isolation dengue virus envelope glycoprotein membranes ...jvi.asm.org/content/18/1/132.full.pdf ·...
TRANSCRIPT

JOURNAL OF ViROLoGy, Apr. 1976, p. 132-140Copyright C 1976 American Society for Microbiology
Vol. 18, No. 1Printed in U.S.A.
Isolation of the Dengue Virus Envelope Glycoprotein fromMembranes of Infected Cells by Concanavalin A Affinity
ChromatographySTEPHEN A. STOHLMAN,l OLLIE R. EYLAR, AND CHARLES L. WISSEMAN, JR.*
Department of Microbiology, University of Maryland, School of Medicine, Baltimore, Maryland 21201
Received for publication 4 August 1975
The membranes isolated from type 2 dengue virus-infected BHK-21/15 cellscontain three glycosylated virus-specified proteins; one corresponds to the virionenvelope glycoprotein, V-3, whereas the other two are nonstructural virus-specified proteins, NV-2 and NV-3. A combination of two nonionic detergents,Nonidet P-40 and Triton X-305, solubilized .80% of the membrane-bound pro-tein and the majority of the type 2 dengue virus complement-fixing antigens.The soluble material was adsorbed by concanavalin A-Sepharose in the presenceof the nonionic detergents, which were subsequently removed by washing withdeoxycholate-containing buffer. Finally, the bound glycoprotein was eluted bythe addition of a-methyl glucopyranoside. V-3 was the only virus-specifiedprotein in the a-methyl glucopyranoside eluate. The V-3-containing fraction didnot cross-react with antisera against other selected Flaviviruses in the comple-ment fixation test. The V-3 contained in the isolated fraction differed from theparent membrane-bound V-3 in two interesting, and as yet unexplained, ways:(i) it exhibited hemagglutinating activity similar to that of the infectious virus,but (ii) it did not block the action of neutralizing antibody.
From an immunological point of view, theenvelope glycoprotein, V-3, of type 2 denguevirus (DEN-2) and other Flavivirus virions ap-pears to be the most important virion structuralcomponent because V-3 interacts with the pop-ulation of antibodies capable of neutralizingviral infectivity (9, 18). V-3 also functions as theviral hemagglutinin (9) and probably as theviral receptor for the cell surface (29). V-3 ofDEN-2 also occurs in the membranes of infectedhost cells, along with the nucleocapsid protein,V-2, and five additional nonstructural virus-specified proteins (24). Two ofthe latter are alsoglycoproteins.
Since large quantities of purified virus aredifficult to obtain, infected host cell membranesmight be an alternate, potentially rich, sourceof DEN-2 V-3. Affinity chromatography, espe-cially with concanavalin A (Con A) as ligand,has been used successfully to isolate the enve-lope glycoproteins from purified viruses (7, 12,20) and glycoprotein antigens from infectedcells (19) and cellular membranes (15). Thispaper reports the successful solubilization ofDEN-2 antigens from infected cell membraneswith nonionic detergents and the isolation of V-
IPresent address: Department of Neurology, McKibbonAnnex, Room 145, University of Southern California Schoolof Medicine, Los Angeles, Calif. 90033.
3 from these extracts by Con A-Sepharose affin-ity chromatography. In addition, some serologi-cal properties of the isolated V-3 are described.
MATERIALS AND METHODSVirus and cells. The procedure for the propaga-
tion, radiolabeling, and maintenance of the BHK-21/15 cells and the production, assay, passage his-tory, radiolabeling, and purification of DEN-2 aredescribed elsewhere (24). For this study, BHK cellswere cultured in 1,400-cm2 roller bottles (Bellco) at34 C, infected with a multiplicity of infection of 0.01to 0.05 PFU/cell, and harvested after 72 h of incuba-tion.
Preparation of cellular membranes. BHK cellu-lar membranes were isolated as previously de-scribed (24). Cells were suspended in 10 mM Trisbuffer, pH 7.4, containing 10 mM NaCl and 1.5 mMMgCl2 and were disrupted by 20 to 25 strokes of atight-fitting dounce homogenizer at 4 C. The homog-enate was centrifuged at 800 x g for 10 min toremove whole cells, nuclei, and large particulatematter. The membranes in the supernatant fluidwere separated from the soluble cytoplasmic compo-nents by centrifugation at 65,000 x g for 15 min at 4C. The pellet, containing the membrane material,was resuspended in 50 mM Tris buffer, pH 7.5, con-taining 50 mM NaCl and 1 mM MnCl2 (CAB) andwas stored at -20 C.
