ion channels in t lymphocytes: an update on facts, mechanisms and therapeutic targeting in...
TRANSCRIPT

C
It
Za
b
U
a
ARRAA
KITII
1
vrmntbvihip
tct[ad
oH
0d
Immunology Letters 130 (2010) 19–25
Contents lists available at ScienceDirect
Immunology Letters
journa l homepage: www.e lsev ier .com/ locate /
urrent views
on channels in T lymphocytes: An update on facts, mechanisms and therapeuticargeting in autoimmune diseases
oltan Vargaa, Peter Hajdua, Gyorgy Panyia,b,∗
University of Debrecen, Department of Biophysics and Cell Biology, 98. Nagyerdei krt, Debrecen 4032, HungaryCell Biology and Signaling Research Group of the Hungarian Academy of Sciences, Department of Biophysics and Cell Biology, Research Center for Molecular Medicine,niversity of Debrecen, Debrecen 4032, Hungary
r t i c l e i n f o
rticle history:eceived 25 November 2009eceived in revised form 8 December 2009ccepted 10 December 2009
a b s t r a c t
During the last quarter of a century a large body of evidence was gathered about the involvement ofion channels in T lymphocyte activation. A series of remarkable findings promoted T cell ion channelsto become potential pharmaceutical targets in the therapy of autoimmune disorders. Numerous com-
vailable online 21 December 2009
eywords:on channelcell activation
mmunosuppression
prehensive reviews describe the types of ion channels found in the plasma membrane of T cells andtheir roles in signaling pathways leading to activation, the changes in the expression of these channelsbrought upon by differentiation to various T cell subsets, the formation and possible functions of signalingmolecular clusters that include ion channels in the immunological synapse, the discovery and refinementof structurally different ion channel blockers and the successful in vivo application of such compoundsto suppress hypersensitivity reactions and autoimmune processes. In this review we wish to provide a
opics
mmunological synapse concise update on these t. Ion channels in T cells
The first ion channel described in T lymphocytes was theoltage-gated Kv1.3 potassium channel, which has a fundamentalole in the activation pathway and presently also seems to be theost important target to control T cell activation [1,2]. Kv1.3 chan-
els are activated by membrane depolarization over −60 mV andheir open probability steeply increases with more positive mem-rane potentials. The opening of Kv1.3 channels at a depolarizedoltage exceeding their activation threshold creates a hyperpolar-zing K+ efflux resulting in a negative feedback loop. Due to the veryigh electrical resistance of the lymphocyte membrane, the open-
ng of a few of these channels is sufficient to regulate the membraneotential.
The other plasma membrane potassium channel of T cells ishe Ca2+ activated KCa3.1 (formerly called IKCa1, for intermediateonductance Ca2+-activated K+ channel), which is activated when
he free intracellular Ca2+ concentration rises above about 200 nM3]. Thus, at resting Ca2+ levels these channels are silent and arectivated only during Ca2+ signaling, when they counteract theepolarizing effect of Ca2+ influx on the membrane potential by∗ Corresponding author at: Department of Biophysics and Cell Biology, Universityf Debrecen, Medical and Health Science Center, 98. Nagyerdei krt, Debrecen 4012,ungary. Tel.: +36 52 412 623; fax: +36 52 412 623.
E-mail address: [email protected] (G. Panyi).
165-2478/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.imlet.2009.12.015
from recent years, highlighting the most notable developments.© 2009 Elsevier B.V. All rights reserved.
allowing K+ efflux. Ca2+ sensing is accomplished by calmodulinmolecules bound to the C terminus of KCa3.1 subunits [4] (Fig. 1).
