biological characteristics of clostridium perfringens type abiological characteristics of...
TRANSCRIPT

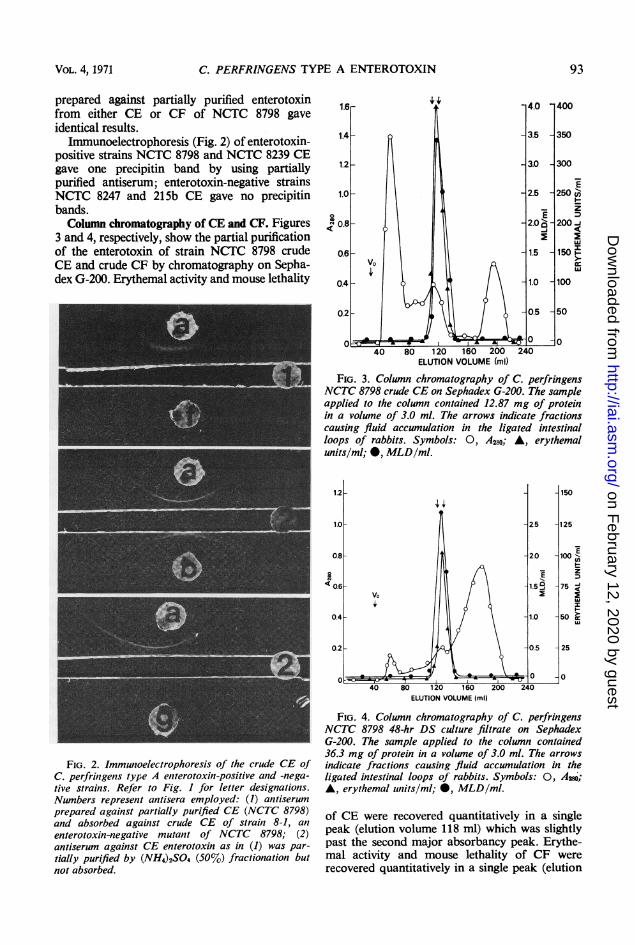
INFECTION AND IMMUNITY, Aug. 1971, p. 89-96Copyright © 1971 American Society for Microbiology
Vol. 4, No. 2Printed in U.S.A.
Biological Characteristics of Clostridium perfringensType A Enterotoxin
RAYMOND L. STARK AND CHARLES L. DUNCANFood Research Inslitute and Department of Bacteriology, University of Wisconsin, Madison, Wisconsin
53706
Received for publication 19 April 1971
An enterotoxin with the ability to induce fluid accumulation in rabbit ileal loops,erythema in the skin of guinea pigs, and lethality in mice appears in cell extracts (CE)and culture filtrates (CF) of sporulating cells of some Clostridiwn perfringens type Astrains. All activities in CE and CF were eluted simultaneously from a SephadexG-200 column. Different elution patterns were obtained for these activities presentin CE and CF. Rabbit immune serum against CF and the active CE fractions elimi-nated the three biological activities in CE and CF. These activities present in CF andCE were not eliminated by any of the known antitoxins present in diagnostic serumagainst C. perfringens types A, B, C, D, and E. Immunodiffusion studies with im-mune serum against active CE fractions and CF indicated a precipitin line of identitybetween CF and CE of NCTC 8798 and other enterotoxin-positive strains but notenterotoxin-negative strains. Disc electrophoresis of active G-200 fractions on7.0% polyacrylamide gels revealed a single area containing erythemal activity andmouse lethality. Immunodiffusion with acrylamide gels, containing crude fraction-ated enterotoxin, and immune serum against partially purified enterotoxin re-vealed a single precipitin band in the same area as the biological activities. Immuno-electrophoresis of CE of enterotoxin-positive and enterotoxin-negative strains alsoshowed one precipitin band which occurred only with enterotoxin-positive strains.These findings suggest that one component is responsible for the biological activitiesattributed to the enterotoxin.
Clostridium perfringens type A causes foodpoisoning in man characterized by diarrhea andabdominal pain (11). These symptoms have re-cently been reproduced in human and monkeysubjects by using culture filtrates (CF) andviable cells of C. perfringens type A enterotoxin-positive strains but not enterotoxin-negativestrains (6, 14). Viable cells, CF, and cell extract(CE) of enterotoxin-positive strains have previ-ously been shown to cause overt diarrhea andfluid accumulation in ligated intestinal loops ofrabbits and lambs (4, 5, 7, 9, 10). The enterotoxinis found early in CE of sporulating cells andlater in the CF when the cells are grown inDuncan-Strong (DS) sporulation medium (5).
Recently it has been shown that the partiallypurified enterotoxin found in CE causes erythemain the skin of guinea pigs and rabbits after intra-dermal injection (8). The enterotoxin of C. per-fringens type A is heat-sensitive, precipitable withammonium sulfate, antigenic, nondialyzable,and inactivated by :Pronase (5, 8). Confirmingwork has shown that the enterotoxin active ininducing fluid accumulation in intestinal loops of
lambs is associated with erythemal activity ofCE and CF of cells grown in vivo and in vitro(Hauschild et al., Can. J. Microbiol., in press).The results indicate that C. perfringens type Aenterotoxin is responsible for fluid accumulationand erythemal activity in guinea pigs. This allowsdirect assay of the enterotoxin based on erythemalactivity.
