links between macroalgal mats, fauna and sediment - gu.se · since the main nutrient source for...
TRANSCRIPT

Links between macroalgal mats, fauna and sediment biogeochemistry
Anna Engelsen
Doctoral Thesis in Marine Ecology
Göteborgs University
Faculty of Science
Department of Marine Ecology
Kristineberg Marine Research station
450 34 Fiskebäckskil Sweden
Akademisk avhandling för filosofie doktorsexamen i Marin Ekologi vid Göteborgs Universitet. Avhandlingen kommer att offentligt försvaras fredagen den 30:e maj 2008, kl. 10.00 i hörsalen, Institutionen för Marin Ekologi, Göteborgs Universitet, Kristinebergs marina forskningsstation, Fiskebäckskil. Examinator: Per Jonsson Fakultetsopponent: Associate Professor, PhD, Marianne Holmer, Biologisk Institut, Syddansk Universitet, Campusvej 55, 5230 Odense

2
Published by the Department of Marine Ecology, Göteborg University © Anna Engelsen, 2008 ISBN: 91-89677-40-4

3
Abstract Fast growing ephemeral green algal mats have become much more common in coastal areas during later decades. It is suggested that shallow-water sediments have been accumulating nutrients for decades and thereby have become a source of nutrients, and now function as a self-regenerating systems during the initiation and growth of ephemeral green algal mats mainly consisting of Ulva spp. Since the main nutrient source for algal mats is suggested to be the sediment, the coupling between benthic inorganic nutrient fluxes and sediment properties were investigated. Also experiments which focused on the role of trophic status (balance between auto - and heterotrophy) as a controlling factor for rates of benthic nutrient fluxes, and the nutrient demand of green algal mats were performed. In addition the grazing effect by Nereis diversicolor on green algal mats and links between grazing, nutrient supply and growth of algal mats were investigated. The results showed that generally, sediments rendered autotrophic by microphytobenthic photosynthesis removed nutrients from the overlying water, while heterotrophic sediments supplied nutrients to the overlying bottom water. Estimations of the green-algal nutrient demand suggested that late in the growth season, nutrient fluxes from net heterotrophic sediments could cover 20 % of the N-demand and 70 % of the P demand. As the benthic trophic status is a functional variable more closely coupled to nutrient fluxes than the comparably conservative structural parameter organic matter content, it is suggested that the trophic status is a more viable parameter to classify sediments and predict benthic nutrient fluxes in shallow-water environments. The results showed that macroalgae grew well without the underlying sediment, where the only supply of nutrients came from the water column. The establishment of a green algal mat was not obvious since cyanobacteria showed to be excellent competitors for inorganic nutrients. Moreover, grazing by N. diversicolor was able to prevent initial algal growth, affect growth capacity and also partly reduce fully - grown algal mats. In addition, the individual weight of N. diversicolor was significantly increased for polychaetes feeding on macroalgae compared to those which had not. There were also indications that a rich nutrient supply per algal biomass counteracted the grazing capacity of N. diversicolor. This was reinforced when the algal mat started to grow after oxygen deficiency due to a combination of higher nutrient release and lack of grazing. Key words: Green algal mats, sediment, trophic state, microphytobenthos, nutrients, macrofauna, grazing, nutrient demand, oxygen deficiency

4

5
Populärvetenskaplig sammanfattning Grönalgsmattor finns över hela världen och utgör ett ekologiskt problem sedan några decennier därför att en algmatta dels konkurrerar ut andra perenna primärproducenter, dels ändrar faunans ekologi, men även utgör ett hot mot kommersiellt viktiga fiskarter som har sin lek och födoplats i grunda havs vikar. Makroalgsmattor växer i grunda (0-1 m) skyddade vikar där vind och vågor har liten inverkan på algmattan. Algmattorna i denna undersökning består till största delen av fintrådiga Ulva spp. som växer till och bildar tjocka mattor vilka kan täcka stora delar av en vik. Algerna har en stor yta i förhållande till sin volym vilket innebär att de snabbare än andra alger kan ta upp näringen ur vattnet som i sin tur leder till en stor konkurrensfördel gentemot exempelvis mikroalger och perenna makroalger. När en matta börjar brytas ner åtgår syre vilket innebär att faunan som lever i viken kan får svårt att klara sig på grund av syrebrist. En grund vik som är skyddad för vind och vågor är också födoplats samt lekplats för bl.a. kommersiella fiskarter, vilka får problem när vikarna växer igen. Ett sediment kan vara autotroft (avger syre och tar upp näring) eller heterotroft (tar upp syre och avger näring). Resultaten i den första undersökningen (uppsats I) visar att trofistatusen är en bättre indikator än organiskt innehåll i sedimentet att använda när man skall förutse näringsflöden mellan sedimentet och vattenpelaren. Tidigare studier har diskuterat och föreslagit att de grunda vikarna fungerar som självgenererande system när det gäller näring och algmattor. Sedimentet skall ha ackumulerat näring genom decennier som nu cirkulerar mellan algmattorna och sedimentet. När algmattan blir gammal bryts den ner och närsalter lagras i sedimentet som senare tas upp vid tillväxt av framtida algmattor. I uppsats I visar resultatet att algernas närsaltsbehov är betydligt större än vad sedimentet kan avge. Sedimentet bidrar endast med 20 % av kvävebehovet och 70 % av fosforbehovet hos algerna och slutsatsen blir att närsalter alltså till stor del måste komma från en annan källa. För att verkligen undersöka sedimentets roll som närsaltskälla gjordes ett experiment med olika sedimenttyper och olika sedimentdjup (uppsats II) och resultatet visade att sedimentet inte var avgörande för algmattornas tillväxt. Det indikerade också att havsborstmasken Nereis diversicolor är en effektiv betare på grönalgsmattor. Detta experiment bekräftade också att konkurrensen från bottenlevande mikroalger var avgörande under initieringen av algmattan. Det tredje experimentet (uppsats III) skulle visa i vilken omfattning N. diversicolor kunde påverka tillväxten av algmattor. Resultatet var entydigt och slutsatsen blev att betningen av N. diversicolor kunde förhindra tillväxt samt reducera stora algmattor. I detta experiment fanns en indikation på att betningen eventuellt kunde bero på närsaltstillgången för algmattan. En större mängd närsalter per gram alger verkade betyda en mindre betningseffekt. Experimentet visade också att de N. diversicolor som hade betat på algmattan ökade sin vikt betydligt mer än de som ej hade tillgång till alger som föda. Det fjärde experimentet (uppsats IV) bekräftade till viss del att näringstillgången spelade en roll för betningseffekten. När algmattan blir stor och nedbrytningen blir hög

6
används större mängder syre och resultatet blir syrebrist. I experimentet framkom att betningseffekten av N. diversicolor minskade eller avtog helt under syrebristen. När det blir syrebrist påverkas sedimentets kemi så att större mängder närsalter avges, främst ammonium och fosfat. Detta tillsammans med minskad betning hos N. diversicolor gynnar nybildandet av en algmatta.

7
Till Christian och Simen

8

9
Table of contents Part A 1 Introduction ............................................................................................................. 11
1.1 Shallow marine sediment habitats.................................................................... 11 1.2 Macroalgal mats ............................................................................................... 12 1.3 Inorganic nutrients and microphytobenthos..................................................... 13 1.4 Algal mats and benthic macrofauna ................................................................. 14 1.5 Nereis diversicolor ........................................................................................... 15
2 Aims of the thesis..................................................................................................... 17 3 Methods .................................................................................................................... 18
3.1 Study area......................................................................................................... 18 3.2 Experimental approach and methodological considerations............................ 19 3.3 Benthic fluxes of oxygen and inorganic nutrients............................................ 20 3.4 Sediment samples............................................................................................. 21
3.4.1 Chlorophyll a.............................................................................................. 21 3.4.2 Infauna – abundance and biomass.............................................................. 21 3.4.3 Characterisation of sediment ...................................................................... 21
3.5 Statistics used in the experiments..................................................................... 22 4 Significance of the results and discussion ............................................................. 23
4.1 Main findings ................................................................................................... 23 4.2 Interactions and feedback mechanisms ............................................................ 24
4.2.1 Sediment characteristics and nutrient fluxes.............................................. 24 4.2.2 Trophic state of the sediment ..................................................................... 26 4.2.3 Nutrient demand of algal mats ................................................................... 28 4.2.4 Green algal mats and benthic primary producers....................................... 29 4.2.5 Grazing – the menace from below ............................................................. 31 4.2.6 Macrofauna, hypoxia, and algal mats......................................................... 32
5 Conclusions and future work ................................................................................. 34 6 Acknowledgments.................................................................................................... 34 7 References ................................................................................................................ 36

10
Part B This thesis is mainly based on investigations presented in the following published/submitted articles and manuscripts, referred to in the text by their roman numerals. The papers are appended at the end of the thesis. List of papers
I. Engelsen A., Hulth S., Pihl L., Sundbäck K. (2008) Benthic trophic status and nutrient fluxes in shallow-water sediments. In press Estuarine, coastal and shelf science.
II. Engelsen A. & Larson F. (submitted) Factors influencing initiation and
growth of green algal mats. Submitted to Journal of experimental marine biology and ecology.
III. Engelsen A., & Pihl L. (2008) Grazing effects by Nereis diversicolor on
development and growth of green algal mats. In press Journal of Sea Research.
IV. Engelsen A., Sundbäck K., Hulth S., Links between oxygen deficiency,
macroalgal growth and grazing by Nereis diversicolor. Manuscript. A doctoral thesis at a university in Sweden is produced either as a monograph or as a collection of papers. In the latter case, the introductory part constitutes the formal thesis, which summarizes the accompanying papers. These have already been published or are manuscripts at different stages (in press, accepted, submitted or manuscript).

11
1 Introduction 1.1 Shallow marine sediment habitats As a direct result from the large concentration of human population in the coastal zone, coastal ecosystems are among the most altered and impacted ecosystems on the globe (Adger et al. 2005, and references therein). Anthropogenic stressors in coastal areas include for example eutrophication from agriculture and industrial discharge (Akkoyunlu & Ileri 2003, Tokarev & Shulman 2007, Aneja et al. 2008) as well as over-fishing (Lotze 2007, Poulsen et al. 2007, Blanchard et al. 2008) and exploitation by various human activities such as increased boat traffic. While coastal zones consist of both rocky and sediment habitats, the Swedish coastline is dominated by sediment habitats. Shallow marine sediment habitats are ecologically important components of the aquatic ecosystem and have a high biological activity in terms of recycling, storing and removal of nutrients (McGlathery et al. 2007). These areas are also highly productive and important nurseries for economically important species of fish and shellfish (Pihl & Rosenberg 1982, Pihl & Wennhage 2002). The sediment is a complex system of biological, chemical and physical components, which all interact with each other and surrounding environments. Macro and meiofauna live in and on the sediment and contribute to physical mixing of the sediment (Aller 1982), affect fluxes of oxygen and nutrients as well as contribute to the content of organic matter in the sediment. Microphytobenthos (MPB) live on or in the sediment. They are either sessile or move up and down in the top 5 mm sediment layer according to the light-conditions (Admiraal 1984). In shallow-water sediments with no seagrass present, the autotrophic status is mainly determined by the photosynthetic activity of MPB (McGlathery et al. 2001, Sundbäck et al. 2003, Tyler et al. 2003) and MPB act as a filter for inorganic nutrients between the sediment and water column (Sundbäck et al. 1991, Saburova & Polikarpov 2003, Thornton et al. 2007). As a consequence, benthic nutrient fluxes correlate better with the net oxygen flux (trophic status) than with the organic matter content of the sediment (Paper I). Diverse seaweed communities have in many coastal areas become replaced by fast growing annual macroalgae that establish dense mats altering the local ecology of benthic systems (Worm et al. 1999). Annual macroalgae are found worldwide (Valiela et al. 1997) and they are dependent on high levels of nutrients (Sundbäck et al. 2003). They grow fast with rich nutrient supply since their morphology give them a large surface to volume ratio (Wallentinus 1984). Although algal mats occur frequently (Pihl et al. 1995), and are considered as a threat to shallow bay communities, there is limited information about their initiation and growth. During the past three decades ephemeral filamentous macroalgal mats have developed in shallow sheltered bays, and consequently the ecology of these bays are changing. Pihl et al. (1999) observed that along the micro-tidal west coast of Sweden, distributions of algal mats were not related to local point sources of nutrient discharge, but rather to the degree of shelter for wind and wave exposure and to the content of organic matter in the sediment. These findings indicate that the sediment may have a major role for the development of algal

12
mats. The biogeochemical coupling between sediment organic content and benthic nutrient fluxes is normally complex. This applies particularly in shallow-water illuminated environments where activities by microphytobenthos are superimposed on mineralization processes mobilizing nutrients to the pore water. In addition to the quantity and biogeochemical composition of the organic material, rates and speciation (e.g. NH4
+, NO2-, NO3
-) of nutrient fluxes are influenced by a variety of factors such as temperature, sediment porosity, physical and biological reworking of surface sediments, availability of oxidants and nutrients in the bottom water and the surface sediment, and benthic microalgal activity (Hansen & Kristensen 1997, Aller 2001, Eyre & Ferguson 2002, Sundbäck & Miles 2002). While bioturbation tends to increase the efflux of nutrients from the surface sediment (Aller 2001, Norling et al. 2007), microalgal activity contributes to the retention of nutrients in the sediment and may remove additional nutrients from the overlying water (Sundbäck & McGlathery 2005). Since the balance between autotrophy and heterotrophy (trophic status) of the sediment system is often governed by activities of microphytobenthos, benthic trophic status may be a general indicator of whether sediments remove or supply essential nutrients. 1.2 Macroalgal mats Mats of macroalgae are a common phenomenon in shallow coastal habitats around the world (Bonsdorff 1992, Valiela et al. 1997, Bäck et al. 2000, Vahteri et al. 2000, Nelson et al. 2003, Sundbäck et al. 2003). On the Swedish west coast, macroalgal mats usually bloom from June to October and mainly consist of filamentous algae such as Ulva spp. (former Enteromorpha spp.) and Cladophora spp. (Pihl et al. 1999, Sundbäck et al. 2003). A coupling between the organic matter content of the surface sediment and the distribution of macroalgal mats accords with the idea that sheltered bays and lagoons with re-occurring algal mats are self-perpetuating systems fuelled by the regeneration of nutrients, which at least partly originate from the algae themselves (Hodgkin & Birch 1986, Pihl et al. 1999). This sediment pool of nutrients becomes available for the algal mats particularly during periods of hypoxia and anoxia, events that often occur under thick algal mats (Thybo-Christesen et al. 1993, Norkko & Bonsdorff 1996a). It has been shown that the highest uptake rate of ammonium is in the lower part of the algal mat, probably depending on a nutrient supply from the sediment (Thybo-Christesen et al. 1993), and the assimilation of nitrogen depends on light conditions (McGlathery et al. 1997). A high irradiance may lead to a high uptake of nutrients and also prevent ammonium diffusion from the benthic nutrient source into the overlying water (McGlathery et al. 1997). Nitrogen seems to recycle in floating mats. In a situation with an oligotrophic water column, the biomass of the green algal mats may have been sustained partly by internal circulation of nutrients within the mat itself (Sundbäck & McGlathery 2005). The filamentous algae have a high surface area to

13
volume ratio and a fast nutrient uptake (Wallentinus 1984) which leads to a rapid growth. They also have low nutrient storage abilities; the acquired nutrients appear to be allocated directly into growth rather than to storage reserves (McGlathery 1992). The result of this is that they outcompete slower growing perennial algae such as e. g. fucoids during conditions of rich nutrient supply, but they need a continuous supply of inorganic nutrients to be able to grow. Thybo-Christesen (1993) concluded that the clear water below the mat was a result of nutrient uptake by the filamentous algae and a result of out-competing of phytoplankton. At high nutrient loadings the growth of phytoplankton was reduced by a factor of 10 in the presence of algal mats (Fong et al. 1993). The competition hierarchy was cyanobacterial mats >> attached green macroalgae > floating green macroalgae > phytoplankton (Fong et al. 1993). The dominance of these algal mats over other benthic macroalgae may in part be due to their rapid rates of growth under nutrient enrichment (Pedersen & Borum 1997). The resultant shading from the opportunistic growth of ephemeral macroalgae further suppress slow-growing benthic macrophytes. Fast-growing benthic macroalgae have a low C/N ratio and decompose rapidly. When macroalgal blooms collapse, nutrients released to the water column temporarily stimulate phytoplankton production, and since ephemeral macroalgae are rich in N and low in structural carbohydrates compared to seagrasses, the decomposition is also likely to stimulate bacterial metabolism (McGlathery 2001, and references therein). 1.3 Inorganic nutrients and microphytobenthos In shallow sediment habitats, biogeochemical processes take place where dense communities of benthic microorganisms drive central ecosystem functions, such as primary production, decomposition and nutrient cycling (Paerl & Pinckney 1996). Benthic microalgal production constitutes the major primary production in shallow sediment areas that lack other macroscopic primary producers. Their production often exceeds that of phytoplankton (Underwood & Kromkamp 1999), and MPB can effectively reduce inorganic nutrients efflux from the sediment during the period that is critical for the initiation of the growth of macroalgal mats (Sundbäck & Miles 2002). These microalgae actually thrive below the mat (Sundbäck et al. 1996), and seem well adapted to withstand the conditions that are established below floating green algal mats. The light conditions below a macroalgal mat are considerably lower than the light conditions on bare sediment surfaces, but MPB can acclimate to lower light conditions. The combination of reduced light below a dense macroalgal mat, and respiration and decomposition within the mat, induce hypoxia or anoxia at the sediment surface during night and also during the day when mats are dense enough (Sundbäck & McGlathery 2005, and references therein). At anoxic conditions, the chemistry of the sediment changes, so that phosphate and ammonium are released into the water column (Blackburn & Henriksen 1983, Sundby et al. 1992), sustaining further algal growth.

14
1.4 Algal mats and benthic macrofauna Despite the obvious link to nutrient availability, the development of macroalgal mats appears to be rather unpredictable, and occasionally they fail to develop in the spring and may be absent during the summer period, suggesting that there are other controlling factors besides nutrients. As algal mats are found in shallow areas, it is logical to expect a strong coupling between the algae and processes occurring in the sediment. Such processes may be closely related to benthic macrofauna, although this functional coupling has gained far less attention than the nutrient-algae link. For example, Hydrobia spp. is shown to function as substrata for green-algal propagules (Schories et al. 2000) and bioturbating infauna is known to increase the efflux of NH4 (Emmerson et al. 2001, Welsh 2003) and macroalgae can be redistributed by macrofauna, such as the polychaete Nereis diversicolor (Nordström et al. 2006). Also macrofaunal grazing on both propagules and established macroalgal mats may be an important controlling factor (Raffaelli 2000, Giannotti & McGlathery 2001, McGlathery 2001) and Paper III. Shallow sediment bays have a high infaunal production and are important nurseries and feeding habitats for epibenthic fauna and fish (Möller et al. 1985). A general response of infauna to a high biomass of algae in shallow bays seems to be vertical migration to the surface to avoid hypoxic conditions in the sediment (Norkko et al. 2000). Some benthic species may also migrate further into the algal mat and reach high abundances among algal strands and filaments (Bonsdorff 1992). Survival of suspension feeding organisms generally decline below macroalgal mats, whereas burrowing detrivores seem to increase (Norkko & Bonsdorff 1996b, Österling & Pihl 2001). Algal mats have also been shown to filter out pelagic larvae of the water column during settlement, which may reduce the recruitment to these populations at high algal disturbance (Olafsson 1988, Bolam et al. 2000). The activities of bioturbating and irrigating benthic infauna tend to stimulate mineralization and coupled nitrification-denitrification in sediments (Gilbert et al. 2003). However, experimental observations and model simulations indicate an optimal nitrification – denitrification and ammonification–denitrification coupling with relative spacing between macrofaunal burrows. At a close distance between burrows (< 5 mm), more or less the complete sediment was oxidized by fauna and denitrification was hindered by O2. If burrows were more separated apart, however, sediments were less oxygenated and denitrification was restricted due to limited availability of nitrate (Gilbert et al., 2003). Dominance by ephemeral macroalgae and the associated changes in benthic fauna in eutrophicated estuaries may also influence the extent to which the system acts as a “filter” to process nitrogen inputs on their transport to the coastal ocean. It is shown that if the cover of filamentous algae are moderate (30-50 %) the species richness and biomass increase, while a high cover (90 %) causes a reduction of the biomass of epibenthic fauna (Pihl et al. 1995). In the Baltic sea there was a massive die-off of benthic fauna populations under drifting algal mats of e.g. the mud snail Hydrobia spp.

15
and the bivalve Macoma baltica, while populations of polychaetes like Nereis diversicolor and tubificid oligochaetes remained stable (Norkko & Bonsdorff 1996a). Also Capitella capitata can withstand long periods of macroalgal mats (Bolam et al. 2000). Shifts in the composition of faunal assemblages may also occur when there is a rich growth of filamentous algae in eelgrass beds. Pihl et al. (1995) found a general reduction in the number of fish species with increasing cover of epiphytic filamentous algae, and the foraging efficiency of juvenile cod and the settling success of plaice were reduced as a response to increasing algal cover (Isaksson et al. 1994, Wennhage & Pihl 1994). By affecting predator-prey relationships, eutrophication-related structural changes in macro-vegetation might cause considerable alterations in trophic relationships in shallow coastal waters. Filamentous drifting algae mats create a mosaic of patches, bare and covered, which increase habitat heterogeneity. Algal mats may induce conditions of hypoxia, but at the same time provide refuge from predation (Norkko 1998), which may result in the algal mat acting as a stabilizer of predator-prey interactions in shallow bays. 1.5 Nereis diversicolor Nereis diversicolor (Fig. 2) has been found to affect the biomass of Ulva sp. both by grazing and by burrowing algal material into the sediment (Hughes 1999, Raffaelli 2000, Nordström et al. 2006). In paper II there was an indication that N. diversicolor may have affected the growth of the algal mats. This polychaete is generally known as a predator or deposit feeder, but can also act as a filter feeder (Vedel & Riisgard 1993, Kristensen & Mikkelsen 2003), but the function as an efficient grazer and moderator of green algal mats has not been discussed before. During the experiments in papers II, III and IV this polychaete has been seen (by eye) grazing on the propagules and filaments in the surface sediment and on the edges of the cores but also in the full grown algal mat. N. diversicolor seems to be indifferent to stress from algal mats (Norkko & Bonsdorff 1996a) and can tolerate low oxygen levels (Janas et al. 2004), which is necessary when living below a macroalgal mat. The results in the studies (Papers II, III and IV) point towards N. diversicolor as an important factor to restraining macroalgal growth.
Fig. 2 Nereis (Hediste) diversicolor

16
The abundance of Nereis diversicolor changes from year to year, which might affect the algal mat differently each year. At the size of 20 to 25 mm N. diversicolor larvae change from a non-feeding period to an active dispersal period in order to find food (Green 1968). During this dispersal period they also compete for space (Möller & Rosenberg 1983) and thereby are exposed to numbers of predators (Crangon crangon, the 0 – group of plaice, birds and cannibalism from adult N. diversicolor) (Pihl & Rosenberg 1984) which can explain the larvae mortality of 97% (Möller, 1985). Also bottom frozen sediments during winters may reduce a whole population (Möller 1985 and references therein), which in turn is of importance for algal growth the following summer. Growth of Ulva spp. spores decreases when salinity is below 5, but is enhanced when it is over 20 (Sousa et al. 2007). N. diversicolor, on the other hand, does not seem to be affected by natural changes in salinity (Kristensen 1983). Ulva spp. spores, however, seem to be sensitive to high concentrations of ammonium and to P limitation (Sousa et al. 2007) which makes them vulnerable to large changes in nutrient concentrations. Inorganic nutrients can in addition to land run off also be released from the sediment by bioturbation (Aller 1982, Hansen & Kristensen 1997, Marinelli & Williams 2003). Temperature is of importance for both germination of propagules and the activity of N. diversicolor. Since polychaetes in cold water have a slower intestinal transit and thereby higher absorption efficiency (Neuhoff 1979), foraging is lower in colder temperatures and stops at 8 °C (Lambert et al. 1992). Germination of Ulva spp. is also favored by warm temperatures with an optimum between 17-22 °C (Sousa et al. 2007). Also the timing between recruitment of N. diversicolor larvae contra nutrient input seems to be of importance for development of algal mats.

17
2 Aims of the thesis Shallow marine sediment habitats where macroalgal mats grow are complex systems. Several studies have been performed in the past that have given valuable information to this field, but still there are some unknown facts that probably will add pieces to the complete understanding of this ecosystem. This thesis focuses on the role of biogeochemical processes in the sediment for the initiation and growth of green macroalgal mats. Field samplings and manipulated experiments were performed during four summer seasons to investigate the processes that are involved in macroalgal growth. All seasons included incubations of sediment-water cores, measurements of oxygen and nutrient fluxes and measurements of macrofauna abundance and biomass and of macroalgal growth. The overall aim for the project was to study interactions and feedback mechanisms between ephemeral floating macroalgal mats and the sediment in shallow habitats. The specific aims were:
• To investigate the coupling between sediment characteristics, trophic state and benthic nutrient fluxes in shallow-water environments.
• To investigate the importance of sediment nutrient release for the initiation of
macroalgal growth.
• To investigate the grazing ability of Nereis diversicolor on developing
macroalgal mats (Ulva spp), • To investigate the link between oxygen deficiency, grazing by Nereis
diversicolor and the growth of macroalgal mats (Ulva spp.)

18
3 Methods 3.1 Study area Macroalgal mats have since the 1980s increased in the Skagerrak archipelago. During the early 1990s most of the bays contained green algal mats that covered 20 – 100 % of the bay, and since then mats seem to appear every year, although unpredictable in which bay or which site. The investigations were carried out in shallow bay areas at the Gullmar fjord entrance and at Sven Love`n Center of Marine Sciences – Kristineberg (58º 15′ N, 11º 30′ E) on the west coast of Sweden. (Fig 1). The land area is characterized by agriculture, rocks and patches of deciduous forest, while the marine area consists of shallow sediment bays and stratified fjords surrounding the archipelago. The area is estuarine and micro-tidal with an average tidal amplitude of 20-30 cm and with influence from the Baltic current. During summer, shallow (0- 1 m) coastal water has a salinity between 20 and 25, and the water temperature ranges from 5 to 15 °C in spring and autumn, and from 15 to 25 °C during summer (Pihl & Rosenberg 1982). Occasionally, the temperature of shallow bays reaches 30 °C. The bottom substrate of the investigated bays varies from sandy to silty sediments, mainly determined by the degree of wave exposure (Pihl 1986). The area is considered as eutrophicated and filamentous ephemeral algae commonly overgrow perennial macroalgae and seagrass meadows, as well as develop into dense mats on bare sediment bottoms.

19
Fig. 1 The sites, marked with arrows, where sediment for experiments in the laboratory were collected. 3.2 Experimental approach and methodological considerations In all experiments, undisturbed sediments were used so that biogeochemical gradients, macrofauna and microphytobenthos could be retained intact. All sediment cores were collected by hand in shallow bays at 30-40 cm water depth. Since green algal mats grow in shallow areas and there is a coupling between the sediment and the mat, the sediment used in the experiment should origin from shallow illuminated areas. Two different types of experimental setups were used, one laboratory incubation system and one flow-through system in a greenhouse. These set-ups differed in the material and size of the cores, light conditions and water circulation. In the laboratory experiment (Paper I) plexiglass cores (8 cm in diameter) were used. In the greenhouse experiments, black cores (ABS plastic), inner diameter 25 cm (Paper II) or 16 cm (Papers III and IV) were used. In the laboratory, the cores were inserted into a tank with circulating in situ water, exposed to a light level that equalled the mean irradiation for the month, with a light-dark cycle of 16/8. The overlying water in the cores was stirred by a magnetic device. In papers II-IV, the cores were inserted into basins in the greenhouse, with a flow-through of seawater (7 l h-1 core -1) that was

20
pumped up from the outside shallow (1 m) bay. In the greenhouse, the sediment was exposed to a natural diurnal light rhythm, although the green house itself reduced the natural outdoor insolation intensity by 25 % and excluded UV–light. By providing the sediment cores with a flow-through of seawater from the outside bay, they were exposed to natural fluctuating nutrient conditions which occur in the bays. The advantage when changing from the laboratory system to a flow-through system was the natural light conditions, with set and dawn and the incoming water from the bay with natural fluctuating conditions in salinity and nutrients. Also, there was an advantage to use larger cores which mirror the sediment systems in the bay better compared to a smaller core. 3.3 Benthic fluxes of oxygen and inorganic nutrients
In general there are two approaches for quantifying chemical fluxes of solutes across the sediment-water interface. Benthic fluxes are either directly measured during sediment-water incubations (Holm 1978, Hall 1984, Hulth 1995) or modelled as diffusive fluxes from concentration gradients within the diffusive boundary layer (Gundersen & Jörgensen 1990) or from gradients within the surficial sediment (Hall 1984, Hulth 1995). Measured fluxes integrate possible contribution from bioirrigating fauna and activities by microphytobenthos (MPB). In the studies reported in this thesis only measured fluxes are used since both fauna and microphytobenthos constitute important factors in the shallow illuminated sediment system. Oxygen was measured together with inorganic nutrients to ensure that the oxygen conditions were not changing more than 20 % in the cores since the behaviour of the inorganic nutrient species then could be due to the oxygen change (Dalsgaard et al. 2000). During the incubation inorganic nutrients and oxygen are released from, or incorporated into the sediment due to several occurring processes in the sediment such as microphytobenthic activity, bioturbation by macrofauna and diffusion. Light and dark incubations were both performed in the same cores, and when dark incubations were made during daytime (Paper I), a one hour preincubation in darkness was done prior to the actual incubation to get the sediment and particularly the MPB community acclimated to darkness. In papers II, III and IV the dark incubations were done at 12.00 p.m. The concentration of oxygen and the inorganic nutrients (ammonium, nitrate, phosphate and silicate) were used to calculate fluxes. In all experiments daily values were calculated since hourly values just give you a snapshot of the activity at the measuring moment. To be able to get an idea of the process in the sediment and predict nutrient availability for macroalgal mats the daily values seem more appropriate to use when e.g. calculating nutrient demand contra nutrient access. Although the daily calculated flux presented in papers II-IV is based on the hourly light values also a light factor, mirroring the sun set and dawn, is included in the calculation for daily oxygen fluxes. For daily fluxes of nutrients, the amounts of light and dark hours were used. In paper I where artificial light was used, the light intensity corresponds to the mean light intensity over the month.

21
To find the trophic state (balance between autotrophy and heterotrophy) of the sediment two methods have been used in this thesis (Paper I). Net daily oxygen fluxes were calculated to show the trophic state by subtracting CR (negative flux) from the dark period from daily NPP. In addition a categorization of the trophic state of the sediment was assessed by applying the Trophic Oxygen Status Index TOSI (Viaroli & Christian 2004), derived from the 4-scale Benthic Trophic Status Index BTSI (Rizzo et al. 1996), where category 3 is fully autotrophic, 2 net autotrophic, 1 net heterotrophic, and 0 fully heterotrophic. In practice, TOSI is derived graphically by plotting NPP against CR (Viaroli & Christian 2004). While the net oxygen fluxes show whether the sediment is autotrophic or heterotrophic, the TOSI index also show the degree of trophy.
3.4 Sediment samples
3.4.1 Chlorophyll a Chlorophyll a was measured as an estimate of microphytobenthic biomass. There are several ways to measure the microphytobenthic biomass (e.g. measuring pigments by High Performance Liquid Chromatography (HPLC) or counting algae under a microscope) and more accurate ways as well, but the chlorophyll a was just used as a rough estimate of the MPB biomass to be able to discuss their possible influence on the system and compare biomasses between treatments. During work with the extraction and absorbance measurements it is of great importance the sample is not exposed to light and is frozen immediately after sampling, since the chlorophylls will degrade and the chlorophyll a concentration in the samples will be underestimated.
3.4.2 Infauna – abundance and biomass Because of their effect on nutrient fluxes from the sediment, but also as grazers on algae, macrofauna was of importance during these studies. Before start (initial cores) and after termination of all incubations the sediment in each core was carefully sieved through a mesh size of 1 mm with running water. The mesh size 1 mm was chosen since the main biomass of the adult macrofauna would be caught in that size. Also, to use a smaller mesh size will be a lot more time consuming and the fauna would probably have broken. The macrofauna were identified to species level when possible, counted and the biomass was estimated through weighting for all species found. According to Rumohr et al. (1987) a deduction of 25 % for shell weight for bivalve and gastropod species was done.
3.4.3 Characterisation of sediment To characterise the sediment the grain size, organic matter, density, porosity, pore-water concentration and C and N content in the sediment were analysed. These

22
analyses were made only in the first paper since it was assumed that these characters were rather constant and at least the joint relationship would last. To analyse the grain size of the sediment surface (0 – 3 cm), the sediment was dried until constant weight at 60 º C and sieved through four different mesh-sizes (500, 250, 100 and 63 µm) corresponding to coarse sand, fine sand, silt and fine silt fractions, respectively. The top two cm of the sediment was also incinerated at 540 º C for 4 hours to analyze the organic content of the sediment. Sediment for pore water and C and N content was manually sampled by inserting Plexiglas cores (diameter 8 cm) ~ 10 cm into the sediment. Samples were taken from two depths in the sediment (0-0.5 and 5-7 cm depth). The pore water obtained after refrigerated centrifugation was filtered on - line and stored frozen at – 80 º C until analysis. Total organic C and N contents of these sediment layers were determined by an elemental analyzer (EA 1108 CHNS-O, Fison, Italy). The porosities for the 0 – 0.5 and 5 – 7 cm sediment layers were estimated by the weight loss after drying wet sediment to constant weight at 60 º C and density of the sediment was calculated from the weight of 5 ml wet sediment. 3.5 Statistics used in the experiments A multivariate analysis (Principal Component Analysis, PCA, SIMCA program) was used to evaluate sediment properties, and to confirm or refute the initial classification of sampling sites made on grounds of basic sediment properties (Paper I). Pearson Correlation was used to evaluate correlations between measured variables (Paper I). One-factor ANOVA and two-factor ANOVA were used to test differences between variances (Papers I, II, III and IV). Analysis of variance was used since it is suitable for planned, manipulative experiments where predictive hypotheses are tested (Underwood 1997). Cochran’s test was performed prior to the analyses to test for heterogeneity of the variances (Underwood 1997). Tukey´s test was used as post-hoc test in cases of significant interactions. Values were considered significant at p ≤ 0.05.

