is the effect of wounding on leaf senescence due to ethylene?
TRANSCRIPT

Plant Science, 47 (1986) 11--14 11 Elsevier Scientific Publishers Ireland Ltd.
IS THE E F F E C T OF W O U N D I N G ON L E A F S E N E S C E N C E DUE TO E T H Y L E N E ?
G. GIRIDHAR a and K.V. THIMANN b
aBiology Department, Wake Forest University, Winston-Salem, NC 27109 and bThimann Laboratories, University of California, Santa Cruz, CA 95064 (U.S.A.)
(Received April 5th, ~.986) (Revision received August 8th, 1986) (Accepted August 8th, 1986)
Wounding delays the loss of chlorophyll (Chl) that normally occurs when oat (Avena sativa L.) leaf segments are held in the dark. There was a continued increase in ethylene production during the senescence of the control segments; in contrast, ethylene production by the wounded segments, although it increased by a factor of 2--3 times, reached its peak in 48 h and then dropped sharply to below the basal level. Added 1-aminocyclopropane- 1-carboxylic acid (ACC) caused a very large increase in ethylene production in both control and wounded seg- ments, but it increased the rate of Chl loss, though only marginally. Aminoethoxyvinylglycine (AVG)inhibited ethylene production by both control and wounded segments and this did decrease the Chl loss, but only in the control segments. In the wounded segments, AVG antagonized the Chl-retaining action of the wound. Since wounding delayed the loss of Chl and yet caused a moderate increase in ethylene production, we conclude that the ethylene production by senescing oat leaves is not the main controlling influence in the wounding effect. The data also throw doubt on the causal participation of ethylene in normal Chl loss by these leaves in darkness.
Key words: oat leaf segments; senescence; wounding; ethylene
I n t r o d u c t i o n
Senescence of de tached leaves is induced by incuba t ion in darkness and is character ized by the loss of Chl and proteins and a con- c o m i t a n t increase in free amino ni t rogen [ 1 - - 6 ] . A m o n g the various factors tha t regulate foliar senescence, plant h o r m o n e s have been p r o m i n e n t [4 - -12] and the role of ABA as a p r o m o t e r of senescence is well established [ 7 , 1 3 ] . The role of e thylene , however , is less clear. Aharon i and Lieberman found tha t dur ing the rapid Chl loss, b o t h e thylene (Eth) p roduc t i on and respirat ion rate increased whereas a decrease in Eth p r o d u c t i o n and respirat ion rate occur red during the slow phase of Chl loss [ 1 4 ] .
Abbreviations: ABA, abscisic acid; ACC, 1-amino- cyclopropane-l-carboxylic acid; AVG, aminoethoxy- vinylglycine; Chl, chlorophyll; Eth, ethylene.
Several o ther studies have po in ted to Eth as a regulator of foliar senescence [ 8 - - 1 7 ] , bu t in several of these reports, the increase in Eth p roduc t i on in presence of its precursors is very great while the increase in the rate of Chl loss is only slight [e.g. 1, 16] . A l though ABA p r o m o t e d senescence in rice leaves it significantly decreased Eth p roduc t ion , and cor respondingly , benzyladenine re tarded senescence bu t p r o m o t e d Eth p roduc t i on [ 1 8 ] . In contras t , in oat leaves, ABA p r o m o t e d senescence and p r o m o t e d Eth p roduc t i on [ 1 6 ] . While ACC had little ef fec t on senes- cence in the dark, kinetin, which re tarded senescence, p r o m o t e d Eth p roduc t i on in bo th light and darkness. However , kinetin decreased Eth p r o d u c t i o n in aquatic plants [ 1 9 ] .
Recen t ly it was shown that mechanical wound ing inhibits the senescence of oat leaf segments held in the dark and the a m o u n t of inhibi t ion is p ropor t ina l to the in tensi ty of wound ing [ 2 0 ] . An increase in p roduc t i on
0618-9452/86/$03.50 © Elsevier Scientific Publishers Ireland Ltd. Printed and Published in Ireland

12
of Eth on wounding fruits, storage tissues and also leaves is well documented and there have been several studies of this 'wound ethylene ' [2 ,21--24] . In fruits the effect can be very large [e.g. 22] . Because the effect of injury is relevant to many types of experiments in which tissues are peeled or scrubbed with abrasive, we felt a need for detailed study on the interaction of wounding and Eth on the oat leaf segments which we have used rout inely for senescence studies. The work was planned to answer the question, is the effect of wounding on the senescence of oat leaf segments in the dark due to Eth?
