journal of invertebrate pathology - sistema de curriculum...
TRANSCRIPT

Journal of Invertebrate Pathology 109 (2012) 307–317
Contents lists available at SciVerse ScienceDirect
Journal of Invertebrate Pathology
journal homepage: www.elsevier .com/ locate/ j ip
Virulence of Vibrio harveyi responsible for the ‘‘Bright-red’’ Syndromein the Pacific white shrimp Litopenaeus vannamei
Sonia A. Soto-Rodriguez a,⇑, Bruno Gomez-Gil a, Rodolfo Lozano a, Rodolfo del Rio-Rodríguez b,Ana L. Diéguez c, Jesús L. Romalde c
a CIAD, A.C. Mazatlan Unit for Aquaculture and Environmental Management, A.P. 711 Mazatlan, Sinaloa 82010, Mexicob Instituto EPOMEX, Universidad Autonoma de Campeche, Av. Agustin Melgar s/n, Col. Buenavista 24039, Campeche, Mexicoc Universidad de Santiago de Compostela, Campus Sur s/n, 15782 Santiago de Compostela, Spain
a r t i c l e i n f o
Article history:Received 14 June 2011Accepted 11 January 2012Available online 28 January 2012
Keywords:‘‘Bright-red’’ SyndromeVibrio harveyiShrimpPathogenicityHistopathologyAntibioticsExtracellular products
0022-2011/$ - see front matter � 2012 Elsevier Inc. Adoi:10.1016/j.jip.2012.01.006
⇑ Corresponding author. Fax: +52 669 989 8701.E-mail address: [email protected] (S.A. Soto-Rodrigue
a b s t r a c t
Vibrio harveyi (Vh) CAIM 1792 strain was isolated from Litopenaeus vannamei affected with ‘‘Bright-red’’Syndrome (BRS). The strain grew in 1–10% NaCl, at 15–35 �C and was resistant to ampicillin (10 lg),carbenicillin (100 lg) and oxytetracycline (30 lg). The lowest MIC was for enrofloxacine (0.5 lg ml�1).The in vivo and in vitro toxicity of bacterial cells and the extracellular products (ECPs) of Vh CAIM1792 grown at 1.0%, 2.0% and 4.0% NaCl were evaluated. Adherence ability, enzymatic activities and sid-erophore production of bacterial cell was tested. The ECPs exhibited several enzymatic activities, such asgelatinase, amylase, lipase, phospholipase and caseinase. These ECPs displayed a strong cytotoxic effecton HELA cell line at 6 and 24 h. Challenges using 103 CFU g�1 caused opacity at the site of injectionand over 80% shrimp mortality before 24 h p.i. (post-injection). Mortality caused by the ECPs was higherthan mortalities with bacteria, especially in the first hours p.i. Bacteria were re-isolated from hemolymphsamples of moribund shrimp and identified as Vh CAIM 1792 by rep-PCR. Histological analysis of shrimpL. vannamei injected with Vh CAIM 1792 revealed generalized necrosis involving skeletal muscle (MU) atthe injection site, the lymphoid organ (LO), heart and connective tissues. Melanization within the MU atthe site of injection was also observed as well as hemocytic nodules within the hearth and MU at 168 hp.i. LO was the target organ of BRS. Necrosis of the MU at the injection site was the main difference incomparison to other shrimp vibriosis.
� 2012 Elsevier Inc. All rights reserved.
1. Introduction
Mexican shrimp aquaculture began developing in the 1980s, butit did not become a well-established industry until about 10 yearsago. Today aquaculture production exceeds the volumes obtainedfrom captures in Mexico, and in 2009 67.84% of shrimp productioncame from aquaculture, which is produced in 71,000 ha (SAGARPA,2009). Most of the Mexican shrimp farming occurs in the northwestpart of the country, where the Pacific white shrimp Litopenaeus van-namei is grown in semi-intensive earthen pond systems. Althoughthe yearly shrimp production has been showing a clear tendencyto increase, the industry has suffered serious economic losses dueto infectious diseases. More than 90% of the shrimp farms in Mexicohave been seriously affected in the last years by the White SpotSyndrome Virus (WSSV), infectious hypodermal and hematopoieticnecrosis virus (IHHNV), Taura syndrome virus (TSV) and bacterialnecrotizing hepatopancreatitis (NHP-B). Vibriosis is another impor-
ll rights reserved.
z).
tant disease affecting the shrimp industry in Mexico, causingrecurrent outbreaks (Soto-Rodriguez et al., 2010a). Some Vibriostrains have proved to be pathogenic to penaeid shrimp (Alapide-Tendencia and Dureza, 1997; Goarant et al., 2006; Jayasree et al.,2006; Soto-Rodriguez et al., 2003, 2010b; Rattanama et al., 2009;Lin et al., 2010).
Recently, Mexican farmers have experienced disease problemsin cultures of L. vannamei associated with signs of diseases notrelated to TSV, WSSV, IHHNV and NHP-B. The industry has sufferedserious economic losses due a new disease called Bright-red Syn-drome (BRS), it has been reported in farms in northwestern Mexicoand the causal agent was identified as a Vh CAIM1 1792 strain(Soto-Rodriguez et al., 2010b). Farmed shrimp affected with BRSbecome lethargic, anorexic, with flaccid body showing multifocalreddish discoloration spots on the abdominal cuticle and sometimesmelanized erosions around spots. Vibrio harveyi has been recognizedas a serious pathogen for a variety of important aquatic organisms inaquaculture around the world. To date, V. harveyi has been the most
1 CAIM is Collection of Aquatic Important Microorganisms.

308 S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317
virulent and prevalent pathogen of larval and grow-out shrimpculture (Lavilla-Pitogo et al., 1990; Liu et al., 1996; Soto-Rodriguezet al., 2010b).
Most studies on pathogenicity of V. harveyi are challenges usingdifferent sizes of fish or shrimp inoculated with a bacterial inocu-lum or extracellular products (ECPs) because they are consideredthe most important virulence determinants of some V. harveyistrains. These ECPs contain proteases, phospholipases, lipases,siderophores, chitinases and hemolysins (Liu et al., 1996; Zhangand Austin, 2000; Soto-Rodriguez et al., 2003; Zhong et al., 2006;Won and Park, 2008; Defoirdt et al., 2010). Vibriosis has beenexperimentally induced in penaeid shrimp by either immersingshrimp in water containing bacteria or by injecting bacteria intothe musculature (de la Peña et al., 1993; Esteve and Herrera,2000; Selvin and Lipton, 2003). These challenge studies reportmainly the mortality of test organisms and sometimes the poten-tial virulence factors of ECPs are evaluated. However, to date, theinfection process of shrimp vibriosis has been little studied dueto the difficulty in reproducing the disease (Roque et al., 1998;Saulnier et al., 2000). Histological techniques afford valuableknowledge regarding the state of health and the physiologicalprocesses of shrimp. More specific cytological and cytochemicalinformation may be obtained by special tissue staining methods.Therefore, the histological study of tissue preparations has becomeone of the routine diagnostic methods in aquaculture for infectionscaused by viruses, bacteria, and parasites. A few studies have eval-uated the histological damage caused by vibrios during experimen-tal infection in the shrimp tissues (Egusa et al., 1988; Esteve andHerrera, 2000).
This paper reports the phenotypic characteristic, antibiotic sen-sibility, adherence ability, cytotoxic and enzymatic activities, andsiderophore production of Vh CAIM 1792 and ECPs from Vh CAIM1792. Histopathology of L. vannamei juveniles shrimp challengedwith bacterial cells and ECPs from bacterial cells was alsoevaluated.
2. Materials and methods
2.1. Phenotypic characterization
Phenotypic identification of Vh CAIM 1792 followed the meth-ods as described by Alsina and Blanch (1994) and Holt et al.(1994), which were Gram stain, growth on thiosulphate citrate bilesalt sucrose agar (TCBS, Bioxon, Mexico), cell morphology, lumi-nescence, oxidase, sensitivity to the vibriostatic agent O/129,motility, O–F test, swarming on triptych soy agar (TSA, Bioxon,Mexico) + 2.0% NaCl, arginine dehydrolase, ornithine decarboxyl-ase and lysine decarboxylase. Growth at 0%, 1%, 2%, 3%, 4%, 5%,6%, 7%, 8%, 9% and 10% NaCl, growth at 4, 15, 25, 30, 35 and40 �C, indole production, gelatinase production, Voges–Proskauer,and utilization of citrate, L-arabinose and D-glucosaminic acid werealso tested. A further characterization was done with the API 20Esystem and different carbon sources were tested too with additionof NaCl to give a final concentration of 2.5%.Vh CAIM 1792 was al-ways incubated at 30 �C for 24–48 h.
