isolation and characterization of marine nonylphenol-degrading bacteria and description of...
TRANSCRIPT

Isolation and Characterization of Marine Nonylphenol-DegradingBacteria and Description of Pseudomaricurvus alkylphenolicusgen. nov., sp. nov.
Hiroaki Iwaki • Makoto Fujioka • Yoshie Hasegawa
Received: 30 April 2013 / Accepted: 30 July 2013 / Published online: 20 September 2013
� Springer Science+Business Media New York 2013
Abstract Two novel aerobic p-n-nonylphenol-degrading
bacterial strains were isolated from seawater obtained from
the coastal region of Ogasawara Islands, Japan. The 16S
rRNA gene sequence analysis indicated that the strains are
affiliated with the order Alteromonadales within the class
Gammaproteobacteria. One isolate, strain KU41G2, is
most closely related to Maricurvus nonylphenolicus (99.2 %
similarity), and is tentatively identified as M. nonylpheno-
licus. The other isolate, strain KU41GT, is also most closely
related to M. nonylphenolicus; however, the 16S rRNA
gene sequence similarity was only 94.7 %. Cells of strain
KU41GT are Gram-negative rods with a single polar fla-
gellum. The predominant respiratory lipoquinone was ubi-
quinone-8, and the major cellular fatty acids were C17:1 x8c
(24.2 %); C15:0 iso 2-OH; and/or C16:1 x7c (16.3 %), C15:0
(10.3 %), C11:0 3-OH (9.5 %), C9:0 3-OH (6.7 %), C10:0
3-OH (6.4 %), and C18:1 x7c (5.5 %). The DNA G?C
content was 53.3 mol%. On the basis of physiological,
chemotaxonomic, and phylogenetic data, strain KU41GT is
suggested to represent a novel species of a new genus, for
which we propose the name Pseudomaricurvus alkylphe-
nolicus gen. nov., sp. nov. The type strain of P. alkylphe-
nolicus is KU41GT (=JCM 19135T = KCTC 32386T).
Introduction
Alkylphenols and alkylphenol polyethoxylates are widely
distributed as pollutants in the environment from a variety
of industries, and contamination of the marine environ-
ment with alkylphenols is of great public concern because
of their toxicity and endocrine-disrupting activity in
humans and marine organisms [1]. Many alkylphenol-
degrading microorganisms have been isolated and char-
acterized [2, 3], and the mechanism of alkylphenol deg-
radation has been studied extensively [4–6]. However,
these organisms have been isolated mainly from terrestrial
or freshwater sites. Recently, we reported the isolation
and characterization of a marine p-n-nonylphenol-
degrading bacterium Maricurvus nonylphenolicus strain
KU41ET [7]; however, information regarding alkylphe-
nol-degrading bacteria from marine environments is rel-
atively scarce. Here, we report on the isolation of two
novel aerobic p-n-nonylphenol-degrading bacteria. Com-
parative 16S rRNA gene sequence analysis indicated that
one of the isolates, strain KU41GT, forms an independent
branch within the order Alteromonadales. Therefore, we
aimed to determine the exact taxonomic position of strain
KU41GT by a polyphasic characterization that included
phenotypic and chemotaxonomic properties and a detailed
phylogenetic analysis based on the 16S rRNA gene
sequence.
Materials and Methods
Strain Isolation
Two p-n-nonylphenol-degrading bacterial strains desig-
nated KU41GT and KU41G2 were isolated from seawater
collected from the coastal region of Chichi-jima Island of
Ogasawara Islands, Japan in August 2011, as described by
Iwaki et al. [7]. The strains KU41GT and KU41G2 were
routinely cultured on Marine Agar 2216 (MA; Becton–
H. Iwaki (&) � M. Fujioka � Y. Hasegawa
Department of Life Science and Biotechnology, Kansai
University, 3-3-35 Yamate-cho, Suita, Osaka 564-8680, Japan
e-mail: [email protected]
123
Curr Microbiol (2014) 68:167–173
DOI 10.1007/s00284-013-0455-x

Dickinson) and stored at -80 �C in the commercial arti-
ficial seawater (Marine Art SF-1; Tomita Pharmaceutical)
and Marine broth 2216 (MB; Becton–Dickinson), respec-
tively, with 10 % (w/v) glycerol.