Materials and methods
Oat seeds (Arena sativa L. cv. Victory) obtained from SvalSf AB, International Division, SvalSv, Sweden were presoaked in water for 1 h and sown in vermiculite. The seedlings were grown at 25°C under con- t inuous light from daylight fluorescent lamps giving 30 pE • m-: s -~ at the plant level. Three-cent imeter long subapical segments from the first leaves of 8-day-old seedlings were used. The wounding was by gently scraping the adaxial surface of the segments with a sharp blade [20] .
Ten segments were placed vertically in 0.5 ml of distilled water or test solution in a glass vial of internal dimensions 7.1 X 1 cm. The air volume after capping the vials with serum caps was 5.0 ml. The vials were kept open in the dark during the senescence period. They were kept closed for 1 h at the appropriate time period, when a 1-ml air sample was with- drawn and injected into a Varian 3700 series Gas Chromatograph equipped with 80--100 mesh alumina in a 6 f t X 1/8 inch stainless steel column and flame ionization detector. After the Eth determinat ion the vials were opened again and kept in the dark until the next Eth measuring time.
For the time course study of the effect of ACC on the loss of Chl, 10 segments (average fresh wt. 16 mg) were f loated on water or 0.3 mM ACC solution in petri dishes and kept in
the dark. Crystalline penicillin G (100 pM) is added routinely in all our long-term experi- ments; there was no visible microbial growth. Samples were taken every 24 h and the Chl extracted and estimated [20] .
Results and conclusions
The hourly product ion of Eth by control and wounded oat leaf segments over a period of 96 h is shown in Table I. The effect of wounding on the loss of Chl is included in the same table. It is seen that, as previously reported [ 20 ], wounding considerably inhibits the loss of Chl by the segments after 4 days in the dark. There is a gradual increase in Eth product ion by the senescing control segments, and at 96 h the rate was 2.7 times that in the first hour. After the same time period the segments had lost more than 75% of their initial content of Chl. But in the wounded segments, where there is clear inhibition of Chl loss, the rate of Eth product ion increased for only 48 h and then dropped sharply to below the coDtrol level afterwards. Thus one effect of wounding is to bring forward the product ion of Eth to an earlier time. (If the product ion rates were to remain constant at the average of the values shown at the begin- ning and end of each 24-h period, the total Eth accumulated in 96 h would be 24 X 11.9 nl for the controls and 24 X 19.7 for the wounded; a very small total increase).
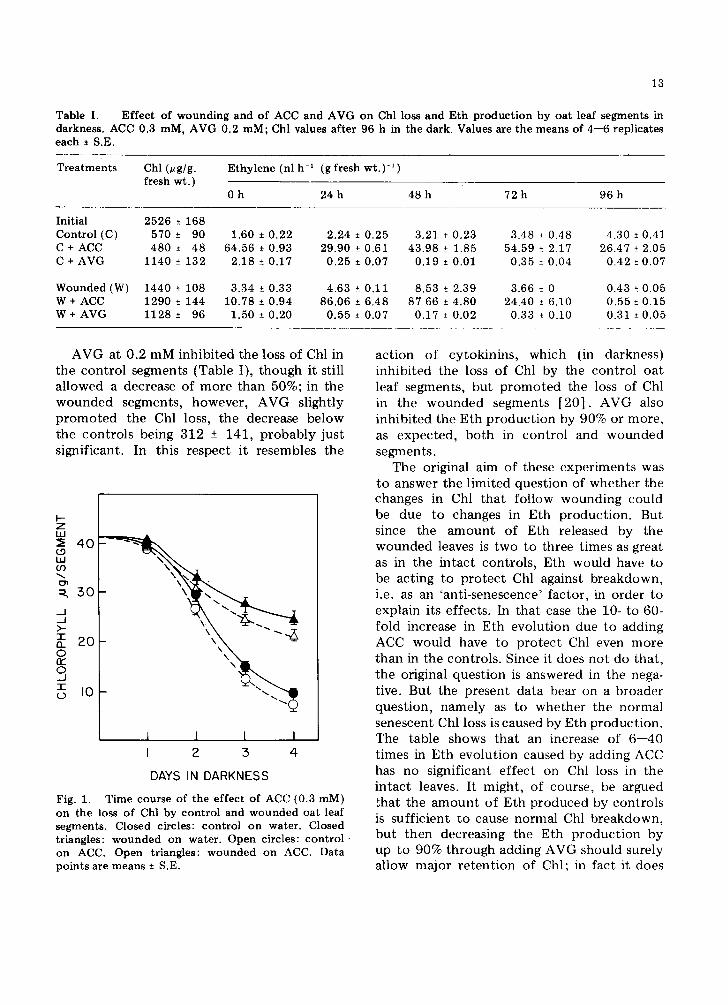
ACC at 0.3 mM concentra t ion caused only a slight increase in the Chl loss from the control segments (Fig. 1). Its effect on Eth product ion by both control and wounded segments, however, was dramatic, as shown in Table I. Even during the first hour ACC en- hanced the rate of Eth product ion in the controls by more than 40-fold. The rate showed a dip at the 24-h period before a second increase in rate at 48 h and 72 h. By 96 h the rate had again dropped to a low level. In the wounded segments, after ACC had caused a 20-fold increase in the rate of Eth product ion, there was a decline in rate, followed by a very sharp drop after 72 h.
