lcwrky5: an unknown function gene from sheepgrass improves drought tolerance in transgenic...
TRANSCRIPT

ORIGINAL PAPER
LcWRKY5: an unknown function gene from sheepgrass improvesdrought tolerance in transgenic Arabidopsis
Tian Ma • Manli Li • Aiguo Zhao • Xing Xu •
Gongshe Liu • Liqin Cheng
Received: 11 April 2014 / Revised: 11 May 2014 / Accepted: 16 May 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract
Key message The expression of LcWRKY5 was induced
significantly by salinity, mannitol and cutting treat-
ments. Arabidopsis-overexpressing LcWRKY5 greatly
increased dehydration tolerance by regulating the
expression of multiple stress-responsive genes.
Abstract Based on the data of sheepgrass 454 high-
throughout sequencing and expression analysis results, a
drought-induced gene LcWRKY5 was isolated and cloned,
and the biological role of the gene has not been reported
until now. Bioinformatics analysis showed that LcWRKY5
contains one conserved WD domain and belongs to the
group II WRKY protein family. LcWRKY5 shows high
sequence identity with predicted or putative protein pro-
ducts of Hordeum vulgare, Aegilops tauschii, Triticum
aestivum, Brachypodium distachyon, Oryza sativa, but it
has low homology with WRKYs from dicotyledonous
plants. Several drought-inducibility, fungal elicitor, MeJA-
responsiveness, endosperm, light, anoxic specific induc-
ibility, and circadian control elements were found in the
promoter region of LcWRKY5. Tissue-specific expression
patterns showed that LcWRKY5 is expressed in roots and
leaves, without expression in other tissues. The expression
of LcWRKY5 was induced significantly under salinity and
mannitol stresses but was not notably changed under cold
and Abscisic acid stress. The LcWRKY5 protein exhibits
transcription activation activity in the yeast one-hybrid
system. Overexpressing LcWRKY5 exhibited increased
rates of cotyledon greening and plant survival in transgenic
Arabidopsis compared with wild-type plants under drought
stress, and the expression levels of DREB2A and RD29A in
transgenic plants were enhanced under drought stress.
These results indicated that LcWRKY5 may play an
important role in drought-response networks through reg-
ulation of the DREB2A pathway. LcWRKY5 can be a
candidate gene for engineering drought tolerance in other
crops.
Keywords Sheepgrass � Transcriptional factor �LcWRKY5 � Drought
Abbreviations
ABA Abscisic acid
CaMV Cauliflower mosaic virus
DREB2A Dehydration-responsive element-binding
protein 2A
GFP Green fluorescent protein
GUS b-Glucuronidase
Communicated by Qiao Zhao.
T. Ma and M. Li contributed equally to this work.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00299-014-1634-3) contains supplementarymaterial, which is available to authorized users.
T. Ma � A. Zhao � G. Liu (&) � L. Cheng (&)
Key Laboratory of Plant Resources, Institute of Botany,
Chinese Academy of Sciences, Beijing 100093,
People’s Republic of China
e-mail: [email protected]
L. Cheng
e-mail: [email protected]
T. Ma � X. Xu
Agricultural College, Ning Xia University, Ningxia 750021,
People’s Republic of China
M. Li
Department of Grassland Science, College of Animal Science
and Technology, China Agricultural University, Beijing 100193,
People’s Republic of China
123
Plant Cell Rep
DOI 10.1007/s00299-014-1634-3

MS Murashige and Skoog
ORF Open reading frame
RT–PCR Reverse transcription–PCR
qRT-PCR Quantitative RT-PCR
TAIL-PCR Thermal asymmetric interlaced PCR
Introduction
Environmental stresses, such as drought, high salinity and
low temperature, have adverse effects on plant growth and
seed production (Shinozaki et al. 2003; Barnabas et al.
2008). Water availability is the major limiting factor for
food production in many countries, with agriculture con-
suming approximately 75 % of the water supply in devel-
oped countries and close to 90 % of the water supply in
many developing countries (Pennisi 2008). Therefore, the
identification and characterization of key genes that
mediate plant responses to drought stress provide a pow-
erful method to select for crop plants with enhanced tol-
erance to drought stress (Kasuga et al. 1999; Valliyodan
and Nguyen 2006).
A number of transcriptional factor (TF) families, such as
MYB, DREB, NAC and WRKY, have been reported to be
involved in the regulation of the plant response to drought
stress (Singh et al. 2002; Shinozaki and Yamaguchi-
Shinozaki 2007). The WRKY family is the largest family of
transcription factors in higher plants and is found through-
out the green lineage (green algae and land plants) (Ulker
and Somssich 2004). WRKY family proteins contain one or
two highly conserved WRKY domains (each spanning
approximately 60 amino acid residues) that are character-
ized by the hallmark heptapeptide WRKYGQK and a zinc-
finger structure (Cx4–5Cx22–23HxH or Cx7Cx23HxC)
distinct from other known zinc-finger motifs. Based on the
number of WRKY domains and pattern of the zinc-finger
motif, the WRKY superfamily members from Arabidopsis
can be classified into three groups: group I, group II, and
group III. Members of group I typically contain two WRKY
domains, whereas most proteins with one WRKY domain
belong to group II. Group III proteins also have a single
WRKY domain, but the pattern of the zinc-finger motif is
unique (Eulgem et al. 2000). The first report concerning the
structure of the WRKY domain investigated the AtWRKY4
protein, which consists of a four-stranded b-sheet and a zinc
binding pocket formed by the conserved Cys/His residues
located at one end of the b-sheet (Yamasaki et al. 2005).
The crystal structure of AtWRKY1-C is composed of a
globular structure with five b-strands, and a novel zinc
binding site is located at one end of the b-sheet between
strands b4 and b5 (Duan et al. 2007). This structural
architecture is required for protein–DNA interaction
(Yamasaki et al. 2005).
In plants, many WRKY proteins are mainly involved in
the biotic stress response or defense (Asai et al. 2002; Kalde
et al. 2003; Rushton et al. 2010). Otherwise, WRKY genes
have also been reported to be involved in responses to the
abiotic stresses of wounding (Cheong et al. 2002), drought,
cold, salinity, heat and ABA signaling (Pnueli et al. 2002;
Zhou et al. 2008; Wang et al. 2009b; Prabu et al. 2011; Niu
et al. 2012; Zhu et al. 2013). In addition, it is also evident
that some members of the WRKY family may play
important regulatory roles in the morphogenesis of tric-
homes (Johnson et al. 2002) and embryos (Alexandrova and
Conger 2002), in senescence (Robatzek and Somssich
2001), and in sugar signaling (Rushton et al. 1995; Sun et al.