Besides these two potassium channels several other channeltypes have been demonstrated or suggested to be expressed byT cells. These include the Ca2+ release-activated Ca2+ (CRAC) chan-nel, the swelling-activated Cl− channel, various members of the TRPchannel family, two-pore K+ channels and even voltage-gated Na+
and Ca2+ channels (reviewed in [5]). The function of some of thesechannels is questionable, but the existence and involvement of theCRAC channel in T cell activation has been known for a long time[6,7]. Despite this, its molecular identity and the coupling betweenendoplasmic reticulum (ER) emptying and activation of the chan-nel remained a mystery up until a few years ago. As it turned out,two proteins, named Orai1 and STIM1 are necessary to produceCRAC current. Orai1 is the pore forming subunit of the channel,while STIM1 is the sensor of Ca2+ depletion in the ER lumen [8–11].STIM1 is found in the ER membrane with an EF-hand in the ERlumen, while Orai1 is located in the plasma membrane mostly asa dimer [12]. Upon store emptying STIM1 senses the drop in Ca2+
concentration and relocalizes to puncta closely associated with theplasma membrane. This translocation brings Orai1 dimers togetherto form tetrameric active CRAC channels that allow the influx of
2+
Ca .The concerted action of the two K+ channels and the CRACchannel achieves a precisely controlled and long lasting Ca2+ sig-nal that is required for the execution of the activation pathway.CRAC channels provide the entry route for Ca2+ while tightly

2 logy Le
rt
2
stsitwsst
bTscemwimbtmirdmbf
scCftiuKintKtdpltraotKsmu
s((l
0 Z. Varga et al. / Immuno
egulated K+ efflux maintains a permissive membrane potential forhe prolonged Ca2+ influx.
. Kv1.3 in autoimmunity: a potential therapeutic target
The sensitivity of the membrane potential and that of the Ca2+
ignal to blockers of the lymphocyte K+ channels immediatelyurned the attention to the application of these molecules for theuppression of immune responses in autoimmune conditions. Andeal immunosuppressant would specifically inhibit the prolifera-ion of T cells being responsible for autoimmune tissue damage butould leave protective immune reactions unharmed. The expres-
ion of Kv1.3 and KCa3.1 channels in naïve, unstimulated cellseemed to contradict the applicability of Kv1.3 or KCa3.1 inhibitorso achieve specific suppression of autoimmune reactions.
This apparent contradiction has been resolved in the last decadey the discovery of the subset specific expression of ion channels inand B lymphocytes [13]. Based on the cell surface marker expres-
ion, cytokine secretion and physiological function T cells can belassified into three major groups [14]. Naïve T cells that have notncountered previously an antigen bear CCR7+CD45RA+ functionalarker expression. Central memory T cells (TCM, CCR7+CD45RA−),hich cells mediate reactive memory, are probably arrested at
ntermediate stages of terminal differentiation to become effectoremory cells (TEM). These cells have little or no effector function,
ut readily proliferate and differentiate to effector cells in responseo antigenic stimulation. Protective memory is governed by effector
emory TEM cells (CCR7−CD45RA+/−). TEM cells display character-stic sets of chemokine receptors and adhesion molecules that areequired for homing to inflamed tissues where they exert imme-iate effector function. In several autoimmune diseases, includingultiple sclerosis (MS) [13], rheumatoid arthritis and type-I dia-
etes mellitus [15], chronically activated TEM cells are responsibleor tissue damage.
Several studies confirmed that these T cell subsets react totimulation with a characteristic change in the expression of K+
hannels [13,16]. Naïve, TCM and TEM cells of either CD4+ orD8+ phenotype express similar number (200–300) of Kv1.3 and
ewer than 30 IKCa1 channels per cell. Upon specific stimula-ion of naïve and TCM cells the number of Kv1.3 channels per cellncreases modestly, ∼1.5-fold, whereas they dramatically upreg-late KCa3.1 channels (500 channel/cell) and thus, acquire anv1.3lowKCa3.1high phenotype. On the contrary, activation of TEM
s accompanied by an increase in the number of Kv1.3 chan-els to ∼1500 per cell without any change in the KCa3.1 levels,hereby the channel phenotype of the activated TEM becomesv1.3highKCa3.1low. Consistent with the ion channel phenotype
he proliferation of Kv1.3highKCa3.1low TEM cells becomes Kv1.3-ependent, i.e. proliferation of these cells can be completely andersistently inhibited by selective Kv1.3 inhibitors (e.g. ShK, see
ater). Although the proliferation of naïve and TCM cells is ini-ially sensitive to Kv1.3 blockers repeated stimulation of the cellselieves the Kv1.3 block-mediated inhibition of proliferation as
consequence of KCa3.1 upregulation [17]. Thus, proliferationf re-activated naïve and TCM becomes sensitive to blockers ofhe KCa3.1 channel. Based on these observations targeting ofv1.3 with high affinity and selectivity inhibitors may result inpecific immunosuppression manifesting in the inhibition of TEM-ediated autoimmune reactions but leaving protective immunity
nharmed.
