is higher biodiversity always leading to better forest ... · first authors observed a positive...
TRANSCRIPT

Is higher biodiversity always leading to
better forest ecosystem functioning? Executive Summary by Romain Cottet, Abigail Fellay, Franziska Scheibler
INTRODUCTION As global change threatens ecosystems and forests worldwide, the functions and services they provide are being jeopardized. Following
the Millenium Assessment (MA), the concern that the loss of biodiversity would affect the provisioning of ecosystem service grew. This
led to the development of a new study field whose goal was to assess the relationship between biodiversity and ecosystem functioning
(BEF). In order to dive into this field, one has to first define two
things: biodiversity and ecosystem functioning. Although
biodiversity will inevitably be mentioned a few times, we will
mainly focus here on the definition of ecosystem functioning as it
has been used in the field of forestry.
STATE OF THE ART
BIODIVERSITY AND ECOSYSTEM FUNCTIONS
The overarching conclusion emerging from this field of study
follows the direction of the concerns of the MA in the sense that
biodiversity is found to be a major driving force in ecosystem functions in general, and this for two main reasons: o Biodiversity provides a range of different functions by exploiting complementary spatial and temporal niches (e.g. nitrogen fixing
trees increase growth of other species) (Brockerhoff et al. 2017; Tilman et al. 2001).
o Biodiversity leads to more resistant and resilient forests, in turn ensuring ecosystem functions over a longer period of time,
especially when facing changing conditions (Insurance Hypothesis) (Bengtsson et al. 2000).
However, most studies focus on productivity as a proxy for ecosystem functioning (Brockerhoff et al. 2017). It has been chosen as a
reference point for forestry because it is a good economic indicator and it is linked to carbon sequestration, the most important
ecosystem function regarding climate change. Productivity can be defined as the increasing amount of plant biomass (growth) which is
derived by using the diameter at breast height (DBH) to compute the volume of a tree or of a whole stand (Vilà et al. 2013; Huang et al.
2018). But it can also be defined using NPP measured with Eddy covariance towers.
BIODIVERSITY AND PRODUCTIVITY
The relationship between productivity and species diversity has been explored through experimental and observational studies as well
as simulation models.
EXPERIMENTAL AND OBSERVATIONAL STUDIES
The main findings are the following:
o In experimental studies, positive relationship between tree species richness and stand basal area or stand volume, two indicators
of productivity are often highlighted (e.g. Huang et al. 2018). This positive effect sometimes appears to grow stronger with time.
o Observational studies using worldwide forest inventories show that biodiversity is not the predominant factor influencing plant
biomass production. Indeed, other abiotic and biotic environmental factors (e.g. climate variables, soil fertility or diseases and
herbivory activity) have been shown to be more effective in measuring productivity (Vilà et al. 2013; Poorter et al. 2015).
Difficulties and limitations:
o The slow growth rate of forests implies that long-term experiments are scarce. Most of them have not been carried out for more
than 10 years so far and are therefore not yet representative of natural dynamics. However, a number of experiments have been
set up and are expected to yield results in coming years (Scherer-Lorenzen et al. 2007).
o Observational studies are often based on forest inventories from around the world. But by doing so, it is impossible to control
for the covariates. Therefore, sophisticated statistical analysis needs to be carried out to highlight the effect of biodiversity on
productivity.
SIMULATION MODELS To overcome the limitations of forest experiments and observational studies, scientists have turned to the power of computers. Forest
models allow us to generate thousands of predictions for many different forests in controlled growing conditions over long periods of
time. Those are mechanistic models, in other words a set of complex equations that try to translate into numbers some natural processes
such as light interception in the canopy (Courbaud et al., 2003) or the allometric relationships of trees (Chave et al. 2014). In order to
assess the relationship between biodiversity and productivity, the number of different species in the modelized forests can be
manipulated. In two studies, using simulations models, Morin et al. (2011) and Bohn and Huth (2017) obtained two different results (Figure 1). The
first authors observed a positive linear relationship between the biodiversity and productivity or net biodiversity effect. This second
Ecosystem function are the ecological (biological, chemical, physical)
mechanisms that support the integrity or maintenance of ecosystems.
This concept is ‘‘ecosystem-centered’’ (Brockerhoff et al. 2017).
Ecosystem services retain a connection to the underlying ecosystem
function that generate them. Ecosystem services represent the
contributions that ecosystems make to human well-being. Therefore,
this concept is ‘‘human-centered’’ (Brockerhoff et al. 2017).
