isolation andpartial characterization of fcy-binding...
TRANSCRIPT

Isolation and Partial Characterization of Fcy-BindingProteins of Human Leukocytes
ANTHONYKULCZYCKI, JR., LATA SOLANKI, and LEONARDCOHEN,Howard HughesMedical Institute and Department of Medicine, Division of Allergy andImmunology, Washington University School of Medicine, St. Louis, Missouri63110
A B S T RA C T Weisolated and partially characterizedthree Fe-binding macromolecules from human leuko-cytes. Mononuclear cells from normal individualsand from five patients with chronic lymphocytic leu-kemia and neutrophils from normal donors were sur-face radiolabeled by using 125I and lactoperoxidase.After detergent solubilization of the cells, Fc,-bind-ing macromolecules were purified by repetitive affinitychromatography under a variety of conditions andanalyzed by sodium dodecyl sulfate (SDS)-polyacryla-mide gel electrophoresis.
Three radiolabeled macromolecules were isolatedthat retained specific ability to bind to Fc fragments.A 52,000-64,000-mol wt macromolecule was isolatedfrom normal mononuclear and polymorphonuclearcells. A43,000-mol wt band was characteristic of mono-nuclear cells, particularly from patients with chroniclymphocytic leukemia. A 33,000-mol wt moleculecould be obtained from normal leukocytes under con-ditions that suggest it might be a proteolytic fragment.
INTRODUCTION
Cell surface receptors that bind the Fc portion ofIgG (Fc, receptors)' are present on a variety of cellsincluding lymphocytes (1-3), monocytes (4, 5), andneutrophils (1, 5). Fce receptors are importantlyinvolved in a variety of cell functions such as antibody-dependent cellular cytotoxicity (6) and the binding andphagocytosis of opsonized particles (7). Fc, receptors
Received for publication 15 May 1981 and in revised form28 July 1981.
1Abbreviations used in this paper: BBS, 0.2 Mborate, 0.15M NaCl, pH 8 buffer; BSA, bovine serum albumin; DNase,deoxyribonuclease I; Fc, receptors, receptors for the Fc por-tion of IgG; NP-40, Nonidet P40; PAGE, polyacrylamide gelelectrophoresis; PMSF, phenylmethyl sulfonyl fluoride;p33, 33,000 mol wt protein; p43, 43,000 mol wt protein; andp52-64, protein(s) in the 52,000- to 64,000-mol wt range.
1558
may also be involved in the regulation of immuneresponses (8) and in intracellular penetration ofautoantibodies (9).
Studies on the isolation and characterization of Fc.,-binding proteins2 have mainly used Fc receptor-bearing cells from experimental animals (10-15) orfrom cell lines (15-23) and numerous, apparently dif-ferent, molecules have been characterized. Rarelyhave Fc.-binding macromolecules been isolated fromhuman peripheral blood leukocytes, either from normalindividuals or from patients, and characterized by poly-acrylamide gel electrophoresis (PAGE) in sodiumdodecyl sulfate. One group has isolated both 120,000-and 60,000-mol wt Fc.,-binding proteins from normalhuman mononuclear cells (24) and human T lympho-cytes (25). But in other studies 28,000- or 30,000-mol wtFc,-binding proteins were purified from patients withchronic lymphocytic leukemia (26, 27).
In this study, we have used an efficient radioiodina-tion method (28) and a repetitive affinity chroma-tography procedure (12, 23) to isolate radiolabeled hu-man Fc.-binding macromolecules. Wehave comparedproperties of Fcy receptors from normal mononuclearcells with Fc, receptors from the mononuclear cells offive patients with chronic lymphocytic leukemia andwith Fcy receptors from normal neutrophils. We iso-lated a broad 52,000-64,000-mol wt band from the nor-mal individuals' mononuclear and polymorphonuclearcells. Wepurified a more distinct 43,000- mol wt bandfrom mononuclear cells of both normal individuals andpatients with chronic lymphocytic leukemia. A 33,000-mol wt band, which retained Fc,-binding ability, couldbe obtained from most cells under conditions thatsuggest that it might be a proteolytic fragment.
2In this paper we interchangeably refer to Fc,-bindingmacromolecules and Fc, receptors. Definitive evidencethat any isolated Fc,-binding macromolecule is a functionalFc, receptor has not yet been demonstrated.
J. Clin. Invest. ©D The American Society for Clinical Investigation, Inc. 0021-9738/81/1211558/08 $1.00Volume 68 December 1981 1558-1565

METHODS
Isolation of leukocytes. Peripheral venous blood of normaldonors, 400 ml, was collected into 100 ml of 0.15 M NaClcontaining 6% (wt/vol) dextran and 5,000 U of heparin. Fol-lowing dextran sedimentation of erythrocytes (at 37°C for 40min) the leukocyte-rich plasma was layered onto Ficoll-Hypaque density gradients (29). After centrifugation at 1,600 gfor 12 min, the interface (typically containing 70-85%lymphocytes and 10-25%i monocytes) was recovered and waswashed twice in 0.01 M phosphate-buffered saline, pH 7.4.Leukocytes that pelleted through the gradient (typically90-95% neutrophils) were similarly washed and contaminat-ing erythrocytes were removed by hypotonic (0.03 M NaCl)lysis for 20 s. An average of 4 x 108 mononuclear cells and 8x 108 polymorphonuclear cells were obtained from each donorand >95% of cells were viable.