Detergent solubilization. For solubilization, 2 to3 mg ofBHK cellular membrane protein per ml (see
132
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

DEN-2 V-3 FROM HOST MEMBRANES 133
below) was placed in a loose-fitting dounce homoge-nizer at 37 C. Nonidet P-40 (NP-40) (Shell Oil Co.)and Triton X-305 (TX-305) (Rohm and Haas) wereeach added to a final concentration of 1%, and theresulting suspension was immediately mixed by ho-mogenization. After 30 min of incubation at 37 C,the remaining insoluble material was separatedfrom the detergent-soluble material (DSM) by cen-trifugation at 65,000 x g for 1 h at 4 C. Both theDSM and the insoluble material were stored at -20C until used.
Affinity chromatography on Con A-Sepharose.Con A-Sepharose (Pharmacia Chemicals) was pre-pared for use by placing the gel in a column, wash-ing successively with CAB, 0.2 M a-methyl gluco-side (a-MG), CAB until no further UV-absorbingmaterial eluted, and finally with approximately 4bed volumes of CAB containing 0.05% NP-40 and0.05% TX-305 (CAB-Det). In some experiments, theDSM was adsorbed directly to a prepared column ofCon A-Sepharose by holding the DSM within the gelbed for 2 h at 4 C before elution. In most experi-ments, however, the adsorption of glycoproteinsfrom the DSM was carried out by a combinationbatch-column procedure. The washed Con A-Sepha-rose was removed from the column, stirred with 1 to2 mg ofDSM protein per ml ofpacked gel beads for 1h at room temperature, and then stirred overnightat 4 C. The beads were then allowed to settle, theunbound DSM was decanted, and the fluid was re-placed with an equal volume of CAB-Det. The beadswere then repacked in the column before elution.When the experiments were monitored at 280 nm, abaseline absorption was established at optical den-sity at 280 nm = 0.20 prior to elution due to thepresence of the nonionic detergents. Elution wascarried out with 2 bed volumes of CAB-Det, 4 bedvolumes of 50 mM Tris buffer, pH 7.5, containing0.1% sodium deoxycholate (TDOC) to establish anew baseline (optical density at 280 nm = 0.00), 4bed volumes of 0.2 M a-MG in TDOC, and finallywith 3 bed volumes of 2.0 M NaCl in 50 mM Trisbuffer, pH 7.5.The peak fractions eluted by a-MG were pooled,
dialyzed 48 h at 4 C against 10 volumes of CAB withthree buffer changes, concentrated to 200 to 400 1Agof protein per ml by pressure dialysis (AmiconCorp.) at 4 C using a UM 10 filter, and stored at -20C. Radiolabeled eluates were concentrated by lyoph-ilization after dialysis.CF tests. Complement fixation (CF) tests were
performed by the microtiter technique (2a). The hy-perimmune mouse sera used were adsorbed in astepwise manner with BHK cellular membranesprepared from uninfected cells (total, 300 ,ug of pro-tein per ml of serum) as previously described (24).Adsorbed antiviral sera were not reactive in the CFtest with membranes prepared from uninfectedcells. For comparison among viruses, each adsorbedserum was first diluted to a CF titer of 1:128, with itshomologous CF antigen prepared from infectedsuckling mouse brain by the method of Hammonand Sather (6). The eluates from the affinity chro-matographic procedures were adjusted to a proteinconcentration of 150 1Lg/ml. The membrane prepara-
tion from infected BHK cells was adjusted to a pro-tein concentration of 300 ,g/ml. Block or "checker-board" titrations were performed in which serialdilutions of antisera were tested against serial dilu-tions of the antigen preparations.
Neutralizing antibody blocking tests. The neu-tralizing antibody blocking activity of the elutedglycoprotein material was determined by themethod of Qureshi and Trent (18). The test materialand serum were diluted in phosphate-buffered sa-line containing 20% heat-inactivated fetal calf se-rum and a-MG at a final concentration of 40 mM(15). An equal volume of virus-specific antiserum, ata final dilution capable of reducing the number ofDEN-2 PFU by 85 to 90%, was added, and the mix-ture was incubated for 1 h at 37 C. After addition ofvirus, the mixture was incubated for 1 h at 37 C, andresidual infectivity was assayed by plaquing onBHK cells. As controls for this test, antibody wasfirst reacted with a membrane preparation derivedfrom uninfected cells (negative control) or mem-branes derived from DEN-2-infected cells (positivecontrol) prior to addition of virus.HA and immunodiffusion tests. The hemaggluti-
nation (HA) test and the preparation of sucklingmouse brain-derived HA antigens were performedby the method of Hammon and Sather (6). Immuno-diffusion was carried out on glass slides using 1.0%agarose prepared in 0.1 M Tris buffer, pH 8.0, con-taining 0.3 M NaCl. Precipitable Con A was quanti-tated by immunodiffusion with anti-Con A rabbitserum (Miles Laboratories) as described by Ponce deLeon et al. (15).PAGE. Samples for polyacrylamide gel electro-
phoresis (PAGE) were adjusted to 1.0% sodium lau-ryl sulfate and 1.0% 2-mercaptoethanol, heated at100 C for 5 min, and then placed on 8% gels (7.0 by0.5 cm). Electrophoresis was carried out at 5 mA/geluntil the dye marker front had migrated 6.0 cm, aspreviously described (24). Gels were cut into 1-mmslices, which were incubated at 37 C overnight in 7.5ml of a mixture of toluene scintillation countingfluid and Soluene 350 (Packard Instruments) prior tothe determination of radioactivity.