Clinical and systemic effects of the enterotoxinhave been determined by intravenous injection ofcrude CE into lambs, rabbits, and guinea pigs(13). Death occurred when the animals weresubjected to large doses of the extract.
This report shows that CE and CF of entero-toxin-positive strains also have the characteristicof being lethal to mice and presents evidence indi-cating that three biological activities, erythemalactivity, ileal loop activity, and lethality, at-tributed to the enterotoxin are due to a commonprotein.
MATERIALS AND METHODS
Cultures. The term rabbit-positive, or enterotoxin-positive, is used to designate strains of C. perfringenstype A, or CE or CF thereof, which repeatedly have
89
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

STARK AND DUNCAN
been shown to produce in young rabbits fluid ac-cumulation in the ligated ileum, overt diarrhea subse-quent to intraileal injection of the nonligated gut, orboth. Conversely, rabbit-negative or enterotoxin-negative denotes the absence of such a response in anyregular pattern.Two enterotoxin-positive strains, NCTC 8239 and
NCTC 8798, were employed. Enterotoxin-negativestrains included NCTC 8247, F42, FD1, 215b,ATCC 3624, and 8-1. Strain ATCC 3624 was origi-nally obtained from American Type Culture Collec-tion. Strain 8-1 is an asporogenous mutant of NCTC8798 obtained after mutagenesis of the wild type withacridine orange (Duncan, Strong, and Sebald,unpublished data). Details of original isolation of theother strains are given elsewhere (14).
Enterotoxin production. Lyophilized CE, CF (from24- or 48-hr-old cultures), or CF concentrated up to33X of all strains were prepared as previously de-scribed by employing DS sporulation medium (5).Both CF and CE were assayed for fluid-accumulationactivity in ligated rabbit ileal loops before testing forother activities.
Partial purification. Lyophilized CF (48 hr) ofstrain NCTC 8798 was reconstituted in distilled waterto a protein content of 45 mg/ml. A volume of 60 mlwas fractionated by ultrafiltration on a Diaflo XM100membrane (Amicon Corp.) having a molecular weightcut-off level of 100,000 by elution at 4 C with 650 mlof distilled water. The ultrafiltrate was then concen-trated by ultrafiltration on an XM50 membrane(cut-off level of 50,000 molecular weight). The mate-rial retained was subjected to diafiltration with a 10-fold volume of distilled water. The final filtrate had aprotein concentration of 5.01 mg/ml. A major portionof the enterotoxin was retained by the XM50 mem-brane, and this material was used as partially purifiedCF.
Antisera. Crude CE, crude CF, partially purifiedCF, and CE partially purified by Sephadex G-200column chromatography (see below) of strain NCTC8798 were used for immunization. Three-month-oldrabbits (New Zealand White) weighing 2.0 to 2.5 kgreceived intramuscular injections of a 1 :1 mixture ofthe challenge and Freund's complete adjuvant(Difco) at weekly intervals for 6 weeks, followed by1- to 2-week rest period. The volumes of the firstthree injections were 0.5 ml each; volumes were thenincreased to 1.0 ml for the subsequent injections. Atthis time animals were given a 1.0-ml intravenous in-jection without adjuvant.
After intravenous injection, immunized rabbitswere rested 2 weeks and then bled by cardiopuncture.Merthiolate, 1:10,000, was added to all immunesera which were subsequently stored at -20 C.
Clostridium diagnostic sera, C. perfringens (welchii)types A, B, C, D, and E, were obtained from Well-come Research Laboratories.
Animal assay. The rabbit ligated intestinal looptechnique was performed as previously described (7).
Erythemal activity was determined by using guineapigs weighing 300 to 400 g. Depilated areas weremarked in squares (2.5 cm2), and 0.05-ml sampleswere injected intradermally. The animals were ob-
served after 18 to 24 hr for erythema of the area. Con-siderable differences in degree of erythemal activityoften were noted with samples injected near the ven-tral side of the animal. Erythema in this area had atendency to become diffuse; therefore, use of this areashould be avoided in quantitating the erythemalactivity. Diameters of the areas of erythema weremeasured by using a vernier caliper. Areas of erythemabetween 0.5 and 1.2 cm in diameter were consideredroughly proportional to the concentration of theenterotoxin in the sample injected. Dilution of theunknown sample in saline was made to obtain areasof erythema with diameters between 0.5 and 1.2 cm.A unit of erythemal activity was defined as that
amount of enterotoxin producing an area of erythema0.8 cm in diameter under the conditions of the experi-ment. The activity of an unknown sample was alwayscompared to a standard injected into the same guineapig. The standard was a partially purified entero-toxin preparation containing 1,000 erythemal unitsper ml. The standard was stored in 1.0-ml amounts at-20 C.