Biodiversity is a multifaceted concept that includes the diversity of life
on different levels of organization from genes, to species, to entire
ecosystems. Biodiversity as such is not an ecosystem function but is
important to many ecosystem functions (Dislich et al. 2017).
>>Because of the inherent difficulties, experimental studies confirming
these results are scarce and not yet representative of reality.
1
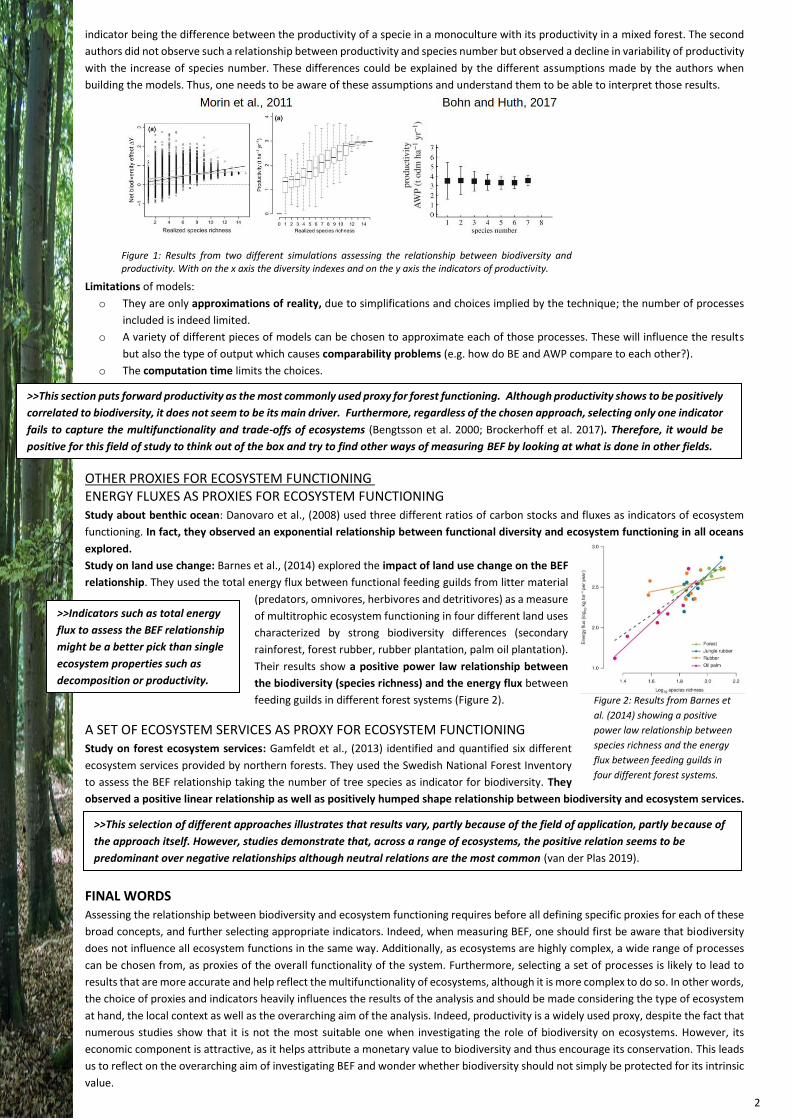
Figure 1: Results from two different simulations assessing the relationship between biodiversity and productivity. With on the x axis the diversity indexes and on the y axis the indicators of productivity.
indicator being the difference between the productivity of a specie in a monoculture with its productivity in a mixed forest. The second
authors did not observe such a relationship between productivity and species number but observed a decline in variability of productivity
with the increase of species number. These differences could be explained by the different assumptions made by the authors when
building the models. Thus, one needs to be aware of these assumptions and understand them to be able to interpret those results.
Limitations of models: o They are only approximations of reality, due to simplifications and choices implied by the technique; the number of processes
included is indeed limited.
o A variety of different pieces of models can be chosen to approximate each of those processes. These will influence the results
but also the type of output which causes comparability problems (e.g. how do BE and AWP compare to each other?).
o The computation time limits the choices.
OTHER PROXIES FOR ECOSYSTEM FUNCTIONING ENERGY FLUXES AS PROXIES FOR ECOSYSTEM FUNCTIONING Study about benthic ocean: Danovaro et al., (2008) used three different ratios of carbon stocks and fluxes as indicators of ecosystem
functioning. In fact, they observed an exponential relationship between functional diversity and ecosystem functioning in all oceans
explored.