Leukocytes were purified in a similar manner from theblood (10-50 ml) of five patients with chronic lymphocyticleukemia. In three experiments with normal donors, mono-cytes were depleted by incubating iron carbonyl particleswith the heparinized blood-dextran mixture at 37°C asdescribed (30) before erythrocyte sedimentation. In twoexperiments, fresh normal thymus tissue, obtained from a 6-mo-old patient and 18-mo-old patient undergoing open heartsurgery, were minced in RPMI 1640 media, and the cell sus-pensions were layered onto Ficoll-Hypaque gradients.Thymocytes isolated from the interface were >95% smalllymphocytes and were -60% viable.
Lymphocytes and neutrophils were identified by usingJenner-Giemsa staining of cytocentrifuged cells. Monocyteswere identified by using the 2,7-fluorenediamine procedure(31). In one experiment, cells that bound a mouse monoclonalanti-(human) monocyte antibody obtained from BethesdaResearch Laboratories (Bethesda, Md.) were identified byusing fluoresceinated antimouse antibody. In addition, es-terase-positive cells were detected by staining with a-naph-thyl acetate. Cell viability was assessed by exclusion of trypanblue dye.
Cell surface radioiodination. Cells were radiolabeledwith 125I using the modifications of the described lactoperoxi-dase method (28) except that each milliliter of iodination mix-ture contained 6 x 107 cells and 0.25 mCi of 1251. The washedradiolabeled cells were solubilized at 0°C in 0.01 M phos-phate, 0.15 M NaCl, pH 7.4 containing 1% (vol/vol) NonidetP40 (NP-40) and 2 mMphenylmethyl sulfonyl fluoride(PMSF) as described (12, 23) with or without such proteinaseinhibitors as: 3 mMEDTA, 10 mMdiisofluorophosphate,one-third trypsin inhibitory U/ml aprotinin, 1 AMpepstatin,1 mM1,10-phenanthroline, and 100 mMiodoacetamide. Aftercentrifugation at 30,000 g x 20 min at 4°C, the supernatantfluid was used for affinity purification.
Imnmunoadsorbents. The preparation of immunoad-sorbents with cyanogen bromide-activated Sepharose 2B wascarried out as described (12) using the following proteins: hu-man IgG, purified from fresh normal donor serum by (NH4)2-S04 precipitation and gel filtration or purchased from N. L.Cappel Laboratories Inc. (Cochranville, Pa.); human (Fab)'2,human Fc, and human IgM, all from N. L. Cappel Labora-tories Inc.; bovine serum albumin, egg albumin, and deoxy-ribonuclease I (DNase), all from Sigma Chemical Co. (St.Louis, Mo.). HumanIgG-Sepharose contained 7-20 mgof pro-tein per milliliter of packed beads; F(ab)'2-Sepharose had 5.9mg/ml beads; Fc-Sepharose had 5.6 mg/ml beads; and IgM/Sepharose had 2.7 mg/ml beads. Albumin-Sepharose immuno-adsorbents contained 4-10 mg of protein per milliliter ofbeads. DNase-Sepharose contained 3.3 mg/ml of beads andhad the capacity to bind at least 0.3 mg of rabbit actin (Sigma
Chemical Co.) per milliliter of immunoadsorbent. Before usethe immunoadsorbents were incubated in 0.2 Mborate 0.15 MNaCl buffer, pH 8 buffer (BBS) containing albumin (10 mg/ml) in order to minimize nonspecific binding (12) and werewashed in BBS containing 1% NP-40.
Purification of Fc, receptor. All steps in receptor purifi-cation were carried out in a 4°C cold room unless specificallynoted. Solubilized extracts of 3 x 108-2 x 109 radiolabeledcells were preincubated with 0.6 ml of an albumin-Sepha-rose immunoadsorbent for 30 min. The supernatant and a 1-ml wash with BBS-1% NP-40 containing the material un-bound to albumin-Sepharose were then incubated with0.6 ml of human IgG-Sepharose on a rotator overnight (unlessotherwise noted). This immunoadsorbent was transferredto a 0.7 x 4-cm glass column and washed with 24 ml of BBScontaining 1% NP-40 (12, 23). The Fc, receptors were ob-tained in their active state by using elution with 0.5 N aceticacid containing 1% NP-40 followed by very rapid neutraliza-tion with 2 M Tris, pH 8.6, containing 1% NP-40 as de-scribed (12, 23).
Assay of receptor activity. The Fc, receptor preparationseluted from human IgG-Sepharose were tested for bind-ing activity at 4°C by measuring the ability of radiolabeled re-ceptor to rebind to fresh human IgG-Sepharose. Nonspecificbinding was estimated both by using albumin-Sepharose as acontrol and by determining the rebinding to human IgG-Sepharose in the presence of an excess of soluble humanIgG. The assay was performed in plastic tubes as described(12, 23) except that human IgG-Sepharose was used. Thisassay system is analogous to the assay of Fc, receptor activity(32), which gave results comparable to two other independentmethods of measuring receptor activity. Tubes containinghighly purified Fc. receptor (receptor that had rebound tohuman IgG-Sepharose) were saved for elution with 1 ml of0.5 N acetic acid or 6 Mguanidine, both containing 1% NP-40(12) and eluted material was analyzed by SDS-PAGE. Toinvestigate specificity other immunoadsorbents were used,or potential inhibitors such as chicken IgG (N. L. CappelLaboratories Inc.) were mixed with human IgG-Sepharose be-fore the addition of labeled receptor.
Gel electrophoresis. SDS-PAGEwas performed accordingto the method of Laemmli (33) in gradient slab gels. Prepara-tion of the 6-18% linear gradient of polyacrylamide, of the 3%stacking gel, and of the samples for electrophoresis have beendescribed (34). Gels were fixed, stained, destained, and au-toradiographed as described (34) except that 50% (vol/vol)methanol, 10% (vol/vol) acetic acid. 3% (vol/vol) glycerol wasused before gel drying. Calibration of molecular weights wasperformed using described protein standards (34) as well asovalbumin (mol wt, 43,000), aldolase (mol wt, 40,000), andrabbit glyceraldehyde-3-phosphate dehydrogenase (mol wt,36,000).
