isolation of specialized transducing bacteriophages carrying the
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Feb. 1978, p. 549-5570021-9193/78/0133-0549$02.00/0Copyright 0 1978 American Society for Microbiology
Vol. 133, No. 2
Printed in U.S.A.
Isolation of Specialized Transducing Bacteriophages Carryingthe Structural Genes of the Hexuronate System in
Escherichia coli K-12: exu RegionMIREILTLE MATA,* MARIE DELSTANCHE, AND JANINE ROBERT-BAUDOUY
Laboratoire de Microbiologie de l'Institut National des Sciences Appliquees de Lyon, Laboratoire Propredu Centre National de la Recherche Scientifique no. 05421, 69621 Villeurbanne, France
Received for publication 1 September 1977
In Escherichia coli HfrH 58, isolated by Shimada et al., a heat-induciblephage has been integrated in a secondary attachment site. We have characterizedthe nature of the A integration. The exuR regulatory gene is inactivated byprophage integration. Genetic and biochemical analysis indicated a gene order:uxaA-uxaC-exuT-(exuR')-ANRAJ (exuR"). By induction of HfrH 58, one classof deletions extending into the exu region was obtained. Analysis ofthese deletionsconfirms the exu region topography and the regulatory mechanism of the hexu-ronate system previously described. It has been possible to regenerate a functionalexuR gene by prophage exision. Various lambda transducing particles, plaque-forming and defective transducing phages carrying the left part or the right partof the exu region, have been derived from the secondary site lysogen HfrH 58. Aphage carrying the entire exuR region was constructed by a cross between thesetwo types of phage. The construction and characterization of these exu transduc-ing phages are presented.
The aldohexuronates glucuronate and galac-turonate are degraded according to the Ashwellpathway (1) (Fig. 1). Fructuronate (D-lyxo-5-hexulosonate) is able to induce the synthesis ofthe five enzymes II, m, m', IV, IV' (14) andthat of the aldohexuronate transport systemTHU (8), whereas tagaturonate (D-arabino-5-hexulosonate) induces only the synthesis of theenzymes of the galacturonate branch (enzymesI,I ml, IV") (14) and that of the aldohexuronatetransport system. Fructuronate and tagaturon-ate are the true inducers. The structural genesof galacturonate catabolizing enzymes are lo-cated in two distinct regions on the chromosome:the exu region (8, 9, 10) (min 66), which includesthe exuR regulatory gene and the three struc-tural genes uxaC (enzyme II), uxaA (enzymeIV"), and exuT (aldohexuronate transport sys-tem); and the uxaB operon (enzyme M') (min52) (Fig. 1). The exu regulon involves all theabove-mentioned genes and is under the nega-tive control of the exuR regulatory gene product(21); (R. Portalier, J. Robert-Baudouy, and F.Stoeber, unpublished data).To study transcription in vitro, we tried to
obtain trnsducing phages carrying differentportions of the exu region. Shimada et al. (18)isolated several lysogens of Escherichia coli toanalyze secondary attachment sites of phage A.In one such lysogen, strain HfrH 58, the A pro-
phage site was located near metC (18), i.e., nearthe exu region. Upon heat induction of thislysogen, we isolated several transducing phagescarrying either the whole or a part of the exuregion. This paper describes the isolation andcharacterization of these phages as well as dele-tions in the exu region obtained by induction ofstrain HfrH 58.
MATERIALS AND METHODSBacterial and phage strains. The genotypes of
the bacterial and phage strains used in this study areshown in Table 1. The bacteriophage Xvir, Ximm21ccLXinm21b2c, and Xsus mutants were from the laboratoryof P. Thomas in Brussels.
Culture media. Media for growth were identicalto those described by Portalier et al. (10). Minimalmedium was M 63, pH 7.2 (20). Solid media containedglucose (5 mg/ml), glycerol (5 mg/ml), tagaturonate(3 mg/ml), and galacturonate, glucuronate, or altron-ate (2.5 mg/ml). Tetrazolium medium (7) containedgalacturonate at 5 mg/ml; aldohexuronate Mac-Conkey media (7) contained these sugars at 15 mg/ml.Enzyme induction and extraction. The condi-
tions for induction and enzyme extraction were out-lined previously (14). The inducer concentration was5 mM.Enzyme assays. Aldonic oxidoreductases and hy-
drolyases were assayed according to previously pub-lished methods (11, 12, 15); hexuronic isomerase wasmeasured by a coupling method described in a pre-vious paper (9).
