la replicazione del dna - elearning.uniroma1.it · ogni 30000 300000 nucleotidi (10.000 100.000 ori...
TRANSCRIPT

La replicazione del DNA


Modelli proposti per la replicazione del DNA
� Alla fine degli anni 1950 erano stati proposti tre modelli per la replicazione del DNA
� Modello Conservativo
� Entrambe le eliche parentali rimangono unite insieme dopo la replicazione
� Modello Semiconservativo
� La doppia elica del DNA dopo la replicazione contiene una elica parentale ed una elica figlia (di nuova sintesi)
� Modello Dispersivo
� Le eliche parentali sono distribuite in maniera dispersa nelle doppie eliche del DNA dopo la replicazione

I tre modelli per la replicazione del DNA

Esperienza di
Meselson e Stahl
(1958)

Interpretazione dei dati
After one generation, DNA is “half-heavy”
After ~ two generations, DNA is of two types: “light” and
“half-heavy ”
This is consistent with only the semi-conservative model

La replicazione in E.coli (Cairns 1963)


La replicazione del DNA in vitro
� Studi pioneristici sulla replicazione in vitro del DNA
Arthur Kornberg - premio nobel nel 1959
Kornberg osservò replicazione utilizzando
� Estratto proteico di E. coli
� DNA stampo� DNA stampo
� Nucleotidi radiomarcati
� Incubazione per consentire la sintesi del DNA
� Aggiunta di acido per precipitare il DNA
� Centrifugazione per separare il DNA dai nucleotidi liberi

Arthur Kornberg
nel 1955-58 isola
la DNA polimerasi I

La sintesi avviene in direzione 3’ -> 5’

L’energia per la
formazione del
legame è portata
dal nucleotide
entrante e non
dalla catena in dalla catena in
crescita
Importante per la
correzione degli
errori

La DNA polimerasi I

La DNA polimerasi I

Attività esonucleasiche della DNA pol IEsonucleasi 3’->5’ proofreading
o correttore di bozze

Attività esonucleasiche della DNA pol IEsonucleasi 5’->3’ sintesi di riparo


Struttura ai raggi X del frammeno di Klenow della DNA-polI di E.coli

La DNA polimerasi I

Lo stesso sito attivo per i 4 dNTP

Gli ioni Mg stabilizzano la struttura dello stato di transizione pentacovalente

La DNA polimerasi I

La DNA polimerasi I
Incorporazione di un nucleotide errato 10000 volte più lenta

La DNA polimerasi I

• cambio conformazionale della
DNA pol I prima della
formazione del legame
covalente
• attività esonucleasica per
correggere l’errore
L’energia per la formazione del
legame è portata dal
nucleotide entrante e non dalla
catena in crescita

Meccanismo di proofreadingCorrezione della incorporazione errata di nucleotidi
(forme tautomeriche dei nucleotidi)




La DNA polimerasi richiede un innescoEstremità 3’-OH libera



La replicazione del DNA

La Replicazione necessita di un apparato
enzimatico complesso:
-DNA Polimerasi
-Proteine iniziatrici
-Primasi e Elicasi;
-Proteine che stabilizzano il DNAss e lo proteggono dalla
degradazione;
-Proteine che stimolano le DNA polimerasi.
-Topoisomerasi che eliminano i superavvolgimenti
-Ligasi


DNA elicasi
DNA elicasi-proteine esameriche processive aprono la doppia elica

DNA elicasi

Topoisomerasi rimuovono
i superavvolgimenti
introdotti dall’azione delle elicasi
DNA topoisomerasi

DNA topoisomerasi

Single strand DNA binding proteins (SSBP)

SSB Proteins
DNA




Filamento leader e filamento ritardato

Leading strand e lagging strand
Frammenti di Okazaki

Processività

Processività


Sliding clamp:
proteine con stuttura esagonale
conservate nell’evoluzione conservate nell’evoluzione
(PCNA negli eucarioti)

Sliding clamp isolate da differenti organismi.
Posizionate sul DNA utilizzando ATP