2003). These reports indicate that WRKY TFs play
important role in regulating different plant processes.
Sheepgrass [Leymus chinensis (Trin.) Tzvel.] is an
important perennial grass of the Poaceae family, and it is
widely distributed on the eastern Eurasian steppe, including
Korea, Eastern Russia, Japan, Mongolia and northern
China (Wang et al. 2010). The plants have diverse adap-
tations to different harsh environments. Sheepgrass has
high tolerance to drought, low temperature and saline-
alkali (Liu et al. 2012). The identification of stress toler-
ance related gene from sheepgrass will provide valuable
gene resources for other crops and forage breeding.
In the present study, we identified a member of the
WRKY family from sheepgrass based on 454 sequence
data, and constructed its phylogenetic relationships and
tried to assign putative orthologs from barley, Arabidopsis
and rice. Further, a drought response gene LcWRKY5 was
characterized using qRT-PCR and transgenic techniques,
these results suggested the involvement of the LcWRKY5
proteins in the response to drought stress.
Materials and methods
Plant materials and treatments
Sheepgrass seedlings (Zhongke No. 2, a variety of the
Institute of Botany, Chinese Academy of Sciences, Beijing,
China) were raised at 25 �C using a 16-h light/8-h dark
photoperiod for 8 weeks before stress treatments. Seedlings
were placed in a growth chamber at 4 �C for cold stress.
For salt and drought stress treatments, seedlings were
irrigated with 400 mM NaCl and 300 mM mannitol,
100 lM ABA, cut off the top 2/3, respectively. The seed-
lings were sampled at 0, 2, 4, 8, 12, 24, 48, and 72 h after
stress treatments. Leaf, rhizome, stem, sheath, and root
tissues were collected from 2-year-old plants grown under
Plant Cell Rep
123

greenhouse conditions as described above. All the samples
were immediately frozen in liquid nitrogen and stored at
-z80 �C for RNA extraction.
Cloning and sequencing analysis of LcWRKY5
According to the data of 454 sequencing and RT-PCR
analysis results, a drought-induced gene (designated as
LcWRKY5) was selected as candidate genes for further
research. To obtain the full-length cDNA of selected
LcWRKY5, total RNA was extracted from 2-week-old
sheepgrass seedlings using TRIzol reagent (TaKaRa)
according to the manufacturer’s instructions. After DNA
was removed using RNase-free DNase I (TaKaRa), the
concentration of RNA was quantified using a Nano spec-
trophotometer. The first-strand cDNAs were synthesized
from 1 lg of total RNA. The full-length cDNAs of
LcWRKY5 were amplified using homologous primers
(forward primer: 50-ATGGAAACGGCGCGGTGGT-30
and 50-CTAATAATCCGGCAGCTTCCGCA-30) based on
sheepgrass LcWRKY5 partial sequences according to 454
sequencing data. The cDNA products were cloned into the
pMD18-T vector (TAKARA) and sequenced. Multiple
sequence alignments and phylogenetic analysis of
LcWRKY5 with complete ORFs were performed using
DNAMAN v5.0 (Lynnon Biosoft Inc., Vandreuil, Quebec,
Canada). Domain architecture analysis was performed
using SMART (http://coot.embl-heidelberg.de/SMART/).
The nuclear localization signal was analyzed by WoLF
SPORT (http://wolfpsort.org/). The 3-D structure was
predicted by CPHmodels-3.0 (http://genome.cbs.dtu.dk/
services/CPHmodels-2_0Server-3D.htm).
Expression pattern of LcWRKY5 in different tissues
and the response to abiotic stresses
Total RNA was extracted from different plant materials
according to the protocols described (TaKaRa). First-strand
cDNA was synthesized using the Primescript II first-strand
cDNA synthesis kit (TaKaRa). LcACTIN and AtACTIN
(Table 1) were used as internal reference genes to assess
gene expression levels from sheepgrass and Arabidopsis.
For semi-quantitative RT-PCR, the cycle parameters were
as follows: initial denaturation at 94 �C for 5 min; 28
cycles at 94 �C for 30 s, 56 �C for 30 s, and 72 �C for 30 s;
and a final extension at 72 �C for 10 min. The PCR pro-
ducts were analyzed by agarose gel electrophoresis. All
quantitative RT-PCR analyses were performed in a 20-ll
volume containing 10 ll of 2 9 SYBR Premix Ex Taq mix
(TaKaRa), 0.2 mM primers, and 2 ll of diluted (1:20 v/v)
first-strand cDNA, with an initial denaturation step (95 �C
for 2 min) followed by 40 cycles at 94 �C for 5 s, 60 �C for
20 s, and 72 �C for 20 s. The relative mRNA ratios were
calculated using the 2-DDCt formula (Livak and Schnittgen
2001). All semi-quantitative RT-PCR and qRT-PCR
experiments described in this section were performed using
three biological replicates. The data were expressed as
mean ± standard error. The primers for semi-quantitative
RT-PCR and qRT-PCR are listed in Table 1.
Isolation and analysis of the LcWRKY5 promoter
To identify the putative cis-acting regulatory elements, the
promoter sequence was isolated using TAIL-PCR (Liu
et al. 1995) with gene-specific primers for SP1 (50-
TATGCTGACGAACCCCATCTGC-30), SP2 (50-
CAGAAAACGAAGCCACGCAGAG-30) and SP3 (50-
AGCGGCAGACAGCACATCAA-30) and three arbitrary
degenerate (AD) primers: AD1 (50-NTCGA(G/C)T(A/
T)T(G/C)G(A/T)GTT-30), AD2 (50-NGTCGA(G/C)(A/T)
GANA(A/T)GAA-30) and AD3 (50-(A/T)GTGNAG(A/
T)ANCANAGA-30). The PCR procedure was performed
as described by Liu et al. (1995). Promoter cis-elements
were identified using the algorithm developed by Higo
et al. (1999) and are available at http://www.dna.affrc.go.