549

550 MATA, DELSTANCHE, AND ROBERT-BAUDOUY
ATHU 11 III
atracLLular sMuT usaC uxuBDglucuronate _ D-glucurn = Dfructuronate = mannonate IV
uA2-I.tol3deoxyDigluconate
A/IV'
agalacturonste - D-pgaLuronh = D4igaturont. = Datronate
extracellutar THU T uxaC uxa a
A
FIG. 1. Degradative pathway of hexuronides and hexuronates in E. coli K-12. Symbols: II, uronateisomerase (EC 5.3.1.12); III, mannonate oxidoreductase (EC 1.1.1.57); IV, mannonate hydrolyase (EC 4.2.1.8);IIr, altronate oxidoreductase (EC 1.1.1.58); IV', altronate hydrolyase (EC 4.2.1.7); A, aldohexuronatestransport system (THU).
TABLE 1. StrainsStrains Relevant genotype Source/reference
E. coliHfrH 58 A(gal-bio) A C1857 integrated at exuR; M. Gottes-
manC600 (P2) Plaques Spi- phage; ThomasEWlb F- his-i toiC argG6 str E. WitneyRP1 Hfr uxaAl metB (10)MH2 Hfr uxaC202 metB (9)TH6 Hfr exuT6 metB (8)TH9 Hfr exut9 metB (8)HfrH Institut PasteurHJ1 Hfr exuRAl metB (14,21)RC1 Hfr exuRAUl metB (14, 21)1573 A(gal-bio) A(exuR'-exuT-uxaC- Heat survivor of HfrH 58; this study
uxaA)1574 A(gal-bio) A(exuR'-exuT-uxaC- Heat survivor of HfrH 58; this study
uxaA)1576 A(gal-bio) A(exuR'-exuT-uxaC- Heat survivor of HfrH 58; this study
uxaA)1693 A(gal-bio) Heat survivor of HfrH 58; this study1694 A(gal-bio) Heat survivor of HfrH 58; this study1576 (P2) This study
BacteriophagesApexul cI857 (exuR") Derived from HfrH 58; this studyXpexu2 cI857 (exuR')-exuT-uxaC-uxaA Derived from HfrH 58; this studyXpeux3 c1857 exuR-exuT-uxaC-uxaA Derived from a cross between Xpexul
and Apexu2; this studyApexu4 cI857 (exuR')-exuT Derived from HfrH 58; this studyApexu5 c1857 (exuR'-exuT-uxaC Derived from HfrH 58; this studyAdexul cI857 (exuR'-exuT-uxaC-uxaA Derived from HfrH 58; this studyAdexu2 cI857 Sam7 (part of exuR)-exuT- Isolated by Schrenk and Weisberg
uxaC-uxaA method
Chemicals. Intermediate substrates of the hexu-ronate pathway (tagaturonate, fructuronate, and al-tronate) were synthesized in our laboratory (9).
Lysogenization and transduction frequencies.Lysogenization frequency was determined as describedby Shimada et al. (18). Transduction frequency was
determined by the same procedure, except that in-fected cells were plated on minimal agar plates.
Preparation of phage lysates. Low-frequencyand high-frequency lysates (LFT and HFT) of lambdalysogens were prepared as described by Miller (7).
Isolation of plaque-forming transducingphages. LFT lysates were plated on a lawn of galac-turonate- recipients, using galacturonate tetrazoliumplates. Plaques formed by nondefective exu transduc-ing phages were easily identified by their halos andcolored centers after 36 h of incubation at 320C.
Isolation of ASpi-. ASpi- (6) were isolated usingfresh LFT lysates derived from HfrH 58, and 0.1 mlof the lysate, containing 10' plaque-forming units, wasmixed with 0.1 ml of an exponential-phase culture ofa P2 lysogen. After 15 min of adsorption, the phage-
J. BATRIUOL.

SPECIALIZED TRANSDUCING PHAGES IN E. COLI
infected cells were poured onto tryptone plates. Afterovernight incubation at 370C, plaques of XSpi- ap-
peared and were isolated as described by Boulter andLee (3).Prophage curing procedure. The method for
curing A lysogens by a thermal treatment (18) was
modified as follows. The A lysogen cultures were
spread at 420C and 32°C either on tryptone plates or
galacturonate MacConkey medium or on minimal me-dia containing galacturonate, altronate, or tagaturon-ate as the sole carbon source. On galacturonateMacConkey, mutants deleted in the exu region are
easily screened. Wild-type survivors are red, whilethose having lost one or several genes, required forthe galacturonate dissimilation, produce white colo-nies. On galacturonate inimal medium, survivorsthat had retained all the genes needed for galacturon-ate metabolism were selected. Altronate and tagatu-ronate media should favor the production of survivorsdisplaying a functional uxaA gene but having lostexuT and uxaC genes.