Fasi della replicazione


Eliminazione del primer
procarioti eucarioti
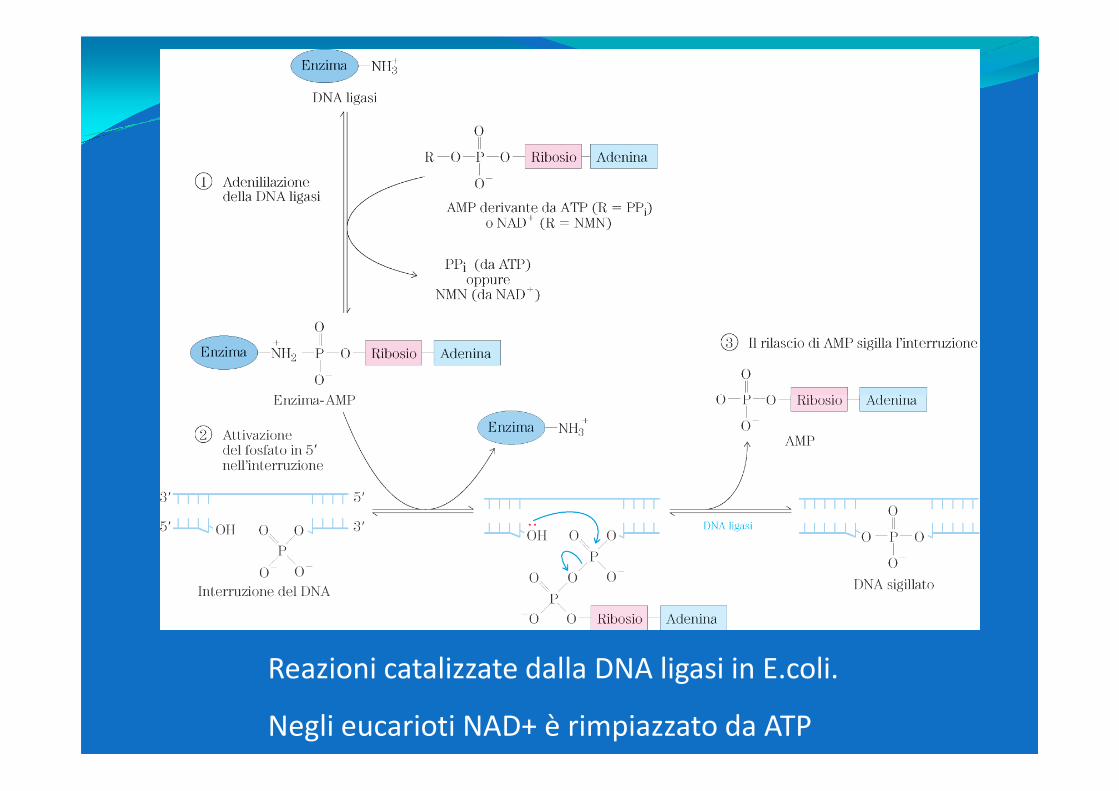
Reazioni catalizzate dalla DNA ligasi in E.coli.
Negli eucarioti NAD+ è rimpiazzato da ATP


LA REPLICAZIONE DEL DNAMeccanismi comuni tra procarioti ed eucarioti
DNA molecola a doppia elica Ogni elica funziona da stampo
Elica parentale Sintesi elica complementare
Sintesi del DNA 5’�3’ DNA-polimerasi
Richiesta di inneschi o primer RNA-polimerasiRichiesta di inneschi o primer RNA-polimerasi
Forca di replicazione Replicazione bidirezionale
Elica parentale 3’�5’ Replicazione continua(leading strand)
Elica parentale 5’�3’ Replicazione discontinua(lagging strand)
Frammenti di Okazaki 100-1000 nt

DNA-POL NEI PROCARIOTIDNA-polimerasi I - Riparo, rimozione primer, solo su doppia elica
(3 attività catalitiche)DNA-polimerasiEsonucleasi (3’�5’) Esonucleasi (5’�3’) Scissione proteolitica produce due frammentiFrammento piccolo (1-323) attività 5’�3’ EsonucleasiFrammento di Klenow (324-928) attività polimerasi e 3’�5’ esonucleasi
dominio 324-517 3’�5’ esonucleasi dominio 517-928 DNA-polimerasi (scissura profonda)
DNA-polimerasi II – RiparoDNA-polimerasi II – Riparo(2 attività catalitiche)
DNA-polimerasiEsonucleasi (3’�5’)
DNA-polimerasi III – Replicazione(2 attività catalitiche)
DNA-polimerasiEsonucleasi (3’�5’) (proofreading)
Cosituita da varie subunità ed un CORE catalitico composto da:Subunità α (Gene polC di E.Coli ) Attività polimerasiSubunità ε Attività 3’�5’ esonucleasiSubunità θ