2.2. Bacterial viability
Two bacterial viability tests were done. In the first test, Vh CAIM1792 was recovered from the cryovials and inoculated in 5.0 ml oftryptic soy broth (TSB, Bioxon, Mexico) + 2.0% NaCl incubated over-night at 30 �C. The bacterial suspension was obtained by inocula-tion of 100 ll in 10.0 ml of TSB (Bioxon, Mexico) + 2.0% NaCl(Bioxon �) and incubated overnight at 30 �C. Tubes with 5.0 ml ofpeptone broth (Dibico�) or 5.0 ml of TSB (Bioxon�) were adjusted
to salinities of 0.5%, 1.0%, 2.0%, 3.0%, 4.0%, 5.0%, 6.0%, 7.0%, 8.0%,9.0% and 10.0% NaCl, including tubes without NaCl with five repli-cates, for the first and second test respectively. Afterwards, alltubes were inoculated with 100 ll of the bacterial suspensionand incubated for 24 h at 30 �C. Serial dilutions were made withSSE and 100 ll of each dilution was inoculated on TCBS (Bioxon�)plates and incubated for 24 h at 30 �C and the absorbance wasmeasured at 600 nm for each tube.
2.3. Susceptibility to antibiotics
The strain Vh CAIM 1792 was analyzed for its susceptibility toantibiotics in vitro using a multidisc system to amikacin (30 lg),ampicillin (10 lg), carbenicillin (100 lg), cephalothin (30 lg), cef-otaxime (30 lg), ceftriaxone (30 lg), chloramphenicol (30 lg),gentamicin (10 lg), netilmicin (30 lg), nitrofurantoin (300 lg),pefloxacin (5 lg), trimethoprim-sulphamethoxazole (25 lg) (SanofiDiagnostics Pasteur, Mexico). For oxytetracycline (OTC, 30 lg)(Sigma–Aldrich Co., Mexico), norfloxacine (10 lg) (Schering-Plough, Mexico), erythromycin (15 lg) (Sigma–Aldrich Co.,Mexico), gentamicin (10 lg) Sigma–Aldrich Co., Mexico, chloram-phenicol (30 lg) (Sigma–Aldrich Co., Mexico) the disk diffusiontest (Bauer et al., 1966) was modified to allow the growth of VhCAIM 1792. The inoculum was brought to an optical density of1.0 at a wavelength of 610 nm, corresponding to a density of1 � 108 bacterial cells ml�1 according to the McFarland scale. Thestrain was suspended in sterile 2.5% NaCl solution and spread onMueller-Hinton agar (MHA; Difco) supplemented with 2.5% NaCl.A 6.0 mm sterile sensidisk (Oxoid, Basingstoke, England) wasinoculated with antibiotic according to Hindler (1992). All concen-trations refer to the concentration per disk. Vh CAIM 1792 wasinoculated in duplicate on the disk, and then the disks were placedon MHA. After incubation (24 h, 30 �C), the inhibition zones weremeasured in millimeters using a transparent ruler.
The minimum inhibitory concentrations (MIC) of enrofloxacine(Cheminova de Mexico, Mexico), fosfomycine (Sigma–Aldrich Co.,Mexico), OTC (Sigma–Aldrich Co., Mexico), flumequine (Sigma–Aldrich Co., Mexico), norfloxacine (Cheminova de Mexico, Mexico)and florfenicol (Schering-Plough, Mexico) to Vh CAIM 1792 wereestimated following the method of Hindler (1992), using mediasupplemented with NaCl at a final concentration of 2.5%.
2.4. Adherence ability
The cell surface hydrophobicity of Vh CAIM 1792 was deter-mined by bacterial adhesion to hydrocarbons (BATH) method ofRosenberg et al. (1980) using the n-octane (Sigma™). The percent-age of partitioning in the hydrocarbon phase was calculated asfollows:
PABH ð%Þ¼A600 original bacterial suspension�A600 aqueous phaseA600 original bacterial suspension
�100
Vh CAIM 1792 was inoculated on TSA previously adjusted 1.0%, 2.0%and 4.0% NaCl. The criteria of hydrophobicity proposed by Santoset al. (1990) were used to evaluate the hydrophobicity of each iso-late tested in this study. Interpretations for the BATH-test (>50%partitioning = strongly hydrophobic, 20–50% partitioning = moder-ately hydrophobic, and <20% partitioning = not hydrophobic).
A biofilms formation assay protocol was adapted from that ofWon and Park (2008). Polystyrene tubes containing 5.0 ml of TSBadjusted 1.0%, 2.0% and 4.0% NaCl were inoculated with overnightcultures of Vh CAIM 1792 and incubated for 48 h at 30 �C withoutagitation. At regular time intervals, the tubes were rinsed thor-oughly with distilled water, and a 1% solution of crystal violet

S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317 309
was added to stain the attached cells. After 15 min of incubation atroom temperature, the tubes were rinsed with distilled water, andthe biomass of attached cells (biofilm) was quantified by solubili-zation of the dye in 3.0 ml of 95% ethanol. The absorbance of thesolubilized dye was measured at 600 nm.
2.5. Enzymatic activities
Bacterial cultures that were grown overnight were spot-inocu-lated onto TSA with 1.0%, 2.0% and 4.0% NaCl that contained 1%gelatin (gelatinase test) or 2.5% skim milk (protease test) as sub-strate. The plates were incubated for 24 h to 48 h at 30 �C, andthe diameter of the lytic halo around each well was measured.Urease activity was examinated in urea broth (Dibico� Mexico)supplement with 1.0%, 2.0% and 4.0% NaCl. Tubes were incubatedfor 24 h at 30 �C. A color change in the medium was indicative ofa positive activity.
2.6. Siderophore production
The universal method of Schwyn and Neilands (1987) and fur-ther modified by Soto-Rodriguez et al. (2003) was employed. Briefly,Vh CAIM 1792 was grown in iron deficient MM9 broth for 48 h at30 �C under constant agitation at 95 rpm. Treatments with six rep-licates were plated directly onto chrome azurol S agar (CAS). Allmedia employed were supplemented with 1.0%, 2.0% and 4.0% NaCl.Incubation in CAS agar was at 30 �C for up to 72 h and orange halosaround colonies were measured at 48 and 72 h by measuring thediameter of the halo around each colony and subtracting from thecolony diameter to give the relative activity (mm). Siderophore pro-duction was considered positive when the halo diameter divided bythe colony diameter exceeded 1.3 (Amaro et al., 1990).
2.7. Bacterial inoculum
The strain Vh CAIM 1792 was previously isolated from hemo-lymph samples taken from shrimp that were affected by BRS(Soto-Rodriguez et al., 2010b). The strain was recovered from thecryovials and inoculated in 10 ml of tryptic soy broth (TSB has a0.5% NaCl, Bioxon, Mexico) supplement with 2.0% NaCl incubatedand plated on TSA (TSA has a 0.5% NaCl, Bioxon, Mexico) supple-ment with 2.0% NaCl and incubated overnight at 30 �C. Colonieswere suspended in sterile 2.5% NaCl and centrifuged at 5724g for10 min at 15 �C. The bacterial suspension was adjusted to an opti-cal density of 1.0 at 610 nm, similar to 0.5 MacFarlane Standard(equal to 108 colony forming units (CFU) ml�1, tested in previoustrials) (Soto-Rodriguez et al., 2003) and serially diluted to achievedensities from 103 to 107 CFU ml�1. These suspensions were platedonto TCBS (Bioxon, Mexico), after serially diluting to determine thereal density of Vh CAIM 1792 inoculated in the challenges.
2.8. ECPs extraction
Bacterial ECPs were obtained by the cellophane plate technique(Zhang and Austin, 2000) by spreading 3 ml of an overnight brothculture of Vh CAIM 1792 over sterilized cellophane sheets placedon TSA plates. TSA agar was previously adjusted to 1.0%, 2.0%and 4.0% NaCl. After incubation for 24 h at 30 �C, bacterial cellswere washed off the cellophane sheets with phosphate bufferedsaline (PBS) at pH 7.0. The bacterial suspensions were centrifugedat 13,000g for 30 min at 4 �C, and the respective supernatants werefiltered through 0.22 lm pore-size membrane filters (Millipore™).All the ECP samples were stored at �20 �C until used. Filtered prod-ucts were subjected to a sterility test by inoculation of 20 ll onTCBS agar (Dibico�) and incubation overnight at 30 �C. The proteinconcentration of the ECPs was determined by the method of
Bradford (1976) using bovine serum albumin (BSA; Sigma) as astandard. To evaluate the total proteolytic activity present in theECP samples, a multiprotein substrate (Azocoll; Sigma–Aldrich)was used following the manufacturer’s instructions. An absorbancereading of 1.0 at 520 nm, after a 30 min assay at 37 �C, was definedas one unit of proteolytic activity (pa).
2.9. Determination of enzymatic activities of ECPs
The enzymatic activities presented in the ECPs obtained fromVh CAIM 1792 cultured at 1.0%, 2.0% and 4.0% NaCl were evaluatedusing the API ZYM system (bioMérieux). Other enzymatic activi-ties, such as caseinase, gelatinase, elastase, amylase, phospholipa-ses, and lipase were determined in plates by a diffusion method.Plate dishes of basal nutrient agar (Difco) were adjusted to 1.0%,2.0% and 4.0% (NaCl, w/v) and supplemented with 1% (w/v) sodiumcaseinate (Difco), 1% (w/v) gelatin (Oxoid), 0.4% (w/v) starch (Dif-co), 2% (v/v) egg yolk emulsion (Sigma-Aldrich) and 1% (v/v)Tween-20/Tween-80 (Sigma–Aldrich), respectively. In all the tests,aliquots of 10 ll of each ECP were placed on the plates and incu-bated at 25 �C for 48 h. The appearance of clear halos around theECP spot indicates a positive result, except in lipase, where positivehalos are opaque.