16S rRNA Gene Sequencing and Phylogenetic Analysis
The 16S rRNA genes were amplified and sequenced as
described by Iwaki et al. [8]. Alignment was carried out
using the CLUSTAL W tool in MEGA version 5.1 [9].
Phylogenetic trees were generated using the neighbor-
joining [10], maximum-parsimony [11], and maximum-
likelihood [12] methods in MEGA version 5.1. The
distance matrix was produced based on Kimura’s 2-parameter
model [13], and the topologies of the resultant trees were
evaluated with bootstrap analysis [14] of 1,000 replicates.
Sequence similarity values were calculated using GEN-
ETYX-MAC version 16 (Genetyx Corporation).
Morphological, Physiological, and Biochemical
Characterization
The cell morphology of strain KU41GT was examined
under a transmission electron microscope (H-7600, Hit-
achi) at 100 kV. Motility was examined on a semisolid MB
medium [15]. Gram staining was performed using a Favor-
G kit (Nissui), and the cells were observed under a light
microscope (BX50F4, Olympus). Catalase and oxidase
tests were performed as described by Barrow and Feltham
[16]. Growth was tested at 25 �C on MA, unless otherwise
stated. Salinity requirements were tested using modified
MA, which had the same composition as MA, except for
the NaCl concentration, i.e., it was supplemented with
0–4 % (w/v) NaCl. The pH range for growth was deter-
mined on MA, and the pH was adjusted to 5.0–10.0. Sus-
ceptibility to antibiotics was determined using the diffu-
sion method with antibiotic disks (Becton–Dickinson), as
Pseudoteredinibacter isoporae SW-11T (FJ347760)Eionea nigra 17X/A02/237T (AY576771)
KU41G2 (AB817936)Maricurvus nonylphenolicus KU41ET (AB626730)Pseudomaricurvus alkylphenolicus KU41GT (AB809161)
Saccharophagus degradans 2-40T (AF055269)Teredinibacter turnerae T7902T (AY028398)
Gilvimarinus chinensis QM42T (DQ822530)Simiduia agarivorans SA1T (EF617350)
Marinimicrobium koreense M9T (AY839869)Marinimicrobium agarilyticum M18T (AY839870)
Melitea salexigens 5IX/A01/131T (AY576729)Spongiibacter marinus HAL40bT (AM117932)
Haliea salexigens 3X/A02/235T (AY576769)Dasania marina KOPRI 20902T (AY771747)
Porticoccus litoralis IMCC 2115T (EF468719)Microbulbifer maritimus TF-17T (AY377986)
Microbulbifer agarilyticus JAMB A3T (AB158515)Microbulbifer hydrolyticus DSM 11525T (AJ608704)
Zooshikella ganghwensis JC2044T (AY130994)Endozoicomonas elysicola MKT110T (AB196667)
Umboniibacter marinipuniceus KMM 3891T (AB467279)Spongiispira norvegica Gp 4 7.1T (AM117931)
Oceanospirillum beijerinckii IFO 15445T (AB006760)Oceanospirillum linum ATCC 11336T (M22365)Neptuniibacter caesariensis MED92T (AY136116)
Amphritea balenae JAMM 1525T (AB330883)Neptunomonas japonica JAMM 0745T (AB288092)
Pseudomonas aeruginosa ATCC 10145T (AF094713)Cellvibrio japonicus NCIMB 10462T (AF452103)
Cellvibrio mixtus ACM 2601T (AF448515)Cellvibrio fulvus NCIMB 8634T (AF448514)
Escherichia coli ATCC 11775T (X80725)
0.01
100
99
99
89
99
100
99
99
100
100
100
97
77Fig. 1 Phylogenetic
relationships between strain
KU41GT and other members of
the class Gammaproteobacteria.
The tree was constructed using
the neighbor-joining algorithm.
Numbers at nodes are bootstrap
percentages based on 1,000
replications; only values[70 %
are shown. Filled circles
indicate that the corresponding
nodes were also recovered in
trees generated with the
maximum-likelihood and
maximum-parsimony
algorithms. Open circles
indicate that the corresponding
nodes were also recovered in the
tree generated with the
maximum-likelihood algorithm.