23
4 Significance of the results and discussion 4.1 Main findings The main findings of the study were.
• The trophic state appeared to be a more viable parameter to classify sediments and predict benthic nutrient fluxes than the organic matter content of the sediment. Sediments rendered autotrophic by microphytobenthos function as sinks and heterotrophic sediments as sources (Paper I).
• Macroalgae (e.g. Ulva spp.) grow well without the underlying sediment, and the
only supply of nutrients may come from the water column (Paper II).
• The green algae were outcompeted for nutrients by cyanobacteria (Paper II) • Grazing by Nereis diversicolor was a main controlling factor of green algal
growth (Paper III).
• The individual weight of N. diversicolor was significantly increased for
polychaetes feeding on macroalgae compared to those which had not (Papers III and IV).
• Periods of benthic oxygen depletion stimulated the growth of green algal mats,
not only by increased availability of nutrients from the sediment, but also by reduced grazing pressure on the algae (Paper IV).
A green algal mat is a link in a complicated system where the sediment system with algal mats includes several interactions and feedback mechanisms. Processes and functions seem to vary according to structural parameters in the sediment such as grain size and porosity (Paper I), biotic factors such as grazing and bioturbation by macrofauna (Papers II, III, IV) and microphytobenthic activity (Papers I and II) and abiotic factors such as nutrient supply (Papers I, II, III and IV). Despite the varying environment in shallow sediment habitats, green algal mats grow well in the bays. The ecosystem of marine shallow sediment habitats harbouring filamentous ephemeral green algae seems to be a multipart system. These studies focused on the trophic state of the sediment to be able to predict nutrient fluxes and nutrient demand of algal mats in addition to grazing activities, initiation of algal growth and hypoxic conditions. The first study (Paper I) suggests that benthic trophic state is a more viable parameter than sediment organic content to classify sediment type and predict benthic nutrient fluxes. Paper I also found that even the most heterotrophic sediment type appeared to provide only a minor part (20 %) of the N demand, but up to 70 % of the P demand for macroalgal mats. However, the second study (Paper II) concluded that macroalgae (e.g. Ulva spp.) grew well without the underlying sediment, and the only supply of

24
nutrients came from the water column. The establishment of a green algal mat is not obvious since for example cyanobacteria showed to be excellent competitors for inorganic nutrients, and grazing by N. diversicolor also seemed to prevent initiation of macroalgal growth. The third experiment further established that N. diversicolor was able to prevent initial algal growth, affect growth capacity and also partly reduce full - grown algal mats. More, the individual weight of N. diversicolor was significantly increased for polychaetes feeding on macroalgae compared to those which had not. There were also indications that a rich nutrient supply per algae biomass counteracted the grazing capacity of N. diversicolor. The indication was reinforced in the forth experiment (Paper IV), where the algal mat started to grow after oxygen deficiency due to a combination of higher nutrient release and lack of grazing. 4.2 Interactions and feedback mechanisms
4.2.1 Sediment characteristics and nutrient fluxes As green algal mats are exclusively observed in shallow areas, a strong coupling between the occurrence of algal mats and benthic organic matter mineralization is expected. The sediment types have different structural characteristics, reflecting, for example, the location of the bay, water movements, organic matter availability, benthic microalgal activity and grain size distributions (Paper I). In paper I the aim was to study the coupling between sediment characteristics and sediment nutrient fluxes. A conceptual model (Fig. 3) summarizes the results found on coupling between sediment characteristics and nutrient fluxes in the investigated sediments. The model suggests that nutrient fluxes are more strongly related to benthic trophic status than to organic matter content of the surface sediment. Without macroscopic vegetation, the autotrophic character of these sediments is directly determined by the activities of microphytobenthos (MPB). Reworking and irrigation by macrofauna (bioturbation) influence rates of benthic oxygen consumption, and may e.g. due to diurnal cycles of bioturbation activities (Karlson 2007b) thereby also, at least indirectly, affect the oxygen balance of sediments. Both microphytobenthos and macrofauna influence sediment–water nutrient fluxes (Sundbäck & Miles 2002, Norling et al. 2007), but while MPB remove nutrients from the pore water and the overlying water, macrofaunal activities often supply nutrients to the overlying water (Aller 1982, Karlson 2007a).

25
Nutrientflux
OM Trophic status MA
MPB
Infauna
Fig. 3 A conceptual model for links between organic matter content (OM), trophic status, nutrient fluxes, macroalgae (MA), infauna and microphytobenthos (MPB) In the first experiment (Paper I), organic matter content (OM) was measured at the start of the project and was used as a base when allocating the different sediment types into three main categories (high, medium and low). There was a vide range in OM content in the solid phase of the sediment ranging from 1 to 18 % (Fig. 4). However, sediment organic matter content is only a rough measure of general biogeochemical properties and not the only structural variable that controls benthic fluxes. For example, in illuminated sediments, the activity of microphytobenthos significantly influences benthic oxygen and nutrient fluxes (McGlathery et al. 2004).This is examplified at one of the medium-OM sites (Paper I), where nutrients were more or less completely retained by the sediment system during incubations. These low nutrient fluxes coincided with a high biomass of MPB and a low biomass of macrofauna and the combination of a high biomass of MPB (active retention of nutrients) and a low abundance of irrigating macrofauna (restricted pore water advection through bioturbation) obviously counterbalanced the solute transport (diffusive and advective) across the sediment-water interface. In addition, to support the suggestion, the PCA model (Paper I) also confirmed that OM was not the only fundamental parameter that determined biogeochemical properties and functions of the sediment at the sampling locations. For example, when compared with the initial categorization based on organic matter content alone, the sediments with a high content of organic matter were more dissimilar and separated apart by the PCA. This deviation supported the idea that additional variables other than OM content (e.g. solid phase C and N, structural and functional properties associated with the macrofaunal community and net oxygen flux) were important for rates and direction of benthic nutrient fluxes observed in the study.

26
Fig. 4 Loss on ignition (LOI, mean + SE, n = 3) in 8 bays.
4.2.2 Trophic state of the sediment Since the balance between autotrophy and heterotrophy (trophic status) of the sediment system is often governed by activities of microphytobenthos, benthic trophic status may be a general indicator of whether sediments remove or supply essential nutrients. The TOSI index (Trophic Oxygen Status Index), where TOSI index is derived graphically by plotting NPP against CR (Viaroli & Christian 2004), was used to determine the trophic state of the sediment (Paper I). When TOSI index was used, (Paper I), sediments with a high OM content were assigned either to categories net heterotrophic or net autotrophic (Fig. 5). Sites with medium organic matter content were either classified in categories net autotrophic or totally autotrophic, while bays with the lowest OM content were fully autotrophic (Fig. 5). While all, except two, sediment types in paper I were net autotrophic in June (Fig 6), sediments with a high OM content turned from autotrophic in spring to heterotrophic in autumn, whereas sediments with a low OM content not only remained their autotrophic character during both spring and autumn, but also increased their autotrophic status in autumn. Therefore, not only the trophic status as such, but also the degree of heterotrophy or autotrophy has to be considered to accurately predict and evaluate benthic nutrient fluxes in shallow-water illuminated environments. Organic matter (OM) content in the sediment has generally been used as an indicator to predict nutrient fluxes, but as the benthic trophic status is a functional variable more closely coupled to nutrient fluxes than the comparably conservative structural parameter organic matter content, it is

27
Fig. 5 Trophic Oxygen Status Index (TOSI), where category 3 is fully autotrophic, 2 net autotrophic, 1 net heterotrophic and 0 fully heterotrophic. Shown are 6 bays in June and in September (Paper I). suggested that the trophic status is a better predictor of benthic nutrient fluxes in these environments (Paper I) (Fig. 4 and 5). Heterotrophic sediments generally release inorganic nutrients which will give basic conditions for algal mats, while the

28
autotrophic sediment probably would not constitute a source since the sediment consumes nutrients.
4.2.3 Nutrient demand of algal mats A key question during this study (Paper I) was whether the trophic state also could be used as a viable parameter to predict the potential for macroalgal growth? It has been suggested that the sediment functions as a self-regenerating system, where decomposed and mineralized macroalgal mats will constitute the nutrient source for coming algal mats (Hodgkin & Birch 1986, Norkko & Bonsdorff 1996b, Pihl et al. 1999). However, in paper I it was found that the nutrient supply coming from the sediment only constituted up to 20 % of the estimated DIN-demand but up to 70 % of the DIP-demand in the late growth season. It seems that the sediment should contribute more to the nutrient supply late in the growth season (August-September) compared to in the spring, since the sediment with a high content of organic matter turned from an autotrophic state in spring to a heterotrophic state in autumn (Fig. 6). However, algal mats in Sweden have a growth season from May to September depending on light, nutrient supply and temperature, and with optimal environmental conditions algal mats can maintenance the biomass also in October. Other factors are probably involved in the nutrient supply, such as pore water and benthic fluxes of DON, which may be important components to understand the importance of benthic nutrient fluxes for macroalgal growth, and to fully account for the macroalgal nutrient demand in micro-tidal environments. However, in paper II, the algal mat grew well without an underlying sediment, which indicates that the nutrient supply from the water column could constitute a sufficient source. The water exchange in shallow bays is high, which results in potential new incoming nutrients during periods. A suggestion is that the mat itself can constitute a nutrient source, through an inner circulation of nutrients (Sundbäck & McGlathery 2005). The algae have no storage ability but are dependent on a continuous supply of nutrients. When the algal mat has grown dense a continuous decomposing and growing inside the mat is going on. This leads to a release of nutrients which are used within the mat, but maybe also leaks out from the mat and is used for germination of propagules outside the algalmat.

29
June
-50
0
50
100
150
200
250
TJA TJB FK RA BA LI BO FI
Net
O2 f
lux
mm
ol m
-2 d
-1
September
-50
0
50
100
150
200
250
FK RA BA LI BO FI
Net
O2 f
lux
mm
ol m
-2 d
-1
Fig. 6 Net O2 fluxes in June and in September. Shown is the balance between autotrophy- heterotrophy. Shown are mean values + SE, n = 3.
4.2.4 Green algal mats and benthic primary producers In paper I it was shown that MPB affected the trophic state, and the trophic state of the sediment was suggested to predict the nutrient supply for algal mats. However, if there is a nutrient supply from the water column, the microphytobenthos (MPB) only have a minor effect, since the calculated sediment supply only provided 20 % N and 70 % P. On the other hand, in paper II, the MPB mainly consisting of cyanobacteria outcompeted the growth of the algal mat. MPB and especially cyanobacteria are excellent competitors for inorganic nutrients (Fong et al. 1993, Sundbäck & Miles 2002), but also other MPB, such as diatoms, have earlier been found to out-compete the opportunistic green algae during the onset of the growth (Sundbäck & Miles 2002,
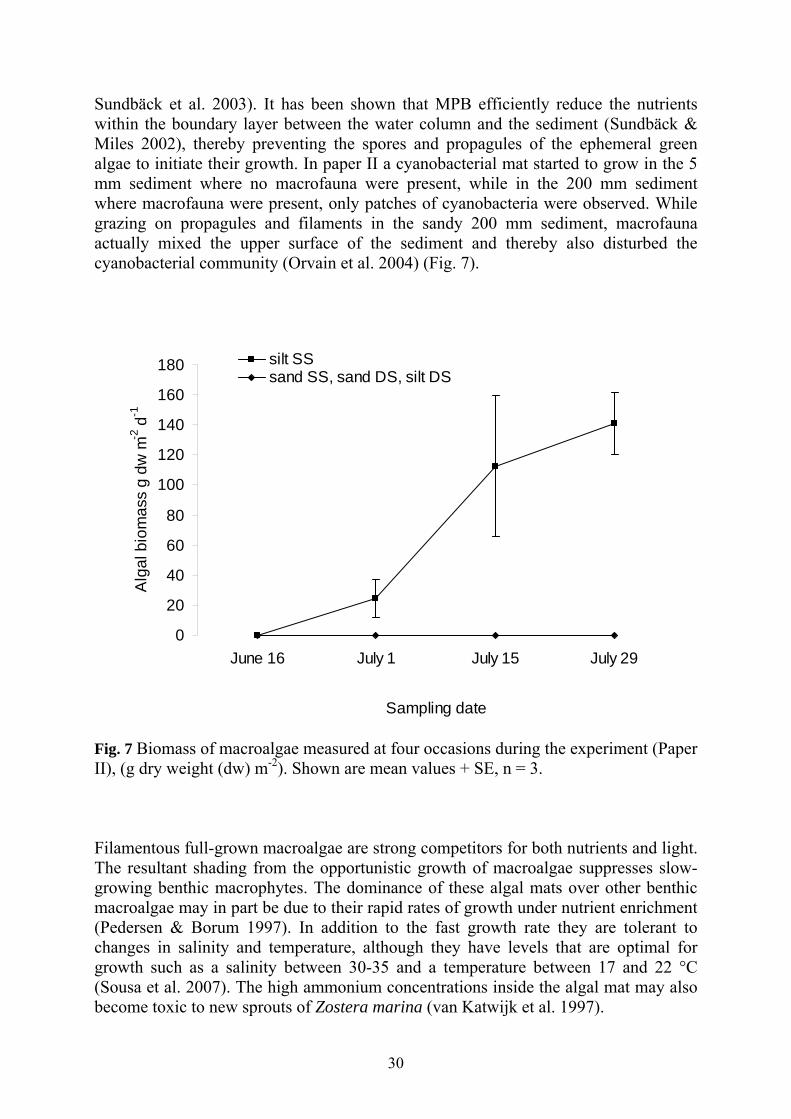
30
Sundbäck et al. 2003). It has been shown that MPB efficiently reduce the nutrients within the boundary layer between the water column and the sediment (Sundbäck & Miles 2002), thereby preventing the spores and propagules of the ephemeral green algae to initiate their growth. In paper II a cyanobacterial mat started to grow in the 5 mm sediment where no macrofauna were present, while in the 200 mm sediment where macrofauna were present, only patches of cyanobacteria were observed. While grazing on propagules and filaments in the sandy 200 mm sediment, macrofauna actually mixed the upper surface of the sediment and thereby also disturbed the cyanobacterial community (Orvain et al. 2004) (Fig. 7).
0
20
40
60
80
100
120
140
160
180
June 16 July 1 July 15 July 29
Sampling date
Alg
al b
iom
ass
g dw
m-2
d-1
silt SSsand SS, sand DS, silt DS
Fig. 7 Biomass of macroalgae measured at four occasions during the experiment (Paper II), (g dry weight (dw) m-2). Shown are mean values + SE, n = 3. Filamentous full-grown macroalgae are strong competitors for both nutrients and light. The resultant shading from the opportunistic growth of macroalgae suppresses slow-growing benthic macrophytes. The dominance of these algal mats over other benthic macroalgae may in part be due to their rapid rates of growth under nutrient enrichment (Pedersen & Borum 1997). In addition to the fast growth rate they are tolerant to changes in salinity and temperature, although they have levels that are optimal for growth such as a salinity between 30-35 and a temperature between 17 and 22 °C (Sousa et al. 2007). The high ammonium concentrations inside the algal mat may also become toxic to new sprouts of Zostera marina (van Katwijk et al. 1997).

31
4.2.5 Grazing – the menace from below The functional coupling between macrofauna and initiation of macroalgal mats has gained far less attention than feedbacks between nutrients and macroalgae. In papers II, III and IV, Nereis diversicolor was found to affect the biomass of Ulva sp. by grazing The results from the investigations in papers II and III and IV indicate that green algal mats should have difficulties to grow in shallow bays due to both grazing by N. diversicolor and competition from microphytobenthos (MPB)! On the contrary, green algal mats still grow well on the west coast of Sweden. However, the occurrence of algae varies between locations and over time, and grazing effects may at least partly explain the shifting growth patterns of macroalgal mats in time and space. It seems that N. diversicolor is able to both reduce fully-grown algal mats and prevent initial algal growth (Papers II and III). Nordström et al. (2006) showed that the limit for significant loss of biomass of green algae promoted in the presences of N. diversicolor seemed to be density dependent, where the threshold is between 35 and 105 individuals of N. diversicolor m-2. In the studies presented in papers II and III, the mean abundance of N. diversicolor exceeded the threshold found by Nordström et al. (2006), which supports the suggestion that the algal growth was inhibited by polychaetes. There is also an indication that the individual weight of N. diversicolor increased for those specimens that had been feeding on filamentous algae when compared with those feeding on propagules only (Papers III and IV). The fact that the worms that had been feeding on the macroalgae seem to grow faster than those feeding in cores with only propagules indicates that these macroalgae either are more nutritious to the worms or are more easy to find and feed on, resulting in higher consumption by N. diversicolor. Also the total energy cost for the worms might be lower when the polychaetes graze on green algae instead of being deposit feeders or predators. In micro-tidal environments the effects from grazing by N. diversicolor are likely most pronounced in regions where N. diversicolor and the algal mats, often floating at the air-water interface, are in close proximity. However, in micro-tidal environments the amplitude is also changed due to high and low air pressure, which can result in low tides for several days where grazing may reduce the algal mat entirely. In tidal environments, feedbacks may be different. For example during low tide, the algal mat will come in close contact with the sediment and N. diversicolor will be able to graze, while during high tide the algal mat will be out of reach of N. diversicolor. The fact that no growth of macroalgae occurred in the sediment cores (100 mm and 200 mm) with macrofauna, but always in the cores with surface sediment (5 mm) without macrofauna, during all experiments (Papers II, III and IV), indicates that the grazing effect may be much stronger when compared with the effect of nutrient supply. If the abundance of Nereis diversicolor is large enough (Nordström et al. 2006) the growth of algal mats will be prevented or reduced.

32
4.2.6 Macrofauna, hypoxia, and algal mats The biogeochemical coupling between sediment organic content and benthic nutrient fluxes is normally complex. If the algal mat grows dense and covers large areas, conditions of low oxygen might appear which in turn will affect the macrofauna negatively resulting in low or lack of bioturbation. The nutrient fluxes will decrease and the algal mat may suffer from nutrient limitation. On the other hand, if the main nutrient source is the water column, the decreased bioturbation will not seriously affect the algal mat. Floating macroalgal mats may create anoxic conditions. In dense mats, the combination of reduced light and enhanced decomposition within the mat induce low oxygen or anoxic conditions during both daylight and night conditions (Sundbäck & McGlathery 2005, and references therein). At hypoxia ([O2] < 40 µM), pulses of above all phosphate and ammonium are released to the bottom water from the sediment, which would benefit initiation of algal growth. During the periods of low oxygen conditions, the activity of grazing by Nereis diversicolor seems to be significantly reduced (Paper IV). The activity by N. diversicolor was affected only for a few days by the anoxic condition (Paper IV). After these days the algal mat started to grow because there were higher nutrient fluxes and lack of grazing. It has been shown that N. diversicolor can withstand anoxic conditions for up to 16 days (Vismann 1990), but if the algal mat creates a severe anoxic situation which lasts for several weeks, no infauna will survive but the sediment will release high amounts of nutrients which results in good growth conditions for the algal mat. In paper IV, the algal mat started to grow due to the combination of higher nutrient supply due to a period of low oxygen and inactivity of N. diversicolor. In the experiment (Paper IV) the sediment system recovered from hypoxia in 10 days or less, which agrees well with the resilience time after short hypoxic periods found by Larson and Sundbäck (2008). Thus, in shallow water illuminated sediments, activities by macrofauna may control benthic nutrient fluxes not only directly by pore water irrigation but also indirectly by grazing of benthic macroalgae. The results of the experiments in paper III indeed suggest that grazing by the polychaete N. diversicolor can control the establishment of green algal mats and that hypoxic events can set off this controlling factor (Paper IV). Although the hypoxic “stímulation” of algal growth is probably a combination of nutrient release from the sediment and release from grazing pressure, grazing, directly on propagules or mature algae, might be occasionally the stronger controller. The quantitative importance of grazing by N. diversicolor seems to be superimposed on the general availability of nutrients. For example, under periodic conditions of elevated nutrient supply from e.g. land run-off, the growth rate of the algae may increase and grazing might not have an impact. Inversely, during calm whether conditions with still water, the nutrient concentration will decrease and grazing by N. diversicolor could be the main factor controlling the existence of the green algal mats.

33
Coupling between N. diversicolor, hypoxia and macroalgal mats (Paper IV) is summarized in the conceptual model (Fig. 8).
Green algal mats
Nereis diversicolor
Inorganicnutrients Microphytobenthos
Oxygen
_ _
_
__
+
+ +
_
+
+
_
Fig. 8 Conceptual model showing couplings between oxygen, Nereis diversicolor, inorganic nutrients and macroalgae for shallow-water illuminated sediments regularly harbouring green-algal mats.

34
5 Conclusions and future work Benthic trophic status can be a more viable parameter than sediment organic content to classify sediment type and predict benthic nutrient fluxes. However, the contribution of nutrients even from a net heterotrophic sediment may not be sufficient to fully support optimal algal growth. Pore water and benthic fluxes of DON, may be key components to understand the importance of benthic nutrient fluxes for macro-algal growth, and to fully account for the macroalgal nutrient demand in micro-tidal environments. However, macroalgae (e.g. Ulva spp.) can grow well without the underlying sediment, with the only supply of nutrients coming from the water column. Also, both competition for nutrients by cyanobacteria and grazing by N. diversicolor seemed to prevent initiation of macroalgal growth. The third experiment further established that N. diversicolor was able to prevent initial algal growth, affect growth capacity and also partly reduce fully - grown algal mats. In addition, the individual weight of N. diversicolor was significantly increased for polychaetes feeding on macroalgae compared to those which had not. There were also indications that a rich nutrient supply per algal biomass counteracted the grazing capacity of N. diversicolor. The indication was reinforced in the forth experiment (Paper IV), where the algal mat started to grow after oxygen deficiency due to a combination of higher nutrient release and lack of grazing. The results from this project reflect only a part of the complex system, and they both agree with and contradict with the results from other investigations. Still, there are questions to answer and investigations to do. Questions related to this project are for example why algal mats increase? What is the main nutrient source for macroalgal mats? How will algal mats develop in the future when considering climate change and global warming? What measures are there to take to improve the shallow bay areas?
6 Acknowledgments Först och främst vill jag tacka mina tre handledare; huvudhandledare Leif Pihl och mina två biträdande handledare Kristina Sundbäck och Stefan Hulth, för att de gav mig chansen att få jobba i ”Baywatch” med vetenskap och som hjälpt och stöttat mig genom min doktorandtid. Leif, dig vill jag tacka för ditt stöd när livet varit hårt, och för att du tvingat mig att bli lite mer strukturerad. Stefan, alltid så entusiastisk och energisk och den som fått mig att tro på mig själv. Kristina, vi har ju kämpat med examensarbeten och Gullock innan detta, och även om du inte tror det så har du fått mig att bli bättre på att skriva! Fredrik Larson, tack för roligt arbete med sediment och alger, och inte lika roligt arbete med att sätta upp ett växthus! Vill även tacka för hjälp att komma hem från konferensen i Savannah, sällskapet under första ridturen i westernsadel under konferensveckan i Utah samt för att du är en trevlig vän. Malin Mohlin, vi har ju gått kurser och rest till Svalbard tillsammans, och du gör livet så roligt med dina galna upptåg och ambitiösa företag. Tack för din härliga vänskap och

35
för att jag fick övernatta i spindelmannen lakan när vi haft undervisning på baskursen. Angela Wulff, dig har jag också en hel del att tacka för. Du är en stor källa för inspiration när det gäller att ta sig fram här i livet. Tack för att du frågade om jag hade lust att jobba mig bort till Grönland och tillbaka och tack för chokladkakorna medan jag jobbade! Cecilia Nyberg, dig vill jag tacka för din vänskap, vår utekväll bland pälsjägarna på Svalbard och härliga turer på Svalbards flata toppar. Tack till mina rumskompisar, Johan Stål och Martin Eriksson, som kommit och gått in och ut i rummet ”längst bort”. Tack för intressanta diskussioner och pratstunder om allt möjligt. Så vill jag även tacka alla andra som är / har varit doktorand under samma tid som jag, samt all personal på Kristineberg för hjälp, pratstunder och glada tillrop. Tack även till alla de som på olika andra sätt bidragit till denna avhandling men som inte nämns här. Christian, dig vill jag ge en varm kram och säga att du är den som ger mig värme och livsgnista. Tack för att du hjälpte mig med layout i denna avhandling. Annars hade det aldrig blivit så fint. Simen, tack för ditt underbara leende, dina små goa’ kramar och för att du har givit mig en annan syn på livet. Min syster Elisabeth, du är min äldsta bästa vän och har alltid stöttat mig i allt jag gjort. Tack för att du finns där och kan lyssna och svara när jag behöver fråga om teknik i ridkonsten och tack för att du granskat en del av mitt arbete. Jag vill också säga tack till mina båda föräldrar som jag vet hade varit stolta. Jag vet att ni finns hos mig. Tack till mina kära hästar för alla skogspromenader, träningar och tävlingar – då jag helt kan koppla bort arbetet. Saga, min lilla tjej, tack för din alltid lika glada och högljudda hälsningsceremoni när jag kommer hem. Utan mina fyrbenta vänner hade det varit lite tomt. De som finansierat min doktorandtjänst, material, konferensresor och kurser är FORMAS, Birger och Birgit Wåhlströms minnesfond, Wilhelm och Martina Lundgrens Vetenskapsfond och Stiftelsen Ymer.

36
7 References Adger WH, Hughes TP, Folke C, Carpenter SR, Rockstrom J (2005) Social-ecological
resilience to coastal disasters. Science 309:1036-1039 Admiraal W (1984) The ecology of estuarine sediment-inhabiting diatoms. Progress in
Phycological Research 3:269-322 Akkoyunlu A, Ileri R (2003) Evaluation of eutrophication process in lake Sapanca
(Sakarya, Turkey). International Journal of Environment and Pollution 19:576-602
Aller RC (1982) The effects of macrobenthos on chemical properties of marine sediment and overlying water. In: McCall, PL, Tevesz, MJS (eds), Animal - Sediment relations, Plenum, NY:53-102
Aller RC (2001) Transport and reactions in the bioirrigated zone. In: Boudreau B, Jorgensen BB (eds) The benthic boundary layer: Transport processes and biogeochemistry Oxford Press:269-301
Aneja VP, Blunden J, James K, Schlesinger WH, Knighton R, Gilliam W, Jennings G, Niyogi D, Cole S (2008) Ammonia assessment from agriculture: US status and needs. Journal of Environmental Quality 37:515-520
Blackburn TH, Henriksen K (1983) Nitrigen cycling in different types of sediments from Danish waters. Limnology and Oceanography 28 (3):477-493
Blanchard JL, Maxwell DL, Jennings S (2008) Power of monitoring surveys to detect abundance trends in depleted populations: the effects of density-dependent habitat use, patchiness, and climate change. Ices Journal of Marine Science 65:111-120
Bolam SG, Fernandes TF, Read P, Raffaelli D (2000) Effects of macroalgal mats on intertidal sandflats: an experimental study. Journal of Experimental Marine Biology and Ecology 249:123-137
Bonsdorff E (1992) Drifting Algae and Zoobenthos - Effects on Settling and Community Structure. Netherlands Journal of Sea Research 30:57-62
Bäck S, Lehvo A, Blomster J (2000) Mass occurrence of unattached Enteromorpha intestinalis on the Finnish Baltic Sea coast. Annales Botanici Fennici 37:155-161
Dalsgaard T, Nielsen LP, Brotas V, Viaroli P, Underwood GJC, Nedwell DB, Sundbäck K, Rysgaard S, Miles A, Bartoli M, Dong L, Thornton DCO, Ottosen LDM, Castaldelli G, Risgaard-Petersen N (2000) Protocol handbook for NICE - Nitrogen Cycling in Estuaries: a project under the EU research program: Marine Science and Technology (MAST III).62
Emmerson MC, Solan M, Emes C, Paterson DM, Raffaelli D (2001) Consistent patterns and the idiosyncratic effects of biodiversity in marine ecosystems. Nature 411:73-77
Eyre BD, Ferguson AJP (2002) Comparison of carbon production and decomposition, benthic nutrient fluxes and denitrification in seagrass, phytoplankton, benthic microalgae- and macroalgae-dominated warm-temperate Australian lagoons. Marine Ecology Progress Series 229:43-59

37
Fong P, Donohoe RM, Zedler JB (1993) Competition with macroalgae and benthic cyanobacterial mats limits phytoplankton abundance in experimental microcosm. Marine Ecology Progress Series 100:97-102
Giannotti AL, McGlathery KJ (2001) Consumption of Ulva lactuca (Chlorophyta) by the omnivorous mud snail Ilyanassa obsoleta (Say). Journal of Phycology 37:209-215
Gilbert F, Aller RC, Hulth S (2003) The influence of macrofaunal burrow spacing and diffusive scaling on sedimentary nitrification and denitrification: An experimental simulation and model approach. Journal of Marine Research 61:101-125
Green J (1968) The biology of estuarine animals. London Sidgwick & Jackson Gundersen JK, Jörgensen BB (1990) Microstructure of diffusive boundary layers and
the oxygen uptake of the sea floor. Nature 345:604-607 Hall POJ (1984) Chemical fluxes at the sediment-water interface; In-situ investigations
with benthic chambers. PhD thesis, Dept of Analytical and Marine Chemistry, University of Göteborg, 1-183
Hansen K, Kristensen E (1997) Impact of macrofaunal recolonization on benthic metabolism and nutrient fluxes in a shallow marine sediment previously overgrown with macroalgal mats. Estuarine Coastal and Shelf Science 45:613-628
Hodgkin EP, Birch PB (1986) No simple solutions - proposing radical management options for an eutrophic estuary. Marine Pollution Bulletin 17:399-404
Holm NG (1978) Phosphorus exchange through the sediment-water interface. Mechanism studies of dynamic processes in the Baltic Sea. PhD thesis, University of Stockholm, 1-149
Hughes RG (1999) Saltmarsh erosion and management of saltmarsh restoration; the effects of infaunal invertebrates. Aquatic Conservation-Marine and Freshwater Ecosystems 9:83-95
Hulth S (1995) Mineralization of biogenic debris in continental shelf and slope sediments. PhD thesis, University of Göteborg, 1-98 and 6 appendices:39-46
Isaksson I, Pihl L, van Montfrans J (1994) Eutrophication-related changes in macrovegetaion and foraging of young cod (Gadus morhua L.): a mesocosm experiment. Journal of Experimental Marine Biology and Ecology 177:203-217
Janas U, Wocial JA, Szaniawska A (2004) Seasonal and annual changes in the macrozoobenthic populations of the Gulf of Gdansk with respect to hypoxia and hydrogen sulphide. Oceanologia 46:85-102
Karlson K (2007a) Diurnal bioturbating activities of Monoporeia affinis: effects on benthic oxygen and nutrient fluxes. Marine Ecology-Progress Series 331:195-205
Karlson K (2007b) Diurnal bioturbating activities of Monoporeia affinis: effects on benthic oxygen and nutrient fluxes. Marine Ecology Progress Series 331:195-205
Kristensen E (1983) Ventilation and oxygen uptake by three species of Nereis (Annelida: Polychaeta). II. Effects of temperature and salinity changes. Marine Ecology Progress Series 12:299-306