All samples were injected in duplicate on the sameguinea pig, and results from duplicate guinea pigs wereaveraged.The higher phosphate concentrations in lyophilized
column fractions concentrated fivefold did not affectthe erythemal response.Mouse lethality was determined by intraperitoneal
injection of 0.5-ml portions of CF or CE preparationsinto each of two mice per sample. Mice (20 to 25 g)of either sex were used to determine lethality. Theanimals were observed at intervals for 72 hr to deter-mine lethality of the samples. The concentrated phos-phate of lyophilized column fractions did not affectthe lethality test. The mouse lethal dose (MLD)//ml isexpressed as the reciprocal of the highest dilutionwhich was lethal to two mice within 72 hr.
Column chromatography. Three milliliters of crudecell extract (24.9 mg of protein) and 3 ml of crudeculture ifitrate (45.4 mg of protein) of strain NCTC8798 were fractionated on a Sephadex G-200 column(2.5 by 35 cm) equilibrated with 0.02 M phosphatebuffer (pH 6.7) at a pressure of 13 cm. Fractions ofapproximately 6 ml were collected, and absorbancewas monitored at 280 nm. Accurate fraction volumeswere determined for each fraction before any assayswere performed. Fractions for injection into ligatedintestinal loops were dialyzed against pH 6.7, 0.002 Mphosphate buffer before concentrating 5X by lyophili-zation. It was necessary to decrease the phosphateconcentration since high phosphate concentrationalone elicited fluid accumulation in the ileal loop.Fractions from the column void volume were used asnegative controls, and crude CE diluted 1:6 was usedas the positive control. Protein determinations weremade by the method of Lowry et al. (12).
Neutralization of biological activity. Culture filtrateand CE at various concentrations in 0.85% NaCl weremixed 1:1 with immune sera of various dilutions.C. perfingens (welchii) diagnostic antitoxins A, B, C,D, and E were diluted 1:4 before mixing 1:1 withvarious' concentrations of the samples. All antigen-
90 INFEC. IMMUN.
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
C. PERFRINGENS TYPE A ENTEROTOXIN
antitoxin mixtures were incubated at 37 C for 30 minbefore animal assay.
Immunodiffusion and immunoelectrophoresis. Im-munodiffusion studies were performed on glass slides(3 by 1 inch, ca. 7.62 by 2.54 cm) with an agar layerconsisting of 1% Noble agar (Difco), 1% NaCl, and1:10,000 Merthiolate. Gelman immunodiffusionequipment was used for holding slides and cutting ofwells (Gelman Instrument Co.). Slides were incubatedfor 24 hr at 37 C in a humidity chamber prior toreading.
Immunoelectrophoresis was conducted by using theequipment and procedure recommended by GelmanInstrument Co. After completion of electrophoresisand addition of antiserum, slides were incubated in ahumidity chamber at 37 C and observed at 12 and24 hr.
Disc electrophoresis. Disc electrophoresis was per-formed by the method of Davis (2). The verticaltubes (0.5 by 7.5 cm) of acrylamide were loaded witha 30-liter sample and were run at 3 ma/tube for 1.5to 2 hr with bromophenol blue as a tracking dye. Theacrylamide separating gel was used at a concentrationof 7.0%, with tris(hydroxymethyl)aminomethane-glycine, pH 9.5, as the running buffer. Fractionatedproteins were stained with coomassie brilliant blue (1).To determine the protein band responsible for
erythema and lethal activity, the gels were slicedlaterally at 1.0-mm intervals after the electrophoreticrun. Each gel slice was eluted with saline for 5 hr at4 C, and the eluate was tested for biological activity.Immunodiffusion studies were also performed byusing the acrylamide gels containing fractionatedprotein. After the electrophoretic run, gels were imme-diately removed from gel tubes and placed in petridishes (100 by 500 mm) containing a total of 15 ml ofmolten agar (1% Noble agar, 1% NaCl). Two acrylam-ide gels per plate were positioned parallel to eachother and 2.1 cm apart. Upon solidification of the
agar, parallel trenches 2.0 mm in width were cut onboth sides and 1.0 cm from the center of each acrylam-ide gel. Antiserum was added to the trenches. Theplates were incubated at 37 C and observed for pre-cipitin bands after 12 and 24 hr.
RESULTSNeutralization of biological activity by immune
sera. CE and CF of C. perfringens type A entero-toxin-positive strains NCTC 8798 and NCTC8239 produced erythemal activity in the skin ofguinea pigs and were lethal to mice. The CE andCF of four of the five enterotoxin-negative strainsalso produced erythemal activity and necrosis insome instances in guinea pigs and lethality inmice. Strain NCTC 8247 had no activity. How-ever, the activities of the negative strains wereeliminated by one or more of the known C. per-fringens antitoxins but not by immune serumprepared against partially purified enterotoxin ofNCTC 8798 CE. The ability of the differentimmune sera to neutralize erythemal activity andmouse lethality of the CF of the strains employedis presented in Tables 1 and 2, respectively. Similarresults were also obtained with CE of thesestrains. The erythemal activity and mouse lethalityof enterotoxin-positive strains were neutralizedby immune serum prepared against partiallypurified NCTC 8798 CE and not by any of theknown C. perfringens types A, B, C, D, and Eantitoxins. Erythemal activity and mouse lethalityof enterotoxin-negative strains were neutralizedby types A, B, C, D, and E antitoxins but not byimmune serum against the NCTC 8798 CE.
