is canine hepatocerebellar degeneration syndrome an animal model for carbohydrate-deficient...
TRANSCRIPT

Is Canine Hepatocerebellar DegenerationSyndrome an Animal Model for Carbohydrate-deficient Glycoprotein Syndrome in Humans? AnExample of Sequencing Glycoprotein Glycanswith Mass Spectrometry
Yi Yang,1† Kaveh D. Arbtan,2 K. Paige Carmichael3 and Ron Orlando1*1Complex Carbohydrate Research Center, and Departments of Biochemistry & Molecular Biology, and Chemistry, Universityof Georgia, 220 Riverbend Road, Athens, GA 30602-4712, USA2Athens Research and Technology, Inc., 183 Paradise Blvd., Athens, GA 30607, USA3Department of Veterinary Pathology, College of Veterinary Medicine, University of Georgia, Athens, GA 30602-7388, USA
The clinical symptoms and morphologic features of canine hepatocerebellar degeneration syndrome (CHD)bear striking resemblance to those of human carbohydrate-deficient glycoprotein syndrome (CDGS). Thecharacteristic biochemical and molecular features of human CDGS lie in the truncated carbohydrate sidechains of serum transferrin and numerous other glycoproteins of affected persons. Therefore, to explore thebiochemical similarities between CHD and CDGS, we compared the structures of the carbohydrate sidechains of canine serum transferrin isolated from a normal and a CHD-affected dog. Because of the verysmall quantity of serum transferrin available from the CHD-affected dog, we used analytical proceduresthat minimize sample consumption. In this scheme, we used microbore liquid chromatography interfaced toelectrospray tandem mass spectrometry to identify and purify the glycopeptides from the tryptic digest ofcanine serum transferrin. This was followed by a series ofexoglycosidase digestions coupled with massspectrometric detection to sequence the carbohydrate side chains of the glycopeptides. With these pro-cedures we completely characterized the carbohydrate chains attached to serum transferrin isolated fromboth a normal and a CHD-affected dog. However, we found no discernible differences in glycosylation of thecarbohydrate side chains of serum transferrin from these two animals, suggesting that the biochemicaldefect in puppies with CHD differs from that in children with CDGS. # 1998 John Wiley & Sons, Ltd.
Received 27 February 1998; Revised 9 March 1998; Accepted 11 March 1998
A kindred of Bernese Mountain dogs affected with caninehepatocerebellar degeneration (CHD) syndrome has re-cently been identified in northeast Georgia.1 This disease ischaracterized by severe central nervous system disorder,growth retardation and liver dysfunction during infancy.Dogs affected with CHD develop normally until approxi-mately five or six weeks of age and then begin developingtremors, ataxia, dysmetria and weakness. By three monthsof age, the dogs are non-ambulatory, although still mentallyalert and responsive. At necropsy, they display mildcerebellar atrophy and dilated lateral ventricles. Micro-scopically, degeneration and loss of purkinje cells in thecerebellum can be seen, with vacuolation and earlydegenerative changes in the olivary nuclei. Based on itsclinical signs and morphologic features, CHD bears a
striking resemblance to an important emerging disorder inchildren, carbohydrate-deficient glycoprotein syndrome(CDGS).1–3 The apparent similarity between CHD andCDGS was further supported by isoelectric focusing,2 atechnique that is widely used to clinically diagnose childrenwith CDGS.4,5 Therefore, CHD-affected puppies appear tobe a spontaneous animal model for studying human CDGSin children.
CDGS in children was first known as disialotransferrindeficiency syndrome, because the primary biochemicalfinding in CDGS is a deficiency in normal disialylatedserum transferrin with a preponderance of monosialylatedglycoproteins.3–5 Further studies of the purified transferrinin affected children showed that the carbohydrate deficiencyaffects not only sialic acid but also the neutral sugarsgalactose andN-acetylglucosamine.4–7 While providing arapid diagnostic test, the differences in glycoform popula-tions of transferrin isolated from normal and CDGS-affectedchildren may also indicate the severity of the disease.5
Therefore, in order to fully understand the biochemical andmolecular characteristics of CHD and its resemblance toCDGS in children, it is important to characterize the glycanstructures of canine serum transferrin from CHD-affecteddogs and compare them with those from clinically normal,age-matched dogs.
The goal of this work was to fully characterize the carbo-hydrate side chains of canine serum transferrin, including
†Current address: SmithKline Beecham Pharmaceuticals, P.O. Box1539, Mail Code UW2920, King of Prussia, PA 19406-0939, USA.*Correspondence to: R. Orlando, Complex Carbohydrate ResearchCenter, and Departments of Biochemistry & Molecular Biology, andChemistry, University of Georgia, 220 Riverbend Road, Athens, GA30602-4712, USA.Contract/grant sponsor: National Institutes of Health; Contract/grantnumber: (2-P41-RR05351).Contract/grant sponsor: National Science Foundation; Contract/grantnumber: (CHE-9626835).Contract/grant sponsor: University of Georgia Research Foundation.Contract/grant sponsor: Veterinary Medical Experiment Station;Contract/grant number: 29-26GR207-002PR97-011.
CCC 0951–4198/98/090571–09 $17.50 # 1998 John Wiley & Sons, Ltd.
RAPID COMMUNICATIONS IN MASS SPECTROMETRYRapid Commun. Mass Spectrom.12, 571–579 (1998)

identifying the number of glycosylation sites, the site-specific carbohydrate side chain structures, and hetero-geneity of the glycoprotein.We alsocompared the resultsfrom a CHD-affectedpuppy with thosefrom a clinicallynormal, age-matched puppy. The primary diffi culty withthis study involved the small amountof serum transferrinavailable from a CHD-affected puppy, approximately100mg. We therefore used a multiple-step analyticalapproachbaseduponvariousmassspectrometric techniquesthat minimize sampleconsumption. First, the molecularmass of the intact glycoproteinwasdetermined by matrix-assistedlaserdesorption/ionization(MAL DI) MS. Next,theglycopeptides werepurified andidentified from the trypticdigest of the glycoprotein by microbore liquid chromato-graphy interfacedto electrospraytandemmassspectrometry(LC/ESI-MS/MS). In this experiment, the ESI tandemMSwasoperated in amodethatwoulddetect theprecursor ionsof the diagnostic sugaroxoniumion HexNAc� at m/z204,which permits the selective detection of glycopeptides.18
The complete structureof the carbohydrate side chainsofthe glycopeptides were then elucidated by a series ofexoglycosidasedigestionscoupledwith massspectrometricdetection usingMALDI -MS andcapillary LC/ESI-MS/MS.This procedurepermittedthecompletestructural character-ization of the glycoprotein from only 35mg of sample,which is approximatelyonethird of theamount of materialavailable from a single puppy.