RESULTSLocation of phage A in the eu region of
strain HfrH 58. Strain HfrH 58, provided byM. Gottesman, was induced at 420C, and severalindependent LFT lysates were obtained. Thesephages transduced various galacturonate- pointand deletion mutants to galacturonate+ (see be-low), but none of them was able to transducethe exuR gene. These results suggest that, instrain HfrH 58, phage A has inserted near or
into the exu region.HfrH 58 grew on aldohexuronates, thus indi-
cating that the presence of the prophage doesnot inactivate any of the exuT, uxaA, and uxaCstructural genes.The level of the different enzymes, II, III',
and IV', involved in the exu regulon was mea-
sured in the presence or absence of the inducerat 320C in strain HfrH 58 (Table 2). Synthesisof the three inducible enzymes, which are regu-lated by the exuR regulatory gene and belongto different operons, is constitutive in that strain.This character shows that exuR expression isaltered by A insertion (Fig. 2).
Isolation ofplaque-forming and defectiveexu transducing phages. Plaque-formingexu phages. LFT lysates made by heat induc-tion of HfrH 58 were used to transduce cells ofTH6 (exuT) at 320C on galacturonate tetrazo-lium plates. Twenty-four galacturonate+ trans-ductants picked up from halos ofcolored plaqueswere purified on homologous media and usedto prepare HFT lysates. Each HFT lysate was
spotted on strain 1576 (class B deletion, see Fig.6). Of the 24 HFT lysates, 6 transduced exuT+(like Xpexu4), 16 transduced exuT-uxaC (likeApexu5), and 2 transduced exuT-uxaC-uxaA(like Apexu2) (Fig. 3 and Table 3). All thesephages were plaque-forming.
TABLE 2. Activity of the exu regulon enzymes in E.coli K-12 in differential rate of synthesis
Relative differential rateof synthesis (%) in en-
Strain Inducer zymea:
II IV' m' THUWild type None 3 6 4 100
Fructuronate 100 100 100
HfrH 58 None 74 70 422Fructuronate 38 49 235
A class A None 4 5 7Fructuronate 102 128 87
A classB None 0 0 398 18Fructuronate 0 0 383
A class C None 75 74 410
TH9 None 18a Percentage of maximal value induced in the wild type.
The activities of strain HfrH grown with fructuronate aretaken as 100 baais for activities of enzymes II, M', and IV'.The wild-type strain for enzyme THU is MH2 (uxaC202) (8).THU activity, which is determined by the amount of intracel-lularly accumulated glucuronate, cannot be measured in HfrHand in class A and C deletions, because in these strains enzymeII synthesis is derepressed, and glucuronate cannot accumu-late.
Defective exu transducing phages. Thesephages were obtained by two different proce-dures. First, an LFT lysate of HfrH 58 was usedto transduce E. coli RP1 (uxaAl) on galacturon-ate MacConkey plates at 320C. One such defec-tive transducing phage, Adexul (Table 3), car-ried the bacterial genes exuT, uxaC, and uxaA.By marker rescue, we have shown that phagemarkers from K to J were deleted (Fig. 3). Thisresult shows that the prophage in HfrH 58 isoriented clockwise and suggests the followinggene sequence: uxaA-uxaC-exuT (exuR')XNRAJ (exuR") (Fig. 2). This is substantiatedby the results obtained with prophage deletions(Fig. 6).The other defective exu transducing phage,
Xdexu2 (Table 3), was obtained by induction ofa mixed culture of abnormal lysogens as de-scribed by Schrenk and Weisberg (16). Themixed lysate was used to transduce strain 1576.The phage Xdexu2 that we obtained transducedexuT, uxaC, and uxaA.ASpi- carrying the right part, (exuR"), of
exuR gene. XSpi- obtained from HfrH 58 byabnormal excision (see above) has lost phagegenes from the left end of the prophage map.The lost phage genes may be replaced by bac-terial DNA adjacent to the right prophage end(6), i.e., the (exuR") part of the exuR gene. Thepresence of (exuR") in the XSpi- was demon-strated by an indirect method (see below).