The presence of Kv1.3highKCa3.1low TEM has been confirmed foreveral human autoimmune diseases, such as multiple sclerosisMS), type 1 diabetes mellitus (T1DM) and rheuamtoid arthritisRA). Human T cell clones generated by repeated antigenic chal-enge of peripheral blood lymphocytes of MS patients become
tters 130 (2010) 19–25
Kv1.3highKCa3.11low TEM only if they were repeatedly stimulatedwith the proper autoantigen (myelin basic protein, MBP) but not ifthe antigenic challenge was with irrelevant antigens such as insulinor ovalbumin [13]. A latter study identified Kv1.3high TEM cells inpost-mortem brain slices of MS patients [18]. These cells werefound in the parenchimal infiltrate of MS plaques. Autoantigen-specific T cells (GAD65 and insulin) were recently isolated from theperipheral blood of T1DM patients either by MHC class II tetramersorting or by the generation of short-term antigen-specific T cellclones [15]. The Kv1.3 expression of these autoantigen-specific TEMcells was significantly higher than that of T cells isolated from T1DMpatients but specific for MBP. One patient in that study sufferedfrom both T1DM and MS and the T cell clones specific for GAD65,insulin and MBP all had Kv1.3high channel phenotype correspond-ing to the presence of TEMs specific for both of these antigens. Incase of RA T cells isolated form the synovial fluid are predominantlyCCR7− TEM cells having Kv1.3high channel phenotype [15]. The accu-mulation of these cells in the autoimmune-based arthritis (RA) wasspecific, the synovial fluid form non-autoimmune arthritis patients(osteoarthritis) contained naïve and TCM cells with Kv1.3low phe-notype. These results clearly indicate a correlation between theautoantigen-specific TEM phenotype and the Kv1.3 expression ofthe cells.
Given that autoantigen-specific TEM cells are Kv1.3high andthat their proliferation is persistently inhibited by selective Kv1.3inhibitors several excellent in vivo animal experiments revealedthe applicability of Kv1.3 inhibitors in the prevention or treatmentof various experimental autoimmune reactions mimicking humandiseases. A common characteristic of these animal experiments isthat they were done in rats where the ion channel expression ofnaïve, TCM and TEM cells is similar to that of the human counter-parts [13,16]. Murine T cells, on the contrary, express several typesof voltage-gated K+ channels in addition to Kv1.3. This, combinedwith the high KCa3.1 channel expression makes murine models ofautoimmune diseases unsuitable for evaluating a Kv1.3 inhibitor-based pharmacological approach.
In line with the above adoptive transfer of experimental autoim-mune encephalomyelitis (AT-EAE), an accepted animal model formultiple sclerosis in Lewis rats, could be prevented or the symp-toms of the disease could be greatly ameliorated using selectiveKv1.3 blockers alone or in combination with a KCa3.1 inhibitor,respectively [16]. Similarly, the symptoms of EAE in Lewis ratswere significantly reduced by inhibiting Kv1.3 function, which wasattributed to the great reduction in the proliferative response ofMBP specific T cells along with a decrease in the production of IL-2,TNF and in Ca2+ influx [19]. The use of high affinity and specificityKv1.3 inhibitors in in vivo treatment and prevention experimentsturned out to be very safe, the amount of the toxin (e.g. ShK(L5), seebelow) needed for therapeutic use does not cause cardiac toxicityassessed by continuous EKG monitoring and does not alter clinicalchemistry and hematological parameters after a 2-week therapy[20]. Several parameters characterizing the pharmacokinetics ofKv1.3 inhibitors were determined and improved for increased sta-bility (e.g. ShK192, see below) [21].