Bar 0.01 substitutions per
nucleotide position
168 H. Iwaki et al.: Pseudomaricurvus alkylphenolicus gen. nov., sp. nov.
123

described by Iwaki et al. [7], and the cell suspension was
adjusted to a 2 McFarland standard. Nitrate reduction,
indole production, acid production from glucose, hydro-
lysis of esculin and gelatin, and the presence of arginine
dihydrolase, urease, and b-galactosidase were tested using
the API 20NE (bioMerieux), according to the manufac-
turer’s instructions, except that the cell suspensions were
prepared using Marine Art SF-1. The results were obtained
after 24 h of incubation at 25 �C. Hydrolysis of casein,
Tween 40, and Tween 80 were tested on MA, using the
substrate concentrations described by Cowan and Steel
[17]. Other enzyme activities were also assayed using API
ZYM (bioMerieux) at 37 �C, according to the manufac-
turer’s instructions. Utilization of various carbon sources
was tested using the commercial artificial seawater med-
ium, Daigo’s IMK-SP (Nihon Pharmaceutical), containing
0.1 % carbon source. Alkylphenol degradation was tested
using solidified Daigo’s IMK-SP, containing 0.1 % alkyl-
phenol; formation of a clear zone around the growing
colonies was considered a positive result.
DNA G?C Content, Isoprenoid Quinone, and Fatty
Acid Analysis
The G?C content of the genomic DNA was determined by
the method used by Katayama-Fujimura et al. [18], using a
high-performance liquid chromatograph (HPLC; Nihon
Waters). Isoprenoid quinones were extracted from freeze-
dried KU41GT cells grown in MB for 4 days at 25 �C
according to the method devised by Nishijima et al. [19]
and analyzed using HPLC (Nihon Waters). Cellular fatty
acids from cells grown on MA for 3 days at 25 �C were
extracted, methylated, and analyzed by gas chromatogra-
phy (Agilent Technologies 7890A GC System equipped
with a HP Ultra2 column) according to the standard pro-
tocol of the Sherlock Microbial Identification System
(MIDI) version 6.0, and identified using the TSBA40
method and TSBA40 library.
Results and Discussion
Phylogenetic Analysis
The nearly complete 1,458 bp 16S rRNA gene sequences
of strains KU41GT and KU41G2 were determined and
deposited in DNA Data Bank of Japan under accession
numbers AB809161 and AB817936, respectively. The 16S
rRNA gene sequence analysis indicated that strains
KU41GT and KU41G2 are phylogenetically affiliated with
the order Alteromonadales within the class Gammaprote-
obacteria. Strain KU41G2 is most closely related to M.
nonylphenolicus with 99.2 % similarity and is tentatively
identified as M. nonylphenolicus. Strain KU41GT is also
most closely related to M. nonylphenolicus; however, the
similarity was only 94.7 %, and the strain forms a distinct
lineage within the order Alteromonadales (Fig. 1). Strain
KU41GT was also found to be related to Pseudoteredi-
nibacter isoporae SW-11T (93.6 % similarity), Teredinib-
acter turnerae T7902T (92.7 %), Eionea nigra 17X/A02/
237T (92.5 %), and Saccharophagus degradans 2-40T
(92.0 %). Therefore, strain KU41GT should be classified as
a novel genus and species in the order Alteromonadales
considering the phylogenetic analyses.
Phenotypic Characteristics
The cells of strain KU41GT were Gram-negative, aerobic,
curved rods (1.5–2.0 lm in length and 0.7–1.0 lm in
width), and motile by a single polar flagellum (Fig. 2), as
with the members of the order Alteromonadales [20]. The
KU41GT cells formed colonies that were pale yellow,
circular, smooth, convex, 1.0–2.0 mm in diameter, and
with an entire margin after 3-days incubation on MA.