Protein determination. Protein content was de-termined by the method of Lowry et al. (11). Thismethod was modified as described by Wang andSmith (26) for the determination of protein in thepresence of nonionic detergents. In all instances,bovine serum albumin was used as standard.
RESULTSDetergent solubilization. For isolation and
study it was necessary to prepare soluble virus-specific glycoproteins, which retained theirstructural and antigenic integrity. A variety ofdetergents were tested for their capacity to solu-bilize virus-specific proteins from cellular mem-branes. Some of the nonionic detergents thatsolubilized DEN-2 CF activity and resulted inlittle or no anticomplementary activity wereexamined more closely for recovery of CF activ-ity in a soluble form. Examination of the insol-
VOL. 18, 1976
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

134 STOHLMAN, EYLAR, AND WISSEMAN
uble material after treatment showsignificant amount of CF activity wa.bilized and the proportion of soluble Cwas related to the ability of the delsolubilize the membrane-bound protefore, nonionic detergents, both singcombination, were tested for their c;solubilize both total protein and DEIStivity. Two nonionic detergents, NP-4305, when used in combination, solubi85% of the membrane-bound protein.son of equal protein concentrations ofand insoluble material for antigenic cthe CF test revealed that both gave eequivalent antigen titers.
Electrophoresis of infected BHEmembranes indicated that the virwcproteins constituted the major specielabeled proteins and glycoproteins (F
6 k V-3
5
4
3
2
0
x
wI-
z
o(-z
0
0
FIGUR
DEN-2 VI
o-o 3H- AMII&- 14C - G LU
V-2
1U L0 30 40 50 60
GEL FRACTION
FIG. 1. Polyacrylamide gel electrophoresis ofDEN-2 virus (Fig. la) and BHK cellular membranes(Fig. I b) isolated from DEN-2-infected cells radiola-labeled with [3H]amino acids (0) and ['4C]-glucosamine (A).
ed that a Only one glycoprotein is found in the intacts not solu- virion (25) (Fig. la), whereas two additional'F activity nonstructural glycoproteins, NV-3 and NV-2,tergent to are found as membrane-bound, virus-specifiedin. There- proteins (Fig. lb). Treatment with NP-40 and,ly and in TX-305 solubilized 85 to 90% of the membrane-apacity to bound [3H]amino acids or [3H]leucine-labeled4-2 CF ac- proteins and 90 to 95% of the ['4C]glucosamineL0 and TX- label. The amount of ['4C]glucosamine-labeledilized 80 to glycolipid was estimated to be s10% by extrac-Compari- tion with chloroform-methanol (2:1). Compari-
f the DSM son of the DSM with the untreated membranescontent by derived from DEN-2-infected cells (Fig. 2)ssentially shows that all of the virus-specified proteins,
including the three virus-specified glycopro-I cellular teins, are rendered soluble by the combineds-specified treatment with NP-40 and TX-305. An addi-s of radio- tional protein, designated "Z" (Fig. 2), which'ig. 1) (24). migrated between V-3 (molecular weight,
59,000) and NV-3 (molecular weight, 48,500),was detected in the DSM and has a calculated
IE 1a molecular weight of 53,000. No Flavivirus-spec-ified proteins with a similar migration patternhave been described in the gel patterns of either
IRUS infected cells or cellular membranes (10, 21, 24,NO ACIDS 27, 28). A PAGE pattern similar to that of theCOSAMINE DSM was obtained by electrophoresis of the
nonionic detergent-insoluble material.Chromatography of DSM on Con A-Sepha-
MARKER rose. Typical elution patterns ofDSM preparedfrom DEN-2-infected BHK cellular membranesare shown in Fig. 3. The DSM eluted with CAB-
V-1 Det contained 95% of the total recoverable pro-
.7z e z
3
E lb
:MBRANES X - 3-S'IDS
w &--& 14C-UNTREATED INFECTEDIDS ~~i...2 MEMBRANES,MINE
z
DYE DYEMARKER Z MRE
U>
NV-i
10 20 30 40 50 60GEL FRACTION
FIG. 2. Polyacrylamide gel electrophoresis of[3H]amino acid-labeled material solubilized fromDEN-2-infected BHK cellular membranes (0)(DSM) and ['4C]amino acid-labeled untreated mem-branes from DEN-2-infected BHK cells (A).