EXPERIME NTAL
Materials
TPCK-treatedtrypsinwaspurchasedfrom Sigma(St.Louis,MO, USA). Glycerol-free recombinant N-glycanase wasobtained from Genzyme (Cambridge, MA, USA). S.tymphimuriuma-2-3sialidasewasagift from NewEnglandBiolabs (Beverly, MA, USA). The exoglycosidasesof A.ureafaciens a-2-3,6,8 sialidase, S. pneumoniae b-1-4galactosidase,andS.pneumoniaeb-1-2 N-acetylhexosami-nidase were obtained from Oxford GlycoSystems (Rose-dale, NY, USA).
A 25-cm length of 0.32 mm i.d.� 0.42mm o.d. fusedsilica reversed-phaseC18capillary columnwasconstructedaccording to the published method.9 The fused silicacapillary tubing waspurchasedfrom Polymicro Technolo-gies (Phoenix, AZ, USA), and the Rainin C18 reversed-phasepacking (5 mm particle, 300A pore) was obtainedfrom the SeparationsGroup(Hesperia,CA, USA).
Isolation and purific ation of intact canine serumtransferrin
Caninetransferrin waspurified from a CHD-affectedandaclinically normal, age-matched puppy using strong ion-exchange chromatography and gel filt ration. The purifiedproteinhadamolecular weightof 77kDaasshownby SDS-PAGE, andformeda single line on immunoelectrophoresisusing an antibodyraisedagainst whole canine serum.TheN-terminal amino acid sequence obtained showed signifi-canthomology with that of humanandrabbit transferrin.
Capillary LC/MS analysisof intact canineserumtransferrin
Samples containing about 5 mg of intact canine serumtransferrin, isolatedfrom bothaCHD-affectedandanormal
puppy,were analyzedusing a PE Sciex API-II I (Ontario,Canada)electrospray triple quadrupole mass spectrometerequippedwith an ionspraysource.The samplewas intro-ducedinto themassspectrometer by anLC pumpthroughaloop injector, with the capillary C18 column directlymounted on the spectrometer (ESI-LC/MS). The HPLCgradientwasperformedusing anApplied Biosystems140Bsolventdelivery system(ABI, Foster City, CA, USA). Themobile phases usedfor gradientelution consisted of bufferA (0.1%aqueous TFA) andbuffer B (80%acetonitrile and20% water containing 0.085%TFA). A fast gradientwasusedfor desalting andthefinal elutionof thesample: 0% Bto 60% B linearly in 20min, then linearly to 100% B in10min. The flow from the HPLC pump(120mL/min) wasdivided into 30:1 (waste:LC)by anAccurate high-pressuresplitter (LC Packings, San Francisco, CA, USA). Thisprovideda flow of approximately 4mL/min to the injectorequippedwith a5 mL loop.Thesolventthenpassedthroughthe capillary column,which wasdirectly connected to themassspectrometer.No UV detector was usedduring thecapillaryLC/MS analysis.TheMS wastunedandcalibratedusingamixtureof polypropyleneglycols(PPG).Theorificepotential was set at 70V. The mass spectrometer wasscannedcontinuously from m/z1400 to 2200 with a scanstepof 0.2 anda 2 msdwell time per step.
Reduction and alkylation of canineserum transferrin
Approximately 25mg of canine serum transferrin(�300pmol) isolatedfrom a CHD-affectedand a controlpuppywasdissolved in 50mL buffer at pH 8.2, containing0.5M Tris-HCl, 6 M guanidine hydrochloride, and 2 mMEDTA. Then1.5mL of 18mM dithiothreitol wasaddedtothereduction buffer.Themixturewaspurged with nitrogenandallowed to react for 1 hr at 37°C. After the reduction,5mL of 18mM iodoacetimide wasadded,and the reactionwas allowed to proceedfor one additional hour at roomtemperature. The reduced and alkylated glycoproteinwasdesaltedfrom the reactionbuffer using a microconultra-filter (3000MW cut-off; Amicon, Beverly, MA, USA) andconcentrated to dryness using a Speed-Vac centrifuge(SavantInstruments, Farmingdale,NY, USA).
Trypsin digestion of canine serum transferrin
Reduced and alkylated canine serum transferrin wasdissolvedin 50mL of 50mM NH4HCO3 buffer at pH 8.5,thendigestedfor 18hr at37°C with TPCK-treatedtrypsinatanenzyme:substrateratio of 1:50 (w/w).
Microbor e LC/UV/ESI-MS/MS and MA LDI-MSanalysesof the trypt ic digest of canineserumtransferrin
The tryptic digest solution of canineserum transferrin wasanalyzedusing a 1.0mm� 25cm C-18 Brownleecolumnconnected to a PESciexAPI-III electrospraymass spectro-meter.The HPLC systemwas operated at a flow rate of50mL/min with a gradientstarting from solventA (0.1%aqueousTFA), which wasincreasedlinearly over50min to50%solventB (20%water,80%acetonitrile, 0.085%TFA),thento 100%solvent B in 15min. The post-columneluentwassplit into 9:1 using a low dead-volumetee(UpchurchScientific,OakHarbor, WA, USA) to divert 45mL/min to aUV detector (� = 214nm) and5 mL/min to theelectrosprayMS. All thepeaksappearing in theUV tracewerecollectedmanually. The Sciex API-III mass spectrometer was
Rapid Commun.MassSpectrom.12, 571–579(1998) # 1998JohnWiley & Sons,Ltd.