Growth occurred at 15–30 �C (optimally at 25–30 �C), at
pH range of at least 7.0 to 10.0 (optimally at pH 8.0), and
with 2.0–3.0 % NaCl. The strain degraded p-n-nonylphe-
nol, p-n-octylphenol, and p-n-heptylphenol, and utilized
these substrates as sole sources of carbon and energy. The
strain did not utilize p-n-hexylphenol, p–t-octylphenol, or
p-nonylphenol (a mixture of branched-chain isomers). The
other phenotypic properties of strain KU41GT are stated in
the genus and species descriptions and those characteristics
that differentiate strain KU41GT from phylogenetically
related taxa are listed in Table 1.
Fig. 2 Transmission electron micrograph of KU41GT cell grown on
MA. Bar 0.5 lm
H. Iwaki et al.: Pseudomaricurvus alkylphenolicus gen. nov., sp. nov. 169
123
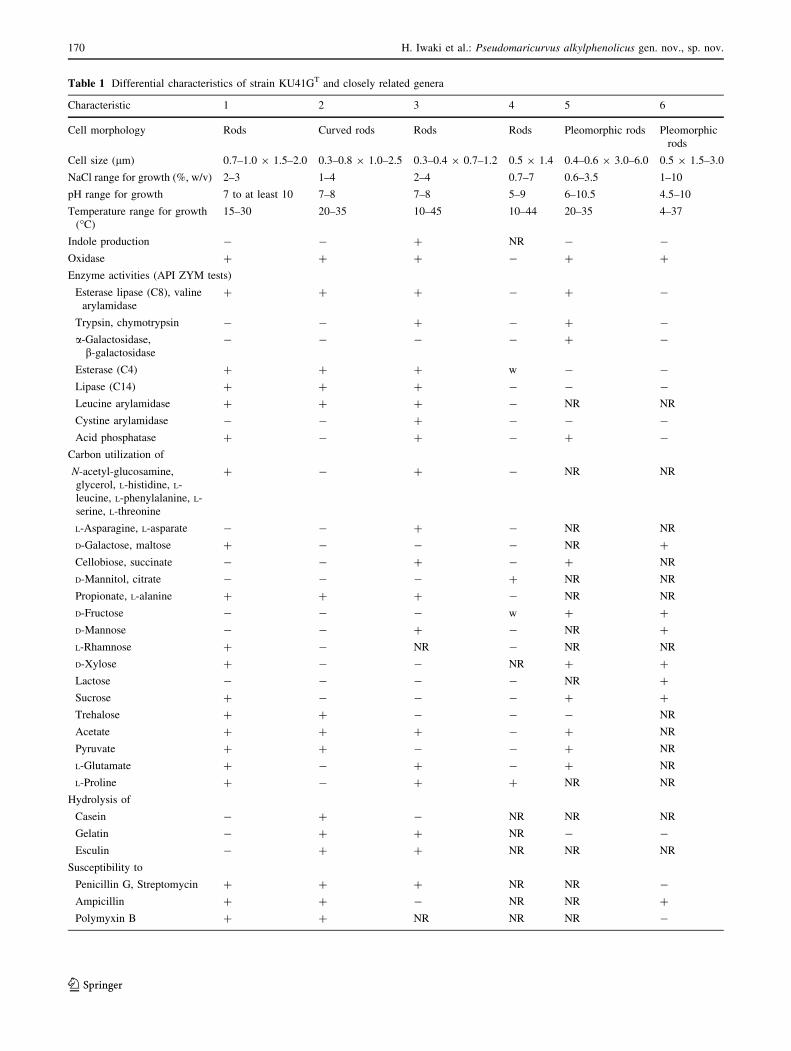
Table 1 Differential characteristics of strain KU41GT and closely related genera
Characteristic 1 2 3 4 5 6
Cell morphology Rods Curved rods Rods Rods Pleomorphic rods Pleomorphic
rods
Cell size (lm) 0.7–1.0 9 1.5–2.0 0.3–0.8 9 1.0–2.5 0.3–0.4 9 0.7–1.2 0.5 9 1.4 0.4–0.6 9 3.0–6.0 0.5 9 1.5–3.0
NaCl range for growth (%, w/v) 2–3 1–4 2–4 0.7–7 0.6–3.5 1–10
pH range for growth 7 to at least 10 7–8 7–8 5–9 6–10.5 4.5–10
Temperature range for growth
(�C)
15–30 20–35 10–45 10–44 20–35 4–37
Indole production - - ? NR - -
Oxidase ? ? ? - ? ?
