lasy i drewno a zmiany klimatyczne: zagroŻenia i … · uprawy i obrót gmo są możliwe w polsce...
TRANSCRIPT

1
Panel Ekspertów „KLIMAT” 1
LASY I DREWNO A ZMIANY KLIMATYCZNE: ZAGROŻENIA I SZANSE
Termin: 18 czerwca 2013
SESJA 2
MOŻLIWOŚCI ZWIĘKSZANIA SEKWESTRACJI WĘGLA W EKOSYSTEMACH
LEŚNYCH: EFEKTYWNOŚĆ FOTOSYNTEZY A ZMIENNOŚĆ
GENO- I EKOTYPOWA GATUNKÓW DRZEWIASTYCH
ZA POMOCĄ INŻYNIERII GENETYCZNEJ
Dr hab. Justyna NOWAKOWSKA, prof. IBL, Zakład Hodowli Lasu i Genetyki Drzew
Leśnych, Instytut Badawczy Leśnictwa
1. Przegląd problematyki z zakresu zwiększania sekwestracji węgla w
ekosystemach leśnych
Lasy na świecie generują znaczne korzyści ekonomiczne i zajmują
niekwestionowane miejsce w dziedzictwie narodowym każdego państwa. Oprócz
dostarczania surowca drzewnego, lasy pełnią ważną funkcję w ochronie jakości
powietrza i wody, niwelują zmiany klimatu poprzez składowanie ogromnych ilości
węgla w biomasie, oraz tworzą warunki bytowania dla wielu gatunków roślin,
mikroorganizmów i zwierząt.
1 Aktualizacja programu będzie prowadzona na stronie http://npl.ibles.pl/klimat

2
Ze względu na produkcyjną wartość sortymentu drzewnego i długą żywotność
drzew, produkty leśne można traktować jako towar wytwarzany w ramach modelu
gospodarstwa rolnego, choć istnieje duża różnica między uprawami drzew leśnych a
uprawami rolnymi - szczególnie wyraźnie widoczna w kontekście zastosowania
rozwiązań nowoczesnej inżynierii genetycznej. Skuteczne wprowadzenie zmian
genetycznych w rolnictwie, choć dla ograniczonej liczby cech i gatunków, może
stanowić wyzwanie dla naukowców i zarządców związanych z sektorem leśnym w
kontekście możliwości zastosowania drzew GMO w produkcji surowca drzewnego.
Temat ten wzbudza jednak gorące dyskusje i gwałtowne reakcje, którym często
brakuje poparcia obiektywnymi informacjami, zwłaszcza gdy informacje płynące ze
świata nauki są ze sobą sprzeczne i podważające własną wiarygodność.
Choć termin biotechnologii w leśnictwie można rozpatrywać pod wieloma
aspektami (genomika, proteomika, metabolomika, klonowanie in vitro, selekcja
drzew oparta na molekularnych markerach), w niniejszym opracowaniu - po krótkim
opisie metodyki tworzenia drzew transgenicznych, przedstawione będą możliwości
inżynierii genetycznej gatunków drzewiastych w Polsce i na świecie, z
uwzględnieniem zwiększenia biomasy i w konsekwencji sekwestracji węgla.
a) Genetyczna transformacja roślin drzewiastych
Modyfikacja genomu roślin drzewiastych, czyli ich genetyczna transformacja,
polega na wprowadzeniu specyficznej sekwencji obcego DNA do genomu komórki w
celu otrzymania osobników o nowych cechach jakościowych. Informacja genetyczna
danego gatunku, zawarta w DNA, może być wzbogacona w dodatkowe sekwencje
genów, które są wprowadzone do komórki na drodze transformacji wektorowej (za
pomocą bakterii Agrobacterium tumefaciens), lub bezwektorowej (przy użyciu
strzelby genowej). Na ogół, produkcja drzew transgenicznych jest ukierunkowana na
wprowadzeniu do komórek drzewa genu, którego produkt (białko) nadaje korzyści
hodowlane.
Ważnym etapem, jaki poprzedza transformację komórek roślin drzewiastych,
jest wyizolowanie sekwencji odpowiedniego genu, np. genu kodującego syntazę
glutaminową lub genu toksyna Bt kodującego toksyczne białko dla owadów z rzędu
Lepidoptera (Gallardo et al. 1999). Poszukiwanie genów odpowiedzialnych za różne
procesy metaboliczne w roślinie jest możliwe dzięki wykorzystaniu techniki
mikromacierzy DNA (DNA microarrays). Technika ta umożliwia identyfikację i

3
wstępną charakterystykę funkcji genów na poziomie transkrypcji. W ten sposób
odkryto m.in. geny Leafy kontrolujące proces kwitnienia u Arabidopsis thaliana, geny
enzymów szlaku syntezy komponentów ściany komórkowej oraz wiele innych genów,
odpowiedzialnych za procesy wzrostu i rozwoju roślin pod wpływem działania
hormonów takich jak auksyny, gibereliny, etylen i kwas abscysynowy (Campbell et al.
2004).
W końcowym etapie, wyhodowane rośliny transgeniczne poddawane są
weryfikacji pod kątem obecności obcego genu w genomie komórek. W zależności od
zastosowanego konstruktu genowego, proces wstępnej weryfikacji może być
przeprowadzany za pomocą testów ekspresji genów kodujących białka markerowe,
np. GUS lub odporność na antybiotyki. Ostateczna weryfikacja uzyskanych linii
transgenicznych oraz ich potomstwa wymaga przeprowadzenia molekularnych analiz
(techniki Southern-, Northern- i Western-blottingu, PCR oraz testów enzymatycznych
i immunochemicznych), potwierdzających integrację transgenu z genomem rośliny
oraz jego ekspresję w roślinie.
b) badania prowadzone w Polsce
Uprawy i obrót GMO są możliwe w Polsce od 28.01.2013 r. dzięki ustawie o
nasiennictwie, oraz ustawie z dnia 22 czerwca 2001 r. o organizmach genetycznie
zmodyfikowanych (Dz. U. z 2001 r. Nr 76, poz. 811; oraz z 2002 r. Nr 25, poz. 253).
W zakresie badań drzew leśnych, prowadzone są m.in. badania nad
wyhodowaniem transgenicznych topoli pod kierownictwem prof. dr hab. Stanisława
Karpińskiego w Katedrze Genetyki, Hodowli i Biotechnologii Roślin SGGW.
Stworzona przez zespół S. Karpińskiego transgeniczna topola Populus trichocarpa
L. posiada zwiększoną zawartość flawonoidów, co ma wpływ na większą
odporność na infekcje patogenów i ochronę nienasyconych kwasów tłuszczowych
przed utlenieniem (Sprawozdanie z kontroli, 2011). W procesie transformacji zostały
wykorzystane: 1) biorca - topola kalifornijska (P. trichocarpa L.); 2) dawca – A.
tumefaciens – wektor insercyjny pH7GWIWG2(I), pCAMBIA; 3) insert – pCAMBIA,
skonstruowany w Instytucie CAMBIA w Australii, pH7GWIWG2(I) - wektor oparty na
systemie Gateway M.
Zamierzone uwolnienie topoli genetycznie zmodyfikowanej planowane jest do
dnia 30 września 2014 r. na polu doświadczalnym „Wolica”, należącym do Katedry