RESULTS
Mononuclear cell Fc,-binding macromolecules.Three discrete radiolabeled Fcy-binding macromole-cules were isolated from human mononuclear cells ofnormal donors by using human IgG-Sepharose im-munoadsorbents (Fig. la). The apparent molecularweights of the Fc5-binding macromolecules were-33,000 (p33), 43,000 (p43), and 52,000-64,000(p52-64) upon analysis in SDS-polyacrylamide gels.The p52-64 band was extremely broad and heterodis-perse compared with the other radiolabeled bands.
Fc,-Binding Proteins from Human Leukocytes 1559

IO0K-
68K-
58K- 1
43K-KS *
40K-36K-
25 7K-
FIGURE 1 A composite of radioautographs illustrating gen-
eral properties of mononuclear cell Fcy-binding macromole-cules. (a) 125I-labeled Fcy-binding molecules obtained froma single IgG-Sepharose affinity column. Neutralized aliquotsfrom (a) were then incubated with: IgG-Sepharose (0.7 mg
IgG bound to 0.1 ml beads), (b) without, or (c) with additionof 3 mg (4.3-fold excess) of soluble IgG. (d) A neutralizedaliquot from (a) was incubated with BSA-Sepharose. Im-munoadsorbents were washed and their eluates were
analyzed by SDS-PAGE.In a separate experiment the solubilized material from
'25I-labeled mononuclear cells was incubated with IgG-Sepharose (e, f) at 4°C, or (g) at 22°C. Track (e) was obtainedfrom a slab gel run without mercaptoethanol, all other tracksin this and subsequent figures were from reduced SDS-gels.
When material eluted from IgG-Sepharose was neu-
tralized and reincubated with fresh IgG-Sepharose,identical bands were again isolated indicating thatsome of their ability to bind ligand (IgG) had been re-
tained (Fig. lb). Specificity for IgG was demonstratedbecause only soluble IgG inhibited rebinding of thesemacromolecules (Fig. lc, Table I below) and becausesuch macromolecules did not rebind to albumin-Sepharose (Fig. ld) or to F(ab)'2-Sepharose (Table Ibelow). Similar gel patterns were obtained whether or
not samples had been reduced with mercaptoethanolbefore SDS-polyacrylamide gel analysis (Fig. le, f).
The specificity of the affinity reagents was alsoassessed by comparing mononuclear cells with celltypes that lack Fc receptors. Human erythrocytes (intwo experiments) and human thymocytes (in two ex-
periments) were radiolabeled and solubilized inparallel with mononuclear cells. Three discrete radio-labeled macromolecules (as described above) were ob-
tained only from the mononuclear cells by using repeti-tive affinity chromatography. Little radioactivity and nodistinct bands were isolated from an equal number orfivefold greater number of labeled erythrocytes orthymocytes.
Quantitation of ligand-binding ability of mononu-clear cell Fc, receptor. The ability of eluted Fc, re-ceptor to specifically rebind to IgG was assayed byusing various protein-Sepharose conjugates. In 12experiments 52.2+4.0% (mean+SEM) of the radio-labeled Fc, receptor preparation bound to human IgG-Sepharose. By comparison, only 16.4+±2.2% of labeledreceptor bound to albumin-Sepharose (P < 0.005,paired t test). In 5 of these experiments an excess ofsoluble human IgG (3 mg) was mixed with IgG-Sepha-rose (0. 1-ml beads coupled to 0.7-0.8 mgIgG), and only18.4±2.5% of radiolabeled Fc, receptor was bound.
Soluble human IgG inhibited receptor binding in adose-dependent manner (Table I). Specificity wasdemonstrated because neither IgM, nor albumin, norchicken IgG inhibited receptor binding, nor did recep-tor preparations bind to F(ab)'2-Sepharose or IgM-Sepharose (Table I). Furthermore, nonquantitativestudies with mononuclear cell Fc,-binding macro-molecules in two experiments demonstrated that eachband bound to IgG-Sepharose and Fc,-Sepharose butnot to F(ab)'2-Sepharose (not shown).
Evidence that p33 may be a fragment of a largermolecule. The relative yields of the three Fc,-bind-ing molecules were related to the temperature at whichsolubilized mononuclear cell extracts and IgG-Sepha-rose were incubated. Whenthe incubation was carriedout overnight at 40C the p43 radiolabeled band wasmore prominent than the p33 band (Fig. lf). Whentheovernight incubation with immunoadsorbent was at220C the p43 band was much less evident than the p33band (Fig. lg). These findings suggested that the p33band might be a fragment of either or both of the largermacromolecules, presumably resulting from endoge-nous proteinase activity.