Study on land use change: Barnes et al., (2014) explored the impact of land use change on the BEF
relationship. They used the total energy flux between functional feeding guilds from litter material
(predators, omnivores, herbivores and detritivores) as a measure
of multitrophic ecosystem functioning in four different land uses
characterized by strong biodiversity differences (secondary
rainforest, forest rubber, rubber plantation, palm oil plantation).
Their results show a positive power law relationship between
the biodiversity (species richness) and the energy flux between
feeding guilds in different forest systems (Figure 2).
A SET OF ECOSYSTEM SERVICES AS PROXY FOR ECOSYSTEM FUNCTIONING Study on forest ecosystem services: Gamfeldt et al., (2013) identified and quantified six different
ecosystem services provided by northern forests. They used the Swedish National Forest Inventory
to assess the BEF relationship taking the number of tree species as indicator for biodiversity. They
observed a positive linear relationship as well as positively humped shape relationship between biodiversity and ecosystem services.
FINAL WORDS
Assessing the relationship between biodiversity and ecosystem functioning requires before all defining specific proxies for each of these
broad concepts, and further selecting appropriate indicators. Indeed, when measuring BEF, one should first be aware that biodiversity
does not influence all ecosystem functions in the same way. Additionally, as ecosystems are highly complex, a wide range of processes
can be chosen from, as proxies of the overall functionality of the system. Furthermore, selecting a set of processes is likely to lead to
results that are more accurate and help reflect the multifunctionality of ecosystems, although it is more complex to do so. In other words,
the choice of proxies and indicators heavily influences the results of the analysis and should be made considering the type of ecosystem
at hand, the local context as well as the overarching aim of the analysis. Indeed, productivity is a widely used proxy, despite the fact that
numerous studies show that it is not the most suitable one when investigating the role of biodiversity on ecosystems. However, its
economic component is attractive, as it helps attribute a monetary value to biodiversity and thus encourage its conservation. This leads
us to reflect on the overarching aim of investigating BEF and wonder whether biodiversity should not simply be protected for its intrinsic
value.
Figure 2: Results from Barnes et
al. (2014) showing a positive
power law relationship between
species richness and the energy
flux between feeding guilds in
four different forest systems.
>>Indicators such as total energy
flux to assess the BEF relationship
might be a better pick than single
ecosystem properties such as
decomposition or productivity.
>>This selection of different approaches illustrates that results vary, partly because of the field of application, partly because of
the approach itself. However, studies demonstrate that, across a range of ecosystems, the positive relation seems to be
predominant over negative relationships although neutral relations are the most common (van der Plas 2019).
>>This section puts forward productivity as the most commonly used proxy for forest functioning. Although productivity shows to be positively
correlated to biodiversity, it does not seem to be its main driver. Furthermore, regardless of the chosen approach, selecting only one indicator
fails to capture the multifunctionality and trade-offs of ecosystems (Bengtsson et al. 2000; Brockerhoff et al. 2017). Therefore, it would be
positive for this field of study to think out of the box and try to find other ways of measuring BEF by looking at what is done in other fields.
2

REFERENCES Barnes, Andrew D., Malte Jochum, Steffen Mumme, Noor Farikhah Haneda, Achmad Farajallah, Tri Heru Widarto, and Ulrich Brose.
2014. ‘Consequences of Tropical Land Use for Multitrophic Biodiversity and Ecosystem Functioning’. Nature Communications 5: 1–7. https://doi.org/10.1038/ncomms6351.
Bengtsson, Jan, Sven G. Nilsson, Alain Franc, and Paolo Menozzi. 2000. ‘Biodiversity, Disturbances, Ecosystem Function and Management of European Forests’. Forest Ecology and Management 132 (1): 39–50. https://doi.org/10.1016/S0378-1127(00)00378-9.
Bohn, Friedrich J., and Andreas Huth. 2017. ‘The Importance of Forest Structure to Biodiversity-Productivity Relationships’. Royal Society Open Science 4 (1). https://doi.org/10.1098/rsos.160521.
Brockerhoff, Eckehard G., Luc Barbaro, Bastien Castagneyrol, David I. Forrester, Barry Gardiner, José Ramón González-Olabarria, Phil O.B. Lyver, et al. 2017. ‘Forest Biodiversity, Ecosystem Functioning and the Provision of Ecosystem Services’. Biodiversity and Conservation 26 (13): 3005–35. https://doi.org/10.1007/s10531-017-1453-2.