J. VIROL.
In 11 Al ,21% A
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

DEN-2 V-3 FROM HOST MEMBRANES
CAB- Dot TDOC I|e -MG | < NaCI
PEAK I
PEAK 3
PEAK I
PEAK 3
PEAK 2
5 15 25 35 65 75 85 95 105 115 125 135
FRACTION NUMBER
FIG. 3. Elution profiles of the protein solubilized by detergent treatment of membranes from DEN-2-infectedBHK cells (DSM). Elution ofthe DSM was monitored by optical density at 280 nm (top) and with both[3H]amino acids (0) and ['4C]glucosamine (A)-labeled DSM (bottom). Column bed volumes varied from 5 to30 ml, and fractions of 1/10 bed volume were collected at a flow rate of 10 to 20 ml/h. All columns were
developed at 4 C. Arrows indicate the change of the elation solutions.
tein and radiolabel. The nonionic detergent inthe CAB-Det was in turn replaced with TDOCto substitute deoxycholate for the nonionic de-tergent mixture. This substitution was moni-tored at 280 nm and was completed before theaddition of a-MG so that the glycoproteineluate would contain a detergent more easilyremoved by dialysis. The nonionic detergentscould also be removed from the column by ex-tensive washing with CAB; however, in this
procedure the material that eluted after theaddition of a-MG was not recovered as a uni-form peak, indicating that the gel was interact-ing with the glycoproteins (15). The addition ofa-MG in TDOC resulted in a sharp peak ofeluted glycoprotein (Fig. 3).Due to the absorption at 280 nm by the non-
ionic detergents, the true protein elution pat-tern was monitored by use of radiolabeled pro-teins solubilized from DEN-2-infected mem-
VOL. 18, 1976
Eo 0.7CaI-,
I
16
X 12
a-x
,n806=C,
4
2
0
135
00
x
0Cal
4
3
2
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

136 STOHLMAN, EYLAR, AND WISSEMAN
branes. This elution pattern (Fig. 3) shows thatsmall amounts of 'H-labeled protein and['4C]glucosamine (' 1% of initial DSM) wereeluted by TDOC. To determine the nature ofthis material, it was rechromatographed on aCon A-Sepharose column equilibrated withTDOC. Deoxycholate does not prevent the bind-ing of glycoproteins by Con A (1); however, theTDOC eluate from the first column was notretained by adsorption on a second Con A-Seph-arose column equilibrated with TDOC. To de-termine if other proteins were bound to theSepharose matrix rather than to the ligand andfailed to elute with a-MG (14), columns werewashed with 2.0 M NaCl (Fig. 3). No additionalprotein or radiolabel was eluted.DSM prepared from uninfected BHK cellular
membranes was chromatographed on Con A-Sepharose to determine the amount of host cellmembrane protein that was adsorbed. All oftheprotein and radiolabel initially adsorbed to theCon A-Sepharose columns was eluted in thefirst peak, and no further material eluted aftertreatment by TDOC, a-MG, or NaCl. The totalrecovery in these and the previous experimentswas 290% of the input protein and radiolabel.
Serological characterization of the glyco-protein eluate. Membranes prepared from in-fected BHK cells (Fig. 4) have DEN-2 CF activ-ity when tested with serum adsorbed to removeantimembrane antibodies. Similar results wereobtained with DSM prepared from infectedmembranes, whereas uninfected membranesand DSM prepared from uninfected membranes
had no DEN-2 CF activity. The membranesderived from DEN-2-infected cells also cross-reacted with adsorbed hyperimmune mouse se-rum prepared against other Flaviviruses, aphenomenon often observed with crude Flavivi-rus antigen preparations. The glycoproteineluate, however, was found to be specific forDEN-2 virus by CF test (Fig. 4) when testedwith the adsorbed hyperimmune serum pre-pared against West Nile, Saint Louis encepha-litis, and Japanese B encephalitis viruses.When used as an immunogen, the mem-
branes derived from infected BHK cells wereable to elicit an antibody response in mice capa-ble of blocking HA and were also able to adsorbHA-inhibiting antibody. The membranes couldnot, however, hemagglutinate goose erythro-cytes. Both DEN-2 virus and a suckling mousebrain-derived DEN-2 HA antigen were used aspositive controls for this test. The a-MG eluate,which contained the envelope glycoprotein in asoluble form, was able to hemagglutinate gooseerythrocytes, exhibited the same pH optimum(pH 6.6) as the seed virus pool, and had an HAtiter of 16 units when tested at a protein concen-tration of 150 ug/ml.
In addition to the HA-inhibiting antibody,the antisera to DEN-2-infected cell membraneswere also reactive in the CF and plaque reduc-tion neutralization tests. Host cell cross-reac-tive antigens were not detected since there wasno diminution of either the neutralizing or CFtiters after adsorption of the serum with mem-branes prepared from uninfected cells. The a-
RECIPROCAL DILUTIONS OF IMMUNE MOUSE SERUM
* DEN-2 WN JBE SLEz , 4 , , 16, 64, 4 . 16. , 64 4 16 64
.....................1-.'..''.'