One alternative possibility, viz. that p33 and p43 areunrelated molecules, each with a maximal bindingaffinity at different temperatures, is excluded by ex-periments such as the one shown in Fig. 2. Fc,-bindingmacromolecules were isolated by using a single IgG-Sepharose column at 40C and then mixed with freshIgG-Sepharose at different temperatures. Both thep43 and p33 macromolecules were rebound to im-munoadsorbent at 40C (Fig. 2d) approximately as wellas at 22°C (Fig. 2e). However, when the mixture ofFc,-binding macromolecules and IgG-Sepharose alsoincluded a solubilized extract of unlabeled mononu-clear cells (to approximate the protease content of theoriginal solubilized cell suspension), less of the p43band and correspondingly more of the p33 band wereisolated at 4°C (Fig. 2b). This effect was extreme at
1560 A. Kulczycki, Jr., L. Solanki, and L. Cohen

TABLE IPercentage of Eluted Mononuclear Cell Fcy Receptor Rebinding to Immunoadsorbents*
Experiment no.Protein-Sepharose
conjugate Soluble proteins 1 2 3 4
mg protein
BSA-Sepharose 25.3±0.7 13.7±1.1 20.6±2.4 25.9±1.0IgG-Sepharose 56.4±0.9 50.8±9.9 67.6±1.3 66.3±2.2IgG-Sepharose IgG (3.0) 13.9±2.5 23.5±2.1IgG-Sepharose IgG (1.0) 25.9±2.0 33.0±1.5IgG-Sepharose IgG (0.3) 48.9±1.4IgG-Sepharose IgM (1.0) 65.4±4.2IgG-Sepharose BSA (3.0) 71.0±4.1IgG-Sepharose Chicken IgG (3.0) 67.7+3.9F(ab)'2-Sepharose 27.1±3.4IgM-Sepharose 24.6±4.1
* Mean values of triplicate determinations ±SD are given. The total radioactivity added to eachimmunoadsorbent in experiments 1-4 was 6,719, 1,646, 2,106, and 1,927 cpm, respectively.
22°C (Fig. 2c), such that the p43 band was barely evi-dent. In the latter case the isolated macromolecules re-sembled the Fcy-binding bands obtained from asingle immunoabsorbent that had been incubated withlabeled solubilized cell extracts only at 22°C (Fig. 2fand Fig. lg).
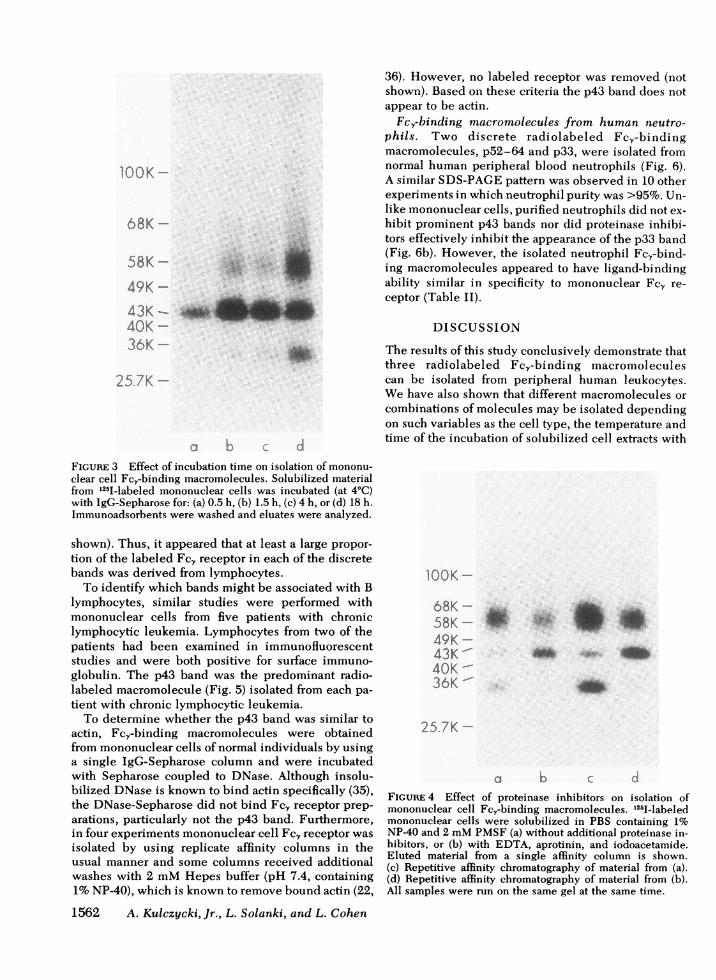
Another variable that affected the relative yield of thethree Fc.-binding macromolecules was the length oftime that solubilized cell extracts were incubated withIgG-Sepharose (Fig. 3). In the briefest incubationperiods (<4 h at 4°C) the p43 band appeared to beselectively isolated.
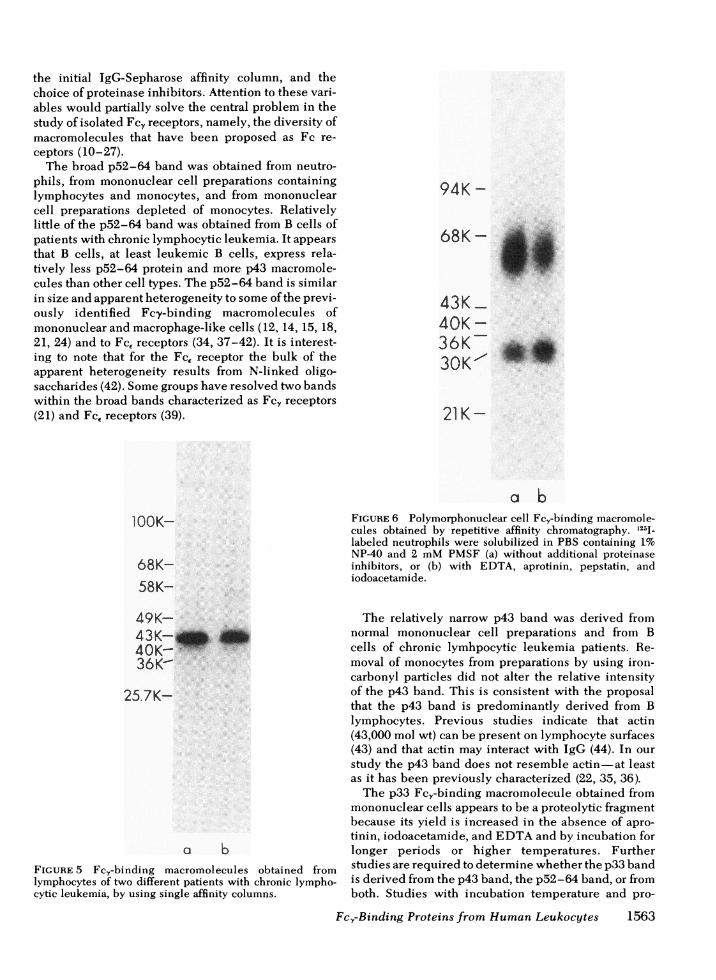
The inclusion of certain proteinase inhibitors in thesolubilization buffer allowed for the recovery of moreradioactivity in the p43 band than in the p33 band. Insix experiments, the solubilization step was evaluatedusing PBS 1% NP-40, 2mMPMSFwith or without acombination of aprotinin, iodoacetamide, and EDTA.As with lowering solubilization temperature, the pro-teinase inhibitors (especially the iodoacetamide) in-creased the amount of radiolabel associated with thep43 band while decreasing the label associated withp33 (Fig. 4). Additional proteinase inhibitors (phenan-throline, diisofluorophosphate, and pepstatin) did notfurther alter the observed pattern nor could any com-bination of inhibitors entirely avoid some copurifica-tion of the p33 band.