jp/PLACE/sigresult.html and Plant CARE (Plant cis-acting
Table 1 Primer used for quantitative real-time PCR
Primer Sequence Primer Sequence Function
LcACTIN-F TGCTGACCGTATGAGCAAAG LcACTIN-R GATTGATCCTCCGATCCAGA qPCR
LcACTIN-F GTGCTTTCCCTCTATGCAAGTGGT LcACTIN-R CTGTTCTTGGCAGTCTCCAGCTC RT-PCR
LcWRKY5-F AAAGGAGCAGGGAGAGCACG LcWRKY5-R CCGCCATTGATACCCGTCTT qPCR
LcWRKY5-F GCAAGAAAAGGAGCAGGGAG LcWRKY5-R TGGTGAGGTCCAGCGTGATG RT-PCR
AtACTIN-F TGCTGACCGTATGAGCAAAG AtACTIN-R GATTGATCCTCCGATCCAGA qPCR
AtABF3-F AACGCTGGGAGAGATGACTTTGGA AtABF3-R TCCCAAGACCTCCATTACTGCCAA qPCR
AtP5CS1-F TAGCACCCGAAGAGCCCCAT AtP5CS1-R TTTCAGTTCCAACGCCAGTAGA qPCR
AtDREB1A-F AGGAGACGTTGGTGGAGGCT AtDREB1A-R ACGTCGTCATCATCGCCGTC qPCR
AtDREB2A-F AAACCTGTCAGCAACAACAGCAGG AtDREB2A-R TTAAGCCTGCAAACACATCGTCGC qPCR
AtRD29A-F TGTGCCGACGGGATTTGACGGA AtRD29A-R TCCGTCTTTGGGTCTCTTCCCAGC qPCR
Plant Cell Rep
123

regulatory element database, http://bioinformatics.psb.
ugent.be/webtools/plantcare/html).
Transcriptional activation activity of the LcWRKY5
protein
The transcription activation activity of the LcWRKY 5
protein was investigated using the yeast one-hybrid system.
The full-length LcWRKY5 cDNA was amplified using the
forward primer 50-CGGAATTCATGGAAACGGCGC
GGTGGTC-30 (EcoRI site underlined) and reverse primer
50-GCGTCGACATAATCCGGCAGCTTCCGCA-30 (SalI
site underlined). The PCR products were inserted into the
corresponding restriction sites of the yeast expression
vector pBridge, containing the GAL4 DNA-binding
domain (BD), to obtain pBD-LcWRKY5. pBD-LcWRKY5,
the positive control pGAL4 (Clontech), and the negative
control pBD vector were all transformed into the yeast
strain AH109 (Clontech). The transformed yeast cells were
selected on SD medium without His and Trp to observe
yeast growth. The colony-lift filter b-galactosidase assay
was performed according to the Yeast Protocols Handbook
(Clontech).
Construction of the expression vector and plant
transformation
The ORF of LcWRKY5 was amplified using the forward
primer 50-GAAGATCTATGGAAACGGCGCGGTGGTC-
30 (BglII site underlined) and the reverse primer 50-CCACTAGTATAATCCGGCAGCTTCCGCA-30 (SpeI
site underlined), and the PCR product was cloned into the
BglII and SpeI sites of the pCAMBIA1302 vector under the
control of the LcWRKY5 promoter (W5P). The recombi-
nant plasmid pBI1302-W5P-LcWRKY5 was electroporated
into A. tumefaciens (EHA105) cells for Arabidopsis
transformation using the floral dipping method (Clough and
Bent 1998). LcWRKY5 transgenic plants were screened on
MS medium containing 50 mg ml/1 hygromycin (Roche).
Two LcWRKY5-overexpressing Arabidopsis lines (L3 and
L6) with progeny segregation ratios of 3:1 were selected
for further stress tolerance studies.
Drought tolerance analysis of transgenic plants
For determination of the seed germination rate of trans-
genic plants, the seeds of WT and transgenic plants were
planted on 1/2MS medium containing 300 mM mannitol.
After germination, the green cotyledon rates were scored at
4 days after sowing. For seedling growth experiments, T2
generation transgenic Arabidopsis seeds were grown on
1/2MS medium in a growth chamber with 16 h of light and
8 h of darkness for 1 week and then transferred to the
1/2MS medium supplemented with 300 mM mannitol for
observation. Three weeks after the transfer, the phenotype
was photographed, and the survival rates were calculated.
For phenotype on soil, WT and LcWRKY5-overexpressing
(L3 and L6) seeds were sown on 1/2MS medium for
7 days, then transferred to pot and placed in greenhouse
with light (16 h) at 25 �C (Unit) for 14 days, then withheld
irrigation for 14 days (D14d), and re-watered for 7 days
(R7d), the phenotype was photographed, and the survival
rates were calculated. The soil relative water content was
100, 3.2 and 75.6 %, respectively. Drought tolerance
experiments were repeated at least three times.
Results
Isolation and sequence analysis of LcWRKY5
from sheepgrass
One hundred twenty-eight LcWRKY ESTs were identified
from the transcription profile data of 454 sequencing.
Based on bioinformatics analysis, 18 were examined the
expression levels in stress by RT-PCR methods, and sev-
eral stress-induced full-length LcWRKY cDNAs were
cloned and identified. Among them, an unknown function
gene, designated LcWRKY5 (GenBank Accession No.
1640665), was selected for further research, because it was
remarkably induced by abiotic stresses. The LcWRKY5
gene is 987 bp in length with a 328-amino acid open
reading frame. The molecular mass of the predicted protein
is approximately 35.6 kDa, and its theoretical isoelectric
point (pI) is 7.5.
The LcWRKY5 protein domains were predicted by
SMART and WoLF SPORT software. The results showed
that LcWRKY5 contained a typical WRKY domain
(169–229 bp) and nuclear localization signals (NLSs;
PVGKKRS: AA 105-111; PVKKKVQ: AA 202-208)
bearing the similar three-dimensional structure as the
classical WRKY. LcWRKY5 belongs to the WRKY group
IIa family with one WRKY domain and contains a poten-
tial leucine zipper (LZ, L–x6–V–x6–L–x6–M–x6–L) motif
in the C-terminus that is present in group II WRKY pro-
teins, such as BhWRKY1, NtWIZZ, OsWRKY76 and At-
WRKY18 (Fig. 1a). NCBI BLAST search results showed
that LcWRKY5 has high identity with predicted protein
products of Hordeum vulgare (BAJ90915.1, 92 %), or
putative Aegilops tauschii (EMT16727.1, 82 %), Triticum
aestivum (EMS54632.1, 77 %), Brachypodium distachyon
(XP_003578125.1, 70 %), Oryza sativa (DAA05141.1,
67 %), Sorghum bicolor (XP_002462384.1, 62 %) and Zea
mays (NP_001120723.1, 59 %), but the function of these
gene products was not reported until now, so we speculated
LcWRKY5 was an unknown function gene with WRKY
Plant Cell Rep
123

domain. Otherwise, LcWRKY5 shows a lower similarity to
the dicotyledon proteins BhWRKY1 (ACI62177.1, 43 %),
NtWIZZ (BAA87058.1, 40 %), and AtWRKY18
(NP_567882.1, 37.7 %).