4 : 1 1N* IL........ fI......
zI-
z
*64z
40 ,,-'','>'... ..,. ..
0 1..i64
FIG. 4. CF analysis of the cross-reactivity of membranes from DEN-2-infected BHK cells and the a-MGCon A affinity column eluate. The hyperimrmune mouse sera tested were prepared against DEN-2, West Nile(WN), Japanese B encephalitis (JBE), and Saint Louis encephalitis (SLE) viruses. Before testing, the serawere adsorbed with membranes from uninfected BHK cells.
J. VIROL.
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
DEN-2 V-3 FROM HOST MEMBRANES
MG eluate, however, failed to block the actionof neutralizing antibody even in the presence ofa-MG. a-MG was added to the neutralizationmixture to reduce the possibility of any residualCon A interacting with the input virus (15).The a-MG eluate from infected membranescontained no host cell membrane antigens de-tectable by either CF or immunodiffusion usinghyperimmune mouse ascitic fluid preparedagainst uninfected BHK cell membranes. Im-munodiffusion utilized to test the a-MG eluatefor the presence of Con A (15) showed that itcontained less than 5 ,ug of precipitable Con Aper ml, and the addition of a-MG to the buffershad no effect on the titers obtained in either theCF or HA tests.
NV -4
FIGURE 5az
V3NV-30
0-5 3H-UNBOUND DSMX 2 NsNFNV-SIW h~14C-UNTREATED INFECTEI
MEMBRANES
z
I-~~~~~~~~~~-
0
0I
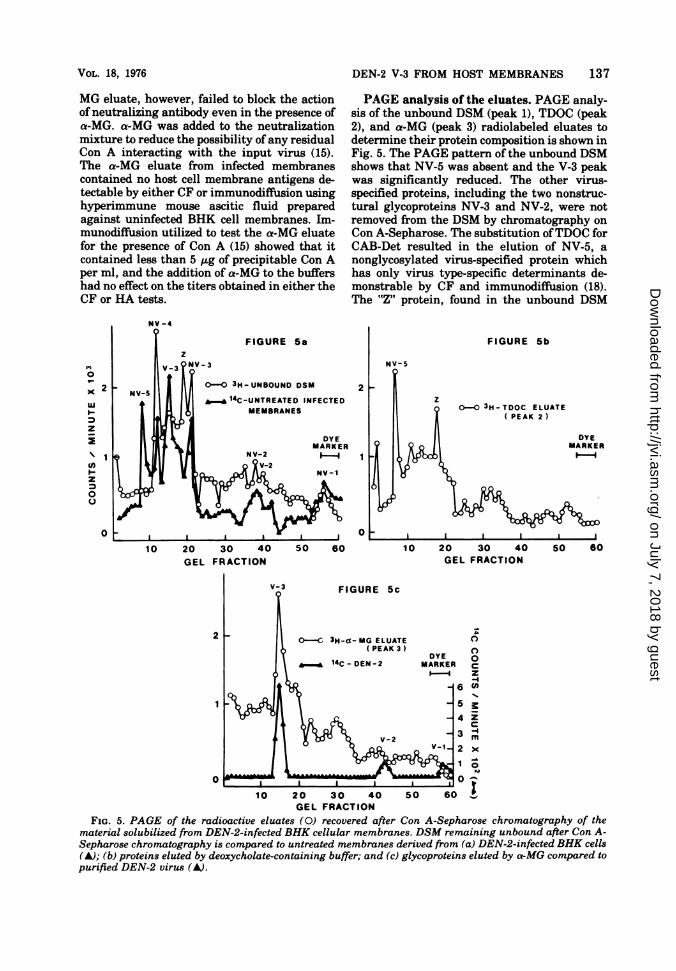
PAGE analysis of the eluates. PAGE analy-sis of the unbound DSM (peak 1), TDOC (peak2), and a-MG (peak 3) radiolabeled eluates todetermine their protein composition is shown inFig. 5. The PAGE pattern of the unbound DSMshows that NV-5 was absent and the V-3 peakwas significantly reduced. The other virus-specified proteins, including the two nonstruc-tural glycoproteins NV-3 and NV-2, were notremoved from the DSM by chromatography onCon A-Sepharose. The substitution ofTDOC forCAB-Det resulted in the elution of NV-5, anonglycosylated virus-specified protein whichhas only virus type-specific determinants de-monstrable by CF and immunodiffusion (18).The "Z" protein, found in the unbound DSM
2
0
FIGURE Sb
00o 3H-TDOC ELUATE( PEAK 2 )
DYEMARKERII-
50 60
2
1
0
FIGURE 5c
O-C 3H-ca-MG ELUATE( PEAK 3 )
14C-DEN-2DYE
MARKER
_ 6
_-4
_ 3V-1-2
1
0
0
0
C
z--4
z
-4
m
0i
10 20 30 40 50 60 "GEL FRACTION
FIG. 5. PAGE of the radioactive eluates (0) recovered after Con A-Sepharose chromatography of thematerial solubilized from DEN-2-infected BHK cellular membranes. DSM remaining unbound after Con A-Sepharose chromatography is compared to untreated membranes derived from (a) DEN-2-infected BHK cells(A); (b) proteins elated by deoxycholate-containing buffer; and (c) glycoproteins elated by o-MG compared topurified DEN-2 virus (A).