Enzyme activities (API ZYM tests)
Esterase lipase (C8), valine
arylamidase
? ? ? - ? -
Trypsin, chymotrypsin - - ? - ? -
a-Galactosidase,
b-galactosidase
- - - - ? -
Esterase (C4) ? ? ? w - -
Lipase (C14) ? ? ? - - -
Leucine arylamidase ? ? ? - NR NR
Cystine arylamidase - - ? - - -
Acid phosphatase ? - ? - ? -
Carbon utilization of
N-acetyl-glucosamine,
glycerol, L-histidine, L-
leucine, L-phenylalanine, L-
serine, L-threonine
? - ? - NR NR
L-Asparagine, L-asparate - - ? - NR NR
D-Galactose, maltose ? - - - NR ?
Cellobiose, succinate - - ? - ? NR
D-Mannitol, citrate - - - ? NR NR
Propionate, L-alanine ? ? ? - NR NR
D-Fructose - - - w ? ?
D-Mannose - - ? - NR ?
L-Rhamnose ? - NR - NR NR
D-Xylose ? - - NR ? ?
Lactose - - - - NR ?
Sucrose ? - - - ? ?
Trehalose ? ? - - - NR
Acetate ? ? ? - ? NR
Pyruvate ? ? - - ? NR
L-Glutamate ? - ? - ? NR
L-Proline ? - ? ? NR NR
Hydrolysis of
Casein - ? - NR NR NR
Gelatin - ? ? NR - -
Esculin - ? ? NR NR NR
Susceptibility to
Penicillin G, Streptomycin ? ? ? NR NR -
Ampicillin ? ? - NR NR ?
Polymyxin B ? ? NR NR NR -
170 H. Iwaki et al.: Pseudomaricurvus alkylphenolicus gen. nov., sp. nov.
123

Chemotaxonomic Characteristics
The G?C content of the genomic DNA was 53.3 mol%,
and was in the range of values 36–54 mol%, which is
characteristic of members of the order Alteromonadales
[20]. The major lipoquinone was ubiquinone-8 (ubiqui-
none-8, 98.5 %; ubiquinone-7, 1.5 %), corresponding to
the members of the order Alteromonadales [20]. The major
cellular fatty acids of strain KU41GT were C17:1 x8c
(24.2 %), summed feature 3 (C15:0 iso 2-OH and/or C16:1
x7c, 16.3 %), C15:0 (10.3 %), C11:0 3-OH (9.5 %), C9:0
3-OH (6.7 %), C10:0 3-OH (6.4 %), and C18:1 x7c (5.5 %)
(Table 2). The fatty acid composition could differentiate
strain KU41GT from the other phylogenetically related
taxa: M. nonylphenolicus KU41ET [7], P. isoporae SW-11T
[21], T. turnerae T7902T [21], E. nigra 17X/A02/237T
[22], and S. degradans 2-40T [21]. These results indicate
that strain KU41GT probably represents an independent
genus of the order Alteromonadales within the class
Gammaproteobacteria.
Taxonomic Conclusions
As shown by the 16S rRNA gene sequence analysis, strain
KU41GT belongs to the order Alteromonadales within the
class Gammaproteobacteria and forms a distinct lineage
from the related genera. Furthermore, strain KU41GT can
be differentiated from closely related genera by the com-
bination of fatty acid composition, G?C content of the
DNA, and phenotypic characteristics. Considering the data
from the polyphasic study, we suggest that strain KU41GT
represents a novel species of a new genus, for which we
propose the name Pseudomaricurvus alkylphenolicus gen.
nov., sp. nov.
Description of Pseudomaricurvus gen. nov.
Pseudomaricurvus (Pseu’do.ma.ri.cur’vus. Gr. adj. pseudes,
false; N. L. masc. n. Maricurvus, a bacterial genus name;
N. L. masc. n. Pseudomaricurvus, false Maricurvus).
Cells are Gram-negative, aerobic rods, and motile by a
single polar flagellum. Sodium ions are required for their
growth. The predominant fatty acids are C17:1 x8c, summed
feature 3 (C15:0 iso 2-OH and/or C16:1 x7c), C15:0, C11:0
3-OH, C9:0 3-OH, C10:0 3-OH, C18:1 x7c. The predominant
respiratory quinone is Q-8. The type species is Pseudomar-
icurvus alkylphenolicus.