Elimination of erythemal activity by immune
TABLE 1. Erythemal activity of culture filtrates: ability of immune sera to neutralize
Immune sera used for neutralizationb
A
NT
B
NT
++
c
NT
++
D
NT
E
NT
Strain
FDI
F42
ATCC 3624
215b
NCTC 8247NCTC 8239
NCTC 8798
Fold concnh
8.252.758.254.138.254.138.252.18.258.252.53.5
Ent
++++++++NT"
Saline control
+c
++
+
a Of 24-hr Duncan-Strong medium culture filtrate.bEnt, antiserum prepared against partially purified strain NCTC 8798 enterotoxin from G-200 column.
A, B, C, D, and E were Clostridium welchii diagnostic sera. See text for neutralization test procedure.c Erythemal activity remaining, +; no erythemal activity, -.J Not tested.
VOL. 4, 1971 91
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
STARK AND DUNCAN
TABLE 2. Mouse lethality of culture filtrates: ability of immune sera to neutralize
MLD/ml Immune sera used for neutralizationbStrain of original
culture filtratea Saline control Ent A B C D E
FDI 0.73 + - - - - -F42 0.48 + + - - - -ATCC 3624 1.46 + + - - - - -215b 0.97 + + _ _ _ - _NCTC 8247 0.0 - NTd NT NT NT NT NTNCTC 8239 2.18 + - + + + + +NCTC 8798 1.78 + _ + + + + +
a MLD, mouse lethal dose.b Ent, antiserum prepared against partially purified strain NCTC 8798 enterotoxin from G-200 column.
A, B, C, D, and E were Clostridium welchii diagnostic sera. See text for neutralization test procedure.c Mouse lethality remaining, +; no mouse lethality,-.d Not tested.
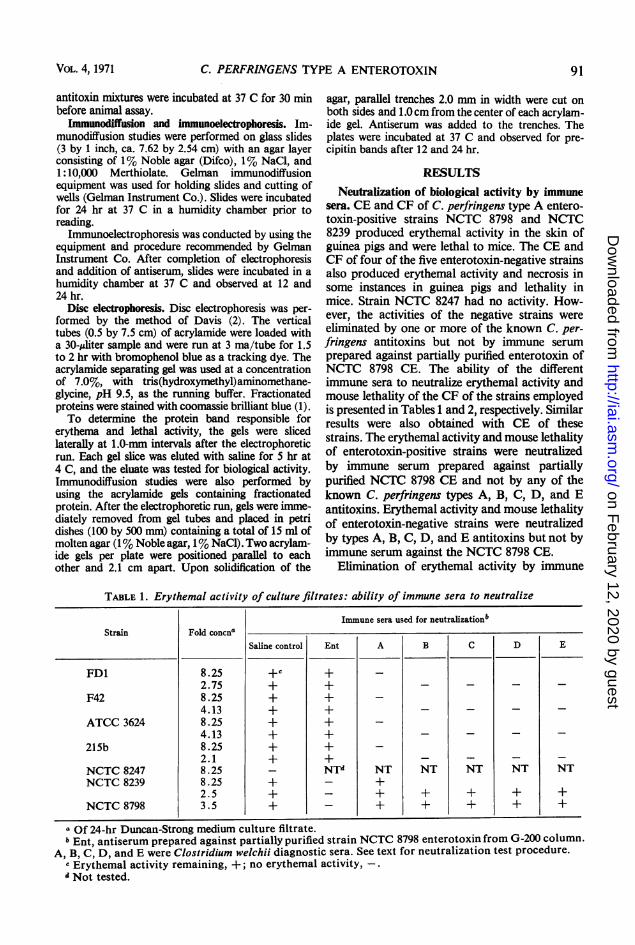
FIG. 1. Immunodiffusiont patternis obtained with enterotoxin-positive anzd -negative strains of C. perfrinzgenzs typeA. Antisera against NCTC 8798 enterotoxin is contained in the center wells with CE and CF of the different strainisin the outside wells. The number 8 antiserum was prepared against the partially purified enterotoxin found in CF anidthe number 9 was prepared against CE enterotoxin fractionated on Sephadex G-200. Inz the outter wells, numbersdesigniate CFand letters CE. Strains NCTC 8798 (1, a); NCTC 8239 (6, b); NCTC 8247 (7, f); ATCC 3624 (2, d);215b (5, g); F42 (3, e); and FDI (4, c) were used.
serum against partially purified NCTC 8798 CEor by heating at 60 C for 10 min also resulted inthe simultaneous elimination of mouse lethalityand the ileal loop fluid-accumulating activity ofCE and CF of enterotoxin-positive strains.