20 30 40 50 60 10 20 30 40GEL FRACTION GEL FRACTION
VOL. 18, 1976 137
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

138 STOHLMAN, EYLAR, AND WISSEMAN
(Fig. 2), although not detectably reduced in theeluted DSM (peak 1), was also detected in theTDOC eluate (peak 2). The addition of a-MG tothe Con A-Sepharose column resulted in theelution of a single major protein, which corre-sponds to the virus envelope glycoprotein, V-3.The gel pattern of the a-MG eluate (peak 3)shows a shoulder on V-3. This shoulder hasbeen demonstrated in the membranes of cellsinfected with DEN-2 and is believed to be a
result of the heterogeneity of V-3 (24).
DISCUSSION
Nonionic detergent solubilization and affin-ity chromatography using Con A as ligand werecombined for the isolation of DEN-2 antigensfrom the membranes of infected BHK cells.Treatment of the BHK cellular membraneswith the combination of nonionic detergents,NP-40 and TX-305, resulted in the solubiliza-tion of greater than 80% of both the membrane-bound protein and DEN-2 CF antigenic activ-ity. The detergent treatment solubilized allseven of the membrane-bound, virus-specifiedproteins, including the three membrane-boundglycoproteins. Coelectrophoresis of the DSMand untreated infected membranes showed thatthey had identical protein compositions, withthe exception of the protein designated "Z."
This protein has a molecular weight of 53,000and, in 8% polyacrylamide gels, migrates be-tween V-3 and NV-3. Although the origin ofthis protein has not been determined, we be-lieve that it is derived from one of the othervirus-specified proteins with a molecularweight greater than 53,000, i.e., V-3, NV-4, or
NV-5, since it is unlikely that an aggregate of alower-molecular-weight protein would be re-
fractory to the denaturing conditions used priorto PAGE.Although Con A is specific for only a few
closely related carbohydrate residues (5, 16, 22,23), it interacts with all of the enveloped vi-ruses thus far tested (2, 12, 13, 14), includingFlaviviruses (29; 0. R. Eylar et al., manuscriptin preparation). An example of this bindingspecificity is apparent from examination of theglycoprotein content of the DSM before andafter adsorption to Con A-Sepharose. Of thethree glycoproteins in the DSM, V-3, NV-3, andNV-2, only V-3 was shown to be adsorbed to theCon A-Sepharose, indicating that NV-3 andNV-2 do not possess exposed areas of glycosyla-tion capable of interaction with the ligand. V-3is presumably bound to the ligand since it iseluted by the addition of the saccharide a-MG.In addition to the adsorption of V-3, NV-5, a
nonglycosylated virus-specified nonstructural
protein, was also shown to be adsorbed by itsdepletion from the unbound DSM. NV-5 wasnot, however, bound to the Con A ligand, sinceit was eluted by the addition of the anionicdetergent deoxycholate. This nonspecific bind-ing was reversed either by the presence of theinorganic ions (15) or by the complete removalof the lipids from the hydrophobic portion oftheNV-5 molecule (3, 8). The TDOC eluate alsocontained the protein designated "Z," suggest-ing that this protein is heterogeneous sinceonly part of the total "Z" protein was bound tothe Con A-Sepharose.
Gel electrophoresis showed that the DEN-2envelope glycoprotein, V-3, was the major spe-cies of virus-specified protein isolated by Con A-Sepharose affinity chromatography. The isola-tion of Flavivirus envelope glycoproteins hasbeen accomplished by other techniques. Eckelset al. (4) have used isoelectric focusing for thepreparation of envelope glycoproteins from in-tact Japanese B encephalitis virions after gelfiltration chromatography, and Qureshi andTrent (17) isolated the V-3 proteins of four Fla-viviruses from infected cells by DEAE-cellulosechromatography after nonionic detergent solu-bilization. These isolated envelope glycopro-teins were cross-reactive in the CF test usingantisera prepared against other Flaviviruses(4, 18), and immunodiffusion analysis showedthat the preparation from infected cells ex-pressed both virus-specific and group-reactiveantigenic determinants (18).The hemagglutinating activity of purified
Flavivirus virions appears to reside in the enve-lope glycoprotein V-3, as does the immunogenthat reacts with neutralizing antibody (9, 29).The location, physical state, and prior treat-ment seem to influence certain biological andserological properties of this molecule, whichare not reflected by changes in PAGE charac-teristics as conventionally determined. Thus,cellular membranes of DEN-2 virus-infectedBHK cells, which contain substantial amountsof V-3 by PAGE analysis, do not agglutinategoose erythrocytes but possess the capacity toadsorb HA-inhibiting antibody activity fromDEN-2 immune serum and, when used as animmunogen, elicit the production of HA-in-hibiting antibodies in mice. However, when sol-ubilized from infected host cell membraneswith nonionic detergents and purified by ConA-Sepharose affinity chromatography, the hostcell membrane-derived, soluble V-3-containingfraction does exhibit hemagglutinating activity.Thus, it is possible that the portion of the V-3molecule responsible for HA is not exposed oravailable while it is still associated with hostcell membranes. This may explain the facts