572 MS SEQUENCING OF GLYCOPROTEINGLYCANS

operatedin thetandemMS modeutili zing theprecursorionscanof the diagnosticsugaroxonium ion of HexNAc� atm/z 204.8 Both the first quadrupole (Q1) and thirdquadrupole(Q3) weretuned andcalibratedusing a mixtureof PPGs.The first quadrupole was scannedcontinuouslyfrom m/z900 to 1900with a scanstep of 1 Da and10 msdwell time perstep.To maximize the ion transmissionandenhance sensitivity, the analyzing quadrupole (Q1) wasoperatedatunit massresolution (50%peakvalleydefinitionat m/z520),while the mass selectingquadrupole(Q3) wasset to passa 2–3Da window aroundm/z204. High purityargonwasusedasthecollision gas,andtheorifice potentialwas set at 55V. The final optimization of the instrumentparameters in precursor-ion scan mode, including thequadrupole offsets and collision gas thickness, wasaccomplished using N-linked glycopeptides isolatedfrombovine fetuin.
The intact glycoproteinandthe glycopeptide-containingHPLCfractionswerealso analyzedusing aHewlett-PackardLDI-1700 XP (Palo Alto, CA, USA) MALDI-TOF massspectrometer. The instrumentwasoperatedat anaccelerat-ing voltageof 30kV, an extractorvoltageof 9 kV, and apressure of �9� 10ÿ7 torr. The sample was desorbed/ionized from the probe tip with a nitrogen laser (� = 337nm) having a pulse width of 3 ns and delivering �12mJenergy/laserpulse.Sinapinicacid was usedas the matrix,andtheinstrument wascalibratedexternally with a mixtureof knownpeptides.
N-Glycanaseand exoglycosidasedigestionof GP-I andGP-II
Theglycopeptide-containing HPLC fractionswere concen-trated to 50mL to hold the final concentration of theglycopeptideat approximately 5 pmol/mL. An aliquotof theglycopeptide(10pmol) was dried on a Speed-Vac centri-fugeandthenreconstituted in 3mL of 25mM NH4HCO3 atpH 7.8.Theglycopeptidewastreatedfor 18hr at37°C with0.1 mU of N-glycanase (0.25 mU/mL), and the resultingdigestionmixture wasanalyzedby MALDI -MS.
A seriesof exoglycosidasedigestionswasperformedwiththeenzymeslisted in Table1. Theexoglycosidasedigestionprotocol was modified: the digestion buffer of 100mMsodium acetate suggested by Oxford GlycoSystemswasreplaced by a volatile 25mM ammoniumacetate solution.Ourprior experimentdemonstratesthata25mM ammoniumacetate buffer both retains the specific activity of theenzyme and allows direct MALDI -TOF analysis of thedigestionmixture without desalting.10 Thespecificitiesandactivities of the exoglycosidaseslisted in Table 1 wereconfirmedby digestingtheglycopeptidespurified from thetryptic digestof bovinefetuin,which is awell-characterizedglycoprotein.Theexoglycosidasedigestioncannormally becompleted in 18 hr at 37°C, provided that the digestionbuffer with theoptimum pH andthe enzyme concentration
are maintained. After exoglycosidase digestion, bothMALDI and capillary LC/MS/MS were used to analyzethe resultingdigestionmixture.
Theglycosylated tryptic fragmentGP-II wassequentiallydigestedwith theexoglycosidasesof S.tymphimuriuma-2-3sialidase,A. ureafaciensa-2-3,6,8sialidase, S.pneumoniae-b-1-4galactosidase,andS. pneumoniaeb-1-2N-acetylhexo-saminidase. The sequentialexoglycosidasedigestionsofGP-II wereperformedin thefollowingway.First,analiquotof GP-II (�40pmol) was dissolved in 5 mL of digestionbuffer and S. tymphimurium a-2-3 sialidasewas added.Af ter overnightdigestion, 0.5mL of the digestionsolutionwas removed for direct MAL DI analysis, while A.ureafaciensa-2-3,6,8sialidasewasaddedto the remainderof the solution to start the second round of digestion,followed by MALDI analysis, and so on. However, if thenextdigestionrequiredtheuseof a buffer with anotherpH,the digestion solution was dried with a Speed-Vac andreconstituted in the buffer with the different pH.
A seriesof four digestionsusingdifferent combinationsof exoglycosidaseswereperformed in parallel to sequencethe N-linked glycan of glycosylatedtryptic fragmentGP-I.In the first experiment, only S. tymphimurium a-2-3sialidase was addedto the exoglycosidasedigest. In thesecond experiment, A. ureafaciensa-2-3,6,8 sialidasewithmore generalspecificity wastheonly exoglycosidaseused.In the third experiment, A. ureafaciensa-2-3,6,8 sialidasewas combined with S. pneumoniaeb-1-4 galactosidasetoremoveNeuAc andb-1-4-linkedGal residues from thenon-reducing termini of the glycan of GP-I. In the fourthexperiment, A. ureafaciensa-2-3,6,8sialidasewas furthercombinedwith S.pneumoniaeb-1-4galactosidaseandb-N-acetylhexosaminidaseto cleave NeuAc, b-1-4-linked Gal,and b-1-2-linked GlcNAc residuesconcurrently from thenon-reducingtermini of thecarbohydrate sidechainof GP-I. Eachdigestionusedapproximately 5–10pmol of sampleandwasperformed in 4 mL of buffer solution. In the thirdand fourth digestions, a buffer with the pH that compro-misestheconditionsfor all enzymesusedwaschosen asthedigestion solution. After each digestion, the resultingdigestion mixture was analyzedby capillary LC/MS/MSutili zing a precursorion scanof HexNAc� at m/z204.
RESULTS AND DISCUSSION
Determination of the molecular massof intact canineserum transferrin
The first stepof our analytical schemewasto analyzethetwo intact glycoproteinsamples primarily to evaluate thepurity andcleanlinessof the samples, sincethese arebothcritical factors in all of the subsequent procedures. Forexample, if the samples containedmultiple glycoproteins,theglycosylation sitemappingprocedurewould havegivenspuriousresults,while thepresenceof contaminantssuchassalts and detergents may have caused problems withsubsequent analytical procedures. In addition, this stepmay haveprovidedevidencethat glycosylation is differentbetween theCHD-affectedandnormalpuppiesby detectingdifferences in the molecular massof the two samplesofcanine serum transferrin. The questionthenshiftedto howto perform this analysis, i.e. MALDI-MS or ESI-MS. WechoseMALDI -MS because it is bettersuitedto analyzingvery heterogeneousglycoproteins.11
A 5 mg sample of intactserum transferrinfrom theCHD-
Table 1. The specificity and experimental conditions for theexoglycosidasedigestions
Enzyme SourceEnzymeconc.