38
Kristensen E, Mikkelsen OL (2003) Impact of the burrow-dwelling polychaete Nereis diversicolor on the degradation of fresh and aged macroalgal detritus in a coastal marine sediment. Marine Ecology Progress Series 265:141-153
Lambert R, Desrosiers G, Retiere C, Miron G (1992) Exploration of Feeding Area by Nereis-Diversicolor (Polychaeta) (Muller,O.F.) - Preliminary Data. Cahiers de Biologie Marine 33 (1):43 - 54
Larson F, Sundbäck K (2008) Role of microphytobenthos in recovery of functions in a shallow-water sediment system after hypoxic events. Marine Ecology Progress Series 357:1-16
Lotze HK (2007) Rise and fall of fishing and marine resource use in the Wadden Sea, southern North Sea. Fisheries Research 87:208-218
Marinelli RL, Williams TJ (2003) Evidence for density-dependent effects of infauna on sediment biogeochemistry and benthic-pelagic coupling in nearshore systems. Estuarine Coastal and Shelf Science 57:179-192
McGlathery KJ (1992) Physiological Controls on the Distribution of the Macroalga Spyridea-Hypnoides - Patterns Along a Eutrophication Gradient in Bermuda. Marine Ecology Progress Series 87:173-182
McGlathery KJ, Krause-Jensenb D, Rysgaardb S, Christensen PB (1997) Patterns of ammonium uptake within dense mats of the filamentous macroalga Chaetomorpha linum. Aquatic Botany 59:99-115
McGlathery KJ (2001) Macroalgal blooms contribute to the decline of seagrass in nutrient-enriched coastal waters. Journal of Phycology 37:453-456
McGlathery KJ, Anderson IC, Tyler AC (2001) Magnitude and variability of benthic and pelagic metabolism in a temperate coastal lagoon. Marine Ecology Progress Series 216:1-15
McGlathery KJ, Sundbäck K, Anderson IC (2004) The importance of primary producers for benthic nitrogen and phosphorus cycling. In: Nielsen SL, Banta GT, Pedersen MF (eds) Estuarine nutrient cycling: The influence of primary producers Aquatic Ecology Ser 2 : 231-261
McGlathery KJ, Sundbäck K, Anderson IC (2007) Eutrophication in shallow coastal bays and lagoons: the role of plants in the coastal filter. Marine Ecology Progress Series 348:1-18
Möller P, Rosenberg R (1983) Recruitement, abundance andproduction of Mya arenaria and Cardium edule in marine shallow waters, western Sweden. Ophelia 22:33-55
Möller P (1985) Production and abundance of juvenile Nereis diversicolor, and oogenic cycles of adults in shallow waters of western Sweden. Journal of the Marine Biological Association of the UK 65:603-616
Möller P, Pihl L, Rosenberg R (1985) Benthic faunal energy flow and biological interaction in some shallow bottom habitats. Marine Ecology Progress Series 27:109-121
Nelson TA, Nelson AV, Tjoelker M (2003) Seasonal and spatial patterns of "Green tides" (Ulvoid algal blooms) and related water quality parameters in the coastal waters of Washington state, USA. Botanica Marina 46:263-275

39
Neuhoff HG (1979) Influence of temperature and salinity on food conversion and growth of different Nereis species (Polychaeta, Annelida). Marine Ecology Progress Series 1:225-262
Nordström M, Bonsdorff E, Salovius S (2006) The impact of infauna (Nereis diversicolor and Saduria entomon) on the redistribution and biomass of macroalgae on marine soft bottoms. Journal of Experimental Marine Biology and Ecology 333:58-70
Norkko A, Bonsdorff E (1996a) Population responses of coastal zoobenthos to stress induced by drifting algal mats. Marine Ecology Progress Series 140:141-151
Norkko A, Bonsdorff E (1996b) Rapid zoobenthic community responses to accumulations of drifting algae. Marine Ecology Progress Series 131:143-157
Norkko A (1998) The impact of loose-lying algal mats and predation by the brown shrimp Crangon crangon (L.) on infaunal prey dispersal and survival. Journal of Experimental Marine Biology and Ecology 221:99-116
Norkko J, Bonsdorff E, Norkko A (2000) Drifting algal mats as an alternative habitat for benthic invertebrates: Species specific responses to a transient resource. Journal of Experimental Marine Biology and Ecology 248:79-104
Norling K, Rosenberg R, Hulth S, Gremare A, Bonsdorff E (2007) Importance of functional biodiversity and species-specific traits of benthic fauna for ecosystem functions in marine sediment. Marine Ecology Progress Series 332:11-23
Olafsson EB (1988) Inhibition of larval settlement to a soft bottom benthic community by drifting algal mats - an experimental test. Marine Biology 97:571-574
Orvain F, Sauriau PG, Sygut A, Joassard L, Le Hir P (2004) Interacting effects of Hydrobia ulvae bioturbation and microphytobenthos on the erodibility of mudflat sediments. Marine Ecology Progress Series 278:205-223
Paerl HW, Pinckney JL (1996) A mini-review of microbial consortia: Their roles in aquatic production and biogeochemical cycling. Microbial Ecology 31:225-247
Pedersen MF, Borum J (1997) Nutrient control of estuarine macroalgae: growth strategy and the balance between nitrogen requirements and uptake. Marine Ecology Progress Series 161:155-163
Pihl L, Rosenberg R (1982) Production, abundance and biomass of mobile epibenthic marine fauna in shallow waters, western Sweden. Journal of Experimental Marine Biology and Ecology 57:273-301
Pihl L, Rosenberg R (1984) Food selection and consumption of the shrimp Crangon crangon in some shallow marine areas in western Sweden. Marine Ecology Progress Series 15:159-168
Pihl L (1986) Exposure, vegetation and sediment as primary factors for mobile epibenthic faunal community structure and production in shallow marine soft bottom areas. Netherlands Journal of Sea Research 20:75-83
Pihl L, Isaksson I, Wennhage H, Moksnes P-O (1995) Recent increase of filamentous algae in shallow Swedish bays: effects on the community structure of epibenthic fauna and fish. Netherlands Journal of Sea Research 29 (3-4):349-358
Pihl L, Svenson A, Moksnes P-O, Wennhage H (1999) Distribution of green algal mats throughout shallow soft bottoms of the Swedish Skagerrak archipelago in

40
relation to nutrient sources and wave exposure. Journal of Sea Research 41:281-294
Pihl L, Wennhage H (2002) Structure and diversity of fish assemblages on rocky and soft bottom shores on the Swedish west coast. Journal of Fish Biology 61:148-166
Poulsen B, Holm P, MacKenzie BR (2007) A long-term (1667-1860) perspective on impacts of fishing and environmental variability on fisheries for herring, eel, and whitefish in the Limfjord, Denmark. Fisheries Research 87:181-195
Raffaelli D (2000) Interactions between macro-algal mats and invertebrates in the Ythan estuary, Aberdeenshire, Scotland. Helgoland Marine Research 54:71-79
Rizzo WM, Dailey SK, Lackey GJ, Christian RR, Berry BE, Wetzel RL (1996) A metabolism-based trophic index for comparing the ecological values of shallow-water sediment habitats. Estuaries 19:247-256
Rumohr H, Brey T, Ankar S (1987) A compilation of biometric conversion factors for benthic invertebrates of the Baltic Sea. Baltic Marine Biologist 9:1-57
Saburova MA, Polikarpov IG (2003) Diatom activity within soft sediments: behavioural and physiological processes. Marine Ecology-Progress Series 251:115-126
Schories D, Anibal J, Chapman AS, Herre E, Isaksson I, Lillebö AI, Pihl L, Reise K, Sprung M, Thiel M (2000) Flagging greens: hydrobiid snails as substrata for the development of green algal mats (Enteromorpha spp) on tidal flats of North Atlantic coasts. Marine Ecology Progress Series 199:127-136
Sousa AI, Martins I, Lillebo AI, Flindt MR, Pardal MA (2007) Influence of salinity, nutrients and light on the germination and growth of Enteromorpha sp spores. Journal of Experimental Marine Biology and Ecology 341:142-150
Sundby B, Gobeil C, Silverberg N, Mucci A (1992) The Phosphorus Cycle in Coastal Marine-Sediments. Limnology and Oceanography 37:1129-1145
Sundbäck K, Enoksson V, Graneli W, Pettersson K (1991) Influence of Sublittoral Microphytobenthos on the Oxygen and Nutrient Flux between Sediment and Water - a Laboratory Continuous-Flow Study. Marine Ecology Progress Series 74:263-279
Sundbäck K, Nilsson P, Nilsson C, Jönsson B (1996) Balance between autotrophic and heterotrophic components and processes in microbenthic communities of sandy sediments: a field study. Estuarine, Coastal and Shelf Science 43:689-706
Sundbäck K, Miles A (2002) Role of microphytobenthos and denitrification for nutrient turnover in embayments with floating macroalgal mats: a spring situation. Aquatic Microbial Ecology 30:91-101
Sundbäck K, Miles A, Hulth S, Pihl L, Engström P, Selander E, Svenson A (2003) Importance of benthic nutrient regeneration during initiation of macroalgal blooms in shallow bays. Marine Ecology Progress Series 246:115-126
Sundbäck K, McGlathery KJ (2005) Interactions between benthic macroalgal and microalgal mats. In Kristensen, E., Haese, R.R., Kostka, J.E. (eds) Interactions between Macro- and Microorganisms in Marine Sediments, AGU series. Coastal and Estuarine Studies 60. Chapter 2:7-29

41
Thornton DCO, Dong LF, Underwood GJC, Nedwell DB (2007) Sediment-water inorganic nutrient exchange and nitrogen budgets in the Colne Estuary, UK. Marine Ecology Progress Series 337:63-77
Thybo-Christesen M, Rasmussen MB, Blackburn TH (1993) Nutrient fluxes and growth of Cladophora sericea in a shallow Danish bay. Marine Ecology Progress Series 100:273-281
Tokarev Y, Shulman G (2007) Biodiversity in the Black Sea: effects of climate and anthropogenic factors. Hydrobiologia 580:23-33
Tyler AC, McGlathery KJ, Anderson IC (2003) Benthic algae control sediment-water column fluxes of organic and inorganic nitrogen compounds in a temperate lagoon. Limnology and Oceanography 48:2125-2137
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge. Cambridge University Press.
Underwood GJC, Kromkamp J (1999) Primary production by phytoplankton and microphytobenthos in estuaries. Advances in Ecological Research 29:93-153
Vahteri P, Makinen A, Salovius S, Vuorinen I (2000) Are drifting algal mats conquering the bottom of the Archipelago Sea, SW Finland? Ambio 29:338-343
Valiela I, McClelland J, Hauxwell J, Behr PJ, Hersh D, Foreman K (1997) Macroalgal blooms in shallow estuaries: Controls and ecophysiological and ecosystem consequences. Limnology and Oceanography 42:1105-1118
Wallentinus I (1984) Comparision of nutrient uptake rates for Baltic macroalgae with different thallus morphologies. Marine Biology 80:215-225
van Katwijk MM, Vergeer LHT, Schmitz GHW, Roelofs JGM (1997) Ammonium toxicity in eelgrass Zostera marina. Marine Ecology Progress Series 57:159-173
Vedel A, Riisgard HU (1993) Filter-feeding in the polychaete Nereis diversicolor: growth and bioenergetics. Marine Ecology Progress Series 100:145-152
Welsh D (2003) It's a dirty job but someone has to do it: The role of marine benthic macrofauna in organic matter turnover and nutrient recycling to the water column. Chemistry and Ecology 19(5):321-342
Wennhage H, Pihl L (1994) Substratum Selection by Juvenile Plaice (Pleuronectes-Platessa L) - Impact of Benthic Microalgae and Filamentous Macroalgae. Netherlands Journal of Sea Research 32:343-351
Viaroli P, Christian RR (2004) Description of trophic status, hyperautotrophy and dystrophy of a coastal lagoon through a potential oxygen production and consumption index--TOSI: Trophic Oxygen Status Index. Ecological Indicators 3:237-250
Vismann B (1990) Sulfide Detoxification and Tolerance in Nereis-(Hediste)-Diversicolor and Nereis-(Neanthes)-Virens (Annelida, Polychaeta). Marine Ecology Progress Series 59:229-238
Worm B, Lotze HK, Boström C, Engkvist R, Labanauskas V, Sommer U (1999) Marine diversity shift linked to interactions among grazers, nutrients and propagule banks. Marine Ecology Progress Series 185:309-314
Österling M, Pihl L (2001) Effects of filamentous green algal mats on benthic macrofaunal functional feeding groups. Journal of Experimental Marine Biology and Ecology 263:159-183

42

Benthic trophic status and nutrient fluxes in shallow-water sediments
Anna Engelsen1, Stefan Hulth2, Leif Pihl1, and Kristina Sundbäck3
1 Department of Marine Ecology, The Sven Lovén Centre for Marine Sciences, University of Gothenburg, SE-
450 34 Fiskebäckskil, Sweden
2 Department of Chemistry, University of Gothenburg, SE-412 96 Göteborg, Sweden
3 Department of Marine Ecology, University of Gothenburg, Box 461; SE-405 30 Göteborg, Sweden
Abstract
Proliferation of fast-growing ephemeral macroalgae in shallow-water embayments constitutes a large-scale environmental change of coastal marine ecosystems. Since inorganic nutrients essential for the initiation and maintenance of macroalgal growth may be supplied from the underlying sediment, we investigated the coupling between benthic inorganic nutrient (mainly N and P) fluxes and sediment properties in 6 bays representing a wide gradient of sediment characteristics (grain size, organic matter content, solid phase C and N). The initial characterization of bays was made in June and also included measurements of oxygen flux and microphytobenthic and macrofaunal biomass. In September, still within the growth season of the macroalgae, complementary experiments with sediment-water incubations for benthic flux measurements of oxygen and nutrients focused on trophic status (balance between auto - and heterotrophy) as a controlling factor for rates of measured benthic nutrient fluxes. Generally, sediments rendered autotrophic by microphytobenthic photosynthesis removed nutrients from the overlying water, while heterotrophic sediments supplied nutrients to the overlying bottom water. Estimations of the green-algal nutrient demand suggested that late in the growth season, net heterotrophic sediments could cover 20 % of the N-demand and 70 % of the P demand. As the benthic trophic status is a functional variable more closely coupled to nutrient fluxes than the comparably conservative structural parameter organic matter content, we suggest that the trophic status is a more viable parameter to classify sediments and predict benthic nutrient fluxes in shallow-water environments. Key words: sediment characteristics, benthic fluxes, autotrophy - heterotrophy, macroalgal growth, benthic microalgae
1. Introduction Shallow bays are highly productive habitats and important sites for organic matter mineralization and nutrient cycling in coastal marine environments (Borum, 1996). In these areas, benthic primary production by seagrasses, macroalgae and benthic microalgae typically exceeds that
*) Corresponding author: [email protected]
of phytoplankton (McGlathery et al., 2004). Because of the shallow water column, key biogeochemical processes, such as organic matter mineralization and nutrient cycling, take place in the surface sediment. An increased supply of nutrients to the coastal zone is currently shifting the dominance of primary producers, from seagrasses to fast-growing, annual macroalgae on the sediment or at the water surface. This phenomenon is observed world-wide (McGlathery et al., 2007; Valiela et al., 1997; Viaroli et al., 1996). Macro-algal

Engelsen et al. / Benthic trophic status and nutrient fluxes 2
mats not only affect benthic macrofauna and food webs (Isaksson et al., 1994; Raffaelli, 2000; Österling and Pihl, 2001), but may also control rates and pathways of organic matter mineralization and thereby nutrient regeneration to the bottom water (Lavery and McComb, 1991; McGlathery et al., 2001b; Sundbäck and McGlathery, 2005). On the west coast of Sweden, green-algal mats consist of filamentous species of the genera Cladophora and Ulva. The mats normally start to grow in May, with a biomass peak in mid-summer (Pihl et al., 1996) when nutrient concentrations in the water column are low. The growth season extends to October. Since the storage capacity of nutrients in these algae is generally low (Pedersen and Borum, 1997), a continuous nutrient supply is needed to maintain their dominance. The surface sediment is often an important nutrient source for sustaining further growth of already established mats of macroalgae (Astill and Lavery, 2001; Thybo-Christesen et al., 1993) and occasionally, nutrient fluxes from the sediment also seem sufficient to trigger the actual onset of macroalgal growth in spring (Sundbäck et al., 2003). The question then arises whether, and to what extent, nutrients provided by surface sediments control the distribution and re-occurrence of macroalgal mats. Pihl et al. (1999) observed that along the micro-tidal west coast of Sweden, distributions of algal mats were not related to local point sources of nutrient discharge, but rather to the degree of shelter for wind and wave exposure and, above all, to the organic content of the sediment. A coupling between the organic matter content of the surface sediment and the distribution of macroalgal mats accords with the idea that sheltered bays and lagoons with re-occurring algal mats are self-perpetuating systems fuelled by the regeneration of nutrients, at least partly originating from the algae themselves (Hodgkin and Birch, 1986; Pihl et al., 1999). The quantitative importance of sediments as a nutrient source for
macroalgal growth has, however, to our knowledge previously not been systematically evaluated. The biogeochemical coupling between sediment organic content and benthic nutrient fluxes is normally complex. This applies particularly in shallow-water illuminated environments where effects from activities by benthic microalgae are superimposed on mineralization processes mobilizing nutrients to the pore water. In addition to the quantity and biogeochemical composition of the organic material, rates and speciation (e.g. NH4
+, NO2-, NO3
-) of nutrient fluxes are influenced by a variety of factors, such as temperature, availability of oxidants and nutrients in the bottom water and the surface sediment, sediment porosity, and physical and biological reworking of surface sediments (Eyre and Ferguson, 2002; Hansen and Kristensen, 1997). While bioturbation tends to increase the efflux of nutrients from the surface sediment (Aller, 2001; Norling et al., 2007), microalgal activity contributes to the retention of nutrients in the sediment and may remove additional nutrients from the overlying water (Sundbäck and McGlathery, 2005). Since the balance between autotrophy and heterotrophy (trophic status) of the sediment system is often governed by activities of benthic microalgae, benthic trophic status may be a general indicator of whether sediments remove or supply essential nutrients. Although there are several observations that net autotrophic sediments retain dissolved inorganic nitrogen from the overlying water (Eyre and Ferguson, 2002; Sundbäck et al., 2004; Tyler et al., 2003), there are only few experimental studies specifically designed to study the relation between benthic trophic status and nutrient fluxes in shallow-water sediments (e.g. Sundbäck, et al., 2003). Our objective was to investigate the coupling between sediment characteristics and benthic nutrient fluxes in shallow-water

3 Engelsen et al. / Benthic trophic status and nutrient fluxes
environments. An important aspect was to investigate if benthic trophic status is a more viable predictor of nutrient fluxes than bulk sediment organic matter. The approach included a survey of several bays representing a broad spectrum of sediment biogeochemical characteristics, such as grain size, organic matter content, solid phase C and N, chlorophyll a, macrofauna, and pore water nutrient concentrations. Oxygen and nutrient fluxes were measured during sediment-water incubations under light and dark conditions. The overriding purpose was to elucidate key mechanisms behind the growth of nuisance macro-algae, particularly in micro-tidal areas. 2. Material and methods 2.1. Study area and sampling sites This investigation was carried out in shallow (< 1 m) micro-tidal (amplitude ~ 0.2 m) bays on the Swedish west coast (58o 15 N, 11o 30´ E) (Fig. 1). The area is significantly influenced by the brackish-water Baltic current and in summer, the salinity of the surface water typically fluctuates between 20 and 25 (Pihl and Rosenberg, 1982). Mean surface-water temperature usually ranges from 5 to 15 °C in spring and autumn, and from 15 to 25 °C during summer. Occasionally, the temperature of shallow bays reaches 30 °C. The bottom substrate of the investigated bays varies from sandy to silty sediments, mainly determined by the degree of wave exposure (Pihl, 1986). This investigation was divided into two experimental parts, performed in June (eight sampling sites) and September (six sampling sites) 2002, respectively. In June, the main emphasis was to characterize sediments with respect to grain size (% < 63 µm), content of organic material (% loss on ignition), porosity, pore water nutrient concentrations (NH4
+, ΣNO3- + NO2
- - referred to as NO3
-, HPO42- - referred to as
dissolved inorganic phosphorous, DIP, and Si(OH)4), solid phase organic carbon (C) and nitrogen (N) content, biomass of macrofauna and microphytobenthos (MPB), consumption and net/gross production of oxygen. Eight bays (Fiskebäckskil, FK, Rågårdsvik, RA, Lindholmen, LI, Bassholmen, BA, Bökevik, BO, Finnsbo, FI, Tjärnö A, (TJA), and Tjärnö B (TJB) with expected differences in OM content (Pihl et al., 1999) were selected for this initial characterization. In September, the main emphasis was to confirm and extend the June investigation to also include quantification of nutrient fluxes in the defined sediment types. Based on multivariate evaluation (principal component analysis; PCA) of sediment properties obtained in June, six bays were selected, representing sediments with high (FK and RA), medium (LI and BA) and low (BO and FI) content of OM (organic matter) in the solid phase, respectively. Due to sampling logistics and similarities in organic content within sites of high organic matter content, the two bays in the Tjärnö area (TJA and TJB) were on the September sampling occasion excluded from further investigations. 2.2. Sediment properties Sediment for pore water and solid phase was manually sampled by inserting Plexiglas cores (diameter 8 cm) ~ 10 cm into the sediment. The cores were transported to the laboratory in refrigerated cool-boxes. The top two cm of the sediment were sampled (n = 2) with a 50 ml cut-off syringe, and incinerated at 540 º C for 4 hours. Surface (0 – 3 cm) sediment for grain size distribution was dried until constant weight at 60 º C and sieved (n = 3) through four different mesh-sizes (500, 250, 100 and 63 µm) corresponding to coarse sand, fine sand, silt and fine silt fractions, respectively. The porosities for the 0 – 0.5 and 5 – 7 cm sediment layers were estimated by the weight loss after drying wet sediment to constant weight at 60 º C.

Engelsen et al. / Benthic trophic status and nutrient fluxes 4
Sweden
Finland
Denmark
Norway
Baltic sea
Gullmar
Fjord
0 200 400
km
0 5 10
km
(FI)(BO)(LI)(FK)(BA)(RA)
(TJA and TJB)
Fig. 1 Map of the location of investigated bays where sediment and in situ water were collected. FI = Finsbo (L), BO = Bökevik (L), LI = Lindholmen (M), FK = Fiskebäckskil (H), BA = Bassholmen (M), RA = Rågårdsvik (H), TJA and TJB = Tjärnö A and B (H) Total organic C and N content of these sediment layers were determined by an elemental analyzer (EA 1108 CHNS-O, Fison, Italy) using 2.5-Bis-[5-tert.-butyl-bensoaxzol-2-yl]-thiophen (BBOT) as standard. Sediment from the intervals 0 – 0.5 cm and 5 – 7 cm was also used for pore-water nutrient distributions. Density of the sediment was calculated from the weight of 5 ml wet sediment. The pore water obtained after refrigerated centrifugation (10°C, 2000 rpm, 30 min) was filtered (0.45 µm, pre-packed cellulose acetate) on - line and stored frozen at – 80 º C until analysis. In June, estimates of chlorophyll a (Chl a) and macrofauna were part of the characterization of the sediment. These parameters were again measured in September as they are important factors for benthic solute fluxes. The top 5 mm of the sediment were sampled with a cut - off 20 ml syringe (i.d. = 12 mm, n = 3) and immediately frozen (– 18 °C) until analysis
for Chl a, a rough estimate of the MPB biomass. The sediment was extracted for 24 hours in 4 ml 90 % acetone at 8 ° C and Chl a determined according to Lorenzen (1967), including correction for pheopigments. The macrofauna was sampled from the sediment cores used for benthic flux incubations (see below). After termination of the flux incubations, the sediment of the bioturbated zone (0–10 cm) was sieved through a 1-mm mesh. Fauna retained on the mesh was preserved in 70 % ethanol until identification to lowest possible taxonomic level. Macrofaunal taxa were divided into functional groups according to feeding mode (suspension feeders, surface detrivores, burrowing detrivores, predators and grazers). Biomass of shell-bearing macrofauna was corrected for shell weight (25 %) (Rumohr et al., 1987). 2.3. Benthic oxygen and nutrient fluxes Benthic oxygen and nutrient fluxes were measured during sediment–water

5 Engelsen et al. / Benthic trophic status and nutrient fluxes
incubations in the laboratory. Oxygen fluxes were measured in June as part of the characterization of sediment properties (i.e. net/gross production of oxygen, n = 4). In September, oxygen fluxes (n = 3) were included with main emphasis to study the influence of trophic status (i.e. balance between daily production and consumption of oxygen) on nutrient fluxes. Sediment with ambient overlying water was sampled by hand using Plexiglas cores (diameter 8 cm, sediment height ~ 24 cm). Additional bottom water was sampled for the benthic flux incubations. After sampling, the cores were transported refrigerated to the laboratory in cool boxes and placed in large containers with a flow-through (20 ml min-
1) system of un-filtered bottom water from the sampling site. The cores were left open without lids over night and the incident light was adjusted to the daily mean light intensity for the month of sampling (530 and 330 µmol photons m -2 s -1 in June and September, respectively). Incubation temperature was the same as the in situ bottom water, i.e. 22 ºC in June and 23 ºC in September. Cores were sealed and pre-incubated for approximately one hour under which oxygen in the overlying water was measured twice to determine sampling intervals (n = 2, start/stop) for the benthic flux incubation. During incubations, oxygen concentrations measured in the overlying bottom water did not deviate by more than 20 % compared to the initial values. The same cores were used for light and dark oxygen and nutrient flux measurements, with 16/8 hrs and 14/10 hrs light/dark cycles in June and September, respectively. Samples for oxygen (20 ml glass syringes) and nutrients (50 ml polypropylene syringes) were removed from the overlying water. The concentration change in the overlying water with time of incubation was used as a direct measure of benthic flux rates (Hulth et al., 1994).
Dissolved oxygen was determined by Winkler titration. Oxygen flux in the light
was used as a measure of net primary production (NPP), and gross primary production (GPP) was calculated by subtracting community respiration (CR, negative flux) in the dark from NPP. For practical reasons, it was assumed that CR in the light and dark were equal. Because the irradiance used for the incubations represented a daily mean of the light hours during the month of the experiment, daily NPP and GPP were calculated by multiplying the hourly values by the number of daylight hours. Daily CR equalled the hourly oxygen flux in the dark times 24. Net oxygen flux over a 24-h period was calculated as hourly light values times the number of light hours (NPP), plus hourly dark values times the number of dark hour (CR, negative flux) A categorization of the trophic status (balance between autotrophy and heterotrophy) of the sediment was assessed by applying the Trophic Oxygen Status Index TOSI (Viaroli and Christian, 2004), derived from the 4-scale Benthic Trophic Status Index BTSI (Rizzo et al., 1996), where category 3 is fully autotrophic, 2 net autotrophic, 1 net heterotrophic, and 0 fully heterotrophic. In practice, TOSI is derived graphically by plotting NPP against CR (Viaroli and Christian, 2004). Samples for nutrients were filtered on-line (0.45 µm cellulose acetate) and frozen immediately at – 80 ° C until analysis. A multi-channel auto-analyzer (TRAACS 800, Bran & Lubbe) was used for colorimetric analysis of inorganic nutrients (Strickland and Parsons, 1972). The daily nutrient flux was obtained by adding the day and the night fluxes. Benthic nutrient (NH4
+, NO3, DIP and Si(OH)4) fluxes (n = 3 cores bay -1) were measured only in September. For a more detailed description of the principles of benthic flux measurements and the system used in this study see (Dalsgaard et al., 2000; Hulth et al., 1994).

Engelsen et al. / Benthic trophic status and nutrient fluxes 6
2.4. Green-algal nutrient demand The quantitative importance of pore water nutrients for macroalgal growth was estimated indirectly using green algae (Ulva sp.) incubated in cores with water from the sampling site, but without sediment (3 cores per bay). The initial biomass of algae added to each core was ca. 15 g wet weight (WW), which corresponds to 2.9 kg WW m-2 (~ 435 g DW m-2), a naturally occurring biomass in the study area (Pihl et al. 1996). The green-algal nutrient (N and P) demand was calculated according to two principles. (1) The measured increase of macroalgal biomass (WW and DW) during an incubation period of 7 days. The increase in DW was converted to a theoretical N demand by applying the measured N content in the algal biomass (mean 2.23 % of DW) and the surface area of the core (the algae covered the complete area). The P content of the algal biomass was not directly measured. Instead, we utilized a mean P content (0.15 % of DW) for August obtained in a previous study of several bays in the same area as that of our study (Melin, 2001). (2) In parallel, the nutrient demand was also calculated from the net oxygen production (NPP) measured during 1-h incubations under the same conditions as those of the nutrient flux measurements. Rates of oxygen production was converted to carbon assimilation (mmol C m-2 day-1) by applying a photosynthetic quotient of 1.2 (Glud et al., 2002) and a measured C/N molar ratio (range 10.2 - 13.8) of the green algae.
2.5. Multivariate and statistical analyses Multivariate analysis (Principal Component Analysis, PCA, SIMCA program) was used to evaluate sediment properties from the June sampling occasion, and to confirm or refute the initial classification of sampling sites made on grounds of basic sediment properties. In addition to sediment OM content, solid phase C, N and C/N ratio, porosity, sediment Chl a, and macrofaunal
abundance and biomass, the PCA also included pore water nutrient (NH4
+, NO3-,
DIP and Si(OH)4) concentrations and benthic oxygen (light, dark, and daily) fluxes.
Pearson Correlation was used to evaluate correlations between measured variables. One-factor ANOVA tested differences in sediment characteristics and nutrient fluxes between sediment types. Two-factor ANOVA tested differences between sampling occasions (June and September) and sediment type for Chl a, oxygen production and benthic macrofauna. Cochran’s test was performed prior to the analyses to test for heterogeneity of the variances (Underwood, 1997). Values were considered significant at p ≤ 0.05. 3. Results 3.1. Sediment characteristics, chlorophyll a and macrofauna The OM content in the solid phase of the sediment ranged from 1 to 18 % (Table 1; Fig. 2), with a significant difference in OM content between the three types of sediments. Highest OM (14 – 18 %) was found for TJA, TJB, FK and RA, with a progressively decreasing content for LI and BA (2.6 and 3.4 %), and BO and FI (1.0 and 1.2 %; Table 1; Fig.2). Locations with the highest OM content were characterized by significantly higher porosity (p = 0.007 for the 0–0.5 cm layer and p = 0.006 for the 5–7 cm layer) when compared with bays with medium and low OM content. There was also a significantly (p = 0.0035) higher content of silt in sediments with a high OM content (Table 1). There was a trend towards a higher C and N content in sediments with a high OM content when compared with medium and low OM content. Solid phase C/N ratios appeared highest for sediments with a low content of OM (Table 1).
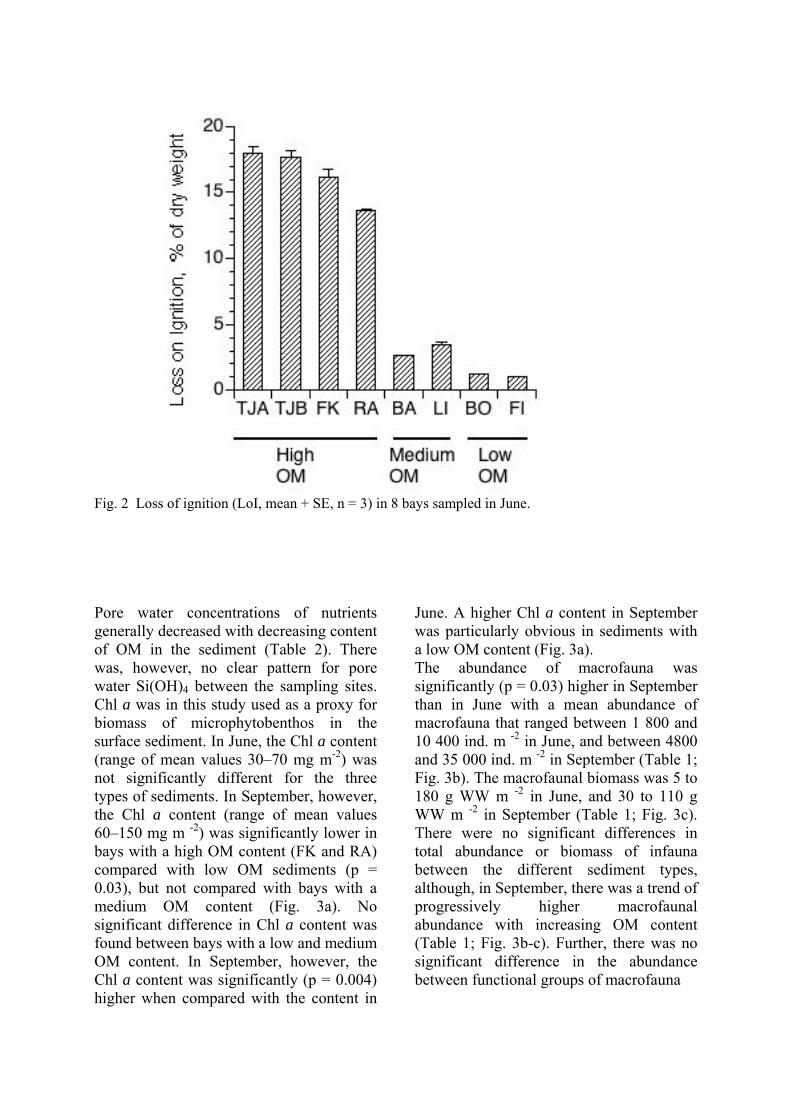
Fig. 2 Loss of ignition (LoI, mean + SE, n = 3) in 8 bays sampled in June. Pore water concentrations of nutrients generally decreased with decreasing content of OM in the sediment (Table 2). There was, however, no clear pattern for pore water Si(OH)4 between the sampling sites. Chl a was in this study used as a proxy for biomass of microphytobenthos in the surface sediment. In June, the Chl a content (range of mean values 30–70 mg m-2) was not significantly different for the three types of sediments. In September, however, the Chl a content (range of mean values 60–150 mg m -2) was significantly lower in bays with a high OM content (FK and RA) compared with low OM sediments (p = 0.03), but not compared with bays with a medium OM content (Fig. 3a). No significant difference in Chl a content was found between bays with a low and medium OM content. In September, however, the Chl a content was significantly (p = 0.004) higher when compared with the content in
June. A higher Chl a content in September was particularly obvious in sediments with a low OM content (Fig. 3a). The abundance of macrofauna was significantly (p = 0.03) higher in September than in June with a mean abundance of macrofauna that ranged between 1 800 and 10 400 ind. m -2 in June, and between 4800 and 35 000 ind. m -2 in September (Table 1; Fig. 3b). The macrofaunal biomass was 5 to 180 g WW m -2 in June, and 30 to 110 g WW m -2 in September (Table 1; Fig. 3c). There were no significant differences in total abundance or biomass of infauna between the different sediment types, although, in September, there was a trend of progressively higher macrofaunal abundance with increasing OM content (Table 1; Fig. 3b-c). Further, there was no significant difference in the abundance between functional groups of macrofauna