J. VIROL.
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

DEN-2 V-3 FROM HOST MEMBRANES
that (i) certain treatments enhance the hemag-glutinating activity of crude infected tissuehomogenates commonly used as a source ofhemagglutinins for serological purposes (6) and(ii) tissue culture cells infected with denguevirus fail to exhibit the phenomenon of hemad-sorption even though the plasma membranecontains substantial quantities of V-3 demon-strable by PAGE analysis (24).Membranes from DEN-2 virus-infected cells,
when used as immunogens, induce the produc-tion of neutralizing antibody (Stohlman et al.,manuscript in preparation), presumably as aresult of the contained V-3 (18). These mem-branes are also capable of removing all DEN-2neutralizing antibody from solution. However,using inhibition of neutralization in the plaquereduction test, it was not possible to demon-strate the determinant, which combines withneutralizing antibody, in the V-3 fraction iso-lated from such infected membranes by the ConA-Sepharose method. Yet, sodium dodecyl sul-fate-polyacrylamide gel electrophoresis has de-tected no gross difference in molecular weightas indicated by migration properties betweenV-3 from the virion and V-3 directly from cellmembranes (24) or after isolation from cellmembranes by the Con A-Sepharose method.
Qureshi and Trent (17) have isolated fourforms of the envelope glycoprotein from St.Louis encephalitis virus. These forms werethought to represent developmental stages ofthe completed molecule; however, all were anti-genically identical by immunodiffusion analy-sis. Since -95% of all radioactivity was re-covered in each of our experiments, it is un-likely that a less reactive population of DEN-2V-3 was isolated by Con A affinity chromatog-raphy. Therefore, unless the "'Z" protein was de-rived from V-3, the vast majority of V-3 addedas DSM was recovered in the a-MG eluate.We have no precise explanation for the ap-
parent loss of antigenic activity of the V-3 iso-lated by the Con A-Sepharose method, i.e.,cross-reactivity in the CF test and blocking ac-tivity in the neutralization test. However, it ispossible that conformational changes may haveoccurred due to the loss of lipids by sequentialtreatment with nonionic and anionic detergents(3) or as a result of the interaction with Con A(29; 0. R. Eylar et al., manuscript in prepara-tion).NV-5 is also a viral-specific Flavivirus CF
antigen (18) and, under the conditions used inthis study, it may be possible to obtain excel-lent yields of both NV-5 and V-3. These prepa-rations would then be suitable for further stud-ies of their immunological as well as biophysi-cal-biochemical characteristics. Additionally,
their potential as specific Flavivirus diagnosticreagents could be evaluated.
ACKNOWLEDGMENTS
We wish to acknowledge Vernon Burnett for excellenttechnical assistance and to thank Lorraine Fiset andCharles Waechter for advice and helpful discussions.
This investigation received partial financial supportfrom contract no. DADA17-71-C-1008 with the U.S. ArmyMedical Research Command, Office of the Surgeon Gen-eral, Department of the Army. S. A. Stohlman was sup-ported as a predoctoral trainee by funds from the NationalInstititute of Allergy and Infectious Disease, training grantT01 AI 0016.
LITERATURE CITED1. Allan, D., J. Auger, and M. J. Crumpton. 1972. Glyco-
protein receptors for concanavalin A isolated from piglymphocyte plasma membrane by affinity chromatog-raphy in sodium deoxycholate. Nature (London) NewBiol. 236:23-25.
2. Calafat, J., and P. C. Hageman. 1972. Binding of con-canavalin A to the envelope of two murine RNAtumour viruses. J. Gen. Virol. 14:103-106.
2a. Casey, H. 1965. Standardized diagnostic complementfixation method and adaptation to micro test. InPublic Health Service Monogr. no. 74, Washington,D.C.
3. Coleman, R. 1973. Membrane-bound enzymes andmembrane ultrastructure. Biochim. Biophys. Acta300:1-30.
4. Eckels, K. H., F. M. Hetrick, and P. K. Russell. 1975.Virion and soluble antigens of Japanese encephalitisvirus. Infect. Immun. 11:1053-1060.
5. Goldstein, I. J., and L. L. So. 1965. Protein-carbohy-drate interaction. III. Agar gel-diffusion studies onthe interaction of concanavalin A, a lectin isolatedfrom jack bean, with polysaccharides. Arch. Bio-chem. Biophys. 111:407-414.