2.10. Cytotoxic activity of the ECPs
Cytotoxicity assays with ECPs obtained from Vh CAIM 1792 cul-tured at 1.0%, 2.0% and 4.0% NaCl were basically carried out asdescribed by Toranzo et al. (1983). HELA cell lines (human cervicalcarcinoma) were employed. Confluent eukaryotic cells monolayerswere prepared in 96-wells plates and maintained at 37 �C. Beforethe test the medium was replaced from the wells with fresh med-ium. Medium was removed from wells containing eukaryotic cells,replaced with fresh medium and, inoculated directly with 50 lldifferent concentrations of ECPs. PBS (pH 7.4) being used as nega-tive control. The development of cytotoxic effects was observed at6 and 24 h post-inoculation.
2.11. Challenges with Vh CAIM 1792 and ECPs from Vh CAIM 1792
Two batches of juvenile shrimp (around 5 and 15 g) were trans-ported from a local farm to the laboratory and acclimatized for1 week in 400 l round tanks filled with filtered seawater, aeratedand held at 27–28 �C with a 400% of daily water exchange. The ani-mals were put in the experimental units and allowed to adapt foranother three days before trials began during which the healthstatus of the shrimp was evaluated through observation. Theexperimental units consisted of 60 l glass aquaria or 54 l fiberglassfilled with filtered (10 lm), UV sterilized and aerated seawater at28–30 �C with 12 h photoperiod. The bacterial water quality wasevaluated by plating a sample onto Marine Agar. Before the chal-lenge with bacteria, hemolymph samples were taken from ran-domly selected shrimp and the total heterotrophic bacteria andvibrios load were quantified on Marine Agar and TCBS respectively.
Five trials with shrimp using Vh CAIM 1792 cells and ECPs fromVh CAIM 1792 were done (Table 1). In all trials shrimp wereremoved from the acclimation tank, weighed, disinfected and in-jected in the third abdominal segment with 100 ll of each treat-ment or injected with 100 ll of sterile 2.5% NaCl for the controlgroup. The fifth challenge was done to evaluate the susceptibilityto bacteria of shrimp acclimated to different salinities, to bacteria.Shrimp were acclimated to 1.0%, 2.0% and 4.0% NaCl water culturebefore challenge with bacteria. Conditions were similar for all tri-als; experimental units were randomly allocated and held for168 h with a 100% of daily water exchange. Shrimp were fed adlibitum twice a day with a commercial diet (protein 35% Purina™).

Table 1Description of the conditions of trials with L. vannamei using Vh CAIM 1792 cells and ECPs from Vh CAIM 1792.
Trial Treatment Dose Weight of shrimp(g)
Replicates/no. ofshrimp
1 Vh CAIM 1792 cells 8.0 � 102, 8.0 � 103 or 8.0 � 104
CFU g�113.66, SD = 0.77 4/6
2 Vh CAIM 1792 cells 2.0 � 102 or 2.0 � 103 CFU g�1 14.90, SD = 0.82 3/103 Vh CAIM 1792 grown in TSA supplemented with 1.0%, 2.0% and 4.0%
NaCl.2.8 � 103 CFU g�1 6.12, SD = 0.77 4/10
4 ECPs from Vh CAIM 1792 cultured at 1.0%, 2.0% and 4.0% NaCl 100 ll per shrimp 4.60, SD = 0.76 3/105 Vh CAIM 1792 cells 2.73 � 103 CFU g�1 7.3, SD = 1.10 3/10
310 S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317
Temperature, nitrates, total ammonia, salinity, and pH were re-corded daily. A record of mortality and moribund organisms foreach aquarium was kept.
2.12. Re-isolation of bacteria and molecular identification
Hemolymph samples from moribund organisms within a periodof 24 h p.i. were inoculated on TCBS agar and incubated at 30 �C for24 h. The shrimp were preserved immediately in Davidson’s fixa-tive for 48 h. Dominant yellow colonies were DNA fingerprintedby rep-PCR (Cabanillas-Beltran et al., 2006). Fixed samples wereprocessed for histological analysis, following standard procedures(Bell and Lightner, 1988). Tissue sections were prepared for hema-toxylin and eosin (H&E) and Gram-Humberstone methods andanalyzed under optical microscopy (Olympus CX31).
3. Results
3.1. Phenotypic characterization and bacterial viability of Vh CAIM1792
Cells were Gram-negative rods that were motile when theywere grown in liquid media. The cells did not swarm on TSA. Col-onies on marine agar (Difco Laboratories, Detroit, Mich.) weresmooth, circular, and off-white and the strain was not luminescent.Colonies on TCBS agar (Bioxon, Mexico) were yellow (sucrose fer-menting), circular with undulate borders and granular surface.
The strain Vh CAIM 1792 was facultatively anaerobic, fer-mented glucose, Kovacs oxidase positive, and was sensitive tothe vibriostatic agent O/129 at 150 mg, Voges–Proskauer and gela-tinase positive, arginine dehydrolase negative, lysine decarboxyl-ase and ornithine decarboxylase positive (Table 2). Thesecharacteristics placed it within the genus Vibrio (Alsina and Blanch,1994). Sodium chloride was required for growth in a wide range;the only salinity at which Vh CAIM 1792 was not able to growwas at 0% NaCl (Fig. 1).
3.2. Susceptibility to antibiotics
The largest inhibition zone (30.0 mm) occurred with chloram-phenicol at 30 lg and the smallest with OTC at 30 lg (7 mm,Table 3). Vh CAIM 1792 was sensitive to most of the tested antibiot-ics and was resistant only to ampicillin (10 lg) and carbenicillin(100 lg). MICs were determined for the antibiotics commonly usedin the Mexican shrimp culture; enrofloxacine MIC 0.50 lg ml�1, OTC2.0 lg ml�1, norfloxacine and florfenicol 4.0 lg ml�1. In additionMICs for flumequine was 2.0 lg ml�1 and fosfomycine >512lg ml�1.
3.3. Adherence ability, enzymatic activities and siderophoreproduction of Vh CAIM 1792
Cell surface hydrophobicity was examined using BATH test(Table 4). Vh CAIM 1792 strain grown at 1.0%, 2.0% and 4.0% NaCl
exhibited strong hydrophobicity using n-octane as a solvent. Thebiofilm formation assay produced absorbance in the range of0.99–1.30 after 48 h. Both tests were not statistically different(p > 0.05) among three salinities. Only a weak caseinase activitywas observed for 1.0 and 2.0% NaCl conditions; meanwhile sidero-phore production was observed in Vh CAIM 1972 grown at threeNaCl concentrations.
3.4. Characterization of ECPs
The enzyme activity, proteolytic activity, and cytotoxic activi-ties of Vh CAIM 1792 ECPs are shown in Table 5. The highestenzyme activity from API ZYM was observed in the ECPS of VhCAIM 1792 grown at 2.0% NaCl. Hydrolytic activity of the othertested enzymes was weak without differences among three treat-ments, only gelatinase, amylase and lipase (tween 20) activitieswere detected in all salinities. ECPs obtained from Vh CAIM 1792grown at 1.0% and 2.0% NaCl had stronger cytotoxic activity. Theobserved morphological changes consisted mainly of vacuoliza-tion, elongation, cellular clumps, detachment and finally mono-layer destruction.
3.5. Challenges with bacterial cells (trial 1 and 2)
The water quality of the experimental units registered was:temperature = 27–29 �C, salinity = 34 g l�1, pH = 8.1 ± 0.2, totalammonium 60.4 mg l�1, nitrites 6 0.2 mg l�1. No vibrios were reg-istered in the water intake nor in the hemolymph samples beforethe challenges. The gross signs of challenged shrimp were lethargy,erratic swimming (whirling movement), anorexia, shell discolor-ation and flaccid body at first hours p.i.
The mortality of the shrimp challenged with 103 CFU g�1 (Fig. 2)was almost 100% before 24 h, with the shrimp showing opacity atsite of injection.
Bacterial density in hemolymph of moribund shrimp before24 h p.i. was 4.80 � 103 for shrimp challenged with 104 CFU g�1.All colonies in TCBS were yellow. At the end of the trial no vibrioswere found in hemolymph samples from the control and chal-lenged groups. In the second trial with Vh CAIM 1792 cells, cumu-lative mortalities were lower than in the first trial with Vh CAIM1792 cells (Table A1 in Appendix), however we used lower doses.