4
Genetyki, Hodowli i Biotechnologii Roślin SGGW. Po zakończonym projekcie,
poznamy wymierny zysk wyhodowanych transgenicznych topoli w naszym kraju.
Brak jest badań odnośnie zwiększenia biomasy innych gatunków lasotwórczych
w oparciu o inżynierię genetyczną.
c) badania prowadzone na świecie
Zastosowanie inżynierii genetycznej w leśnictwie dotyczy w pierwszym rzędzie
upraw roślin szybko rosnących (topola), o dużym znaczeniu gospodarczym i
stosunkowo łatwym sposobie rozmnażania. Dzięki transformacji roślin drzewiastych,
możemy o wiele szybciej uzyskać pożądany efekt jakościowy przez np. zwiększenie
odporności na herbicydy i szkodliwe owady w porównaniu do tradycyjnych metod
selekcji. Dzięki tym metodom, drzewa na plantacjach lepiej przyrastają i
magazynują dwutlenek węgla w tkankach, bez strat spowodowanych przez
chwasty i żery owadów.
I tak, w USA powstała m.in. hodowla transgenicznych siewek topoli (Populus
trichocarpa x P. deltoides), odpornych na środki ochrony roślin takie, jak herbicyd
Roundup, powszechnie stosowany do zwalczania chwastów szerokolistnych i
trawiastych (Strauss et al. 2001). Aktywny składnik tego preparatu – glifosat, blokuje
kluczowy enzym – syntazę kwasu 5-endo-pirogronoszikimo-3-fosforanowego
(EPSPS) w biosyntezie aminokwasów aromatycznych w komórkach roślin wyższych.
W celu uzyskania odporności siewek topoli na glifosat, wyizolowano z komórek
bakteryjnych (Salmonella typhimurium) gen aroA, który koduje zmutowane białko
EPSPS i wprowadzono go do genomu rośliny (Campbell et al. 2003). Nadekspresja
genu aroA pod kontrolą stałego promotora CaMV 35S u topoli prowadzi do
akumulacji zmutowanego białka EPSPS w komórkach, co jest przyczyną
zmniejszenia powinowactwa białek docelowych dla herbicydu, bez wpływu na
biosyntezę aminokwasów aromatycznych. Dzięki temu, stosowany w uprawach topoli
herbicyd, hamuje selektywnie wzrost chwastów jednoliściennych, nie wpływając na
rozwój siewek topoli. Podobnie, otrzymano odporność na glifosat u siewek
modrzewia (Larix decidua Mill.) oraz mieszańca topoli (Populus tremula x alba)
(Campbell et al. 2003).
Innym ważnym zastosowaniem inżynierii genetycznej w leśnictwie jest hodowla
transgenicznych siewek topoli (Populus trichocarpa x P. deltoides) odpornych na