REPLICAZIONE IN E.ColiInizio della replicazione nel locus OriC (~250bp AT-rich)
La proteina DnaA si lega ad OriC (richiesto ATP)Si associa la proteien DnaB (elicasi) esamerica –> unwinding (richiesto ATP)DNA-girasi (topoII) elimina supercoilingSSB proteins mantengono il DNA a singola elica
Sintesi dei primers o inneschiSintesi primer a RNA sulle leading e ladding strandsPrimosome (600 kDa contiene DnaB-elicasi e DnaG-primasi)
Sintesi del DNALa sintesi è catalizzata dalla DNApol-IIILa sintesi è catalizzata dalla DNApol-IIIReplisoma (2 DNA-pol-III una per lading ed una per lagging strand)2 subunità β formano un anello che scorre lungo il DNALa leading strand viene duplicata in maniera continua (1000 nt/sec) La lagging strand forma un loop e richiede un nuovo primer ogni 1000 nt
frammenti di OkazakiRimozione dei primers
DNA-pol-I rimuove i primers e sintetizza il DNAUnione dei frammenti
DNA-ligasi unisce le interruzioniTermine della replicazione
Regione terminatore, opposta a OriC (sequenze terminatrici TerE-F)Proteine Tus si legano ai siti Ter impedendo l’apertura dell’elica del DNA


replicatore: sequenze agenti in cis ricche in AT
verde:siti di legame dell’iniziatore
blu:elementi di apertura della doppia elica

l’iniziatore di E.coli, la proteina DnaA,è regolata da ATP e da SeqA
Iniziatore interagisce con il DNA e altri fattori

Inizio della replicazione in E.coli


Oloenzima della DNA pol III


REPLICAZIONE IN E.Coli

REPLICAZIONE IN E.Coli






Rimozione dei primer e completamento delle
interruzioni

Il problema della ri-replicazione

Controllo della ri-replicazione


Termine della replicazione
� Il termine della replicazione avviene a livello della
regione ter del cromosoma di E. coli
� La regione ter è ricca in G e T e segnala il termine
della replicazionedella replicazione
� La regione terminatore ter è specificamente legata
dal fattore Tus
� Tus impedisce il passaggio della forca di replicazione
inibendo l’attività elicasica

Terminazione della replicazione in E.coli
Proteina Tus-terminator utilization substance




DNA-POL NEGLI EUCARIOTIDNA polimerasi α: NUCLEARE
Attività primasiAttività esonucleasica assente (no proofreading)Processività moderata: 100 nt/secCoinvolta nella replicazione -> Replicazione lagging strand
DNA polimerasi β: NUCLEAREPartecipa al riparo del DNA
DNA polimerasi γ: MITOCONDRIALESimile nei cloroplasti -> Replicazione DNA non-nucleare
DNA polimerasi δ: NUCLEAREDNA polimerasi δ: NUCLEAREPriva di attività primasiSolo attività 3’�5’ esonucleasiProcessività elevata specie se associata a PCNA
PROLIFERATING CELL NUCLEAR ANTIGEN (PCNA)3 subunità a forma di anelloSimile alla subunità β della DNApol-III
Complesso PCNA-DNAPol δ -> Replicazione lagging/leading strand
DNA polimerasi ε: NUCLEAREElevata processività anche in assenza di PCNASolo attività 3’�5’ esonucleasiRiparo danni da radiazioni UV (Replicazione lagging strand)

REPLICAZIONE NEGLI EUCARIOTIInizio della replicazione
Siti di origine multipli (ogni 3-300 kb)Replicazione non simultanea in tutte le originiUnità di replicazione -> REPLICON con una origineAttivazione sequenziale di gruppi costituiti da 20-80 replicons adiacentiMeccanismo non completamente notoNel lievito:
Inizio replicazione -> ARS (Autonomously replicating sequences) Sequenze di 11bp adiacenti a regioni di DNA facilmente denaturabiliRichiesta attività elicasi (MCM)
Problemi correlati alla replicazione del DNA eucarioticoSincronizzazione della replicazione
Sincronizzazione con il ciclo cellulareStruttura cromatina
DNA eucariotico organizzzato in livelli strutturaliPrimo livello -> nucleosomiDisassemblaggio nucleosomi prima dell’apertura della doppia elicaRiassemblaggio dei nucleosomi dopo la duplicazione
Replicazione estremità 5’Incompleta duplicazione estremità 5’ terminaliTelomeri (sequenze ripetitive)Telomerasi -> duplicazione estremità terminali