(mU/mL)Buffer
pH Linkagespecificity
Neuraminidase S.tymphimurium 500 6.0 NeuAca-2-3Sialidase A. ureafaciens 1 5.0 NeuAca-2-3/6b-Galactosidase S.pneumoniae 0.2 6.0 Gal b-1-4b-GlcNAcase S.pneumoniae 0.009 6.0 GlcNAc b-1-2
# 1998JohnWiley & Sons,Ltd. Rapid Commun.MassSpectrom.12, 571–579(1998)
MS SEQUENCING OF GLYCOPROTEIN GLYCANS 573
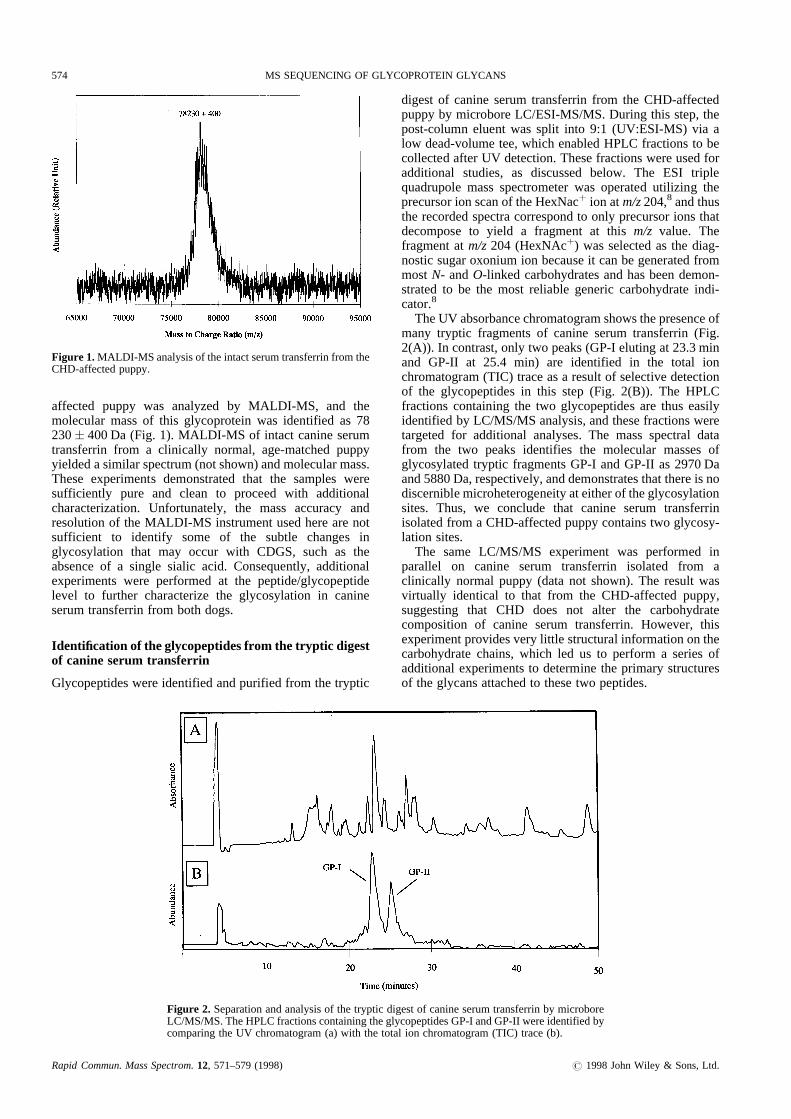
affected puppy was analyzed by MALDI-M S, and themolecular massof this glycoprotein was identified as 78230� 400Da (Fig. 1). MALDI -MS of intact canineserumtransferrin from a clinically normal, age-matchedpuppyyieldedasimilar spectrum (notshown) andmolecular mass.These experiments demonstratedthat the samplesweresufficiently pure and clean to proceed with additionalcharacterization. Unfortunately, the mass accuracy andresolution of the MAL DI-MS instrumentusedherearenotsufficient to identify some of the subtle changes inglycosylation that may occur with CDGS, such as theabsence of a single sialic acid. Consequently, additionalexperiments were performedat the peptide/glycopeptidelevel to further characterize the glycosylation in canineserum transferrin from bothdogs.
Identification of the glycopeptidesfrom the tryptic digestof canineserum transferrin
Glycopeptideswereidentified andpurifiedfrom thetryptic
digestof canineserum transferrin from the CHD-affectedpuppyby microboreLC/ESI-MS/MS. During this step, thepost-column eluent was split into 9:1 (UV:ESI-MS) via alow dead-volumetee,which enabledHPLC fractionsto becollected afterUV detection.Thesefractionswereusedforadditional studies, as discussed below. The ESI triplequadrupole massspectrometer was operated utilizing theprecursor ion scanof theHexNac� ion atm/z204,8 andthusthe recordedspectracorrespondto only precursorions thatdecompose to yield a fragment at this m/z value. Thefragmentat m/z204 (HexNAc�) wasselectedasthe diag-nosticsugaroxonium ion becauseit canbegeneratedfrommostN- andO-linked carbohydratesandhasbeendemon-stratedto be the most reliable genericcarbohydrateindi-cator.8
TheUV absorbancechromatogramshowsthepresenceofmany tryptic fragmentsof canineserumtransferrin (Fig.2(A)). In contrast,only two peaks(GP-I elutingat 23.3minand GP-II at 25.4 min) are identified in the total ionchromatogram(TIC) traceasa result of selective detectionof the glycopeptides in this step (Fig. 2(B)). The HPLCfractionscontaining the two glycopeptides are thus easilyidentifiedby LC/MS/MS analysis,andthesefractionsweretargetedfor additional analyses. The massspectral datafrom the two peaks identifies the molecular massesofglycosylatedtryptic fragments GP-I andGP-II as2970Daand5880Da,respectively, anddemonstratesthatthereis nodiscerniblemicroheterogeneityateitherof theglycosylationsites. Thus, we conclude that canine serum transferrinisolatedfrom a CHD-affectedpuppycontainstwo glycosy-lation sites.