Table 1. Mean values of basic sediment (June) and biological characteristics (June and September) in eight bays on the Swedish west coast. Bays were allocated to three main categories based on organic matter content (high, medium and low). For acronyms of bays, see material and methods (n.d. = no data). High OM Medium OM Low OM
Sediment characteristics TJA TJ B FK RA LI BA BO FI
Organic matter, % 18 18 16 14 3.4 2.6 1.2 1.0 Grain size, % < 63µm 25 27 24 33 11 16 5.7 7.5 Density, 0–0.5 cm, g m-3 1.2 1.2 1.2 1.2 1.3 1.3 1.9 1.9 Density, 5–7 cm, g m-3 1.2 1.2 1.3 1.3 1.6 1.5 1.9 2.0 Porosity, 0–0.5 cm 0.9 0.9 0.8 0.9 0.7 0.6 0.5 0.5 Porosity, 5–7 cm 0.8 0.9 0.9 0.9 0.4 0.5 0.4 0.4 C, 0 – 0.5 cm, % 6.8 7.4 4.0 4.8 2.4 1.3 0.4 0.2 N, 0 – 0.5 cm, % 1.1 1.1 0.1 0.1 0.4 0.2 0.04 0.03 C/N, 0 – 0.5 cm, molar 7.3 8.1 6.5 7.8 7.9 7.8 11 8.9 C/N, 5 – 7 cm, molar 7.6 10 8.7 8.5 - 12 12 10
Biological characteristics June
Chlorophyll a, mg m -2 51 45 67 58 28 59 69 36 Macrofauna, ind. m -2 10400 8500 1800 5500 8700 5050 2050 5450 Macrofauna, g WW m -2 66 180 35 120 40 86 5 54
Functional groups Suspension feeders, ind.m -2 n.d. n.d. 400 600 750 250 0 0 Surface detrivores, ind.m -2 n.d. n.d. 350 1200 400 900 0 2200 Burrowing detrivores, ind.m -2 n.d. n.d. 850 2800 4550 1850 1250 2050 Predators, ind.m -2 n.d. n.d. 0 350 3000 1100 750 950 Grazers, ind.m -2 n.d. n.d. 200 550 0 650 50 200
September Chlorophyll a, mg m -2 n.d. n.d. 68 63 102 76 149 122 Macrofauna, ind. m - 2 n.d. n.d. 35350 15300 7900 20000 4800 13600 Macrofauna, g W.W. m -2 n.d. n.d. 85 110 33 97 31 112
Functional groups Suspension feeders, ind.m -2 n.d. n.d. 150 70 550 530 0 2400 Surface detrivores, ind.m -2 n.d. n.d. 200 450 250 250 70 800 Burrowing detrivores, ind.m -2 n.d. n.d. 32800 14400 6900 17850 4450 8600 Predators, ind.m -2 n.d. n.d. 0 250 150 150 150 1000 Grazers, ind.m -2 n.d. n.d. 2200 70 70 1050 150 800
(suspension feeders, surface detrivores, burrowing detrivores, predators and grazers) for the three sediment types in June (Table 1). In September, however, there was a significantly (p = 0.0001) higher abundance of burrowing detrivores (mainly caused by the colonisation of
Chironomidae), especially at the sites with a high and medium content of OM. In June, all bays (except for TJA and TJB that were excluded from the September sampling; see above) were autotrophic during daytime (i.e. NPP > 0; Fig. 4), as well as over a 24-hour period (i.e. net O2 flux > 0; Fig. 4).

Fig. 3 Chlorophyll a content, abundance and biomass (excluding shell weights; mean values + SE, n = 3) of macrofauna in 8 bays sampled in June and 6 bays sampled in September.

Engelsen et al. / Benthic trophic status and nutrient fluxes 10
3.2. Oxygen fluxes and trophic status In September, bays (FK and RA) with a high organic matter content (> 13 %) were heterotrophic (net O2 flux < 0), while bays (FI and BO) with a low organic matter content (< 2 %) were autotrophic. Bays with an intermediate OM content of the sediment (~ 3 %) were either autotrophic (LI) or heterotrophic (BA) during the September sampling occasion (Fig. 4). Net oxygen fluxes in September were significantly negatively correlated to the OM content (r = – 0.65, p = 0.05, n = 12). This correlation was not significant in June.
When applying the Trophic Oxygen Status Index (TOSI) to the September sampling, sediments with a high OM content (FK and RA) were assigned either to categories 1 (net heterotrophic) or 2 (net autotrophic), mainly depending on the variation between individual cores. Sites with medium organic matter content were either classified in category 2 (BA) or 3 (totally autotrophic, LI). Bays with the lowest OM content (BO and FI) were fully autotrophic (category 3). Using the TOSI, in essence, there was a progressive shift from category 1 to 3 with decreasing OM content.
Fig. 4 Daily (24-h) gross primary production (GPP), net primary production (NPP) and community respiration (CR) (top panels), and daily net oxygen flux (bottom panels) in June (8 bays) and September (6 bays). Shown are mean values + SE (n = 3).

11 Engelsen et al. / Benthic trophic status and nutrient fluxes
A significant correlation between sediment Chl a and net O2 flux (r = 0.940, p = 0.001) indicated that biomass of benthic microalgae strongly regulated the trophic status of the sediment system. There was a negative trend between community respiration and net O2 flux (r = –0.44, p = 0.1). Neither biomass nor abundance of macrofauna were significantly correlated to net oxygen flux or community respiration in September. In June, however, abundance, but not biomass, was significantly negatively correlated (r = – 0.745, p = 0.05) to the net O2 flux. Although not significant, there was a positive trend between faunal abundance and community respiration. 3.3. Benthic nutrient fluxes As a direct consequence of organic matter mineralization and solute mobilization to the pore water, NH4
+, Si(OH)4 and DIP fluxes were in general directed from the sediment to the overlying water. NO3
-, on the other hand, was mainly consumed by the sediment (Fig. 5). Fluxes of NH4
+ to the overlying water decreased and fluxes of NO3
- into the sediment increased, along the gradient of decreasing sediment OM content (Fig. 5). The highest efflux of NH4
+ was measured at FK (TOSI = 1), and the highest flux of NO3
- into the sediment was measured at FI (TOSI = 3). There was a significant positive correlation between fluxes (–600 to 4000 µmol m -2 d -1) of dissolved inorganic nitrogen (DIN = Σ NH4
+ +NO3- + NO2
-) and OM content (r = 0.599, p = 0.05, n = 12). Thus, the importance of the sediment as a source for DIN to the overlying bottom water increased with increasing content of organic matter. In accordance, there was also a trend of increasing DIN fluxes into the sediment with decreasing organic carbon (TOC) content of the sediment (not shown). However, no general correlation between OM content and DIP fluxes (–60 to 200 µmol m-2 d-1) was found, partly because a remarkably large (~ 200 µmol m -2 d -1) efflux of DIP was measured at FI, one of
the bays with a low organic matter content (Fig. 5). Si(OH)4 fluxes ranged between 130 and 6000 µmol m -2 d -1, with no obvious pattern between the sampling sites (Fig. 5). At LI, all daily nutrient fluxes were consistently low (DIN < 50 µmol m -2 d -1, DIP ~ 0 µmol m -2 d -1 , Si(OH)4 < 150 µmol m -2 d -1) (Fig. 5).
There were significant negative correlations between the net oxygen flux, and fluxes of NH4
+, NO3-, DIN and DIP (Fig. 6). The
correlation was, however, not significant for Si(OH)4. Further, there were significant negative correlations between Chl a, and benthic fluxes of DIN and DIP (Fig. 6). In addition, different rates of nutrient fluxes during day and night conditions further confirmed the importance of MPB for regulating sediment–water nutrient fluxes. There was a significant positive correlation between abundance (but not biomass) of benthic infauna, and fluxes of NH4
+ and Si(OH)4 (Fig. 7). In contrast, DIP fluxes correlated significantly with the biomass of infauna (r = 0.586, p = 0.05), but not with macrofaunal abundance. 3.4. Multivariate analysis – PCA Although based on 26 variables related to properties and functions of the shallow-water system rather than OM content alone (PCA n × m = 8 stations × 26 variables), the PCA model confirmed the initial general allocation of bays into three main categories (Fig. 8). The model made the largest distinction between BO and TJB, while TJA and TJB, and LI and BA, respectively, and formed two categories closely grouped together. Compared to the initial categorization based on OM content, RA and FK were more separated apart using the PCA evaluation. In fact, FK was similar to LI and BA in the first principal component (Fig. 8). More than 74 % of model results were explained by the first and second principal components.

Fig. 5 Daily (24-h) benthic fluxes of ammonium, nitrate, dissolved inorganic nitrogen (DIN), phosphate (DIP) and silicate (mean + SE, n = 3) in 6 bays in September.
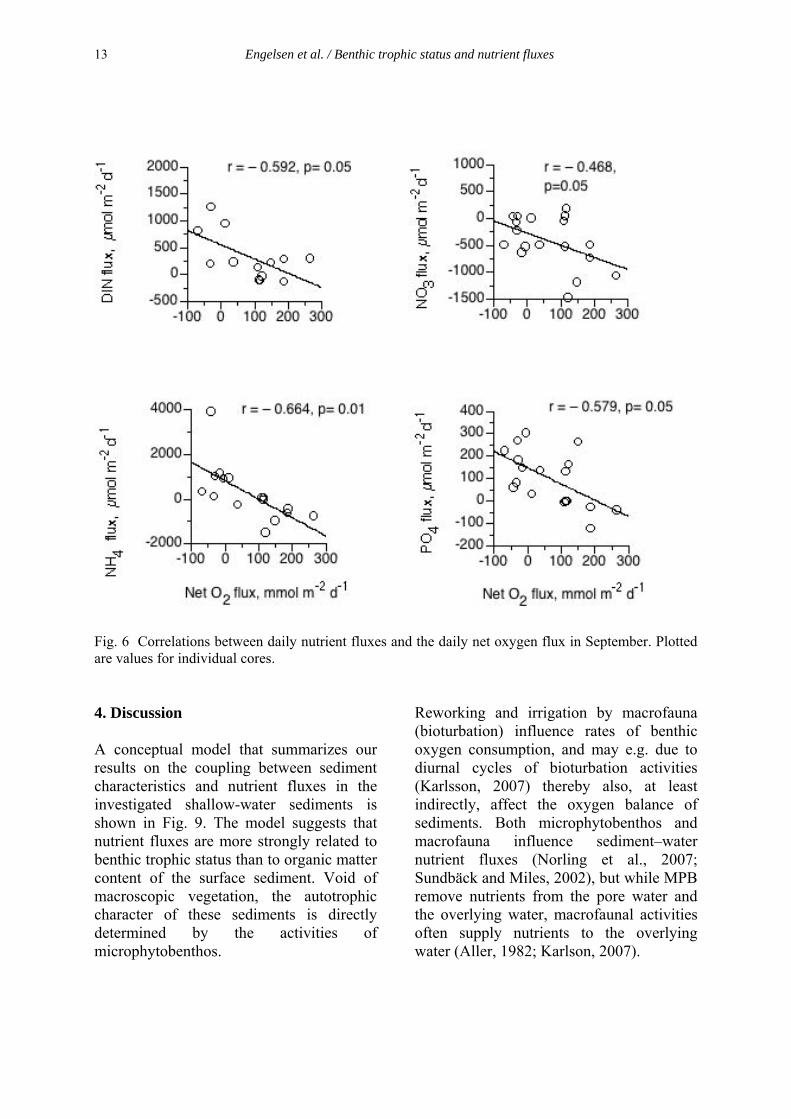
13 Engelsen et al. / Benthic trophic status and nutrient fluxes
Fig. 6 Correlations between daily nutrient fluxes and the daily net oxygen flux in September. Plotted are values for individual cores. 4. Discussion A conceptual model that summarizes our results on the coupling between sediment characteristics and nutrient fluxes in the investigated shallow-water sediments is shown in Fig. 9. The model suggests that nutrient fluxes are more strongly related to benthic trophic status than to organic matter content of the surface sediment. Void of macroscopic vegetation, the autotrophic character of these sediments is directly determined by the activities of microphytobenthos.
Reworking and irrigation by macrofauna (bioturbation) influence rates of benthic oxygen consumption, and may e.g. due to diurnal cycles of bioturbation activities (Karlsson, 2007) thereby also, at least indirectly, affect the oxygen balance of sediments. Both microphytobenthos and macrofauna influence sediment–water nutrient fluxes (Norling et al., 2007; Sundbäck and Miles, 2002), but while MPB remove nutrients from the pore water and the overlying water, macrofaunal activities often supply nutrients to the overlying water (Aller, 1982; Karlson, 2007).
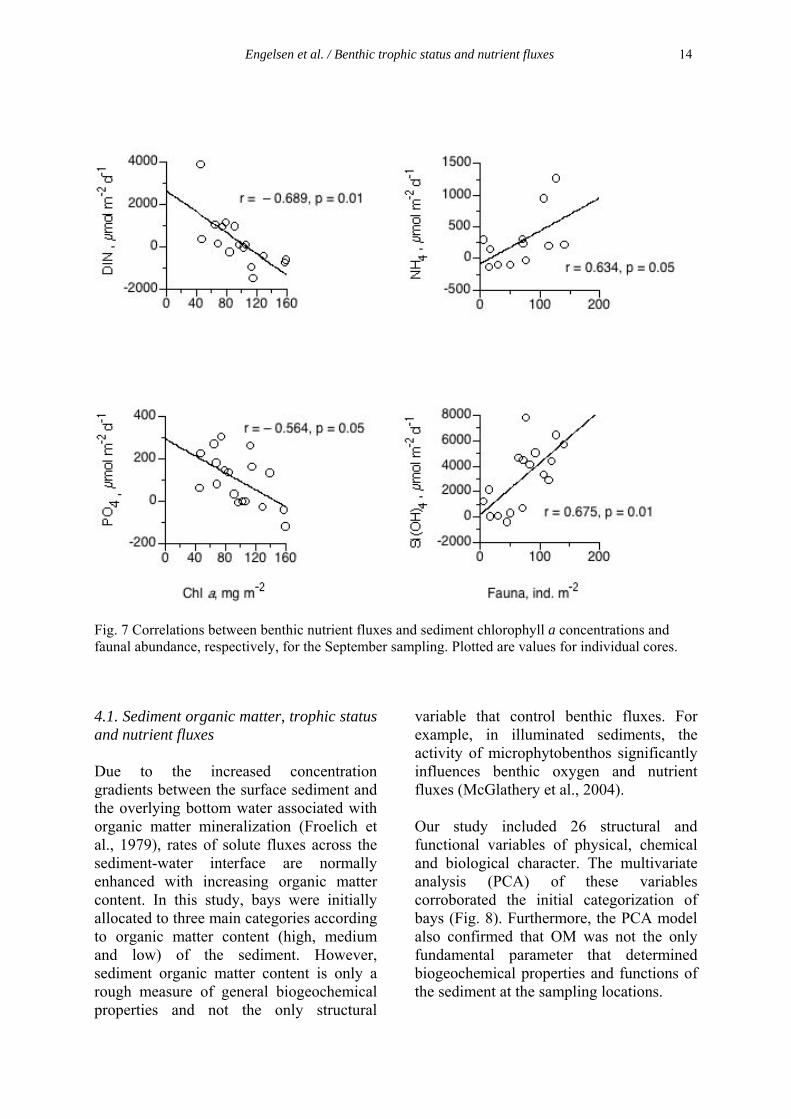
Engelsen et al. / Benthic trophic status and nutrient fluxes 14
Fig. 7 Correlations between benthic nutrient fluxes and sediment chlorophyll a concentrations and faunal abundance, respectively, for the September sampling. Plotted are values for individual cores. 4.1. Sediment organic matter, trophic status and nutrient fluxes Due to the increased concentration gradients between the surface sediment and the overlying bottom water associated with organic matter mineralization (Froelich et al., 1979), rates of solute fluxes across the sediment-water interface are normally enhanced with increasing organic matter content. In this study, bays were initially allocated to three main categories according to organic matter content (high, medium and low) of the sediment. However, sediment organic matter content is only a rough measure of general biogeochemical properties and not the only structural
variable that control benthic fluxes. For example, in illuminated sediments, the activity of microphytobenthos significantly influences benthic oxygen and nutrient fluxes (McGlathery et al., 2004). Our study included 26 structural and functional variables of physical, chemical and biological character. The multivariate analysis (PCA) of these variables corroborated the initial categorization of bays (Fig. 8). Furthermore, the PCA model also confirmed that OM was not the only fundamental parameter that determined biogeochemical properties and functions of the sediment at the sampling locations.

Fig. 8 Principal coordinates of the 8 stations investigated in June. The PCA (Principle Component Analysis) included all variables measured in both June and September.
For example, when compared with the initial categorization based on organic matter content alone, RA and FK were more dissimilar and separated apart by the PCA. This deviation supported the idea that
additional variables other than OM content (e.g. solid phase C and N, structural and functional properties associated with the macrofaunal community and net oxygen flux)
Nutrientflux
OM MA
MPB
Infauna
Trophicstate
Nutrientflux
OM MA
MPB
Infauna
Trophicstate
Fig. 9 Conceptual model for links between organic matter (OM) content, trophic status, benthic nutrient fluxes, macroalgae (MA), infauna and microphytobenthos (MPB) for shallow-water illuminated sediments regularly harbouring green-algal mats. The relative importance of suggested pathways is indicated by arrows of different thickness.
TJA
TJB
FK
RA
LIBA
BO
FI
-5
-3
-1
1
3
5
-5 -3 -1 1 3 5 PCA 1

were important for rates and direction of benthic nutrient fluxes observed in our study. Benthic trophic status, i.e. the balance between oxygen production and consumption in surface sediments, has during the last few years been suggested as a versatile indicator for evaluating whether sediments are net sources or sinks of nutrients (McGlathery et al., 2001a; Sundbäck et al., 2003; Tyler et al., 2003). In our study, nutrient fluxes correlated better with the net oxygen flux (trophic status) than with the organic matter content of the sediment. In these shallow-water sediments with no seagrass present, the autotrophic status is mainly determined by the photosynthetic activity of MPB. There was, for example, a significant negative correlation between the net oxygen flux and fluxes of DIN and DIP (Fig. 6). A significant negative correlation between biomass of MPB and fluxes of DIN and DIP further highlighted the importance of MPB for nutrient fluxes in these environments. Our study supports the idea that benthic trophic status, which reflects the photosynthetic activity of MPB, is a major property that determines benthic nutrient fluxes in shallow-water illuminated sediments. At our high latitude (59°N), however, such environments are strongly affected by seasonal variations in, for example, light and temperature. While all sediment types (except TJA and TJB) were net autotrophic in June, some were autotrophic (LI, BO and FI) and some (net) heterotrophic (FK, RA and BA) in September. Thus, sediments with a high OM content turned from autotrophic in spring to heterotrophic in autumn, whereas sediments with a low OM content seemed to remain their autotrophic character during both spring and autumn. These latter locations even increased their autotrophic status (to TOSI category 3) in autumn. This increase in the autotrophic status in September paralleled a higher Chl a content in the sandy sediments when compared
with the more silty locations, or when compared with the Chl a content in June. Therefore, not only the trophic status as such, but also the degree of heterotrophy or autotrophy (determined e.g. by the TOSI index) (Viaroli and Christian, 2004) has to be considered to accurately predict and evaluate benthic nutrient fluxes in shallow-water illuminated environments. As the benthic trophic status is a functional variable more closely coupled to nutrient fluxes than the comparably conservative structural parameter organic matter content, we suggest that the trophic status is a better predictor of benthic nutrient fluxes in these environments. 4.2. Macrofauna, MPB and benthic fluxes A primary effect of macrofaunal irrigation of tubes and burrows is to create diffusive sources or sinks for pore water solutes within otherwise isolated portions of sediments (Aller, 2001). Although there are conflicting observations in abandoned and infilled burrows e.g. of the polychaete worm Nepthys incise (Zhu et al., 2006), burrows not irrigated by fauna have normally no obvious effects on pore water distributions. Irrigated burrows, however, maintain local oxygenated conditions and diffusive concentration gradients between burrows and the surrounding sediment (Hulth et al., 2002; Mermillod-Blondin et al., 2005). In addition to a response pattern of solute distributions in the pore water, macrofaunal irrigation and reworking activities often enhance benthic fluxes (Emmerson et al., 2001; Marinelli and Williams, 2003). Although abundance, biomass or functional groups of macrofauna did not differ significantly between the three defined sediment types, there was still a positive correlation between macrofaunal abundance and fluxes of Si(OH)4 and NH4
+ at the investigated sites (Fig. 7). In non-illuminated sediments, Si(OH)4 is generally considered a conservative parameter, with concentration patterns mainly governed by

Table 2. Mean pore water concentrations (µM) of inorganic nutrients in the bays FK, BA, and FI representing low, medium and high organic matter content, respectively. NH4
+ NO3- DIP Si(OH)4
Sediment layer 0 – 0.5 5 – 7 0 – 0.5 5 – 7 0 – 0.5 5 – 7 0 – 0.5 5 – 7
High OM 21 40 24 13 1.0 2.0 61 67
Medium OM 11 8.0 14 7.0 0 0 16 94
Low OM 4.0 - 8.0 - 0 - 62 - the dissolution/precipitation of biogenic opal (SiO2) and diffusion/advection of Si(OH)4. Therefore, benthic Si(OH)4 fluxes are in bioturbated deposits commonly used as a viable measure of macrofaunal irrigation of tubes and burrows (Aller, 2001; Hammond et al., 1999; Marinelli, 1992). There was also a significantly higher abundance of macrofauna in September than in June, and macrofaunal abundance in September seemed to increase with increasing OM content of the sampling sites (Fig. 3b). This latter trend was associated with a considerable colonization of chironomid larvae at locations with a high OM content in the surface sediment. Grazing and the direct removal of MPB by chironomids may explain the lower NPP measured in September at the high-OM sites (Table 2, Fig. 4). Not only macrofaunal abundance, biomass, biovolume and functional biodiversity (Aller, 1982; Gilbert et al., 2007; Norling et al., 2007), but also the geometric scaling and diffusive distance between burrows are important parameters for organic matter mineralization and solute mobilization. For example, in the study by (Gilbert et al., 2003), both experimental and model observations demonstrated that rates of N mineralization, as well as the balance between stimulation and inhibition of denitrification, were highly related to the biogenic structure and the particular geometries of irrigated burrow distributions.
Benthic DIN fluxes were consistent with the general observation that high-OM sediments released and low-OM sediments retained nutrients from the overlying water. However, fluxes of DIP and Si(OH)4 did not follow this correlation, at least partly due to large variations in DIP and Si(OH)4 fluxes between bays within each category (Fig. 5). Although Si(OH)4 is often considered a conservative parameter for water transport, in shallow-water sediments, benthic Si(OH)4 fluxes are commonly influenced by activities of benthic diatoms (Marinelli, 1994). In our study, a significant positive correlation between fluxes of Si(OH)4 and macrofaunal abundance confirmed the importance of macrofaunal reworking activities for benthic Si(OH)4 fluxes (Fig. 7). A stronger correlation between macrofaunal abundance, rather than biomass, and benthic Si(OH)4 fluxes again suggested that geometric scaling and diffusive distance between burrows (Gilbert et al., 2003) are important for benthic nutrient fluxes. In contrast, a significant negative correlation between DIP fluxes and net oxygen fluxes (Fig. 6), as well as between DIP fluxes and Chl a content (Fig. 7), suggested that activities by MPB and the overall oxygen content controlled benthic DIP fluxes. In addition to direct utilization of DIP by MPB, oxygenation of the surface sediment and the formation of Fe(III)-oxide surfaces may have stimulated phosphate adsorption and the reversible removal of phosphate

Engelsen et al. / Benthic trophic status and nutrient fluxes 18
from the pore water (Sundby et al., 1992; Thamdrup, 2000). Thereby, DIP fluxes across the sediment-water interface were significantly reduced. The importance of MPB and benthic macrofauna for benthic nutrient fluxes was further exemplified at LI, one of the medium-OM sites. At this site, nutrients were more or less completely retained by the sediment system during incubations (e.g. [DIN] < 0.3 µM, [DIP] ~ 0 µM, [Si(OH)4] < 0.4 µM). Based on concentration gradients between the overlying water and the pore water (Table 2), a transport of solutes across the sediment-water interface was to be expected (at least for NH4
+, NO3-, and
Si(OH)4). However, net nutrient fluxes at LI were ~ 0. These low nutrient fluxes also coincided with a high biomass of MPB and a low biomass of macrofauna (mainly oligochaetes) (Table 2). The combination of a high biomass of MPB (active retention of nutrients) and a low abundance of irrigating macrofauna (restricted pore water advection through bioturbation) obviously counterbalanced the solute transport (diffusive and advective) across the sediment-water interface. 4.3. Benthic nutrient fluxes and macroalgal growth Benthic trophic status, described by the TOSI index or by the net oxygen flux, seemed a viable indicator of benthic nutrient fluxes in the investigated sediments (Fig. 6). A key question during our study was whether the trophic status also could be used as a viable parameter to predict the potential for macroalgal growth? Our study was made in September, i.e. during the late part of the normal growth season. The low DIN/DIP molar ratios (1 - 6) of the overlying water suggested conditions of N limitation, except in one bay (LI). Both N and P nutrient concentrations were, however, extremely low at LI. Further, the sites categorized as fully autotrophic
(category 3) did not supply DIN to the overlying water. The highest DIN flux from the sediment was observed at one of the net heterotrophic sites (FK; category 1; Fig. 5). None of the investigated shallow-water illuminated sediments were fully heterotrophic (TOSI category 0), as MPB were active during the light period (i.e. NPP > 0) at all sites. The actual uptake of inorganic nutrients by the green-algal mats was not measured directly, (e.g. by 15N; Cohen and Fong, 2005), but rather indirectly estimated from the increase of algal biomass in cores with only algae and overlying water from the sampling site (see Material and methods). As a net increase of biomass was not found at all sites, the algal nutrient demand was also calculated from daily NPP measured in the same cores. Both approaches yielded similar results, where the calculated nutrient demand for macroalgal growth varied between 8 and 28 mmol N m -2 d -1 and 0.3 and 1.1 mmol P m-2 d-1 (Table 3). For comparison, flux rates of DIN from the sediment ranged from 0 to 4.3 mmol N m-2 d-1, and from 0 to 0.2 mmol P m-2 d-1 (Fig. 5, Table 3). The proportion of DIN supplied from the sediment to support the calculated algal N demand was < 20 %. The highest contribution was found at FK, one of the net heterotrophic sites, where, depending on the mode of calculation, 15–20 % of the N demand was supplied from the sediment. The autotrophic sediments did not provide DIN to the overlying water. For comparison, utilizing the NPP measured in the sediment cores, a photosynthetic quotient (PQ) of 1.2, and a C/N ratio of 9 (e.g. Sundbäck et al. 2003), the estimated DIN demand of microphytobenthos was similar to that estimated for the macroalgae (9–25 mmol N m-2 d-1). The proportion of pore water DIP to support the algal P-demand was larger (9-70 %), except at two sites (LI and BO) where no efflux of P was observed (Table 3).