“ “ “ “

Origini di replicazione multiple negli eucarioti
ogni 30000 300000 nucleotidi (10.000 100.000 ori nell’uomo)

E.coliLievito
1500
4200 kb40 kb
50.000 bp/min3.600 bp/min
Organismo No. direpliconi
Lunghezzamedia
velocità dellaforca
I repliconi dei genomi di vari organismi
LievitoDrosophilaXenopusTopoVicia faba
5003.500
15.00025.00035.000
40 kb40 kb
200 kb150 kb300 kb
3.600 bp/min2.600 bp/min
500 bp/min2.200 bp/min

Origini multiple della replicazione negli eucarioti

ORC lega e idrolizza ATP

ORC lega e idrolizza ATP



Sintesi del primer
negli eucarioti
switching della polimerasi


Struttura ai raggi X dell’antigene nucleare di proliferazione cellulare PCNA

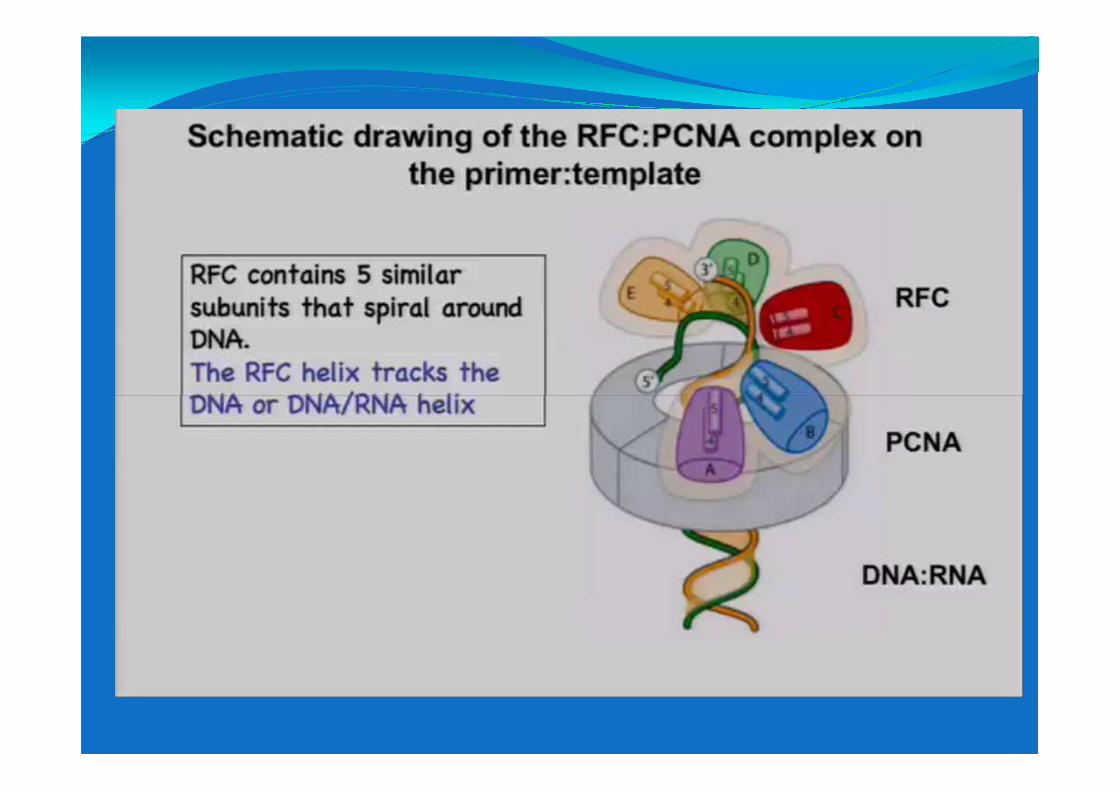

Allungamento





I nucleosomi durante la replicazione



ORC: origin recognition complex
Cdc6, Cdt1: caricatori dell’elicasi
Cdk: chinasi ciclina dipendenti

Attivazione della CDK chinasi ciclina dipendente







I TELOMERIReplicazione estremità 5’ del lagging strand
L’ultimo primer utilizzato non può essere rimosso
Accorciamento del DNA alle estremità
• costituiti da ∼1000 ripetizioni di una piccola sequenza G-RICH (GGGGTTA)estensione a singola elica di 12–16 nucleotidi all’estremità 3’ (G-tail)
• G-tail funziona da stampo per il primer che inizia l’ultimo frammento di Okazaki della lagging strand
alle estremità
TELOMERI: Estremità di ciscun cromosoma

