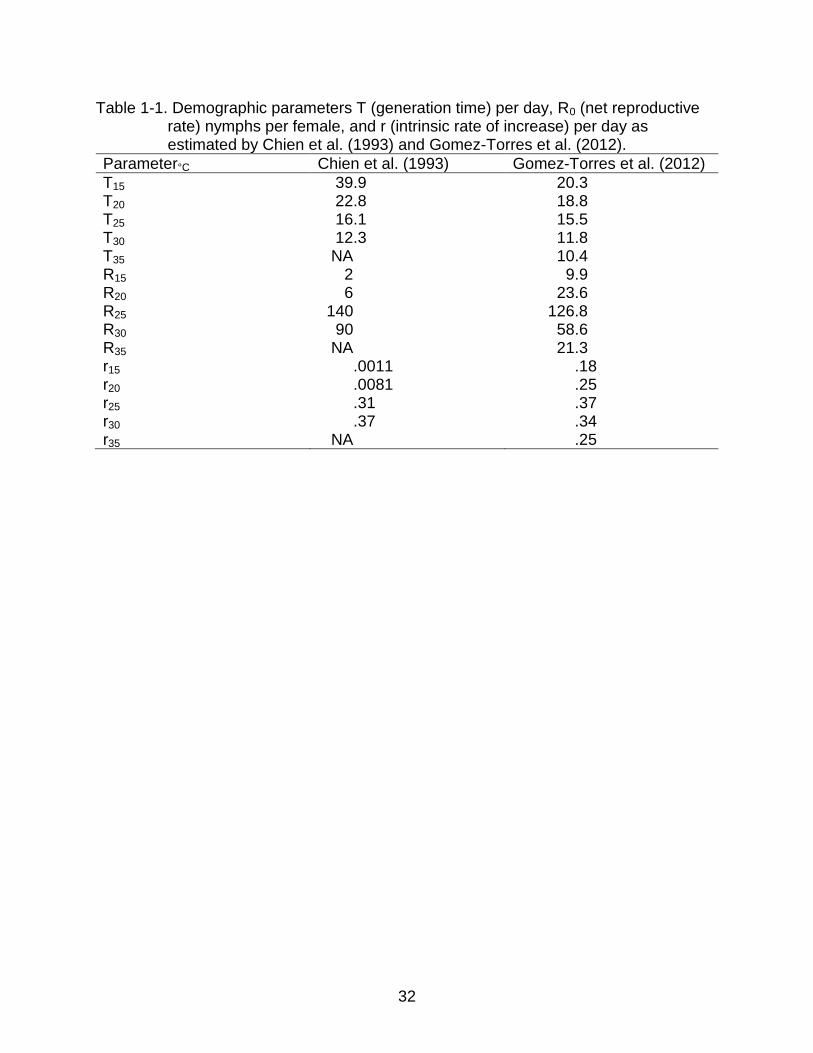
or 30, 5th instar nymphal hosts by Chien et al. (1993) and Gomez-Torres (2012)
respectively varied considerably, especially at low temperatures (Table 1-1). Different
results may have been due to different conditions during these two studies. Chien et al.
(1993) did theirs at host density 20 and 100% RH for five replications whereas Gomez-
Torres et al. (2012) conducted their study at density 30, 70 ± 10% RH for ten
replications, which may have led to the different results. Differences could also be
inherent in the races of T. radiata tested from Taiwan and Brazil respectively.
Skelley and Hoy (2004) showed that T. radiata stored for up to 35 d at 17 C with
honey and yeast suffered less than 5% mortality. Chien et al. (1994a) reported that
females stored for 20 d at 25 C were able to lay a total of 156 eggs, compared with 98
eggs when stored at 15 C. We may conclude that less mortality was experienced at low
temperature but that production suffered. Therefore, ideal storage temperature should
be determined according to specific objectives (establishment or augmentation) and
conditions (host availability).
32
Table 1-1. Demographic parameters T (generation time) per day, R0 (net reproductive rate) nymphs per female, and r (intrinsic rate of increase) per day as estimated by Chien et al. (1993) and Gomez-Torres et al. (2012).
Parameter°C Chien et al. (1993) Gomez-Torres et al. (2012)
T15 39.9 20.3 T20 22.8 18.8 T25 16.1 15.5 T30 12.3 11.8 T35 NA 10.4 R15 2 9.9 R20 6 23.6 R25 140 126.8 R30 90 58.6 R35 NA 21.3 r15 .0011 .18 r20 .0081 .25 r25 .31 .37 r30 .37 .34 r35 NA .25

33
CHAPTER 2 CARBON DIOXIDE ANESTHESIA OF TAMARIXIA RADIATA (WATERSTON)
(HYMENOPTERA: EULOPHIDAE) PARASITOID OF DIAPHORINA CITRI (HEMIPTERA: PSYLLIDAE)
Introduction
Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae), is an arrhenotokous
ectoparasite of the Asian citrus psyllid (ACP) Diaphorina citri (Kuwayama) (Hemiptera:
Psyllidae), vector of citrus greening disease or huanglongbing (HLB). The parasitoid is
reported to have controlled ACP populations to low levels on the islands of Réunion,
Guadeloupe and Puerto Rico (Aubert & Quilici 1984; Etienne et al. 2001, Pluke et al.
2008). Tamarixia radiata was first imported to Florida from Taiwan and Vietnam in 1998
and released in 1999-2001 (Hoy & Nguyen 2001). A survey conducted in 2006-07
determined that T. radiata was well distributed in citrus orchards throughout the state
(Qureshi et al. 2009). However, incidence of parasitism was generally low, especially
early in the growing season, suggesting a need for augmenting parasitoid populations at
that critical time as component of an integrated management program (Qureshi et al.
2007, 2009). Studies of T. radiata biology and current efforts at mass rearing and
release of this species might benefit from an ability to inactivate adults by CO2
anesthetization, including separation of emergent wasps and psyllids.
Carbon dioxide (CO2) is widely used to anesthetize insects, but may also cause
deleterious side effects on biology and behavior. (Brooks 1957) found that development
rate of German cockroach, Blattella germanica L. (Blattodea: Blattellidae), nymphs
decreased 53% when exposed weekly to high CO2 concentrations for 3 min. (Crystal
1967) reported significantly decreased survival rates and fertility of New World
screwworm, Cochliomyia hominivorax (Coquerel) (Diptera: Calliphoridae), exposed to

34
100% CO2 for 30 min. Sherman (1953) reported that CO2 anesthesia of Mediterranean
fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae), led to increased mortality.
This study was undertaken as a first step toward possible use of anesthesia for
mass rearing by evaluating the response of T. radiata to CO2 exposure. The objective
was to test the effectiveness of CO2 anesthesia of T. radiata and to determine the
incidence and severity of side effects of CO2 anesthesia on longevity, parasitism rate
and sex ratio.
Materials and Methods
Colonies
A T. radiata colony was maintained at FDACS-DPI in Gainesville on ACP
nymphs using orange jasmine, Murraya paniculata (l.) Jack (Sapindales: Rutaceae), as
plant host. Six newly trimmed plants with new growth were held in an acrylic 62 cm
cubic cage and 600 D. citri adults were released for 72 h for oviposition in a greenhouse
under natural sunlight 25 ± 5 C and 50%~ 70% RH. Adults were removed and plants
held for 10 d until 4th instar nymphs were available. Plants were moved into another
clean cage of the same type for 20 days into which 100 T. radiata were released. Adult
progeny were later collected daily until no more emerged.
Gas Chamber
A gas chamber was constructed consisting of a vial, 6.50 cm in diam and 12 cm
in height (Fisher Scientific, Pittsburgh, Pennsylvania), provided with two 0.50 cm diam
holes in the lid into each of which was fitted a 0.5 cm plastic tube inserted either 1 cm or
11 cm into the chamber for a gas outlet and inlet respectively (Fig 2-1). Plasticine
modeling clay (Flair Leisure Products, Cheam, Surrey, England) was molded around the
openings of the lid to prevent leakage.

35
Flow time of gas at 3.8 kpa (2 psi) needed to displace all air in the chamber was
assessed by filling the vial with water and then replacing with CO2 through the inlet. All
the water was displaced in 10 s. A CO2 sensor (K-33 ICB 30% CO2 Sensor, CO2Meter
Inc., Ormond Beach, Florida, USA) was used to determine that 3 s of flow time were
necessary to attain a 30% CO2 concentration confirming the earlier result. The CO2
sensor was also used to test for leaks by confirming that a given concentration
remained constant over several min.
Recovery Time
Five wasps having emerged within 24 h or less were placed in the chamber. The
lower ¾ of the vial was covered with black cloth to induce the wasps to walk to the top
and thus avoid injury from inrushing gas. Gas was introduced through an inlet from a
CO2 tank at 3.8 kpa for 15 s to exchange all the air, and then the 2 tubes closed with
metal clamps. Wasps were removed after a 5 min exposure and observed with the
naked eye using a stop watch to record recovery time (normal movement). Males and
females were treated separately, each with eight replications so that a total of 80 wasps
was used.
Survival Rate
To evaluate survival, 5 anesthetized wasps were collected into each of 6 small
glass vials (1.5 cm in diam, 5.3 cm high) and provided pure honey on a tissue paper
strip. On the same day control wasps not anesthetized were placed in 6 other vials.
Vials with wasps were held in a growth chamber at 25 C, 14:10 h L: D and 60 ± 5% RH,
and checked daily, noting sex of all cadavers until all had died.

