kyc/teaching/files/543-05/2010... · web viewthis causes the her2 to become constitutively active;...
TRANSCRIPT

Chapter 26: Cancer and Growth ControlChapter 26: Cancer and Growth Control
By Lisa JablonskiBy Lisa Jablonski Chemistry 544 – Chemistry 544 – Chemical Biology Chemical Biology Professor K.Y. Chen Professor K.Y. Chen
May 3, 2010 May 3, 2010
Cancer results from abnormal proliferation (increase in number) of cells in the body. A tumor, also called a neoplasm, is any abnormal proliferation of cells which may be benign or malignant. A benign tumor stays confined to its original location; it neither invades neighboring normal tissue nor spreads to distant body sites. On the other hand, a malignant tumor is able to both invade neighboring normal tissue and spread throughout the body via the circulatory or lymphatic systems. The spreading of malignant tumors to distant body sites is called metastasis. Cancer refers specifically to malignant tumors or cells.1
Carcinomas are malignancies of epithelial cells.1 Lung, breast and colon are the most frequent carcinomas in the United States.2 Sarcomas are solid tumors of connective tissues, including muscle, bone, fat, cartilage and fibrous tissue.1,2 Leukemias are malignancies of blood-forming cells that grow in the bone marrow.1,2 Lymphomas are malignanies of immune cells that grow in the lymph nodes.1,2 Tumors are further classified according to the origin tissue and the type of cell involved.1
1
OutlineOutline What is cancer?What is cancer? What are the types of cancer?What are the types of cancer? How does cancer develop?How does cancer develop? What are the key properties of cancer?What are the key properties of cancer? How do cancer cells behave in culture?How do cancer cells behave in culture? Proto-oncogenes, oncogenes, tumor suppressor genes and DNA Proto-oncogenes, oncogenes, tumor suppressor genes and DNA
repair genesrepair genes Oncogenes: conversion from proto-oncogenes by gain-of-function Oncogenes: conversion from proto-oncogenes by gain-of-function
mutations; identification; activation by cancer-causing virusesmutations; identification; activation by cancer-causing viruses Tumor suppressor genes: deactivation by loss-of-function Tumor suppressor genes: deactivation by loss-of-function
mutations; identificationmutations; identification Examples of oncoproteinsExamples of oncoproteins Examples of deactivation of tumor suppressor genesExamples of deactivation of tumor suppressor genes Examples of defects in DNA repair genes that lead to cancerExamples of defects in DNA repair genes that lead to cancer Chromosomal abnormalities in cancerChromosomal abnormalities in cancer A brief look at cancer drugs that target oncoproteinsA brief look at cancer drugs that target oncoproteins ConclusionConclusion ReferencesReferences
What is cancer?What is cancer?
What are the types of cancer?What are the types of cancer?

Most cancers occur when a person is older, indicating that the development of cancer requires several mutations (Figure 1).4
Cancer begins with a mutation that provides a selective advantage for a cell, such as increased ability to proliferate or survive; this process is called tumor initiation. That cell will divide and create a population of clonally derived tumors cells; this process is called tumor clonality. Further mutations occur within the cells of the tumor population, leading to tumor progression. If one of the mutations provides a selective advantage, then cells with that mutation will more dominantly proliferate; this process is called clonal selection. Further mutations occur that lead to a more malignant and fast-growing tumor1. Figure 2 shows this process of cancer development. Transformation is the change that a normal cell undergoes as it becomes cancerous.5 Carcinogenesis is the process by which normal cells are transformed into cancer cells.5
2
Figure 2: Process of cancer development.1
Figure 1: Epidemiology of human cancers.4
How does cancer develop?How does cancer develop?