To examine the phylogenetic relationship of the
LcWRKY5 protein with representative members from
other plants, we performed multiple sequence alignment of
the LcWRKY5 protein. A phylogenetic tree of LcWRKY5
and stress-related representative WRKY proteins from
other plants was constructed. As shown in Fig. 1b, the
WRKY proteins were of polyphyletic origin and were
distributed among three groups, group II was further
splitted into five subgroups, namely a, b, c, d and e
according to the analysis by Rushton et al. (2010).
LcWRKY5, OsWRKY76, NtWIZZ, BhWRKY1 and At-
WRKY18 fell into the WRKY group IIa family, although
they have different identities (38–92 %) and diverse
function. Wheat WRKY2, 14, 17, 19, 27, 33, HvSUSIBA2,
Fig. 1 Domain features by SMART and phylogeny of LcWRKY5
with orthologs. a Predicted protein structure of LcWRKY5 by
SMART. The WRKY domain (AA 169-229), DNA-binding domains
(E value 1.39e–34); the WRKY DNA-binding domain [pfam03106,
E value 1.61e–31]; nuclear localization signal (NLSs, PVGKKRS:
AA 105-111; PVKKKVQ: AA 202-208); Leucine Zipper motif (AA
51-83); Alignment of the LcWRKY5 protein with the deduced amino
acid sequences from other species. b Phylogeny of LcWRKY5 was
constructed based on amino acid sequences from stress response
WRKY family members with DNAMAN tree program; c the
predicted dimensional structure of LcWRKY5 by CPHmodels-3.0.
Blue Arrow b-strand; Single line turn
Plant Cell Rep
123

IbSPF1 and AtWRKY1 belonged to group I, they have
different biological functions. TaWRKY11 and TaW-
RKY13 were classified into group III. The 3D structure of
LcWRKY5 protein contains 4 b-sheets by CPHmodels-3.0
(Fig. 1c), a characteristic that is similar to group II WRKY
proteins.
Analysis of the LcWRKY5 promoter
The promoter sequence was isolated and cloned by TAIL-
PCR method using sheepgrass genomic DNA as template
(Supplementary material 2). The PLACE (Plant Cis-acting
regulatory DNA elements) and Plant CARE (Plant cis-
acting regulatory element) databases were used to search
the cis-acting regulatory DNA elements in the promoters of
LcWRKY5. The predicted cis-acting elements in the
LcWRKY5 promoter include: a number of fungal elicitor,
MeJA, auxin, defense and stress, drought, anoxic, GA,
light, ABA responsiveness, circadian control and meristem
expression elements (Table 2). The fungal elicitor, MeJA,
drought inducibility elements appeared more times than
others in the LcWRKY5 promoter. We speculated that
LcWRKY5 might play a role in different signal pathways of
plant growth, development and response to environmental
stimuli processes.
Expression of LcWRKY5 genes under various stresses
and in tissues
The expression patterns of LcWRKY5 in different tissues
and organs, as well as with various treatments, were
investigated using RT-PCR and qRT-PCR. The results
showed that the expression patterns differed in each type of
stress. The expression of LcWRKY5 was induced signifi-
cantly by drought, NaCl and cutting treatments in sheep-
grass but was induced weakly by cold and ABA (Fig. 2a–
e). The expression of LcWRKY5 was induced and reached a
maximum 2 h after cold treatment (Fig. 2a). LcWRKY5
transcripts accumulated quickly in response to salt and
reached its peaks twice after 4 and 48 h of salt treatment
(Fig. 2b). LcWRKY5 transcripts were distinctly induced
and reached a maximum at 48 h with mannitol treatment
(Fig. 2c). The expression of LcWRKY5 was induced
weakly at 4 h after ABA treatment (Fig. 2d). LcWRKY5
Fig. 1 continued
Table 2 Predicted cis-acting elements of LcWRKY5 promoter
Cis-acting elements Function of cis-acting elements Number
TGAC Fungal elicitor responsive element 29
CAANNNNATC Involved in circadian control 1
TGACG MeJA-responsiveness 8
AACGAC Auxin-responsive element 1
ATTTTCTTCA Defense and stress responsiveness 1
GTCAT Required for endosperm expression 3
CAACTG Involved in drought-inducibility 6
CCCCCG Anoxic specific inducibility 3
AAACAGA Gibberellin-responsive element 1
CACGAC Light responsiveness 4
CACGTG Abscisic acid responsiveness 2
TGGTTT Essential for the anaerobic induction 2
GCCACT Related to meristem expression 2
Plant Cell Rep
123

transcripts were significantly induced and reached a max-
imum at 8 h with cutting treatment (Fig. 2e). The expres-
sion of LcWRKY5 was detected in roots and leaves but was
not found in other organs (Fig. 2f).
Transcriptional activation activity of the LcWRKY5
protein
The transcriptional activation activity of LcWRKY5 was
tested using the yeast one-hybrid system. All the plasmids,
pBridge-BD-WRKY5, pBridge-BD-GAL4 (positive con-
trol) and pBridge-BD (negative control), were transformed
into the yeast strain AH109 containing the upstream acti-
vating sequence (UAS), which could be specifically bound
by the GAL4-binding domain. AH109 containing pBridge-
BD could not grow on SD medium without His and Trp
(SD/-His-Trp). However, the cells harboring the pBridge-
BD-WRKY5 and pBridge-BD-GAL4 plasmids could grow
normally on the same medium and exhibited blue signals in
b-galactosidase assays upon addition of X-gal to the
Whatman filter paper (Fig. 3a–c). These results indicated
that LcWRKY5 is a transcriptional activator.