36
Percent Parasitism
Six newly trimmed plants were held in a ventilated 62 cm acrylic cubic cage until
there were at least 3 new shoots 3 cm in length upon which to evaluate parasitism.
Plants were infested by releasing 600 ACP adults for a 24-h oviposition period. ACP
adults were removed and the plants were held for 9 days in a rearing room at 25 ℃ and
60 ± 5% RH. A small brush was used to remove nymphs until exactly 120 fourth instars
remained on each plant. Each plant was then placed individually into a clear acrylic
cylinder (12.5 cm diam, 43 cm high) into which 3 T. radiata females and 2 males were
released. Cages were randomly selected to receive either anesthetized or untreated
wasps (N = 8). Newly emerged T. radiata offspring were collected daily from day-7 until
day-19 after which no more new progeny were found. Progeny were counted and sexed
and parasitism rate calculated based on 120 original hosts.
Results
Seventy percent of T. radiata females recovered from anesthesia with CO2 within
4 min, males recovered about as quickly. Indeed, there was no significant difference
between male and female recovery time (2 = 13.04, df = 7, P = 0.071, Fig 1). It was
noted that a wasp often would recover immediately after being crawled over by another
recovering individual.
Survival rate for the treated wasps was consistently lower than the control over
the entire study period (Fig. 2). Insect-days, the area under the curve of insect numbers
by time (Ruppel 1983), was significantly less for the CO2 treatment (3445.3 ± 348.6)
than the control (5610.5 ± 836.6) (t = 2.39, df = 7, P ˂ 0.05).
A mean of 59.0 ± 3.8 wasps emerged over 12 days from 120 fourth instar hosts
exposed to 3 female and 2 male T. radiata treated with CO2 compared to a mean of

37
85.2 ± 4.5 in the control. This corresponded to a parasitism rate of 49.2 ± 3.2% for
treated wasps compared to 71.0 ± 3.7% for untreated wasps (t = 4.46, df = 14, P =
0.00054). There was no significant difference in progeny sex ratio (t = 1.03, df = 14, P =
0.32) between the treated and the control.
Discussion
Carbon dioxide anesthesia is a convenient tool for manipulating insects, but can
cause deleterious side effects. In this case, a 5 min exposure of Tamarixia radiata
adults to 100% CO2 concentration caused a knockdown of about 4 min, significantly
reduced survivorship and fecundity, but did not affect the sex ratio of progeny from
treated adults. Future research will focus on using less concentrated doses or shorter
exposure times to inactivate the wasps in order to improve survival and fecundity.
38
Figure 2-1. Carbon dioxide gas chamber (Photo courtesy of Xulin Chen)
Figure 2-2. Tamarixia radiata female and male recovery time frequency (%) distribution

39
Figure 2-3. Mean (SEM) survivorship of CO2 treated and untreated Tamarixia radiata adults.

40
CHAPTER 3 FUNCTIONAL RESPONSE OF TAMARIXIA RADIATA (HYMENOPTERA:
EULOPHIDAE) TO DENSITIES OF ITS HOST, DIAPHORINA CITRI (HEMIPTERA: PSYLLIDAE)
Introduction
Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae), is an arrhenotokous
ectoparasite of the Asian citrus psyllid (ACP) Diaphorina citri (Kuwayama) (Hemiptera:
Psyllidae), vector of citrus greening disease or huanglongbing (HLB). The parasitoid is
reported to have controlled ACP populations to low levels on the islands of Réunion,
Guadeloupe and Puerto Rico (Aubert & Quilici 1984, Etienne et al. 2001, Pluke et al.
2008). Tamarixia radiata was first imported to Florida from Taiwan and Vietnam in 1998
and released in 1999-2001 (Hoy & Nguyen 2001). A survey conducted in 2006-07
determined that T. radiata was well distributed in citrus orchards throughout the state.
However, incidence of parasitism was generally low, especially early in the growing
season, suggesting a need for augmenting parasitoid populations at that critical time as
component of an integrated management program (Qureshi et al. 2007, 2009). The
success of such a program would depend, in part, on the efficiency of mass-rearing the
parasitoid. Thus optimization of the host: parasitoid ratio is of critical importance.
Optimal host: parasitoid ratio may also be an important consideration for guiding field
release strategies.
It has been reported that the fecundity of T. radiata is highly correlated with host
density (Chu and Chien 1991, Chien et al. 1991a 1995). The relationship between both
daily and lifetime fecundity with host density showed similar parabolic responses,
meaning that female fecundity ascends with host density to a peak of 40 hosts, and
then decreases as host density increases (Chien et al. 1995). The decline in fecundity

41
at high host density could be due to host defense or excessive honey dew. In fact, he
experiments were conducted at 100% RH which would increase honeydew and
consequent sooty mold that would interfere with the T. radiata female’s search for hosts.
Another issue may be that late 5th instar nymphs are not suitable hosts especially when
they are about to emerge as adults. Similar behaviors have been observed in
Eretmocerus mundus (Mercet) (Hymenoptera: Aphelinidae), parasitoid of whitefly
Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). The female E. mundus never
attacked late 4th instar white fly nymphs once adult development was initiated (Gelman
et al. 2005). Therefore, it is necessary to repeat these experiments under more
favorable conditions of relative humidity and host availability.
The relationship between number of hosts parasitized per parasitoid and host
density is known as functional response. It has been used to illustrate the complex
interactions between parasitoids and their hosts, and is an important character
influencing biocontrol success (Jones et al. 2003). Functional response has been
described as Holling (1959) Type I, II or III, characterized by linear, decelerating and
sigmoidal responses of parasitism rate to increasing host density. Most laboratory
studies show that parasitoids exhibit Type II functional response, Type III functional
responses are also common (Holling 1959). The shape of the functional response curve
varies with parasitoid species, and also parasitoid age (Jones et al. 2003).
My objective was to evaluate the influence of host density on percent parasitism
and the number of hosts parasitized by T. radiata (functional response), to help optimize
rearing conditions in the laboratory as well as release conditions in the field.

42
Materials and Methods
Colonies
Murraya paniculata (L.) Jack (Rutaceae) contained in 3.92 L pots with 40: 60 mix
of Canadian sphagnum peat (Fafard 4P mix Professional Growing Mix soil, Conrad
Fafard Inc.) was used to rear D. citri. Each plant selected had 2-5 shoots of new growth,
with each new shoot about 4 cm in length. Plants were maintained using M-pede soap
(Dow AgroSciences LLC) as a contact chemical to control unwanted psyllids and other
pests, soap sprayed plants cannot be used within 3 d. All plants used in this experiment
were grown in an unheated screen houses (hoop style trussed greenhouse with insect
screen mesh) with natural ventilation and sunlight.
A wood framed cubic cage was used (62 cm in each dimension) 3 sides of which
were covered with fine mesh for ventilation with the other 3 sides enclosed with clear
acrylic. Six flushing M. paniculata plants were placed in the cage and 600 D. citri
released and held there for 24 h for oviposition. Plants were then moved to a similar
clean cage for about seven days until the eggs hatched and nymphs developed to the
4th instar. All cages were held in an air-conditioned rearing room at 25-30 C, 60-80%
RH (Extech RH10 humidity and temperature datalogger, Grainger, Inc) and L: D=14:10.
Murraya paniculata with 4th nymphal instar psyllids were transferred into a clear
acrylic cylinder (12.5 cm diam, 43 cm high) with mesh on top in a growth chamber
(Percival model I36LLC8, Perry, Iowa) at 24 ± 4 C, RH 60-80%, L: D=14:10 (Extech
RH10 humidity and temperature datalogger, Grainger, Inc). Five pairs of newly emerged
T. radiata were released into this cylinder for three days to allow the parasitoids to
mature.

43
Arenas
Arenas were prepared using centrifuge tubes (11 cm long and 3 cm diam). Into
each arena was placed a 7 cm young shoot of M. paniculata infested with 10, 20, 30,
40, 50, or 60, 4th instar psyllid nymphs; the number of nymphs was controlled using a
small paintbrush. Optimal young shoots were chosen as those with more than the
number of hosts needed. A small paintbrush was used to remove extra nymphs to
achieve the desired density. Placing additional nymphs onto the young growth was
avoided, because the newly placed nymphs always crawled off the new shoot. One
randomly-chosen 3-d old parasitoid pair was released into each centrifuge tube and
sealed with Parafilm provided with small holes made by a No.1 insect pin for ventilation.
Arenas were placed in the growth chamber and shoots replaced every 24 h. Exposed
nymphs were inverted under a stereoscopic microscope to check for parasitoid eggs.
Each pair of T. radiata was held in the arena for 5-d, and six replications were
completed for each host density.
Statistical Analysis
Data on number and proportion of parasitized and superparasitized hosts in each
arena among six host densities over the 5-d period were transformed using square root
for normality fitness and then subjected to Analysis of Repeated Measures using JMP
software (SAS Institute, 2012) with mean separation by Fisher’s LSD (P < 0.05). A
polynomial regression of proportion of hosts consumed versus density was used to
distinguish between Type II and Type III functional response (Juliano 2001). A
significant positive linear term coefficient indicates Type III whereas a significant
negative linear term coefficient implies a Type II functional response. Data were then
fitted to the corresponding functional response equation using nonlinear regression in