Throughout the cancer development process, mutations cause cancer cells to have increased loss of normal growth control.6
The development and metastasis of human colorectal cancer illustrates how multiple mutations lead to tumor progression and malignancy. The specific stages in tumor progression of colon cancer, and the mutations that cause each stage, were determined largely by Bert Vogelstein (Johns Hopkins University) and his colleagues. The order of the mutations, and not just their accumulation, determines tumor initiation and progression (Figure 4).7
Individuals with inherited mutations in the APC gene, a gene that is involved in limiting cell growth, have a hereditary predisposition for colon cancer.4 For example, familial adenomatous polyposis (FAP) is an inherited predisposition for colon cancer in which individuals form thousands of adenomas during their 20’s and 30’s.7 Individuals with FAP have an inherited defect in the APC gene. The adenomas are benign, but because there are so many of them, there is a high chance that one will become malignant.4 FAP is an example of a “gatekeeper gene defect;” 4 gatekeeper genes regulate growth and differentiation pathways.9 Hereditary non-polyposis colon cancer (HNPCC) is caused by defective mismatch repair genes. Patients with HNPCC form less adenomas, but these adenomas are more likely to
3
NormalEpithelium
Intermediate Adenoma
Lateadenoma
Metastases
Carcinoma
5qMutation or loss
APC gene
DNAhypomethylation
12pMutation
K-ras18qLossDDC17pLossp53
Other alterations
HyperproliferativeEpithelium
Earlyadenoma
Figure 4a (top left): Mutations and their clinical effects in colon cancer. Figure 4b (top right): A flow chart illustrating the same mutations and their clinical effects.4
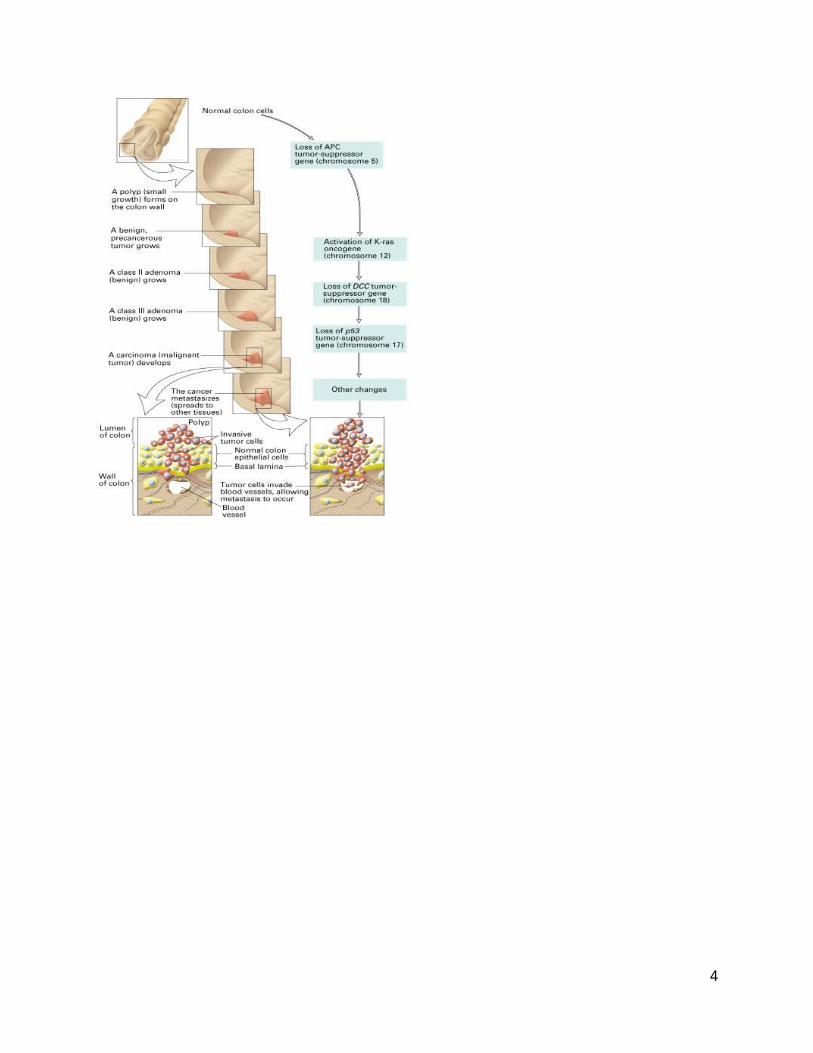
become cancerous because the defective mismatch repair genes cause increased mutations.4 HNPCC is an example of a “caretaker gene defect;”4 caretaker genes maintain genetic stability and minimize mutation rates.8 Juvenile polyposis syndrome (JPS) and ulcerative colitis (UC) are conditions in which stroma cells become abnormally proliferative, forming hamartomatous polyps. These polyps cause nearby epithelial cells to become cancerous due to an abnormal microenvironment.4 JPS and UC are examples of “landscaper gene defects;”4 landscaper genes encode proteins involved in the stromal environment.9 The risks of developing colon cancer are heightened by possessing gatekeeper, caretaker or landscaper defects (Figure 5).
In their paper “The hallmarks of cancer” published in 2000, Douglas Hanahan and Robert A. Weinberg describe the key acquired capabilities of cancer:10
1) Self-sufficiency in growth signals. Cancer cells have reduced dependency on extracellular growth factors. Why? Some cancer cells are able to produce their own growth factors, leading to autocrine growth stimulation (the continuous autostimulation of cell division).1 Some cancer cells overexpress growth factor receptors (such as tyrosine kinase receptors) or constitutively express different proteins in signal transduction pathways involved in cell growth and proliferation.1,10
2) Insensitivity to antigrowth signals. Cancer cells lack density-dependent inhibition, the process by which normal cells proliferate. In density-dependent inhibition, normal cells will duplicate until they reach a certain cell density. Anti-growth signals will tell the cells to stop proliferating and become quiescent (remain in G0 of cell cycle).1,10 By lacking density-dependent inhibition, cancer cells are insensitive to antigrowth signals and will continue to proliferate. Cancer cells also lack the ability to terminally differentiate.10 Most fully differentiated normal cells will either stop dividing or divide only rarely. Cancer cells fail to undergo terminal differentiation, leading to abnormal proliferation.1
4
Figure 5: Types of defects associated with colon cancer, compared to sporadic defects, and the probability of developing colon cancer.4
What are the key properties of cancer?What are the key properties of cancer?