5
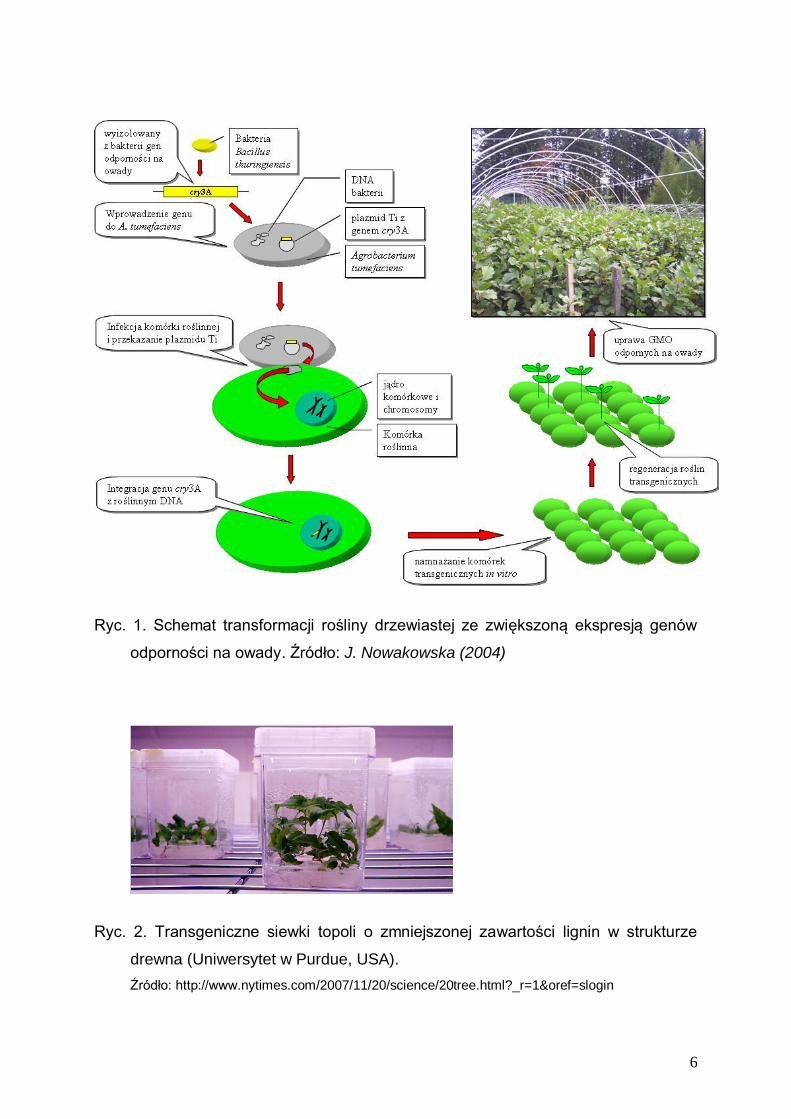
owady z rodziny stonkowatych, takich jak Chrysomela scripta (ryc. 1). Aby temu
zapobiec, zwiększono odporność mieszańca topoli Populus trichocarpa x P. deltoides
przez wprowadzenie do embriogennych komórek roślinnych konstruktu genowego
zawierającego gen kodujący -endotoksyny cryIA(a), cryIA(b) lub cryIA(c) z Bacillus
thuringiensis (Gelernter et al. 1993). Zintegrowany z DNA roślinnym gen Bt koduje
toksyczne białko dla żerujących na blaszkach liściowych larw. W te sposób,
transformacja topoli genem Bt umożliwiła uprawę tego gatunku na plantacjach bez
zastosowania chemicznych środków ochrony roślin - insektycydów.
6
zależności między sygnałami kwantowymi generowanymi w obrębie
fotosystemów a statusem redoks-hormonalnym komórki w odpowiedzi na stresy
biotyczne i abiotyczne. Badania te mają na celu zwiększenie produkcji biomasy
(plonowania) w naturalnych warunkach suszy glebowej.
poziomie ekspresji genów kodujących białka PSII u mutanta mpk4 A. thaliana, którą obroniłem w marcu 2012. Aktualnie zajmuję się badaniem interakcji MAP kinazy MPK4, ze specyficznymi fosfatazami, w komórkach topoli (Populus trichocarpa).
Ryc. 1. Schemat transformacji rośliny drzewiastej ze zwiększoną ekspresją genów
odporności na owady. Źródło: J. Nowakowska (2004)
Ryc. 2. Transgeniczne siewki topoli o zmniejszonej zawartości lignin w strukturze
drewna (Uniwersytet w Purdue, USA).
Źródło: http://www.nytimes.com/2007/11/20/science/20tree.html?_r=1&oref=slogin

7
Jak dotąd, jedynie u roślin modelowych (zielnych i uprawnych), prowadzone
są badania nad wprowadzeniem genów odpowiedzialnych za wzrost aktywności
fotosyntetycznej. Jest to możliwe dzięki modyfikacji szlaku fotosyntezy typu C3 (95
% roślin lądowych, do których przynależą również drzewa leśne), oraz szlaku C4 i
CAM.
Wzrost biomasy może być osiągnięty poprzez zwiększenie wydajności
fotosyntetycznej w chloroplastach, lub zmniejszenie procesu fotooddychania.
Fotosynteza typu C3 jest ograniczona aktywnością kluczowego enzymu rybulozo-
1,5-bisfosforanu karboksylazy-oksygenazy (Rubisco), oraz utratą cząsteczek CO2
przez fotooddychanie. W obecnym składzie atmosfery (21% O2 i 0,035% CO2), tlen
O2 w 25% obniża wydajność procesu fotosyntezy. Dodatkowo, spadek wydajności
fotosyntezy pogłębia się wraz z deficytem wodnym (suszą) i wysoką temperaturą.
Teoretycznie, modyfikacja aktywności enzymu Rubisco na rzecz zwiększonej
aktywności karboksylowej kosztem oksydacyjnej, może prowadzić do zwiększenia
efektywności fotosyntezy poprzez większe przyswajanie CO2 z atmosfery. Pierwsze
takie próby podjęto przez zmianę struktury miejsca aktywnego enzymu via
mutagenezę sterowaną genu Rubisco, oraz przez wprowadzenie większej ilości kopii
tego genu do rośliny. Niestety, nie uzyskano pozytywnych wyników (Normile 1999;
Zelitch 1989), ponieważ mechanizmy działania Rubisco nie są jeszcze gruntownie
poznane u roślin wyższych.
U roślin typu C4, cząsteczki CO2 są magazynowane podczas dnia, kiedy
aktywność oksydacyjna Rubisco jest największa, a uwalniane nocą, kiedy przeważa
aktywność karbokslazy Rubisco. Prowadzi to do większej wydajności fotosyntezy.
Pierwsze próby zmiany systemu asymilacji CO2 u roślin typu C3 na typ C4,
wykonano zwiększając w tkankach ryżu ekspresję karboksylazy
fosfoenolopirogronianowej (PEPC), która w roślinach C4 bierze udział w
przyswajaniu dwutlenku węgla. Podobnie u ziemniaka, zwiększona ekspresja genu
PEPC i dehydrogenazy NADH jabłczanowej, zahamowała aktywność oksygenazy u
Rubisco i zmniejszyła popyt na elektrony fotosystemie PSII przy podwyższonej
temperaturze, przez co wydajność fotosyntezy się zwiększyła (Lipka et al. 1999).
Na podstawie badań aktywności fotosyntetycznej u glonów, stwierdzono
zwiększenie stężenia cząsteczek CO2 wokół przestrzeni komórkowej, co wpłynęło na
większą aktywność Rubisco i w konsekwencji spowodowało wzrost aktywności
fotosyntetycznej (Lieman-Hurwitz et al. 2003).