Table 3 Estimated range of N and P demand of ephemeral filamentous green algae (Ulva sp.), and sediment–water inorganic nitrogen (DIN) and phosphorus (DIP) fluxes obtained during 4 investigations on the west coast of Sweden. The macro-algal N and P demand was calculated from the increase of macro-algal biomass and measured algal N content. In the present study, assimilation of N was also calculated from net primary production (NPP), measured as oxygen production during sediment-water incubations. n.m. = not measured.
N demand (mmol N m-2 d-1)
Benthic flux (mmol N m-2 d-1)
N demand from the sediment (%)
P demand (mmol P m-2 d-1)
Benthic flux (mmol P m-2 d-1)
P demand from the sediment (%)
Reference Remarks
20–50 n.m 0.09–0.2 n.m. Pihl et al. (1996) Seasonal in situ growth (biomass)
5.3 n.m. 0.09 –0.2 n.m. Pihl et al. (1996) Seasonal growth (biomass) in field cages
2.3 1–2.9 42–100 0.08 0–0.06 0–16 Sundbäck et al. (2003) May–June, in situ growth, (biomass)
16–22
0–4.3
0–19 1.0–1.1 0–0.2 0.3–19 This study September, laboratory growth (biomass) under simulated in situ conditions
8–28 (mean 16.6)
0–4.3 0–15 0.3–1.0 (mean 0.62)
0–0.2
0–70 This study September, NPP (O2) in the laboratory under simulated in situ conditions

Our current estimations on the importance of sediments as a net source for nutrients to support macroalgal growth are slightly different compared with a study in the same area, conducted in spring and early summer. (Sundbäck et al., 2003) suggested that more or less the complete macroalgal N (but not P) demand could be covered by nutrients from heterotrophic sediments. The nutrient demand in spring and early summer was considerably lower than in our September study, but within the same range as has been estimated for macroalgal growth in several micro-tidal illuminated environments on the Swedish west coast (Table 3). The lower estimated uptake rate in spring was probably caused by the fact that algal growth was based on in situ increase of biomass over several weeks, thereby likely to a larger extent also including effects from grazing and degradation than was observed during our laboratory incubations. Our study suggests that, during the prevailing oligotrophic conditions, sediments may provide only part (but still significant) of the green-algal nutrient demand. In addition to nutrients supplied from e.g. adjacent water bodies, the biomass of green-algal mats may be sustained partly by internal circulation of nutrients within the mat itself (e.g. Sundbäck and MacGlathery 2005). However, for net growth additional nutrients have to be supplied from elsewhere. Sediments may provide an important nutrient source to ephemeral algae also under particular circumstances, such as low oxygen and anoxic conditions, e.g. under thick algal mats. Under temporary anoxic conditions in otherwise oxic bottom waters, surface sediments frequently release significant amounts of DIP and DIN (above all as NH4
+) stored in the surface sediment (Sundbäck and McGlathery, 2005). Since our incubations lasted ~ 1 hour in light and dark, respectively, pulses of inorganic
nutrients released from the sediment may have been excluded and not accounted for during the benthic flux measurements. Further, under natural conditions, periodic irrigation of macrofaunal burrows and hydrodynamic reworking of surface sediments may cause significant temporal variations during a 24-h period, including large pulses of pore water nutrients (Thybo-Christesen et al., 1993). Furthermore, dissolved organic nitrogen (DON) makes up a large fraction of the total dissolved nitrogen pool in shallow coastal waters (Sundbäck et al., 2006; Tyler et al., 2005; Welsh, 2003). This DON pool was not considered in our study as a source of nutrients for macro-algal growth. At low concentrations of DIN, macroalgae can effectively utilize DON as a source of nitrogen. (Tyler et al., 2005) showed that the green alga Ulva lactuca and the red alga Gracilaria spp. have a high affinity for urea and dissolved free amino acids. Similarly, DON may under low DIN concentrations also provide an important N-source for MPB (Sundbäck et al., 2006). Therefore, DON has to be taken into account in future studies when considering sediment–water coupling and algal nutrient demand in shallow waters. 5. Conclusion Our study suggests that benthic trophic status is a more viable parameter than sediment organic content to classify sediment type and predict benthic nutrient fluxes. Overall, autotrophic sediments remove nutrients from the overlying water column due to the activity of microphytobenthos, while net heterotrophic sediments supply nutrients to the overlying water. During the N-limited late growing season, even heterotrophic sediments appeared to provide only part of the N demand (at most 20 %), but up to 70 % of the P demand. Pore water and benthic fluxes of DON may be key components to understand the importance of benthic nutrient fluxes for macro-algal growth, and

21 Engelsen et al. / Benthic trophic status and nutrient fluxes
to fully account for the macro-algal nutrient demand in micro-tidal environments. Acknowledgements Financial support was received from the Swedish Research Council for Environment, Agricultural Science and Special Planning (FORMAS), the Swedish Research Council (VR), and the Marine Research Center of the Göteborg University (GMF). Jenny Krönström assisted during field sampling. References Aller, R.C., 1982. The effects of
macrobenthos on chemical properties of marine sediment and overlying water. In: McCall, P.L., Tevesz, M.J.S. (eds), Animal - Sediment relations, Plenum, NY 53-102.
Aller, R.C., 2001. Transport and reactions in the bioirrigated zone. In: Boudreau B., Jorgensen B.B. (eds) The benthic boundary layer: Transport processes and biogeochemistry. Oxford Press 269-301.
Astill, H. Lavery, P.S., 2001. The dynamics of unattached benthic macroalgal accumulations in the Swan-Canning Estuary. Hydrological Processes 15, 2387-2399.
Borum, J., 1996. Shallow waters and Land/Sea Boundaries. In: Eutrophication in Coastal Marine Ecosystems 179-204.
Cohen, R.A. Fong, P., 2005. Experimental evidence supports the use of delta N-15 content of the opportunistic green macroalga Enteromorpha intestinalis (Chlorophyta) to determine nitrogen sources to estuaries. Journal of Phycology 41, 287-293.
Dalsgaard, T., Nielsen, L.P., Brotas, V., Viaroli, P., Underwood, G.J.C., Nedwell, D.B., Sundbäck, K.,
Rysgaard, S., Miles, A., Bartoli, M., Dong, L., Thornton, D.C.O., Ottosen, L.D.M., Castaldelli, G. Risgaard-Petersen, N., 2000. Protocol handbook for NICE - Nitrogen Cycling in Estuaries: a project under the EU research program: Marine Science and Technology (MAST III). 62.
Emmerson, M.C., Solan, M., Emes, C., Paterson, D.M. Raffaelli, D., 2001. Consistent patterns and the idiosyncratic effects of biodiversity in marine ecosystems. Nature 411, 73-77.
Eyre, B.D. Ferguson, A.J.P., 2002. Comparison of carbon production and decomposition, benthic nutrient fluxes and denitrification in seagrass, phytoplankton, benthic microalgae- and macroalgae-dominated warm-temperate Australian lagoons. Marine Ecology Progress Series 229, 43-59.
Froelich, P.N., Klinkhammer, G.P., Bender, M.L., Luedtke, N.A., Heath, G.R., Cullen, D., Dauphin, P., Hammond, D., Hartman, B. Maynard, V., 1979. Early oxidation of organic matter in pelagic sediments of the eastern equatorial Atlantic: suboxic diagenesis. Geochimica et Cosmochimica Acta 43, 1075-1090.
Gilbert, F., Aller, R.C. Hulth, S., 2003. The influence of macrofaunal burrow spacing and diffusive scaling on sedimentary nitrification and denitrification: An experimental simulation and model approach. Journal of Marine Research 61, 101-125.
Gilbert, F., Hulth, S., Grossi, V., Poggiale, J.C., Desrosiers, G., Rosenberg, R., Gerino, M., Francois-Carcaillet, F., Michaud, E. Stora, G., 2007. Sediment reworking by marine benthic species from the Gullmar Fjord (Western Sweden): Importance of faunal biovolume.

Engelsen et al. / Benthic trophic status and nutrient fluxes 22
Journal of Experimental Marine Biology and Ecology 348, 133-144.
Glud, R.N., Kühl, M., Wenzhöfer, F. Rysgaard, S., 2002. Benthic diatoms of a high Arctic fjord (Young Sound, NE Greenland): importance for ecosystem primary production. Marine Ecology Progress Series 238, 15-29.
Hammond, D.E., Giordani, P., Berelson, W.M. Poletti, R., 1999. Diagenesis of carbon and nutrients and benthic exchange in sediments of the Northern Adriatic Sea. Marine Chemistry 66, 53-79.
Hansen, K. Kristensen, E., 1997. Impact of macrofaunal recolonization on benthic metabolism and nutrient fluxes in a shallow marine sediment previously overgrown with macroalgal mats. Estuarine Coastal and Shelf Science 45, 613-628.
Hodgkin, E.P. Birch, P.B., 1986. No simple solutions - proposing radical management options for an eutrophic estuary. Marine Pollution Bulletin 17, 399-404.
Hulth, S., Blackburn, T.H. Hall, P.O.J., 1994. Arctic Sediments (Svalbard) - Consumption and Microdistribution of Oxygen. Marine Chemistry 46, 293-316.
Hulth, S., Aller, R.C., Engstrom, P. Selander, E., 2002. A pH plate fluorosensor (optode) for early diagenetic studies of marine sediments. Limnology and Oceanography 47, 212-220.
Isaksson, I., Pihl, L. van Montfrans, J., 1994. Eutrophication-related changes in macrovegetaion and foraging of young cod (Gadus morhua L.): a mesocosm experiment. Journal of Experimental Marine Biology and Ecology 177, 203-217.
Karlson, K., 2007. Diurnal bioturbating activities of Monoporeia affinis: effects on benthic oxygen and
nutrient fluxes. Marine Ecology-Progress Series 331, 195-205.
Lavery, P.S. McComb, A.J., 1991. The nutritional eco-physiology of Chaetomorpha linum and Ulva rigida in Peel Inlet, western Australia. Botanica Marina 34, 251-260.
Lorenzen, C.J., 1967. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations. Limnology and Oceanography 12, 343-346.
Marinelli, R.L., 1992. Effects of polychaetes on silicate dynamics and fluxes in sediments -importance of species, animal activity and polychaete effects on benthic diatoms. Journal of Marine Research 50, 745-779.
Marinelli, R.L., 1994. Effects of burrow ventilation on activities of a terebellid polychaete and silicate removal from sediment pore waters. Limnol. Oceanogr 39, 303-317.
Marinelli, R.L. Williams, T.J., 2003. Evidence for density-dependent effects of infauna on sediment biogeochemistry and benthic-pelagic coupling in nearshore systems. Estuarine Coastal and Shelf Science 57, 179-192.
McGlathery, K.J., Anderson, I.C. Tyler, A.C., 2001a. Magnitude and variability of benthic and pelagic metabolism in a temperate coastal lagoon. Marine Ecology Progress Series 216, 1-15.
McGlathery, K.J., Berg, P. Marino, R., 2001b. Using porewater profiles to assess nutrient availability in seagrass-vegetated carbonate sediments. Biogeochemistry 56, 239-263.
McGlathery, K.J., Sundbäck, K. Anderson, I.C., 2004. The importance of primary producers for benthic nitrogen and phosphorus cycling. In: Nielsen SL, Banta GT, Pedersen MF (eds) Estuarine nutrient cycling: The

23 Engelsen et al. / Benthic trophic status and nutrient fluxes
influence of primary producers. Aquatic Ecology Ser 2 : 231-261
McGlathery, K.J., Sundback, K. Anderson, I.C., 2007. Eutrophication in shallow coastal bays and lagoons: the role of plants in the coastal filter. Marine Ecology-Progress Series 348, 1-18.
Melin, Y., 2001. Can marine filamentous algae be used as fertiliser? MSc thesis, Göteborg University
Mermillod-Blondin, F., Francois-Carcaillet, F. Rosenberg, R., 2005. Biodiversity of benthic invertebrates and organic matter processing in shallow marine sediments: an experimental study. Journal of Experimental Marine Biology and Ecology 315, 187-209.
Norling, K., Rosenberg, R., Hulth, S., Gremare, A. Bonsdorff, E., 2007. Importance of functional biodiversity and species-specific traits of benthic fauna for ecosystem functions in marine sediment. Marine Ecology Progress Series 332, 11-23.
Pedersen, M.F. Borum, J., 1997. Nutrient control of estuarine macroalgae: growth strategy and the balance between nitrogen requirements and uptake. Marine Ecology Progress Series 161, 155-163.
Pihl, L. Rosenberg, R., 1982. Production, abundance and biomass of mobile epibenthic marine fauna in shallow waters, western Sweden. Journal of Experimental Marine Biology and Ecology 57, 273-301.
Pihl, L., 1986. Exposure, vegetation and sediment as primary factors for mobile epibenthic faunal community structure and production in shallow marine soft bottom areas. Netherlands Journal of Sea Research 20, 75-83.
Pihl, L., Magnusson, G., Isaksson, I. Wallentinus, I., 1996. Distribution and growth dynamics of ephemeral macroalgae in shallow bays on the
Swedish west coast. Journal of Sea Research 35 (1 - 3), 169-180.
Pihl, L., Svenson, A., Moksnes, P.-O. Wennhage, H., 1999. Distribution of green algal mats throughout shallow soft bottoms of the Swedish Skagerrak archipelago in relation to nutrient sources and wave exposure. Journal of Sea Research 41, 281-294.
Raffaelli, D., 2000. Interactions between macro-algal mats and invertebrates in the Ythan estuary, Aberdeenshire, Scotland. Helgoland Marine Research 54, 71-79.
Rizzo, W.M., Dailey, S.K., Lackey, G.J., Christian, R.R., Berry, B.E. Wetzel, R.L., 1996. A metabolism-based trophic index for comparing the ecological values of shallow-water sediment habitats. Estuaries 19, 247-256.
Rumohr, H., Brey, T. Ankar, S., 1987. A compilation of biometric conversion factors for benthic invertebrates of the Baltic Sea. Baltic Marine Biologist 9, 1-57.
Strickland, J.D. Parsons, T.R., 1972. A practical handbook of seawater analysis. Bulletin of Fisheries Reasearch Board of Canada 169, 1-311.
Sundby, B., Gobeil, C., Silverberg, N. Mucci, A., 1992. The Phosphorus Cycle in Coastal Marine-Sediments. Limnology and Oceanography 37, 1129-1145.
Sundbäck, K. Miles, A., 2002. Role of microphytobenthos and denitrification for nutrient turnover in embayments with floating macroalgal mats: a spring situation. Aquatic Microbial Ecology 30, 91-101.
Sundbäck, K., Miles, A., Hulth, S., Pihl, L., Engström, P., Selander, E. Svenson, A., 2003. Importance of benthic nutrient regeneration during initiation of macroalgal blooms in

Engelsen et al. / Benthic trophic status and nutrient fluxes 24
shallow bays. Marine Ecology Progress Series 246, 115-126.
Sundbäck, K., Linares, F., Larson, F., Wulff, A. Engelsen, A., 2004. Benthic nitrogen fluxes along a depth gradient in a microtidal fjord: The role of denitrification and microphytobenthos. Limnology and Oceanography 49, 1095-1107.
Sundbäck, K. McGlathery, K.J., 2005. Interactions between benthic macroalgal and microalgal mats. In Kristensen, E., Haese, R.R., Kostka, J.E. (eds) Interactions between Macro- and Microorganisms in Marine Sediments, AGU series. Coastal and Estuarine Studies 60. Chapter 2, 7-29.
Sundbäck, K., Miles, A. Linares, F., 2006. Nitrogen dynamics in nontidal littoral sediments: Role of microphytobenthos and denitrification. Estuaries and Coasts 29, 6B, 1196-1211.
Thamdrup, B., 2000. Bacterial manganese and iron reduction in aquatic sediments. Advances in Microbial Ecology 16, 41-84.
Thybo-Christesen, M., Rasmussen, M.B. Blackburn, T.H., 1993. Nutrient fluxes and growth of Cladophora sericea in a shallow Danish bay. Marine Ecology Progress Series 100, 273-281.
Tyler, A.C., McGlathery, K.J. Anderson, I.C., 2003. Benthic algae control sediment-water column fluxes of organic and inorganic nitrogen compounds in a temperate lagoon. Limnology and Oceanography 48, 2125-2137.
Tyler, A.C., McGlathery, K.J. Macko, S.A., 2005. Uptake of urea and amino acids by the macroalgae Ulva lactuca (Chlorophyta) and Gracilaria vermiculophylla (Rhodophyta). Marine Ecology Progress Series 294, 161-172.
Underwood, A.J., 1997. Experiments in ecology: their logical design and
interpretation using analysis of variance. Cambridge. Cambridge University Press.
Valiela, I., McClelland, J., Hauxwell, J., Behr, P.J., Hersh, D. Foreman, K., 1997. Macroalgal blooms in shallow estuaries: Controls and ecophysiological and ecosystem consequences. Limnology and Oceanography 42, 1105-1118.
Welsh, D., 2003. It's a dirty job but someone has to do it: The role of marine benthic macrofauna in organic matter turnover and nutrient recycling to the water column. Chemistry and Ecology 19(5), 321-342.
Viaroli, P., Bartoli, M., Bondavalli, C., Christian, R.R., Giordani, G. Naldi, M., 1996. Macrophyte communities and their impact on benthic fluxes of oxygen, sulphide and nutrients in shallow eutrophic environments. Hydrobiologia 329, 105-119.
Viaroli, P. Christian, R.R., 2004. Description of trophic status, hyperautotrophy and dystrophy of a coastal lagoon through a potential oxygen production and consumption index--TOSI: Trophic Oxygen Status Index. Ecological Indicators 3, 237-250.
Zhu, Q.Z., Aller, R.C. Fan, Y.Z., 2006. Two-dimensional pH distributions and dynamics in bioturbated marine sediments. Geochimica et Cosmochimica Acta 70, 4933-4949.
Österling, M. Pihl, L., 2001. Effects of filamentous green algal mats on benthic macrofaunal functional feeding groups. Journal of Experimental Marine Biology and Ecology 263, 159-183.

- 1 -
Factors influencing initiation and growth of greenalgal mats
Anna Engelsen1 & Fredrik Larson2
1 Department of Marine Ecology, Göteborg University, Kristineberg Marine Research Station, 450 34
Fiskebäckskil, Sweden 2 Department of Marine Ecology, marine botany, Göteborg University, P.O. box 461, 405 30 Göteborg, Sweden
ABSTRACT: Fast growing ephemeral green algal mats have become much more common in coastal areas during later decades. Despite efforts to decrease anthropogenic nutrient loading, algal mats are still increasing. One suggestion is that shallow-water sediments have been accumulating nutrients for decades and thereby have become a source of nutrients, which function as a self-regenerating system during the initiation and growth of ephemeral green algal mats. The hypothesis that sediments are more important as a nutrient source than the water column was tested in a flow-through system using 200 mm and 5 mm deep sediments, with two types of sediment; high (silty) respectively low (sandy) content of organic matter. Growth of green algae, sediment-water nutrient and oxygen fluxes, benthic microalgal biomass, infauna, meiofauna and sediment characteristics were measured during six weeks. The results showed an intense growth of green algae only in the treatment with silty surface (5 mm) sediment of high organic matter content. Possible explanations are the exclusion of potential grazers, such as larger infauna, in the 5 mm sediment and/or nutrient competition with benthic microalgae and cyanobacteria in the 200 mm thick sediment. The conclusion from our data is that ephemeral green algae are able to grow intensely with generally low amounts of nutrients supplied by the water column and grazing by Nereis diversicolor and competition from benthic microalgae for nutrients are factors in the sediment that may prevent initiation of growth of these green algal mats. KEYWORDS: Ephemeral green algal mats, sediment, Nereis diversicolor, grazing, nutrient fluxes, benthic microalgae
1. Introduction Mats of green algae are a common
phenomenon in shallow coastal habitats around the world (Bonsdorff, 1992; Valiela, et al., 1997; Bäck, et al., 2000; Vahteri, et al., 2000; Nelson, et al., 2003; Sundbäck, et al., 2003).
*) Corresponding author: [email protected]
On the Swedish west coast, the mats usually bloom from June to September and mainly consist of filamentous Ulva spp. (former Enteromorpha spp.) and Cladophora spp. (Pihl, et al., 1999; Sundbäck, et al., 2003). It has been suggested that embayments harbouring these algal mats turn into self-generating systems (Norkko and Bonsdorff, 1996a; Pihl, et al., 1996; Sundbäck, et al., 2003) where nutrients are released from the

INITIATION OF ALGAL MATS
- 2 -
sediment, and incorporated into the biomass of algae during the summer bloom. Later, the algae are remineralised via decay in the sediment, where the nutrients constitute a nutrient pool for future production of algal mats.
It has been shown that the highest uptake rate of ammonium is in the lower part of the algal mat, probably depending on a nutrient supply from the sediment (Thybo-Christensen, et al., 1993), and the assimilation of nitrogen depends on light conditions (McGlathery, et al., 1997).
A high irradiance may lead to an uptake as high as 900 µmol NH4 m-2 h-1 and also prevent ammonium diffusion from the benthic nutrient source into the overlying water (McGlathery, et al., 1997). These filamentous algae also have low nutrient storage abilities; the acquired nutrients appear to be allocated directly into growth rather than to storage reserves (McGlathery, 1992). They also have a high surface area to volume ratio and a fast nutrient uptake (Wallentinus, 1984) which leads to a rapid growth. The result of this is that they outcompete slower growing perennial algae such as e. g. fucoids, and they need a continuous supply of inorganic nutrients to be able to grow. Benthic microalgae (BMA) can effectively reduce inorganic nutrients efflux from the sediment during the period that is critical for the initiation of the growth of macroalgal mats (Sundbäck and Miles, 2002).
If the development of algal mats is
mainly dependent on the sediment efflux of remineralised nutrients, the benthic microalgae (BMA) might delay, or even prevent, the mat to develop (Sundbäck and Miles, 2002).
It is well known that animals through
their bioturbation stimulate decomposition of organic material and enhance the microbial activities in the sediment (Jensen, 1996; Aller and Blair, 2004; Karlson, et al., 2005; Michaud, et al.,
2006). Algal mats may create conditions of hypoxia/anoxia (Sundbäck, et al., 1990; Thybo-Christesen, et al., 1993; Cummins, et al., 2004), defaunating areas for long periods. This may lead to high pore-water pools of mineralization products, e.g. ammonium, in the sediment (Hansen and Kristensen, 1997) and after recolonization of macrofauna the bioturbation and irrigation will, for shorter periods, result in massive effluxes of nutrient rich pore-water (Hansen and Kristensen, 1997). It is still not known whether algal mats during the initiation mainly depend on nutrients coming from the sediment or from the water column, but there are indications that the algae need the regenerated inorganic nutrients that come from the sediment (Hodgkin and Birch, 1986; Pihl, et al., 1999).
Our aim was to investigate the
importance of sediment nutrient release for the initiation of growth of algal mats and to study the effect of grazers. Our first hypothesis is that the sediment is the main source of inorganic nutrients during the initiation of algal growth. The idea is that remineralised inorganic nutrients during spring, when macrofauna activities start and temperature and light increase, are released from the sediment and constitute the main nutrient source. Our second hypothesis was that efficient grazing from macrofauna could control the initiation of macroalgal mats.
We used a flow-trough system, two types
of sediment and two different thicknesses (5 mm and 200 mm) of the sediments as treatments, where the 200 mm represented the natural sediment with a nutrient source and fauna and the 5 mm sediment represented the sediment without fauna and a nutrient source in the sediment. Variables measured were growth rate of green algae, primary production (oxygen release) sediment-water fluxes of inorganic nutrients, biomass of BMA, abundance and biomass of macrofauna and meiofauna, as

INITIATION OF ALGAL MATS
- 3 -
well as sediment characteristics. The experiment lasted for 8 weeks.
2. Materials and Methods The study took place on the west coast of
Sweden (58º 15′ N, 11º 30′ E) from early June until the end of July in 2003. The area is micro-tidal with average tidal amplitude of 20 – 30 cm. During summer, shallow (0 – 1 m) coastal waters on the west coast of Sweden have a salinity between 20 and 28, and the water temperature is approximately 20 to 25°C (Pihl and Rosenberg, 1982). Undisturbed sediments were collected in the beginning of June from two different bays; one sandy bay with low (1 %) content of organic material in the sediment and a C/N of 7.2, and one silty bay with high (14 %) content of organic material and C/N of 8.6 (Engelsen et al. submitted). From each bay, 12 cores (i.d. = 250 mm, height 270 mm) holding sediment with a height of 200 mm (deep sediment, DS), and 12 cores (i.d. = 250, height 270 mm) with surface sediment (SS) of just 5 mm was sampled. The SS cores had the upper 5 mm sliced off and spread out on a plate which then was placed at the same height as the sediment surfaces of the DS cores, so that all sediment cores had the same height of the overlying water column.
The DS cores contained natural densities of benthic macrofauna, whereas macrofauna was excluded from SS cores. All cores were randomly placed in shallow basins with a flow-trough system with in situ water (10 L h-1).Seawater was pumped from 1 m depth in the adjacent bay to get an “ambient” content of inorganic nutrients. Each core was supplied with an individual flow of water so that in every core there was a water column of 5 cm overlying the sediment. Through turbulence the individual flow also mixed the water column in each core. The water column in each core overflowed the edge of the core and ended up in the basin. The flow-through system also functioned as a cooling system so that the temperature (22°
C) of the sediment was maintained during the experiment. The whole system was kept in a greenhouse in ambient light conditions (the light was reduced by 25 %) with dawn and twilight.. The experiment started on the 6th of June with an initial sampling and then was left for three weeks before the second sampling; only photos were taken every week. During the experiment, there were six sampling occasions, i.e. one per week, including four destructive sediment samplings and two water samplings. During the destructive sampling, both water samples and samples from the sediment were taken and after sampling the sediment was sieved. Water sampling only included samples for nutrient and oxygen fluxes. At each sampling, six cores from each sediment type were randomly chosen; three DS and three SS cores.
Each core from both SS and DS treatments had a natural distribution of propagules from the start. Percentage macroalgal cover was observed visually and through photo documentation. Photos were taken 6 times during the development of algal growth; 3 with just a few days between to monitor the start of the growth and 3 with approximately one week in between when the development had slowed down. At each destructive sampling also the wet weight, and after drying, the dry weight, were used for algal biomass estimation. The macroalgae were dried at 60 °C for 6 days. In addition, macroalgal nutrient demand was calculated based on net primary production (NPP) where oxygen was converted to C by using the photosynthetic quotient of 1.2 (Glud, et al., 2002).
Sediment cores from each bay were sampled for flux measurements of oxygen and inorganic nutrients (NH4
+, NO32-,
PO43- and Si(OH)4). The same cores were
used for both oxygen and nutrient fluxes which were measured both during the daylight (~12.00 a.m.) and during the dark period (~12.00 p.m.). During incubations,

INITIATION OF ALGAL MATS
- 4 -
cores were closed by Plexiglas lids and the individual water flow of each core was turned off. The incubation period lasted for approximately one hour during the light period, and two hours during the dark period. Oxygen concentrations were not allowed to change > 20 % during incubation, to avoid oxygen-induced changes in nutrient fluxes. Sundbäck et al. (2003) showed that in situ incubations gave similar results as incubations in the laboratory, and for practical reasons the incubations were therefore performed in the laboratory. Glass syringes (20 ml) were used to take oxygen samples, and disposable plastic syringes (20 ml) were used to take nutrient samples. Both types of syringes had tubing that fitted a hole in the transparent Plexiglas lid. Samples for time zero were taken from open cylinders and at the end of the incubation through a 5 mm opening in the lid. The method and process is described in Dalsgaard et al. (2000).
The water samples were analysed for dissolved oxygen by Winkler titration (Strickland and Parsons, 1972), while the samples for nutrients were filtered on-line (0.45 µm Sartorius syringe filters), and stored frozen (-80ºC) until analysis by wet-chemistry methods with a nutrient auto-analyzer (TrAAcs 2000, Bran+Luebbe, Norderstedt, Germany). Daily rates of nutrient fluxes were calculated by multiplying the hourly rate in light times the number of daylight hours and the hourly rate in the dark times the number of dark hours and adding these two values. Daily net primary production (NPP) was calculated by multiplying daytime flux rates per incubation time by a light factor calculated as a ratio between total daily irradiance (light data from the Swedish Meteorological and Hydrological Institute) and irradiance during the incubation period. Measurements of inorganic nutrients in the incoming water from the bay were analysed twice a week by sampling water from the common inlet for all cores.
After sampling for nutrients and oxygen fluxes, chlorophyll a and meiofauna, benthic macrofauna was collected from the cores by sieving the sediment through a mesh size of 1 mm. The fauna was identified to lowest taxonomic level possible, counted for abundance and the wet weight was used for biomass estimation. Biomass of shell-bearing macrofauna was corrected for shell weight (25%). Values were recalculated to show abundance and biomass per square meter.
Meiofauna was sampled with a cut-off 2 ml syringe (i.d. = 8.7 mm) from all treatments (n = 3). The top 5 mm from both the 200 mm and the 5 mm sediment cores were sampled and put in 4% borax buffered formalin containing rose Bengal and stored at +8°C until analysis. The organisms were extracted by decantation. The supernatant was poured through a sieve with mesh sizes of 100 µm. The meiofauna was counted under a dissecting microscope and sorted into major taxa.
Chlorophyll a was measured as a rough estimate of the benthic microalgal biomass (n = 3). The top 5 mm of the sediment was sampled with a cut-off 2 ml syringe (i.d. = 8.7 mm) and immediately frozen (-18°C) until analysis. All samples were extracted for 24 hours in 4 ml 90 % acetone in the fridge (+8°C), thereafter centrifuged. Analysis of the supernatant was carried out according to Lorenzen (1967) and Bartoli et al. (2003). For more details see Dalsgaard et al. (2000).
Samples for benthic microalgal (BMA) species were taken with a cut-off 2 ml syringe (i.d. = 8.7 mm) (n = 3), preserved in glutaraldehyde (2.5%) and put in the fridge (+8°C) until analysis. Microalgal composition was studied under a microscope and classified to genera.
Two-way ANOVA with day and treatment as fixed factors was used and in cases of significant interactions, Tukey´s post-hoc test was used to test on what days there were differences between the

INITIATION OF ALGAL MATS
- 5 -
treatments. Tests were done between sediment thicknesses within each type of sediment. Homogeneity of variances was checked using Cochran’s test. Differences were accepted as significant at p < 0.05.
3. Results
During the experiment, a macroalgal (Ulva spp.) mat developed only in the cores that contained 5 mm of silty surface sediment (silty SS cores). No macroalgal mat was observed in sandy SS, sandy DS or silty DS (Fig. 1, “July 23”). The silty SS had an early initiation of growth (day 20), and over time a thick green algal mat developed. The sandy SS, however, showed during the later part of the experiment a cyanobacterial growth that gave a blue-green colour to the surface (Fig. 1, “July 7”). The macroalgal mats in the silty SS had a fresh green colour during the beginning of the experiment. Oxygen bubbles were formed resulting in mats that were floating (Fig 1, “July 15).Towards the end of the experiment, when the macroalgal mat had become dense it turned yellow and started to degrade (Fig. 1, “July 23”). At this stage the macroalgal mat was dense and connected, and below the mat the sediment turned from a previously brownish to a pale grey colouration.
The growth of macroalgae in the silty SS started in the end of June (Fig. 1, “June 27”), three weeks after the start of the experiment, and within 4 days it covered between 2-10 % of the surface (Fig. 1, “July 1”). Two weeks from the start of the growth, the percentage of surface cover had increased up to 40-80% (Fig. 1, “July 3”), and after further one week the coverage was 100% in all silty SS replicates (Fig. 1, “July 7”). During the following two weeks the mat grew thicker (Fig. 1, “July 15”) and finally degraded and started to break up into fragmented filaments (Fig. 1, “July 23”). During the first two weeks the mean daily growth was 1.3 g m-2 d-1, the second two weeks the
mean daily growth increased to 6.3 g m-2 d-
1 and thereafter decreased to 2.03 g m-2 d-1 the last two weeks (Fig. 2).
Oxygen fluxes (NPP) differed significantly between treatments (DS and SS) and over time for silty sediments but only between treatments for sandy sediments) (Table 1). All treatments except silty DS showed oxygen production throughout the experiment (Fig. 3), where silty SS had the highest production during the last three samplings. Silty DS mainly showed oxygen consumption on the three first sampling occasions, but turned into production on the three last sampling occasions. Both silty and sandy SS appeared to have higher production compared to silty and sandy DS (Fig. 3). Community respiration did not differ significantly between treatments or over time. Highest respiration rate was found on the fourth sampling (9th of July) in all treatments, and in general, higher CR occurred at the end of the experiment.
Daily nutrient fluxes differed significantly between treatments and over time except for nitrate, where no significance could be found (Table 1). A consistent pattern was the release of ammonium throughout the experimental period. Also sandy and silty DS mainly showed a release of silicate, while SS treatments showed a low release in the beginning of the study which increased at the end DS (Fig. 4). The sandy SS showed an uptake of ammonium during the experimental period while the silty SS that had a small outflow (Fig. 4). Most significant differences were found during the light period , where interactions appeared between time and treatment (Table 1).

INITIATION OF ALGAL MATS
- 6 -
Fig. 1. Photos of a time series showing the absence/presence of growth of ephemeral green
algae and cyanobacteria from sandy and silty DS (deep sediment, 200 mm) and SS (surface sediment, 5 mm) cores.

INITIATION OF ALGAL MATS
- 7 -
0
20
40
60
80
100
120
140
160
June 16 July 1 July 15 July 29
Alg
al b
iom
ass
g dw
m-2
Sampling day
sand
silt
Fig. 2. The biomass and growth rate of the ephemeral green algae, both based on
dry weight, from the 16th of June until the 29th of July. Shown for the biomass are mean values + SE (n = 3).
-400
0
400
800
1200
1600
2000 Sand (DS)
-400
0
400
800
1200
1600
2000 Sand (SS)
-400
0
400
800
1200
1600
2000
17 24 1 9 15 23
Silt (DS)
-400
0
400
800
1200
1600
2000
17 24 1 9 15 23
Silt (SS)
NPP
mm
ol O
2m
-2d-1
Date
Fig. 3a. Daily net primary production (NPP) for sandy and silty DS and SS cores (see legend for Fig. 1). Shown are mean values + SE (n = 3).