3) Evading apoptosis (programmed cell death). Cancer cells can survive in environments that would trigger a normal cell to go through apoptosis.10
4) Limitless replicative potential. Cancer cells show telomere maintenance, allowing them to multiply without limit.11
5) Sustained angiogenesis (formation of new blood vessels). In normal adults angiogenesis is infrequent, occurring in women during their menstrual cycle, and in both men and women during wound healing.11 Cancer cells use angiogenesis to continue growing and spreading.12
6) Tissue invasion and metastasis. Cancer cells are less adhesive than normal cells, which helps them to invade other tissues and metastasize.1 Cancer cells also lack contact inhibition, the process by which normal cells migrate. Normal cells will move until they make contact with a neighboring cell. Once contact is made, further cell migration is inhibited, and cells will adhere to each other. Because cancer cells lack contact inhibition, they will move after contact is made with a neighboring cell and potentially move into surrounding normal tissue.1
Cancer cells also bypass normal checkpoints that prevent mutations, resulting in further downstream genetic instability.10
In his paper “What are the hallmarks of cancer?” published in 2010, ten years after Hanahan and Weinberg’s paper, Yuri Lazebnik points out that self-sufficiency in growth signals, insensitivity to antigrowth signals, evasion of apoptosis, angiogenesis and limitless replicative potential are features of both benign and malignant tumors. The property that uniquely defines malignant tumors is their ability to invade tissue and metastasize.12
Leukemias are an example of how the inability to fully differentiate leads to cancer. Normal red blood cells are derived from a common stem cell (a hematopoietic stem cell), and will follow a differentiation pathway through different progenitor cell types until they become fully differentiated.4 Once completely differentiated, the red blood cells will stop dividing.1 Leukemia occurs when a red blood cell does becomes “stuck” in a premature cell state where it can continue dividing.1 A mutation in a progenitor cell or a mutation in a signaling protein telling a progenitor cell to differentiate can lead to various types of leukemia (Figure 6).4
5
Figure 6: Red blood cell differentiation pathways. Red and pink are stem cells, green are progenitor cells.4