6. Hammon, W. M., and G. E. Sather. 1969. Arboviruses,p. 227-280. In E. H. Lennette and N. J. Schmidt (ed.),Diagnostic procedures for viral and rickettsial infec-tions. American Public Health Association, Inc., NewYork.
7. Hayman, M. J., J. Skehel, and M. J. Crumpton. 1973.Purification of virus glycoproteins by affinity chro-matography using Les culinaris phytohaemagglu-tinin. FEBS Lett. 29:185-188.
8. Helenius, A., and K. Simons. 1972. The binding ofdetergents to lipophilic and hydrophilic proteins. J.Biol. Chem. 247:3656-3661.
9. Kitano, T., K. Suzuki, and T. Yamaguchi. 1974. Mor-phological, chemical, and biological characterizationof Japanese encephalitis virus virion and its hemag-glutinin. J. Virol. 14:631-639.
10. Kos, K. A., D. Shapiro, Z. Vaituzis, and P. K. Russell.1975. Viral polypeptide composition of Japanese en-cephalitis virus-infected cell membranes. Arch. Vi-rol. 47:217-224.
11. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
12. Moennig, V., H. Frank, G. Hunsmann, I. Schneider,and W. Schafer. 1974. Properties of mouse leukemiaviruses. VII. The major viral glycoprotein of Friendleukemia virus. Isolation and physiochemical proper-ties. Virology 61:100-111.
13. Okada, Y., and J. Kim. 1972. Interaction of concana-valin A with enveloped viruses and host cells. Virol-ogy 50:507-515.
14. Oram, J. D., D. C. Ellwood, G. Appleyard, and J. L.
VOL. 18, 1976 139
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

140 STOHLMAN, EYLAR, AND WISSEMAN
Stanely. 1971. Agglutination of an arbovirus by con-
canavalin A. Nature (London) New Biol. 233:50-51.15. Ponce de Leon, M., H. Hessle, and G. H. Cohen. 1973.
Separation of herpes simplex virus-induced antigensby concanavalin A affinity chromatography. J. Virol.12:766-774.
16. Poretz, R. D., and I. J. Goldstein. 1970. An examina-tion of the topography of the saccharide binding sitesof concanavalin A and the forces involved in complex-ation. Biochemistry 9:2890-2896.
17. Qureshi, A. A., and D. W. Trent. 1973. Group B arbovi-rus structural and nonstructural antigens. I. Serolog-ical identification of Saint Louis encephalitis virussoluble antigens. Infect. Immun. 7:242-248.
18. Qureshi, A. A., and D. W. Trent. 1973. Group B arbovi-rus structural and nonstructural antigens. III. Sero-logical specificity of solubilized intracellular viralproteins. Infect. Immun. 8:993-999.
19. Ross, L. J. N. 1974. Comparison of antigenic glycopro-teins and glycoprotein receptors of concanavalin Aisolated from duck embryo cells infected with Marek'sdisease virus and a herpes virus of turkeys (StrainFC126). J. Gen. Virol. 24:549-562.
20. Scheid, A., and P. W. Choppin. 1974. The hemaggluti-nating and neuraminidase protein of a Paramyxovi-rus: interaction with neuraminic acid in affinity chro-matography. Virology 62:125-133.
21. Shapiro, D., K. Kos, W. E. Brandt, and P. K. Russell.1972. Membrane-bound proteins of Japanese enceph-alitis virus-infected chick embryo cells. Virology48:360-372.
22. Sharon, N., and H. Lis. 1972. Lectins: cell-agglutinat-ing and sugar-specific proteins. Science 177:949-959.
23. So, L. L., and I. J. Goldstein. 1967. Protein-carbohy-drate interaction. IV. Application of quantitativeprecipitin method to polysaccharide-concanavalin Ainteraction. J. Biol. Chem. 242:1617-1622.
24. Stohlman, S. A., C. L. Wisseman, 0. R. Eylar, and D.J. Silverman. 1975. Dengue virus induced modifica-tions of host cell membranes. J. Virol. 16:1017-1026.
25. Stollar, V. 1969. Studies on the nature of dengue vi-ruses. IV. The structural proteins of type 2 denguevirus. Virology 39:426-438.
26. Wang, C., and R. L. Smith. 1975. Lowry determinationof protein in the presence of Triton X-100. Anal. Bio-chem. 63:414-417.
27. Westaway, E. G. 1973. Proteins specified by group BTogaviruses in mammalian cells during productiveinfections. Virology 51:454-465.
28. Westaway, E. G. 1975. The proteins of Murray Valleyencephalitis virus. J. Gen. Virol. 27:283-292.
29. Yoshinaka, Y., and T. Shiomi. 1975. Agglutination ofJapanese encephalitis virus with concanavalin A. J.Virol. 15:671-674.
J. VIROL.
on July 7, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from