13
Table I. Effect of wounding and of ACC and AVG on Chl loss and Eth production by oat leaf segments in darkness. ACC 0.3 mM, AVG 0.2 raM; Chl values after 96 h in the dark. Values are the means of 4--6 replicates each , S.E.
T r e a t m e n t s Chl (#g/g . f resh wt . )
E t h y l e n e (nl h - ' (g f r e sh w t . ) -~ )
0 h 24 h 48 h 72 h 96 h
Init ia l 2 5 2 6 ± 168 C o n t r o l (C) 570 ± 90 1.60 ± 0 .22 2 .24 ± 0 .25 3.21 + 0 .23 3.48 * 0 .48 4.30 ± 0.41 C + A C C 480 ± 48 64 .56 ± 0 .93 29 .90 ± 0 . 6 1 43 .98 ± 1.85 54.59_* 2.17 2 6 . 4 7 ± 2 . 0 5 C + A V G 1140 ± 132 2 .18 ± 0 .17 0 .25 ± 0 .07 0 .19 ± 0 .01 0 .35 + 0 .04 0 .42 ~_ 0 .07
W o u n d e d (W) 1440 ± 108 3 .34 _* 0.33 4 .63 +_ 0 .11 8 .53 ± 2.39 3.66 _+ 0 0 .43 ~ 0 .05 W + ACC 1290 ± 144 10 .78 ± 0 .94 86 .06 ± 6 .48 87 66 ± 4 .80 24 .40 *_ 6 .10 0 .55 ± 0 .15 W + A V G 1128 ± 96 1 .50 ± 0 . 2 0 0 . 5 5 ± 0 .07 0 .17 ± 0 .02 0 .33 ± 0 . 1 0 0 . 3 1 ± 0 . 0 5
A V G at 0.2 mM inh ib i ted the loss of Chl in the con t ro l segments (Table I), t hough it still a l lowed a decrease of m o r e t han 50%; in the w o u n d e d segments , however , A V G slightly p r o m o t e d the Chl loss, the decrease be low the con t ro l s being 312 + 141, p r o b a b l y jus t s ignif icant . In this r e spec t it r esembles the
b- Z LtJ
4 0 o I,I
~, 5O
._.1 __1
I a_ 20 O n- O _.1 T
Fig. 1.
- \ \
I I I I I 2 5 4-
DAYS IN DARKNESS
T i m e cou r se o f t he e f fec t o f ACC (0.3 raM) o n the loss o f Chl by c o n t r o l and w o u n d e d oa t leaf s e g m e n t s . C l o s e d c irc les : c o n t r o l o n wa te r . Closed t r iangles : w o u n d e d o n w a t e r . O p e n circles: c o n t r o l - o n ACC. O p e n t r iangles : w o u n d e d o n ACC. Da ta p o i n t s are m e a n s ± S.E.
ac t ion o f cy tok in ins , which (in darkness) inh ib i ted the loss of Chl by the con t ro l oa t leaf segments , bu t p r o m o t e d the loss of Chl in the w o u n d e d segmen t s [ 2 0 ] . A V G also inh ib i ted the E th p r o d u c t i o n by 90% or m o r e , as e x p e c t e d , b o t h in con t ro l and w o u n d e d segments .