Description of Pseudomaricurvus alkylphenolicus sp.
nov.
Pseudomaricurvus alkylphenolicus (al.kyl.phe.no’li.cus N.
L. n. alkylphenolis, alkylphenol; L. suff. -icus -a -um suffix
used with the sense of belonging to; N. L. masc. adj. al-
kylphenolicus referring to the substrate alkylphenol that
can be utilized by the species).
The description is identical to that for the genus, with
the following additions. Cells are 1.5–2.0 lm in length and
0.7–1.0 lm in width. Colonies are pale yellow, circular,
smooth, convex, 1.0–2.0 mm in diameter, and with an
entire margin after 3-days incubation on MA. Oxidase- and
catalase-positive. Growth is observed between 15 and
30 �C, with optimum growth occurring at 25–30 �C, a pH
range from at least 7.0 to 10.0, with optimum growth
occurring at pH 8.0 and at NaCl concentrations of 2–3 %.
The cells are susceptible to ampicillin (10 lg), chloram-
phenicol (30 lg), gentamicin (10 lg), kanamycin (30 lg),
nalidixic acid (30 lg), novobiocin (30 lg), penicillin G
(10 U), polymyxin B (300 U), rifampicin (5 lg), strepto-
mycin (10 lg), and tetracycline (30 lg), but not to linco-
mycin (2 lg). In API ZYM system, cells are positive for
alkaline phosphatase, esterase (C4), esterase lipase (C8),
lipase (C14), leucine arylamidase, valine arylamidase, acid
phosphatase, and naphthol-AS-BI-phosphohydrolase, but
negative for all other enzymes. Hydrolysis of Tween 40 and
Tween 80 are positive. Hydrolysis of casein, esculin, and gelatin,
nitrate reduction, indole production, acid production from glu-
cose (fermentation), arginine dihydrolase, urease, and b-galac-
tosidase are negative. The cells utilize the following compounds
as sole carbon and energy sources: p-n-nonylphenol, p-n-octyl-
phenol, p-n-heptylphenol, N-acetyl-glucosamine, D-galactose, D-
glucose, L-rhamnose, D-xylose, D-maltose, sucrose, trehalose,
glycerol, acetate, caprate, n-hexanoate, propionate, pyruvate,
benzoate, 4-hydroxybenzoate, L-alanine, L-arginine, L-gluta-
mine, L-glutamate, L-histidine, L-isoleucine, L-leucine, L-lysine,
L-phenylalanine, L-proline, L-serine, L-threonine, L-tryptophan,
Table 1 continued
Characteristic 1 2 3 4 5 6
Major quinone Q-8 Q-8 Q-9 (79 %), Q-8
(21 %)
Q-8 Q-8 Q-8
DNA G?C content (mol%) 53.3 48.6 51.6 47.5 49–51 45.8
Genera 1 strain KU41GT, 2 M. nonylphenolicus KU41ET [7], 3 P. isoporae SW-11T [21], 4 E. nigra 17X/A02/237T [3], 5 T. turnerae T7902T [3,
21, 23], 6 S. degradans 2-40T [3, 21, 24, 25], ? positive reaction, - negative reaction, w weakly positive reaction, NR not reported
H. Iwaki et al.: Pseudomaricurvus alkylphenolicus gen. nov., sp. nov. 171
123

and L-valine, but not the following compounds: L-arabinose, D-
arabitol, D-fructose, D-mannitol, D-mannose, D-ribose, D-sorbitol,
myo-inositol, cellobiose, lactose, adipate, citrate, gluconate,
formate, DL-malate, succinate, L-asparagine, L-asparate, L-cys-
teine, L-glycine, and L-methionine. The DNA G?C content is
53.3 mol%.
The type strain, KU41GT (=JCM 19135T = KCTC
32386T), was isolated from seawater obtained from the coastal
region of Chichi-jima Island of Ogasawara Islands, Japan.