The same LC/MS/MS experiment was performed inparallel on canine serum transferrin isolated from aclinically normal puppy (datanot shown). The result wasvirtually identical to that from the CHD-affected puppy,suggesting that CHD does not alter the carbohydratecomposition of canine serum transferrin. However, thisexperimentprovidesvery little structural informationonthecarbohydrate chains,which led us to perform a seriesofadditionalexperimentsto determine the primary structuresof the glycansattachedto thesetwo peptides.
Figure 1. MALDI-MS analysisof theintactserumtransferrinfrom theCHD-affectedpuppy.
Figure 2. Separationandanalysisof the tryptic digestof canineserumtransferrinby microboreLC/MS/MS.TheHPLCfractionscontainingtheglycopeptidesGP-IandGP-II wereidentifiedbycomparingthe UV chromatogram(a) with the total ion chromatogram(TIC) trace(b).
Rapid Commun.MassSpectrom.12, 571–579(1998) # 1998JohnWiley & Sons,Ltd.
574 MS SEQUENCING OF GLYCOPROTEINGLYCANS
Sequencing the oligosaccharide chain of GP-II(Mr = 5088Da)
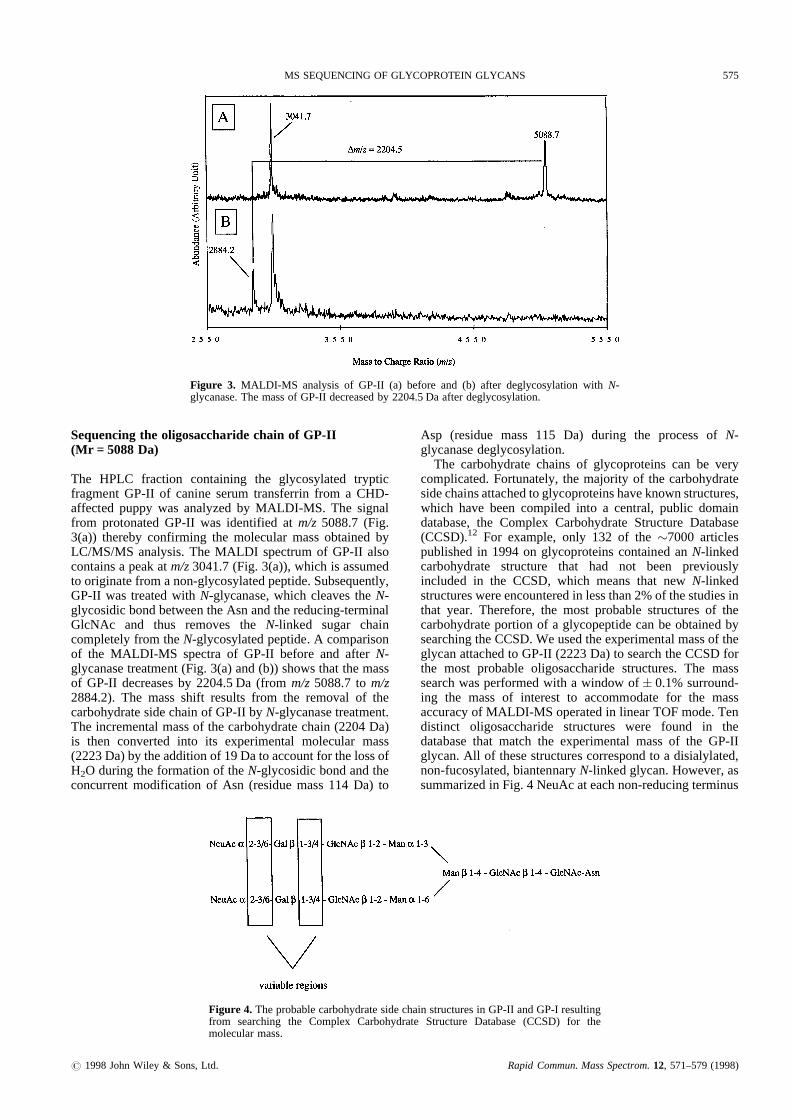
The HPLC fraction containing the glycosylated trypticfragment GP-II of canine serumtransferrin from a CHD-affected puppy was analyzed by MALDI-MS. The signalfrom protonatedGP-II was identified at m/z 5088.7(Fig.3(a)) therebyconfirming the molecular mass obtainedbyLC/MS/MS analysis.The MALDI spectrum of GP-II alsocontainsa peakat m/z3041.7(Fig. 3(a)),which is assumedto originate from anon-glycosylatedpeptide.Subsequently,GP-II was treatedwith N-glycanase,which cleavesthe N-glycosidicbondbetween theAsn andthereducing-terminalGlcNAc and thus removes the N-linked sugar chaincompletely from theN-glycosylatedpeptide. A comparisonof the MALDI -MS spectraof GP-II before and after N-glycanasetreatment (Fig. 3(a)and(b)) showsthat themassof GP-II decreases by 2204.5Da (from m/z5088.7 to m/z2884.2). The massshift results from the removal of thecarbohydratesidechainof GP-II by N-glycanasetreatment.The incrementalmassof thecarbohydratechain(2204Da)is then converted into its experimental molecular mass(2223Da)by theadditionof 19Dato accountfor thelossofH2O duringtheformation of theN-glycosidic bondandtheconcurrent modification of Asn (residue mass 114 Da) to
Asp (residue mass 115 Da) during the processof N-glycanasedeglycosylation.
The carbohydrate chainsof glycoproteinscan be verycomplicated.Fortunately,the majority of the carbohydratesidechainsattachedto glycoproteinshaveknownstructures,which have beencompiled into a central, public domaindatabase,the Complex Carbohydrate Structure Database(CCSD).12 For example,only 132 of the �7000 articlespublished in 1994on glycoproteinscontained an N-linkedcarbohydrate structure that had not been previouslyincluded in the CCSD, which meansthat new N-linkedstructureswereencounteredin lessthan2%of thestudiesinthat year. Therefore, the most probable structures of thecarbohydrate portion of a glycopeptide canbe obtained bysearchingtheCCSD.We usedtheexperimental massof theglycanattachedto GP-II (2223Da) to searchtheCCSDforthe most probable oligosaccharide structures. The masssearch wasperformedwith a window of� 0.1%surround-ing the mass of interest to accommodate for the massaccuracyof MALDI-MS operatedin linearTOF mode. Tendistinct oligosaccharide structures were found in thedatabasethat match the experimental massof the GP-IIglycan.All of thesestructures correspondto a disialylated,non-fucosylated,biantennaryN-linked glycan.However, assummarizedin Fig. 4 NeuAcat eachnon-reducing terminus
Figure 3. MALDI-MS analysisof GP-II (a) before and (b) after deglycosylationwith N-glycanase.Themassof GP-II decreasedby 2204.5Da after deglycosylation.