Fcy-binding macromolecules from human lympho-cytes. Because both lymphocytes and monocyteswere present in the "mononuclear cell preparations,"the cell type that produced the p43 and p52-64 bandswas not established. In three experiments, the periph-eral blood was divided into two equal samples, one ofwhich was incubated with iron carbonyl particles,which resulted in removal of >80% of the monocytesfrom mononuclear cell preparations. Then from both
samples, 125I-labeled Fc,-binding macromoleculeswere isolated. The amount of Fc, receptor and its SDS-PAGE pattern were similar from each sample (not
lOOK-
68K -58K-
49K-43K -
40K-36K-
4,
4,
25.7K-9
a b c d e fFIGURE 2 Effect of temperature on isolation of mononuclearcell Fc,-binding macromolecules. (a) 125I-labeled FcY-binding proteins obtained from a single affinity column (afterincubation overnight at 4°C). Neutralized aliquots from (a)containing partially purified Fce receptor were premixed (5min at 4°C) with: (b, c) 1 ml containing solubilized mono-nuclear cells (108 cell equivalents), or (d, e) 1 ml of solubili-zation buffer. These mixtures were then incubated with IgG-Sepharose overnight at 4°C (b, d), or at 22°C (c, e). Immuno-adsorbents were washed and their eluates were analyzed bySDS-PAGE. (f) For comparison, Fcy binding macromole-cules obtained from a single affinity column as in (a) exceptthat incubation was at 22°C instead of 4°C.
Fc,-Binding Proteins from Human Leukocytes 1561
36). However, no labeled receptor was removed (notshown). Based on these criteria the p43 band does notappear to be actin.
Fc,-binding macromolecules from human neutro-phils. Two discrete radiolabeled Fc,-bindingmacromolecules, p52-64 and p33, were isolated fromnormal human peripheral blood neutrophils (Fig. 6).A similar SDS-PAGEpattern was observed in 10 otherexperiments in which neutrophil purity was >95%. Un-like mononuclear cells, purified neutrophils did not ex-hibit prominent p43 bands nor did proteinase inhibi-tors effectively inhibit the appearance of the p33 band(Fig. 6b). However, the isolated neutrophil Fc,-bind-ing macromolecules appeared to have ligand-bindingability similar in specificity to mononuclear Fc, re-ceptor (Table II).
DISCUSSION
The results of this study conclusively demonstrate thatthree radiolabeled Fc,-binding macromoleculescan be isolated from peripheral human leukocytes.Wehave also shown that different macromolecules orcombinations of molecules may be isolated dependingon such variables as the cell type, the temperature andtime of the incubation of solubilized cell extracts with
FIGURE 3 Effect of incubation time on isolation of mononu-
clear cell Fc,-binding macromolecules. Solubilized materialfrom 125I-labeled mononuclear cells was incubated (at 4°C)with IgG-Sepharose for: (a) 0.5 h, (b) 1.5 h, (c) 4 h, or (d) 18 h.Immunoadsorbents were washed and eluates were analyzed.
shown). Thus, it appeared that at least a large propor-tion of the labeled Fc, receptor in each of the discretebands was derived from lymphocytes.
To identify which bands might be associated with Blymphocytes, similar studies were performed withmononuclear cells from five patients with chroniclymphocytic leukemia. Lymphocytes from two of thepatients had been examined in immunofluorescentstudies and were both positive for surface immuno-globulin. The p43 band was the predominant radio-labeled macromolecule (Fig. 5) isolated from each pa-tient with chronic lymphocytic leukemia.
To determine whether the p43 band was similar toactin, Fc,-binding macromolecules were obtainedfrom mononuclear cells of normal individuals by usinga single IgG-Sepharose column and were incubatedwith Sepharose coupled to DNase. Although insolu-bilized DNase is known to bind actin specifically (35),the DNase-Sepharose did not bind Fc, receptor prep-arations, particularly not the p43 band. Furthermore,in four experiments mononuclear cell Fcy receptor was
isolated by using replicate affinity columns in theusual manner and some columns received additionalwashes with 2 mMHepes buffer (pH 7.4, containing1%NP-40), which is known to remove bound actin (22,
1562 A. Kulezycki, Jr., L. Solanki, and L. Cohen
IOOK-
68K58K-49K-43K40K -36K
A.IKh .-.