Acknowledgments This research was financially supported in part
by the Kansai University Grant-in-Aid for progress of research in
graduate course, 2013.
References
1. David A, Fenet H, Gomez E (2009) Alkylphenols in marine
environments: distribution monitoring strategies and detection
considerations. Mar Pollut Bull 58:953–960
2. Fujii K, Urano N, Ushio H, Satomi M, Kimura S (2001)
Sphingomonas cloacae sp. nov., a nonylphenol-degrading bac-
terium isolated from wastewater of a sewage-treatment plant in
Tokyo. Int J Syst Evol Microbiol 51:603–610
3. Ushiba Y, Takahara Y, Ohta H (2003) Sphingobium amiense sp.
nov., a novel nonylphenol-degrading bacterium isolated from a
river sediment. Int J Syst Evol Microbiol 53:2045–2048
4. Corvini PF, Schaffer A, Schlosser D (2006) Microbial degrada-
tion of nonylphenol and other alkylphenols—our evolving view.
Appl Microbiol Biotechnol 72:223–243
5. Porter AW, Hay AG (2007) Identification of opdA, a gene
involved in biodegradation of the endocrine disrupter octylphe-
nol. Appl Environ Microbiol 73:7373–7379
6. Takeo M, Prabu SK, Kitamura C, Hirai M, Takahashi H, Kato D,
Negoro S (2006) Characterization of alkylphenol degradation
gene cluster in Pseudomonas putida MT4 and evidence of oxi-
dation of alkylphenols and alkylcatechols with medium-length
alkyl chain. J Biosci Bioeng 102:352–361
7. Iwaki H, Takada K, Hasegawa Y (2012) Maricurvus nonylphe-
nolicus gen. nov., sp. nov., a nonylphenol-degrading bacterium
isolated from seawater. FEMS Microbiol Lett 327:142–147
8. Iwaki H, Yasukawa N, Fujioka M, Takada K, Hasegawa Y (2013)
Isolation and characterization of a marine cyclohexylacetate-
degrading bacterium Lutimaribacter litoralis sp. nov., and
reclassification of Oceanicola pacificus as Lutimaribacter paci-
ficus comb. nov. Curr Microbiol 66:588–593
9. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S
(2011) MEGA5: molecular evolutionary genetics analysis using
maximum likelihood, evolutionary distance, and maximum par-
simony methods. Mol Biol Evol 28:2731–2739
10. Saitou N, Nei M (1987) The neighbor-joining method: a new
method for reconstructing phylogenetic trees. Mol Biol Evol
4:406–425
11. Fitch WM (1971) Toward defining the course of evolution:
minimum change for a specific tree topology. Syst Zool
20:406–416
12. Felsenstein J (1981) Evolutionary trees from DNA sequences: a
maximum likelihood approach. J Mol Evol 17:368–376
13. Kimura M (1980) A simple method of estimating evolutionary
rates of base substitutions through comparative studies of
nucleotide sequences. J Mol Evol 16:111–120
14. Felsenstein J (1985) Confidence limits on phylogenies: an
approach using the bootstrap. Evolution 39:783–791
15. Tittsler RP, Sandholzer LA (1936) The use of semi-solid agar for
the detection of bacterial motility. J Bacteriol 31:575–580
16. Barrow GI, Feltham RKA (1993) Cowan and steel’s manual for
the identification of medical bacteria, 3rd edn. Cambridge Uni-
versity Press, Cambridge
17. Cowan ST, Steel KJ (1965) Manual for the identification of
medical bacteria. Cambridge University Press, London
18. Katayama-Fujimura Y, Komatsu Y, Kuraishi H, Kaneko T (1984)
Estimation of DNA base composition by high performance liquid
chromatography of its nuclease P1 hydrolysate. Agric Biol Chem
48:3169–3172
19. Nishijima M, Araki-Sakai M, Sano H (1997) Identification of
isoprenoid quinones by frit-FAB liquid chromatography-mass
spectrometry for the chemotaxonomy of microorganisms.