REPLICAZIONE TELOMERICAMentre la forca di replicazione si muove verso l’estremità del cromosoma la replicazione della lagging C-strand genera un DNA con una estremità 3’-sporgente a singolo filamento detta G-tail come risultato della eliminazione del primer di RNA dell’ultimo frammento di Okazaki.La replicazione della leading G-strand può duplicare completamente il DNA completamente il DNA lasciando una estremità piatta che però non è substrato della Telomersi. In questo caso l’estremità 3-sporgente (G-tail) viene generata mediante degradazione esonucleasica della C-strandLa telomerasi estende la G-strand mentre la DNA-polimerasi completa la sua duplicazione (attraverso il meccanismo convenzionale)
Le freccie indicano le estremita 3’
Le linee azzurre indicano la replicazione della leading e lagging strands
Le barre azzurre rappresentano i primers di RNA
Le linee rosse indicano l'allungamento prodotto dalla telomerasi

LA TELOMERASISintetizza e mantiene il DNA telomerico• È una ribonucleoproteina• La molecola di RNA contiene una sequenza (UAACCCC nei mammiferi)
complementare all’unità ripetitica telomerica (GGGGTTA nei mammiferi) • La subunita TERT ("TElomere Reverse Transcriptase") possiede l’attività catalitica• La telomerasi è una trascrittasi inversa (sintetizza DNA su stampo di RNA)
Mantenimento del DNA telomerico:
•G-rich strandTelomerasi
Rif (Rap1 interacting factors)
Cdc13 e Stn1G-tail binding
proteins
•C-rich strandDNA-polimerasi
RPAssDNA binding
protein
Rap1(repressor/activator-site
binding protein) in S. cerevisiae
TRF1(telomeric-repeat binding factor-1)
in humans
Rap1/TRF1Sequence specific
dsDNA binding protein


LA TELOMERASI


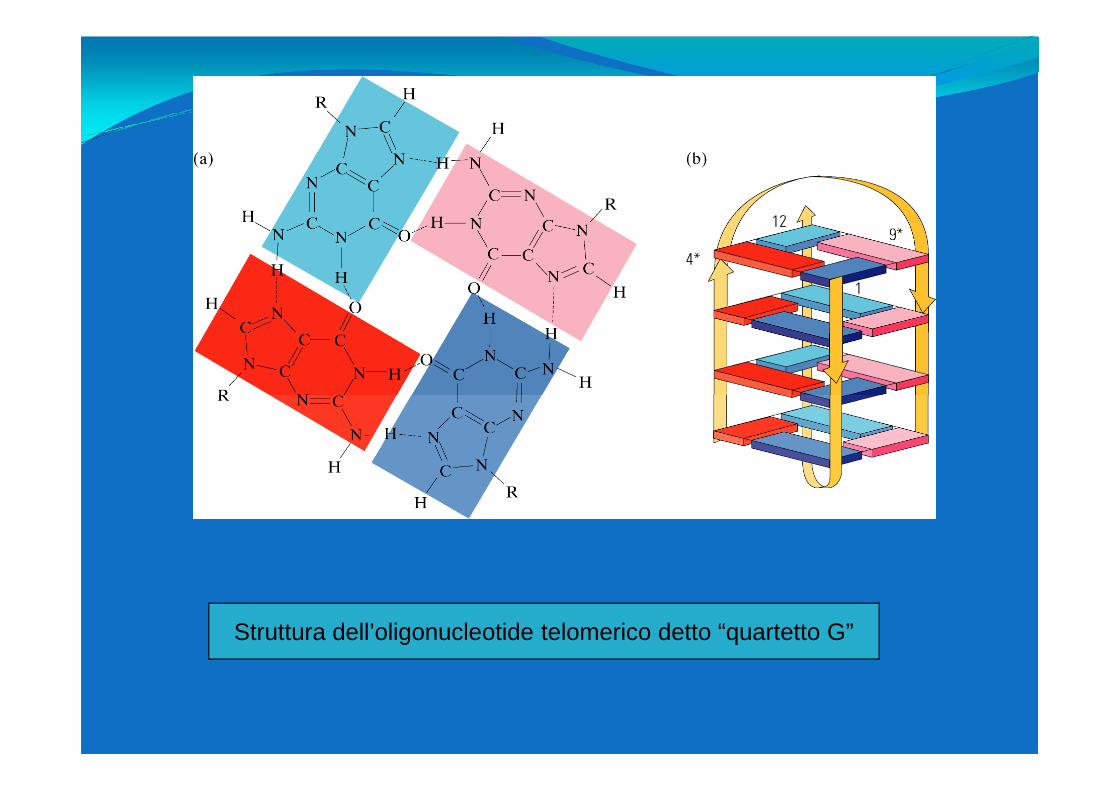
Struttura dell’oligonucleotide telomerico detto “quartetto G”