8
Inne możliwości zwiększenia aktywności fotosyntezy u drzew leśnych
prowadzone są przez blokowanie procesu fotooddychania i wprowadzanie genów
kodujących białka odpowiedzialne za większą przyswajalność cząsteczek wody
przez roślinę. Zmniejszenie procesu fotooddychania może mieć miejsce poprzez
zablokowanie genów biorących udział w biosyntezie fosfoglikolanu – pierwszego
produktu fotooddychania w komórkach chloroplastowych Arabidopsis thaliana
(Kebeish 2006). Natomiast Almeida et al. (2007), wprowadzając gen AtTPS1 z
rzodkiewnika pospolitego do tytoniu, uzyskali lepszą przyswajalność wody z podłoża i
wzrost aktywności fotosyntetycznej na poziomie fotosystemu PSII.
Istotnym problemem, wymagającym dodatkowego rozpoznania efektywności
procesu fotosyntezy, jest dokładne poznanie biosyntezy i regulacji ekspresji
kluczowego enzymu fotosyntezy u roślin C3, tj. Rubisco.
U gatunków drzewiastych, obecnie prowadzone są badania genetyczne nad
zmianami w strukturze wiązań fenylopropenowych w ścianach komórkowych, co
narusza chemiczną strukturę ligniny, usieciowanej wiązaniami eterowymi i
kowalencyjnymi typu węgiel-węgiel (C−C). Lignina jest polimerem, w skład którego
wchodzą monomery - związki organiczne, będące pochodnymi alkoholi fenolowych,
m.in. alkohol koniferylowy, alkohol synapinowy, alkohol kumarylowy.
W ostatnich dziesięcioleciach, wiele uwagi poświęcono badaniu genów
uszkadzających strukturę monomerów lignin (Tabela 1). W Belgii, Francji i USA
prowadzone są badania nad transgenicznymi gatunkami drzew leśnych o
zmniejszonej zawartości lignin w ścianach komórkowych dzięki zmienionym
genotypom enzymów kluczowych w łańcuchu biosyntezy lignin (ligaza C4L,
dehydrogenaza CAD, liaza PAL) (Hu et al. 1999, Peña et Séguin 2001). Zmiana w
ekspresji genu ligazy kumarynowej C4L, która jest kluczowym enzymem w
biosyntezie lignin, prowadzi np. do zwiększenia o 30% zawartości włókien celulozy w
komórkach produkowanego drewna (ryc. 2), co umożliwia zastosowanie mniej
szkodliwych dla środowiska procesów technologicznych w przemyśle papierniczym.
W mieszańcach topoli (np. P. tremula x alba), obniżenie zawartości lignin
uzyskano przez zablokowanie genu COMT, poddanemu regulacji konstytutywnego
promotora 35S z kalafiora (Jouanin et al. 2000).

9
Tabela 1: Spis podstawowych modyfikacji genetycznych u roślin, ukierunkowanych
na biosyntezę lignin

10
Zablokowanie biosyntezy koenzymu CCoAOMT umożliwiło wyhodowanie
transgenicznej topoli o 12% zmniejszonej zawartości lignin. Ekspresja genu białka
F5H (ferulate-5-hydroxylaza) z Arabidopsis pod kontrolą promotora C4H, umożliwiła
Franke et al. (2000), wyhodowanie P. tremula × P. alba o zwiększonej zawartości
monomerów, co daje łatwiejszą delignifikację w trakcie produkcji pulpy. Zawartość
lignin można obniżyć aż do 52% przez wprowadzenie genu hydrolazy CAld5H do
komórek P. tremuloides.
Modyfikacja zawartości składu polisacharydów w ścianie komórkowej
prowadzi do zwiększenia zawartości celulozy, a w związku z tym wiązania węgla w
materii organicznej (Tabela 2). W transgenicznej topoli P. tremula, do której
wprowadzono genami endoglukanazy z rzodkiewnika pospolitego, zmniejszenie
zawartości lignin spowodowało zwiększenie zawartości celulozy ksyloglukanów
nawet o 10% (Shani et al. 2004). W 2007 r., Coleman et al. wyhodowali
transgenicznego mieszańca P. alba × P. grandidentata z wprowadzonym genem
bakteryjnym fosforylazy UDP-glukozowej (UGPase), choć drzewa te rosły jednak
wolniej niż kontrolne.
Nadal trwają badania nad zmianami w strukturze ściany komórkowej u takich
gatunków drzewiastych, jak: Populus sp., Pinus radiata i Eucalyptus.
Również zastosowanie techniki blokowania microRNA w tworzeniu drewna
reakcyjnego u drzew, umożliwia zwiększenie biosyntezy celulozy w ścianie
komórkowej (Lu et al. 2005).
Zwiększenie wzrostu biomasy jest jednym z priorytetowych zadań gospodarki
leśnej. Inżynieria genetyczna dotyczy tu głównie modyfikacji genomów drzew przez
wprowadzenie genów (tabela 2), biorących udział w kodowaniu białek,
odpowiedzialnych za syntezę i transport hormonów wzrostu np. auksyn (gen ugt,
acb). Wprowadzanie tych genów do roślin modelowych powoduje wzrost długości
korzeni u topoli (Salyaev et al. 2006). Transgeniczna topola z genem ekspansyny
PttEXPA1 miała zwiększone długości międzywęźli oraz większą powierzchnię
blaszek liściowych. Są to jednak badania tylko w fazie eksperymentalnej i potrzeba
jeszcze dalszych prac, zanim zostaną wyprodukowane stabilne linie transgenicznych
drzew o tendencji zwiększonego przyrostu biomasy i dobrej jakości włókien drewna.

11
Zakończenie sekwencjonowania innych genomów drzew, oraz ukończony w
2006 roku projekt sekwencjonowania całego genomu topoli P. trichocarpa przez
grupę badaczy Tuskana et al. (2006), przyczyni się do lepszego poznania
genetycznego podłoża regulacji ekspresji informacji genetycznej ukierunkowanej na
większą aktywność fotosyntezy w komórkach, co spowoduje szybszy przyrost masy
drewna i jego dobrą jakość.
Tabela 2. Podsumowanie głównych modyfikacji genetycznych prowadzonych w celu
zwiększenia wzrostu drzew
Genetycznie modyfikowane drzewa, ukierunkowane na produkcję biomasy na
plantacjach drzew leśnych, przyczynią się do większego pochłaniania dwutlenku
węgla, poprzez proces sekwestracji węgla w większych ilościach w częściach
nadziemnych drzew i w glebie (Kellison 2007).