Normal cells, as described earlier, migrate according to contact inhibition and multiply following density-dependent inhibition. When transformed, however, they lose contact inhibition and density-dependent inhibition, and will grow and move on top of one another, forming 3-D clusters of cells called focuses.1,4 The cancer cells are less attached to each other than normal cells,4 and also appear rounder than normal because of their reduced attachment with neighboring cells1 (Figure 7).
Figure 7a (top left): Scanning EM of normal 3T3 (mouse fibroblast) cells. Figure 7b (top right): Scanning EM of 3T3 cells transformed by the Rous sarcoma virus.
We now know that the key properties of cancer, and that multiple mutations cause these changes to occur. What genes are mutated?
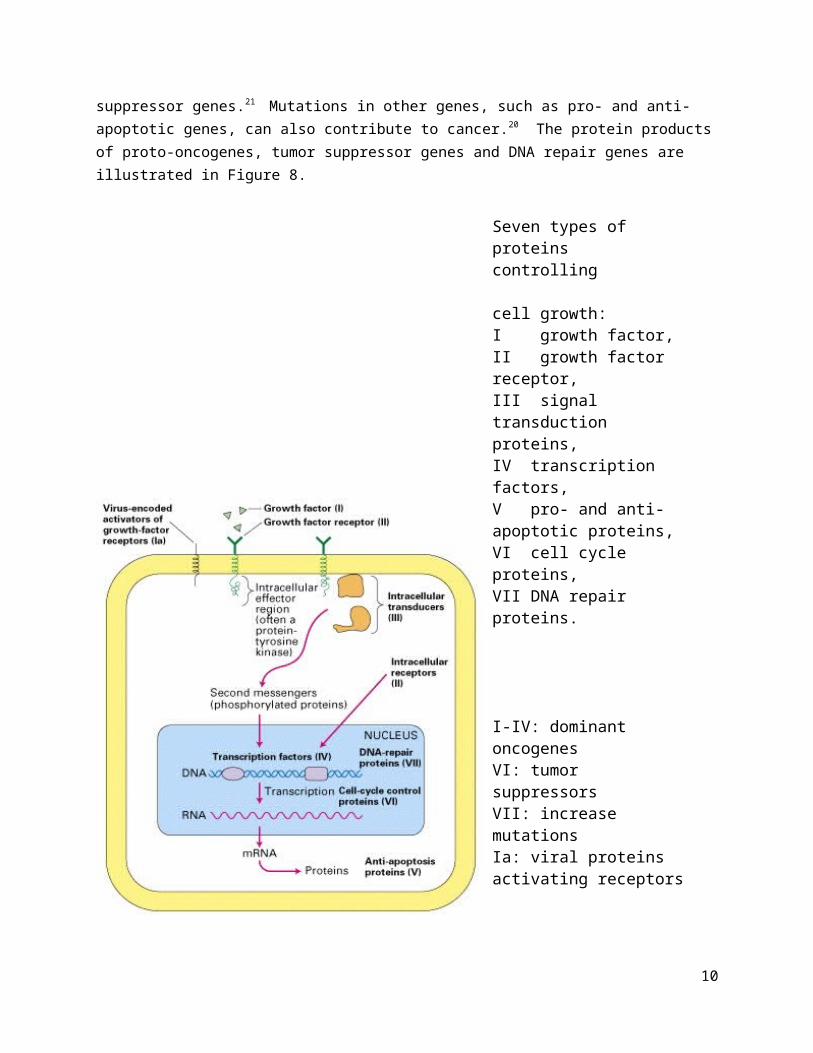
The main genes mutated in cancer are proto-oncogenes, tumor suppressor genes and DNA repair genes.14 Proto-oncogenes are genes that encode proteins that support cell growth and proliferation, such as growth factors, growth factor receptors, signaling proteins and transcription factors.4,15 Mutant, overactive forms of proto-oncogenes are called oncogenes;16 oncogenes encode for abnormal versions or quantities of growth-control proteins that cause the growth-signaling pathway to become overactive.17 Oncoproteins are the proteins encoded by an oncogene that cause transformation.21 Tumor suppressor genes are genes that encode proteins that limit cell growth and division, such as cell cycle regulatory proteins. When tumor suppressor genes are mutated, they are unable to stop cell growth, allowing the cell to grow and divide.18 DNA repair genes are genes that encode proteins that correct errors that occur during cell division, such as DNA mismatch repair genes.19 Mutations in DNA repair genes causes increased mutations that can activate proto-oncogenes into oncogenes or deactivate tumor suppressor genes.21 Mutations in other genes, such as pro- and anti-apoptotic genes, can also contribute to cancer.20 The protein products of proto-oncogenes, tumor suppressor genes and DNA repair genes are illustrated in Figure 8.
6
How do cancer cells behave in culture?How do cancer cells behave in culture?
Proto-oncogenes, oncogenes, tumor suppressor genes and DNA repair genesProto-oncogenes, oncogenes, tumor suppressor genes and DNA repair genes