3.6. Challenges with Vh CAIM 1792 grown at different salinities (trial3) and ECPs from Vh CAIM 1792 (trial 4)
Challenges were done to evaluate the pathogenicity of Vh CAIM1792 and ECPs from Vh CAIM 1792 grown at 1.0%, 2.0% and 4.0%NaCl. A wide variability in mortality with bacteria challenge com-pared with ECPs was observed. Mortalities caused by the ECPs inthe first hours were higher than mortalities with bacteria (Fig. 3).Treatment of 4.0% NaCl was more pathogenic before 24 h p.i. forboth bacteria and ECPs with mortalities of 70% and 100% respec-tively. After 48 h p.i., no statistically differences among three salin-ities (p > 0.05) were observed in challenge with bacteria (see

Table 2Morphological and physiological characteristics of VhCAIM 1792 isolated from L. vannamei with ‘‘Bright-red’’Syndrome. +: positive; �: negative; Y: yellow colonies.
Characteristic CAIM 1792
Gram staining –Cell morphology RodMotility +Swarming on TSA (2% NaCl) –Pigment –Growth on TCBS YLuminescence –Sensitivity to vibriostatic 0/129 +Oxidase +Arginine dehydrolase –Ornithine decarboxilase +Lysine decarboxilase +O–F +
Production ofH2S –Nitrate reduction +Indole –Methyl red +Gas from D-glucose –Voges–Proskauer +b-Galactosidase (OPNG test) –Tryptophan deaminase –Gelatinase +Urease –Triple sugar iron (TSI) +
Temperature tolerance4 �C –15 �C +25 �C +30 �C +35 �C +40 �C –
Utilization ofAcetate +++
L-Alanine ++
Amygdalin +c-Aminobutyrate ++
L-Arabinose –
L-Arginine +
L-Aspartate ++
Cellobiose –Citrate ++Fructose +
D-Galactose –
D-Galacturonate –
D-Gluconate –
D-Glucose +
D-Glucosamine –
D-Glucuronate +
L-Glutamate +++
Glutarate +
L-Glutamine +++
Glycerol –Glycine ++
L-Histidine –
p-Hydroxybenzoate +b-Hydroxybutirate ++myo-Inositol –a-Ketoglutarate +++
DL-Lactate +++
Lactose –
L-Leucine –
L-Lisina +
DL-Malate –
D-Mannitol +
D-Mannose +
Melibiose –Methanol ++
Table 2 (continued)
Characteristic CAIM 1792
L-Ornithine –
Propionate –Putresine +Pyruvate +++
L-Rhamnose –
D-Ribose +
Salicin –
D-Sorbitol –
Sucrose +Succinate +++Trehalose ++
L-Threonine +
D-Xylose –
S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317 311
Table A2 in Appendix). In contrast, challenge with ECPs obtained at1.0% NaCl was statistically different compared with 2.0% and 4.0%NaCl.
In the fifth trial, shrimp were acclimated to different watersalinity conditions; the highest mortality was observed at 4.0%and 1.0% (see Table A3 in Appendix).
Before the challenges with bacterial cells, bacteria were re-iso-lated from hemolymph samples of moribund shrimp and finger-printed by rep-PCR (Cabanillas-Beltran et al., 2006) showing thesame electrophoretic band pattern corresponding to Vh CAIM1792.
3.7. Histopathological analysis (trial 1 and 2)
In the first trial (168 h of exposure) Vh CAIM 1792 caused a gen-eral necrosis in the lymphoid organ (LO), striated muscle, heart,antennal gland and gills of L. vannamei with the LO as the main af-fected organ. Around the site of injection, the bacteria caused a se-vere necrosis of the muscle fibers with melanin deposits after168 h p.i. (Fig. 4a and b). During the first hours p.i. the hemocytesshowed a tendency to agglutinate within the heart chambers,probably as a first response to the infection (Fig. 4c); at 168 h hem-ocytic nodules could be observed within the heart muscle (Fig. 4d).LO tubules gradually lost their structure during the first hours p.i.(Fig. 5a), associated with the presence of Gram-negative bacteriaclumps dispersed in the stromal matrix (Fig. 5d). A week later,the LO was severely necrotic and bacterial clumps could still be ob-served within the tubules (Fig. 5c). The antennal gland epitheliumand gill lamellae were necrotic in the first hours p.i. with Vh CAIM1792 (Fig. 6a–c); meanwhile after 168 h p.i., formation of hemocy-tic nodules was evident (Fig. 6d). Histopathological observationswere similar in the second trial.
Histopathological analysis of L. vannamei challenged with V.harveyi CAIM 1792 (trial 3) and ECPs from Vh CAIM 1792 (trial 4)at different salinities.
No significant histological differences were observed between L.vannamei challenged with Vh CAIM 1792 or ECPs grown 1.0%, 2.0%and 4.0% NaCl. Damage was mainly observed at the site of injec-tion, LO, heart and connective tissue (Fig. 7). From 9 to 13 h p.i., se-vere necrosis of the skeletal muscle (MU) with bacterial masseswas observed. At 48 h p.i. there was a decrease of bacterial masseswithin MU, showing melanizated hemocytic nodules and hemocy-tic infiltration.
As infection progressed, hemocytic infiltration and formation ofmultifocal hemocytic nodules was observed (Fig. 7a) culminatingwith the formation of granulomas within the MU (Fig. 7c). ECPscaused severe acute necrosis at the site of injection site, without

A
B
Fig. 1. Bacterial viability of Vh CAIM 1792 grown in (A) peptone broth and (B) TSBat different salinity conditions. Bars indicate standard deviation.
Table 4Cell surface hydrophobicity, biofilm formation, hydrolytic activities, and siderophore prodUre: urease; Sid: siderophores.
NaCl(%)
% Part. BATH(S.D.)
Biofilm Abs600
(S.D.)Hydro
Cas (4
1.0 93.9 (3.1)a 0.99 (0.06)a 1.7 (w2.0 84.6 (10.7)a 1.30 (0.15)a 1.5 (w4.0 93.0 (3.3)a 1.00 (0.13)a –
BATH: bacterial adhesion to hydrocarbons test: >50% partitioning = strong hydrophobic+ = positive activity.
a Treatments with the same letter are not statistically different (p > 0.05, for BATH an
Table 3Sensitivity of Vh CAIM 1792 to antimicrobial agents.
Antibiotic Concentration(lg)
Halo(mm)
Sensitivity*
Chloranphenicol 30 30 SCeftriaxone 30 28 SCefotaxime 30 27 STrimethoprim-
sulphamethoxazole25 26 S
Pefloxacin 5 23 SNorfloxacine 10 21.5 n.d.Netilmicin 30 21 SNitrofurantoin 300 20 SErythromycin 15 18.5 SGentamicin 10 18 SAmikacin 30 17 SCephalothin 30 16 IOxytetracycline 30 7 n.d.Ampicillin 10 0 RCarbenicillin 100 0 R
* The sensitivity criterion used was taken from human or veterinary medicine,since there are no criteria defined for shrimp bacterial pathogens (Giono Cerezo,1983; Lynch and Raphael, 1983; Barry and Thornsberry, 1985). S: sensitive, R:resistant; n.d. no data.
Table 5Characterization of the ECPs of Vh CAIM 1792 grown at three salinities.
NaCl (%)
1.0 2.0 4.0
API ZYMAlkaline phosphatase +++ +++ ++Esterase – ++ +Esterase lipase – ++ –Lipase – – –Leucine arylamidase – + –Valine arylamidase – – –Cystine arylamidase – – –Trypsin ++ +++ ++a-chymotrypsin – + ++Acid phosphatase +++ +++ +++Naphthol-AS-BI-phosphohydrolase ++ ++ –a-galactosidase – – –b-galactosidase – – –b-glucuronidase – – –a-glucosidase – – –b-glucosidase – – –N-Acetyl-b-glucosaminidase + + –a-mannosidase – – –a-fucosidase – – –Gelatinase +(17 mm) +(15 mm) +(14 mm)Amylase +(7 mm) +(7 mm) +(5 mm)Esculin – – –Lipase Tween 80 +(5 mm) +(9 mm) –Lipase Tween 20 +(5 mm) +(9 mm) +(5 mm)Caseinase +(10 mm) – –DNase – – –Elastase – – –Urease – – –Total protein (lg ml�1) 90 119 27.5Proteolytic activity (ua ml�1) 4.72 2.70 2.49Cytotoxic activity (HELA cells) 6/24 hCrude ECPs ++/++ ++/++ ++/++1:2 Dilution ++/++ ++/++ +/+1:5 Dilution +/+ (+)/(+) �/�
312 S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317
hemocytic infiltration (Fig. 7b). Accumulation of hemocytic nod-ules, bacterial masses and melanized hemocytic nodules were alsofound within the LO and the heart which are lesions typically ob-served as consequence of systemic vibriosis in these animals (seeFig. A1a and b in Appendix).