Serological relationships. Immunodiffusion
data presented in Fig. 1 show a precipitin line ofidentity between the CE of NCTC 8798 andNCTC 8239, the CF of these two strains, and alsobetween the CF and CE of each strain. Precipitinbands were not obtained with any of the entero-toxin-negative strains tested. Immune serum
92 INFEC. IMMUN.
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
C. PERFRINGENS TYPE A ENTEROTOXIN
prepared against partially purified enterotoxinfrom either CE or CF of NCTC 8798 gaveidentical results.
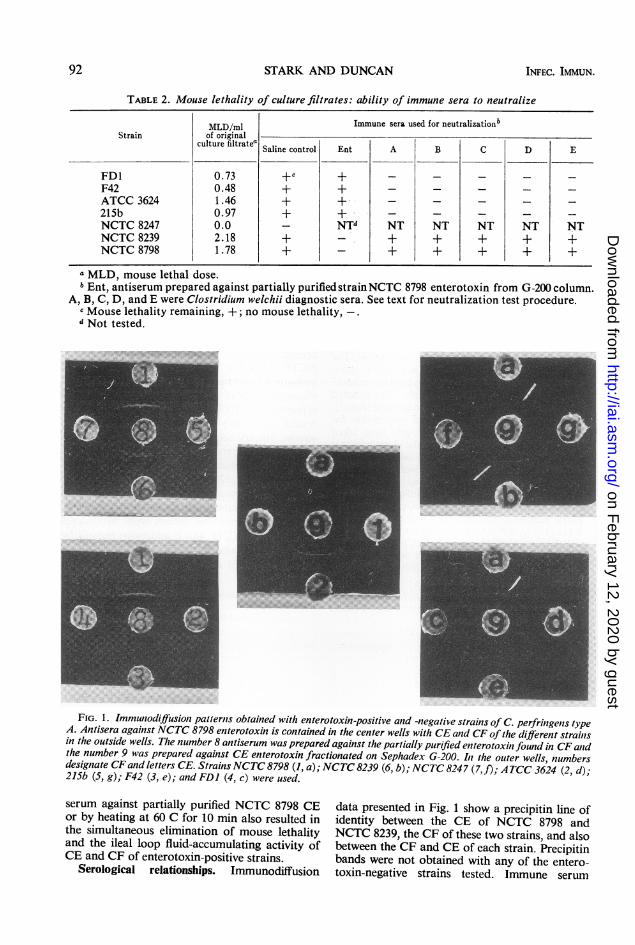
Immunoelectrophoresis (Fig. 2) of enterotoxin-positive strains NCTC 8798 and NCTC 8239 CEgave one precipitin band by using partiallypurified antiserum; enterotoxin-negative strainsNCTC 8247 and 215b CE gave no precipitinbands.Column chromatography of CE and CF. Figures
3 and 4, respectively, show the partial purificationof the enterotoxin of strain NCTC 8798 crudeCE and crude CF by chromatography on Sepha-dex G-200. Erythemal activity and mouse lethality
ELUTION VOLUME (ml)
FiG. 3. Column chromatography of C. perfringensNCTC 8798 crude CE on Sephadex G-200. The sampleapplied to the column contained 12.87 mg of proteinin a volume of 3.0 ml. The arrows indicate fractionscausing fluid accumulation in the ligated intestinalloops of rabbits. Symbols: 0, A2so; A, erythemalunits/ml; 0, MLD/ml.
FIG. 2. Immunoelectrophoresis of the crude CE ofC. perfringens type A enterotoxin-positive and -nega-tive strains. Refer to Fig. I for letter designations.Numbers represent antisera employed: (1) antiserumprepared against partially purified CE (NCTC 8798)and absorbed against crude CE of strain 8-1, an
enterotoxin-negative mutant of NCTC 8798; (2)antiserum against CE enterotoxin as in (1) was par-tially purified by (NH4)2SO4 (50%) fractionation butnot absorbed.
Q.8 2.0 100
z
0.2-0.5 25
0 040 80 120 180 200 240
ELUTION VOLUME (ml)
FIG. 4. Column chromatography of C. perfringensNCTC 8798 48-hr DS culture filtrate on SephadexG-200. The sample applied to the column contained36.3 mg of protein in a volume of 3.0 ml. The arrowsindicate fractions causing fluid accumulation in theligated intestinal loops of rabbits. Symbols: 0, A,20,;A, erythemal units/ml; 0, MLD/ml.
of CE were recovered quantitatively in a singlepeak (elution volume 118 ml) which was slightlypast the second major absorbancy peak. Erythe-mal activity and mouse lethality of CF wererecovered quantitatively in a single peak (elution
VOL. 4, 1971 93
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

STARK AND DUNCAN
volume 128 ml) coinciding with the second ab-sorbancy peak. Fractions having less than 25 to30 erythemal units per ml were concentrated 3 xby lyophilization before determining MLD/ml inmice.