12
Ciekawym osiągnięciem było uzyskanie przez prof. Straussa w 2001 roku w
USA sadzonek transgenicznej topoli (Populus trichocarpa), o zwiększonej asymilacji
jonów amonowych NH4+ (ryc. 3).
Ryc. 3. Modyfikacje prowadzące do wzrostu biomasy. A: zablokowanie ekspresji
genu 4CL powoduje wzrost biomasy transgenicznej topoli w porównaniu do kontroli.
B: zwiększona powierzchnia blaszki liściowej (po prawej) transgenicznej topoli w
odniesieniu do kontroli (po lewej). Źródło: University of California, Davis, 2006
2. Koncepcje rozwiązania, doskonalenia w biotechnologii leśnej
Biotechnologia otwiera szerokie możliwości dla modyfikacji cech genetycznych
drzew leśnych. Wiele jednostek naukowych pracuje nad optymalizacją metod
transformacji różnych gatunków drzew leśnych, w celu zwiększenia odporności na
jeden lub kilka czynników biotycznych i abiotycznych. Poznanie procesów
biochemicznych, regulujących mechanizm wegetatywnego rozmnażania gatunków
drzew leśnych stanowi cel badań wielu ośrodków naukowych. Sam proces
transformacji genetycznej powoduje bowiem zwykle zmniejszenie efektywności
regeneracji zarodków somatycznych z komórek, np. sosny i brzozy in vitro (Aronen i
Häggman 1995). W przypadku Pinus taeda, zauważono, iż proces transformacji
komórek kalusa powoduje zmniejszenie efektywności regeneracji o 40% (Wenk et al.
1999). Rozmnażanie sosny (Pinus taeda L.) i świerka (Picea abies L. Karst.) in vitro
nie jest jeszcze w sposób zadawalający opracowane, a napotkane trudności mają
B

13
podłoże genetyczne (ekspresja genów) lub fizjologiczne (białka kontrolujące proces
embriogenezy, jak np. reduktaza glutationowa). W związku z tym, trwają intensywne
badania nad poznaniem mechanizmów indukujących powstawanie zarodków z
dojrzałych komórek sosny i świerka, tak aby można było rozmnażać wegetatywnie
drzewa elitarne ww. gatunków (Lelu et al. 1999). Rozmnażanie cennych linii
hodowlanych różnych gatunków drzew leśnych drogą somatycznej embriogenezy
pozwala uzyskać dużą ilość rozmnażanego materiału o wyselekcjonowanym
genotypie. Jak dotąd, poznano np. czynnik transkrypcyjny CHAP3a, który indukuje
ekspresję genów biorących udział w regeneracji komórek świerka in vitro (Rutledge
et al., dane nie publikowane) i można przypuszczać, że podobny mechanizm
występuje również u innych gatunków drzew leśnych. Dalsze prace będą
postępować w kierunku identyfikacji innych czynników transkrypcyjnych i lokalizacji
genów odpowiedzialnych za indukcję procesu somatycznej embriogenezy, oraz ich
aktywność w czasie formowania się zarodków drzew iglastych.
Na szczególną uwagę zasługuje również udoskonalenie metod weryfikacji
stabilności transgenów w liniach zmodyfikowanych genetycznie.
Obecnie, w wielu laboratoriach trwają prace nad transformacją ważnych
przemysłowo gatunków drzew leśnych, w celu nadania odporności na herbicydy oraz
czynniki chorobotwórcze takie jak grzyby, owady, wirusy (Tzfira et al. 1998). W
Stanach Zjednoczonych i w Hiszpanii hodowane są w warunkach szklarniowych
transgeniczne odmiany brzozy i sosny, odporne na owady z rzędów Lepidoptera,
Diptera i Coleoptera (Peña i Séguin 2001). W Finlandii uzyskano już pozytywną
transformację komórek brzozy Betula pendula Roth., do której wprowadzono gen
transferazy metylowej PtCOMT z Populus tremuloides. Otrzymane tą drogą
transgeniczne brzozy wykazują zwiększoną odporność na owady z rzędu Coleoptera
w warunkach szklarniowych. Transgeniczna brzoza brodawkowata, posiadająca gen
chitynazy IV, jest odporna na wiele szczepów bakteryjnych (m.in. Streptomyces) i
grzybowych (np. Phytophthora infestans) (Lohtander et al., dane nie publikowane). W
Szwecji, zmodyfikowano genetycznie osobniki mieszańca P. tremula x P. tremuloides
w celu poprawy jakości drewna, zwiększenia przemiany węglowodanów w roślinie,
zwiększonej syntezy giberelin oraz nadanie odporności na owady z rzędów
Coleoptera, Lepidoptera i Diptera poprzez ekspresję toksyny Bt.

14
W Stanach Zjednoczonych i w Izraelu opracowywana jest metoda uzyskania
odporności na niekorzystne warunki klimatyczne (mróz, susza) i środowiskowe
(zasolenie, wysokie stężenie jonów rtęci w podłożu) oraz o zwiększonej asymilacji
jonów amonowych (NH4+) u topoli i sosny (Tzfira et al. 1998).
W Belgii i Francji prowadzone są badania nad transgeniczną topolą o
zmienionym składzie wiązań fenylopropenowych lignin, co umożliwi zastosowanie
mniej szkodliwych dla środowiska procesów technologicznych w przemyśle
papierniczym (Hu et al. 1999). Założone w 2004 roku plantacje topoli transgenicznej
odpornej na Melampsora sp. w INRA pod Orleanem (Francja), zostały niestety
zniszczone przez okoliczną ludność, więc na wyniki tych badań trzeba jeszcze
poczekać (informacja własna).
Transformacje genetyczne nad wprowadzeniem zmian metabolizmu
komórkowego są prowadzone głównie u takich gatunków drzew leśnych jak Pinus
taeda, P. radiata, P. sylvestris, Picea abies, Betula pendula, Pseudotsuga menziesii,
Populus tremula x P. tremuloides i Populus nigra. Jakkolwiek w większości prace te
dotyczą transformacji i ekspresji genów markerowych (GUS), to rozpoczęto już próby
wprowadzenia do komórek drzew leśnych takich genów jak: geny odporności na
herbicydy (BASTA), geny kodujące enzymy szlaku biosyntezy terpenów, których
ekspresja w roślinach transgenicznych zwiększa odporność drzew na szkodniki i
patogeny grzybowe u świerka, np. Phytium dimorphum i Ceratocystis polonica
(Fossdal et al. 2001), geny cyklazy karboksylowej (ACC), które wprowadzone do
komórek sosny zwyczajnej i brzozy brodawkowatej, przyczyniają się do zwiększonej
produkcji etylenu i indukcji dojrzewania zarodków somatycznych in vitro (Lu et al.,
dane nie publikowane).
Ważnym osiągnięciem w hodowli zmodyfikowanych genetycznie drzew będzie
wyhodowanie osobników charakteryzujących się kontrolowanym procesem
kwitnienia. Proces ten można przyspieszać powodując nadekspresję genów LEAFY
lub APETALA u topoli, lub blokować rozwój pąków kwiatowych poprzez wyciszanie
genów (Tzfira et al. 1998). Zahamowanie procesu kwitnienia w roślinach
transgenicznych eliminuje ryzyko uwolnienia transgenu do środowiska.
Wiele przedsiębiorstw przemysłu drzewnego, takich jak SweTreeTechnology AB
(Szwecja), ArborGen llc. (USA), Rubicon Ltd. (USA), Trees and Technology (USA),
Genesis Research and Development Corp. (Nowa Zelandia) oraz międzynarodowa