4. Discussion
Variable phenotypic results, e.g., for arginine dihydrolase, ni-trate reduction, Voges–Proskauer, indol, cellobiose, citrate, sucrose,lactose, luminescence of V. harveyi strains isolates from shrimp,lobster and coral, have been reported (Karunasagar et al., 1996;Diggles et al., 2000; Stabili et al., 2006; Najiah et al., 2008). For in-stance, Vibrio campbellii, V. harveyi and Vibrio rotiferianus havenearly indistinguishable phenotypes (Gomez-Gil et al., 2003,2004). Due to the economic importance of V. harveyi infections,
uction of Vh CAIM 1792 grown at different salinities. Cas: caseianse; Gel: gelatinase;
lytic activities (diam., mm)
8 h) Gel (48 h) Ure (24 h) Sid (72 h)
) – – +) – – +
– – +
ity. Hydrolytic activities: w = weak; – = negative activity. Siderophore production:
d biofilms test n = 3; for hydrolytic activities and siderophores, n = 6).

Fig. 2. Cumulative mortality in the first trial of L. vannamei challenged with threedoses of Vh CAIM 1792 during 168 h. Contl: control group. Bars indicate standarddeviation.
A
B
Fig. 3. Cumulative mortality of L. vannamei challenged with 2.8 � 103 CFU g�1 of VhCAIM 1792 grown at 1.0%, 2.0% and 4.0% NaCl. Live cells (A) and ECPs (B). Contl:control group. Bars indicate standard deviation.
S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317 313
there is considerable interest in methods not only to phenotypingV. harveyi-related populations associated with marine rearedanimals; it is also important to study the mechanisms of virulence.Phenotypic differences were also registered in NaCl tolerance,where Vh CAIM 1792 grew from 1% to 10% NaCl. More than 80%of V. harveyi isolates from Penaeus monodon grew only at 0% and5% NaCl (Najiah et al., 2008). This viability in a wide range of salin-ity means that Vh CAIM 1792 could proliferate in extreme environ-mental conditions that occur during the shrimp farming season inMexico, which include brackish water during the rainy season andhyper saline water during the dry season.
Regarding the sensibility tests, Vh CAIM 1792 was resistant toampicillin and carbenicillin, the latter used by farmers in Thailandto combat shrimp vibriosis (Nakayama et al., 2006). This resultcoincides with other works where 70–100% of V. harveyi strainsisolated from farmed shrimp were resistant to both antibiotics(Baticados et al., 1990; Abraham et al., 1997; Roque et al., 2001;
Najiah et al., 2008). The resistance against b-lactam antibioticsmight be due to the natural occurrence of b-lactamases in V. har-veyi strains (Teo et al., 2002). This work found that Vh CAIM1792 was also resistant to OTC such as most of V. harveyi strainsisolated from diseased shrimp (Baticados et al., 1990; Chanratcha-kool et al., 1995; Soto-Rodriguez et al., 2006; Manilal et al., 2010).The MIC for OTC against Vibrio sp. isolated from diseased shrimpwas the highest reported (Soto-Rodriguez et al., 2008). Even so,currently, OTC is amongst the most commonly used antibiotic byshrimp farmers worldwide, including Mexico (Soto-Rodriguezet al., 2010a). Interestingly, Vh CAIM 1792 was found to be sensi-tive to most of the antibiotics tested; in contrast, other works havefound multiple resistances of V. harveyi strains to antibiotics(Tendencia and de la Pena, 2001). In this work, the lowest MIC va-lue was for enrofloxacine, currently one of the most used antibiot-ics in Mexico (Soto-Rodriguez et al., 2010a).
Bacterial adhesion to host surfaces has been described as one ofthe initial steps in microbial pathogenesis. It has been suggestedthat hydrophobicity and biofilm formation are the determiningfactors in the adhesive process and in the survival of pathogensin cells (Daly and Stevenson, 1987). Vh CAIM 1792 grown at threesalinities exhibited strong hydrophobicity in the BATH assay. Bio-film formation is known as one of the virulent factors of V. harveyi(Karunasagar et al., 1996; Won and Park, 2008). We observed thatbiofilm formation of Vh CAIM 1792 grown at three salinities werenot statistically different (p > 0.05). Other virulent factors associ-ated with the pathogenicity of this bacterium were identified bySoto-Rodriguez et al. (2003) and Owens et al. (1996), who demon-strated the role of siderophore production in V. harveyi virulence.We found siderophore production by Vh CAIM 1792 grown at threesalinities, but they did not show differences among treatments. Theproduction of extracellular enzymes by bacterial fish pathogenshas been widely observed (Amaro et al., 1992; Alcaide et al.,1999); however, the role of these products in pathogenesis is stillnot clear. ECPs are considered to be one of the most importantdeterminants of virulence in V. harveyi (Austin and Zhang, 2006;Won and Park, 2008; Manilal et al., 2010). In our results, ECPs ofVh CAIM 1792 grown at 2.0% NaCl exhibited the highest enzymeactivity compared with ECPs obtained from 1.0% and 4.0% NaClby API ZYM test. Only gelatinase, amylase and lipase activitieswere detected in the three salinities, the hydrolytic activity ofthe other enzymes was weak without differences among treat-ments. Several studies consider the hemolytic activity as a majorvirulence factor for some V. harveyi strains (Zhong et al., 2006;Haldar et al., 2010; Rattanama et al., 2009) but Vh CAIM 1792 doesnot have it (data not shown). There was no correlation betweenArtemia mortality and hemolytic activity of V. harveyi strains(Soto-Rodriguez et al., 2003). However, we found serine and Znproteases in the Vh CAIM 1792 genome by analysis in silico (datanot shown) which coincide with Won and Park (2008) who re-vealed that the major protease of V. harveyi ECPs isolated frommarine fishes was serine protease, meanwhile Liu and Lee (1999)and Liu et al. (1997) considered a cysteine protease as the majorexotoxin. Based on these observations, we suggest that the majorprotease of V. harveyi ECPs is strain-dependent. Tissue cultureshave played a crucial role in investigating bacterial–host interac-tions due to the easy manipulation of cells and their maintenanceunder controlled conditions. Ideally, we should use crustacean celllines to test cytotoxic activities of pathogenic vibrios, but to datewe have not that methodology. In the present study, severe dam-age to HELA cell line for 6 and 24 h was shown in ECPs samplesobtained from three salinities, although ECPs from Vh CAIM 1792grown at 1.0% and 2.0% NaCl had stronger cytotoxic activity. Mon-tero and Austin (1999) revealed that V. harveyi isolated from in-fected shrimp, L. vannamei exhibited a strong cytotoxic activity1 h after inoculation.
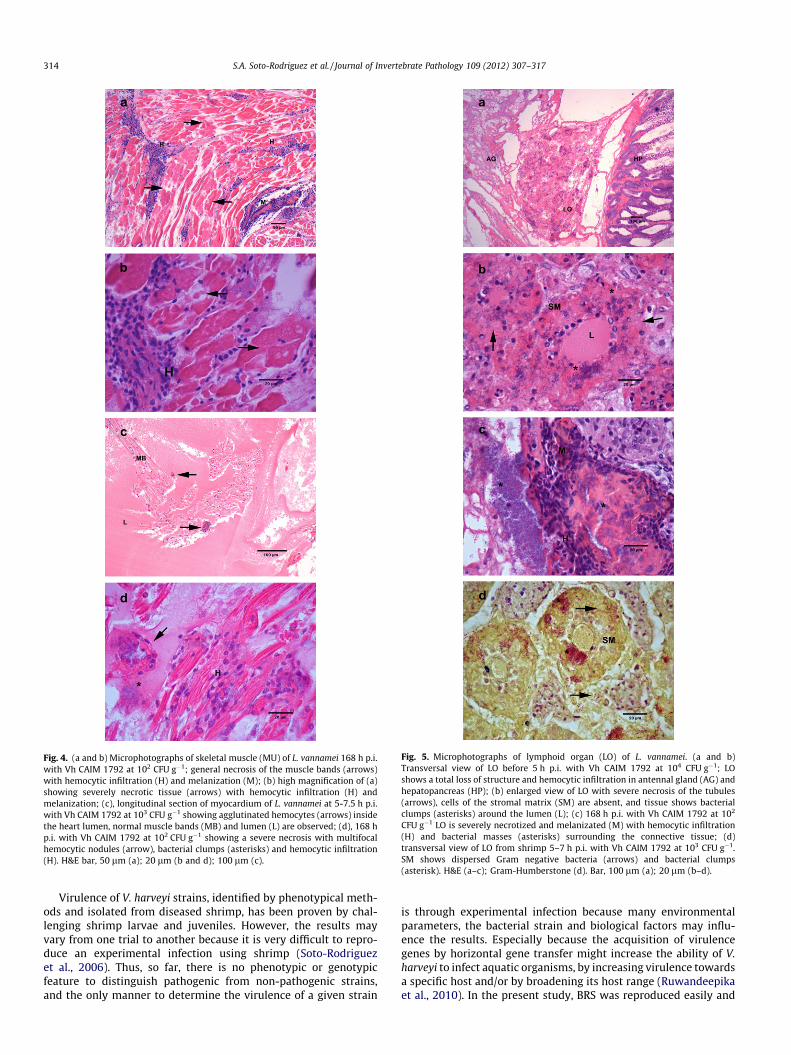
Fig. 4. (a and b) Microphotographs of skeletal muscle (MU) of L. vannamei 168 h p.i.with Vh CAIM 1792 at 102 CFU g�1; general necrosis of the muscle bands (arrows)with hemocytic infiltration (H) and melanization (M); (b) high magnification of (a)showing severely necrotic tissue (arrows) with hemocytic infiltration (H) andmelanization; (c), longitudinal section of myocardium of L. vannamei at 5-7.5 h p.i.with Vh CAIM 1792 at 103 CFU g�1 showing agglutinated hemocytes (arrows) insidethe heart lumen, normal muscle bands (MB) and lumen (L) are observed; (d), 168 hp.i. with Vh CAIM 1792 at 102 CFU g�1 showing a severe necrosis with multifocalhemocytic nodules (arrow), bacterial clumps (asterisks) and hemocytic infiltration(H). H&E bar, 50 lm (a); 20 lm (b and d); 100 lm (c).