Drought tolerance analysis of LcWRKY5 transgenic
plants
To further investigate the biological function of LcWRKY5,
transgenic Arabidopsis plants were created with the
LcWRKY5 gene construct under the control of the W5P
promoter. Because previous expression analysis results
have shown that the mRNA of LcWRKY5 was significantly
induced by mannitol stress, we examined the seed germi-
nation rate of WT and LcWRKY5-overexpressing plants in
normal 1/2MS medium and 1/2MS medium supplemented
with 300 mM mannitol. No significant difference was
observed in the cotyledon greening rate between wild-type
Fig. 2 Expression patterns of
LcWRKY5 in response to
various stress treatments in
sheepgrass. a Expression
patterns of LcWRKY5 in
4-week-old sheepgrass
seedlings in low temperature
(4 �C) by qPCR. b 400 mM
NaCl/L. c 300 mM Mannitol.
d 100Lm ABA. e Cut off the
top 2/3. The samples were
collected for each condition at
0, 2, 4, 8, 12, 24, 48 and 72 h,
respectively. f Expression
profilings of LcWRKY5 in
different organs of 2-year-old
sheepgrass at the flowering
stage
Fig. 3 Transactivational ability assay of LcWRKY5. a The trans-
formed RH109 yeasts with pBD-LcWRKY5, pGAL4 (positive
control), pBridge (pBD) (negative control) were selected on SD-
Trp-His media. b b-Galactosidase activity assay (The LacZ marker
gene was examined by X-gal assay). c The position of each
transformed yeast cell
Plant Cell Rep
123
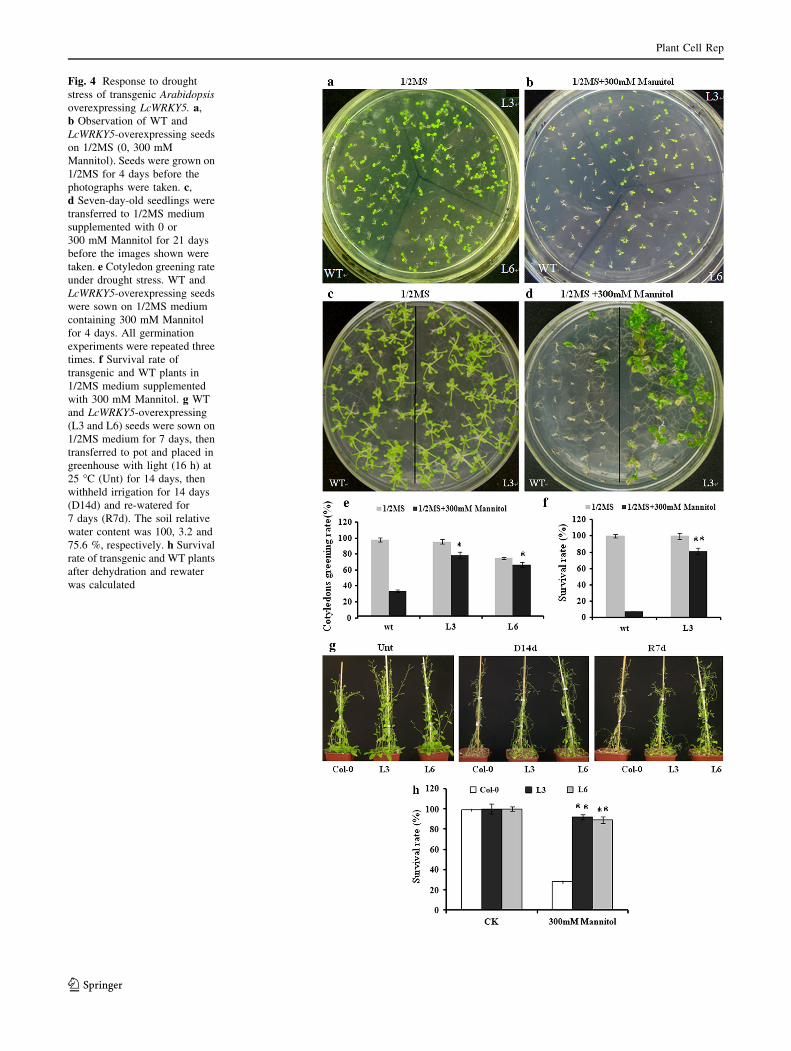
Fig. 4 Response to drought
stress of transgenic Arabidopsis
overexpressing LcWRKY5. a,
b Observation of WT and
LcWRKY5-overexpressing seeds
on 1/2MS (0, 300 mM
Mannitol). Seeds were grown on
1/2MS for 4 days before the
photographs were taken. c,
d Seven-day-old seedlings were
transferred to 1/2MS medium
supplemented with 0 or
300 mM Mannitol for 21 days
before the images shown were
taken. e Cotyledon greening rate
under drought stress. WT and
LcWRKY5-overexpressing seeds
were sown on 1/2MS medium
containing 300 mM Mannitol
for 4 days. All germination
experiments were repeated three
times. f Survival rate of
transgenic and WT plants in
1/2MS medium supplemented
with 300 mM Mannitol. g WT
and LcWRKY5-overexpressing
(L3 and L6) seeds were sown on
1/2MS medium for 7 days, then
transferred to pot and placed in
greenhouse with light (16 h) at
25 �C (Unt) for 14 days, then
withheld irrigation for 14 days
(D14d) and re-watered for
7 days (R7d). The soil relative
water content was 100, 3.2 and
75.6 %, respectively. h Survival
rate of transgenic and WT plants
after dehydration and rewater
was calculated
Plant Cell Rep
123

and transgenic plants (L3) without drought stress, though
some seeds of line L6 did not germinate normally (Fig. 4a).
However, the LcWRKY5-overexpressing plants (L3 and L6)
showed significantly higher cotyledon greening rates (79
and 67 %, respectively) than those of the WT plants (34 %)
under mannitol stress (Fig. 4b, e). The 7-day-old seedlings
of transgenic and wild-type (WT) plants were transferred to
the 1/2MS medium supplemented with 300 mM mannitol
after germination in 1/2MS medium. Images of both plants
were taken 21 days after movement to the mannitol stress
medium (Fig. 4c, d). The survival rates of transgenic plants
were more than 80 % under mannitol stress; a finding that
was significantly higher than that of WT (8 %) (Fig. 4f).
Others, transgenic lines of LcWRKY5 in reproductive stage
Arabidopsis in pot also exhibited more tolerance to water-
deficit stress than wild-type plants, and the survival rates of
overexpression lines were more than 90 %; this was dra-
matically higher than the 30 % survival rate of WT after
drought stress (Fig. 4g, h). These results indicated that the
transgenic plants were more tolerant to drought than the
WT plants. No significant difference was found in devel-
opment and growth between the WT and transgenic plants
in 1/2MS medium without stress.