551VOL. 133, 1978
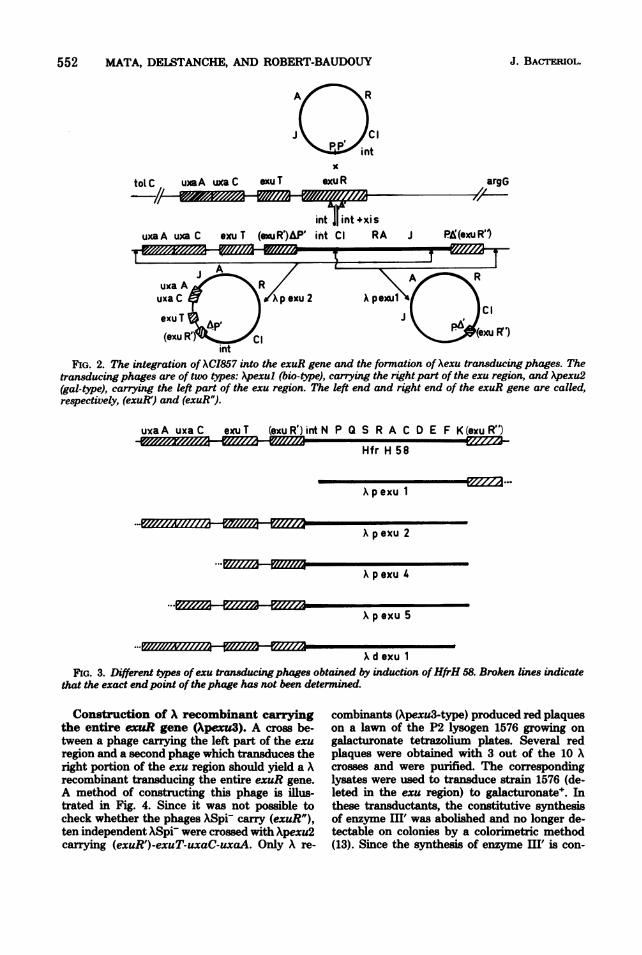
552 MATA, DELSTANCHE, AND ROBERT-BAUDOUY
:1
toL C uaA uxaC exuT exu R argG
uxa A Ruxa C a
exuT(9'(exuR) Iint
FIG. 2. The integration of ACI857 into the exuR gene and the formation of Aexu transducing phages. Thetransducing phages are of two types: Apexul (bio-type), carrying the right part of the exu region, and Apexu2(gal-type), carrying the left part of the exu region. The left end and right end of the exuR gene are called,respectively, (exuR') and (exuR").
uxa A uxa C exu T (exu R') int N P Q S R A C D E F K(exuR")
Hfr H 58
Xpexu 1
... iz::i iX::-?~p exu 2
Apxu 4
..._
p exu S
x d exu 1FIG. 3. Different types ofexu transducing phages obtained by induction ofHfrH 58. Broken lines indicate
that the exact endpoint of thephage has not been determined.
Construction of X recombinant carryingthe entire exuR gene (Xpexu3). A cross be-tween a phage carrying the left part of the exuregion and a second phage which transduces theright portion of the exu region should yield a Arecombinant tranaducing the entire exuR gene.A method of constructing this phage is illus-trated in Fig. 4. Since it was not possible tocheck whether the phages XSpi- carry (exuR"),ten independent XSpi- were crossed with Xpexu2carrying (exuR')-exuT-uxaC-uxaA. Only A re-
combinants (Apexu3-type) produced red plaqueson a lawn of the P2 lysogen 1576 growing ongalacturonate tetrazolium plates. Several redplaques were obtained with 3 out of the 10 Acrosses and were purified. The correspondinglysates were used to transduce strain 1576 (de-leted in the exu region) to galacturonatel. Inthese transductants, the constitutive synthesisof enzyme Il' was abolished and no longer de-tectable on colonies by a colorimetric method(13). Since the synthesis of enzyme m' is con-
-simEmo-
.
J. BACTER1OL.

SPECIALIZED TRANSDUCING PHAGES IN E. COLI
stitutive in strain 1576 (see class B deletion,below), the presence of a functional exuR geneshould render this strain inducible. Table 4shows the level of enzymes II, III', and IV' instrain 1576 transduced by Apexu3, measured inthe presence and absence of galacturonate asinducer. The synthesis of these enzymes is nolonger constitutive, but they present a wild-typeinduction pattern, thus proving that a functionalexuR gene has been restored.A recombinants (Apexu3-like phages) carry the
entire exuR gene and the exuT-uxaC-uxaA
TABLE 3. Mapping data for the Xexu transducingphagese
Galac- Galac- Galac- Galac- Glucu-turon- turon- turon- turon- ronate
Phage ate, ate, ate, ate, 1576a,TH6, MH2, RP1, 1576 A clam BexuT uxaC uxaA classB clasB
Xpexul 0 0 0 0Xpexu2 + + + +Apexu3 + + + +Xpexu4 + 0 0 0 0Xpexu5 + + 0 0 +Xdexul + + + + +Xdexu2 + + + + +
a Xpexul, -2, 4, and -5 and Adexul were obtainedby induction of HfrH 58 (see Fig. 3), and Xdexu2 wasisolated as described by Schrenk and Weisberg (16).Apexu3 is a recombinant from Xpexu2 and Xpexul. Afresh lysate containing 1iO to 1010 plaque-forming unitsper ml was prepared for each ransducing phage andwas spotted on galacturonate or glucuronate minimalplate seeded with E. coli K-12 galacturonate- mutants.The plates were examined for galacturonate+ or glu-curonate+ transductants after 48 h of growth at 32°C.Symbols: 0, no transductants obtained; +, tranaduc-tants obtained. The transduction frequency of thepoint mutants Aatt+ (TH6-MH2-RP1) with Xpexu2,-4, and -5 was about 1%. The transduction frequencyof mutant 1576 with the same phages was less than0.1%.