Figure 4. Theprobablecarbohydratesidechainstructuresin GP-II andGP-I resultingfrom searching the Complex CarbohydrateStructure Database(CCSD) for themolecularmass.
# 1998JohnWiley & Sons,Ltd. Rapid Commun.MassSpectrom.12, 571–579(1998)
MS SEQUENCING OF GLYCOPROTEIN GLYCANS 575

is reported to be eithera–2-3- or a–2-6-linked to Gal, andGal is eitherb–1-4-or b–1-3-linked to GlcNAc.
A seriesof exoglycosidasedigestions was designed tocharacterizetheoligosaccharidechainin GP-II on thebasisof the structure of a disialylated biantennary N-linkedglycan.Thisprocessuseshighly specificexoglycosidasestoselectively release monosaccharidesfrom thenon-reducingtermini basedon their stereochemistry, anomericconfig-uration, and linkage to the remainderof the carbohydratesidechain.10,13–15After eachexoglycosidasetreatment,thedigestionproductwasanalyzedby MS to determinetheshiftin molecularmassof the glycopeptide, which providesthenumber of monosaccharidesreleased.
GP-II wassequentially digestedwith theexoglycosidasesof S. tymphimurium a-2-3 sialidase, A. ureafaciensa-2-3,6,8 sialidase,S. pneumoniaeb-1-4 galactosidase,and S.pneumoniaeb-1-2N-acetylhexosaminidase.Duringthefirstdigestion,S.tymphimuriumsialidasehadno effectonGP-II(Fig. 5(b)), suggesting that NeuAc is not a-2-3-linked toGal. The second digestionconsistedof treatment with A.ureafaciensa-2-3,6,8 sialidase, decreasingthe molecularmass of GP-II by 582Da (from m/z5089.5to 4508.3). Thisdecreaseis indicative of therelease of two NeuAc residues,which, basedon the specificity of this exoglycosidase,canbelinkeda-2-3, a-2-6, and/ora-2-8 to theremainderof theglycan (Fig. 5(c)). As shown by the first digestion (Fig.5(b)), theNeuAcresidues in GP-II cannotbeattachedby a-2-3 linkages. Furthermore, the CCSDsearchrevealed thatNeuAc residues havenot beenpreviously foundattachedtoglycansof thismolecular massvia a-2-8 linkages.Thus,we
concludedthat the two NeuAcresidues area-2-6-linked tothe Gal residues
Thethird digestionwasperformedwith S.pneumoniaeb-1-4 galactosidase and caused the molecular mass ofdesialylated GP-II to decreaseby �324Da (from m/z4508.3to 4184.2). This massshift indicatesthe lossof two-1-4Gal residues(Fig. 5(d))andthereforedemonstratesthatthe two Gal residuesin GP-II areb-1-4-linked to GlcNAc.During the fourth experiment, S. pneumoniae b-1-2 N-acetylhexosaminidasewasaddedto thedigestion,andGP-IIyieldedanothermassshift of �406Da (from m/z4184.2to3778.1).This decreasecorrespondsto the lossof two b-1-2GlcNAc residues(Fig. 5(e))andconfirmsthatbothGlcNAcresiduesin the chains of GP-II are attached to the coreoligosaccharide by b-1-2 linkages. The sequentialexogly-cosidasedigestionshavethus cleavedthe glycan in GP-IIdown to the corepentasaccharideof the N-linked carbohy-dratesidechain.In summary,thecarbohydratesidechainofGP-II from canineserum transferrin isolatedfrom a CHD-affectedpuppyis determinedto bethea-2-6disialoGalb-1-4-GlcNAc b-1-2 biantennary, non-fucosylated, complex-type N-linked oligosaccharide (Fig. 6).
This same set of digestionswas usedto show that theoligosaccharide chain in GP-II of canineserumtransferrinisolatedfrom theclinically normalpuppywasalsoof thea-2-6 disialo b-1-4-GlcNAc b-1-2 biantennary, non-fucosy-lated,complex-typeN-linked structure(Fig. 6).
Sequencing the oligosaccharide chain of GP-I(Mr = 2970Da)
We attempted to sequencethe glycan of the glycosylatedtryptic fragmentGP-1isolatedfrom a CHD-affectedpuppyusing an experimental approach similar to that illustrated.However, GP-I could not be detected by MALDI-MSanalysisof its corresponding HPLC fraction (Fig. 7(a)),presumably due to the presenceof severalpeptides in thisfraction, combined with the tendency of non-glycosylatedpeptidesto bepreferentially ionizedin amixtureof peptidesandglycopeptides.
GP-I wastreatedwith N-glycanase,andthe correspond-ing MALDI spectrum is shown in Fig. 7(b). A newpeakatm/z766.2 is identified, and it corresponds to the deglyco-sylatedform of GP-I. As determined by ESI-MS/MS, theprotonatedform of GP-I hasa molecular massof 2971Da.Therefore, the mass shift of GP-I resulting from theremovedN-glycan was determined to be 2204.8Da (from2971Da to 766.2). The result from the subsequent masssearch(Mr = 2224� 2 Da) of the CCSDsuggests that thecarbohydratesidechain in GP-I is of the disialylated,non-fucosylated,biantennary,complex-type N-linked structure,which is identical to the glycan in GP-II.
A seriesof exoglycosidasedigestionswasthenperformedto elucidate the linkages and anomeric centers of theperipheralNeuAc andGal residues presentin GP-I. SinceGP-I wasnot detectedby MAL DI from the corresponding
Figure 5.MALDI analysisof (a) intactGP-II,andafterexoglycosidasetreatmentwith (b) a-2-3sialidase,(c) a-2-3,6,8sialidase,(d) a-2-3,6,8sialidaseand b-1-4 galactosidase,and (e) a-2-3,6,8sialidase,b-1-4galactosidase,andb-1-2 GlcNAcase.