44
JMP and the functional response parameters: searching efficiency (a) and handing time
(TH) were reported from the regression.
Results
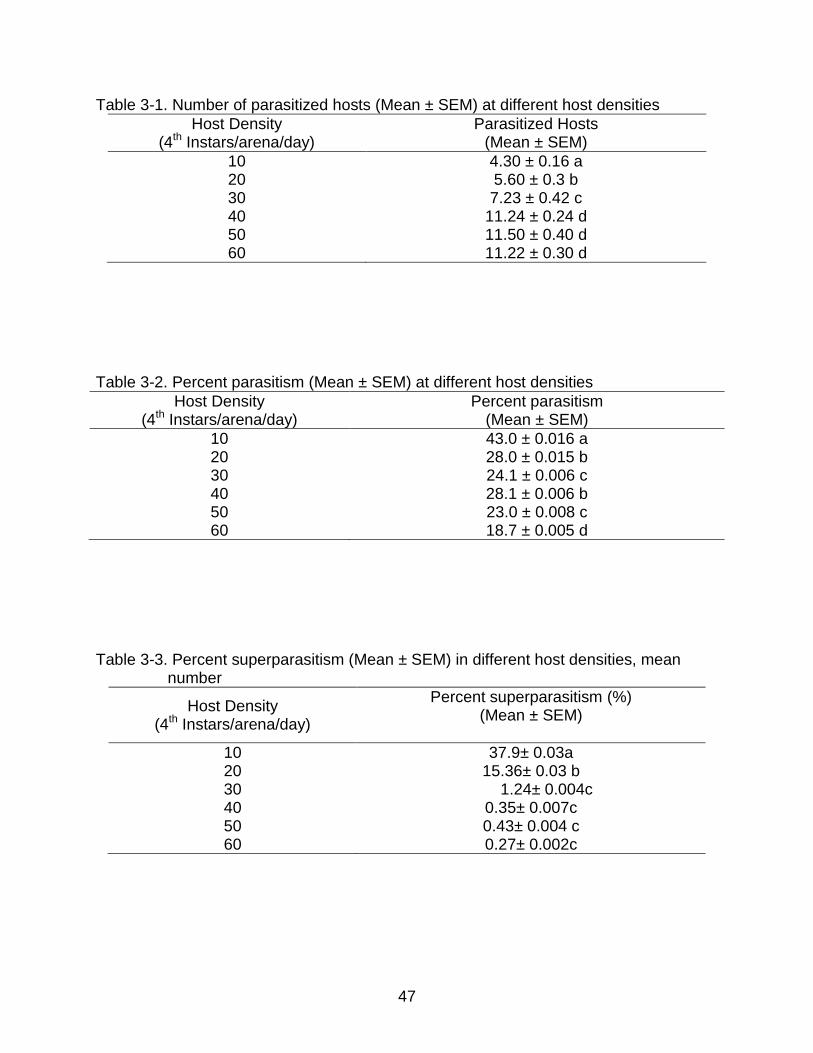
Fecundity, Percent Parasitism and Percent Superparasitism
Inspection of the box plot revealed no outliers among the data, and no significant
differences were observed among replications (F = 0.6461, df = 5, P = 0.6648).
Fecundity was significantly different among the six host densities (F = 111.2432, df = 5,
P < 0.0001), and it increased with host density to a maximum of 11 to 12 eggs per day
with no significant differences among host densities of 40, 50, and 60 (Table 3-1 and
Fig 3-1).
Percent parasitism was significantly different among six host densities (F =
49.6352, df = 5, P < 0.0001): highest at 43% with 10 hosts and least at 18.7% with 60
hosts (Table 2, Fig. 2). Percent superparasitism (superparasitized hosts number over
parasitized hosts) was also significantly different among the six host densities (F =
140.7006, df = 5, P < 0.0001); highest (37.9%) at the lowest host density level of 10
hosts per female, followed by 15.4% at 20 hosts per female. Superparasitism was
negligible at densities over the range of 30 to 60 hosts per female (Table 3)
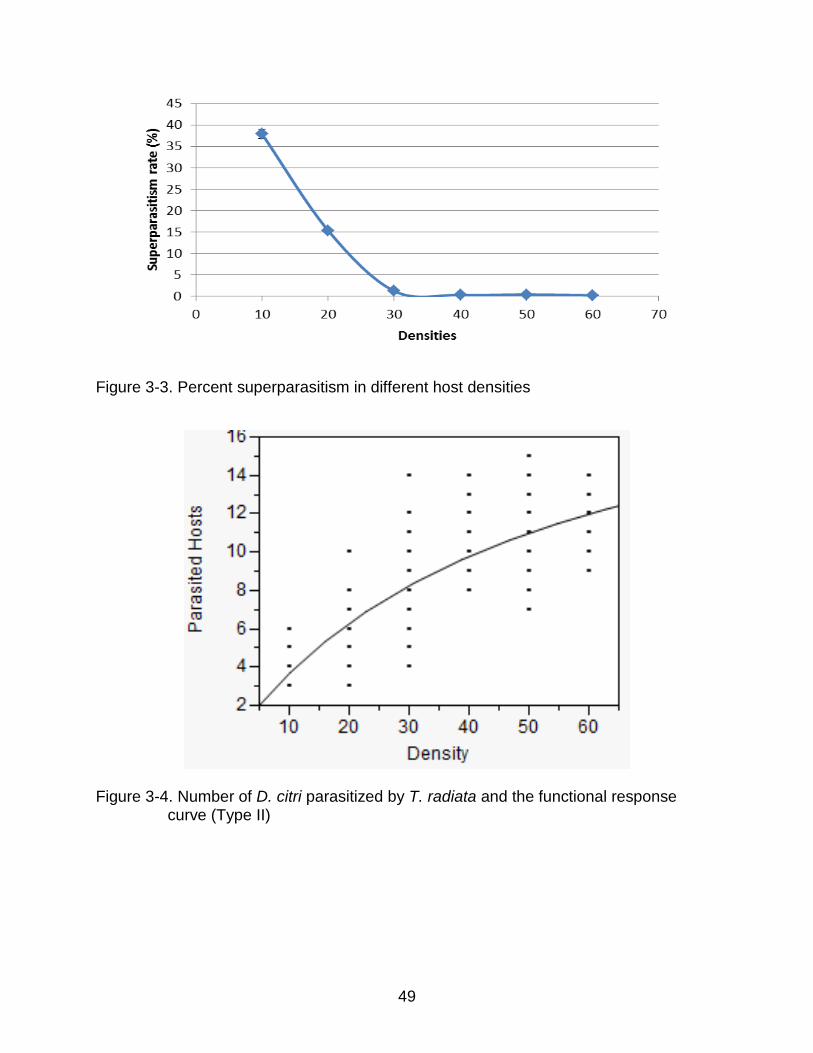
Functional Response
The estimated linear term coefficient for the polynomial regression was -0.0099 ±
0.0015 and significant (t = 4.01, df = 177, P < 0.001), indicating a type II functional
response (Juliano 2001). When the data were fitted to the disc equation (Holling 1959),
the estimated searching efficiency (a) was 0.442 ± 0.036 per day (95% confidence
interval was 0.378~0.520) and estimated handing time (TH) was 0.045 ± 0.008 d (95%
confidence interval was 0.032 ~ 0.063), which were all significant (Fig 4).

45
Discussion
Percent parasitism and percent superparasitism changed with host density. The
number of hosts parasitized increased from density 10 to 40 per female, after which it
remained constant. Host availablility was limited at low host densities, indicating that
fecundity was constrained by lack of hosts. The number of eggs laid did not increase
beyond 40 hosts per female, indicating that the upper limit had been reached.
Parasitism rate was highest at density 10, but 37.9% of the parasitized hosts were
superparasitized, which meant the hosts were not used effectively as only one
parasitoid adult can emerge and supernumerary parasitoids are thus wasted (Chien et
al. 1991a). Therefore a host density of 10 per female would not be efficient for
parasitoid mass-rearing.
A maximum number of hosts were parasitized a host density of 40, and percent
parasitism was highest at densities of 20 and 40 hosts per female. However, 15.4%
superparasitism was observed at 20 hosts per female whereas superparasitism was
negligible at a host density of 40. Therefore a density of 40 host per female optimal
because it maximized number of percentage of hosts parasitized with minimal
superparasitism. When host density exceeded 40, the number of hosts parasitized did
not increase but the percent parasitism declined. However, no decrease in number of
hosts parasitized was observed at host densities above 40 per female as reported by
Chien et al. (1995). In conclusion, a host density of 40 was the optimal choice maximal
progeny and the best for usage of D. citri.
Superparasitism was highest at a host density of 10, and was not significantly
different when the density exceeded 30. Chien et al. (1991) reported that female T.
radiata were able to discriminate between parasitized and non-parasitized hosts. But

46
gravid females still laid eggs even though there were not enough suitable hosts
available resulting in superparasitism. Superparasitism is wasteful because only one
egg can mature to the adult and any others will die.(Chien et al. 1991a) To avoid
superparasitism, host number should be controlled in excess of 20 per female.
The pattern of hosts parasitized by T. radiata over the first five days conformed to
a functional response Type II, which means the searching efficiency and the handing
time were all constant in different host densities. These two parameters will be useful to
compare T. radiata with other parasitoids, and they may help to make a better study of
the behavior of T. radiata in the future.
47
Table 3-1. Number of parasitized hosts (Mean ± SEM) at different host densities
Host Density (4th Instars/arena/day)
Parasitized Hosts (Mean ± SEM)
10 4.30 ± 0.16 a 20 5.60 ± 0.3 b 30 7.23 ± 0.42 c 40 11.24 ± 0.24 d 50 11.50 ± 0.40 d 60 11.22 ± 0.30 d
Table 3-2. Percent parasitism (Mean ± SEM) at different host densities
Host Density (4th Instars/arena/day)
Percent parasitism (Mean ± SEM)
10 43.0 ± 0.016 a 20 28.0 ± 0.015 b 30 24.1 ± 0.006 c 40 28.1 ± 0.006 b 50 23.0 ± 0.008 c 60 18.7 ± 0.005 d
Table 3-3. Percent superparasitism (Mean ± SEM) in different host densities, mean
number
Host Density (4th Instars/arena/day)
Percent superparasitism (%) (Mean ± SEM)
10 37.9± 0.03a 20 15.36± 0.03 b 30 1.24± 0.004c 40 0.35± 0.007c 50 0.43± 0.004 c 60 0.27± 0.002c