genes. This result proves that the three ASpi-which are able to recombine carry the right part,(exuR"), of the exuR gene (Apexul-type, see
above).Lysogenization of strain 1576 by phages
Apexu2 and Adexu2. Strain 1576 lysogenizedby Apexu2 presents two genotypes as describedin Fig. 5. Two different events lead to lysogeni-zation of strain 1576. On the one hand, integra-tion at specific sites restores a whole and func-tional exuR gene (93% of lysogens). On the otherhand, integration by homology between se-
quences on the phage and the chromosomeleaves exuR nonfunctional (57% of lysogens),and enzyme III' is synthetized constitutively. Inthe second alternative, 100% of the transduc-tants obtained by transduction of 1576 withAdexu2, isolated by the method of Schrenk andWeisberg (16), retain constitutivity for the syn-
thesis of enzyme III'. At this point, we cannotconclude that Adexu2 carries a part of exuR,but, if it does, this part is probably differentfrom (exuR') because the entire exuR gene isnever restored by integration of Xdexu2.exu transducing phage and the exuR su-
perrepressible mutations. Two types of su-
perrepressible mutation have been isolated forthe exu regulon, type RC1 (genotype exuRAUl)and type HJ1 (genotype exuRAl) (14, 21; Por-talier et al., unpublished data). These pleiotropicmutations have been located in the exuR gene.Phage Apexu2 (gal-type) and Apexul (bio-type)carry, respectively, the left part and the rightpart ofthe exuR gene. To determine the positionof these exuR transdominant mutations in one
or the other part of exuR, strains RC1 and HJ1were infected by the two phage types, and re-
combinants able to grow on galacturonate min-imal medium were selected. Reversion fre-quency of these exuR mutations was 6 x 10'.Revertants were distinguished from wild-typerecombinants by estimation of enzyme III' con-
A J uxa A uxa C exu T exu R ) int N Cl R
X pexu 2P
A J b2 PI& N Cl R
opexul (exR
A J uxaA uxaC exuT exuR NCI R
XpLxu 3FIG. 4. Diagrammatic representation of the phage cross Apexul x Apexu2 yielding Apexu3 carrying exuR-
exuT-uxaC-uxaA.
553VOL. 133, 1978

554 MATA, DELSTANCHE, AND ROBERT-BAUDOUY
stitutivity. For both exuR mutations, recombi-nants were obtained with Xpexu2 but not withXpexul. This result suggests that the two exuRmutations were located in the left part, (exuR'),of the exuR gene.The same experiment was carried out with
Xdexu2. Recombinants were obtained with RC1infected by Xdexu2. HJ1 gave no recombinants.This result is consistent with the view thatXdexu2 carries a part of the exuR gene but thatthis portion is smaller than the (exuR') part.These two exuR mutations are not located atthe same point of the exuR gene, which corrob-orates unpublished results (Portalier et al. andRobert-Baudouy et al., unpublished data) andplaces the exuRAUl mutation closer to exuTthan exuRAl.
TABLE 4. Activity of the exu regulon enzymes indifferential rates of synthesis (percent ofmaximalvalue induced in the wild type) in strain 1576 beforeand after transduction by Apexu3 (k recombinant
phage carrying exuR-exuT-uxaA-uxaC)Relative differentialrate of synthesis (%)
Strain Inducer in enzyme:
II IV' IHlHfrH None 3 6 4
Galacturonate 100 100 100
1576 None 0 0 395Galacturonate 0 0 320
1576, None 0 4 2(Apexu3) Galacturonate 113 110 112
1576
exuTApexu2
uxaC IuxaA
Deletion mutants formed on induction ofHfrH 58. When HfrH 58 is incubated at 420C,most cells die because phage functions lethal tothe host are induced. In the rare heat-resistantsurvivors, phage genes specifying or controllingthese lethal functions have been deleted (17).Among the survivors, one would expect to finddeletions extending into adjacent bacterialgenes, i.e., in the exu region. For such an exper-iment, a Xvir-resistant derivative of strain HfrH58 was examined. Survivors were selected ontryptone broth or galacturonate MacConkeymedium or on minimal media supplementedwith galacturonate, tagaturonate, or altronateas the sole carbon source. The growth of deletionmutants on these minimal media requires thepresence of genes involved in hexuronate dissi-milation. Deletions to various lengths in the exuregion were expected by this selection (seeabove), but it was not possible to obtain dele-tions extending into exuT or exuT-uxaC whilekeeping uxaA undamaged. To explain this be-havior, hypotheses are given in the discussionsection.The frequency of thermal curing of HfrH 58
was about 10- for survivors selected on richmedia and about 10' for survivors selected onminimal media.