Figure 6.Thededucedstructureof thecarbohydratesidechainsof GP-I andGP-II in canineserumtransferrin.
Rapid Commun.MassSpectrom.12, 571–579(1998) # 1998JohnWiley & Sons,Ltd.
576 MS SEQUENCING OF GLYCOPROTEINGLYCANS

HPLC fraction, capillary LC/MS/MS, due to its highselectivity, was chosen as the MS detectionmethod forthe analysis of exoglycosidasedigestion product. BothMALDI and capillary LC/MS/MS are very sensitive MStechniques.MALDI providesa rapid and facile meanstoobtain rough qualitative identification of many differentkinds of samples.However, very diversemixturestend toproducesignalsuppressionof somecomponentsin MALDI-MS. For a complex glycopeptide/peptide mixture, theglycopeptideis very often suppressed relative to peptides.In contrast, the glycopeptide can easily be detectedbytandemMS utilizing theprecursor ion scanof HexNAc� atm/z204.The useof LC/MS/MS alsoautomatically desaltsthe sample during analysis,should any buffer bepresent.
Unlike the sequential digestions(i.e. the additionof oneenzymeatatime in theexperimentusedto sequenceGP-II),aseriesof four digestionswasperformedin parallelto probethe structure of the carbohydrate sidechainon GP-I. Thiswasdoneto decreasethe overall experimental time. Af terthe digestion, all the digestsolution was injected into theMS for analysis. Since each exoglycosidase can onlyremove one kind of monosaccharide and each digestionresults in acertainamountof massshift of theglycopeptide,oneneedonly monitor the masswindows surrounding theexpecteddigestionproduct. Forexample,afterthedigestionby sialidase, theglycopeptide GP-I (Mr = 2970)is expectedto loseeithernone,one,or two sialic acid residues,whichwould decreasethe massof GP-I by 0, 291, or 582 Da,respectively.Therefore,only themasswindows(6 Dawide)around theseexpectedions (doubly charged ions at m/z1195,1341,and1486,and triply chargedions at m/z797,894,and991) wereselectively monitoredduring this step.The selectedion monitoring (SIM) techniqueundoubtedlyprovidesbettersensitivity for capillary LC/MS/MS analysis,sincemoretimeis spentdetecting theionsof interest in eachscanwithoutextending the total scantime.
The reconstructed molecular massplots from the ESI-MS/MS analysis of GP-I after a seriesof exoglycosidasearraydigestionsareshown in Fig. 8. Most of theglycansinGP-I had two sialic acids attached (Mr = 2970), while a
small percentage had only one sialic acid attached(Mr = 2680,Fig. 8(a)).Digestion with S. tymphimuriuma-2-3sialidaseresultedin nocarbohydratesidechain cleavage(Fig. 8(b)), while digestionwith A. ureafaciensa-2-3,6,8sialidasecausedthe loss of two NeuAc residues (582Da,from Mr 2970to 2388,Fig. 8(c)).Theseresultssuggestthatbothof theNeuAc residuesareattachedvia a-2-6 linkages,asdiscussed abovefor GP-II. Digestion with A. ureafaciens-2-3,6,8sialidaseandS.pneumoniaeb-1-4galactosidase,inthethird experiment(Fig. 8(d)) decreasedthemassof GP-Iby 906Da (from Mr 2970 to 2064) from the loss of twoNeuAc (582Da) andtwo b-1-4 Gal (324Da) residues.Theresults from this experiment demonstrate the both Galresidues areattachedby b-1-4 linkages. Digestion with theA ureafaciens a-2-3,6,8, sialidase,S. pneumoniae b-1-4galactosidase,and b-1-2 N-acetylhexosaminidase in thefourth experiment(Fig. 8(e))decreased themassof GP-I by1312 Da (from Mr 2970 to 1658) from the release of twoNeuAc (582 Da), two b-1-4 Gal (324 Da), and two b-1-2GlcNAc (406 Da) residues. This last experiment demon-strates that the 2 GlcNAc resides are attached to thepentasaccharide core by b-1-2 linkages. Therefore, thecarbohydrate chain of GP-I of canine serum transferrinisolatedfrom aCHD-affectedpuppywasidentifiedasana-2-6 disialo Gal b-1-4-GlcNAc b-1-2 biantennary, non-fucosylated,complex-type oligosaccharide (Fig. 6). This isthesamecarbohydratestructurefoundon GP-II (Figs 5 and8).Furthermore,theoligosaccharidechainof GP-I of canineserum transferrin isolated from a clinically normal puppywas also determined to be of a-2-6 disialo Gal b-1-4-GlcNAc b-1-2 biantennary,non-fucosylated,complex-typestructure.
Analyzing the intact glycoproteins by ESI-MS
The apparentlack of carbohydrate heterogeneity in theserum transferrin from the two sourcessuggests that directESI-MS analysisof the intact glycoproteinshouldbe fairlystraightforward. Furthermore,this techniquehasthelevel ofmass accuracy and resolution to determine very subtle
Figure 7. MALDI-MS analysisof GP-I (a)beforeand(b) afterdeglycosylationby N-glycanasetreatment.A newpeakat m/z766.2wasidentified asthe deglycosylatedform of GP-I.
# 1998JohnWiley & Sons,Ltd. Rapid Commun.MassSpectrom.12, 571–579(1998)
MS SEQUENCING OF GLYCOPROTEIN GLYCANS 577

changes in glycosylation. ESI-MS analysisof intact canineserum transferrin from a CHD-affected puppy showedasingle seriesof peaks in the m/z region of 1400–2200,confirming that thereis virtually no microheterogeneity intheoligosaccharidesidechains(Fig. 9).Themolecular mass
of canineserumtransferrin as determined by ESI-MS isshownat 78 316� 3.1Da (inset in Fig. 9). Intact canineserum transferrin from a clinically normal, age-matchedpuppywasalsoanalyzedby ESI-MS(spectrumnot shown).Essentiallythe sameion distribution patternandmolecularmasswere found as in the ESI-MS spectrum of canineserum transferrin from a CHD-affected puppy (Fig. 9).Theseresults further support the finding that there is nodifferencein glycosylationbetween the CHD-affectedandthe clinically normalpuppies.