25 7K -
a b
FIGURE 4 Effect of proteinase inhibitors on isolation ofmononuclear cell Fcy-binding macromolecules. 12,5-labeledmononuclear cells were solubilized in PBS containing 1%NP40 and 2 mMPMSF(a) without additional proteinase in-hibitors, or (b) with EDTA, aprotinin, and iodoacetamide.Eluted material from a single affinity column is shown.(c) Repetitive affinity chromatography of material from (a).(d) Repetitive affinity chromatography of material from (b).All samples were run on the same gel at the same time.
'00K -
68K -
58K -
49K -
43K-40K -
:36K --
5 7K-
d
.u,.
Wli
r* w
Amik"A&&", .-dmmkft
op -11
the initial IgG-Sepharose affinity column, and thechoice of proteinase inhibitors. Attention to these vari-ables would partially solve the central problem in thestudy of isolated Fc, receptors, namely, the diversity ofmacromolecules that have been proposed as Fc re-ceptors (10-27).
The broad p52-64 band was obtained from neutro-phils, from mononuclear cell preparations containinglymphocytes and monocytes, and from mononuclearcell preparations depleted of monocytes. Relativelylittle of the p52-64 band was obtained from B cells ofpatients with chronic lymphocytic leukemia. It appearsthat B cells, at least leukemic B cells, express rela-tively less p52-64 protein and more p43 macromole-cules than other cell types. The p52-64 band is similarin size and apparent heterogeneity to some of the previ-ously identified Fcy-binding macromolecules ofmononuclear and macrophage-like cells (12, 14, 15, 18,21, 24) and to Fc, receptors (34, 37-42). It is interest-ing to note that for the Fc, receptor the bulk of theapparent heterogeneity results from N-linked oligo-saccharides (42). Somegroups have resolved two bandswithin the broad bands characterized as Fcy receptors(21) and Fc, receptors (39).
lOOK-
68K-58K-
49K-43K- _40K-36K-
25.7K-
a bFIGURE 5 Fcy-binding macromolecules obtained fromlymphocytes of two different patients with chronic lympho-cytic leukemia, by using single affinity columns.
94K -
68K- *6_43K40K36K30K' S.
21 K-
a bFIGURE 6 Polymorphonuclear cell Fc,-binding macromole-cules obtained by repetitive affinity chromatography. 125j_labeled neutrophils were solubilized in PBS containing 1%NP-40 and 2 mMPMSF (a) without additional proteinaseinhibitors, or (b) with EDTA, aprotinin, pepstatin, andiodoacetamide.
The relatively narrow p43 band was derived fromnormal mononuclear cell preparations and from Bcells of chronic lymhpocytic leukemia patients. Re-moval of monocytes from preparations by using iron-carbonyl particles did not alter the relative intensityof the p43 band. This is consistent with the proposalthat the p43 band is predominantly derived from Blymphocytes. Previous studies indicate that actin(43,000 mol wt) can be present on lymphocyte surfaces(43) and that actin may interact with IgG (44). In ourstudy the p43 band does not resemble actin-at leastas it has been previously characterized (22, 35, 36).
The p33 Fc,-binding macromolecule obtained frommononuclear cells appears to be a proteolytic fragmentbecause its yield is increased in the absence of apro-tinin, iodoacetamide, and EDTAand by incubation forlonger periods or higher temperatures. Furtherstudies are required to determine whether the p33 bandis derived from the p43 band, the p52-64 band, or fromboth. Studies with incubation temperature and pro-
Fcr-Binding Proteins from HumanLeukocytes 1563

TABLE IIPercentage of Eluted Neutrophil Fc, Receptor Rebinding to Immunoadsorbents*
Experiment no.Protein-Sepharose
conjugate Soluble proteins 1 2 3 4
mg protein
BSA-Sepharose 12.5+0.6 12.2±2.0 16.3±2.4 24.4±3.2IgG-Sepharose 64.3+1.3 36.1±9.0 54.9±1.6 41.9+0.1IgG-Sepharose IgG (3.0) 12.6±0.9 19.9+0.2IgG-Sepharose IgG (1.0) 24.4±4.7 24.4±2.3IgG-Sepharose IgG (0.3) 33.3+1.7IgG-Sepharose IgM (1.0) 41.9±2.0IgG-Sepharose BSA (3.0) 41.1±2.4IgG-Sepharose Chicken IgG (3.0) 38.3+2.8F(ab)'2-Sepharose 16.6± 1.2IgM-Sepharose 18.5±+1.7* Mean values of triplicate determinations ±SD are given. The total radioactivity added to eachimmunoadsorbent in experiments 1-4 was 4,383, 21,146, 3,903, and 5,324 cpm, respectively.
teinase inhibitors (Figs. 2 and 4) suggest an inverse re-lationship between the p33 and p43 bands; however,little or no p33 band was obtained from chroniclymphocytic leukemia cells (Fig. 5). Fc,-bindingmacromolecules in the 33,000-mol wt region werealways obtained from polymorphonuclear cells.3Neutrophils contain a large amount and variety of pro-teolytic enzymes and presumably proteolysis occurseither before solubilization or in spite of proteinase in-hibitors during solubilization.
Wehave not isolated macromolecules smaller than33,000 daltons as reported in previous studies withmononuclear cells of normal donors (24) and chroniclymphocytic leukemia patients (26, 27). We suspectthat the previous studies may have proteolytically de-graded that Fc, receptor because cell lysates were in-cubated at 37°C (24, 26) or were subjected to freezingand thawing and overnight dialysis (27).