Survivors were purified and analyzed for re-sidual phage and hexuronate genes (Fig. 6).Their growth phenotypes were examined, andactivities of enzymes II, m', and IV' were mea-sured in the presence or absence of inducer(Table 2). Three classes of survivors, A, B, andC, were obtained. Class B mutants were isolated
deletionex F AexuR"
uxaC
R, bexuT
exu R
F J uxaA uxaC exuT (exuR) int Cl RA J (exuW)
FIG. 5. Diagrammatic representation of the events leading to lysogenization of strain 1576 by phageApexu2. Integration occurs (a) by reciprocal recombination between AP on the phage and PA', nucleotidesequence of strain 1576; or (b) by a reciprocal recombinational event involving homologous sequences on thephage and chromosome (F to J).
J. BACTERIOL.

SPECIALIZED TRANSDUCING PHAGES IN E. COLI
HfrH uxaA uxaC exuT (exuR')N P Q S R A C D E F K(exuR")
classE_ _ _ _-.E1+ + + B1576
RT + + + + 4-1573 B
1574 XE- _
1693, 4 +1694 '
_ _ _4_+ + +4 + + + 4 4+
+ N+r- + + +
B
C
FIG. 6. Prophage deletion mapping of HfrH 58. Derivatives of strain HfrH 58 able to grow at 42°C werescored for phage and bacterial markers as follows. Detection of residual phage markers is as described byShimada et aL (18). The bacterial DNA deletions were deternined by the growth phenotype on hexuronatesand by measure of enzymes II, IIr, IV' activity (Table 2). Class B deletion was studied by cross-streakingthe deleted mutants with mutants containing various point mutations in the structural genes of the exuregion. *, Part of this gene is present.
only on galacturonate MacConkey medium, andclasses A and C could be obtained on eachmedium mentioned above.Class A (95% of survivors). These mutants
have wild-type behavior and catabolize galactu-ronate and glucuronate. The product of theexuR gene is functional; enzymes II, 111, andIV' are inducible by fructuronate, and their dif-ferential rates of synthesis are identical to thoseobtained with a wild-type strain (Table 2). Thedeletion has eliminated the entire prophage.Class B (0.6% of survivors). This class
grows on fructuronate but not on aldohexuron-ates and tagaturonate. Enzymes II and IV' arenot inducible by fructuronate, and the synthesisof enzyme IH' is constitutive (Table 2). Thealdohexuronate transport activity is much lowerthan that of the wild-type strain (18%). Thisbasal level is identical to that of a strain bearinga mutation in the structural gene of the per-mease (8).For genetic analysis, 15 independent mutants
were examined. The mutations were transferredin a F- strain, EWlb. Presence of bacterial genesuxaA, uxaC, and exuT was tested by cross-streaking the F- class B strains with Hfr mutantsbearing various independent point mutations inthe structural genes of the exu region; 10 uxaA,4 uxaC, and 11 exuT Hfr were used (8, 9, 10).Recombinants were selected on galacturonateand glucuronate minimal media. F- class B mu-tants were unable to recombine with uxaC andexuT Hfr. The class B deletion extends at leastinto exuTand uxaC genes. The same F- deletionmutants crossed with uxaA- Hfr gave no recom-binants able to grow on galacturonate, but re-combinants able to grow on glucuronate were
obtained. Class B strains transduced by a Apexucarrying exuT and uxaC but not uxaA genes(Xpexu5, Table 1) have the same behavior, trans-ductants catabolize glucuronate but not galac-turonate.
All these data indicate that the class B dele-tion encompasses exuT, uxaC, and uxaA (Fig.6). Presence of phage markers was analyzed forthree class B strains (Fig. 6). Some phage geneswere rescued in tests with Xsus lines; N, P, andQ A genes at least were deleted. Consequently,the left end, (exuR'), of the exuR gene, whichis located between exuT and X insertion, is en-compassed by the deletion.Class C (4% of survivors). This class can
grow on aldohexuronates, thus indicating thatnone of the exuT, uxaA, and uxaC structuralgenes is deleted. The synthesis of enzymes II,I', and IV' is constitutive and identical to thatof HfrH 58. Some phage markers are deleted(Fig. 3), and the exuR gene is inactivated as inHfrH 58 by the residual X genes.