Kv1.3 inhibitors also ameliorate disease in rat models of RAand T1DM. In the pristane-induced MHC class II-restricted chronicarthritis model (model of RA) Kv1.3 inhibitor-treated (ShK(L5)-amide treatment) rats had significantly fewer affected joints duringthe 21 days of the treatment than the control group of animals[15]. A prevention trial in MHC class II-restricted DP-BB/W rats,a standard model for T1DM, showed that the development ofexperimental autoimmune diabetes (EAD) in the Kv1.3 specific
inhibitor-treated (PAP-1 treatment, see below) group was reducedto ∼50% of the control group. The prevention of EAD in PAP-1-treated rats is likely to be the result of specific immunosuppressionmanifesting in a decreased intraislet T cell and macrophage infiltra-
ogy Le
tc
tiia�ati
ttscitttiC
aiis
3
iaaiidntbhm
lbalatavito
wmattpstft
Z. Varga et al. / Immunol
ion and reduced � cell destruction compared with vehicle-treatedontrols [15].
In a rat model of allergic contact dermatitis (ACD) the infil-rating cells are CD8+ Kv1.3high TEM cells [22]. A PAP-1-meditatednhibition of the Kv1.3 activity potently suppressed oxazolone-nduced inflammation by inhibiting the infiltration of CD8+ T cellsnd reducing the production of the inflammatory cytokines IFN-, IL-2, and IL-17. Interestingly, topical application of PAP-1 wass effective as intraperitoneal or oral administration, which openshe possibility of developing a drug for the topical treatment ofnflammatory skin diseases such as psoriasis.
The application of cyclosporine (CsA) in the long-term preven-ion of kidney transplant rejection is limited by its severe kidneyoxicity which emphasizes the need for novel immunosuppressivetrategies. A recent study addressed this issue by substituting aombination of Kv1.3 and KCa3.1 inhibitors for CsA following thenitial period of CsA application for 7 days [23]. Allografts wereransplanted from Fisher to Lewis rats and hystopathology showedhat the K+ channel inhibitor-based treatment is equally potent tohe CsA-treatment in the reduction of interstitial mononuclear cellnfiltration, the number of CD43+ T cells, CD8+ cytotoxic T cells andD68+ macrophages in the allografts.
Parallel to the discovery of the therapeutic applicability innimal experiments the pharmacological properties of Kv1.3nhibitors were also dramatically improved resulting in high affin-ty, high specificity and stable Kv1.3 inhibitors. These efforts areummarized in the next section.
. High affinity and specificity blockers of Kv1.3
The discovery that lymphocyte activation in vitro can be inhib-ted by agents that block their potassium channels is nearly as olds the discovery of the channels themselves [1,2]. The first in vivopplication of a high affinity Kv1.3 blocker with the purpose ofmmunosuppression came more than a decade later [24]. The find-ng that the activation of TEM cells involved in several autoimmuneiseases relies heavily on Kv1.3 channels while that of TCM andaïve cells does not, gave new momentum to the search for selec-ive Kv1.3 blockers [13]. During the last decade several compoundslocking Kv1.3 channels were shown to prevent the delayed typeypersensitivity reaction and improve the symptoms of variousodels of autoimmune diseases [15,16,19,25].The molecules shown to block Kv1.3 channels belong in two
arge classes: peptide toxins and small molecule blockers. Kv1.3-locking peptide toxins from scorpions, sea anemones or othernimals typically consist of 30–40 residues and have a molecu-ar weight of about 4 kDa. They carry a large positive net chargend consequently are impermeant through cell membranes. Pep-ide toxins utilize a “cork in the bottle” blocking strategy, coveringnd plugging the external end of the channel pore, thereby pre-enting the efflux of K+ ions. Their binding relies on a fairly largenteracting surface with the channel, where several correctly posi-ioned residue pairs contribute to the high affinity and selectivityf the binding [26].