Figure 8: Protein products of proto-oncogenes, tumor suppressor genes and DNA repair genes in controlling cell growth.21
Gain-of-function mutations convert proto-oncogenes into oncogenes.21 In “Molecular Cell Biology,” Lodish et al. describe three mechanisms by which proto-oncogenes are converted into oncogenes:
• “Point mutations in a proto-oncogene that result in a constitutively acting protein product.• Localized reduplication (gene amplification) of a DNA segment that includes a proto-
oncogene, leading to overexpression of the encoded protein.• Chromosomal translocation that brings a growth-regulatory gene under the control of
different promoter and that causes inappropriate expression of the gene.”21
For all three mechanisms, a mutation in only one of the two alleles for the gene can cause cancer.21
Oncogenes were first identified in cancer-causing retroviruses. Peyton Rous discovered the Rous sarcoma virus (RSV) in 1911. The Rous virus contains a gene, src, that is required for
7
Seven types of proteins controlling cell growth:I growth factor, II growth factor receptor, III signal transduction proteins,IV transcription factors, V pro- and anti-apoptotic proteins,VI cell cycle proteins, VII DNA repair proteins.
I-IV: dominant oncogenesVI: tumor suppressorsVII: increase mutationsIa: viral proteins activating receptors
Oncogenes: conversion from proto-oncogenes by gain-of-funtion mutations; identification; Oncogenes: conversion from proto-oncogenes by gain-of-funtion mutations; identification; activation by cancer-causing virusesactivation by cancer-causing viruses

cancer initiation but not for viral function.4 In 1977, through experiments with the Rous virus, J. Michael Bishop and Harold Varmus concluded that the src gene in the virus wasn’t an original viral gene, but was a cellular gene that the virus acquired during replication in the host cell.22 The normal cellular gene is called c-src and is the proto-oncogene; the viral gene, v-src, is the oncogene. 21
Unlike the Rous sarcoma virus, which is a fast-acting virus, most cancer-causing retroviruses are slow-acting and cause cancer only have months or years have passed. These slow-acting retroviruses do not carry oncogenes, but integrate their DNA into host genomes at locations that affect cellular proto-oncogenes.21 For example, a retrovirus can insert its genes upstream of c-myc, a cellular gene that encodes a transcription factor involved in cell growth. If there is a defect that prevents the right-hand viral LTR (long terminal repeats) from being transcribed, then the right-hand viral LTR can act as a promoter to turn on c-myc (Figure 9a). Another possibility is that the retrovirus inserts its genes upstream of c-myc, but in an opposite transcriptional direction; in this case the viral LTR can act as an enhancer of c-myc (Figure 9b).
Oncogenes can be identified by adding DNA from human cancer cells to a culture of mouse 3T3 cells (Figure 10). DNA from human cancer cells is added to a culture of mouse 3T3 cells. About one mouse cell in a million will become transformed and form a focus. DNA from the focus is isolated and transferred to another group of mouse 3T3 cells in order to isolate the oncogene. The DNA from the second cycle is cloned into phages. Most human genes have Alu sequences, which are repetitive DNA sequences, nearby. An Alu probe is added to the phages, and it hybridizes only with the phage that received human DNA. The phage with the DNA also contains the oncogene.21
8
Figure 9: Activation of the c-myc proto-oncogene due to integrated retrovirus DNA.