Enterotoxin-containing fractions having ap-proximately 300 to 400 erythemal units per mgof protein were lethal to mice within 3 to 6 hr,and fractions with 200 to 300 erythemal units permg of protein were usually lethal in 24 to 72 hr.Little difference was noted between two identi-cally injected mice if high concentrations ofenterotoxin were present. Only those fractionshaving erythemal activity and mouse lethalityelicited fluid accumulation in the rabbit ileal loopupon 5 X concentration. These data confirm pre-vious work showing simultaneous elution oferythemal activity and ileal loop activity in CEon Sephadex G-100 columns (8).
Separation and identification of enterotoxin bypolyacrylamide disc electrophoresis. Further proofthat the same protein was responsible for erythe-mal activity, ileal loop fluid-accumulation ac-tivity, and mouse lethality was obtained by discelectrophoresis on 7.0% polyacrylamide gels ofpartially purified and crude CE of NCTC 8798.Figure 5 shows the patterns of protein bandsobtained with partially purified and crude CE.Upon lateral slicing of gels containing frac-tionated proteins and elution of the proteins fromthe gel slices, it was found that only one proteinband would elicit the erythemal response. Thisband was located 2.1 cm from the tracking dye atthe anode end. To obtain enough protein fromgels for a lethal dose, 50 Aliters (0.041 mg ofprotein) of partially purified enterotoxin andcrude CE (0.425 mg of protein) each were appliedto each of six gel tubes. One tube of each serieswas stained with coomassie brilliant blue toidentify protein bands, and the other five gelswere sliced laterally into 2-mm slices. The identi-cal slices were pooled, and the protein was elutedfor 5 hr at 4 C with 1.0 ml of saline per five pooledslices. Samples containing 250 uliters of partiallypurified enterotoxin (0.205 mg of total protein)or crude CE enterotoxin (2.12 mg of total pro-tein) were used as positive controls for thelethality test. Lateral slices from gel regionsshowing no erythemal activity were pooled,eluted, and used as negative controls. Only theprotein band possessing erythemal activity waslethal to mice.
Figure 6 shows the results obtained whenacrylamide gels containing fractionated proteinsof crude CE from strains NCTC 8798 andNCTC 8239 were subjected to immunodiffusion.When antiserum prepared against partiallypurified enterotoxin of CE NCTC 8798 (anti-
FIG. 5. Polyacrylamide disc electrophoresis ofpartially purified NCTC 8798 enterotoxin found incell extract (a) and crude cell extract (b). Arrowsindicate protein band having biological activity inguinea pigs and mice. Anode end was at the bottom ofthe gels.
serum 4) was used, two precipitin bands wereobtained with NCTC 8798 fractionated proteins;when the same antiserum was absorbed againstCE of enterotoxin-negative strain 8-1 (antiserum1), only one precipitin band was obtained withNCTC 8239. When the antiserum was subjectedto ammonium sulfate fractionation (antiserum2), only one band was obtained with the entero-toxin-positive strains. Only one precipitin bandwas obtained with both NCTC 8798 and NCTC8239 CE fractionated proteins when antiserumprepared against partially purified enterotoxin ofNCTC 8798 CF (XM 50 retained material) andabsorbed against CF of strain 8-1 was used(antiserum 3).The center of the precipitin band for both
strains was an identical 2.1-cm distance from theanode end of the gel as measured from the dyemarker. This was at the same location as thebiological activity (Fig. 5).The heavy diffuse band obtained with strain
NCTC 8239 would indicate more enterotoxin permilligram of protein applied than strain NCTC8798. Total protein concentrations applied to the
i.