15
spółka CellFor Inc., wspierają finansowo badania naukowe w poszukiwaniu genów
odpowiedzialnych za cenne przemysłowo cechy drewna oraz finansuje wdrożenie
uzyskanych gatunków transgenicznych drzew do produkcji. Dla przykładu, spółka
ArborGen llc. przeprowadza testy hodowlane transgenicznej topoli na 50 poletkach
doświadczalnych głównie w USA, Brazylii i Nowej Zelandii (dla gatunków Pinus i
Populus), gdzie produkcja transgenicznych sadzonek P. taeda wynosi ok. 150000
sztuk rocznie.
Główne przeszkody (finansowe) napotykane w hodowli i obrocie drzew
genetycznie zmodyfikowanych dotyczą następujących kwestii:
1) koszt wyprodukowania transgenicznych linii w laboratorium jest duży, gdyż
wymaga często prowadzenia badań podstawowych na temat struktury i funkcji
genu, który ma być przeniesiony z obcego organizmu (bakterii, innej rośliny) do
komórki np. topoli,
2) złożony wpływ transgenu na ekologię gatunku w warunkach polowych
wymaga długoterminowych, kosztownych badań. Wraz z niekontrolowanym
przepływem pyłku, może nastąpić krzyżowanie transgenu z formami dzikimi
gatunku, dlatego wymagane są badania prowadzone na szeroką skalę nad
zasięgiem, efektywnością zapylenia, żywotnością mieszańców (modelowanie
komputerowe)
3) koszt patentów, licencji, zgody na publikacje przy wykonywaniu badań z
transgenem niosącym korzyści handlowe (np. przyrostu biomasy), jeśli wymaga
tego prawo,
4) koszty przechowywania dokumentacji i zgodności z przepisami, które mogą
być bardzo wymagające i prawnie ryzykowne dla złożonych programów
obejmujących wiele lat i miejsc objętych uprawą,
5) brak gwarancji, że ustalenia prawne nie ulegną zmianie (np. w interpretacji) w
trakcie długiego testowania lub wprowadzania w obrót handlowy produktu GMO,
6) ryzyko nieumyślnego uwolnienia do środowiska transgenów, które może mieć
miejsce podczas przedwcześnie zakończonych doświadczeń polowych,
pozostawienia plantacji na długi czas po osiągnięciu przez gatunek fazy
kwitnienia, używanie genotypów niekomercyjnych, lub prowadzenia plantacji na

16
geograficznie odległych siedliskach, bez uprzedniego zbadania wpływu
transgenu na środowisko,
7) czynniki zniechęcające do podejmowania kosztownych i ryzykownych badań,
w przypadku ewentualnego odrzucenia popytu rynku na dany produkt GMO i
koszty rozwiązania umowy wynikające z własności genów i metod transferu
genów, będących zazwyczaj w rękach hodowców i producentów drzew, którzy
w większości pokrywają koszty badań w terenie.
Przeszkody te, napotykane również w hodowli roślin uprawnych, wymusiły na
przedsiębiorstwach i rządach państw zaostrzenie przepisów prawnych, które stawiają
szereg wymogów odnośnie zastosowania biotechnologii w środowisku rolnym. Na
przykład w USA, gdzie jest najmniej sprzeciwu społecznego wobec upraw GMO,
planowane jest wprowadzenie obostrzeń warunków upraw polowych w tzw. modelu
eko-genetycznym, czyli po kilkudziesięcioletnim badaniu wpływu transgenu na
lokalne genotypy gatunku w konkretnych warunkach środowiska.
Pierwszym krokiem, do 2030 roku w hodowli drzew transgenicznych powinno
być uzyskanie sterylnych linii, u których nie dochodzi do wytworzenia kwiatów, pyłku i
nasion. Obecnie, wciąż trwają prace nad wyhodowaniem sterylnych mutantów u
roślin modelowych, w tym również topoli (Brunner et al. 2007). Średni koszt
wyhodowania jednej stabilnej linii mutantów (kilka lat) to około 10 000 USD.
Następnym etapem badań, do 2080 roku, będzie doskonalenie produktu –
zmodyfikowanego drzewa, w celu uzyskania precyzyjnych cech hodowlanych i
eliminacja genu markerowego (kodującego odporność na antybiotyki). Jeżeli celem
będzie długoterminowa uprawa leśna, przekraczająca wiek wejścia drzew w fazę
kwitnienia, w USA planuje się zastosowanie transgenów kodujących białka o wąskim
spektrum odporności, np. na patogenny grzyb Cryphonectria parasitica, który jest
główną przyczyną zamierania kasztanowca od początku 1900 w USA, lub na
chrząszcza z rodziny kózkowatych Anoplophora glabripennis pochodzącego z Chin.
Wprowadzenie transgenów hamujących rozwój dużej liczby gatunków – patogenów i
szkodników, niesie ryzyko eliminacji również pożytecznych gatunków bytujących na
drzewach.
Inny ważny aspekt to stabilność wprowadzonego genu do rośliny, gdyż często
fragmenty obcego DNA są eliminowane z jądra komórkowego w mechanizmach
segregacji genów w trakcie mejozy. Następne pokolenie nie posiada już