Loss-of-function mutations deactivate tumor suppressor genes.21 Loss-of-function mutations occur when a point mutation in a tumor suppressor gene prevents expression of the tumor suppressor protein product, or causes a nonfunctional protein product.21 A deletion of a DNA segment that includes a tumor suppressor gene will also lead to lack of expression of the tumor suppressor protein.4,21
For loss-of-function mutations, both alleles for the tumor suppressor gene must be mutated.21
Chromosomal missegregation or mitotic recombination can cause both chromosomes to have the mutant allele. This process is called loss of heterozygosity and leads to loss-of-function of the tumor suppressor gene (Figure 11).4
9
Figure 11: Chromosomal missegregation or mitotic recombination can cause a chromosome pair to become homozygous for mutant allele.4
Figure 10: The identification and cloning of an oncogene.21
Tumor suppressor genes: deactivation by loss-of-function mutations; identificationTumor suppressor genes: deactivation by loss-of-function mutations; identification
The first tumor suppressor gene to be identified was the Rb gene, identified in patients with inherited retinoblastoma. Individuals with this disease inherit a single mutant copy of the Rb gene, and develop retinal tumors when they are younger and usually in both eyes. Individuals are likely to develop tumors when they are younger and in both eyes because they already have one mutant allele present. Tumors occur when a somatic mutation occurs in the other Rb allele, causing loss-of-function of the gene. Figure 12 shows a pedigree of hereditary retinoblastoma.21 In sporadic retinoblastoma, individuals inherit two normal copies of the Rb gene; the copies eventually both become mutant through somatic mutations, leading to loss-of-function. Sporadic retinoblastoma is rarer than inherited retinoblastoma, occurs later in life and usually only occurs in one eye. 21
Many human tumors contain mutant Rb alleles arising from somatic mutations.4 Another tumor suppressor gene is BRCA1; women who inherit one mutant allele of BRCA1 have a much higher probability of developing breast cancer.21
Alfred Knudson studied inherited and sporadic retinoblastoma and determined that two mutation “hits” are required for loss-of-function of the Rb gene; his model of cancer causation by two mutations is called the Knudson two-hit hypothesis.23
Following are examples of oncoproteins that include virus-encoded activators of growth-factor receptors; growth factor receptors; signal induction proteins; and transcription factors. Overexpression of these oncoproteins can lead to cancer.
1) Virus-encoded activators of growth-factor receptors act as oncoproteins
Some viruses create proteins that can activate growth-factor receptors, causing overexpression of the growth-factor receptor. These viral proteins act as oncoproteins, since they behave like growth factors and cause overexpression of the growth-factor receptor.21
Spleen focus-forming virus (SFFV) is a retrovirus that disrupts the erythroid progenitor differentiation pathway. For normal erythroid progenitors to become differentiated red blood cells, the ligand Epo is needed to bind to the Epo receptor. SFFV encodes a mutant retrovirus envelope glycoprotein, gp55, that is able to bind to and activate Epo receptors, causing abnormal stimulation of proliferation of erythroid progenitors. This can lead to erythroleukemia in mice (Figure 13).21
10
Figure 12: Pedigree of hereditary retinoblastoma. Individuals in red inherited one mutant allele of Rb.21
Examples of oncoproteinsExamples of oncoproteins

The human papillomavirus (HPV) encodes a protein E5 that can bind to platelet-derived
growth factor (PDGF) receptors, causing the PDGF receptor to become activated; this can lead
to cervical cancer.21
2) Overexpression of growth-factor receptors
A point mutation coding for the transmembrane region of Her2, a growth-factor receptor, causes
Her2 receptors to dimerize and become activated even when not bound to a ligand. This
causes the Her2 to become constitutively active; the oncoprotein form of Her2 is called Neu
(Figure 14). The point mutation codes for glutamine instead of valine.21 Many human breast
cancers overexpress Her2.4 A point mutation for the epidermal growth factor (EGF) receptor
also causes overexpression. The mutation causes part of the ligand-binding domain of the EGF
receptor to be deleted, causing the EGF receptor to be constitutively activated (Figure 14).21
11
Figure 13: Virus-encoded glycoprotein gp55 acts as an oncoprotein.21
Figure 14: Point mutations causing growth-factor receptors Her2 and EGF to become overexpressed.21

A chromosomal translocation causes a different growth receptor, the Trk receptor, to become an
oncoprotein. Normal Trk receptors are activated by nerve growth factor (NGF). A chromosomal
translocation causes the extracellular domain of the Trk receptor to be fused with part of the
sequences encoding the tropomyosin protein; this causes dimerization of two Trk receptors
bound to tropomyosin and the activation of the receptors (Figure 15).21
3) Overexpression of signal induction proteins
Src is a signaling molecule with kinase activity that is involved in the cell growth and differentiation pathway26. Normal Src (c-Src) is inactivated when its tyrosine residue at position 527 is phosphorylated by another kinase, Csk. c-Src is activated when tyrosine 527 is dephosphorylated by a phosphatase enzyme. In an oncogenic form of Src, such as v-Src, tyrosine 527 is missing or altered; therefore, the oncogenic form of Src is constitutively active (Figure 16).21
12
Figure 15: Chromosomal translocation causing Trk to become an oncoprotein.21
Figure 16: Comparison of structure of c-Src and v-Src. v-Src is missing the region containing tyrosine 527, and is therefore constitutively active.21