INITIATION OF ALGAL MATS
- 8 -
-1200
-1000
-800
-600
-400
-200
0
Sand (DS)-1200
-1000
-800
-600
-400
-200
0
timeSand (SS)
CRm
mol
O2
m-2
d-1
-1200
-1000
-800
-600
-400
-200
017 24 1 9 15 23
Silt (DS)-1200
-1000
-800
-600
-400
-200
0
Silt (SS)17 24 1 9 15 23 17 24 1 9 15 23
Date Fig. 3b Daily community respiration (CR) for sandy and silty DS and SS cores (see legend
for Fig. 1). Shown are mean values + SE (n = 3). Table 1. Results from 2-way ANOVA with day (D) and treatment (T) as factors where the
treatment is thickness of sediment (5 or 200 mm) within each sediment type. In cases of significant interactions between treatment and day, Tukey´s post-hoc test indicates on which days there were significant effects.
SILT SAND Functions Light
Factor F p Significant on days:
Factor F p Significant on days:
Ammonium T 12.67 0.02 T 11.484 0.02 Nitrate ns ns Phosphate DxT 2.675 0.05 4, 6 ns Silicate DxT 2.732 0.04 1 ns NPP DxT 4.052 0.008 1, 5, 6 T 34.682 0.0001 Functions Dark
Ammonium DxT 11.695 0.0001 1,3,4,5,6 T 4.498 0.044 Nitrate ns ns Phosphate T 6.89 0.015 ns Silicate ns ns Respiration ns ns
Structure Chlorophyll a DxT 10.69 0.002 5, 6 DxT 6.508 0.012 6 Meiofauna T 4.733 0.05 ns

INITIATION OF ALGAL MATS
- 9 -
The incoming bay-water had nutrient levels of <1 µM nitrate, <0.1 µM ammonium, <0.1 µM phosphate, which given the flow of 10 L cyl.-1 h-1 and a cylinder area of 0.049 m2, gives a contribution of <500 µmol m-2 d-1 of NO3
-, <50 µmol m-2 d-1 of NH4
+ and <50 µmol m-2 d-1 of PO4
3-. The calculated N demand for the macroalgae was between 7 and 30 mmol N m-2 d-1.
Chl a, which was used as a rough measure of benthic microalgal (BMA) biomass, differed significantly between the two sediment types. Differences were found between sediment thickness of sand and silt as well as over time for sand DS, sand SS, and silt DS but not for silt SS which had an invariable biomass of BMA throughout the experiment. Sandy SS had an increase in Chl a concentration at the end of the investigation, whereas concentrations in the sandy DS first decreased but again increased in the end. Silty SS retained the biomass during the experiment, while in silty DS it decreased (Fig. 5).
Cyanobacteria dominated the BMA at
the sediment surface of the sandy sediment; both in DS and SS. Common were Oscillatoria sp., Merismopedia sp. and sp. Also large sigmoid diatoms were found. Both silty SS and DS sediment where dominated by naviculoid attached diatoms of different size classes, but no cyanobacteria were found.
Main groups of meiofauna (> 100 µm) found included polychaete larvae, nematodes, harpacticoid copepods, ostracodes and foraminifers. Highest abundance was found for nematodes (1100 - 1550 ind. m-2) and foraminifers (470 - 3900 ind. m-2), followed by harpacticoid copepods (340 - 1270 ind. m-2) and
ostracodes (50 – 165 ind. m-2) whereas polychaete larvae only showed 2 – 30 ind. m-2 (Fig. 6). Significant differences were found between treatments in the silty sediment for both DS and SS, concerning % of total abundance (Table 1). Sandy sediments showed a significantly higher percent of foraminifers (7-8 %) and harpacticoid copepods (2-3 %), while the silty sediment had a significantly higher percentage of ostracodes (6-9 %). No difference in meiofauna abundance between sediment depths was found (Table 1).
The dominating species of macrofauna was Nereis diversicolor, which had the significantly highest mean abundance of macrofauna. N. diversicolor mean abundance for sandy DS was approximately 450 individuals m-2 and for silty DS 350 individuals m-2 (Fig. 7). The mean biomass of N. diversicolor reached 60 g m-2 in the sandy DS and 100 g m-2 in the silty DS. The three bivalve molluscs Cerastoderma edule, Macoma baltica and M. arenaria had a mean abundance between 20 and 40 individuals m-2 in the sandy DS and between 0 and 60 individuals m-2 in the silty DS M. arenaria had the highest mean biomass; 550 g m-2 in the sandy DS and 700 g m-2 in the silty DS.
The other two bivalve species had mean
biomasses ranging from 70 to 100 g m-2.. The polychaete Pygospio elegans was found only in sandy DS and had a mean abundance of almost 300 individuals m-2. The biomass of P. elegans was 20 g m-2 in the sandy DS. Species from the gastropod family Hydrobidae, and crustacean family Mysidae and the genus Corophium was mainly found in the silty DS.

- 10 -
Ammonium
Nitrate
Silicate
-1500
-1000
-500
0
500
1000
1500um
ol N
H4
m-2
d-1
Sand DSSand SS
-100100300500700900
110013001500 Silt DS
Silt SS
-900
-700
-500
-300
-100
100
300
umol
NO
3m
-2d-
1
-900
-700
-500
-300
-100
100
300
-350
-250
-150
-50
50
150
umol
PO
4m
-2 d
-1
Phosphate
-350
-250
-150
-50
50
150
-1500
500
2500
4500
6500
8500
17 24 1 9 15 23
umol
Si(O
H) 4
m-2
d-1
Date
-1500
500
2500
4500
6500
8500
17 24 1 9 15 23Date
Fig. 4. Daily fluxes of nutrients (NH4
+, NO3-, PO4
3- and Si(OH)4) for sandy and silty DS and SS cores (see ledgend for Fig. 1). Shown are mean values + SE (n = 3).

INITIATION OF ALGAL MATS
- 11 -
Chlorophyll a in sediment
mg
Chl
am
-2
June 16 July 1 July 15 July 29 June 16 July 1 July 15 July 29
Fig. 5. Chlorophyll a concentration of the top 5 mm sediment for sandy and silty DS and SS
cores (see ledgend for Fig. 1). Shown are mean values + SE (n = 3).
0
1000
2000
3000
4000
5000
6000
Polych
aeta
Nematoda
Harpacti
coid
cope
pod
Ostrac
oda
Foraminife
ra
Mei
ofau
na, i
nd. m
-2
silt SS silt DS sand SS sand DS
Fig. 6 The abundance of major meiofauna groups in the sandy and silty DS and SS cores
(see legend for Fig. 1). Shown are mean values +SE (n = 3).

INITIATION OF ALGAL MATS
- 12 -
0
100
200
300
400
500
600
Nereis d
iversi
color
Cerastode
rma e
dule
Mya ar
enaria
Macom
a balt
ica
Hinia retic
ulata
Pygosp
io elega
ns
Mac
rofu
na, i
nd. m
-2
SandSilt
Fig. 7 The abundance of the major macrofauna species in the sandy and silty DS sediment
(see legend for Fig. 1). Shown are mean values +SE (n = 3).
4. Discussion
Macroalgal mats started to grow in the silty SS cores (Fig 1). This was surprising, since the expectation was to find growth in the silty DS cores because of the nutrient release from the sediment. The macroalgal mats in the silty SS cores started to grow in early July, which is normal for algal mats in situ on the west coast of Sweden (Pihl, et al., 1996; Pihl, et al., 1999). No macroalgal growth, however, was seen in any other treatment.
The two sediment types gave fluxes of nitrogen and phosphate, which were within the same range as measured fluxes in other studies from the area (Sundbäck, et al., 2000; Sundbäck, et al., 2003). The incoming bay-water generally had low nutrient levels, rendering input rates of <500 µmol m-2 d-1 of NO3
-, <50 µmol m-2 d-1 of NH4+ and <50
µmol m-2 d-1 of PO43-), but compared to the
release from e.g. the silty DS (60 µmol m-2 d-1 of NO3
-, 700 µmol m-2 d-1 of NH4+, and
25 µmol m-2 d-1 of PO43-) the incoming water
had higher amounts of NO3- and PO4
3-. Silty DS seems to provide higher amounts of ammonium than the incoming water, but not high enough to be able to entirely support the algal growth. The calculated N demand in this study was between 7 and 30 mmol N m-2 d-1 during the initiation of growth. Other studies have shown similar (2 – 50 mmol N m-2 d-1) demands based on calculations (see summary in Engelsen et al. submitted). It seems that the N demand of green macroalgae is higher than the supply from the sediment.
However, other studies have shown that the sediment is capable of being the main source of nutrients for ephemeral algal mats (Hodgkin and Birch, 1986; Astill and Lavery, 2001; Sundbäck, et al., 2003). The different views could be a result of variations in measured flux rates. Flux rates of nutrients from the sediment varies due to infaunal activity, resulting in periods of large pulses of ammonium (Hansen and Kristensen, 1997). The growth of macroalgae might be dependent on both larger pulses of nutrients coming from the

INITIATION OF ALGAL MATS
- 13 -
sediment, but also on continuous water exchange in shallow areas and water runoff after periods of rain. The water column in shallow areas on the west coast of Sweden normally have very fluctuating nutrient concentrations (Pihl, et al., 1996), since the exchange of water varies during the day mainly depending on tidal exchange, air pressure and wind driven currents (Söderkvist, 1997; Dubnyak and Timchenko, 2000; Valle-Levinson, et al., 2001).
Macroalgal mats did not develop on sandy
or silty DS. Both treatments contained high densities of Nereis diversicolor, and the lack of algal growth could be due to grazing by these polychaetes. Some studies point to N. diversicolor as a herbivore, feeding on fresh green algae (Meziane and Retiere, 2002; Nordström, et al., 2006). The limit for significant loss of biomass of green algae promoted in presences of N. diversicolor seemed to be density dependent, where the threshold is between 35 and 105 ind. m-2
(Nordström, et al., 2006). In this study the mean abundance of N. diversicolor was 350 (sandy sediments) and 450 (silty sediments) ind. m-2, which supports the suggestion that the algal growth was inhibited by polychaetes. Also a pilot study (Engelsen and Larson, unpublished) showed significant grazing on green algae with small variations in both 200 mm (DS) and 5 mm (SS cores) sediments in presence of N. diversicolor. Although the biomass of N. diversicolor has been related to macroalgal biomass (Raffaelli, 2000; Nordström, et al., 2006) and increases in biomass of adult worms living below algal mats has been recorded (Norkko and Bonsdorff, 1996b), the impact of N. diversicolor as a modulator on green algal mats has not yet been investigated or discussed.
The cores with sandy SS and DS did not show any growth of macroalgae, but instead a cyanobacterial mat developed on the sandy sediment. It has been found that Benthic microalgae (BMA) (Sundbäck and Miles, 2002), and especially cyanobacteria (Fong,
et al., 1993) are good competitors for nutrients, and are able to outcompete fast growing green algae. The cyanobacterial community was not affected by the grazing by Nereis diversicolor, since they have been found to reject cyanobacteria as a food source (Vedel and Riisgard, 1993; Olivier, et al., 1995). It is also possible that BMA efficiently reduce the nutrients within the boundary layer between the water column and the sediment, thereby preventing the spores and propagules of the ephemeral green algae to initiate their growth. Thereby, green algal mats actually are subjected to both the threat of grazing from N. diversicolor and competition for inorganic nutrients from BMA, and especially from cyanobacteria. The cyanobacterial mat increased over time in the sandy SS, while cyanobacterias in the sandy DS decreased but, again increased at the end of the experiment (Fig. 5). One explanation to the more visible cyanobacterial mat and higher biomass in cores of sandy SS compared to sandy DS, could be that sediment surfaces without influence from macrofauna (SS) became an excellent habitat to grow in, whereas a sediment surface with grazing and bioturbation (DS) constitutes a habitat with physical disturbance and surface destabilisation resulting in lower biomass of cyanobacteria. A further explanation to the development of a cyanobacterial mat in sandy sediments could be due to the high abundance of foraminifers since these have been found to graze on diatoms (Austin, et al., 2005), and thereby reduce the biomass of diatoms in favour to cyanobacteria.
The results from this investigation indicate
that green algal mats should have difficulties to grow in shallow bays. On the contrary, the distribution of green algal mats has actually increased (Pihl, et al., 1995) on the Swedish west coast. However, the occurrence of algae varies between areas and over time. Inorganic nutrients used during initiation of growth of the mats seem to originate both from the sediment and the water column. Grazing by Nereis diversicolor and

INITIATION OF ALGAL MATS
- 14 -
competition from BMA may be the explanations, at least partly, to the shifting growth patterns of green algal mats at some locations, or during some periods of time. If algal mats develop late in the spring or early summer, Nereis larvae might already have settled and are capable of ingesting germinating propagules, but if mats start to grow early in the spring (growth can start in April; Pihl et al. 1999), the Nereis larvae might settle too late to be able to effectively control the fast-growing green algal mat. Further, some winters are cold enough to bottom-freeze the shallow areas, and thereby the infauna, including polychaetes, is severely reduced in densities in spring (Reiss, et al., 2006). Propagules may also attach to moving objects e.g. the small gastropod Hydrobia spp. (Schories, et al., 2000), or to stones or shells etc. (Pihl, et al., 1999), where they are out of reach of grazing by N. diversicolor, and finally can start to grow and develop into a fully grown mat.
This study shows that filamentous green algae can grow with low concentrations of nutrients in the water column if no grazers are present. Even though these algae are fast in consuming nutrients, (Wallentinus, 1984) and allocate it directly into growth rather than to storage reserves (McGlathery, 1992), N. diversicolor and benthic cyanobacterial mats seem to have great impact on growth of green algal mats.
Conclusions The results from this study points to at
least two aspects which have to be taken into consideration when studying the initiation and growth of green algal mats; grazing by Nereis diversicolor and competition for nutrients with benthic microalgae. This study concludes that macroalgae (e.g. Ulva spp.) grew well without the underlying sediment. The only supply of nutrients came from the water column. Cyanobacteria seemed to be excellent competitors for inorganic nutrients, and thereby were able to prevented macroalgal growth. The grazing
of N. diversicolor also seemed to prevent initiation of macroalgal growth.
Future outlook Although competition for nutrients with
benthic micoalgae and grazing by Nereis diversicolor have been reported from other studies, the role of grazing by N. diversicolor during the actual initiation and growth of green algal mats need more studying. There has not yet been discussed whether fully grown algal mats are affected, or what interactions there are with inorganic nutrients and grazing by N. diversicolor. Questions arise whether this is a common phenomenon, and if N. diversicolor can control the growth of these green algal mats in situ.
Acknowledgements. The study was financially supported by grants from FORMAS, The foundation in memory of Birgit and Birger Wåhlström, The foundation of Captain Stenholm, The foundation of Wilhelm and Martina Lundgren and The royal Swedish Academy of Sciences. Thanks to professors Leif Pihl and Kristina Sundbäck, Göteborg University, for valuable comments on this manuscript.
LITERATURE CITED Aller, R.C., Blair, N.E., 2004. Early
diagenetic remineralization of sedimentary organic C in the Gulf of Papua deltaic complex (Papua New Guinea): Net loss of terrestrial C and diagenetic fractionation of C isotopes. Geochim. Cosmochim. 68, 1815-1825.
Astill, H., Lavery, P.S., 2001. The dynamics of unattached benthic macroalgal accumulations in the Swan-Canning Estuary. Hydrol. Proc. 15, 2387-2399.
Austin, H.A., Austin, W.E., Paterson, D.M., 2005. Extracellular cracking and content removal of the benthic diatom Pleurosigma angulatum (Quekett) by the benthic foraminifera Haynesina

INITIATION OF ALGAL MATS
- 15 -
germanica (Ehrenberg). Mar. Micropal. 57, 68-73.
Bartoli, M., Nizzoli, D., Viaroli, P., 2003. Microphytobenthos activity and fluxes at the sediment-water interface: interactions and spatial variability. Aqua. Bot. 37, 341-349.
Bonsdorff, E., 1992. Drifting Algae and Zoobenthos - Effects on Settling and Community Structure. Neth. J. Sea Res. 30, 57-62.
Bäck, S., Lehvo, A., Blomster, J., 2000. Mass occurrence of unattached Enteromorpha intestinalis on the Finnish Baltic Sea coast. Ann. Bot. Fenn. 37, 155-161.
Cummins, S.P., Roberts, D.E., Zimmerman, K.D., 2004. Effects of the green macroalga Enteromorpha intestinalis on macrobenthic and seagrass assemblages in a shallow coastal estuary. Mar. Ecol. Prog. Ser. 266, 77-87.
Dalsgaard, T., Nielsen, L.P., Brotas, V., Viaroli, P., Underwood, G.J.C., Nedwell, D.B., Sundbäck, K., Rysgaard, S., Miles, A., Bartoli, M., Dong, L., Thornton, D.C.O., Ottosen, L.D.M., Castaldelli, G., Risgaard-Petersen, N., 2000. Protocol handbook for NICE - Nitrogen Cycling in Estuaries: a project under the EU research program: Marine Science and Technology (MAST III). National Environmental Research Institute, Denmark, 62 pp.
Dubnyak, S., Timchenko, V., 2000. Ecological role of hydrodynamic processes in the Dnieper reservoirs. Ecol. Eng. 16, 181-188.
Fong, P., Donohoe, R.M., Zedler, J.B., 1993. Competition with macroalgae and benthic cyanobacterial mats limits phytoplankton abundance in experimental microcosm. Mar. Ecol. Prog. Ser. 100, 97-102.
Glud, R.N., Rysgaard, S., Kuhl, M., 2002. A laboratory study on O-2 dynamics and photosynthesis in ice algal communities: quantification by microsensors,O-2 exchange rates, C-14 incubations and a PAM fluorometer. Aquat. Microb. Ecol. 27, 301-311.
Hansen, K., Kristensen, E., 1997. Impact of macrofaunal recolonization on benthic metabolism and nutrient fluxes in a shallow marine sediment previously overgrown with macroalgal mats. Estuar. Coast. Shelf. Sci. 45, 613-628.
Hodgkin, E.P., Birch, P.B., 1986. No simple solutions - proposing radical management options for an eutrophic estuary. Mar. Poll. Bull. 17, 399-404.
Jensen, P., 1996. Burrows of marine nematodes as centres for microbial growth. Nematol. 42, 320-329.
Karlson, K., Hulth, S., Ringdahl, K., Rosenberg, R., 2005. Experimental recolonisation of Baltic Sea reduced sediments: survival of benthic macrofauna and effects on nutrient cycling. Mar. Ecol. Prog. Ser. 294, 35-49.
Lorenzen, C.J., 1967. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations. Limnol. Oceanogr. 12, 343-346.
Mcglathery, K.J., 1992. Physiological Controls on the Distribution of the Macroalga Spyridea hypnoides - Patterns Along a Eutrophication Gradient in Bermuda. Mar. Ecol. Prog. Ser. 87, 173-182.
Mcglathery, K.J., Krause-Jensenb, D., Rysgaardb, S., Christensen, P.B., 1997. Patterns of ammonium uptake within dense mats of the filamentous macroalga Chaetomorpha linum. Aquat. Bot. 59, 99-115.
Meziane, T., Retiere, C., 2002. Growth of Nereis diversicolor (L.) juveniles fed with detritus of halophytes. Ocean. Acta 25, 119-124.
Michaud, E., Desrosiers, G., Mermillod-Blondin, F., Sundby, B., Stora, G., 2006. The functional group approach to bioturbation: II. The effects of the Macoma balthica community on fluxes of nutrients and dissolved organic carbon across the sediment-water interface. J. Exp. Mar. Biol. Ecol. 337, 178-189.
Nelson, T.A., Nelson, A.V., Tjoelker, M., 2003. Seasonal and spatial patterns of "Green tides" (Ulvoid algal blooms) and

INITIATION OF ALGAL MATS
- 16 -
related water quality parameters in the coastal waters of Washington state, USA. Bot. Mar. 46, 263-275.
Nordström, M., Bonsdorff, E., Salovius, S., 2006. The impact of infauna (Nereis diversicolor and Saduria entomon) on the redistribution and biomass of macroalgae on marine soft bottoms. J. Exp. Mar. Biol. Ecol. 333, 58-70.
Norkko, A., Bonsdorff, E., 1996a. Rapid zoobenthic community responses to accumulations of drifting algae. Mar. Ecol. Prog. Ser. 131, 143-157.
Norkko, A., Bonsdorff, E., 1996b. Population responses of coastal zoobenthos to stress induced by drifting algal mats. Mar. Ecol. Prog. Ser. 140, 141-151.
Olivier, M., Desrosiers, G., Caron, A., Retiere, C., Caillou, A., 1995. Behavioural responses of the polychaetes Nereis diversicolor (O.F. Muller) and Nereis virens (Sars) to alimentary stimuli: use of particular organic matter, algae and halophytes. Can. J. Zoo. 73, 2307-2317.
Pihl, L., Rosenberg, R., 1982. Production, abundance and biomass of mobile epibenthic marine fauna in shallow waters, western Sweden. J. Exp. Mar. Biol. Ecol. 57, 273-301.
Pihl, L., Isaksson, I., Wennhage, H., Moksnes, P.-O., 1995. Recent increase of filamentous algae in shallow Swedish bays: effects on the community structure of epibenthic fauna and fish. Neth. J. Sea Res. 29 (3-4), 349-358.
Pihl, L., Magnusson, G., Isaksson, I., Wallentinus, I., 1996. Distribution and growth dynamics of ephemeral macroalgae in shallow bays on the Swedish west coast. J. Sea Res. 35 (1 - 3), 169-180.
Pihl, L., Svenson, A., Moksnes, P.-O., Wennhage, H., 1999. Distribution of green algal mats throughout shallow soft bottoms of the Swedish Skagerrak archipelago in relation to nutrient sources and wave exposure. J. Sea Res. 41, 281-294.
Raffaelli, D., 2000. Interactions between macro-algal mats and invertebrates in the Ythan estuary, Aberdeenshire, Scotland. Helgol. Mar. Res. 54, 71-79.
Reiss, H., Meybohm, K., Kroncke, I., 2006. Cold winter effects on benthic macrofauna communities in near- and offshore regions of the North Sea. Helgol. Mar. Res. 60, 224-238.
Schories, D., Anibal, J., Chapman, A.S., Herre, E., Isaksson, I., Lillebö, A.I., Pihl, L., Reise, K., Sprung, M., Thiel, M., 2000. Flagging greens: hydrobiid snails as substrata for the development of green algal mats (Enteromorpha spp) on tidal flats of North Atlantic coasts. Mar. Ecol. Prog. Ser. 199, 127-136.
Strickland, J.D., Parsons, T.R., 1972. A practical handbook of seawater analysis. Bull. Fish Res. Board Can. 167.
Sundbäck, K., Miles, A., 2002. Role of microphytobenthos and denitrification for nutrient turnover in embayments with floating macroalgal mats: a spring situation. Aquat. Microb. Ecol. 30, 91-101.
Sundbäck, K., Miles, A., Goransson, E., 2000. Nitrogen fluxes, denitrification and the role of microphytobenthos in microtidal shallow-water sediments: an annual study. Mar. Ecol. Prog. Ser. 200, 59-76.
Sundbäck, K., Jönsson, B., Nilsson, P., Lindström, I., 1990. Impact of accumulating drifting macroalgae on a shallow-water sediment system: an experimental study. Mar. Ecol. Prog. Ser. 58, 261-274.
Sundbäck, K., Miles, A., Hulth, S., Pihl, L., Engström, P., Selander, E., Svenson, A., 2003. Importance of benthic nutrient regeneration during initiation of macroalgal blooms in shallow bays. Mar. Ecol. Prog. Ser. 246, 115-126.
Söderkvist, J., 1997. Water exchange in a shallow bay. Master thesis, Gothenborg University, Sweden.
Thybo-Christensen, M., Rasmussen, M.B., Blackburn, T.H., 1993. Nutrient fluxes and growth of Cladophora sericea in a

INITIATION OF ALGAL MATS
- 17 -
shallow Danish bay. Mar Ecol-Prog Ser 100, 273-281.
Thybo-Christesen, M., Rasmussen, M.B., Blackburn, T.H., 1993. Nutrient fluxes and growth of Cladophora sericea in a shallow Danish bay. Mar. Ecol. Prog. Ser. 100, 273-281.
Vahteri, P., Makinen, A., Salovius, S., Vuorinen, I., 2000. Are drifting algal mats conquering the bottom of the Archipelago Sea, SW Finland? Ambio 29, 338-343.
Valiela, I., Mcclelland, J., Hauxwell, J., Behr, P.J., Hersh, D., Foreman, K., 1997. Macroalgal blooms in shallow estuaries: controls and ecophysiological and ecosystem consequences. Limnol. Oceanogr 42, 1105-1118.
Valle-Levinson, A., Delgado, J.A., Atkinson, L.P., 2001. Reversing water exchange patterns at the entrance to a semiarid coastal lagoon. Estuar. Coast. Shelf. Sci. 53, 825-838.
Wallentinus, I., 1984. Comparision of nutrient uptake rates for Baltic macroalgae with different thallus morphologies. Mar. Biol. 80, 215-225.
Vedel, A., Riisgard, H.U., 1993. Filter-feeding in the polychaete Nereis diversicolor: growth and bioenergetics. Mar. Ecol. Prog. Ser. 100, 145-152.

Grazing effects by Nereis diversicolor on development and growth of green algal mats.
Anna Engelsen and Leif Pihl
Department of Marine Ecology, Kristineberg Marine Research Station, Göteborg University,
SE-45034 Fiskebäckskil, Sweden
Abstract
Nereis diversicolor is generally considered to be a predator and deposit feeder, but have also been found to graze on benthic algae in shallow coastal areas. In this study we investigated the grazing effects on the development and growth of green algae, Ulva spp. Algal growth was studied in an experiment including two levels of sediment thickness; 100 mm sediment including macrofauna and 5 mm sediment without macrofauna, and three treatments of varying algal biomass; sediment with propagules, sediment with low algal biomass (120 g dry weight (dwt) m-2) and sediment with high algal biomass (240 g dwt m-2). In the 100 mm sediment, with a natural population of macrofauna, N. diversicolor was the dominating (60 % of total biomass) species. After three weeks of experimentation the result showed that N. diversicolor was able to prevent initial algal growth, affect growth capacity and also partly reduce full - grown algal mats. The weight of N. diversicolor was significantly higher for polychaetes in treatments with algae added compared to non - algal treatments. There were also indications that a rich nutrient supply per algae biomass counteracted the grazing capacity of N. diversicolor. Keywords: Nereis diversicolor, Ulva spp., grazing, nutrients, eutrophication
1. Introduction Diverse seaweed communities have in many coastal areas become replaced by fast growing annual macroalgae that establish dense mats altering the local ecology of benthic systems (Worm et al., 1999). Human influences such as climate change, eutrophication and food web alterations due to over fishing support the development of macro algal mats in shallow coastal areas and thereby constitutes a threat to these communities (Lotze and Worm, 2002). Annual macroalgae are found worldwide (Valiela et al., 1997) and they are dependent on high levels of nutrients (Sundbäck et al., 2003), *) Corresponding author: [email protected]
since their storage capacity is low (McGlathery, 1992)an appropriate substrate to attach to (Schories et al., 2000; Stål, 2001) and locations with low wind and wave exposure (Pihl et al., 1996; Pihl et al., 1999). They grow fast with rich nutrient supply since their morphology give them a large surface to volume ratio (Wallentinus, 1984). Although algal mats occur frequently (Pihl et al., 1995), and are considered as a threat to shallow bay communities, still little is understood about their initiation and growth. Further, Worm et al. (1999) and Lotze et al. (1999) found that grazing by different herbivores on green algae (e.g. Ulva spp.) may be of significance for the algal growth. Lotze and Worm (2000) showed that a high density of epibentic herbivores, such as e.g. Idothea

Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats 2
spp., Gammarus spp. and Littorina spp could effectively control macroalgal blooms in areas with eutrophication. Also infauna species could potentially be grazing, and it is found that N. diversicolor could feed on green algae (Hughes, 1999; Raffaelli, 2000; Nordström et al., 2006). N. diversicolor is generally known as a predator or deposit feeder (Kristensen and Mikkelsen, 2003), but can also act as a filter feeder (Vedel and Riisgard, 1993; Kristensen and Mikkelsen, 2003). This opens the question weather N. diversicolor has an impact on the development of algal mats, or even is able to reduce full grown mats. There are few studies so far, that concern the impact of N. diversicolor on the growth of macro algal mats on shallow shores, although N. diversicolor seems to constitute a potential grazer on Ulva spp. in such areas (Hughes, 1999; Raffaelli, 2000; Nordström et al., 2006) Since macro algal mats seem to vary in biomass from year to year and Nereis diversicolor has been found to graze on filamentous algae, the aim of this study was to investigate the grazing capacity on developing macro algal mats (e. g. Ulva spp). The hypothesis was that N. diversicolor is able to prevent initial algal growth, affect growth capacity and also reduce full-grown algal mats. Natural undisturbed sediments of two different thicknesses (100 mm and 5 mm) were exposed to three different treatments: natural density of propagules, addition of low biomass of green algae and addition of high biomass of green algae. Experiments were conducted in small mesocosms with a flow through system during a period of three weeks. 2. Materials and Methods 2.1 Experimental design The investigation was carried out from 16 June to 7 July 2005 at Kristineberg Marine Research Station on the west coast of Sweden (58º 15′ N, 11º 30′ E). The area is micro-tidal with average amplitude of 20
cm. During summer shallow (0.5 m) coastal water has a salinity in situ between 20 and 25, and the water temperature range between 18 and 28°C (Pihl and Rosenberg, 1982). Experiments were carried out in an out door green house laboratory. Sixteen undisturbed deep sediment cores (DS) (internal diameter (i.d.) 165 mm and height 100 mm) were sampled 16 of June in a bay having silty sediment (grain size fraction < 63 µm was 35 %), and with an organic matter content of 14%. Sampled DS cores contained natural densities of benthic macrofauna. In addition, 16 sediment cores were sampled from which the surface sediment (SS; i.d. 165 mm and height 5 mm) was extracted by slicing the upper 5 mm of the sediment, and thereby excluding the major part of the macrofauna from the samples. Further, the extracted surface sediment was examined by eye to remove potentially remaining macrofauna by hand. All sampled cores were transported to the laboratory and randomly allocated in basins with flow - through sea water, taken from 6 m depth. The 5 mm surface sediments were spread out on plates which were placed at the same height as the surfaces of the DS cores, so that all sediment cores had the same light conditions. Each core was supplied with an individual flow of water so that in every core there was a water column of 5 cm overlying the sediment. The water column in each core overflowed the edge of the core and drained into in the basin. The flow-through system also functioned as a cooling system so that ambient temperature of the sediment was maintained during the experiment. The collections of DS and SS samples were both separated into four groups; one initial (control) group with four replicates (DS init; SS init) and three treatments each with four replicates: natural distribution of propagules (DS prop; SS prop), added low biomass (corresponding to 120 g dwt m-2) of algae (DS low; SS low), and added high biomass (corresponding to 240 g dwt m-2) of algae (DS high; SS high) (Se Table 1 for design). The added algae consisted of Ulva spp. with dominance of

3 Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats
U. compressa. To all treatments nutrient enrichment were added every 5th day to give optimal growth conditions for the algae. Non - coated fertilizer pellets (Hydro Agri OptiCrop NPK 21 – 3 – 10), normally used in agriculture, were placed in mesh bags at the water inlet of each core. The amount of fertilizer pellets used was chosen according to Worm et al. (2000). Initial (control) sampling was performed 24 hours after experimental set-up. Thereafter samples were taken on two occasions, 10 days apart. The initial (17 June) and last (7 July) sampling included destructive samplings for macro algal biomass, and benthic fauna abundance and biomass. Water samples for measurements of oxygen and nutrient fluxes were taken on all three occasions. Three cores out of four from each treatment were randomly chosen at each sampling. 2.2 Macroalgal growth The degree of macro algal cover in the cores was assessed every week by visual observations and photo documentation. During the last (7 July) destructive sampling macroalgae were removed from all cores and biomass (dwt.) was determined for calculations of algal growth over the 20 days of investigation. 2.3 Benthic oxygen- and nutrient fluxes
To calculate oxygen and inorganic nutrient fluxes, concentrations of O2, NH4
+, NO3, PO4
3- and Si(OH)4) were measured in samples taken from the water column of each core, both during light (12.00 a. m.) and dark conditions (12.00 p.m.). During incubations, cores were closed by acrylic lids and the individual water flow at each core was turned off. Oxygen concentrations were not allowed to change > 20 % during incubation, to avoid oxygen-induced changes in nutrient fluxes (Dalsgaard et al., 2000).The incubation period lasted for approximately one hour during the light period and two hours during the dark period. The same cores were used for both oxygen and nutrient fluxes. Glass syringes (20 - ml) were used to take oxygen samples, and disposable plastic syringes (20 ml) were used to take nutrient samples. Samples for time zero were taken from open cylinders and at the end of the incubation through a 5 mm opening in the lid. Samples for nutrients were filtered on-line (0.45 µm Sartorius syringe filters) and frozen immediately at - 80 °C until analysis. A multi-channel auto-analyzer (TRAACS 800, Bran & Lubbe) was used for colorimetric analysis of inorganic nutrients (Strickland and Parsons, 1972). Daily nutrient fluxes were calculated by multiplying the hourly light fluxes by the number of daylight hours (day),
Table 1. Overview over names, treatments, algal biomass and nutrients added and number of replicates. DS = sediment 100 mm and SS = sediment 5 mm. DS and SS low correspond to an algal biomass of 120 g dry weight (dwt), DS and SS high correspond to an algal biomass of 240 g dwt, DS and SS prop are sediments with only propagules. Treatment Sediment thickness Algal biomass
added Nutrients added
Replicates
DS Init 100 mm natural sediment - - 4 DS prop 100 mm natural sediment propagules X 4 DS low 100 mm natural sediment (120 g dwt) X 4 DS high 100 mm natural sediment (240 g dwt) X 4 SS prop 5 mm surface sediment propagules X 4 SS low 5 mm surface sediment (120 g dwt) X 4 SS high 5 mm surface sediment (240 g dwt) X 4

Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats 4
and the hourly night fluxes by the number of dark hours, and then adding these two numbers. The method and process is described in Dalsgaard et al. (2000). Oxygen concentrations were measured immediately after sampling using Winkler titration (Strickland and Parsons, 1972). Daily net primary production (NPP) was calculated by multiplying flux rates per incubation time by a light factor calculated as a ratio between total daily irradiance and irradiance during the incubation period (light data from the Swedish Meteorological and Hydrological Institute). 2.4 Benthic macrofauna During the destructive sampling the sediment of each core was wet-sieved and the benthic macrofauna was retrieved by a 1 mm mesh sized sieve. The fauna was identified to lowest taxonomic level possible, counted for abundance and the wet weight was used for biomass estimation. 2.5 Statistical analyses A one - factor ANOVA tested differences in Nereis diversicolor abundance and biomass between DS treatments (propagules, low algal biomass and high algal biomass). A two-factor ANOVA tested differences in oxygen and nutrient fluxes and algal growth between treatments, with sediment thickness (DS and SS cores) and algal biomass (propagules, low algal biomass and high algal biomass) as fixed factors. Homogeneity of variance was tested by Levene’s test of equality of error variances. When there was a significant interaction between factors, comparisons of the means were made using Tukey test. Differences were accepted as significant at p < 0.05. 3. Results 3.1 Macroalgal growth No growth of macroalgae could be detected in the DS prop treatment during the
experimental period (Fig. 1). However, on the SS prop cores propagules developed during the 20 days of experimentation into an algal mat corresponding to a mean biomass of 105 g dwt. m-2. In treatments with low biomass added, there was a trend of increased algal growth in both DS and SS cores (not statistically confirmed) (Table 2). In DS cores where high biomass of algae was added the algal biomass decreased significantly (p = 0.001) compared to start biomass, and there was also a reduction in macro algal biomass in SS high cores, although not statistically confirmed (Fig. 1; Table 2). Reduction in biomass was double as high in DS high cores as in the SS high cores, and the difference between treatments was bordering significance (p=0.07). A significant interaction (p = 0.035) was found between algal biomass (prop, low and high) and sediment depth (DS and SS). Tukey test showed significant differences in algal growth between DS high and DS prop (p = 0.01) and between DS low and DS prop (p = 0.002). DS prop also differed significantly from SS high (p = 0.001) and SS low (p = 0.001). No significant differences were found within SS treatments, and therefore the mean algal biomass for all SS cores could be considered as a reference measure of algae unaffected by grazing. This measure could therefore be used to compare the effect of grazing by Nereis diversicolor in the three DS treatments (Table 2). 3.2 Fluxes of oxygen and nutrients The highest net primary production (NPP) among DS cores was found in DS low (150 and 250 mmol O2 m-2 d-1), while the highest NPP in SS cores was found in SS low (230 and 270 mmol O2 m-2 d-1) and SS prop (230 and 300 mmol O2 m-2 d-1) (Fig. 2). NPP differed significantly between sediment types (p = 0.01) and between algal biomass (p = 0.003), and a significant (p = 0.001) interaction was shown (Table 2).