48
Figure 3-1. Number of parasitized host at different host densities, error bar stands for
SEM
Figure 3-2. Percent parasitism at different host densities, error bar stands for SEM
49
Figure 3-3. Percent superparasitism in different host densities
Figure 3-4. Number of D. citri parasitized by T. radiata and the functional response curve (Type II)

50
CHAPTER 4 THE INFLUENCE OF DIET ON EGG FORMATION IN TAMARIXIA RADIATA (HYMENOPTERA: EULOPHIDAE), A PARASITOID OF DIAPHORINA CITRI
(HEMIPTERA: PSYLLIDAE)
Introduction
Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae), is an arrhenotokous
ectoparasite of the Asian citrus psyllid (ACP), Diaphorina citri (Kuwayama) (Hemiptera:
Psyllidae), vector of citrus greening disease or huanglongbing (HLB). The parasitoid is
reported to have controlled ACP populations to low levels on the islands of Réunion,
Guadeloupe, and Puerto Rico (Aubert & Quilici 1984, Etienne et al. 2001, Pluke et al.
2008). Tamarixia radiata was first imported to Florida from Taiwan and Vietnam in 1998
and released in 1999-2001 (Hoy & Nguyen 2001). A survey conducted in 2006-07
determined that T. radiata was well distributed in citrus orchards throughout the state.
However, incidence of parasitism was generally low, especially early in the growing
season, suggesting a need for augmenting parasitoid populations at that critical time as
component of an integrated management program (Qureshi et al. 2009). It would thus
be necessary to mass-rear parasitoids to obtain adequate numbers for release,
requiring temporary storage. During the holding period, food provided to females may
affect the number of eggs formed in ovaries, which may influence their efficiency as a
biocontrol agent upon release.
Oogenesis is a nutrition-limited process, and nutrition is obtained during the
larval or adult stage; insufficient nutrients always affect egg production (Wheeler 1996).
Carbohydrate is the major energy source for most insects; but it is not the main nutrition
that triggers egg formation in female insects. Varley and Edwards (1957) reported that
female Nasonia vitripennis (Wlkr.) (Pteromalidae) fed on carbohydrates only survived,

51
but resorbed eggs resulting in gradual decreaseof eggs in the ovaries. (Bownes and
Blair 1986) found that when Drosophila were feeding on sugar diets, the number of
eggs laid and the number of vitellogenic oocytes in ovaries were reduced significantly.
This was due to reduced transcription of yolk protein in the fat bodies resulting in
reduced availability of yolk protein for oogenesis.
Host hemolymph is rich in protein and an important source of nutrients to many
hymenopterous female parasites for increasing fecundity (Howard 1910, Rockwood
1917). Varley and Edwards (1957) observed egg development by female N.
vitripennis,was initiated and/or accelerated shortly after host hemolymph was. (Leius
1961) reported that female Itoplectis conquisitor (Say) (Hymenoptera: Ichneumonidae)
which fed on host body fluid laid more eggs than those fed on carbohydrates alone.
Additionally, Leius (1961) found that pollen in diets significantly increased female I.
conquisitor fecundity, and females fed on a mixed diet (host body fluid and
carbohydrates) deposited even more eggs.
The objective of this experiment was to investigate which foods promote egg
production during storage, presumable improving oviposition rate upon release.
Materials and Methods
Colonies
Murraya paniculata (L.) Jack (Rutaceae) was used as a host plant to maintain a
D. citri colony. Plants were grown in 3.9 L pots using 40: 60 mix of Canadian sphagnum
peat potting soil (Fafard 4P Professional Growing Mix soil, Conrad Fafard Inc.) in a
screen house (hoop style trussed, with insect screen mesh) under natural sunlight and
passive ventillation. Plants sprayed with 35% M-pede soap (Dow AgroSciences LLC) as
needed to control psyllids insects and mites.

52
The D. citri colony was maintained in an air-conditioned rearing room at 28± 2 C,
60-80% RH (Extech RH10 humidity and temperature datalogger, Grainger, Inc), and L:
D= 14: 10. Six M. paniculata plants, each with five to eight, 4 cm long new sprouts were
placed a wood frame cage (60 cm 60 cm 120 cm) covered all three sides with fine
mesh for ventillation. Six hundred D. citri adults were released in each cage for 24 h for
oviposition. All adult psyllids were then collected and removed. Plants were held for
approximately 10 d (depending on temperature and humidity until all nymphs reached
4th instar.
Six M. paniculata plants with 4th instar nymphs were transferred into a clear
acrylic cage (60 cm 80 cm 90 cm) with 3 sides of fine mesh in an air conditioned
greenhouse maintained at 28± 2 C, 75± 5% RH. Fifty T. radiata females and 30 males
were released in the cage for 48 hours to parasitize hosts, then collected and removed.
Plants were held for another 6 d until the parasitized hosts d mummified. Then, the
young shoots with nymphs were excised and nymphs were examined under a
microscope. When a nymph was found parasitized with its parasitoid wasp developed
almost to the pupal stage, that part of the shoot (about 2 cm) was excised and placed in
a glass tube (75 mm long, 12 mm in diameter, Fisher Scientific, Pyrex 9820) in an air-
conditioned rearing room at 28±2 ℃, 60-80% RH (Extech RH10 humidity and
temperature datalogger, Grainger, Inc). Tubes were checked frequently, and once the
wasp removed immediately after emerge.
Diets
Each pair of newly emerged T. radiata was placed in a 50 ml centrifuge tube
(Kendall Labware, Mansfield, MA), sealed with Parafilm (Pechiney Plastic Packaging

53
Company, Chicago, Illinois) and stored in a growth chamber (Percival Scientific, model
I36LLC8,Perry Iowa) at 17 C, 75~ 85% RH , and L: D = 14:10. Wasp were provided
with 8 different diet treatments: water alone, honey, Nu-Lure (Miller Chemical &
Fertilizer Corporation, Hanover, Pennsylvania), host nymphs, honey+ Nu-Lure, honey+
host nymphs, Nu-Lure+ host nymphs, and honey+ Nu-Lure + host nymphs (water stripe
was provided in every treatment). Nu-Lure (Miller Chemical and Fertilizer, Hanover, Pa.)
is a proteinaceous dark brown liquid made from 44% hydrolyzed corn gluten meal, and
56% inert ingredients. Water, honey, and N-Lure were all provided on 8 cm long, 1.5 cm
wide cellulose strips (Wypall-L30, Kimberly-Clark Professional). Host nymphs were
provided on a fresh cut shout, on which 2nd, 3rd, 4th instar nymphs were mixed together.
Water strips were changed every 24 h, and honey, Nu-Lure strips, and host nymphs
were changed every 48 hours to make sure they did not dry out and continued providing
efficient food sources. Wasps in each treatment were held separately for 5, 10, 15, and
20 d. There were 10 replications of every treatment day combination.
Egg Load
T. radiata females under each different treatment were dissected after 5, 10, 15,
and 20 d as well as newly emerged T. radiata under a dissecting microscope (Olympus
SZX 16, Olympus Corporation). Pictures were taken using an Olympus DP 21, and the
number of eggs was recorded.
Statistical Analysis
Data from the eight treatments at each 5 day dissection interval were compared
using Tukey’s Student’s t-test (JMP, SAS Institute Inc. 2012).

54
Results
Newly emerged T. radiata females averaged 4.6 eggs each. Egg loads were
significantly different among the eight treatments at 5 days (F = 11.1702, df = 55, P <
0.0001), 10 days (F = 12.72, df = 40, P < 0.0001), 15 days (F = 7.8989, df = 35, P <
0.0001), and 20 days (F = 10.28, df = 34, P < 0.0001). Wasps fed on water and Nu-Lure
all died within 10 days. At each of the five-day checks, females fed on honey had
significantly fewer eggs compared with the other treatments except for water and Nu-
Lure. During the first five days, females fed on nymphs formed the same number of
eggs as honey + Nu-Lure, but with time , females fed on honey+ Nu-Lure formed fewer
and fewer eggs until the 20th day, when the difference was significant compared to
those fed on nymphs. Females fed on Nu-Lure + nymphs, and honey+ Nu-Lure+
nymphs formed significantly more eggs than those provided with nymphs alone, and a
little more than those on honey + nymphs, although the difference was not significant
(Table 4-1). Wasps fed on honey alone formed fewer and fewer eggs after 10 days;
wasps fed on honey +Nu-Lure also formed gradually fewer eggs
Discussion
An average of 4.6 eggs were observed in ovaries of newly emerged female T.
radiata, despite not feeding on any diets (Fig 4-1, 4-2), so they may use reserved
nutrition from larval stages to form the first clutch of eggs. Mating was not necessary for
egg formation.
This study showed that honey alone was enough to keep wasp females alive, but
egg resorption took place within 5 days after emergence (Fig 4-3). This may mean that
T. radiata females lack the ability to transfer carbohydrates into amino acids to form