J Microbiol Met 28:113–122
20. Bowman JP, McMeekin TA (2005) Order X. Alteromonadales
ord. nov. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM (eds)
Table 2 Cellular fatty acid composition (%) of strain KU41GT and
closely related genera
Fatty acid 1 2 3 4 5 6
C9:0 4.1 1.0 – – – –
C10:0 2.2 6.4 3.5 – 3.2 –
C9:0 3-OH 6.7 –
C10:0 3-OH 6.4 9.4 3.2 6.7 1.6 8.3
C11:0 – – 2.5 – – 2.4
C11:0 3-OH 9.5 – 4.0 – – –
C12:0 – – 3.0 – 2.5 3.0
C12:0 anteiso – – – – – 8.4
C12:0 3-OH – – 2.4 – 4.7 –
C12:1 3-OH – – – – – 4.3
C13:0 anteiso – – – – – 4.1
C14:0 – 1.3 1.1 – 4.0 5.1
C15:0 10.3 1.8 – – – –
C15:1 x6c 2.4 – – – – –
C15:1 x8c 1.2 0.1 – – – –
C16:0 3.5 17.0 10.3 17.8 20.9 28.9
C16:0 3-OH – – – 1.0 – –
C17:0 2.4 1.8 5.1 1.3 1.3 17.3
C17:1 x6c 3.0 – 1.2 – – –
C17:1 x8c 24.2 5.6 13.4 – – –
C17:1 anteiso x9c – – 10.0 – – –
C18:0 – 1.6 – 6.8 4.2 5.1
C18:1 x6c – 3.0 13.1b – 36.7b 9.9b
C18:1 x7c 5.5 19.8 29.0
Summed feature 3a 16.3c 28.4c 22.3d 36.8c 18.1d 3.4d
Values are percentages of the total fatty acids; fatty acids that make up
\1 % of the total are not shown or indicated by ‘‘-‘‘
Genera 1 strain KU41GT, 2 M. nonylphenolicus KU41ET [7], 3 P.
isoporae SW-11T [21], 4 E. nigra 17X/A02/237T [3], 5 T. turnerae
T7902T [21], 6 S. degradans 2-40T [21]a, c, d Summed features are groups of two fatty acids that cannot be
separated by GLC using the MIDI system. Summed feature 3 com-
prises cC15:0 iso 2-OH and/or C16:1 x7c; dC16:1 x7c and/or C16:1 x6cb C18:1 x6c and/or C18:1 x7c
172 H. Iwaki et al.: Pseudomaricurvus alkylphenolicus gen. nov., sp. nov.
123

Bergey’s manual of systematic bacteriology, vol 2, 2nd edn.
Springer, New York, p 443
21. Chen MH, Sheu SY, Arun AB, Young CC, Chen CA, Wang JT,
Chen WM (2011) Pseudoteredinibacter isoporae gen. nov., sp.
nov., a marine bacterium, isolated from the reef-building coral
Isopora palifera. Int J Syst Evol Microbiol 61:1887–1893. doi:10.
1099/ijs.0.024190-0
22. Urios L, Intertaglia L, Lesongeur F, Lebaron P (2011) Eionea
nigra gen. nov., sp. nov., a gammaproteobacterium from the
Mediterranean Sea. Int J Syst Evol Microbiol 61:1677–1681
23. Distel DL, Morrill W, MacLaren-Toussaint N, Franks D,
Waterbury J (2002) Teredinibacter turnerae gen. nov., sp. nov., a
dinitrogen-fixing, cellulolytic, endosymbiotic gamma-proteobac-
terium isolated from the gills of wood-boring molluscs (Bivalvia:
Teredinidae). Int J Syst Evol Microbiol 52:2261–2269
24. Ekborg NA, Gonzalez JM, Howard MB, Taylor LE, Hutcheson
SW, Weiner RM (2005) Saccharophagus degradans gen. nov.,
sp. nov., a versatile marine degrader of complex polysaccharides.
Int J Syst Evol Microbiol 55:1545–1549
25. Gonzalez JM, Weiner RM (2000) Phylogenetic characterization
of marine bacterium strain 2–40, a degrader of complex poly-
saccharides. Int J Syst Evol Microbiol 50:831–834
H. Iwaki et al.: Pseudomaricurvus alkylphenolicus gen. nov., sp. nov. 173
123