7
i,ii
k
94 INFEC. IMMUN.
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
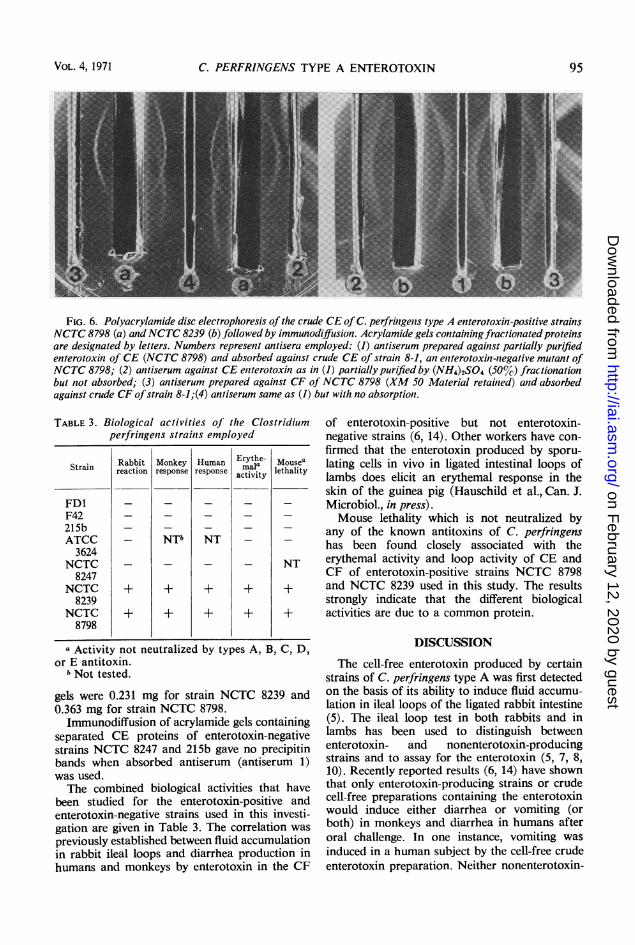
C. PERFRINGENS TYPE A ENTEROTOXIN
FIG. 6. Polyacrylamide disc electrophoresis of the crude CE of C. perfringenis type A enterotoxinl-positive strainsNCTC 8798 (a) and NCTC 8239 (b) followed by immunodiffusion. Acrylamide gels containingfraclionatedproteinsare designated by letters. Numbers represenit antisera employed: (1) antiserum prepared again1st partially purifiedenterotoxin of CE (NCTC 8798) anid absorbed againist crude CE of strain 8-1, an enterotoxin-negalive mutant ofNCTC 8798; (2) antiserum against CE eniterotoxin as in (1) partially purified by (NH4)2S04 (50%'7) fractionationbut not absorbed; (3) antiserum prepared against CF of NCTC 8798 (XM 50 Material retainted) anid absorbedagainst crude CF ofstrain 8-1;(4) antiserum same as (1) but with no absorption.
TABLE 3. Biological activities of the Clostridiumperfringenis strainis employed
Strain
FD1F42215bATCC3624
NCTC8247
NCTC8239
NCTC8798
Rabbit Monkey Human Erythe-reaction response response activity
+
NTb
+
NT
+ +
aActivity not neutralized by types A,or E antitoxin.bNot tested.
B, C, D,
gels were 0.231 mg for strain NCTC 8239 and0.363 mg for strain NCTC 8798.
Immunodiffusion of acrylamide gels containingseparated CE proteins of enterotoxin-negativestrains NCTC 8247 and 215b gave no precipitinbands when absorbed antiserum (antiserum 1)was used.The combined biological activities that have
been studied for the enterotoxin-positive andenterotoxin-negative strains used in this investi-gation are given in Table 3. The correlation was
previously established between fluid accumulationin rabbit ileal loops and diarrhea production inhumans and monkeys by enterotoxin in the CF
of enterotoxin-positive but not enterotoxin-negative strains (6, 14). Other workers have con-firmed that the enterotoxin produced by sporu-lating cells in vivo in ligated intestinal loops oflambs does elicit an erythemal response in theskin of the guinea pig (Hauschild et al., Can. J.Microbiol., in press).Mouse lethality which is not neutralized by
any of the known antitoxins of C. perfringenshas been found closely associated with theerythemal activity and loop activity of CE andCF of enterotoxin-positive strains NCTC 8798and NCTC 8239 used in this study. The resultsstrongly indicate that the different biologicalactivities are due to a common protein.
DISCUSSION
The cell-free enterotoxin produced by certainstrains of C. perfringens type A was first detectedon the basis of its ability to induce fluid accumu-lation in ileal loops of the ligated rabbit intestine(5). The ileal loop test in both rabbits and inlambs has been used to distinguish betweenenterotoxin- and nonenterotoxin-producingstrains and to assay for the enterotoxin (5, 7, 8,10). Recently reported results (6, 14) have shownthat only enterotoxin-producing strains or crudecell-free preparations containing the enterotoxinwould induce either diarrhea or vomiting (orboth) in monkeys and diarrhea in humans afteroral challenge. In one instance, vomiting wasinduced in a human subject by the cell-free crudeenterotoxin preparation. Neither nonenterotoxin-
VOL. 4, 1971 95
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

96 STARK ANI
producing strains nor their crude cell-freepreparations were active in monkeys or humans.The present results confirm the report that the
enterotoxin also possesses erythemal activitywhen injected into the skin of guinea pigs orrabbits (8) and further shows that it is lethal tomice after intraperitoneal challenge. Thus, ilealloop fluid-accumulating activity, erythemalactivity, and lethality are all biological charac-teristics of the enterotoxin active in monkeys andhumans. The three biological activities are foundin both CE and CF of the enterotoxin-producingstrains when they are grown in DS sporulationmedium (3) but not when they are grown in fluidthioglycolate medium. The relationship betweenproduction of the enterotoxin and sporulationwill be presented in a separate communication.The activities of the enterotoxin are similarlyheat-sensitive, nondialyzable, and neutralized byimmune sera prepared against the enterotoxin.All three activities appear simultaneously whenchromatographed on Sephadex G-200. However,slightly different elution profiles for the biologicalactivities present in CE and CF indicate possiblemolecular weight differences in the active proteinpresent in the two different preparations. Thisobservation is currently under further investi-gation.The biological activities of the enterotoxin were
not neutralized by any of the known antitoxinspresent in C. perfringens types A, B, C, D, or Ediagnostic sera. Although CF and CE of entero-toxin-negative strains were lethal to mice andhad erythemal activity that was frequently accom-panied by necrosis, these activities were neu-tralized by type A, B, C, D, and E diagnosticserum but not by immune serum prepared againstenterotoxin found in CE of strain NCTC 8798.The enterotoxin is obviously distinct from anyof the previously recognized major toxins of C.perfringens. The fact that the CE and CF ofnonenterotoxin-producing strains are usuallylethal and possess erythemal and necrotic ac-tivity emphasizes the need for the proper antiseracontrols when testing crude preparations for theseactivities.Immunodiffusion studies showed a precipitin
line of identity between the enterotoxin found inCE and CF of both enterotoxin-positive strainsand between the two strains when antiserumabsorbed against CE of an enterotoxin-negativestrain was employed. By using the same anti-serum, precipitin bands were not obtained withany of the enterotoxin-negative strains. Immuno-electrophoresis also showed one precipitin bandidentical to both enterotoxin-positive strains andno precipitin bands with enterotoxin-negativestrains.