55
eggs, or oogenesis requires essential amino acids which cannot be transferred from
carbohydrates.
Nu-Lure alone was not sufficient to keep the females alive for more than 5 days,
which indicated that carbohydrate is a necessary energy resource needed for survival,
and they may lack the capability of transforming amino acids into carbohydrate.
The combination of honey + Nu-Lure resulted in female survivorship similar to a
diet of host nymphs, but egg formation was still less than if provided with nymphs (Fig 4-
4). Females with access to host nymphs (Fig 4-5) matured significantly more eggs than
those fed on honey, Nu-Lure, or honey+ Nu-Lure, indicating that host body fluid
contained essential nutrients not available in the food supplements. However, the
combinations of honey+ Nu-Lure+ nymphs (Fig 4-8) and Nu-Lure+ nymphs (Fig 4-7)
resulted in significantly higher fecundity than a diet of nymphs alone, honey + nymphs
(Fig 4-6) was a little better, but not significant (Table 4-1). I concluded that host nymphs’
body fluid is an irreplaceable nutrition source for females’ egg formation. However, the
extra source of carbohydrates and amino acids provided by the supplements had a
positive effect both in the absence and presence of nymphs. In future studies, it will be
interesting to investigate which amino acids or other constituents of hemolymph are
important for egg formation.
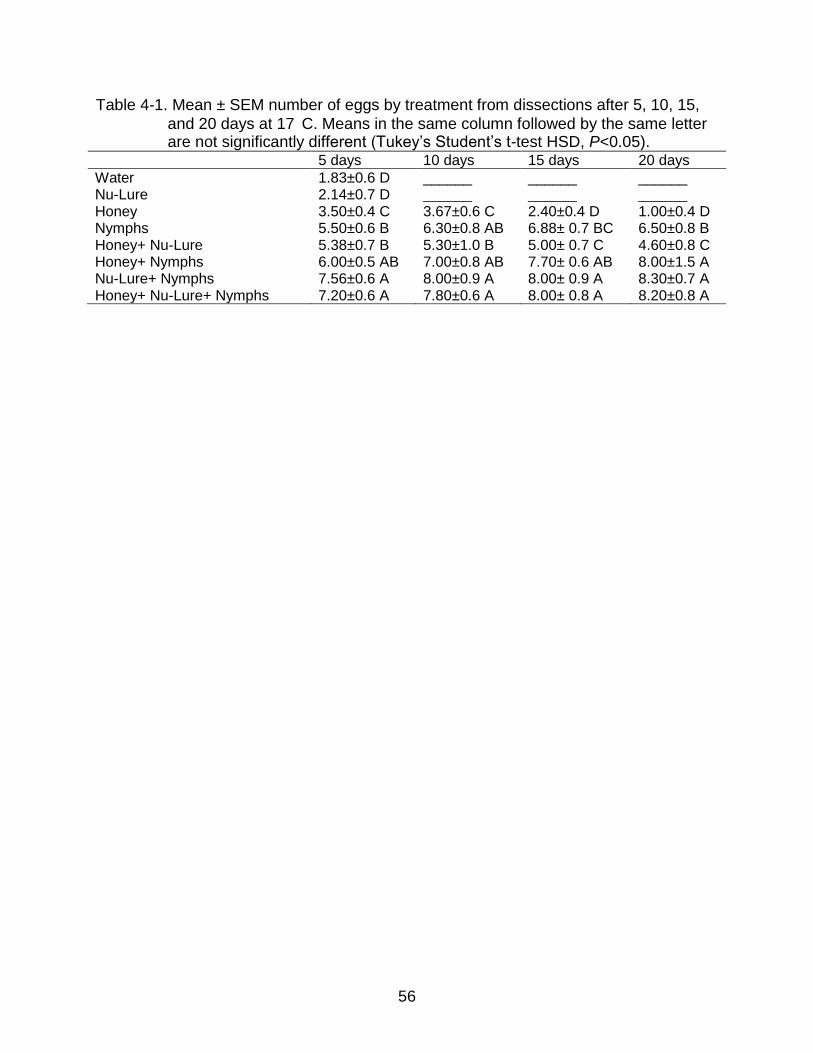
56
Table 4-1. Mean ± SEM number of eggs by treatment from dissections after 5, 10, 15,
and 20 days at 17 C. Means in the same column followed by the same letter are not significantly different (Tukey’s Student’s t-test HSD, P<0.05).
5 days 10 days 15 days 20 days
Water 1.83±0.6 D ______ ______ ______ Nu-Lure 2.14±0.7 D ______ ______ ______ Honey 3.50±0.4 C 3.67±0.6 C 2.40±0.4 D 1.00±0.4 D Nymphs 5.50±0.6 B 6.30±0.8 AB 6.88± 0.7 BC 6.50±0.8 B Honey+ Nu-Lure 5.38±0.7 B 5.30±1.0 B 5.00± 0.7 C 4.60±0.8 C Honey+ Nymphs 6.00±0.5 AB 7.00±0.8 AB 7.70± 0.6 AB 8.00±1.5 A Nu-Lure+ Nymphs 7.56±0.6 A 8.00±0.9 A 8.00± 0.9 A 8.30±0.7 A Honey+ Nu-Lure+ Nymphs 7.20±0.6 A 7.80±0.6 A 8.00± 0.8 A 8.20±0.8 A

57
Figure 4-1. Newly emerged (unfed) T. radiata female digestive system (Photo courtesy of Xulin Chen)
Figure 4-2. Newly emerged (unfed) T. radiata female (Photo courtesy of Xulin Chen)
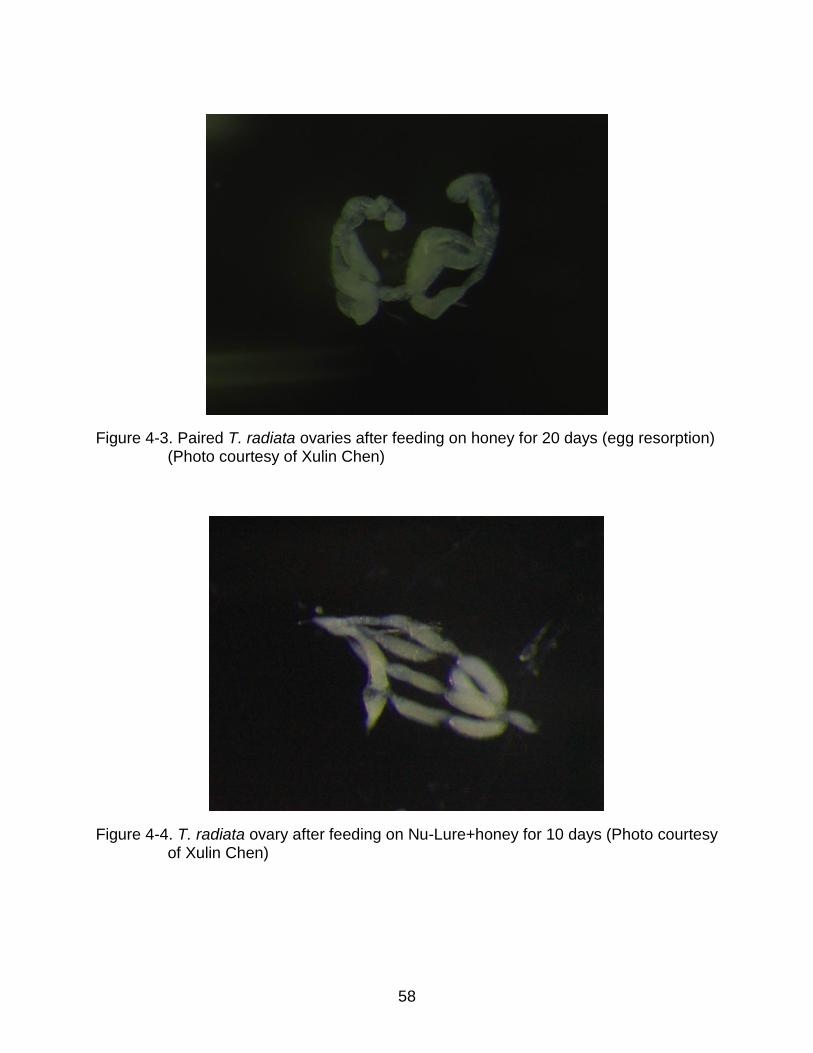
58
Figure 4-3. Paired T. radiata ovaries after feeding on honey for 20 days (egg resorption) (Photo courtesy of Xulin Chen)
Figure 4-4. T. radiata ovary after feeding on Nu-Lure+honey for 10 days (Photo courtesy of Xulin Chen)