DISCUSSIONWe have demonstrated that, in strain HfrH
58, phage A was inserted into the exuR geneand rendered it nonfunctional. Two classes oftransducing phage were isolated by abnormalexcision, Agal-type, transducing the left part ofthe exu region, and Xbio-type, transducing theright part. We isolated aX recombinant carryingthe entire exu region (Xpexu3) by a cross be-tween these two types of phage. The Xpexu3isolation is sinilar to that described by Shimadaet al. (19) for XptrpEDCBA phage. The forma-tion of this phage occurs by an int-xis promotedrecombination between secondary attachment
555VOL. 133, 1978

556 MATA, DEISTANCHE, AND ROBERT-BAUDOUY
sites of the bacterium and phage chromosome,AP' and PA'. The length of bacterial DNA car-ried by each type of phage is being investigatedusing restriction enzymes and heteroduplexmapping.Our results indicate that the ?gal-type trans-
ducing phages derived from HfrH 58 and Xdexu2isolated by Schrenk's method both carry a dif-ferent portion of the exuR gene. It appears thatphage X can integrate in the exuR gene at twodifferent locations as in the araB gene (3).
Isolation of X exu transducing particles anddeletions is proving to be extremely useful instudies on the expression of the exu regulon.We have shown that strain 1576 (class B dele-
tion) transduced by Xpexu5 (XpexuT-uxaC) isunable to dissimilate galacturonate because itlacks the uxaA gene, but grows on glucuronate.Since metabolism of glucuronate requires theexpression of isomerase (enzyme II) coded forby the uxaC gene, the growth of 1576 (Xpexu5)on glucuronate suggests that the promoter ofthe uxaC-uxaA operon is located between exuTand uxaC, and that the transcription directionmight be from uxaC to uxaA.Heat survivors selected from HfrH 58 were
able to regenerate a functional exuR gene (con-sistent with a mechanism of int-promoted re-combination [4]) or provided large deletions(class B) in the exu region involving (exuR')-exuT-uxaC and uxaA with variable end pointsinto the prophage. Another set of deletions ex-tending only in (exuR')-exuT-uxaC was ex-pected by selection on tagaturonate or altronate.Growth on these sugars requires only the expres-sion of the uxaA gene. We assume that thisselection was inadequate, since our results sug-gest that the promotor ofthe uxaC-uxaA operonis located between exuT and uxaC. It shouldbe possible to obtain every kind of deletion ongalacturonate MacConkey medium. We did notsucceed in selecting them because the numberof deletions analyzed was too small (geneticanalysis was carried out on for 15 independentdeletions).
Class B deletion was unable to catabolize gal-acturonate, tagaturonate, and glucuronate, butgrew on fructuronate. This result confirms thatthe fructuronate transport system, which hasnot yet been located, is not affected by thedeletion. In this same class of deletion, the basallevel of enzyme THU is identical to that of astrain bearing a mutation in the structural geneof the transport system. Among negative exuTpoint mutations, none was found to completelyabolish hexuronate transport activity (8). Sinceall the deletions also exhibit a residual THUactivity, the result would suggest that another
transport system is responsible for slow aldo-hexuronate uptake in the cells.
In the deletion strains as well as in strainHfrH 58, the activity of enzyme Il' is stronglyderepressed, corresponding to a three- to four-fold higher level than the fully induced wild-type strain (Table 2). This suggests that thewhole regulatory system involved in the synthe-sis of that enzyme is not functional in thesestrains. Consequently, the transcription of thestructural gene of enzyme III' is probably sub-jected to the action of the exuR product.
ACKNOWLEDGMaENSThis work was supported by gants from the Centre Na-
tional de la Recherche Scientifique (laboratoire propre duC.N.R.S. no. 05421) and from the Fondation pour la Re-cherche Medicale Fran9aise.We thank F. Stoeber for the encouragement he has given
to this study. We are very grateful to M. E. Gottesman forthe strain HfrH 58 and for helpful disctussions. We also thankC. Marchal and M. Hofnung of Institut Pasteur for fruitfuladvice, D. Atlan and S. Ottomani for skillful technical assist-ance, and Le Thy Bich Thuy.
LITERATURE CTE1. Ashwell, G. 1962. Enzymes ofglucuronic and galacturonic
acid metabolism in bacteria. Methods Enzymol.5:190-208.