The original aim of these e x p e r i m e n t s was to answer the l imi ted ques t ion of w h e t h e r the changes in Chl t ha t fo l low wound ing could be due to changes in Eth p roduc t i on . But since the a m o u n t of E th released by the w o u n d e d leaves is t w o to three t imes as great as in the in tac t con t ro l s , E th would have to be act ing to p r o t e c t Chl against b r e a k d o w n , i.e. as an ' an t i - senescence ' fac tor , in order to expla in its effects . In t ha t case the 10- to 60- fo ld increase in E th evo lu t ion due to adding ACC would have to p r o t e c t Chl even m o r e t h a n in the cont ro ls . Since it does no t do tha t , the original ques t ion is answered in the nega- tive. But the p resen t da ta bear on a b roade r ques t ion , n a m e l y as to whe the r the no rma l senescent Chl loss is caused by Eth p roduc t i on . The tab le shows tha t an increase of 6 - -40 t imes in E th evo lu t ion caused by adding ACC has no s ignif icant e f f ec t on Chl loss in the in tac t leaves. I t might , o f course , be argued ~hat the a m o u n t o f E th p r o d u c e d by con t ro l s is suf f ic ien t to cause no rma l Chl b r e a k d o w n , bu t t hen decreas ing the Eth p r o d u c t i o n by up to 90% th rough adding A V G should surely al low m a j o r r e t e n t i o n of Chl; in fac t it does

14
i n c r e a s e Chl r e t e n t i o n b y a m o d e s t a m o u n t , s o m e 3 0 % in t h e i n t a c t c o n t r o l s , b u t has n o such e f f e c t in t h e w o u n d e d leaves , even s o m e - w h a t i n c r e a s i n g t h e Chl loss . P u t t i n g all t h e d a t a t o g e t h e r , we c o n c l u d e t h a t t h e p r o d u c t i o n o f E t h b y s e n e s c i n g o a t l e a f s e g m e n t s in d a r k - ness c a n h a r d l y be t h e m a j o r c o n t r o l l i n g f a c t o r in t h e i r s e n e s c e n c e . I t m a y be a c o n t r i b u t o r y f a c t o r , s i n c e e x o g e n o u s E t h d o e s p r o m o t e Chl loss in l i g h t [ 1 6 ] . In t h i s r e s p e c t o a t l e a f s e g m e n t s s h o w a s h a r p c o n t r a s t w i t h f r u i t s , f o r t h e a m o u n t o f E t h p r o d u c e d f r o m a d d e d A C C {Tab le I) is o f a m a g n i t u d e c o m p a r a b l e w i t h t h a t f o r m e d w i t h o u t a d d e d A C C in m a n y e d i b l e f r u i t s , y e t in such f r u i t s E t h is i n d e e d t h e m a j o r c o n t r o l l i n g f a c t o r in m a - t u r a t i o n a n d s e n e s c e n c e .
A c k n o w l e d g m e n t
We t h a n k Dr . T . L . D a v e n p o r t , in w h o s e l a b o r a t o r y m o s t o f t h e w o r k was c a r r i e d o u t .
R e f e r e n c e s
1 P.K. Bassi and M.S. Spencer, Plant Physiol., 73 (1983) 758.
2 M. Esaka, T. Takahashi and T. Asahi, Plant Cell Physiol., 24 (1983) 625.
3 R.B. Popovic, D.J. Kyle, A.S. Cohen and S. Zalik, Plant Physiol., 64 (1979) 721.
4 K.V. Thimann, in: Senescence in Plants, CRC Press, Boca Raton, FL, 1982, p. 86.
5 H.B. Vickery, G.W. Pucher, A.J. Wakeman and C.S. Leavenworth, Conn. Agr. Exp. Sta. New Haven Bull., 399 (1937) 757.
6 A.C. Leopold, Science, 1334 {1961) 1727. 7 F.T. Addicott and J.L. Lyon, Ann. Rev. Plant
Physiol., 20 (1969) 139. 8 N. Aharoni and M. Lieberman, Plant Physiol., 20
(1969) 139. 9 R.A. Fletcher and D.J. Osborne, Can. J. Bot., 44
(1966) 739. 10 A.E. Richmond and A. Lang, Science, 125 (1957)
650. 11 J.A. Sacher, Science, 125 (1957) 1199. 12 J.A. Sacher, Plant Physiol., 13 (1959) 365. 13 H.M.M. E1-Antably, P.F. Wareing and J. Hillman,
Planta, 73 (1967) 74. 14 N. Aharoni and M. Lieberman, Plant Physiol., 64
(1979) 796. 15 C.R. Barmore, Hort. Sci., 10 (1975) 595. 16 S. Gepstein and K.V. Thimann, Plant Physiol., 68
{1981) 349. 17 P.W. Morgan and J.I. Durham, Plant Physiol., 66
(1980) 88. 18 C.H. Kao and S.F. Yang, Plant Physiol., 73 (1983)
881. 19 S. Jana and M.A. Choudhuri, Aquat. Bot., 13
(1982) 359. 20 G. Giridhar and K.V. Thimann, Plant Physiol.,
78 (1985) 29. 21 T. Boller and H. Kende, Nature, 286 (1980) 259. 22 N.E. Hoffman and S.F. Yang, Plant Physiol.,
69 (1982) 317. 23 H. Imasaki, M. Uchiyama and I. Uritani, Agric.
Biol. Chem., 32 (1968) 387. 24 H. Kende and T. Boller, Planta, 151 (1981) 476.