59
Figure 4-5. Paired T. radiata ovaries after feeding on nymphs for 5 days (Photo courtesy of Xulin Chen)
Figure 4-6. Paired T. radiata ovaries after feeding on honey +nymphs for 10 days (Photo courtesy of Xulin Chen)
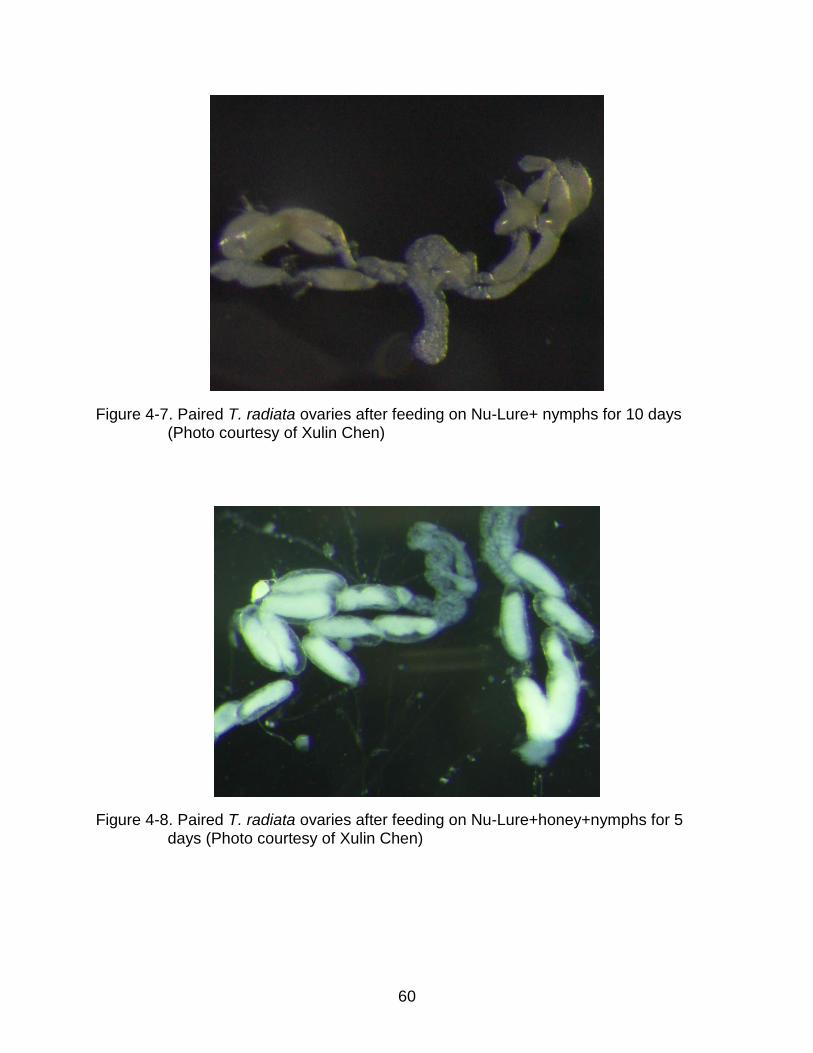
60
Figure 4-7. Paired T. radiata ovaries after feeding on Nu-Lure+ nymphs for 10 days (Photo courtesy of Xulin Chen)
Figure 4-8. Paired T. radiata ovaries after feeding on Nu-Lure+honey+nymphs for 5 days (Photo courtesy of Xulin Chen)

61
CHAPTER 5 DISCUSSION AND CONCLUSIONS
Tamarixia radiata, is an arrhenotokous ectoparasite of the Asian citrus psyllid
(ACP) Diaphorina citri, vector of citrus greening disease or huanglongbing (HLB).
Tamarixia radiata is being tried as an augmentive biological control agent, since the
number present in the field reduced in winter and by insecticide applications. Any
program aimed at augmentation of a natural enemy would require an efficient system of
mass production. Key components would include methods of manipulating, rearing and
storing large populations of insects.
To make counting easily and separating D. citri and T. radiata after collection,
carbon dioxide was used as anesthesia. Carbon dioxide anesthesia can be a
convenient tool for manipulating insects, but can also cause deleterious side effects. In
this study, a 5 min exposure of Tamarixia radiata adults to 100% CO2 concentration
caused a knockdown of about 4 min, and significantly reduced survivorship and
fecundity, but did not affect the sex ratio of progeny from treated adults.
The harmful effects of CO2 proscribe its routine under the tested exposure
conditions for rearing T. radiate. Nevertheless the wasp showed surprising tolerance to
high CO2 concentration: the wasps acted normally when sealed in a gas chamber with
30% CO2. Perhaps lower concentrations and/or exposure times would provide more
acceptable alternatives. Also, the physiological mechanism of CO2 catabolism could be
an interesting research aspect. The effects of chilling to immobilize the insects should
also be tested.
Host density is an important factor influencing mass rearing efficiency. Fourth
and early 5th instars are the preferred stage for oviposition by T. radiata. My study

62
showed that a density 40, 4th instar nymphs per female optimized fecundity, maximized
incidence of parasitism and minimized incidence of superparasitism. This result could
be used to determine the idea number of parasitoids to be released into a rearing cage.
The number of flushes in the rearing cage can be counted, and the number of proper
stage nymphs on one shoot can be estimated to calculate the number of nymphs in the
whole rearing cage. Then, knowing the sex ratio and the ideal 4th instar host to female
ratio of 1:40, the best number of T. radiata females to release will be clear.
The pattern of parasitism observed over the first five days conformed to a Type II
functional response, estimated searching efficiency was 0.442 ± 0.036 per day and
estimated handing time was 0.045 ± 0.008 days. These two parameters are important in
comparing a T. radiata female’s parasitism efficiency with other parasitoids, to evaluate
its parasitic capacity.
When T. radiata were stored for release, certain nutritional food is necessary to
maintain fecundity. My results showed that, T. radiata formed more eggs feeding on
mixed diets (Nu-Lure+ honey+ nymphs or Nu-Lure+ nymphs) compared to nymphs
alone. Until now, no diet has been found to completely substitute nymphal hemolymph.
The present practice is to hold wasps in centrifuge tubes provided with honey on
paper strips preparatory to release. My research would indicate that wasps should also
be provided with a supplementary protein source such as Nu-Lure, and also with hosts
in order to maintain the fecundity.
Tamaraxia radiata is an efficient biological control agent of D. citri under optimal
temperature, humidity, host densities and nutritional supplement in lab conditions. More

63
work need to be done to apply these lab results in field to get a more optimal parasitism
capacity of D. citri.

64
LIST OF REFERENCES
ANONYMOUS. 2004. Estudos indicam que a nova doença tem relação com o greening. Fundecitrus. http://www.fundecitrus.com.
AUBERT, B., GRISONI, M., VILLEMIN, M., ROSSOLIN, G., DA GRACA, J. V., MORENO, P., AND
YOKOMI, R.K. 1996. A case study of Huanglongbing (greening) control in Reunion. Proceedings of the 13th Conference of the International Organization of Citrus Virologists, p. 276-278.
AUBERT, B., AND QUILICI, S. 1984. Biological control of the African and Asian citrus psyllids (Homoptera : Psylloidea) through eulophid and encyrtid parasites (Hymenoptera: Chalcidoidea) in Reunion Island. Proceedings from the 9th Conference of the International Organization of Citrus Virologists, p.100-108.
AUBERT, B., AND HUA, X. Y. 1990. Monitoring flight activity of Diaphorina citri on citrus and Murraya canopies, Rehabilitation of citrus industry in the Asia Pacific Region. Proc. 4th International Asia Pacific Conference on Citrus Rehabilitation, Chiang Mai, Thailand, pp. 181-187.
AUBERT, B., AND QUILICI, S. 1988. Monitoring adult psyllas on yellow traps in Reunion Island. Proceedings of the 10th Conference of International Organization of Citrus Virologists. International Organization of Citrus Virologists, Riverside, CA, pp. 249-254.
BATOOL, A., IFTIKHAR, Y., MUGHAL, S. M., KHAN, M. M., JASKANI, M. J., ABBAS, M., AND
KHAN. I. A. 2007. Citrus greening disease a major cause of citrus decline in the worlda review. Hort. Sci. 34: 159-166.
BOVÉ, J. M. 2005. First report of a huanglongbing-like disease of citrus in São Paulo State, Brazil and association of a new Liberibacter species, “Candidatus Liberibacter americanus”, with the disease. Plant Dis. 89: 107.
BOVÉ, J. M. 2006. Huanglongbing: a destructive, newly emerging, century-old disease of citrus. J. Plant Pathol. 88: 7-37.
BOWNES, M., AND BLAIR, M. 1986. The effects of a sugar diet and hormones on the expression of the drosophila yolk-protein genes. J.Insect Physiol. 32: 493-501.
BROOKS, M. A. 1957. Growth-retarding effect of carbon-dioxide anaesthesia on the German cockroach. J. Insect Physiol. 1: 76-84.
BROOKS, M. A. 1965. The effects of repeated anesthesia on the biology of Blattella germanica (Linnaeus.). Entomol. Exp. Appl. 8: 39-48.
BUITENDAG, C. H., AND BROEMBSEN, L. A. 1993. Living with citrus greening in South Africa. Citrus J. 3: 29-32.

65
CAPOOR, S. P., RAO, D. G., AND VISWANATH, S. M. 1974. Greening disease of citrus in the Deccan Trap Country and its relationship with the vector, Diaphorina citri Kuwayama. Proceedings of the 6th Conference of the International Organization of Citrus Virologists, Swaziland. Division of Agricultural Science, University of California, Berkeley, CA. pp. 43-49.
CHIEN, C. C., CHIU, S. C., AND KU, S. C. 1989. Biological control of Diaphorina citri in Taiwan. Fruits (Paris) 44: 401-407.
CHIEN, C. C., CHU, Y. I., AND KU, S. C. 1991a. Parasitic strategy, morphology and life history of Tamarixia radiata (Hymenoptera: Eulophidae). Chinese Insect 11: 264-281. (In Chinese).
CHIEN, C. C., CHU, Y. I., AND KU, S. C. 1991b. Biological control of citrus psyllid, Diaphorina citri in Taiwan. Evaluation of Tamarixia radiata and Diaphorencyrtus diaphorinae for the control of Diaphorina citri. Chinese Insect 11: 25-38. (In Chinese).
CHIEN, C. C., CHU, Y. I., AND KU, S. C. 1993. Influence of temperature on the population increase, host killing capacity and the storage of the Tamarixia radiata. Chinese Insect 13: 111-123. (In Chinese).
CHIEN, C. C., CHU, Y. I., AND KU, S. C. 1994a. Oosorption and ovopisition regulating capability of the Eulophid wasp, Tamarixia radiata and its internal reproductive organs. Plant Prot. Bull. 36: 97-105. (In Chinese).
CHIEN, C. C., CHU, Y. I., AND KU, S. C. 1994b. Influence of food on longevity, egg production and population increase of the Eulophid wasp, Tamarixia radiata. Plant Prot. Bull. 36: 97-105. (In Chinese).
CHIEN, C. C., CHU, Y. I., AND KU, S. C. 1995. Influence of host densities on the population increase of the Eulophid wasp, Tamarixia radiata, and its host killing ability. Plant Prot. Bull. 37: 81-96. (In Chinese).
CHIEN, C. C., AND CHU, Y. I. 1996. Biological control of citrus psyllid, Diaphorina citri in Taiwan. Biological pest control in systems of integrated pest management. Proceedings of the International Symposium on "The use of Biological Control Agents under Integrated Pest Management". 93-105.
CHU, Y. I., AND CHIEN, C. C. 1991. Utilization of natural enemies to control psyllid vectors transmitting citrus greening. Integrated control of plant virus diseases. Proceedings of the International Workshop TARI, Taichung, Taiwan, April 9- 14, 1990: 135- 145.
COLETTA-FILHO, H. D., TARGON, M. L. P. N., TAKITA, M. A., DE NEGRI, J. D., POMPEU, J., MACHADO, M. A., AMARAL, A. M., AND MULLER, G. W. 2004. First report of the causal agent of Huanglongbing (“Candidatus Liberibacter asiaticus”) in Brazil. Plant Dis. 88: 1382.