Expression of abiotic stress response genes
in transgenic plants
To further elucidate the mechanism of the LcWRKY5
transgenic lines underlying the enhanced tolerance to
drought stress, the expression levels of several known
stress-responsive marker genes were analyzed in the
transgenic and WT Arabidopsis plants. The transcription
factor DREB2A and the functional genes RD29A, P5CS1
exhibited increased expression levels in transgenic plants
under drought stress treatments, but the expressions of
DREB1A, P5CS2, RAB18 were lower than that in
LcWRKY5 overexpression plants compared with WT plants
(Fig. 5).
Discussion
Drought stress is one of the biggest environmental threats to
agriculture and crop production. Thus, it is pertinent to
identify new genes that confer drought tolerance to enable
crops to better survive water-deficit stress. Sheepgrass is an
important monocotyledonous forage grass, and it plays an
important role in economic construction and ecological
protection because of its diverse adaptability and good
nutritional value (Bai et al. 2010; Wang et al. 2009a).
Investigation of the molecular mechanism of sheepgrass
adapting to adverse environments provides for potential gene
resources to improve the breeding of other crops by genetic
engineering. We were interested in transcription factors,
particularly those encoded by sheepgrass WRKY family
genes related to various stresses. Analysis of the functions of
these genes is critical to advance our understanding of the
molecular mechanisms governing plant stress response and
Fig. 5 Expression level of
drought-responsive genes in
WT and transgenic plants Total
RNA was extracted from WT
and transgenic seedlings grown
in 1/2MS medium for 2 weeks
and transferred to 1/2MS
supplemented with 300 mM
Mannitol. The samples were
taken at 0, 6, 12, and 24 h after
drought stress treatment. The
transcript levels were measured
by real-time RT-PCR. Actin
was used as an internal control.
The expression levels of
drought-responsive genes
DREB2A, RD29A, DREB1A,
P5CS1, P5CS2 and RAB18 in
WT plants and transgenic plant
lines L3 and L6 under drought
stress, respectively
Plant Cell Rep
123

tolerance, ultimately leading to enhancement of stress tol-
erance in crops through genetic manipulation.
In the current study, we isolated a WRKY family gene
(LcWRKY5) from sheepgrass based on transcriptional
profiles data using 454-sequencing. Based on BLAST
searches in GenBank and multiple sequence alignments
with other WRKY proteins, we found that the predicted
protein contains a typical WRKY domain with a
WRKYGQK motif, which has been found in Arabidopsis,
rice, wheat and barley (Zhang and Wang 2005; Mangelsen
et al. 2008; Zhu et al. 2013). The LcWRKY5 protein showed
high identity with predicted protein products of Hordeum
vulgare (92 %), putative WRKY transcription factor 40
Aegilops tauschii (82 %), Brachypodium distachyon
(70 %), Triticum aestivum (77 %), Oryza sativa (67 %),
Sorghum bicolor (62 %) and Zea mays (59 %), but these
protein products are predicted, putative or hypothetical,
and their function is not reported until now. The LcWRKY5
showed lower similarity to the dicotyledon BhWRKY1
(43 %), NtWIZZ (40 %), and AtWRKY18 (38 %)(Fig. 1). In
previous studies, BhWRKY1 participates in dehydration
tolerance by binding to the W-box elements of the ga-
lactinol synthase (BhGolS1) promoter (Wang et al. 2009b),
NtWIZZ (WRKY-type transcription regulator) plays an
important role in the immediate-early wounding response
(Hara et al. 2000), OsWRKY76 (Ryu et al. 2006), and At-
WRKY18 (Chen et al. 2002) is induced by pathogens,
indicating that the group IIa WRKYs have diverse func-
tions in plants. Wheat TaWRKY2 and TaWRKY19 were
induced by drought, salt and ABA; TaWRKY19 can also be
induced by cold, but TaWRKY2 not response to cold stress.
Transgenic Arabidopsis plants overexpressing TaWRKY2
exhibited salt and drought tolerance compared with con-
trols. Overexpression of TaWRKY19 conferred tolerance to
salt, drought and freezing stresses in transgenic plants.
LcWRKY5 has low identity with TaWRKY2 (13 %) and
TaWRKY19 (15 %), so we speculate LcWRKY5 is an
unknown function protein that may participate in diverse
pathway in plant stress response. In addition, LcWRKY5
has transactivation activity in the yeast one-hybrid system
(Fig. 3), indicating that it is a transcription activator.
More than half of these drought-inducible genes were
also reported to be induced by high salinity and/or ABA
treatment, implicating significant cross-talk between the
drought, high salinity, and ABA response pathways. By
contrast, only 10 % of the drought-inducible genes were
also induced by cold stress (Shinozaki and Yamaguchi-
Shinozaki 2007). Expression pattern analysis showed that
the expression of LcWRKY5 is detected in roots and leaves
and induced significantly by salinity and mannitol treat-
ment but only weakly induced under cold stress treatments,
indicating that LcWRKY5 might play an important role in
the response to drought and salt stress (Fig. 2).
Gene expression is often regulated by the promoters of
genes upstream, and most gene function analyses have
been conducted under the CaMV 35S promoter. Some-
times, overexpression of genes under a component pro-
moter can lead to death or negative effects on plant growth
and development, such as the dwarf or retardation pheno-
type observed in transgenic rice and other plants over-
expressing abiotic stress-related genes (Shen et al. 2003;
Wang et al. 2008). Thus, we constructed the pCAM-
BIA1302-W5P-LcWRKY5 vector for transformation using
the LcWRKY5 promoter (W5P) to obtain precise under-
standing of LcWRKY5 gene function. Overexpression of
LcWRKY5 did not affect notably normal growth in trans-
genic Arabidopsis on 1/2MS medium without stress and
improved the drought tolerance of transgenic plants under
drought stress (Fig. 4). Our results suggested LcWRKY5
improves drought tolerance during germination and seed-
ling development in transgenic Arabidopsis. Otherwise, we
found that there were no notable changes between trans-
genic lines and WT plants after salt stress treatments
(Supplementary material Figure S1).
In previous reports, AtP5CS1 was demonstrated to be
mainly responsible for proline accumulation during salt
and drought stress (Szabados and Savoure 2010). P5CS2
can be activated by avirulent bacteria, salicylic acid (SA)
and ROS signals (Fabro et al. 2004). Our results showed
that the expression of AtP5CS1 was enhanced in transgenic
lines compared with the WT plants in drought stress. The
expression level of AtP5CS2 was lower in transgenic plants
than in WT plants under stress (Fig. 5). Thus, we speculate
the reasons for high drought stress tolerance in transgenic
Arabidopsis might be achieved by increasing the accumu-
lation of AtP5CS1, preceding the raise of proline amount
under drought.