Polyacrylamide disc electrophoresis of NCTC8798 CE followed by lateral slicing and elution of
DIDUNCAN INFEC. IMMUN.
the protein bands revealed that a single proteinband possessed both erythemal activity and mouselethal activity. Immunodiffusion studies with theacrylamide gels containing the CE fractionatedproteins showed one strong precipitin band in thesame region as the biological activity.These results strongly indicate that the biologi-
cal activities attributed to enterotoxin-containingpreparations that are active in inducing diarrheaand vomiting in humans and in monkeys, i.e.,ileal loop fluid accumulation, erythemal activity,and lethality, are due to a common protein.
ACKNOWLEDGMENTS
This research was supported by the College of Agricultural andLife Sciences, University of Wisconsin, Madison, by Public HealthService grant number I-ROI-CC-00554-01, 1-R01-FD-00203-01,and by contributions to the Food Research Institute by inemberindustries.
LITERATURE CITED
1. Chrambach, A., R. A. Reisfeld, M. Wyckoff, and J. Zaccari.1967. A procedure for rapid and sensitive staining of proteinfractionated by polyacrylamnide gel elecrophoresis. Anal.Biochem. 20:150-154.
2. Davis, B. J. 1964. Disc electrophoresis. II. Method and appli-cation to human serumn proteins. Ann. N.Y. Acad. Sci. 121:404-427.
3. Duncan, C. L., and D. H. Strong. 1968. Improved medium forsporulation of Clostridiurn perfringens. Appl. Microbiol.16:82-89.
4. Duncan, C. L., and D. H. Strong. 1969. Experimental pro-duction of diarrhea in rabbits by cell-free products of Clos-tridiu,nii perfiilngenis. Can. J. Microbiol. 15:765-770.
5. Duncan, C. L., and D. H. Strong. 1969. Ileal loop fluid ac-cumulation and production of diarrhea in rabbits by cellfree products of Clostridiuiti perfrin?gens. J. Bacteriol. 100:86-94.
6. Duncan, C. L., and D. H. Strong. 1971. Clostridiun, perfirin-geilts type A food poisoning. I. Response of the rabbit ileumas an indication of enteropathogenicity of strains of Clos-tridiurn perJiringenis in imionkeys. Infec. Immun. 3:167-170.
7. Duncan, C. L., H. Sugiyama, and D. H. Strong. 1968. Rabbitileal loop response to strains of Clostridiuin perfrinigens. J.Bacteriol. 95:1560-1566.
8. Hauschild, A. H. W. 1970. Erythemal activity of the cellularenteropathogenic factor of Clostridium perfrinigens type A.Can. J. Microbiol. 16:651-654.
9. Hauschild, A. H. W., L. Niilo, and W. J. Dorward. 1967. Ex-perimental enteritis with food poisoning and classical strainsof Clostridiuni perfrinlgelns type A in lambs. J. Infec. Dis.117:379-386.
10. Hauschild, A. H. W., L. Niilo, and W. J. Dorward. 1968.Clostridium perfringens type A infection of ligated intestinalloops in lambs. Appl. Microbiol. 16:1235-1239.
11. Hobbs, B. C., M. E. Smithi, C. L. Oakley, G. H. Warrack, antidJ. C. Cruickshank. 1953. Clostr idiun,z welehii food poisoning.J. Hyg. 51:75-101.
12. Lowry, 0. H., N. J. Rosenbrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with Folin phenolreagent. J. Biol. Chem. 193:265-275.
13. Niilo, L. 1971. Mechanism of action of the enlteropathogenicfactor of Clostridium perfringenis type A. Infec. Immun. 3:100-106.
14. Strong, D. H., C. L. Duncan, and G. Perna. 197 1. Clostridiul'perfringens type A food poisoning. II. Response of therabbit ileutm as an indication of enteropathogenicity ofstrains of Clostridiutn perfrin?gen?s in human beings. Infec.Immun. 3:171-178.
on February 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from