4) Overexpression of transcription factors
The overexpression of transcription factors can also cause transformation.21 When quiescent fibroblasts are stimulated to proliferate by addition of serum containing PDGF, the c-Fos and c-Myc genes are activated (Figure 17).21 c-Fos and c-Myc are proto-oncogenes that code for c-Fos and c-Myc, transcription factors that activate genes that code for proteins involved in cell growth (specifically, proteins that cause cells to enter G1 and transition from G1 to S). Therefore, if c-Fos and c-Myc are overexpressed, the cells are stimulated to grow and replicate.21
In some cancers, multiple oncogenes are expressed that increase tumor formation (Figure 18).4
Following are examples of the deactivation of tumor suppressor genes involved in the cell cycle pathway, anti-growth signaling, and pro-apoptotic signaling.
1) Loss of activity of tumor suppressor proteins involved in the cell cycle pathway
Cells that enter the G1 phase of the cell cycle must past through a restriction point before they enter the S phase. Passage through the restriction point is regulated by cyclins, cyclin-
13
Figure 17: Proto-oncogenes activated by addition of PDGF in serum.21
Examples of deactivation of tumor suppressor genesExamples of deactivation of tumor suppressor genes
Figure 18: The effect of multiple oncogenes on tumor formation.4

dependent kinases (Cdks) and the Rb protein (Figure 18). If proteins that prevent the passage of the cell from G1 to S are mutated, this can lead to uncontrolled cell growth.21 For example, loss-of-function of Rb allows the release of E2F, a transcription factor that activates DNA synthesis genes. Loss-of-function of p16, a cyclin-kinase inhibitors, leads to overexpression of cyclin D, which causes hyperphosporylation of Rb and the relase of E2F. Both Rb and p16 are tumor suppressor proteins. Some viruses also create proteins that can bind to Rb and inhibit it (HPV - protein E7; SV40 virus – protein “T antigen”).21
2) Loss of activity of tumor suppressor proteins that act as anti-growth factors
Tumorderived growth factor β (TGFβ) is a growth factor that inhibits the growth of different cells. When TGFβ is deactivated, this contributes to the development of tumors. The binding of TGFβ to receptors leads to the activation of Smad proteins, which move to the nucleus and activate transcription of genes including p15. p15 functions to keep cells in G1 (Figure 19). Therefore, if TGFβ or Smad proteins are deactivated, p15 is no longer expressed and cells can move past G1. Most human pancreatic cancers contain deletions for the one of the Smad proteins, Smad4.21
14
Figure 18: Proteins involved in restriction point control. Mutations in the proteins highlighted in pink occur commonly in human cancers.21
Figure 19: The TGFβ pathway.21

3) Deactivation of tumor suppressor genes that promote pro-apoptotic functions
p53 is a protein that stops cells in G1 with damaged DNA from proceeding to the S phase. More than 50 percent of human cancers have mutations in the p53 gene. When DNA damage occurs, kinases phosphorylate p53 to stabilize it and therefore increase its concentration. p53 activates transcription of p21, a cyclin-kinase inhibitor, that is involved in arresting cells in G1. The increase in concentration of p53 also leads to the creation of proteins that cause apoptosis. Therefore, if p53 is mutated, p21 is not activated and the cells can move into the S phase; pro-apoptosis signals are also not activated, even if cells contain damaged DNA.21
When conditions inside the cell are normal, the protein MDM2 binds to p53 to deactivate it. Therefore if MDM2 is upregulated, this can lead to loss-of-function of p53. Similar to the case of Rb, some viruses also create proteins that can bind to p53 and inhibit it (HPV - protein E6, SV40 virus - protein “T antigen”). Certain carcinogens such as benzo(a)pyrene (in cigarette smoke) also cause mutations in p53.21
Defects in DNA repair genes, such as excision-repair and mismatch repair genes, can lead to cancer. Indiviudals with genetic defects in certain DNA repair genes have a larger chance of developing cancer; some of these genetic diseases and the types of cancer associated with them are listed in Figure 20.21
Tumor cells often have chromosomal abnormalities, such aneuploity (an abnormal number of chromosomes) and translocations (elements from different chromosomes joined together). For example, in Burkitt’s lymphoma, the c-myc gene is translocated from chromosome 8 to
15
Figure 20: Human hereditary diseases associated with DNA-repair defects.4
Examples of defects in DNA repair genes that lead to cancerExamples of defects in DNA repair genes that lead to cancer
Chromosomal abnormalities in cancerChromosomal abnormalities in cancer