Several groups have used antigen-antibody precipi-tates or aggregated IgG coupled to Sepharose for isola-tion of Fcy receptors from human and animal cells, andthey obtain macromolecules in the 100,000-130,000-mol wt range (11, 13, 20, 24, 25). In those studies,however, "control" immunoprecipitates and aggre-gates formed with F(ab)'2 fragments may have beensignificantly smaller than those formed with IgG, allow-ing some doubt as to specificity of the putative recep-tor. Such large macromolecules have not been ob-tained when Fcy-binding macromolecules were puri-fied by using unaggregated IgG coupled to Sepharose(12, 18, 21, 23) or by using an antibody to Fc, recep-tor (15).
In this study specificity is documented because
3The present experiments do not establish whether the33,000-mol wt molecules from neutrophils are identical to the33,000-mol wt macromolecules obtained from mononuclearcells.
1564 A. Kulczycki, Jr., L. Solanki, and L. Cohen
radiolabeled Fc,-binding macromolecules bind toIgG-Sepharose and, in some studies, to Fc,-Sepha-rose but not to F(ab)'2-Sepharose nor to other immuno-adsorbents. Soluble IgG interacts with soluble recep-tor as assayed by inhibition of subsequent binding toimmunoadsorbents, but F(ab)'2 fragments, a non-mammalian IgG, and other proteins do not. Thisstudy demonstrates that three macromolecules that re-tain ligand binding activity for Fc, fragments can beisolated from human leukocytes and further investiga-tion is required to determine their functional sig-nificance.
ACKNOWLEDGMENTS
Wethank Dr. J. P. Atkinson and Dr. B. L. Hempstead for theirhelpful advice, Ms. Jane Gold for technical assistance, and Ms.Mamie Tomich for secretarial assistance.
This work was supported in part by National Institutes ofHealth grants RO1AI 16946, T32 AI 00112, and P50 AI 15322.
REFERENCES1. Uhr, J. W., and J. M. Phillips. 1966. In vitro sensitization
of phagocytes and lymphocytes by antigen-antibodycomplexes. Ann. N. Y. Acad. Sci. 129: 793-798.
2. Bianco, C., R. Patrick, and V. Nussensweig. 1970. A popu-lation of lymphocytes bearing a membrane receptor forantigen-antibody-complement complexes. I. Separationand characterization. J. Exp. Med. 132: 702-720.
3. Basten, A., J. F. A. P. Miller, J. Sprent, and J. Pye. 1972.A receptor for antibody on B lymphocytes. I. Method ofdetection and functional significance. J. Exp. Med. 135:610-626.
4. Huber, H., and H. H. Fudenberg. 1968. Receptor sites ofhuman monocytes for IgG. Int. Arch. Allergy Appl.Immunol. 34: 18-31.
5. Messner, R. P., and J. Jelinek. 1970. Receptors for humanyG globulin on human neutrophils. J. Clin. Invest. 49:2165-2171.
6. MacLennan, I. C. M. 1972. Antibody in the induction andinhibition of lymphocyte cytotoxicity. Transplant. Rev.13: 67-90.
7. Mantovani, B., M. Rabinovitch, and V. Nussenzweig.

1972. Phagocytosis of immune complexes by macro-phages. Different roles of the macrophage receptor sitesfor C3 and IgG.J. Exp. Mled. 135: 780-792.
8. Dickler, H. B. 1976. Lymphocyte receptors for immuno-globulin. Adv. I t7mt7un1ol. 24: 167-2 14.
9. Alarcon-Segovia, D., A. Ruiz-Arguelles, and E. Fishbein.1978. Antibody to nuclear ribonucleoprotein penetrateslive human mononuclear cells through Fc receptors.Nature (Lond.). 271: 67-69.
10. Rask, L., L. Klareskog, L. Ostberg, and P. A. Peterson.1975. Isolation and properties of a murine spleen cellFc receptor. Nature (Lond.). 257: 231-233.
11. Bourgois, A., E. R. Abney, and R. M. E. Parkhouse. 1977.Structure of mouse Fc receptor. Eur. J. Imnmunol. 7:691-695.
12. Kulczvcki, A., Jr., V. Krause, C. Chew Killion, and J. P.Atkinson. 1980. Purification of Fcy receptor fromrabbit alveolar macrophages that retains ligand-bindingactivity. J. Immt77utnol. 124: 2772-2779.
13. Sire, J., B. Kahn-Perles, A. Colle, and A. Bourgois. 1980.Biochemical characterization of an Fc receptor of rabbitlymphocytes. Eur. J. Im)71ut7ol. 10: 116-121.
14. Lane, B. C., J. Kan-NMitchell, NI. S. Mitchell, and S. M.Cooper. 1980. Structural evidence for distinct IgG sub-class-specific Fc receptors on mouse peritonealmacrophages. J. ExP. Med. 152: 1147-1161.
15. Mellman, I. S., and J. C. Unkeless. 1980. Purification ofa functional mouse Fc receptor through the use of amionoclonal antibody. J. Exp. Med. 152: 1048-1069.
16. Wernet, P., and H. G. Kunkel. 1975. Immunochemicalaspects of Ia type B cell membrane specificities (HL-B)as candidates for a human MILC stimulator antigen sys-tem. NIolecular heterogeneity and distinction from theFc receptors. In Histocompatibility Testing. F. Kissmeyer-Nielsen, editor. Munksgaard, Copenhagen, Denmark.731-734.
17. Cooper, S. NI., and Y. Sambray. 1977. Characterization ofthe Fc receptors of the murine leukemia L1210. J.Supramol. Struct. 6: 591-597.
18. Loube, S. R., T. C. McNabb, and K. J. Dorrington. 1978.Isolation of an Fc-binding protein from the cell mem-brane of a macrophage-like cell line (P388D,) after deter-gent solubilization. J. Immnunol. 120: 709-715.