It has been reported that there are ABA-dependent and
ABA-independent pathways exist in drought and salt
stresses. DREB2s are pivotal transcription factors in ABA-
independent pathway, other several stress-responsive genes
AtRD29A, AtRAB18 and AtDREB1A also play important
roles in stress response pathways (Shinozaki and Yamag-
uchi-Shinozaki 2007), and often as marker genes in stress
response. Wheat TaWRKYs affect stress tolerance through
regulation of different downstream genes, such as the
overexpression of wheat TaWRKY2 enhances STZ expres-
sion, whereas TaWRKY19 promotes DREB2A-mediated
activation of RD29A, RD29B and Cor6.6 (Niu et al. 2012).
Here, the expression of AtDREB2A, AtRD29A, AtRAB18
and AtDREB1A was analyzed between LcWRKY5 OE lines
and the wild type under drought. The results showed the
expression levels of DREB2A and RD29A were increased
in transgenic plants under drought stress, but the expres-
sions of DREB1A and RAB18 in LcWRKY5-overexpression
plants were lower than that in WT plants; these results
Plant Cell Rep
123

suggest that the increase of drought stress tolerance in
LcWRKY5-overexpressing Arabidopsis maybe the results
of enhanced expression of transcription factors (DREB2A)
and functional genes (P5CS1 and RD29A).
The predicted cis-acting elements in the LcWRKY5
promoter showed the fungal elicitor, MeJA, drought
inducibility elements appeared more times than others in
the LcWRKY5 promoter (Table 2). The results on expres-
sion analysis and overexpression in Arabidposis indicated
the LcWRKY5 plays important role in drought stress
response. Whether LcWRKY5 plays the role on response or
resistance to fungi and MeJA stress need further
investigation.
In conclusion, we isolated and identified an unknown
function LcWRKY5 gene from sheepgrass. LcWRKY5 is
differentially up-regulated by salinity, dehydration and
cutting. Overexpression of LcWRKY5 could enhance the
DREB2A and RD29A expression levels, and improved
tolerance to drought stress in transgenic Arabidopsis,
indicating that LcWRKY5 functions as a transcriptional
activator for the expression of downstream genes that
contribute to drought tolerance in transgenic Arabidopsis
by ABA-independent signaling pathways. These results
demonstrate that the LcWRKY5 gene plays important roles
in drought stress tolerance processes.
Acknowledgments This work was supported by the National Basic
Research Program of China (‘‘973’’, 2014CB138704), the Project of
Ningxia 3 Agricultural Comprehensive Development Office (NTKJ-
2013-03(1)), the National Natural Science Foundation of China
(31170316), the National High Technology Research and Develop-
ment Program of China (‘‘863’’, 2011AA100209), and the Ministry of
Agriculture of China (2009ZX08009-097B).
Conflict of interest The authors declare no conflict of interest.
References
Alexandrova KS, Conger BV (2002) Isolation of two somatic
embryogenesis-related genes from orchardgrass (Dactylis glom-
erata). Plant Sci 162:301–307
Asai T, Tena G, Plotnikova J, Willmann MR, Chiu WL, Gomez-
Gomez L, Boller T, Ausubel FM, Sheen J (2002) Map kinase
signalling cascade in Arabidopsis innate immunity. Nature
415:977–983
Bai WM, Xun F, Li Y, Zhang WH, Li LH (2010) Rhizome severing
increases root lifespan of Leymus chinensis in a typical steppe of
inner 456 Mongolia. PLoS One 5:e12125
Barnabas B, Jager K, Feher A (2008) The effect of drought and heat
stress on reproductive processes in cereals. Plant Cell Environ
31:11–38
Chen W, Provart NJ, Glazebrook J, Katagiri F, Chang HS, Eulgem T
et al (2002) Expression profile matrix of Arabidopsis transcrip-
tion factor genes suggests their putative functions in response to
environmental stresses. Plant Cell 14:559–574
Cheong YH, Chang HS, Gupta R, Wang X, Zhu T, Luan S (2002)
Transcriptional profiling reveals novel interactions between
wounding, pathogen, abiotic stress, and hormonal responses in
Arabidopsis. Plant Physiol 129:661–677
Clough SJ, Bent AF (1998) Floral dip: a simplified method for
Agrobacterium-mediated transformation of Arabidopsis thali-
ana. Plant J 16:735–743
Duan MR, Nan J, Liang YH, Mao P, Lu L, Li L, Wei C, Lai L, Li Y,
Su XD (2007) DNA binding mechanism revealed by high
resolution crystal structure of Arabidopsis thaliana WRKY1
protein. Nucleic Acids Res 35:1145–1154
Eulgem T, Rushton PJ, Robatzek S, Somssich IE (2000) The WRKY
superfamily of plant transcription factors. Trends Plant Sci
5:199–206
Fabro G, Kovacs I, Pavet V, Szabados L and Alvarez ME (2004)
Proline accumulation and AtP5CS2 gene activation are induced
by plant-pathogen incompatible interactions in Arabidopsis. Mol
Plant Microbe Interact 17:343–350
Hara K, Yagi M, Kusano T, Sano H (2000) Rapid systemic
accumulation of transcripts encoding a tobacco WRKY tran-
scription factor upon wounding. Mol Gen Genet 263:30–37
Higo K, Ugawa Y, Iwamot M, Korenaga T (1999) Plant cis-acting
regulatory DNA elements (PLACE) database: 1999. Nucleic
Acids Res 27:297–300
Johnson CS, Kolevski B, Smyth DR (2002) TRANSPARENT TESTA
GLABRA2, a trichome and seed coat development gene of
Arabidopsis, encodes a WRKY transcription factor. Plant Cell
14:1359–1375
Kalde M, Barth M, Somssich IE, Lippok B (2003) Members of the
Arabidopsis WRKY group III transcription factors are part of
different plant defense signaling pathways. Mol Plant Microbe
Interact 16:295–305
Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, Shinozaki K
(1999) Improving plant drought, salt, and freezing tolerance by
gene transfer of a single stress-inducible transcription factor. Nat
Biotechnol 17:287–291
Liu YG, Mitsukawa N, Oosumi T, Whittier RF (1995) Efficient
isolation and mapping of Arabidopsis thaliana T-DNA insert
junctions by thermal asymmetric interlaced PCR. Plant J
8:457–463
Liu JS, Wang L, Wang DL, Bonser SP, Sun F, Zhou YF, Gao Y, Teng
X (2012) Plants can benefit from herbivory: stimulatory effects
of sheep saliva on growth of Leymus chinensis. PLoS One
7:e29259
Livak KJ, Schnittgen TD (2001) Analysis of relative gene expression
data using real-time quantitative PCR and the 2(-Delta Delta C
(T)). Methods 25:402–408
Mangelsen E, Kilian J, Berendzen KW, Kolukisaoglu UH, Harter K,
Jansson C, Wanke D (2008) Phylogenetic and comparative gene
expression analysis of barley (Hordeum vulgare) WRKY
transcription factor family reveals putatively retained functions
between monocots and dicots. BMC Genom 9:194–210
Niu CF, Wei W, Zhou QY, Tian AG, Hao YJ, Zhang WK, Ma B, Lin
Q, Zhang ZB, Zhang JS, Chen SY (2012) Wheat WRKY genes
TaWRKY2 and TaWRKY19 regulate abiotic stress tolerance in
transgenic Arabidopsis plants. Plant Cell Environ 35:1156–1170
Pennisi E (2008) The blue revolution, drop by drop, gene by gene.