Fig. 5. Microphotographs of lymphoid organ (LO) of L. vannamei. (a and b)Transversal view of LO before 5 h p.i. with Vh CAIM 1792 at 104 CFU g�1; LOshows a total loss of structure and hemocytic infiltration in antennal gland (AG) andhepatopancreas (HP); (b) enlarged view of LO with severe necrosis of the tubules(arrows), cells of the stromal matrix (SM) are absent, and tissue shows bacterialclumps (asterisks) around the lumen (L); (c) 168 h p.i. with Vh CAIM 1792 at 102
CFU g�1 LO is severely necrotized and melanizated (M) with hemocytic infiltration(H) and bacterial masses (asterisks) surrounding the connective tissue; (d)transversal view of LO from shrimp 5–7 h p.i. with Vh CAIM 1792 at 103 CFU g�1.SM shows dispersed Gram negative bacteria (arrows) and bacterial clumps(asterisk). H&E (a–c); Gram-Humberstone (d). Bar, 100 lm (a); 20 lm (b–d).
314 S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317
Virulence of V. harveyi strains, identified by phenotypical meth-ods and isolated from diseased shrimp, has been proven by chal-lenging shrimp larvae and juveniles. However, the results mayvary from one trial to another because it is very difficult to repro-duce an experimental infection using shrimp (Soto-Rodriguezet al., 2006). Thus, so far, there is no phenotypic or genotypicfeature to distinguish pathogenic from non-pathogenic strains,and the only manner to determine the virulence of a given strain
is through experimental infection because many environmentalparameters, the bacterial strain and biological factors may influ-ence the results. Especially because the acquisition of virulencegenes by horizontal gene transfer might increase the ability of V.harveyi to infect aquatic organisms, by increasing virulence towardsa specific host and/or by broadening its host range (Ruwandeepikaet al., 2010). In the present study, BRS was reproduced easily and

Fig. 6. Microphotographs of antennal gland (AG) of L. vannamei. (a) Before 5 h p.i.with Vh CAIM 1792 at 104 CFU g�1 antennal glandular epithelium (GE) showsnecrosis and pyknotic hemocytes in the hemolymph sinuses as a response tobacterial masses (asterisk); (b) AG 5–7.5 h p.i. with Vh CAIM 1792 at 103 CFU g�1 GEshows necrosis with hemocytic infiltration (H) inside haemal spaces. L: lumen ofthe glandular tubules; (c) necrosis of gill lamellae of L. vannamei before 5 h p.i. withVh CAIM 1792 at 104 CFU g�1 where bacterial masses were observed (asterisk); (d)necrosis (asterisk) of gill lamellae 168 h p.i. with Vh CAIM 1792 at 102 CFU g�1 withformation of hemocytic nodules (arrow). H&E bar, 20 lm (a, c, and d); 50 lm (b).
Fig. 7. Microphotography of skeletal muscle (MU) of L. vannamei on the site ofinjection. (a and c) Shrimp challenge with Vh CAIM 1792 at 2.8 � 103 CFU g�1
grown at 4.0% and 1.0% NaCl respectively. (a) 19 h p.i. showed necrosis (N) of theskeletal muscle with an increased inflammatory response (H), starting theformation of hemocytic nodules; (b) L. vannamei injected with ECPs of Vh CAIM1792 grown to 1.0% NaCl at 8 h p.i. showing a severe necrosis of muscle fibers (N)around the site of injection; (c) 168 h p.i. showing a chronic lesion with a decreasedinflammatory response (H). A melanizated (M) granuloma with necrotic materialinside was too observed in the tissue. H&E bar, 100 lm.
S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317 315
the gross signs in challenged shrimp were similar to shrimp vibrio-sis (Alapide-Tendencia and Dureza, 1997; Manilal et al., 2010) butthe opacity at the site of injection has never been reported. Musclenecrosis in cultured shrimp is commonly observed and, often re-sults in the appearance of white and opaque lesions in the abdom-inal MU in response to environmental factors, such as suddenchanges in temperature, salinity, or other stress. During the pastfew years, virus infections also have been found to cause musclenecrosis in cultured shrimp. A nodavirus (tentatively named PvNV,
Penaeus vannamei nodavirus) that causes MU necrosis in P. vanna-mei was found in Belize in 2004 (Tang et al., 2007). Infectious myo-necrosis virus (IMNV) is another virus that causes muscle necrosisin L. vannamei cultured in Brazil and Indonesia (Poulos et al.,2006). The virus causes abdominal muscle necrosis with particu-larly emphasis on the sixth abdominal segment. Similarly, the maingross signs of BRS are opacity on the abdominal muscle and reddiscoloration spots on the abdominal cuticle of L. vannamei; how-ever the difference with virus infections is the presence of melan-ization around the red spots (Soto-Rodriguez et al., 2010b). Thesimilarity between gross signs observed during these viral infec-tions and BRS could be confusing, therefore it is very important toconfirm the findings by using histology and/or molecular methods.

316 S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317
In the present work, Vh CAIM 1792 was pathogenic to shrimpcausing more than 80% mortality before 24 h p.i. at dose of103 CFU g�1. Challenges with V. harveyi strains reported mortalitiesof about 50% at doses exceeding 104 CFU g�1 (Liu et al., 1996; Alap-ide-Tendencia and Dureza, 1997; Jayasree et al., 2006; Rattanamaet al., 2009). These findings clearly demonstrate differences inthe virulence of V. harveyi strains. In general, challenges with ECPswere more toxic than live cells, especially in the first hours. It hasbeen suggested that salinity concentration of bacteriological mediaof V. harveyi strains might affect their virulence (Prayitno andLatchford, 1995; Shivappa, 1997). ECPs obtained at 2.0% and 4.0%NaCl caused the highest mortalities. Before 24 h p.i., live cellsgrown at 4.0% NaCl and corresponding ECPs were more pathogeniccausing mortalities that ranged from 70% to 100%. In contrast, after48 h p.i., no differences were found in mortality between chal-lenges with bacteria at the same salinities (see Table A2 in Appen-dix). Our results suggest no clear relationship between NaClconcentration and virulence of Vh CAIM 1792.
There have been numerous studies describing morphologicalchanges of shrimp tissues infected with Vibrio, but informationon experimentally induced infection with V. harveyi is scarce.Histopathology of affected shrimp with Vibrio sp., including V. har-veyi, has always reported the hepatopancreas as the target organ ofinfection (Martin et al., 2004). Histological analysis carried out onshrimp infected with Vh CAIM 1792 revealed the LO as the targetorgan of infection; in the first hours p.i., a total loss of structure oc-curred. At 168 h p.i., severe necrosis of the LO, with formation ofhemocyitic nodules was observed. Egusa et al. (1988) reportedsimilar pathological changes in the LO of Penaeus japonicus exper-imentally infected with Vibrio sp. where no extensive necrotic le-sions were found in other organs. It is believed that the LO isessentially a filtering organ that clears the hemolymph of infec-tious agents as well as for homeostatic improvement of the hemo-lymph filtrate (Van de Braak et al., 2002; Duangsuwan et al., 2008).In the present study, severe necrosis was also observed withinmuscle fibers at the site of injection and surrounding connectivetissue as well as antennal gland and gill filaments. Bacteremiawas evident throughout the hemocele.
No histopathological differences were observed among L. van-namei challenged with Vh CAIM 1792 or injected with ECPs andthat were grown at 1.0%, 2.0% and 4.0% NaCl. Damage again, wasmainly observed in the site of injection at the first hours p.i., LOand heart that could be attributed to the enzyme activity of theECPs. Later, hemocytic inflammation with formation of hemocyticnodules and granulomas was observed within the muscle. Addi-tionally, ECPs also caused acute severe necrosis at the site of injec-tion, without hemocytic infiltration.