19. D'Urso-Coward, M., and R. E. Cone. 1978. Membraneproteins of the P388Di macrophage cell line: isolation ofmembrane polypeptidles that bind to the Fc portion ofaggregated IgG. J. Imm71unol. 121: 1973-1980.
20. Kahn-Perles, B., J. Sire, A. Boned, and A. Bourgois. 1980.Putative conformation of mouse Fc,-receptor. J.Imtmt71unol. 125: 1360-1366.
21. Loube, S. R., and K. J. Dorrington. 1980. Isolation of bio-synthetically-labeled Fc-binding proteins from deter-gent lysates and spent culture fluid of a macrophage-likecell line (P388D,). J. Imrminunol. 125: 970-975.
22. Takacs, B. J. 1980. Characterization of functional Fcreceptor material from human lymphoblastoid cell lines.I. Large scale purification and biochemical analysis.Molecular Imtmt71unol. 17: 1293-1314.
23. Schneider, R. J., J. P. Atkinson, V. Krause, and A.Kulczycki, Jr. Characterization of ligand-binding activityof isolated murine Fc- receptor. J. Imtnunol. 126:735-740.
24. Cunningham-Rundles, C., Siegal, F. P., and R. A. Good.1978. Isolation and characterization of a human mono-nuclear cell Fc receptor. Immtunochemnistry. 15:365-370.
25. Cunningham-Rundles, C., D. Lawless, S. Gupta, C.Galanos, and R. A. Good. 1980. Isolation and partialchemical characterization of the IgG Fc receptors of hu-
man T lymphocytes and production of an antiserum.Proc. Natl. Acad. Sci. U. S. A. 77: 3645-3648.
26. Thoenes, J., and H. Stein. 1979. Properties of an Fc,-binding protein isolated from human leukemia B cells.
J. Exp. Med. 150: 1049-1066.27. Suzuki, T., T. Taki, K. Hachimine, and R. Sadasivan.
1981. Biochemical properties of biologically active Fc,receptors of human B lymphocytes. Molecular Immunol.18: 55-65.
28. Kulczycki, A., Jr., V. Krause, C. Chew Killion, and J. P.Atkinson. 1980. Improved cell surface radioiodination ofmacrophages.J. Immunol. Methods. 37: 133-138.
29. Eisen, S. A., H. J. Wedner, and C. W. Parker. 1972. Isola-tion of pure human peripheral blood T-lymphocytes usingnylon wool columns. Immunol. Comnmun. 1: 571-577.
30. Faguet, G. B. 1974. Lymphocyte purification: an im-proved method. Qualitative and quantitative evaluation.Biomedicine (Paris). 21: 153-157.
31. Benavides, I., and D. Catovsky. 1978. Myeloperoxidasecytochemistry using 2,7-fluorenediamine. J. Clin. Path.31: 1114-1116.
32. Kulczycki, A., Jr., B. L. Hempstead, S. L. Hofmann, E. W.Wood, and C. W. Parker. 1979. The cell surface receptorfor IgE. II. Properties of purified biologically activereceptor.J. Biol. Chem. 254: 3194-3200.
33. Laemmli, U. K. 1970. Cleavage of structural proteinsduring the assembly of the head of bacteriophage T4.Nature (Lond.). 227: 680-685.
34. Hempstead, B. L., C. W. Parker, and A. Kulczycki, Jr.1979. Characterization of the IgE receptor isolated fromhuman basophils. J. Immunol. 123: 2283-2291.
35. Lazarides, E., and U. Lindberg. 1974. Actin is thenaturally occurring inhibitor of deoxyribonuclease I.Proc. Natl. Acad. Sci. U. S. A. 71: 4742-4746.
36. Moosic, J. P., A. Nilson, G. J. Hammerling, and D. J.McKean. 1980. Biochemical characterization of Ia anti-gens.J. Imnmunol. 125: 1463-1469.
37. Conrad, D. A., and A. Froese. 1976. Characterization ofthe target cell receptor for IgE. II. Polyacrylamide gelanalysis of the surface IgE receptor from normal rat mastcells and from rat basophilic leukemia cells. J. Immunol.116: 319-326.
38. Kulczycki, A., Jr., T. A. McNearney, and C. W. Parker.1976. The rat basophilic leukemia cell receptor for IgE.I. Characterization as a glycoprotein. J. Imrnunol.117: 661-665.
39. Conrad, D. H., and A. Froese. 1978. Characterization ofthe target cell receptor for IgE. III. Properties of thereceptor isolated from rat basophilic leukemia cells byaffinity chromatography. J. Immunol. 120: 429-437.
40. Kulczycki, A., Jr., and C. W. Parker. 1979. The cell-surface receptor for immunoglobulin E. I. The use ofrepetitive affinity chromatography for the purification ofa mammalian receptor. J. Biol. Chem. 254: 3187-3193.
41. Kanellopoulos, J., G. Rossi, and H. Metzger. 1979. Pre-parative isolation of the cell receptor for IgE. J. Biol.Chem. 254: 7691-7697.
42. Hempstead, B. L., C. W. Parker, and A. Kulczycki, Jr.The cell surface receptor for IgE. Effect of tunicamycinon molecular properties of receptor from RBL-1cells.J. Biol. Chem. 256: 10717-10723.
43. Owen, M. J., A. Auger, B. H. Barber, A. J. Edwards, F. S.Walsh, and M. J. Crumpton. 1978. Actin may be presentoni the lymphocyte surface. Proc. Natl. Acad. Sci. U. S. A.75: 4484-4488.
44. Fechheimer, M., J. L. Daiss, and J. J. Cebra. 1979. Inter-action of immunoglobulin with actin. Molecular Im-munol. 16: 881-888.
Fc,-Binding Proteins from Human Leukocytes 1565