CapillaryLC/MS waschosen overdirect infusionfor theESI analysis of the intact glycoprotein, as it offers severaladvantages.16 Comparedwith direct infusion,capillary LC/MS requiresaboutfivefold less total sample. In fact, directinfusion requires the injection of at least15mL of samplesolution of low pmol concentration, while only 1–3mL ofsamplesolution of the sameconcentration is neededforcapillary LC/MS analysis, which alsotypically results in aspectrum of higher quality. Additionally, the desaltingcapabilities of reversed-phaseHPLC allow us to obtain aspectrumof the glycoprotein free from salt interference.Moreover, the retention of thesampleon theC18capillarycolumnandits subsequentelutionasa narrowpeakprovidea way to concentratethe sample.
CONCLUSIONS
Two N-linked glycosylation sites have beenidentified incanineserumtransferrin by LC/ESI-MS/MS analysis of thetrypsin digestion product of glycoprotein. A series ofexoglycosidasedigestionscoupledwith massspectrometricdetection (MALDI and capillary LC/ESI-MS/MS) haveshownthat the predominant form of the carbohydrate sidechainsattachedto bothof theseglycosylationsiteshavethe-2-6 disialo Gal b-1-4-GlcNAc b-1-2 biantennary, non-fucosylated, complex-type structure (Fig. 6). To ourknowledge, the present work is the first study into theglycanstructureof canineserum transferrin.PreviousNMRstudieshave shown that the biantennary structure(a-2-6disialo Gal b-1-4-GlcNAc b-1-2 biantennary, non-fucosy-
Figure 8. The reconstructedmolecularmassplots from the capillaryLC/ESI-MS/MSanalysisof (a) intact GP-I, andafter exoglycosidasetreatmentwith (b) a-2-3sialidase,(c) a-2-3,6,8sialidase,(d) a-2-3,6,8sialidaseand b-1-4 galactosidase,and (e) a-2-3,6,8sialidase,b-1-4galactosidase,andb-1-2 GlcNAcase.
Figure 9. Capillary LC/ESI-MSanalysisof the intact serumtransferrinfrom theCHD-affectedpuppy.The reconstructedmolecularmassof the glycoproteinis shownin theinset.
Rapid Commun.MassSpectrom.12, 571–579(1998) # 1998JohnWiley & Sons,Ltd.
578 MS SEQUENCING OF GLYCOPROTEINGLYCANS

lated,complex-type) is themajor glycoform (85%to 90%)in human serumtransferrin.17,18
No discernibledifferenceswerefoundin thestructuresofthe carbohydrate side chainsof canine serum transferrinisolated from a CHD-affected and a clinically normalpuppy.Our result thus suggeststhat CHD in puppiesandCDGS in children do not have similar biochemical ormolecular origins. However, asidefrom transferrin, manyother serum glycoproteins have also beenreported to beabnormal with respect to their glycoform populations inCDGS-affected children, and these include glycoproteinhormones, complement factors, enzymes and enzymeinhibitors.3–7 Therefore, more research involving compara-tive structural studies on the other serum glycoproteinsisolatedfrom normal andCHD-affectedpuppiesis requiredbeforea final determinationcanbemadeof thesimilaritiesbetween CHD in puppiesandCDGSin children.
Acknowledgements
This work was supportedby grantsfrom the National InstitutesofHealth(2-P41-RR05351,RO),theNationalScienceFoundation(CHE-9626835,RO),Universityof GeorgiaResearchFoundation(RO),andthe VeterinaryMedical ExperimentStation(29-26GR207-002PR97-011,KPC).
REFERENCES
1. K. P. Carmichael,M. A. Miller, B. E. Miller, C. A. Rawlings,A.
Fisher and J. E. Oliver, J. Am. Med. Vet. Assoc.208, 166–168(1996).
2. K. P. Carmichael,R. Orlando, K. Arbtan, M. Miller and J. E.Oliver, Vet.Pathol.32, 575(1995).
3. J. Jaeken,B. Hagbergand P. Stromme,Acta. Paediatr. Scand.Suppl.375,6–13(1991).
4. J. Jaeken,H. G. vanEijk, C. vanderHeul, L. Corbeel,R. EeckelsandE. Eggermont,Clin. Chim.Acta.144,245–247(1984).
5. J. Jaeken,H. Carchonand H. Stibler, Glycobiology3, 423–428(1993).
6. K. Yamashita,T. Ohkura,H. Ideo, K. Ohno and M. Kanai, J.Biochem.114,766–769(1993).
7. K. Yamashita,H. Ideo, T. Ohkura,K. Fukushima,I. Yuasa,K.OhnoandK. Takeshita,J. Biol. Chem.268,5783–5789(1993).
8. S.A. Carr,M. J.HuddlestonandM. F. Bean,ProteinSci.2, 183–196(1993).
9. M. T. DavisandT. D. Lee,Protein Sci.1, 935–944(1992).10. Y. YangandR. Orlando,Anal. Chem.68, 570–572(1996).11. R. Orlando and Y. Yang, ‘Analysis of glycoproteins’ in Mass
Spectrometryof Biological Materials, B. S. Larsen and C. N.McEwen,(Eds)Marcel Dekker,New York, 215–246(1998).
12. S. DoubetandP. Albersheim,Glycobiology 2, 505–507(1992).13. C. W. Sutton,J.A. O’Neill andJ.S.Cottrell,Anal.Biochem.218,
34–46(1994).14. G.D. Roberts,W. P.Johnson,S.Burman,K. R.AnumulaandS.A.
Carr,Anal. Chem.67, 3613–3625(1995).15. P. A. Schindler,C. A. Settiner,X. Collet, C. J. FieldingandA. L.
Burlingame,Protein Sci.4, 791–803(1995).16. H. Zhang,P.E. AndrenandR. M. Caprioli,J. Mass.Spectrom.30,
1768–1771(1995).17. K. Yamashita,N. Koide,T. Endo,Y. Iwaki andA. Kobata,J. Biol.
Chem.264,2415–2423(1989).18. D. Fu andH. vanHalbeek,Anal. Biochem.206,53–63(1992).
# 1998JohnWiley & Sons,Ltd. Rapid Commun.MassSpectrom.12, 571–579(1998)
MS SEQUENCING OF GLYCOPROTEIN GLYCANS 579