5 Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats
Table 2. Summary of analyses of variance (ANOVA) where the one - factor ANOVA used algal biomass (prop, low and high) as fixed factor and the two - factor ANOVA used algal biomass (prop, low and high) and sediment thickness (DS and SS) as fixed factors. Differences were accepted as significant at p < 0.05. Significance is highlighted in bold text in the table.
df MS F p Nereis diversicolor (abundance) Algal biomass 2 15184 1.45 0.254 Nereis diversicolor (biomass) Algal biomass 2 4473 3.54 0.045 Algal growth Algal biomass 2 44315 14.08 0.001 Sediment depth 1 6093 1.93 0.181 Sediment depth*algal biomass. 2 5819 3.70 0.035 Net primary production (O2) Algal biomass 2 56772 6.90 0.003 Sediment depth 1 102381 12.44 0.001 Sediment depth*algal biomass. 2 70385 8.55 0.001 NH4 Algal biomass 2 4033139 3.23 0.051 Sediment depth 1 3543066 2.84 0.101 Sediment depth*algal biomass. 2 3728166 2.99 0.063 NO3 Algal biomass 2 39361 0.11 0.892 Sediment depth 1 1781052 5.17 0.029 Sediment depth*algal biomass. 2 2658 0.01 0.992 PO4 Algal biomass 2 453770 0.69 0.507 Sediment depth 1 8165585 12.47 0.001 Sediment depth*algal biomass. 2 204925 0.31 0.733 Si(OH)4 Algal biomass 2 349358 0.65 0.526 Sediment depth 1 188470 0.35 0.556 Sediment depth*algal biomass. 2 501690 0.94 0.400

Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats 6
0
50
100
150
200
250
300
DS low DS high DS prop SS low SS high SS prop
Mac
roal
gal b
iom
ass
g dw
t m-2
17 Jun07-jul
Fig. 1 Macroalgal biomass, dry weight, (dwt) at start (17 June) and stop (7 July) of the
experiment. DS = 100 mm sediment and SS = 5 mm sediment. DS and SS low correspond to an addition of 120 g dry weight (dwt) of macroalgae, DS and SS high correspond to an addition of 240 g (dwt) of algae and DS and SS prop correspond to sediments with propagules only. At start, replicates had identical biomass. At stop mean and SE are given (n = 4).
Tukey test showed that NPP was significantly lower for DS prop compared to DS low (p = 0.004), SS prop (p = 0.003) and SS low (p = 0.003) (Fig. 2). Also DS high differed significantly (p = 0.03) from
SS prop. Generally, there was a daily net release of NH4, PO4 and Si(OH)4 from the sediments, whereas for NO3 both an uptake and release was observed (Fig. 3).
0
50
100
150
200
250
300
350
400
DS low DS high DS prop SS low SS high SS prop
NPP
mm
ol O
2 m-2
d-1
17-jun 27-jun 07-jul
Fig. 2 Daily (24 h) net primary production (NPP) during the experiment period, where DS =
100 mm sediment and SS = 5 mm sediment. DS and SS low correspond to an addition of 120 g dry weight (dwt) of macroalgae, DS and SS high correspond to an addition of 240 g (dwt) of algae and DS and SS prop correspond to sediments with propagules only. Mean and SE are given (n = 4).

7 Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats
Highest fluxes for PO4 were shown the in the end of the investigation, although not statistical confirmed. Ammonium fluxes seemed to be stable throughout the investigation, except for SS prop, which had high values the second sampling (27 June). Nitrate showed a pattern of an uptake in DS cores and release in SS cores the first sampling (17 June). During the second
sampling (27 June) there was mainly a release of NO3, which turned into an uptake the third (7 July) sampling except for DS low and SS high (Fig.3). Nitrate and phosphate both showed significant (NO3; p = 0.029, PO4; p = 0.001) differences in flux rate between sediment thickness (DS and SS) (Fig. 3, Table 2).
-2000
-1000
0
1000
2000
3000
4000
5000
umol
PO
4 m
-2 d
-1
-1000-800-600-400-200
0200400600800
1000
DS low DS high DS prop SS low SS high SS prop
umol
NO
3 m-2
d-1
-1000
-500
0
500
1000
1500
2000
2500
DS low DS high DS prop SS low SS high SS prop
umol
Si(O
H)4
m-2
d-1
-2000
-1000
0
1000
2000
3000
4000
5000
6000
umol
NH
4 m
-2 d
-1
17-jun 27-jun 07-jul
Fig. 3 Daily (24 h) fluxes of inorganic nitrogen (NH4 and NO3), phosphorous and silicon.
DS = 100 mm sediment and SS = 5 mm sediment. DS and SS low correspond to an addition of 120 g dry weight (dwt) of macroalgae, DS and SS high correspond to an addition of 240 g (dwt) of algae and DS and SS prop correspond to sediments with propagules only. Mean and SE are given (n = 4).
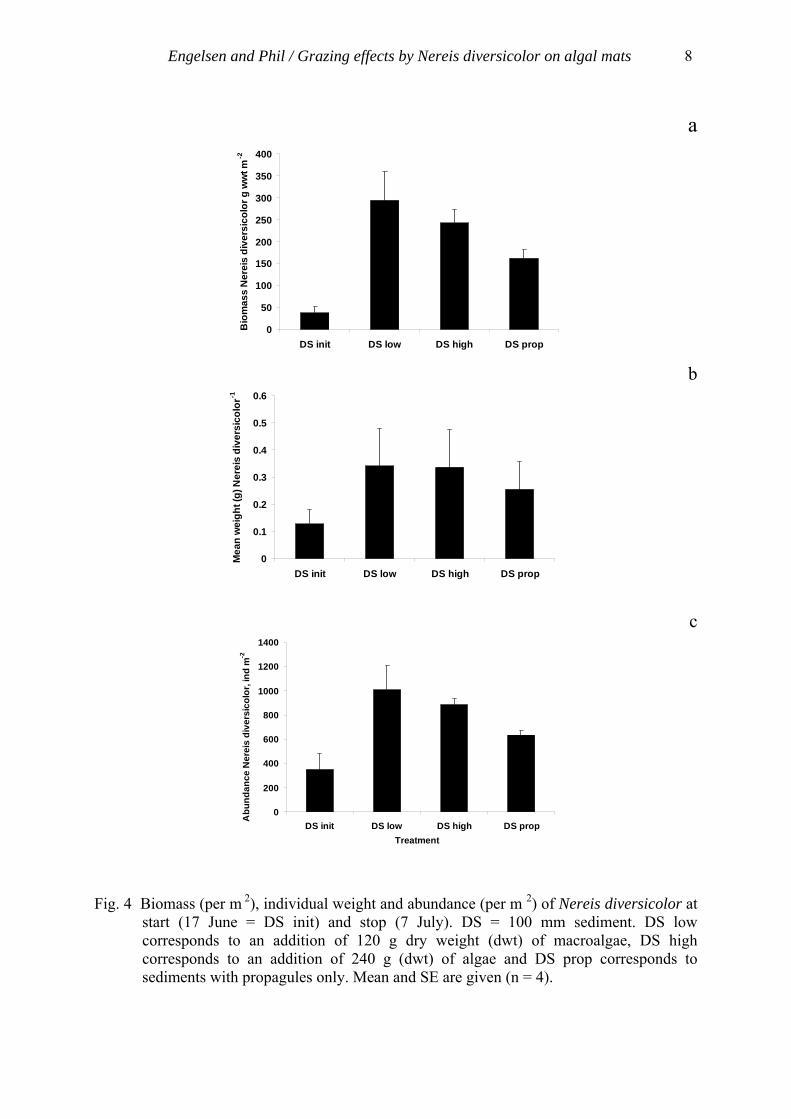
Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats 8
a
0
50
100
150
200
250
300
350
400
DS init DS low DS high DS prop
Bio
mas
s N
erei
s di
vers
icol
or g
ww
t m-2
b
0
0.1
0.2
0.3
0.4
0.5
0.6
DS init DS low DS high DS prop
Mea
n w
eigh
t (g)
Ner
eis
dive
rsic
olor
-1
c
0
200
400
600
800
1000
1200
1400
DS init DS low DS high DS propTreatment
Abu
ndan
ce N
erei
s di
vers
icol
or, i
nd m
-2
Fig. 4 Biomass (per m 2), individual weight and abundance (per m 2) of Nereis diversicolor at
start (17 June = DS init) and stop (7 July). DS = 100 mm sediment. DS low corresponds to an addition of 120 g dry weight (dwt) of macroalgae, DS high corresponds to an addition of 240 g (dwt) of algae and DS prop corresponds to sediments with propagules only. Mean and SE are given (n = 4).

9 Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats
3.3 Macrofauna - Nereis diversicolor The benthic macrofauna in the DS cores consisted mainly of Macoma baltica, Mya arenaria, Nereis virens, Nereis diversicolor, Pygospio elegans and chironomides, where N. diversicolor constituted up to 60 % of total biomass in replicate samples. The biomass of N. diversicolor increased significantly (p = 0.0006) in all treatments from start to the last sampling, and it was significantly (p = 0.045) higher in treatments where algae were added compared to sediment with propagules only (Fig. 4a). There was also a trend towards an increase in individual weight of N. diversicolor in treatments where algae were added (DS low and DS high), compared to the bare sediment treatment (DS prop) although not statistical confirmed (Fig. 4b). However, no significant differences were found in abundance of N. diversicolor between treatments (Fig. 4c). From visual observations and photo documentation N. diversicolor was seen grazing on propagules, algal filaments and full grown algae. Markings of grazing could be seen on the edges of the cores (DS prop, DS low and DS high) where filaments were growing, and N. diversicolor were seen grazing on the sediment surface, and in the algal mats. 4. Discussion 4.1 Impact of Nereis diversicolor on macroalgal growth The results from this study show that Nereis diversicolor is able to prevent initial algal growth, affect growth capacity and also partly reduce full-grown algal mats. The major difference between DS and SS cores was the presence of macrofauna, and in the DS sediments N. diversicolor was the dominating macrofaunal species. Propagules did not develop into an algal mat in presence of N. diversicolor, and there was a strong trend bordering
significance (p=0.07) of a higher reduction in algal biomass in DS compared to SS cores. There are few studies so far, concerning the impact of N. diversicolor on the growth of macro algal, although N. diversicolor seems to constitute a potential grazer on Ulva spp. in such areas (Hughes, 1999; Raffaelli, 2000; Nordström et al., 2006). Our study suggests that N. diversicolor could control the growth of algal mats in shallow habitats and also show that the growth of N. diversicolor increases for those polychaetes that had been grazing on green algae. The fact that worms that had been feeding on the macroalgae seems to grew faster than those feeding in cores with only propagules, indicates that these macroalgae either are more nutritious to the worms or are more easy to find and feed on, resulting in higher consumptions by N. diversicolor. Also the total energy cost for the worms might be lower when the polychaetes graze on green algae instead of being deposit feeders or predators. 4.2 The role of nutrients for growth of macroalgae The nutrient supply has a major impact on the growth of macroalgae. The supply of nutrients per biomass algae is essential for the rate of development of these macro algal mats since threadlike green algae have a fast uptake of inorganic nutrients (Wallentinus, 1984) but low storage ability (McGlathery, 1992). The prerequisite for fluxes of nutrients in this experiment may differ due to the absence of macrofaunal bioturbation in the SS treatments. However, the addition of nutrients and degradation of the macroalgae may reduce the differences between DS and SS treatments. In the DS and SS low treatments algal biomass increased, probably as a result of rich nutrient supply in relation to algal biomass. In contrast, the large algal mat in DS and SS high treatments probably suffered from N-limitation, since both NH4 and NO3 were low (Fig. 3). The observed reduction in algal biomass might be due to degrading

Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats 10
algae when the nutrient conditions for the algae are unfavourable. The increase in algal biomass was larger in SS low compared to DS low, and the decrease in SS high was less than that observed in the DS high treatment. These differences are probably a result of grazing by Nereis diversicolor, which counter balance algal growth. In DS low the algal growth rate could be faster than the grazing rate of worms and the algal biomass increase. In contrast, algae in the DS high treatment probably suffered from N-limitation in addition to grazing, resulting in a reduction in algal biomass (Fig. 1). These two scenarios indicate that a fully grown large mat of algae is more vulnerable to a limited nutrient supply than a smaller mat. If nutrient concentrations are low N. diversicolor could consume a considerable part of the large mat, whereas a smaller mat probably could grow faster than the grazing rate at the time, due to a higher nutrient supply per biomass algae. Another suggestion for the reduction of algal biomass in DS high treatments is that N. diversicolor rather chose macroalgae as a food source when there is a large supply (functional response), while they chose other food sources when the algal mat is smaller (DS low). 4.3 Net primary production The highest net primary production (NPP) was found for DS low, SS low and SS prop treatments, where high macroalgal biomass was found at the end of the experiment (7 Jul). The DS prop treatment, where no germination of macroalgae occurred, had a low NPP resulting only from benthic microalgae. There was a pronounced decrease in NPP during the second sampling for DS high and SS high, which might mirror the reduction in macroalgal biomass. The following increase in NPP (last sampling), could be the result of benthic microalgae that flourished due to excellent nutrient conditions for benthic microalgae, when the macroalgal mat degraded. This temporal variation in NPP
could also be a result of N-limitation during the second sampling since NH4 concentrations were low both in DS high and SS high treatments. During the last sampling, though, NH4 was produced which supplied the algal mat with nitrogen. The explanation for the low NPP during the second and last sampling in the DS prop treatments could be that algal propagules were lost due to grazing and the actual NPP came from the microphytobenthic community. The SS prop treatments showed a trend of increase in the NPP during the second and last sampling, probably due to the development of a new algal mat. 4.4 Establishment of a green algal mat In addition to inorganic nutrients, green algal propagules need a substrate to attach to for germination. The propagules attach to grains > 1 mm (Stål, 2001) in sandy sediments, but in silty sediments where the grain size is smaller (< 63 µm), propagules may attach to e.g. Hydrobia spp. (Schories et al., 2000). Since the gastropods move around, these propagules might have a better chance to grow and develop out of reach of Nereis diversicolor. However, in shallow tidal areas Hydrobia spp. take shelter in the sediment during low tide, and the algal filaments may be exposed to grazing. In very shallow areas (10 – 20 cm) the polychaetes are able to reach algal filaments more efficiently (Raffaelli, 2000) and there could be difficulties in establishing a macroalgal mat. The polychaetes can graze on the algal filaments during the time they still are attached to the bottom substrate, but the question arises how and if the polychaetes can reach the floating mat? A likely scenario would be that N. diversicolor starts to graze on propagules and continue to feed on algae until the mat starts to float out of its reach. The floating algal mats, however, will in shallow (20 – 30 cm) areas again suffer from grazing by polychaetes during low tide if the mat lies on the sediment.

11 Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats
4.5 Factors affecting the growth and grazing of green algal mats Salinity and temperature have a major role during initiation and growth of macro algal mats, partly due to indirect effects from grazing by Nereis diversicolor. Growth of Ulva spp. spores decreases when salinity is below 5, but is enhanced when it is over 20 (Sousa et al., 2007). This will result in less germination during rainy periods when fresh water mixes into the sea reducing the salinity. N. diversicolor, on the other hand, does not seem to be affected by natural changes in salinity (Kristensen, 1983). More run off from land will also result in higher concentrations of inorganic nutrients in the surface water, which will benefit the growth of germinated spores. Ulva spp. spores, however, seem to be sensitive to N toxicity and to P limitation (Sousa et al., 2007) which makes them vulnerable to large changes in nutrient concentrations. Inorganic nutrients can in addition to land run off also be released from the sediment by bioturbation (Aller, 1982; Hansen and Kristensen, 1997; Marinelli and Williams, 2003). Fluxes of PO4 and Si(OH)4 were highest during the last sampling (Fig. 3), which probably is a result of higher biomass of macrofauna since the mean individual weight of N. diversicolor increased between 90 and 130 % (Fig. 4b) during the study. A high bioturbation could then lead to higher concentrations of inorganic nutrients in the water which may result in a higher algal growth rate, but a high bioturbation also mean that e.g. N. diversicolor have a high biomass or abundance and are able to graze more efficiently. Temperature is of importance for both germination of propagules and the activity of N. diversicolor. Since polychaetes in cold water have a slower intestinal transit and thereby higher absorption efficiency (Neuhoff, 1979), foraging is lower in colder temperatures and stops at 8 °C (Lambert et al., 1992). This means that grazing does not start or is slow until temperature rises. Germination of Ulva spp. is also favored by
warm temperatures with an optimum between 17-22 °C (Sousa et al., 2007). This implies that grazing by N. diversicolor either starts earlier than, or simultaneous to germination of Ulva spp. Also the timing between recruitment of N. diversicolor larvae contra nutrient input seems to be of importance for development of algal mats. If polychaete larvae have started to grow and are capable of grazing before the propagules have nutrients enough to germinate, the development of an algal mat may be prevented. However, if the opposite situation occurs with an already established mat and high nutrient concentration when N. diversicolor juveniles start to graze, the impact on algal growth might be less severe. The ecosystem around green algal mats is complex and there are both abiotic and biotic factors influencing the potential growth of macroalgal mats in shallow marine areas. The abundance of Nereis diversicolor changes from year to year. At the size of 20 to 25 mm N. diversicolor larvae change from a non-feeding period to an active dispersal period in order to find food (Green, 1968). During this dispersal period they also compete for space (Möller and Rosenberg, 1983) and thereby are exposed to numbers of predators (Crangon crangon, the 0 – group of plaice, birds and cannibalism from adult N. diversicolor) (Pihl and Rosenberg, 1984) which can explain the larvae mortality of 97% (Möller, 1985). Also bottom frozen sediments during winters may reduce a whole population (Möller, 1985 and references therein). The existence of macroalgal mats depends on, not only abiotic factors such as nutrient release, temperature and salinity, but also on how the winter conditions have affected the entire macrofauna, since the population of N. diversicolor depends e.g. on the population dynamic of other macrofaunal species. Conclusion

Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats 12
Nereis diversicolor is able to reduce the growth of macro algal mats as well as prevent an algal mat of Ulva spp. to develop. Nereis diversicolor seems to grow faster and gain a larger biomass if they feed on Ulva spp. There seem to be an interaction between grazing pressure and algal growth rate, and a large macroalgal mat is more vulnerable to grazing with limited nutrient concentrations in the water. Acknowledgments Fundings was received from the Swedish Research Council for Environment, Agricultural Science and Special Planning (FORMAS), the Memorial Fund of Birgit and Birger Wåhlström and the Captain Stenholm Fund. We would also like to thank Linda Ottosson (Göteborg University) for all help during field and laboratory work and Karl Norling for comments on the design of the experiment. References Aller, R.C., 1982. The effects of
macrobenthos on chemical properties of marine sediment and overlying water. In: McCall, P.L., Tevesz, M.J.S. (eds), Animal - Sediment relations, Plenum, NY. 53-102.
Dalsgaard, T., Nielsen, L.P., Brotas, V., Viaroli, P., Underwood, G.J.C., Nedwtell, D.B., Sundbäck, K., Rysgaard, S., Miles, A., Bartoli, M., Dong, L., Thornton, D.C.O., Ottosen, L.D.M., Castaldelli, G., Risgaard – Petersen, N., 2000. Protocol handbook for NICE – Nitrogen Cycling in Estuaries: a project under the EU research program: Marine Science and Tecnology (MAST III). National Environmental Research Institute, Denmark, 62 pp.
Green, J., 1968. The biology of estuarine animals. London. Sidgwick & Jackson.
Hansen, K., Kristensen, E., 1997. Impact of macrofaunal recolonization on benthic metabolism and nutrient fluxes in a
shallow marine sediment previously overgrown with macroalgal mats. Estuar. Coast. Shelf. Sci. 45, 613-628.
Hughes, R.G., 1999. Saltmarsh erosion and management of saltmarsh restoration; the effects of infaunal invertebrates. Aqua. Cons. Mar. Freshwat. Ecosyst. 9, 83-95.
Kristensen, E., 1983. Ventilation and oxygen uptake by three species of Nereis (Annelida: Polychaeta). II. Effects of temperature and salinity changes. Mar. Ecol. Prog. Ser. 12, 299-306.
Kristensen, E., Mikkelsen, O.L., 2003. Impact of the burrow-dwelling polychaete Nereis diversicolor on the degradation of fresh and aged macroalgal detritus in a coastal marine sediment. Mar. Ecol. Prog. Ser. 265, 141-153.
Lambert, R., Desrosiers, G., Retiere, C., Miron, G., 1992. Exploration of Feeding Area by Nereis-Diversicolor (Polychaeta) (Muller,O.F.) - Preliminary Data. Cahie. Biol. Mar. 33 (1), 43 - 54.
Lotze, H.K., Schramm, W., Schories, D., Worm, B., 1999. Control of macroalgal blooms at early developmental stages: Pilayella littoralis versus Enteromorpha spp. Oecologia. 119, 46-54.
Lotze, H.K., Worm, B., 2000. Variable and complementary effects of herbivores on different life stages of bloom-forming macroalgae. Mar. Ecol. Prog. Ser. 200, 167-175.
Lotze, H.K., Worm, B., 2002. Complex interactions of climatic and ecological controls on macroalgal recruitment. Limnol. Oceanogr. 47 (6), 1734 - 1741.
Marinelli, R.L., Williams, T.J., 2003. Evidence for density-dependent effects of infauna on sediment biogeochemistry and benthic-pelagic coupling in nearshore systems. Estuar. Coast. Shelf. Sci. 57, 179-192.
Möller, P., Rosenberg, R., 1983. Recruitement, abundance andproduction of Mya arenaria and Cardium edule in marine shallow waters, western Sweden. Ophelia. 22, 33-55.

13 Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats
Möller, P., 1985. Production and abundance of juvenile Nereis diversicolor, and oogenic cycles of adults in shallow waters of western Sweden. J. Mar. Biol. Ass. U.K. 65, 603-616.
Neuhoff, H.G., 1979. Influence of temperature and salinity on food conversion and growth of different Nereis species (Polychaeta, Annelida). Mar. Ecol. Prog. Ser. 1, 225-262.
Nordström, M., Bonsdorff, E., Salovius, S., 2006. The impact of infauna (Nereis diversicolor and Saduria entomon) on the redistribution and biomass of macroalgae on marine soft bottoms. J. Exp. Mar. Biol. Ecol. 333, 58-70.
Pihl, L., Rosenberg, R., 1982. Production, abundance and biomass of mobile epibenthic marine fauna in shallow waters, western Sweden. J. Exp. Mar. Biol. Ecol. 57, 273-301.
Pihl, L., Rosenberg, R., 1984. Food selection and consumption of the shrimp Crangon crangon in some shallow marine areas in western Sweden. Mar. Ecol. Prog. Ser. 15, 159-168.
Pihl, L., Isaksson, I., Wennhage, H., Moksnes, P.-O., 1995. Recent increase of filamentous algae in shallow Swedish bays: effects on the community structure of epibenthic fauna and fish. Neth. J. Sea Res. 29 (3-4), 349-358.
Pihl, L., Magnusson, G., Isaksson, I., Wallentinus, I., 1996. Distribution and growth dynamics of ephemeral macroalgae in shallow bays on the Swedish west coast. J. Sea Res. 35 (1 - 3), 169-180.
Pihl, L., Svenson, A., Moksnes, P.-O., Wennhage, H., 1999. Distribution of green algal mats throughout shallow soft bottoms of the Swedish Skagerrak archipelago in relation to nutrient sources and wave exposure. J. Sea Res. 41, 281-294.
Raffaelli, D., 2000. Interactions between macro-algal mats and invertebrates in the Ythan estuary, Aberdeenshire, Scotland. Helgol. Mar. Res. 54, 71-79.
Schories, D., Anibal, J., Chapman, A.S., Herre, E., Isaksson, I., Lillebö, A.I., Pihl, L., Reise, K., Sprung, M., Thiel, M., 2000. Flagging greens: hydrobiid snails as substrata for the development of green algal mats (Enteromorpha spp) on tidal flats of North Atlantic coasts. Mar. Ecol. Prog. Ser. 199, 127-136.
Sousa, A.I., Martins, I., Lillebo, A.I., Flindt, M.R., Pardal, M.A., 2007. Influence of salinity, nutrients and light on the germination and growth of Enteromorpha sp spores. J. Exp. Mar. Biol. Ecol. 341, 142-150.
Strickland, J.D., Parsons, T.R., 1972. A practical handbook of seawater analysis. Bull. Fish Res. Board Can. 169, 1-311.
Stål, J., 2001. Importance of substrate structure for initiation and growth of filamentous algae (in Swedish). Master thesis, Göteborg University. 126, 1-18.
Sundbäck, K., Miles, A., Hulth, S., Pihl, L., Engström, P., Selander, E., Svenson, A., 2003. Importance of benthic nutrient regeneration during initiation of macroalgal blooms in shallow bays. Mar. Ecol. Prog. Ser. 246, 115-126.
Valiela, I., McClelland, J., hauxwell, J., Behr, P.J., Hersh, D., Foreman, K., 1997. Macroalgal blooms in shallow estuaries: controls and ecophysiological and ecosystem consequences. Limnol. Oceanogr. 42, 1105-1118.
Wallentinus, I., 1984. Comparision of nutrient uptake rates for Baltic macroalgae with different thallus morphologies. Mar. Biol. 80, 215-225.
Vedel, A., Riisgard, H.U., 1993. Filter-feeding in the polychaete Nereis diversicolor: growth and bioenergetics. Mar. Ecol. Prog. Ser. 100, 145-152.
Worm, B., Lotze, H.K., Boström, C., Engkvist, R., Labanauskas, V., Sommer, U., 1999. Marine diversity shift linked to interactions among grazers, nutrients and propagule banks. Mar. Ecol. Prog. Ser. 185, 309-314.
Worm, B., Reusch, T.B.H., Lotze, H.K., 2000. In situ nutrient enrichment:

Engelsen and Phil / Grazing effects by Nereis diversicolor on algal mats 14
Methods for marine benthic ecology. Int. Rew. Hydrobiol. 85, 359-375.

- 1 -
Links between oxygen deficiency, macroalgal growth and grazing by the polychaete Nereis
diversicolor
Anna Engelsen1, Kristina Sundbäck2 and Stefan Hulth3
1 Department of Marine Ecology, The Sven Lovén Centre for Marine Sciences - Kristineberg, University of Gothenburg, SE-450 34 Fiskebäckskil, Sweden
2 Department of Marine Ecology, University of Gothenburg, Box 461; SE-405 30 Göteborg, Sweden
3 Department of Chemistry, University of Gothenburg, SE-412 96 Göteborg, Sweden
ABSTRACT: The proliferation of green-algal mats in coastal embayments is generally considered a symptom of excessive nutrient supply. As these nuisance blooms are exclusively found in shallow areas, a strong coupling between their occurrence and benthic processes can be expected. Significant research efforts have been focused to the relation between green-algal mats and nutrients supplied from the pore-water to the water column, particularly under hypoxic and anoxic events in surface sediments. The functional role of benthic macrofauna for the initiation and sustenance of macroalgal mats is, however, less well known. The biogeochemical coupling between bottom-water oxygen deficiency and grazing of filamentous Ulva sp. by the polychaete Nereis diversicolor was in this study experimentally investigated in flow-through incubations (33 days) of undisturbed and manipulated sediment under close to natural light and temperature conditions. Three treatments were used: cores with the top 10 cm of natural sediment (NatSed; both Ulva propagules and macrofauna present), similar cores exposed to 5 days of anoxia (AnoxSed), and cores with only the top 5 mm natural sediment (SurfSed; Ulva propagules, no macrofauna). In addition to sediment–water oxygen and nutrient fluxes, macroalgal biomass, sediment chlorophyll a, and biomass of macrofauna were measured. A thick greenalgal mat developed in the SurfSed treatment. In AnoxSed, a mat started to develop shortly (4 days) after the anoxic period, but disappeared after additional 10 days. No mat was observed in the NatSed treatment. These results suggest that grazing by N. diversicolor is an important controlling factor for the initiation and sustenance of green-algal mats in shallow-water environments. Periods of benthic oxygen depletion seem to stimulate the growth of green-algal mats, not only by the increased availability of nutrients from the sediment (enhanced fluxes of ammonium and phosphate were observed during anoxia), but also by a reduced macrofaunal grazing pressure on the algae. KEY WORDS: Nereis diversicolor, grazing, sediment, macro-algal growth, anoxia
Introduction Proliferation of filamentous, or sheet-like, mats of green algae in shallow-water embayments is often considered a direct symptom of excessive nutrient supply to coastal waters (Valiela et al. 1997; Pihl et al. 1999). *) Corresponding author: [email protected]
Filamentous algae normally need a continuous supply of nutrients, since their storage capacity for nutrients is low due to their thallus morphology {Campbell, 2001 #535; Pihl, 1996 #207}. Consequently, to understand what factors that control the initiation and sustenance of these often nuisance blooms of filamentous green-algae, experimental research has mainly focused on the availability of nutrients, supplied either from external (e.g. land

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 2 -
run-off and adjacent water bodies) or internal (regeneration from local sediments) nutrient sources. On the micro-tidal west coast of Sweden, green-algal mats start to grow in spring, and during late summer, up to 50% of shallow embayments can be covered by algal-mats floating at the water surface (Pihl et al. 1999). A positive correlation has been observed between organic matter content in the sediment and recurrent green-algal mats (Pihl et al. 1999). Such immediate feedback suggests that sheltered bays may serve as self-perpetuating systems through benthic regeneration of pore-water nutrients (Trimmer et al. 2000; Sundbäck et al. 2003). This sediment pool of nutrients, particularly pronounced in organic-rich environments, is made available for macro-algal growth above all during low oxygen (hypoxic) and anoxic conditions, events frequently observed under thick algal mats (Thybo-Christesen and Blackburn 1993; Norkko and Bonsdorff 1996). In dense macro-algal mats, the combination of reduced light and enhanced organic matter mineralization within the mat induces low oxygen or anoxic conditions during both day and night conditions (Sundbäck and McGlathery 2005, and references therein). Under reducing conditions, pulses of above all phosphate and ammonium are released to the bottom water from the surface sediment (Blackburn and Henriksen 1983; Sundby et al. 1992). However, despite this obvious link between oxygen conditions in the bottom water, nutrient availability in the pore water and potential macroalgal growth, the development of macroalgal mats seems an irregular feature of shallow-water environments. Although otherwise favourable environmental conditions (e.g. high temperature, enhanced nutrient availability, low wind-speed and limited water movements), algal mats may not develop in the spring as predicted. Besides nutrients, there thus seem to be additional factors that control the initiation and
sustenance of green-algal mats in shallow-water embayments. As green-algal mats are exclusively observed in shallow areas, a strong coupling between the occurrence of algal mats and benthic processes mobilizing nutrients to the pore water and the overlying water is to be expected. The functional role of benthic macrofauna for this coupling between pore water nutrients and green-algal mats has largely been overlooked. Rates and pathways of organic matter mineralization in bioturbated sediments are closely related to activities by benthic macrofauna (i.e. bioturbation). Positive feedbacks between bioturbation and organic matter mineralization include, for example, particle manipulation, irrigation and removal of toxic metabolites, and transport of particles across redox boundaries (Aller 2001). Mineralization and reaction coupling may, however, also be constrained e.g. due to construction of macrofaunal tubes and linings of burrows with restricted gas and solute permeability. The polychaete Nereis diversicolor has been found to affect the biomass of above-sediment Ulva spp. both by grazing and by redistributing algal material deeper into the sediment (Hughes 1999; Raffaelli 2000; Nordström et al. 2006). Macrofaunal grazing and the removal of both propagules and established macroalgal mats (Raffaelli 2000; Giannotti and McGlathery 2001) constitute potentially important, but previously overlooked, controls for macroalgal growth in shallow-water embayments. The aim of our study was to experimentally investigate links between bottom-water oxygen deficiency, grazing by Nereis diversicolor and the growth of green-algal mats. We hypothesise that events of oxygen deficiency and anoxia in the bottom water stimulate the growth of green-algal mats due the combined effects from i) a reduced grazing pressure by N. diversicolor, and ii) a stimulated supply of

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 3 -
bioavailable nutrients from the sediment-pore water system. Under oxygenated conditions, on the other hand, grazing and organic matter redistribution by N. diversicolor control – and may even inhibit – the growth of algal mats. To study effects from low oxygen concentrations (including anoxia) and grazing by Nereis diversicolor on the growth of green-algal mats (Ulva sp.; formerly Enteromorpha), we performed sediment-water incubations of undisturbed and manipulated (including exposure to anoxic overlying water) surface sediment in a flow-through system under close to natural light and temperature conditions. Materials & Methods Experimental design and general overview A schematic overview of the experimental design and applied treatments is shown in Fig. 1. In brief, three types of treatments
were used: (i) sediment cores with the top 10 cm of natural sediment including indigenous macrofauna (NatSed); (ii) sediment cores as in (i) exposed to oxygen deficiency for 5 days during the experiment (AnoxSed); and (iii) sediment cores with only the upper-most 5 mm of natural sediment, i.e. no visible macrofauna (SurfSed). The experimental set-up also included cores with intact sediment from the sampling site (InitialSed). The sediment was incubated (total incubation period of 20 days) together with overlying water for benthic oxygen and nutrient fluxes. Additional samples were taken for green-algal and microalgal biomass, macrofaunal abundance and biomass. Sampling of sediment and benthic fluxes The experiment was performed during summer (June and July) 2005. Sediment
´
Day
ActivityFinal sampling for macrofauna
337 9 11 20 211810
Fieldcollec-tion
2
FluxesNatSed and SurfSed
Fluxes + destr. sampling
Fluxes + algalsampling
Start growthg. a. Hypox
Nereis activeagainHypox
Growth g. a. decreasedHypox
Start growthg. a. SurfSed
4
Anoxicperiod
FluxesAnoxSed
Observations
NatSed n = 4 AnoxSed n = 4 SurfSed n = 4InitialSed n = 4
Fig. 1 Treatments (a) and timing of sampling activities (above time scale) and main
observations (below time scale) during the experiment. NatSed (10-cm deep undisturbed sediment); AnoxSed (10-cm deep undisturbed sediment exposed to anoxia for 5 days; SurfSed (5-mm surface sediment, no macrofauna). n = 3+ SE; g.a.= green algae.