Discheratosi congenita: perdita della funzione della telomerasi

MALFUNZIONAMENTO NEI TELOMERI
La degradazione delle estremità 5’ non controllata provoca un aumento della estensione a singola elica in 3’ coperta da ssDNA binding proteins che può portare ad una esposizione delle sequenze telomeriche prima a doppia elica Una azione incontrollata della telomerasi può alterare le funzioni di capping a livello dei telomeri o sbilanciare la sintesi delle G.rich e C-rich strands, che possono portare all’accumulo di DNA in 3’

Controllo della lunghezza dei telomeriPOT:proteine di protezione dei telomeriTRF: fattori che legano le ripetizioni telomeriche




Farmaci antivirali
Molti antivirali interferiscono con gli enzimi coinvolti nel processovirale
Gli Inibitori della DNA polimerasi virale sono anche composti adattività antitumorale.
l'Ara-A o vidarabina è prevalentemente un antivirale, il suo analogoAra-C o citarabina è principalmente un farmaco antineoplastico;Ara-C o citarabina è principalmente un farmaco antineoplastico;
Anche la Timidilato Sintasi è bersaglio antivirale: questa può legare gliantimetaboliti, al posto dei metaboliti veri, incorporarli nel DNA eRNA virale con l'intervento di un apposito enzima provocando la suadistruzione, quindi eliminando l'infezione.
.

Nucleosidi purinici modificati allo zucchero(nucleosidi aciclici)
I nucleosidi aciclici nascono dall’esigenza di trovare inibitori della adenosinadeaminasi da somministrare assieme a Ara-A, per migliorarne l’effetto. Ilcomposto 9-(2-idrossietossimetil)adenina si rivelò essere un substrato perl’adenosina deaminasi. Il fatto che questo spezzone zuccherino potesseessere riconosciuto da un enzima fece ipotizzare che potesse esserericonosciuto anche da altri enzimi. Nacquero così gli aciclonucleosidi, il più
importante dei quali è l’aciclovir.importante dei quali è l’aciclovir.
Aciclovir : [9-(2'-idrossietossimetil)guanina], usato come tale o sottoformadi sale sodico. Agisce sulla DNA polimerasi virale. Struttura zuccherinaprofondamente modificata. L'aciclovir è un nucleoside ad ampio spettro e agrande selettività.

HN
NH
O
O
F
HN
N
F
O
OOHOH2C
HO
F
O
N
N
H3C
OH OH
HN CH3(CH2)4O
O
C
O
Fluorouracile Floxuridina Capecitabina
Formule di struttura di analoghi pirimidinici
OOHOH2C
HO
N
N
F
F
NH2
OOHOH2C
HO
HO
N
N
NH2
Fluorouracile Floxuridina Capecitabina
Citarabina Gemcitabina

Nucleosidi derivati dall’ adenina
N
N
NH2
N
N
HO H
H OH
OHO
HN
N
O
N
N
HO H
H OH
OOP
O
OH
HO
HO HO
VIDARABINA (AraA) Ara-Hx-MPUtilizzati contro Herpes simplex e varicella zosterAra A agisce con diversi meccanismi d’azione diminuisce resistenza ma aumenta tossicità.1) Come trifosfato inibisce DNA polimerasi2) Incorporata nel DNA (virus e ospite)3) Inibisce reazioni transmetilazione (metil transferasi)4) Inibisce dideossiadenosindifosfato da ADP (↓ ATP)

Nucleosidi pirimidinici modificati allo zucchero e alla base
N
N
NH2
OHO
I
O
HO H
H F
FIAC (5-iodo-1(2’-deossi-2’-fluoro-βD-arabinosofuranosil)citosina
Attivo contro HSV-1; HSV-2; citomegalovirus; Epstein Barr virus. Inibisce DNA polimerasi

Nucleosidi pirimidinici modificati allo zucchero
HN
N
O
H F
OHO
CH3
O
FMAU 1(2-deossi-2-fluoro-βD-arabinofuranosil)timina
Attivo contro HSV-1; HSV-2; citomegalovirus; Inibisce DNA polimerasi
HO H