Chave, Jérôme, Maxime Réjou-Méchain, Alberto Búrquez, Emmanuel Chidumayo, Matthew S. Colgan, Welington B.C. Delitti, Alvaro Duque, et al. 2014. ‘Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees’. Global Change Biology 20 (10): 3177–90. https://doi.org/10.1111/gcb.12629.
Courbaud, Benoît, François De Coligny, and Thomas Cordonnier. 2003. ‘Simulating Radiation Distribution in a Heterogeneous Norway Spruce Forest on a Slope’. Agricultural and Forest Meteorology 116 (1–2): 1–18. https://doi.org/10.1016/S0168-1923(02)00254-X.
Danovaro, Roberto, Cristina Gambi, Antonio Dell’Anno, Cinzia Corinaldesi, Simonetta Fraschetti, Ann Vanreusel, Magda Vincx, and Andrew J. Gooday. 2008. ‘Exponential Decline of Deep-Sea Ecosystem Functioning Linked to Benthic Biodiversity Loss’. Current Biology 18 (1): 1–8. https://doi.org/10.1016/j.cub.2007.11.056.
Dislich, Claudia, Alexander C. Keyel, Jan Salecker, Yael Kisel, Katrin M. Meyer, Mark Auliya, Andrew D. Barnes, et al. 2017. ‘A Review of the Ecosystem Functions in Oil Palm Plantations, Using Forests as a Reference System’. Biological Reviews 92 (3): 1539–69. https://doi.org/10.1111/brv.12295.
Gamfeldt, Lars, Tord Snäll, Robert Bagchi, Micael Jonsson, Lena Gustafsson, Petter Kjellander, María C. Ruiz-Jaen, et al. 2013. ‘Higher Levels of Multiple Ecosystem Services Are Found in Forests with More Tree Species’. Nature Communications 4. https://doi.org/10.1038/ncomms2328.
Huang, Yuanyuan, Yuxin Chen, Nadia Castro-Izaguirre, Martin Baruffol, Matteo Brezzi, Anne Lang, Ying Li, et al. 2018. ‘Impacts of Species Richness on Productivity in a Large-Scale Subtropical Forest Experiment’. Science 362 (6410): 80–83. https://doi.org/10.1126/science.aat6405.
Morin, Xavier, Lorenz Fahse, Michael Scherer-Lorenzen, and Harald Bugmann. 2011. ‘Tree Species Richness Promotes Productivity in Temperate Forests through Strong Complementarity between Species’. Ecology Letters 14 (12): 1211–19. https://doi.org/10.1111/j.1461-0248.2011.01691.x.
Plas, Fons van der. 2019. ‘Biodiversity and Ecosystem Functioning in Naturally Assembled Communities’. Biological Reviews 94 (4): 1220–45. https://doi.org/10.1111/brv.12499.
Poorter, L., M. T. van der Sande, J. Thompson, E. J.M.M. Arets, A. Alarcón, J. Álvarez-Sánchez, N. Ascarrunz, et al. 2015. ‘Diversity Enhances Carbon Storage in Tropical Forests’. Global Ecology and Biogeography 24 (11): 1314–28. https://doi.org/10.1111/geb.12364.
Scherer-Lorenzen, Michael, Ernst Detlef Schulze, Axel Don, Jens Schumacher, and Eberhard Weller. 2007. ‘Exploring the Functional Significance of Forest Diversity: A New Long-Term Experiment with Temperate Tree Species (BIOTREE)’. Perspectives in Plant Ecology, Evolution and Systematics 9 (2): 53–70. https://doi.org/10.1016/j.ppees.2007.08.002.
Tilman, D., P. B. Reich, J. Knops, D. Wedin, T. Mielke, and C. Lehman. 2001. ‘Diversity and Productivity in a Long-Term Grassland Experiment’. Science 294 (5543): 843–45. https://doi.org/10.1126/science.1060391.
Vilà, Montserrat, Amparo Carrillo-Gavilán, Jordi Vayreda, Harald Bugmann, Jonas Fridman, Wojciech Grodzki, Josephine Haase, Georges Kunstler, Mart Jan Schelhaas, and Antoni Trasobares. 2013. ‘Disentangling Biodiversity and Climatic Determinants of Wood Production’. PLoS ONE 8 (2). https://doi.org/10.1371/journal.pone.0053530.
3