17
wprowadzonej nowej cechy do genomu i kosztowną procedurę transformacji należy
rozpocząć od nowa. W badaniu transmisji obcych genów w transgenicznym tytoniu
Nicotiana tabacum L., uzyskuje się tylko około 10% linii stabilnych genetycznie, tzn.
takich, których nasiona zawierają wprowadzony gen (obserwacje własne). Ośmioletni
monitoring 384 drzew transgenicznej topoli z genem BAR (odporność na herbicydy)
na plantacji w USA wykazał, że tylko 32 klony zachowały obcy gen w swoich
komórkach. Zachowanie transgenu w topoli może być jak dotąd jedynie
zagwarantowane dzięki wegetatywnemu rozmnażaniu (Brunner et al. 2007).
3. Propozycje zmian w istniejących regulacjach i systemach zarządzania
Nie proponuje się żadnych zmian z zapisie dokumentów regulujących
gospodarkę leśną w Polsce, gdyż potrzeba jeszcze wiele lat badań
interdyscyplinarnych zespołów naukowych aby wytworzyć stabilną, sterylną linię
transgenicznych drzew m.in. o właściwościach odporności na szkodliwe czynniki
biotyczne, o zwiększonej aktywności oksydazy enzymu Rubisco i większej
zawartości polisacharydów w ścianie komórkowej.
Zaproponować można jedynie działania edukacyjne i promocyjne w ośrodkach
prowadzących badania z zakresu biotechnologii drzew leśnych w Polsce, w celu
przybliżenia tej tematyki szerokiemu gronu odbiorców.
4. Podsumowanie
Wraz z pojawieniem się biotechnologii w sektorze leśnym, możliwe jest
zastosowanie rozwiązań genetycznych, ukierunkowanych na produkcję drzew o
zwiększonej biomasie, oraz odpornych na szkodliwe czynniki biotyczne (patogeny i
owady) i abiotyczne (np. wynikające z nagromadzenia składników odżywczych w
glebie). Dzięki wyhodowaniu odmian drzew zmodyfikowanych genetycznie, możliwa
będzie produkcja surowca drzewnego na terenach poddanych antropopresji, przy
zachowaniu pozaprodukcyjnych funkcji lasu, w tym: bytowanie innych gatunków
roślin i drzew, ochrona przed wiatrami, walory rekreacyjne, oraz sekwestracja
dwutlenku węgla w biomasie.

18
Obecnie, 35 krajów prowadzi badania nad 19 gatunkami GMT, z czego 2/3 ma
miejsce w USA. W Chinach jest ponad 500 ha plantacji klonów Populus nigra z
wprowadzonym genem toksyny Bt. W Europie, w prowadzeniu badań i hodowli GMO
obowiązują przepisy prawne: Protokół Kartageński, Konwencja IPPC i Dyrektywa UE
2003/556/EC.
W Polsce, aspekty prawne regulujące zakres badań laboratoryjnych i polowych,
oraz obrót materiałem modyfikowanym genetycznie zawiera ustawa z dnia 22
czerwca 2001 r. o organizmach genetycznie zmodyfikowanych (Dz. U. z 2007 r. Nr
36, poz. 233 z późn. zm.), w szczególności ustawa ta ukierunkowana jest na
czynności kontrolne takie, jak:
sprawdzenie danych użytkownika przeprowadzającego zamierzone
uwolnienie;
sprawdzenie zgodności miejsca zamierzonego uwolnienia z miejscem
określonym w decyzji w sprawie zamierzonego uwolnienia oraz
oznaczenia tego miejsca;
sprawdzenie izolacji przestrzennej miejsca dokonywania zamierzonego
uwolnienia;
sprawdzenie dokumentów związanych z prowadzeniem zamierzonego
uwalniania roślin genetycznie zmodyfikowanych;
sprawdzenie sposobu realizacji przez wnioskodawców warunków
wynikających z posiadanych decyzji Ministra Środowiska.
Warunki przekazywania sprawozdań z przebiegu prowadzonych doświadczeń
polowych są podane na formularzach określonych w załączniku do decyzji Komisji
2003/701/WE z dnia 29 września 2003 r., ustanawiającej zgodnie z dyrektywą
2001/18/WE Parlamentu Europejskiego i Rady UE formularz w odniesieniu do
przedstawiania wyników zamierzonego uwalniania do środowiska zmodyfikowanych
genetycznie roślin wyższych do celów innych niż wprowadzanie do obrotu (Dz. Urz.
UE L 254).
Rekomendacją do Narodowego Programu Leśnego jest utworzenie platformy
badań w zakresie:

19
1) genomiki funkcjonalnej gatunków drzewiastych, ukierunkowanej na
doskonalenie warsztatu (stabilność transgenu, metoda transformacji,
sterylność drzew),
2) opracowania intelektualnego prawa własności transgenu (fragmentu DNA) i
drzewa GMO. Obecnie, każdy element DNA użytego do konstrukcji transgenu
(sekwencje promotora, genu, terminatora) są własnością intelektualną
podmiotów, które nie biorą udziału w komercjalizacji końcowego produktu,
3) prowadzenia "Zarządzania adaptacyjnego" i określenia kosztów licencji w
związku z wysokimi kosztami i długimi terminami prowadzenia upraw drzew
transgenicznych,
4) przejrzystości polityki w prowadzeniu upraw genetycznie zmodyfikowanych
drzew, informowaniu społeczeństwa o prowadzeniu takich upraw,
przeprowadzeniu bilansu zysków i strat (ekonomicznych, ekologicznych i
społecznych).
Literatura źródłowa:
Almeida A.M., Silva A.B., Arau S. S., Cardoso L.A., Santos D.M., Torne J.M., Silva J.M., Paul M.J.,
Fevereiro P.S. 2007. Responses to water withdrawal of tobacco plants genetically engineered with the AtTPS1 gene: a special reference to photosynthetic parameters. Euphytica 154: 113–126.
Aronen T., Häggman H. 1995. Eur J. For. Pathology 25: 197-213.
Brunner A.M., Li J., DiFazio S.P., Shevchenko O., Montgomery B.E., Mohamed R., Wei H., Ma C., Elias A.A., VanWormer K., Strauss S.H. 2007. Genetic containment of forest plantations. Tree Genetics & Genomes DOI 10.1007/s11295-006-0067-8.
Campbell M. M., Brunner A. M., Jones H. M., Strauss S. H. (2003), Plant Biotechnol. J. 1: 141-154.
Coleman H.D., Canam T., Kang K.Y., Ellis D.D., Mansfield S.D. 2007. Overexpression of UDP-glucose pyrophosphorylase in hybrid poplar affects carbon allocation. Journal of Experimental Botany 58: 4257-4268.
Forest and genetically modified trees. Food and Agriculture Organization of the United Nations (FAO), Rome, 2010.
Fossdal C. G., Sharma P., Lönnenborg A. (2001), Plant Mol. Biol. 47(3): 423-435.
Franke R., McMichael C.M., Meyer K., Shirley A.M., Cusumano J.C., Chapple C. 2000. Modified lignin in tobacco and poplar plants overexpressing the Arabidopsis gene encoding ferulate 5-hydroxylase. The Plant Journal 22: 223-234.
Gallardo F., Fu J., Cantón F. R., Garcia-Gutiérrez A., Cánovas F. M., Kirby E. G. 1999. Planta 210: 19-26.
Gelernter W., Schwab G. E. 1993. Bacillus thuringiensis, an environmental biopesticide: theory and practice. Edited by P. F. Entwistle, J. S. Corry, M. J. Ailey and S. Higgs. 1993 John Wiley & Sons Ltd., pp. 90-104.
Hu W. J., Harding S. A., Lung J., Popko J. L., Ralph J., Stokke D. D., Tsai C. J., Chiang V. L. 1999. Nature Biotechnol. 17: 808-812.

20
Jouanin L., Goujon T., de Nadai V., Martin M.-T., Mila I., Vallet C., Pollet B., Yoshinaga A., Chabbert B., Petit-Conil M., Lapierre C. 2000. Lignification in transgenic poplars with extremely reduced caffeic acid O-methyltransferase activity. Plant Physiology 123: 1363-1374.
Kebeish R. M. A. 2006. Construction and molecular analysis of genetically modified C3 plants expressing a glycolate oxidizing pathway inside the chloroplast. Rozpr. Dokt. Institute for Biology I (Botany and Molecular Genetics), Aachen, Niemcy, 182 pp.
Kellison R. 2007. Value-added products from forest biotechnology. Euphytica 154: 279–288. Kellison R.C., Balocchi C.E., Valenzuela S., Rodriguez J. 2007. Forest biotechnology: an extension of
tree improvement. International Journal of Biotechnology 9: 448-459.
Lelu M. A., Bastien C., Drugeault A., Gouez A. L., Klimaszewska K. 1999. Physiol. Plantarum 105: 719-728.
Lieman-Hurwitz J., Rachmilevitch S., Mittler R., Marcus Y., Kaplan A. 2003. Enhanced photosynthesis and growth of transgenic plants that express ictB, a gene involved in HCO3 accumulation in cyanobacteria. Plant Biotechnology 1: 43-50.
Lipka V., Häusler R. E., Rademacher T., Li J., Hirsch H. J., Kreuzaler, F. 1999. Solanum tuberosum double transgenic expressing phosphoenolpyruvate carboxylase and NADP-malic enzyme display reduced electron requirement for CO2 fixation. Plant Sci. 144: 93-105.
Lu S., Sun Y.-H., Shi R., Clark C., Li L., Chiang V.L. 2005. Novel and mechanical stressresponsive microRNAs in Populus trichocarpa that are absent from Arabidopsis. Plant Cell 17: 2186-2203.
Normile D. 1999. Genetic engineers aim to soup up crop photosynthesis. Science 283: 314-316. Nowakowska J. 2004. Zastosowanie roślin transgenicznych w leśnictwie – perspektywy i zagrożenia.
Biotechnologia 3(66): 78-86. Nowakowska J. 2011. Drzewa GMO leśnictwie – iluzja czy przyszłość ? Oikos 12/2011: 16-17. Nowakowska J. A. 2008. Czy w naszych lasach będą rosły drzewa GMO ? Notatnik Naukowy IBL
1(80)/2008(XVI), 4 pp. Nowakowska J.A. 2006. Detekcja ekspresji genów drzew leśnych za pomocą mikromacierzy DNA.
Sylwan 4: 33-43. Peña L., Séguin A. 2001. Trends Biotechnol. 19(12): 500-506.
Salyaev R., Rekoslavskaya N., Chepinoga A., Mapelli S., Pacovsky R. 2006. Transgenic poplar with enhanced growth by introduction of the ugt and acb genes. New Forests 32: 211-229.
Shani Z., Dekel M., Tsabary G., Goren R., Shoseyov O. 2004. Growth enhancement of transgenic poplar plants by overexpression of Arabidopsis thaliana endo-1,4-b-glucanase (cel1). Molecular Breeding 14: 321-330.
Strauss S. H., Campbell M. M., Pryor S. N., Coventry P., Burley J. 2001. J. of Forestry 99(12): 4-7.
Tuskan G.A., DiFazio S., Jansson S. et al. 2006. The genome of black cottonwood, Populus trichocarpa (Torr & Gray). Science 313(5793): 1596-1604.
Tzfira T., Zuker A., Altman A. 1998. Trends Biotechnol. 16: 439-446.
Wenk A. R., Quinn M., Whetten R. W., Pullman G., Sederoff R. 1999. Plant Mol. Biol. 39: 407-416.
Zelitch I. 1989. Selection and characterization of tobacco plants with novel O2 resistant photosynthesis. Plant Physiology 90: 1457-1464.