chromosome 14. On chromosome 14 it is near an enhancer region and is therefore overexpressed (Figure 21).21
Cancer cells also duplicate a DNA sequence, causing multiple (up to as many as 100) copies of the DNA sequence to be present. When duplicated DNA sequences are lined up in the same region of a chromosome, they form a homogeneously staining region (HSR) (Figure 22a). The DNA duplicates can also exist as independent structures outside the chromosome; these structures are called double minute chromosomes (Figure 22b).21
Cancer is treated with chemotherapy, radiation therapy, surgery and other methods. Gene therapy chemotherapeutic drugs have been developed to target oncoproteins. Some examples are Trastuzumab (Herceptin®), which targets the Neu protein in breast cancer patients, and imatinib (Gleevec®), which targets the oncoprotein BCR-ABL in patients with chronic myeloid leukemia (CML).24 Certain cancer drugs also target biomarkers on tumor cell; depending on the types of biomarkers on a patient’s tumor cells, the drugs used to target those biomarkers can be personalized.25
Cancer is complex, involving both overexpression of genes and proteins that stimulate cell growth and underexpression of genes and proteins that limit cell growth. The loss-of-function of one tumor suppressor gene may not cause cancer if other pathways are still intact to make up
16
Figure 21: Chromosomal translocation in Burkitt’s lymphoma.21
Figure 22a: Homogeneously staining regions. Figure 22b: Double minute chromosomes.4
A brief look at cancer drugs that target oncoproteinsA brief look at cancer drugs that target oncoproteins
ConclusionConclusion

for the tumor suppressor gene’s loss-of-function. But, as multiple mutations accumulate, they increase the chance of developing cancer by disrupting more parts of the cell growth control pathway.
1 Cooper, Geoffrey M. The Cell: A Molecular Approach. Second Edition. Copyright 2000. Accessed from http://www.ncbi.nlm.nih.gov/bookshelf/.2 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide23 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide34 Lectures notes “Cancer and growth control.” Dr K.Y. Chen, Rutgers University, May 2010.5 http://www.cancer.gov/dictionary/
6 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide537 Lecture notes “Colon Cancer.” Dr. Brian Keith, University of Pennsylvania, November 2003.8 http://www.cancernetwork.com/display/article/10165/634589 Michor, F. et al (2004). “Dynamics of Cancer progression." Nature Reviews Cancer, 4:197-205.10 Hanahan, Douglas and Weinberg, Robert A (2000). “The hallmarks of cancer.” Cell, 100 (1): 57-70.11 http://www.cancer.gov/cancertopics/understandingcancer/angiogenesis/Slide512 http://www.cancer.gov/cancertopics/understandingcancer/angiogenesis/Slide813 Lazebnik, Yuri. “What are the hallmarks of cancer?” Nature Reviews Cancer, 10:232-233.14 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide5015 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide4316 Alberts, Bruce et al. Molecular Biology of The Cell. Fourth Edition. Copyright 2002.17 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide4418 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide4619 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide4820 http://www.cancer.gov/cancertopics/understandingcancer/cancer/Slide5021 Lodish, Harvey et al. Molecular Cell Biology. Fourth Edition. Copyright 2000. Accessed from http://www.ncbi.nlm.nih.gov/bookshelf/.22 http://nobelprize.org/nobel_prizes/medicine/laureates/1989/press.html23 Knudson, Alfred (1971). “Mutations and Cancer: Statistical Study of Retinoblastoma.” Proceedings of the National Academy of Sciences, 68 (4):820-823.24 http://www.cancer.org/docroot/ETO/content/ETO_1_4x_oncogenes_and_tumor_suppressor_genes.asp25 Thayer, Ann M (2010). “Biomarkers Help Hit The Bull’s-Eye.” Chemical & Engineering News, 88 (17):8.26 Martin, Steven G (2001). “The hunting of the Src.” Nature Reviews Molecular Cell Biology, 2:467-475.
17
ReferencesReferences