In conclusion, phenotypic characteristics of Vh CAIM 1792 weredifferent to others V. harveyi strains. These include tolerance to awide range of salinities, such as extreme environments, wherethe viability of the bacteria could be retained and remain infec-tious. Fortunately, Vh CAIM 1792 was sensitive to most antibioticsused in shrimp culture, so if an outbreak by BRS occurs, it can becontrolled. Virulence of Vh CAIM 1792 reflects the synergisticinteractions among virulence factors mainly the hydrophobicity,biofilm formation, siderophores production, and the proteolytic,enzymatic causing degradation of the shrimp tissue. No clear rela-tionship was found between salinity growth condition of bacteriaor ECPs and virulence. Further studies will be required to deter-mine the relationship, if any, between the pathogenicity of VhCAIM 1792 and natural infection conditions such as water temper-ature, host species, infection route, and the existence of stress. Atpresent, the authors are looking for, in silico, virulence genes ofVh CAIM 1792. Future work is necessary to prove if those potentialvirulence genes are responsible for virulence in its natural host, thepenaeid shrimp, considering that the acquisition of virulence genes
by horizontal gene transfer might increase the ability of Vh CAIM1792 to infect aquatic organisms, by increasing its virulence to-wards a specific host and/or by broadening its host range.
Acknowledgments
This study was in part supported by shrimp farmer’s contribu-tions. We thank C. Bolan and F. Marrujo for technical assistance,S. Abad for the microphotographs, N. Duncan for reviewing thegrammar, and C. Cabrera for donating L. vannamei.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.jip.2012.01.006.
References
Abraham, T.J., Manley, R., Palaniappan, R., Dhevendaran, K., 1997. Pathogenicity andantibiotic sensitivity of luminous Vibrio harveyi isolated from diseased penaeidshrimp. J. Aquat. Tropics 121, 1–8.
Alapide-Tendencia, E.V., Dureza, L.A., 1997. Isolation of Vibrio spp. from Penaeusmonodon (Fabricius) with red disease syndrome. Aquaculture 154, 107–114.
Alcaide, E., Amaro, C., Todolí, R., Oltra, R., 1999. Isolation and characterization ofVibrio parahaemolyticus causing infection in Iberian toothcarp Aphanius iberus.Dis. Aquat. Org. 35, 77–90.
Alsina, M., Blanch, A.R., 1994. Improvement and update of set of keys forbiochemical identification of environmental Vibrio species. J. Appl. Bacteriol.77, 719–721.
Amaro, C., Aznar, R., Alcaide, E., Lemos, M.L., 1990. Iron-binding compounds andrelated outer membrane proteins in Vibrio cholerae non-O1 strains from aquaticenvironments. Appl. Environ. Microbiol. 56, 2410–2416.
Amaro, C., Biosca, E.F., Esteve, C., Fouz, B., Toranzo, A.E., 1992. Comparative study ofphenotypic and virulence properties in Vibrio vulnificus biotypes 1 y 2 obtainedfrom an European eel farm experiencing mortalities. Dis. Aquat. Org. 13, 29–32.
Austin, B., Zhang, X.H., 2006. Vibrio harveyi: a significant pathogen of marinevertebrates and invertebrates. Lett. Appl. Microbiol. 43, 119–124.
Barry, A., Thornsberry, C., 1985. Susceptibility test diffusion test procedures. In:Lennette, E. (ed.), Manual of Clinical microbiology, 4th. Washington D.C.,American Society for Microbiology, pp. 978–987.
Baticados, M.C.L., Lavilla-Pitogo, C.R., Cruz-Lacierda, E.R., de la Pena, L.D., Sunaz,N.A., 1990. Studies on the chemical control of luminous bacteria Vibrio harveyiand V. splendidus isolated from diseased Penaeus monodon larvae and rearingwater. Dis. Aquat. Org. 9, 133–139.
Bauer, A.W., Kirby, W.M.M., Sherris, J.C., Turck, M., 1966. Antibiotics susceptibilitytesting by a standardized single disk method. Am. J. Clin. Pathol. 45, 493–496.
Bell, T.A., Lightner, D.V., 1988. A Handbook of Normal Penaeid Shrimp Histology.World Aquaculture Society, Baton Rouge, LA, USA.
Bradford, M.M., 1976. A rapid a sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein–dye binding. Anal.Biochem. 72, 248–254.
Cabanillas-Beltran, H., LLausas-Magaña, E., Romero, R., Espinoza, A., Garcia-Gasca,A., Nishibuchi, M., Ishibashi, M., Gomez-Gil, B., 2006. Outbreak of gastroenteritiscaused by the pandemicVibrio parahaemolyticus O3: K6 in Mexico. FEMSMicrobiol. Lett. 265, 76–80.
Chanratchakool, P., Pearson, M., Limsuwan, C., Roberts, R.J., 1995. Oxytetracyclinesensitivity of Vibrio species isolated from diseased black tiger shrimp, Penaeusmonodon Fabricius. J. Fish Dis. 18, 79–82.
de la Peña, L., Tamaki, T., Monoyama, K., Nakai, T., Muroga, K., 1993. Characteristicsof the causative bacterium of vibriosis in the kuruma prawn, Penaeus japonicus.Aquaculture 115, 1–12.
Daly, J.G., Stevenson, R.M.W., 1987. Hydrophobic and haemagglutination propertiesof Renibacterium salmoninarum. J. Gen. Microbiol. 133, 3575–3580.
Defoirdt, T., Darshanee Ruwandeepika, H.A., Karunasagar, I., Boon, N., Bossier, P.,2010. Quorum sensing negatively regulates chitinase in Vibrio harveyi. Environ.Microbiol. Rep. 2, 44–49.
Diggles, B.K., Moss, G.A., Carson, J., Anderson, C.D., 2000. Luminous vibriosis in rocklobster Jasus verreauxi (Decapoda: Palinuridae) phyllosoma larvae associatedwith infection by Vibrio harveyi. Dis. Aquat. Org. 43, 127–137.
Duangsuwan, P., Phoungpetchara, I., Tinikul, Y., Poljaroen, J., Wanichanon, C.,Sobhon, P., 2008. Histological and three dimensional organizations of lymphoidtubules in normal lymphoid organ of Penaeus monodon. Fish Shellfish Immunol.24, 426–435.
Egusa, S., Takahashi, Y., Itami, T., Momoyama, K., 1988. Histopathology of vibriosisin kuruma prawn Penaeus japonicus Bate. Fish Pathol. 23, 59–65.
Esteve, C., Herrera, F.C., 2000. Hepatopancreatic alterations in Litopenaeus vannamei(Boone, 1931) (crustacea: decapoda: penaeidae) experimentally infected withVibrio alginolyticus strain. J. Invertebr. Pathol. 76, 1–5.
Giono Cerezo, S., 1983. Prueba de Kirby-Bauer para sensibilidad a losantimicrobianos. Infectol. 3, 325.

S.A. Soto-Rodriguez et al. / Journal of Invertebrate Pathology 109 (2012) 307–317 317
Goarant, C., Reynaud, Y., Ansquer, D., de Decker, S., Saulnier, D., le Roux, F., 2006.Molecular epidemiology of Vibrio nigripulchritudo, a pathogen of culturedpenaeid shrimp (Litopenaeus stylirostris) in New Caledonia. Syst. Appl.Microbiol. 29, 570–580.
Gomez-Gil, B., Thompson, F.L., Thompson, C.C., Swings, J., 2003. Vibrio rotiferianussp. nov., isolated from cultures of the rotifer Brachionus plicatilis. Int. J. Syst. Evol.Microbiol. 53, 239–243.
Gomez-Gil, B., Soto-Rodriguez, S., Garcia-Gasca, A., Roque, A., Vazquez-Juarez, R.,Thompson, F.L., Swings, J., 2004. Molecular identification of Vibrio harveyi-related isolates associated with diseased aquatic organisms. Microbiology 150,1769–1777.
Haldar, S., Neogi, S.B., Kogure, K., Chatterjee, S., Chowdhury, N., Hinenoya, A.,Asakura, M., Yamasaki, S., 2010. Development of a haemolysin gene-basedmultiplex PCR for simultaneous detection of Vibrio campbellii, Vibrio harveyi andVibrio parahaemolyticus. Lett. Appl. Microbiol. 50, 146–152.
Hindler J., 1992. Clinical Procedures Handbook I. American Society for MicrobiologySection 5, Washington, DC.
Holt, J.G., Krieg, N.R., Sneath, P.H.A., Staley, T., Williams, S.T., 1994. Bergey’s Manualof Determinative Bacteriology, ninth ed. Williams and Wilkins, Baltimore.
Jayasree, L., Janakiram, P., Madhavi, R.T., 2006. Characterization of Vibrio spp.associated with diseased shrimp from culture ponds of Andhra Pradesh (India).J. World Aquacult. Soc. 37, 523–532.
Karunasagar, I., Otta, S.K., Karunasagar, I., 1996. Biofilm formation by Vibrio harveyion various surfaces. Aquaculture 140, 241–245.
Lavilla-Pitogo, C.R., Baticados, M.C.L., Cruz-Lacierda, E.R., de la Pena, L.D., 1990.Occurrence of luminous bacterial disease of Penaeus monodon larvae in thePhillipines. Aquaculture 91, 1–13.
Lin, B., Wang, Z., Malanoski, A.P., O’Grady, E.A., Wimpee, C.F., Vuddhakul, V., Alves Jr,N., Thompson, F.L., Gomez-Gil, B., Vora, G.L., 2010. Comparative genomicanalyses identify the Vibrio harveyi genome sequenced strains BAA-1116 andHY01 as Vibrio campbellii. Environ. Microbiol. Rep. 2, 81–89.