66
CRYSTAL, M. M. 1967. Carbon dioxide anesthesia of untreated and chemosterilant-treated screw-worm flies, Cochliomyia hominivorax (Coquerel) (Diptera:Calliphoridae). J. Med. Entomol. 4: 415-418.
ÉTIENNE, J., QUILICI, S., MARIVAL, D., AND FRANCK, A. 2001. Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits 56: 307-315.
FRENCH, J. V., KAHLKE, C. J., AND DA GRAÇA, J. V. 2001. First record of the Asian citrus psylla, Diaphorina citri Kuwayama (Homoptera: Psyllidae), in Texas. Subtropical Plant Sci. 53: 14-15.
FUNG, Y. C., AND CHEN, C. N. 2006. Effects of temperature and host plant on population parameters of the citrus psyllid (Diaphorina citri Kuwayama). Formosan Entomol. 26: 109-123.
GARNIER, M., AND BOVÉ, J. M. 1993. Citrus greening disease and the greening bacterium. Proc. of the 12th Conf. Intl. Organ. Citrus Virologists. P. Moreno, J. V. da Graça, and L. W. Timmer, eds. IOCV, Riverside, CA. P 212-219.
GARNIER, M., DANEL, N., AND BOVÉ, J. M. 1984. Transmission of citrus greening disease from sweet orange to periwinkle (Vinca rosea) by dodder (Cuscuta campestris) and proof of the bacterial nature of the organism. Yale J. Biol. Med. 57: 904-904.
GELMAN, D., GERLING, B. D., AND BLACKBURN, M. A. 2005. Host-parasitoid interactions relating to penetration of the whitefly, Bemisia tabaci, by the parasitoid wasp, Eretmocerus mundus. J. Insect Sci. 5: 46.
GOTTWALD, T. R., GRAÇA, J. V., AND BASSANEZI, R. B. 2007. Citrus Huanglongbing: The Pathogen and Its Impact http://www.apsnet.org/publications/apsnetfeatures/Pages/HuanglongbingImpact.aspx.
GRAFTON-CARDWELL, E. E., STELINSKI, L. L., AND STANSLY, P. A. 2013. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 58: 413-32.
HALBERT, S. E. 2005. The discovery of huanglongbing in Florida. In Proceedings of the 2nd International Citrus Canker and Huanglongbing Research Workshop. 2005. p: 1-6.
HALBERT, S. E. 2006. Asian citrus psyllid - A serious exotic pest of FL citrus. http://www.doacs.state.fl.us/pi/enpp/ento/dcitri.htm.
HALBERT, S. E. 2010. Pest alert- Citrus greening/Huanglongbing. http://swfrec.ifas.ufl.edu/hlb/database/pdf/00002355.pdf

67
HALBERT, S. E., AND NÚÑEZ, C. A. 2004. Distribution of the Asian citrus psyllid, Diaphorina citri Kuwayama (Rhynchota: Psyllidae) in the Caribbean basin. Florida Entomol. 87(3): 401-402.
HALBERT, S. E., AND MANJUNATH, K. L. 2004. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: a literature review and assessment of risk in Florida. Florida Entomol. 87: 330-353.
HALBERT, S. E., SUN, X., AND DIXON, W. 1998. Current Florida Update on Asian citrus psyllid http://swfrec.ifas.ufl.edu/hlb/database/pdf/00001807.pdf (18 November 1998).
HALBERT, S. E., SUN, X., AND DIXON, W. 2001. Asian citrus psyllid update. http:///www.freshfromflorida.com/pi/enpp/ento/asian-citrus-psyllid.htm (14 October 2002).
HALL, D. G. 2008a. Biology, history and world status of Diaphorina citri. North American Plant Protection Organization Workshop, 2008. p: 1-11.
HALL, D. G. 2008b. Biological control of Diaphorina citri. North American Plant Protection Organization Workshop, 7- 9 May (2008). p: 1-7.
HALL, D. G., AND ALBRIGO, L. G. 2007. Estimating the relative abundance of flush shoots in citrus, with implications on monitoring insects associated with flush. Hort. Science 42: 364-368.
HODDLE, M. S. 2012. Foreign exploration for natural enemies of Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae), in the Punjab of Pakistan for use in a classical biological control program in California USA. Pakistan Entomol. 34: 1-5.
HOLLING, C. S. 1959. Some characteristics of simple types of predation and parasitism. Canadian Entomol. 91: 385-398.
HOOPER, G. H. S. 1970. Use of carbon dioxide, nitrogen and cold to immobilize adults of the Mediterranean fruit fly. J. Econ. Entomol. 63: 1962-1963.
HOWARD, L. O. 1910. On the habit with certain Chalcidoidea of feeding at puncture holes made by the ovipositor. J. Econ. Entomol. 3: 257-260.
HOY, M., AND NGUYEN, R. 2001. Long PCR is a sensitive method for detecting Liberobacter asiaticum in parasitoids undergoing risk assessment in quarantine. Biol. Control. 22: 278-287.
HUNG, T. H., HUNG, S. C., CHEN, C. N., HSU, M. H., AND SU, H. J. 2004. Detection by PCR of Candidatus Liberibacter asiaticus, the bacterium causing citrus huanglongbing in vector psyllids: application to the study of vector-pathogen relationships. Plant Pathol. 53: 96-102.

68
HUSAIN, M. A., AND NATH, D. 1923. The Life-history of Tetrastichus radiatus parasitic on Euphalerm citri Kuw. and its hyperparasite. Report from the Proceedings of the 5th Entomological Meeting, Pusa, February 1923 (1924), p.122-128.
HUSAIN, M. A., AND NATH, D. 1927. The citrus psylla (Diaphorina citri, Kuw.) [Psyllidae: Homoptera]. Mem. Dept. Agric. India, Entomol. Ser. 10(2): 27.
JAGOUEIX, S., BOVÉ, J. M., AND GARNIER, M. 1994. The phloem-limited bacterium of greening disease of citrus is a member of alpha subdivision of Proteobacteria. Int. J. Syst. Bacteriol. 44: 379-386.
JAGOUEIX, S., BOVÉ, J. M., AND GARNIER, M. 1996. PCR detection of the Candidatus Liberibacter species associated with greening disease of citrus. Mol. Cell Probes10: 43-50.
JONES, D. B., GILES, K. L., BERBERET, R. C. , ROYER, T. A., ELLIOTT, N. C., AND PAYTON, M. E. 2003. Functional responses of an introduced parasitoid and an indigenous parasitoid on greenbug at four temperatures. Environ. Entomol. 32: 425-432.
JUAN-BLASCO, M., QURESHI, J. A., URBANEJA, A., AND STANSLY, P. A. 2012. Predatory mite, Amblyseius swirskii (Acari: Phytoseiidae), for biological control of Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Florida Entomol. 95:543-551.
JULIANO, S. A. Nonlinear curve fitting. Predation and functional response curve. In: Scheiner SM, Gurevitch J (eds) Design and analysis of ecological experiments. 2nd edn. Oxford University Press, New York, pp 178-196.
LEIUS, K. 1961. Influence of food on fecundity and longevity of adults of Itoplectis conquisitor (Say) (Hymenoptera: Ichneumonidae). Canadian Entomol. 93: 771-780.
LIMA, A. C. 1942. Insetos do Brasil, Homopteros. Serie didatica Escola Nacional de Agronomia 3: 327.
LIU, Y. H., AND TSAI, J. H. 2000. Effects of temperature on biology and life table parameters of the Asian citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Ann. Appl. Biol. 137: 201-206.
MANJUNATH, K. L., HALBERT, S. E., RAMADUGU, C., WEBB, S., AND LEE, R. F. 2008. Detection of Candidatus Liberibacter asiaticus in Diaphorina citri and its importance in the management of citrus huanglongbing in Florida. Phytopathology 98: 387-396.
MANN, R. S., QURESHI, J. A., STANSLY, P. A., AND STELINSKI, L. L. 2010. Behavioral Response of Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae) to Volatiles Emanating from Diaphorina citri Kuwayama (Hemiptera: Psyllidae) and Citrus. J. Insect Behav. 23: 447-458.