- 4 -
was collected in a shallow silty bay (Rågårdsvik, 58º 15′ N, 11º 30′ E) from a micro-tidal (tidal amplitude of ~20 cm) area on the west coast of Sweden. In summer, the shallow bays have a salinity between 20 and 25, and the water temperature varies normally between 22 and 25°C (Pihl and Rosenberg 1982). Twelve plexiglass cores (i.d. = 165 mm, sediment height ~ 10 cm) were carefully inserted into the sediment by hand. The cores with sediment and overlying water were transported to the Sven Lovén Centre for Marine Sciences - Kristineberg and incubated in a flow-through experimental system placed in a green-house for natural light conditions. Ambient light was therefore reduced by ~ 25 % and wavelengths within the UV-region were more or less completely excluded. For the treatment with only surface sediment (SurfSed), sediment was sampled using additional 10 - cm plexiglass cores. For this treatment, the top 5 mm layer of the surface sediment was removed and spread on the surface of a PVC plate (Fig. 1). The plate was positioned at the same level as the sediment surface of the other treatments, i.e. the volume of overlying water was approximately the same for all treatments. In the SurfSed treatment, concentrations of nutrients in the pore water were assumed to be close to those of the overlying water. Prior to the start of experiments, five specimens of Nereis diversicolor (~ 4 - 5 cm long) were added to the NatSed and the AnoxSed treatments to adjust for natural variations in the abundance of polychaetes. All treatments included the natural pool of macroalgal propagules and benthic microalgae. The cores were placed randomly in flumes in the greenhouse and each core was connected to an individual flow of surface water, directly supplied on-line from a nearby bay. The water was allowed to overflow the edge of the cores and drain into the flume. The temperature of the
cores was thereby maintained close to that of the surface water throughout the experiment (20 °C). A continuous water flow also ensured appropriate mixing of the overlying water. One day after the field sampling (day 1), initial samples were taken for oxygen and nutrients (NH4
+, NO3-, PO4
3- and Si(OH)4) in the overlying water, and for chlorophyll a, benthic microalgae, meiofauna, macrofauna and macroalgae from the sediment (n = 4 cores; InitialSed). Benthic flux incubations were initiated in that the water flow was stopped and the cores were sealed with transparent Plexiglas lids. Immediately after sealing the cores and after ~ one hour of closed incubation, samples for oxygen (20-ml glass syringes) and nutrients (50 ml polypropylene syringes) were removed from the overlying water through a port in the lid. Benthic flux rates were calculated from the concentration change in the overlying water with time of incubation (Hulth et al. 1994; Dalsgaard et al. 2000). During incubations, concentrations of oxygen measured in the overlying water did not deviate by more than 20 % from the initial values. Two days after the field sampling, oxygen was depleted in the overlying water of the AnoxSed treatment by sealing the cores with plexiglass lids and covering with black plastic. Anoxic conditions prevailed after ~ 2.5 hours. After five days of anoxic incubation (day 7), the lids were removed and the water supply was re-established. In addition to benthic fluxes measured on day 1, benthic fluxes of oxygen and nutrients (n = 3 cores per treatment and each sampling occasion) were determined on day 7 (AnoxSed), on day 9 (NatSed and SurfSed) and on day 20 (all treatments) using the same principles as described for the initial benthic flux measurement. The sediment cores were sectioned for macrofauna on day 33 (Fig. 1).

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 5 -
The same cores were used for light and dark fluxes of oxygen and nutrients. Oxygen flux under light conditions was used as a measure of net primary production (NPP), calculated by multiplying flux rates per incubation time by a light factor calculated as a ratio between total daily irradiance and irradiance during the incubation period (light data from the Swedish Meteorological and Hydrological Institute). Daily community respiration (CR) was estimated from the oxygen flux in the dark calculated over 24 hours. Daily rates of nutrient fluxes were calculated by multiplying the hourly rate under light conditions with the number of daylight hours, and the hourly rate in the dark times the number of dark hours, and adding these two values. Samples for dissolved oxygen were immediately precipitated and analysed by Winkler titration within 1 hour. Samples for nutrients were immediately frozen (-80ºC) and stored until analysis by a nutrient auto analyzer (TRAACS 2000, Bran-Luebbe) using standard colorimetric protocols (Strickland and Parsons 1972). Biological sampling Macroalgal dry weight (DW) was estimated from measured wet weight (day 20; Fig. 1) using a factor of 2.5 (Österling and Pihl 2001). The total coverage and progressive development of green-algal growth was followed by weekly estimate the macroalgal cover by eye Chlorophyll a content (Chl a) of the sediment was used as a rough estimate of the microphytobenthic biomass. After the flux measurements, the top 5 mm of the sediment was sampled with a cut-off 2 ml syringe (i.d. = 8.7 mm) and immediately frozen (-18 °C) until analysis. Six sub-samples were taken from each core, of
which two samples were pooled (i.e. there were three pseudo-replicates from each core). The samples were extracted in 4 ml 90 % acetone for 24 hours (+8 °C) after which Chl a was determined (Lorenzen 1967; Bartoli et al. 2003). During the destructive sampling at the end of experiments, the sediment (AnoxSed and NatSed) was sieved through a 1 mm mesh to recover benthic macrofauna. Fauna was identified to species level and counted for abundance. The wet weight, corrected for shell weight (25%) (Rumohr et al. 1987), was used for biomass estimation. Statistical analyses A one factor ANOVA tested the differences in Nereis diversicolor abundance and biomass and macroalgal biomass between NatSed, SurfSed and AnoxSed. A two-factor ANOVA was used to evaluate the differences in oxygen and nutrient fluxes between treatments (NatSed, AnoxSed and SurfSed). Time was used as a fixed factor and the same start values (InitSed) were used for all treatments. Homogeneity of variances was tested using Cochran’s test. Differences were considered significant at p ≤ 0.05.
Results Macroalgae and benthic microalgae A green-algal mat started to grow in the cores containing 5 mm surface sediment (SurfSed) on day 4 of the experiment. In cores exposed to anoxic conditions for ~ 5 days (AnoxSed), the growth of green-algae started 4 days after the lids were re-opened and the overlying water was re-oxygenated, i.e. on day 11. After additional ~ 10 days (days 19-22), however, the algae progressively disappeared from the sediment surface.

- 6 -
0
100
200
300
400
500
NatSed AnoxSed SurfSed
Gre
en a
lgal
bio
mas
s (g
dw
t m-2
)
Fig. 2 Biomass of green algae on Day 20 in the three treatments. NatSed (10-cm deep
undisturbed sediment); AnoxSed (10-cm deep undisturbed sediment exposed to anoxia for 5 days; SurfSed (5-mm surface sediment, no macrofauna) n = 3+ SE
In contrast, no growth of macroalgae was observed in the NatSed treatment. On day 20 (Fig. 2), there was a significantly (p = 0.03) higher mean algal biomass in the SurfSed (282 g DW m-2) treatment compared with the AnoxSed (37 g dwt. m-2 ) and the NatSed (0 g DW m-2 ) treatments (Fig. 2). There was a 50 – 80% cover of macroalgae in cores with only 5 mm surface sediment (SurfSed) and 10-15 % in the AnoxSed treatment. Benthic microalgae were at the start of the experiment dominated by pennate diatoms such as Navicula sp. and Nitzschia sp. During the experiment, different species of cyanobacteria (e.g. the genera Oscillatoria, Anabaena and Merismopedia) started to grow in all cores. On day 20, only cynobacterial mats were found in the cores with natural sediment including fauna (NatSed). The cyanobacterial mats progressively increased in the NatSed treatment, and were also observed (day 20) in cores where macroalgae were temporarily removed for quantification. Mats of cyanobacteria were simultaneously observed in situ in a nearby bay with environmental conditions similar to those
in this study (A. Norkko, pers comm.). Chl a was similar between treatments and did not change significantly over time. However, on day 20, the biomass seemed higher (not statistically confirmed) in the AnoxSed treatment than in the NatSed and the SurfSed treatments (Fig. 3). Fauna The polychaete Nereis diversicolor was the dominant species of macrofauna throughout the experiment. Other types of fauna were found in the NatSed and in the AnoxSed treatments, including e.g. chironomides, the bivalves Mya arenaria and Macoma baltica, the amphipod Corophium sp., and the gastropod Hydrobia sp., but N. diversicolor was the only species that survived the anoxic treatment. Individuals of Nereis diversicolor were frequently seen feeding on aggregates of green-algae on the sediment surface. This observation was particularly evident in the NatSed and the AnoxSed treatment before and 11 days after the anoxic period.

- 7 -
0
5
10
15
20
25
30
35
40
45
50
InitialSed NatSed AnoxSed SurfSed
Chl
a, m
g m
-2
Fig. 3 Chlorophyll a content in the sediment at start (InitSed) and at the end of the experiment
(NatSed ,10-cm deep undisturbed sediment; AnoxSed (10-cm deep undisturbed sediment exposed to anoxia for 5 days; SurfSed, 5-mm surface sediment, no macrofauna). n = 3 + SE.
The abundance and biomass of N. diversicolor were similar between treatments (mean abundance 580 - 700 ind. m-2 and mean biomass 145 – 150 g WW m-2, respectively) (Fig 4a). However, the biomass increased (p = 0.05) during the experimental period for the AnoxSed and the NatSed treatments. There was no visual confirmation of faunal activity on the sediment surface during the week following the anoxic treatment in the AnoxSed treatment. It was not until 11 days after the anoxic period that N. diversicolor was again seen on the sediment surface. For comparison, faunal activity was during this time continuously observed in the NatSed treatment. Harpacticoid copepods and different species of nematodes dominated the benthic meiofauna and constituted between 40 and 50%, while Ostracodes made up between 5 and 20% of the total meiofauna. No meiofauna was found in the cores exposed for anoxic conditions.
Benthic fluxes Community respiration (CR) was similar between treatments and over time, although there was a tendency of lower rates of oxygen consumption in the SurfSed compared with the AnoxSed and the NatSed treatments (Fig. 5). Mean net primary production (NPP) was highest during the initial sampling (310 mmol m-2 d-1, Day 1). On the subsequent sampling occasions (Day 7, 9 and 20), mean NPP was highest in the SurfSed treatment (121 mmol m-2 d-1day 7 and 9 and 261 mmol m-
2 d-1day 20, Fig. 5). Excluding the initial sampling, highest NPP was observed for the SurfSed treatment on the last sampling occasion (261 mmol m-2 d-1). NPP seemed higher on day 20 compared with day 7 and 9, particularly for the AnoxSed treatment where a net consumption (negative NPP) turned to a net production of oxygen following the anoxic period (Fig 6). Differences in NPP were not statistically confirmed.

- 8 -
a
0
100
200
300
400
500
600
700
800
InitialSed NatSed AnoxSed
Abu
ndan
ce (i
nd. m
-2)
0
20
4060
80
100
120140
160
180
Biom
ass (g w.w
. m-2)
b
0
0.05
0.1
0.15
0.2
0.25
0.3
InitSed NatSed AnoxSed
Ind.
wei
ght o
f Ner
eis
dive
rsic
olor
Fig. 4 (a) Abundance (left axis) and biomass (right axis) of live Nereis diversicolor on Day
33 in the three treatments. NatSed (10-cm deep undisturbed sediment); AnoxSed (10-cm deep undisturbed sediment exposed to anoxia for 5 days; SurfSed (5-mm surface sediment, no macrofauna). n = 3 + SE. (b) Individual weight of Nereis diversicolor on Day 1 InitSed and on Day 33 in NatSed and AnoxSed. NatSed (10-cm deep undisturbed sediment); AnoxSed (10-cm deep undisturbed sediment exposed to anoxia for 5 days. n = 4 + SE.
At the initial sampling, NH4
+ (934 µmol m-
2 d-1), DIP (52 µmol m-2 d-1) and Si(OH)4 (617 µmol m-2 d-1) were released to, while NO3
- (- 753 µmol m-2 d-1) was removed from the overlying water (Fig. 6). The largest effect from the experimental treatments seemed to occur on the second
sampling occasion. For example, DIP was released from the sediment (150 µmol m-2 d-1) following the anoxic period (AnoxSed, day 7), while DIP was removed from the overlying water in the two other treatments (111µmol m-2 d-1 (NatSed) and 216µmol m-2 d-1 (SurfSed). .

- 9 -
-100
0
100
200
300
400
500
Initial Day 7 and 9 Day 20
NP
P (m
mol
O2
m-2
d-1
)
NatSed AnoxSed SurfSed
-500
-400
-300
-200
-100
0
Initial Day 7 and 9 Day 20
CR
(mm
ol O
2 m
-2 d
-1)
Fig. 5 Daily net primary production (NPP) and community respiration (CR) in the initial
cores and in the three treatments. NatSed (10-cm deep undisturbed sediment); AnoxSed (10-cm deep undisturbed sediment exposed to anoxia for 5 days; SurfSed (5-mm surface sediment, no macrofauna). n = 3 + SE.
For the NatSed and the AnoxSed treatments, benthic NH4
+ fluxes were not statistically different between the initial and the second sampling occasions (p = 0.87), while significantly (p = 0.0002, AnoxSed and p = 0.03, NatSed) lower on day 20. In contrast, although not statistically confirmed, NH4
+ fluxes for the SurfSed treatment seemed higher at the last (1092 µmol m-2 d-1) compared with the second (224 µmol m-2 d-1) sampling. NO3
-
was taken up by the sediment, except for the SurfSed and the AnoxSed treatments during the second sampling where mean fluxes indicated a small release of NO3
- to the overlying water. Fluxes of Si(OH)4 were highly variable over time (Fig. 6). At the initial and the last sampling, Si(OH)4 was released to the overlying water for all treatments. In contrast, Si(OH)4 was taken up by the sediment at the second sampling.

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 10 -
0
500
1000
1500
2000
NH
4 (u
mol
m-2
d-1
)
NatSed AnoxSed SurfSed
-1000
-500
0
500
1000
1500
2000
NO
3 (u
mol
m-2
d-1
)
-500
0
500
1000
1500
2000
DIP
(um
ol m
-2 d
-1)
-1500
-1000
-500
0
500
1000
1500
2000
Initial 7 and 9 20
Si(O
H) 4
(um
ol m
-2 d
-1)
Fig. 6 Daily (24-h) fluxes of inorganic nitrogen (NH4+ and NO3
-), phosphorus and silica for the initial sediment and the three treatments. NatSed (10-cm deep undisturbed sediment); AnoxSed (10-cm deep undisturbed sediment exposed to anoxia for 5 days; SurfSed (5-mm surface sediment, no macrofauna). n = 3 + SE.

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 11 -
Discussion Our main hypothesis was that periods of oxygen depletion stimulate the growth of green-algal mats, and that this stimulation is not only caused by an increased availability of reactive nutrients (above all NH4
+ and DIP) but may also occur as a consequence from a reduced macrofaunal grazing-pressure on the algae. The results of our experiment indeed supported the idea that grazing by the polychaete Nereis diversicolor provides an important control for the establishment of green-algal mats in shallow-water environments. The experiment further confirmed that periods of anoxia and direct feedbacks between oxygen availability and macrofaunal activities govern macrofaunal grazing. Although the oxygen-related stimulation of macroalgal growth is probably a combined effect from an enhanced nutrient release from the sediment and a reduced grazing pressure, obtained results suggest that macrofaunal grazing (either directly on propagules or on mature algae) may be the stronger controller. Generally, nutrients originating from the sediment pore water are important for the growth of macroalgal mats (for references see Sundbäck and McGlathery 2005). In our experiment, the release of nutrients (mainly NH4
+ and DIP) from the sediment increased after the onset of anoxia, likely a direct consequence from a restricted pathway of sediment nitrification and coupled nitrification/denitrification, and the reduction of Fe-oxides in the surface sediment, respectively {Gilbert, 2003 #292; Thamdrup, 2002 #537}. Simultaneously, there was no activity by Nereis diversicolor during the anoxic period. After one week, the growth of propagules and macroalgal filaments started. After additional 10 days, N. diversicolor was again observed to feed on the sediment surface and the newly formed macroalgal mat was more or less completely consumed. The question arises
whether the development of macro-algal mats, in this study continuously observed in the SurfSed treatment and 4 days after termination of the anoxic period in the AnoxSed treatment, was a consequence from a reduced grazing pressure by N. diversicolor or a higher availability of inorganic nutrients? A high nutrient supply seems to counteract the grazing pressure through a fast growth rate of the algae. It is well established that Ulva sp. has a morphology that enables high rates of nutrient incorporation (Wallentinus 1984). Pulses of pore water nutrients to the overlying water may therefore favour opportunistic species such as Ulva spp. compared to, for example, Fucus spp. On the other hand, an established algal mat of Ulva sp. exposed to low nutrient concentrations may be more vulnerable to macrofaunal grazing due to slow (or no) net growth. A preferential grazing of N. diversicolor on established macroalgal mats was in this experiment supported by the weight of individual specimens of N. diversicolor. Mean individual biomass was larger (not statistically confirmed) for individuals feeding on fully-grown green algae (AnoxSed) when compared with those feeding on propagules (NatSed) (Fig. 4b). The increased growth of N. diversicolor after digesting Ulva sp. was also observed in an accompanying study (Engelsen & Pihl, in press). The statistically non-significant results of this study may be related to the comparably short incubation (i.e. grazing) time (10 days). During periods of limited oxygen availability (AnoxSed treatment), grazing by Nereis diversicolor was significantly reduced. The sensitivity of N. diversicolor to low oxygen conditions has previously been investigated (Gamenick et al. 1996; Hansen and Kristensen 1997; SaizSalinas and FrancesZubillaga 1997; Janas et al. 2004), and the polychaete has been described as both an oxy-conformer and oxy-regulator (Fritzsche and Vonoertzen

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 12 -
1995, and references therein), depending on the activity level of the polychaete (Kristensen 1983). Observations suggest that N. diversicolor can withstand up to 16 days of hypoxia (Vismann 1990). In our experiment, individuals resumed a general activity, and grazing on green-algae in particular, approximately 10 days after termination of the anoxic period when the overlying bottom water was re-oxygenated. Our experiment therefore confirms a less active, or non-feeding, period of Nereis diversicolor during anoxia, rather than faunal asphyxiation caused by oxygen limitation in the bottom water. Similar conditions may apply, at least locally and on a small scale, following organic matter
mineralization within or below green-algal mats. Immediately after the anoxic event, benthic nutrient fluxes were different in the AnoxSed compared to the other treatments (Fig. 6). On the last sampling, however, benthic fluxes of nitrate (into the sediment), phosphate and silicate were similar for all treatments. Fluxes of ammonium seemed larger for the SurfSed treatment, but there were significant differences between replicates. The recovery of nutrient fluxes following a period of bottom water anoxia agrees with the observations by (Larson and Sundbäck 2008).
Green algal mats
Nereis diversicolor
Inorganicnutrients Microphytobenthos
Oxygen
_ _
_
__
+
+ +
_
+
+
_
Fig. 7 Conceptual model for the couplings between Nereis diversicolor, anoxia, inorganic
nutrients, microphytobenthos and green algal mats in shallow-water areas. + positive effect, – negative effect.

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 13 -
The release of NO3 immediately after the anoxic period in the AnoxSed cores indicates a rapid re-oxygenation of the sediment surface and a stimulated sediment nitrification. In addition to oxygen supplied from the overlying water, a rapid re-oxygenation of the sediment surface is supported by the photosynthetic activities of benthic diatoms (Larson and Sundbäck 2008). A stimulated activity by diatoms after the anoxic event was in this study also evident by the removal of silicate from the overlying water. We have summarized the main results from our experiment and the suggested links between organic matter grazing by Nereis diversicolor, bottom-water anoxia and the development of macroalgal mats in a conceptual model (Fig. 7). The green algal mat appears to be a key player in a complex system of sediment biogeochemistry (organic matter mineralization and mobilization of nutrients to the pore water), physical processes (diffusion/advection of pore water and bottom water), and activities by larger benthic organisms (above all macrofauna and MPB). Important prerequisites for N. diversicolor controlling the growth of macroalgal mats by grazing include, for example, the close proximity and availability of propagules, as well as the timing between greenalgal propagules and dense populations of N. diversicolor. In micro-tidal environments, mature green-algal mats are mostly at the water surface. Therefore, the most pronounced effect of N. diversicolor grazing can be expected at the time when green-algal propagules are found on the sediment surface, or when mature mats are close to the sediment, e.g. during low tide {see e.g. Sundbäck, 2005 #396}. Our results thus support the hypothesis that grazing by N. diversicolor may control the initiation and maintenance of macroalgal mats. However, the quantitative
importance of grazing by Nereis is superimposed on the availability of nutrients. For example, under conditions of elevated nutrient supply from land run-off (e.g. during periods of heavy rainy and snow-melt), the growth rate of macroalgal mats is likely enhanced. Such stimulated growth periods associated with an elevated supply of nutrients seem to overrule grazing by Nereis diversicolor. Inversely, during dry periods and calm weather conditions, nutrients are often limiting for macroalgal growth. Under these conditions, grazing by Nereis diversicolor could be the main control for green algal mats. Acknowledgments Fundings was received from the Swedish Research Council for Environment, Agricultural Science and Special Planning (FORMAS), the Memorial Fund of Birgit and Birger Wåhlström and the Captain Stenholm Fund. We would also like to thank Linda Ottosson (Göteborg University) for all help during field and laboratory work and Karl Norling for comments on the design of the experiment. Literature cited Aller, R. C. 2001. Transport and Reactions
in the Bioirrigated Zone. In: Boudreau B., Jorgensen B.B. (eds) The benthic boundary layer: Transport processes and biogeochemistry. Oxford Press 269-301
Bartoli, M., Nizzoli, D. and Viaroli, P. 2003. Microphytobenthos Activity and Fluxes at the Sediment-Water Interface: Interactions and Spatial Variability. Aquatic botany 37:341-349
Blackburn, T. H. and Henriksen, K. 1983. Nitrigen Cycling in Different Types of Sediments from Danish Waters.

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 14 -
Limnology and Oceanography 28 (3):477-493
Dalsgaard, T., Nielsen, L. P., Brotas, V., Viaroli, P., Underwood, G. J. C., Nedwell, D. B., Sundbäck, K., Rysgaard, S., Miles, A., Bartoli, M., Dong, L., Thornton, D. C. O., Ottosen, L. D. M., Castaldelli, G. and Risgaard-Petersen, N. 2000. Protocol Handbook for Nice - Nitrogen Cycling in Estuaries: A Project under the Eu Research Program: Marine Science and Technology (Mast Iii). 62
Fritzsche, D. and Vonoertzen, J. A. 1995. Metabolic Responses to Changing Environmental-Conditions in the Brackish-Water Polychaetes Marenzelleria-Viridis and Hediste-Diversicolor. Marine Biology 121:693-699
Gamenick, I., Jahn, A., Vopel, K. and Giere, O. 1996. Hypoxia and Sulphide as Structuring Factors in a Macrozoobenthic Community on the Baltic Sea Shore: Colonisation Studies and Tolerance Experiments. Marine Ecology Progress Series 144:73-85
Giannotti, A. L. and Mcglathery, K. J. 2001. Consumption of Ulva Lactuca (Chlorophyta) by the Omnivorous Mud Snail Ilyanassa Obsoleta (Say). Journal of Phycology 37:209-215
Hansen, K. and Kristensen, E. 1997. Impact of Macrofaunal Recolonization on Benthic Metabolism and Nutrient Fluxes in a Shallow Marine Sediment Previously Overgrown with Macroalgal Mats. Estuarine Coastal and Shelf Science 45:613-628
Hughes, R. G. 1999. Saltmarsh Erosion and Management of Saltmarsh Restoration; the Effects of Infaunal Invertebrates. Aquatic Conservation-Marine and Freshwater Ecosystems 9:83-95
Hulth, S., Blackburn, T. H. and Hall, P. O. J. 1994. Arctic Sediments (Svalbard) - Consumption and Microdistribution of Oxygen. Marine Chemistry 46:293-316
Janas, U., Wocial, J. A. and Szaniawska, A. 2004. Seasonal and Annual Changes in the Macrozoobenthic Populations of the Gulf of Gdansk with Respect to Hypoxia and Hydrogen Sulphide. Oceanologia 46:85-102
Kristensen, E. 1983. Ventilation and Oxygen Uptake by Three Species of Nereis (Annelida: Polychaeta). I. Effects of Hypoxia. Marine Ecology Progress Series 12:289-297
Larson, F. and Sundbäck, K. 2008. Role of Microphytobenthos in Recovery of Functions in a Shallow-Water Sediment System after Hypoxic Events. Marine Ecology Progress Series 357:1-16
Lorenzen, C. J. 1967. Determination of Chlorophyll and Pheo-Pigments: Spectrophotometric Equations. Limnology and Oceanography 12:343-346
Nordström, M., Bonsdorff, E. and Salovius, S. 2006. The Impact of Infauna (Nereis Diversicolor and Saduria Entomon) on the Redistribution and Biomass of Macroalgae on Marine Soft Bottoms. Journal of Experimental Marine Biology and Ecology 333:58-70
Norkko, A. and Bonsdorff, E. 1996. Population Responses of Coastal Zoobenthos to Stress Induced by Drifting Algal Mats. Marine Ecology Progress Series 140:141-151
Pihl, L. and Rosenberg, R. 1982. Production, Abundance and Biomass of Mobile Epibenthic Marine Fauna in Shallow Waters, Western Sweden. Journal of

OXYGEN DEFICIENCY AND MACROFAUNAL GRAZING ON GREEN-ALGAL MATS
- 15 -
Experimental Marine Biology and Ecology 57:273-301
Pihl, L., Svenson, A., Moksnes, P.-O. and Wennhage, H. 1999. Distribution of Green Algal Mats Throughout Shallow Soft Bottoms of the Swedish Skagerrak Archipelago in Relation to Nutrient Sources and Wave Exposure. Journal of Sea Research 41:281-294
Raffaelli, D. 2000. Interactions between Macro-Algal Mats and Invertebrates in the Ythan Estuary, Aberdeenshire, Scotland. Helgoland Marine Research 54:71-79
Rumohr, H., Brey, T. and Ankar, S. 1987. A Compilation of Biometric Conversion Factors for Benthic Invertebrates of the Baltic Sea. Baltic Marine Biologist 9:1-57
Saizsalinas, J. I. and Franceszubillaga, G. 1997. Tidal Zonation on Mud Flats in a Polluted Estuary Caused by Oxygen-Depleted Water. Journal of Experimental Marine Biology and Ecology 209:157-170
Strickland, J. D. and Parsons, T. R. 1972. A Practical Handbook of Seawater Analysis. Bulletin of Fisheries Reasearch Board of Canada 169:1-311
Sundby, B., Gobeil, C., Silverberg, N. and Mucci, A. 1992. The Phosphorus Cycle in Coastal Marine-Sediments. Limnology and Oceanography 37:1129-1145
Sundbäck, K. and Mcglathery, K. J. 2005. Interactions between Benthic Macroalgal and Microalgal Mats. In Kristensen, E., Haese, R.R., Kostka, J.E. (Eds) Interactions between Macro- and Microorganisms in Marine Sediments, Agu Series. Coastal and Estuarine Studies 60. Chapter 2:7-29
Sundbäck, K., Miles, A., Hulth, S., Pihl, L., Engström, P., Selander, E. and Svenson, A. 2003. Importance of
Benthic Nutrient Regeneration During Initiation of Macroalgal Blooms in Shallow Bays. Marine Ecology Progress Series 246:115-126
Thybo-Christesen, M. and Blackburn, T. H. 1993. Internal N-Cycling, Measured by 15nh4
+ Dilution, in Cladophora Sericea in a Shallow Danish Bay. Marine Ecology Progress Series 100:283-286
Trimmer, M., Nedwell, D. B., Sivyer, D. B. and Malcolm, S. J. 2000. Seasonal Organic Mineralisation and Denitrification in Intertidal Sediments and Their Relationship to the Abundance of Enteromorpha Sp and Ulva Sp. Marine Ecology Progress Series 203:67-80
Valiela, I., Mcclelland, J., Hauxwell, J., Behr, P. J., Hersh, D. and Foreman, K. 1997. Macroalgal Blooms in Shallow Estuaries: Controls and Ecophysiological and Ecosystem Consequences. Limnology and Oceanography 42:1105-1118
Wallentinus, I. 1984. Comparision of Nutrient Uptake Rates for Baltic Macroalgae with Different Thallus Morphologies. Marine Biology 80:215-225
Vismann, B. 1990. Sulfide Detoxification and Tolerance in Nereis-(Hediste)-Diversicolor and Nereis-(Neanthes)-Virens (Annelida, Polychaeta). Marine Ecology Progress Series 59:229-238
Österling, M. and Pihl, L. 2001. Effects of Filamentous Green Algal Mats on Benthic Macrofaunal Functional Feeding Groups. Journal of Experimental Marine Biology and Ecology 263:159-183