Science 320:171–173
Pnueli L, Hallak-Herr E, Rozenberg M, Cohen M, Goloubinoff P,
Kaplan A, Mittler R (2002) Molecular and biochemical mech-
anisms associated with dormancy and drought tolerance in the
desert legume Retama raetam. Plant J 31:319–330
Prabu G, Kawar PG, Pagariya MC, Prasad DT (2011) Identification of
water deficit stress upregulated genes in sugarcane. Plant Mol
Biol Rep 29:291–304
Plant Cell Rep
123

Robatzek S, Somssich IE (2001) A new member of the Arabidopsis
WRKY transcription factor family, AtWRKY6, is associated with
both senescence- and defence-related processes. Plant J
28:123–133
Rushton PJ, Macdonald H, Huttly AK, Lazarus CM, Hooley R (1995)
Members of a new family of DNA-binding proteins bind to a
conserved cis-element in the promoters of alpha-Amy2 genes.
Plant Mol Biol 29:691–702
Rushton PJ, Somssich IE, Ringler P, Shen QJ (2010) WRKY
transcription factors. Trends Plant Sci 15:247–258
Ryu HS, Han M, Lee SK, Cho JI, Ryoo N, Heu S, Lee YH, Bhoo SH,
Wang GL, Hahn TR, Jeon JS (2006) A comprehensive expres-
sion analysis of the WRKY gene superfamily in rice plants during
defense response. Plant Cell Rep 25:836–847
Shen YG, Zhang WK, He SJ, Zhang JS, Liu Q, Chen SY (2003) An
EREBP/AP2-type protein in Triticum aestivum was a DRE
binding transcription factor induced by cold, dehydration and
ABA stress. Theor Appl Genet 106:923–930
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks
involved in drought stress response and tolerance. J Exp Bot
58:221–227
Shinozaki K, Yamaguchi-Shinozaki K, Seki M (2003) Regulatory
network of gene expression in the drought and cold stress
responses. Curr Opin Plant Biol 6:410–417
Singh K, Foley RC, Onate-Sanchez L (2002) Transcription factors in
plant defense and stress responses. Curr Opin Plant Biol
5:430–436
Sun C, Palmqvist S, Olsson H, Boren M, Ahlandsberg S, Jansson C
(2003) A novel WRKY transcription factor, SUSIBA2, partic-
ipates in sugar signaling in barley by binding to the sugar-
responsive elements of the iso1 promoter. Plant Cell
15:2076–2092
Szabados L, Savoure A (2010) Proline: a multifunctional amino acid.
Trends Plant Sci 15:89–97
Ulker B, Somssich IE (2004) WRKY transcription factors: from DNA
binding towards biological function. Curr Opin Plant Biol
7:491–498
Valliyodan B, Nguyen HT (2006) Understanding regulatory networks
and engineering for enhanced drought tolerance in plants. Curr
Opin Plant Biol 9:189–195
Wang QY, Guan YC, Wu YR, Chen HL, Chen F, Chu CC (2008)
Overexpression of a rice OSDREB1F gene increases salt,
drought, and low temperature tolerance in both Arabidopsis
and rice. Plant Mol Biol 67:589–602
Wang LJ, Li XF, Chen SY, Liu GS (2009a) Enhanced drought
tolerance in transgenic Leymus chinensis plants with constitu-
tively expressed wheat TaLEA3. Biotechnol Lett 31:313–319
Wang Z, Zhu Y, Wang LL, Liu X, Liu YX, Phillips J, Deng X
(2009b) A WRKY transcription factor participates in dehydra-
tion tolerance in Boea hygrometrica by binding to the W-box
elements of the galactinol synthase (BhGolS1) promoter. Planta
230:1155–1166
Wang L, Wang DL, He ZB, Liu GF, Hodgkinson KC (2010)
Mechanisms linking plant species richness to foraging of a large
herbivore. J Appl Ecol 47:868–875
Yamasaki K, Kigawa T, Inoue M, Tateno M, Yamasaki T, Yabuki T,
Aoki M, Seki E, Matsuda T, Tomo Y, Hayami N, Terada T,
Shirouzu M, Tanaka A, Seki M, Shinozaki K (2005) Yokoyama
S. Solution structure of an Arabidopsis WRKY DNA binding
domain. Plant Cell 17:944–956
Zhang YJ, Wang LJ (2005) The WRKY transcription factor
superfamily: its origin in eukaryotes and expansion in plants.
BMC Evol Biol 5:1–12
Zhou QY, Tian AG, Zou HF, Xie ZM, Lei G, Huang J, Wang CM,
Wang HW, Zhang JS, Chen SY (2008) Soybean WRKY-type
transcription factor genes, GmWRKY13, GmWRKY21, and
GmWRKY54, confer differential tolerance to abiotic stresses in
transgenic Arabidopsis plants. Plant Biotechnol J 6:486–503
Zhu XL, Liu SW, Meng C, Qin LM, Kong LN, Xia GM (2013)
WRKY Transcription factors in wheat and their induction by
biotic and abiotic stress. Plant Mol Biol Rep 31:1053–1067
Plant Cell Rep
123