Liu, P.C., Lee, K.K., Chen, S.N., 1996. Pathogenicity of different isolates of Vibrioharveyi in tiger prawn, Penaeus monodon. Lett. Appl. Microbiol. 22, 413–416.
Liu, P.C., Lee, K.K., 1999. Cysteine protease is a major exotoxin of pathogenicluminous Vibrio harveyi in the tiger prawn, Penaeus monodon. Lett. Appl.Microbiol. 28, 428–430.
Liu, P.C., Lee, K.K., Tu, C.C., Chen, S.N., 1997. Purification and characterization of acysteine protease produced by pathogenic luminous Vibrio harveyi. Curr.Microbiol. 35, 32–39.
Manilal, A., Sujith, S., Selvin, J., Shakir, C., Gandhimathi, R., Kiran, G.S., 2010.Virulence of vibrios isolated from diseased Black Tiger Shrimp, Penaeusmonodon, Fabricius. J. World Aquacult. Soc. 41, 332–343.
Martin, G.G., Rubin, N., Swanson, E., 2004. Vibrio parahaemolyticus and V. harveyicause detachment of the epithelium from the midgut trunk of the penaeidshrimp Sicyonia ingentis. Dis. Aquat. Org. 60, 21–29.
Montero, A.B., Austin, B., 1999. Characterization of extracellular products from anisolate of Vibrio harveyi recovered from diseased post-larval Penaeus vannamei(Bonne). J. Fish Dis. 22, 377–386.
Najiah, M., Lee, S.W., Wendy, W., 2008. Phenotypic and genotypic characteristics ofVibrio harveyi isolated from Black Tiger Shrimp (Penaeus Monodon). World Appl.Sci. J. 3, 885–902.
Nakayama, T., Ito, E., Nomura, N., Nomura, N., Matsumura, M., 2006. Comparisonof Vibrio harveyi strains isolated from shrimp farms and from culture collectionin terms of toxicity and antibiotic resistance. FEMS Microbiol. Lett. 258,194–199.
Owens, L., Austin, D.A., Austin, B., 1996. Effect of strain origin on siderophoreproduction in Vibrio harveyi isolates. Dis. Aquat. Org. 27, 157–160.
Poulos, B.T., Tang, K.F.J., Pantoja, C.R., Bonami, J.R., Lightner, D.V., 2006. Purificationand characterization of infectious myonecrosis virus of penaeid shrimp. J. Gen.Virol. 87, 987–996.
Prayitno, S.B., Latchford, J.W., 1995. Experimental infections of crustaceans withluminous bacteria related to Photobacterium and Vibrio: effect of salinity and pHon infectiosity. Aquaculture 132, 105–112.
Lynch, M.J., Raphael, S.S., 1983. Lynch’s Medical Laboratory Technology, fourth ed.W.B. Saunders, Philadelphia.
Rattanama, P., Srinitiwarawong, K., Thompson, J.R., Pomwised, R., Supamattaya, K.,Vuddhakul, V., 2009. Shrimp pathogenicity, hemolysis, and the presence ofhemolysin and TTSS genes in Vibrio harveyi isolated from Thailand. Dis. Aquat.Org. 86, 113–122.
Roque, A., Turnbull, J.F., Escalante, G., Gomez-Gil, B., Alday Sanz, M.V., 1998.Development of a bath challenge for the marine shrimp Penaeus vannameiBoone, 1931. Aquaculture 169, 283–290.
Roque, A., Molina-Aja, A., Bolan-Mejia, C., Gomez-Gil, B., 2001. In vitro susceptibilityto 15 antibiotics of vibrios isolated from penaeid shrimps in NorthwesternMexico. Intl. J. Antimicrob. Agents 17, 383–387.
Rosenberg, M., Gutnick, D., Rosenberg, E., 1980. Adherence of bacteria tohydrocarbons: a simple method for measuring cell surface hydrophobicity.FEMS Microbiol. Lett. 9, 29–33.
Ruwandeepika, H.A.D., Defoirdt, T., Bhowmick, P.P., Shekar, M., Bossier, P.,Karunasagar, I., 2010. Presence of typical and atypical virulence genes invibrio isolates belonging to the Harveyi clade. J. Appl. Microbiol. 109, 888–899.
Santos, Y., Bandin, I., Bruno, D.W., Ellis, A.E., Toranzo, A., 1990. Comparison of thecell surface hydrophobicity of bacterial fish pathogens by different procedures.In: Perkins, O.F., Chen, T.C. (Eds.), Pathology in Marine Sciences. Academic Press,San Diego, pp. 101–115.
Saulnier, D., Haffner, P., Goarant, C., Levy, P., Ansquer, D., 2000. Experimentalinfection models for shrimp vibriosis studies: a review. Aquaculture 191, 133–144.
Schwyn, B., Neilands, J.B., 1987. Universal chemical assay for detection anddetermination of siderophores. Anal. Biochem. 160, 47–56.
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y AlimentaciónSAGARPA, 2009. Anuario Estadístico de Acuacultura y Pesca 2009. <http://www.conapesca.sagarpa.gob.mx/wb/cona/anuario_2009>.
Selvin, J., Lipton, A.P., 2003. Vibrio alginolyticus associated with white spot disease ofPenaeus monodon. Dis. Aquat. Org. 57, 147–150.
Shivappa, R.B., 1997. Efficacy of Probiotics and Disinfectants in ControllingLuminescent Bacteria in Shrimp Post Larvae under Normal and StressedConditions. MSc Thesis, Asian Institute of Technology, Bangkok.
Soto-Rodriguez, S.A., Roque, A., Lizarraga Partida, M.L., Guerra Flores, A.L., GomezGil, B., 2003. Virulence of luminous Vibrios to Artemia franciscana Kellog, 1906,nauplii. Dis. Aquat. Org. 53, 231–240.
Soto-Rodriguez, S.A., Simoes, N., Roque, A., Gomez Gil, B., 2006. Pathogenicity andcolonization of Litopenaeus vannamei larvae by luminescent vibrios.Aquaculture 258, 109–115.
Soto-Rodriguez, S.A., Gomez Gil, B., Lozano, R., Roque, A., 2008. MICs to antibiotics ofVibrio spp. isolated from the Pacific White Shrimp Litopenaeus vannameicultivated in Mexico. Panorama Acuicola Mag. 11, 52–57.
Soto-Rodriguez, S.A., Gomez-Gil, B., Lozano, R., Roque, A., 2010a. Density of vibriosin hemolymph and hepatopancreas of diseased Pacific White Shrimp,Litopenaeus vannamei, from northwestern Mexico. J. World Aquacult. Soc. 41,76–83.
Soto-Rodriguez, S.A., Gomez-Gil, B., Lozano, R., 2010b. ‘‘Bright-red’’ Syndrome inPacific white shrimp Litopenaeus vannamei is caused by Vibrio harveyi. Dis.Aquat. Org. 92, 11–19.
Stabili, L., Gravili, C., Piraino, S., Boero, F., Alifano, P., 2006. Vibrio harveyi associatedwith Aglaophenia octodonta (Hydrozoa, Cnidaria). Microb. Ecol. 52, 603–608.
Tang, K.F.J., Pantoja, C.R., Redman, R.M., Lightner, D.V., 2007. Development of in situhybridization and RT-PCR assay for the detection of a nodavirus (PvNV) thatcauses muscle necrosis in Penaeus vannamei. Dis. Aquat. Org. 75, 183–190.
Teo, J.W.P., Tan, T.M.C., Poh, C.L., 2002. Genetic determinants of tetracyclineresistance in Vibrio harveyi. Antimicrob. Agents Chemother. 46, 1038–1045.
Tendencia, E.A., de la Pena, L., 2001. Antibiotic resistance of bacteria from shrimpponds. Aquaculture 195, 193–204.
Toranzo, A.E., Barja, J.L., Colwell, R.R., Hetrick, F.M., Crosa, J.H., 1983.Haemaglutination, haemolytic and cytotoxic activities of Vibrio anguillarumand related vibrios isolated from striped bass on Atlantic coast. FEMS Microbiol.Lett. 18, 257–262.
Van de Braak, C.B.T., Botterblom, M.H.A., Taverne, N., van Muiswinkel, W.B.,Rombout, J.H.W.M., van der Knaap, W.P.W., 2002. The roles of haemocytes andthe lymphoid organ in the clearance of injected Vibrio bacteria in Penaeusmonodon shrimp. Fish Shellfish Immunol. 13, 293–309.
Won, K.M., Park, S.I., 2008. Pathogenicity of Vibrio harveyi to cultured marine fishesin Korea. Aquaculture 285, 8–13.
Zhang, X.H., Austin, B., 2000. Pathogenicity of Vibrio harveyi to salmonids. J. Fish Dis.23, 93–102.
Zhong, Y.B., Zhang, X.H., Chen, J.X., Chi, Z.H., Sun, B.G., Li, Y., Austin, B., 2006.Overexpression, purification, characterization, and pathogenicity of Vibrioharveyi hemolysin VHH. Infect. Immun. 74, 6001–6005.