2. Bachmann, B. J., K. B. Low, and A. L Taylor. 1976.Recalibrated linkage map of Escherichia coli K-12.Bacteriol. Rev. 40:116-167.
3. Boulter, J., and N. Lee. 1975. Isolation of specializedtransducing bacteriophage lambda carrying genes ofthe L-srabinose operon of Escherichia coli B/r. J. Bac-teriol. 123:1043-1054.
4. Franklin, N., W. Dove, and C. Yanofsky. 1965. Thelinear insertion of a prophage into the chromosome ofE. coli by deletion mapping. Biochem. Biophys. Res.Commun. 18:910-923.
5. Kayajanlian, G. 1968. Studies in the genetics of biotin-transducing, defective variants of bacteriophage A. Vi-rology 36:30-41.
6. Lindahl, G., G. Sironi, IL Bialy, and R. Calendar.1970. Bacteriophage lambda; abortive infection of bac-teria lysogenic for phage P2. Proc. Natl. Acad. Sci.U.SAL 66:587-594.
7. Miller, J. H. 1972. Experiments in molecular genetics.Cold Spnng Harbor Laboratory, Cold Spnng Harbor,N.Y.
8. Nemoz, G., J. Robert-Baudouy, and F. Stoeber. 1976.Physiological and genetic regulation of the aldohexu-ronate transport system in Escherichia coli. J. Bacte-riol. 127:706-718.
9. Portalier, R., J. Robert-Baudouy, and G. Nemoz.1974. Etude de mutations affectant les genes de struc-ture de l'isom6rase uronique et de l'oxydor6ductasealtronique chez Escherichia coli K 12. Mol. Gen. Genet.128:301-319.
10. Portalier, R., J. Robert-Baudouy, and F. Stoeber.1972. Localisation genetique et caract6risation biochi-mique de mutations affectant le gene de structure del'hydrolyase altronique chez Escherichia coli K 12.Mol. Gen. Genet. 118:335-350.
11. Portalier, R., and F. Stoeber. 1974. La D-altronate:NAD-oxydoreductase d'Escherichia coli K 12: purifi-cation, proprietes et individualite. Eur. J. Biochem.26:50-61.
J. BACTERIOL.

SPECIALIZED TRANSDUCING PHAGES IN E. COLI 557
12. Portalier, R., and F. Stoeber. 1972. La D-mannonate:NAD-oxydoreductase d'Escherichia coli K 12: purifi-cation, propri6tes et individualite. Eur. J. Biochem.26:290-300.
13. Portalier, R., and F. Stoeber. 1972. Dosages colorimet-riques des oxydor6ductases aldoniques d'Escherichiacoli K 12: applications. Biochim. Biophys. Acta289:19-27.
14. Robert-Baudouy, J., RE Portalier, and F. Stoeber.1974. R6gulation du metabolisme des hexuronates chezEscherichia coli K 12: modalit6s de l'induction desenzymes du systeme hexuronate. Eur. J. Biochem.43:1-15.
15. Robert-Baudouy, J., and F. Stoeber. 1973. Purificationet proprietes de la D-mannonate hydrolyased'Escherichia coli K 12. Biochim. Biophys. Acta309:474-485.
16. Schrenk, W. J., and R. A. Weisberg. 1975. A simplemethod for making new transducing lines of coliphageA. Mol. Gen. Genet. 137:101-107.
17. Shapiro, J. A., and S. L Adhya. 1969. The galactose
operon of E. coli K 12. 1. A deletion analysis of operonstructure and polarity. Genetics 62:249-264.
18. Shimada, K., R. A. Weisberg, and KL E. Gottesman.1972. Prophage lambda at unusual chromosomal loca-tions. I. Location of the secondary attachment sitesand the properties of the lysogens. J. Mol. Biol.63:483-503.
19. Shimada K., R. A. Weisberg, and K. E. Gottesman.1973. Prophage lambda at unusual locations. II. Muta-tions induced by bacteriophage lambda in Escherichiacoli K 12. J. Mol. Biol. 80:297-314.
20. Sistrom, W. R. 1958. On the physical state of the intra-cellularly accumulated substrates of ,B-galactoside per-mease in Escherichia coli. Biochim. Biophys. Acta29:579-587.
21. Stoeber, F., A. Lagarde, G. Nemoz, G. Novel, M.Novel, R. Portalier, J. Pouysegur, and J. Robert-Baudouy. 1974. Le metabolisme des hexuronides etdes hexuronates chez Escherichia coli K 12: aspectsphysiologiques et genetique de sa regulation. Biochimie56:199-213.
VOL. 133, 1978