69
MASSONIE, G., GARNIER, M., AND BOVÉ, J. M. 1976. Transmission of Indian greening by Trioza erytreae (Del Guercio), the vector of South African greening. Pages 18-20 in: Proc. of the 7th Conf. Intl. Organ. Citrus Virol. E. C. Calavan, ed. IOCV, Riverside, CA.
MCFARLAND, C. D., AND HOY, M. A. 2001. Survival of Diaphorina citri (Homoptera: Psyllidae) and its two parasitoids, Tamarixia radiata (Hymenoptera: Eulophidae) and Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae), under different relative humidities and temperature regimes. Florida Entomol. 84: 227-233.
MEAD, F.W. 1977. The Asiatic citrus psyllid, Diaphorina citri Kuwayama (Homoptera: Psyllidae). Entomol.Circ. 180. Florida Dept. Agric. Consum. Serv. Plant Industry Ind. 4 pp.
MEAD, F. W., AND FASULO, T. R. 2010. Asian Citrus Psyllid, Diaphorina citri Kuwayama (Insecta: Hemiptera: Psyllidae). Florida Coop. Extn. Serv., Inst. Food Agric. Sci., Univ. Florida. Gainesville, FL, EENY-033 (2010), p.1-8.
MERCET, R. G. 1924. Eulophidos de Espana (1ª nota) Bol. Soc. R. Española Hist. Nat. 24: 57-59.
MICHAUD, J. P. 2004. Natural mortality of Asian citrus psyllid (Homoptera : Psyllidae) in central Florida. Biol. Control 29: 260-269.
MOLL, J. N., AND VAN VUUREN., S. P. 1977. Greening disease in Africa. Proc. Int. Soc. Citriculture 3: 903-912.
MORRIS, R. A., ERICK, C., AND ESTES, M. 2009a. Greening infection at 1.6%, survey to estimate the rate of greening and canker infection in Florida citrus groves. Citrus Ind. 90: 16-18.
MORRIS, R. A., ERICK, C., AND ESTES, M. 2009b. The incidence of greening and canker infection in Florida citrus groves from September 2007 through August 2008 (EDIS document FE823). University of Florida, Institute of Food and Agricultural Sciences, Gainesville, FL.
NAVA, D. E., TORRES, M. L. G., RODRIGUES, M. D. L., BENTO, J. M. S., AND PARRA, J. R. P. 2007. Biology of Diaphorina citri (Hemiptera, Psyllidae) on different hosts and at different temperatures. J. Appl. Entomol. 131: 709-715.
PAIVA, P. E. B., AND PARRA, J. R. P. 2012. Life table analysis of Diaphorina citri (Hemiptera: Psyllidae) infesting sweet orange (Citrus sinensis) in São Paulo. Florida Entomol. 95 (2): 278-284.
PLUKE, R. W. H., QURESHI, J. A., AND STANSLY, P. A. 2008. Citrus flushing patterns, Diaphorina citri (Hemiptera : Psyllidae) populations and parasitism by Tamarixia radiata (Hymenoptera : Eulophidae) in Puerto Rico. Florida Entomol. 91: 36-42.

70
PELZ-STELINSKI, K. S., BRLANSKY, R. H., EBERT, T. A., AND ROGERS, M. E. 2010. Transmission parameters for Candidatus Liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae). J. Econ. Entomol. 103: 1531-1541.
PRUTHI, H. S. AND BATRA, H. N. 1938. A preliminary annotated list of fruit pests of the North-West Frontier Province. Misc. Bull. Imp. Council Agric. Res. India 19: 10-12.
QUILICI, S., AND FAUVERGUE, X. 1990. Studies on the biology of Tamarixia radiata Waterston Hymenoptera: Eulophidae. Proceedings of the 4th International Asia-Pacific Conference on Citrus Rehabilitation 4-10 February (1990), p. 251-256.
QURESHI, J. A., AND STANSLY, P. A. 2007. Integrated approaches for managing the Asian citrus psyllid Diaphorina citri (Homoptera: Psyllidae) in Florida. Proc. Florida State Hort. Soc. 120: 110–115.
QURESHI, J. A., AND STANSLY, P. A. 2009. Exclusion techniques reveal significant biotic mortality suffered by Asian citrus psyllid Diaphorina citri (Hemiptera: Psyllidae) populations in Florida citrus. Biol. Control 50: 129-136.
ROCKWOOD, L. P. 1917. An aphis parasite feeding at puncture holes made by the ovipositor. J. Econ Entomol 10: 415.
ROHRIG, E., SHIRK, P. D., HALL, D. G., AND STANSLY, P. A. 2011. Larval Development of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae), an Endoparasitoid of Diaphorina citri (Hemiptera: Psyllidae). Ann. Entomol. Soc. America 104: 50-58.
ROISTACHER, C. N., DA GRAÇA., J. V., MORENO., P., AND YOKOMI, R. K. 1996. The economics of living with citrus diseases: Huanglongbing (greening) in Thailand. Proceedings of the 13th Conference of the International Organization of Citrus Virologists (IOCV) (1996), p.279-285.
RUPPEL, R. F. 1983. Cumulative insect-days as an index of crop protection. J. Econ. Entomol. 76: 375-377.
SAGARAM, U. M., DEANGELIS, K. M., TRIVEDI, P., ANDERSEN, G. L., LU, S. AND WANG. N. 2009. Bacterial diversity analysis of huanglongbing pathogen-infected citrus, using phylochip arrays and 16s rRNA gene clone library sequencing. Appl. Environ. Microbiol. 75: 1566-1574.
SKELLEY, L. H., AND HOY, M. A. 2004. A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine. Biol. Control 29: 14-23.
TANG, Y. Q. 1990. On the parasite complex of Diaphorina citri Kuwayama (Homoptera: Psyllidae) in Asian-Pacific and other areas. Proceedings of the 4th International Conference on Citrus Rehabilitation: Proceedings of the Asia Pacific International Conference on Citriculture, 4-10 February (1990), p.240-245.

71
TANG, Y. Q., AND AUBERT, B. 1990. An illustrated guide to the identification of parasitic wasps associated with Diaphorina citri Kuwayama in the Asian-Pacific region. Rehabilitation of Citrus Industry in the Asia Pacific Region: Proceedings of the 4th International Conference on Citrus Rehabilitation, 4- 10 February (1990), p.228-239.
TANG, Y. Q. AND WU, M. X. 1991. Interspecific host discrimination between two primary parasites of Asian citrus psyllid Diaphorina citri (Kuwayama). Proceedings of the 6th International Asia Pacific Workshop on Integrated Citrus Health Management, 24-30 June (1991), p.99-104.
TANG, Y. Q., AND HUANG, Z. P. 1991. Studies on the biology of two primary parasites of Diaphorina citri Kuwayama (Homoptera: Psyllidae) Proceedings of the 6th International Asia Pacific Workshop on Integrated Citrus Health Management, 24-30 June (1991), p.91-98.
TEIXEIRA, D. D., SAILLARD, C., EVEILLARD, S., DANET, J. L., DA COSTA, P. I., AYRES, A. J., AND BOVÉ, J. M. 2005. Candidatus Liberibacter americanus, associated with citrus huanglongbing (greening disease) in São Paulo State, Brazil. Int. J. Syst. Evol. Microbiol. 55: 1857-1862.
TSAI, J. H., AND LIU, Y. H. 2000. Biology of Diaphorina citri (Homoptera: Psyllidae) on four host plants. J. Econ. Entomol. 93 (6): 1721-1725.
TSAI, J. H., WANG, J. J., AND LIU, Y. H. 2002. Seasonal abundance of the Asian citrus psyllid, Diaphorina citri (Homoptera : Psyllidae) in southern Florida. Fla. Entomol 85: 446-451.
VARLEY, G. C., AND EDWARDS, R. L. 1957. The bearing of parasite behavior on the dynamics of insect host and parasite populations. J. Anim. Ecol. 26: 471-477.
WALTER, A. J., HALL, D. G., AND DUAN, Y. P. 2012. Low Incidence of 'Candidatus Liberibacter asiaticus' in Murraya paniculata and Associated Diaphorina citri. Plant Dis. 96: 827-832.
WANG, Z., YIN, Y., HU, H., YUAN, Q., PENG, G., AND XIA, Y. 2006. Development and application of molecular-based diagnosis for Candidatus Liberibacter asiaticus, the causal pathogen of citrus huanglongbing. Plant Pathol. 55: 630-638.
WATERSTON, J. 1922. On the chalcidoid parasites of psyllids (Hemiptera, Homoptera). Bull. Entomol. Res. 13: 41-58.
WENNINGER, E. J., AND HALL, D.G. 2007. Daily timing and age at mating in the Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Florida Entomol. 90: 715-722.
WHEELER, D. 1996. The role of nourishment in oogenesis. Annu. Rev. Entomol. 41: 407-431.

72
XU, C. F., XIA, Y. H., LI, K. B. AND KE, C. 1988. Further study of the transmission of citrus huanglongbing by a psyllid, Diaphorina citri Kuwayama. In: Timmer, L. W., Garnsey, S. M., and L. Navarro (eds.), Proceedings of the 9th Conference of International Organization of Citrus Virologists. International Organization of Citrus Virologists, Valencia, Spain, pp. 100-108.
XU, J. H., AND TANG, Y. Q. 1993. The immature development and morphology of Tamarixia radiata. Fujian Agric. Dept.Acad. J. 22: 311-316. (In Chinese).
YANG, Y. B. 1989. Influence of light, temperature and humidity on the development reproduction and survival of citrus psyllids. Chinese J. Ecol. 9(4): 126-133.

73
BIOGRAPHICAL SKETCH
Xulin Chen was born in Shandong Province, China. She began her
undergraduate study in Shandong Agriculture University majoring in Plant Quarantine in
2007. After she graduated in July, 2011, she started her graduate study in University of
Florida, Entomology and Nematology Department under the supervision of Dr. Phil
Stansly.