In contrast, small molecule blockers typically have a moleculareight of less than 800 Da, are hydrophobic and thus easily per-eate through cell membranes. In the water-filled cavity located
t the middle of the channel they bind from the intracellular sideo the internal end of the selectivity filter. Due to their smaller sizehese molecules have fewer contact points with the channel than
eptide toxins, which generally results in lower affinity and poorerelectivity [27]. Furthermore, as opposed to the external mouth ofhe pore, which is practically isolated from gating-associated con-ormational changes, the internal end of the selectivity filter andhe cavity undergo significant structural rearrangements when thetters 130 (2010) 19–25 21
channel shifts between gating states. As a result, external block bypeptide toxins is state-independent, while internal block by smallmolecules is state-dependent, usually preferring the inactivatedstate of Kv1.3 channels [28]. At the resting membrane potentialof T cells the majority, but not all, of Kv1.3 channels are in the inac-tivated state. A notable exception to the typical small moleculebehavior is the traditional K+ channel blocker tetraethylammo-nium, which is capable of blocking the channel from both sides,but is membrane-impermeant.
From a pharmaceutical aspect Kv1.3-blocking small moleculesare more desirable than peptides because of easier and less costlyproduction and more easily attainable oral bioavailability. Althoughless stressful delivery systems are being developed, the adminis-tration of peptide drugs usually requires injections and peptidesare also cleared from the body more quickly than small molecules.Moreover, peptides are more likely to trigger an immune reac-tion in the patient and may be less stable chemically [21]. On theother hand, due to their higher affinity and selectivity for Kv1.3,much lower concentrations of peptide toxins may be needed fora therapeutic effect, the possibility of unspecific binding to othermolecular structures (and therefore the possibility of side effects)is minimized, and the interaction with other drugs is also less likely.
When the structure of a natural lead peptide that blocks Kv1.3channels with high affinity is altered with the purpose of develop-ing it into a therapeutically usable drug, achieving high selectivityfor Kv1.3 over other channels in vitro is the primary step. If thetoxin blocks multiple channels then the comparison of sequencesand affinities, structure determination by X-ray crystallography orNMR, thermodynamic mutant cycle analysis and in silico dock-ing simulations can identify the interacting amino acid pairs inthe channel and the toxin that determine the toxin’s selectivityamong the various channels [29–32]. By introducing point muta-tions at these locations toxin selectivity can be shifted towardthe desired target channel [33,34]. However, for the peptide tobe a safe and effective drug all of the aspects mentioned abovemust be considered. The evolution of a native peptide into apotent, highly selective, chemically stable, non-toxic and non-immunogenic molecule is best exemplified by the sea anemonetoxin ShK from Stichodactyla helianthus. The native peptide potentlyblocks Kv1.3 channels along with Kv1.1, Kv1.4, Kv1.6 and Kv3.2channels and in an effort to improve its selectivity the lysineresidue that is highly conserved among K+ channel blocking tox-ins was replaced by the non-natural amino acid diaminopropionicacid [35–37]. Although the mutant toxin (ShK-Dap22) displayedimproved selectivity for Kv1.3, its efficacy to inhibit T cell activa-tion was later shown to be much lower than that of ShK, and italso turned out to block heterotetramers of Kv1.1 and 1.2 subunits[38]. Two other ShK mutants, ShK-F6CA containing a fluoresceinmoiety attached to its N-terminus, and ShK(L5) containing an l-phosphotyrosine at the same position were produced with greatlyimproved properties [20,39]. The latter blocks Kv1.3 channels andsuppresses the proliferation of TEM cells in picomolar concentra-tions and is at least 100-fold less effective on other related channels.This peptide was shown not to be cytotoxic or mutagenic on mam-malian cell lines, yet remained stable in the plasma of rats anddid not influence EKG records, blood chemistry or hematologicalparameters even during prolonged administration. ShK(L5) effec-tively prevented the delayed type hypersensitivity reaction andimproved the symptoms of acute adoptive experimental autoim-mune encephalomyelitis in rats. However, at acidic pH and elevatedtemperatures dephosphorylation of the tyrosine residue and oxi-
dation of a methionine residue was observed indicating instabilityof the molecule [21]. The peptide was also degraded by proteasesruling out the possibility of oral administration in that form. Toovercome stability problems the phosphotyrosine was exchangedfor a non-hydrolysable replacement group, the methionine for nor-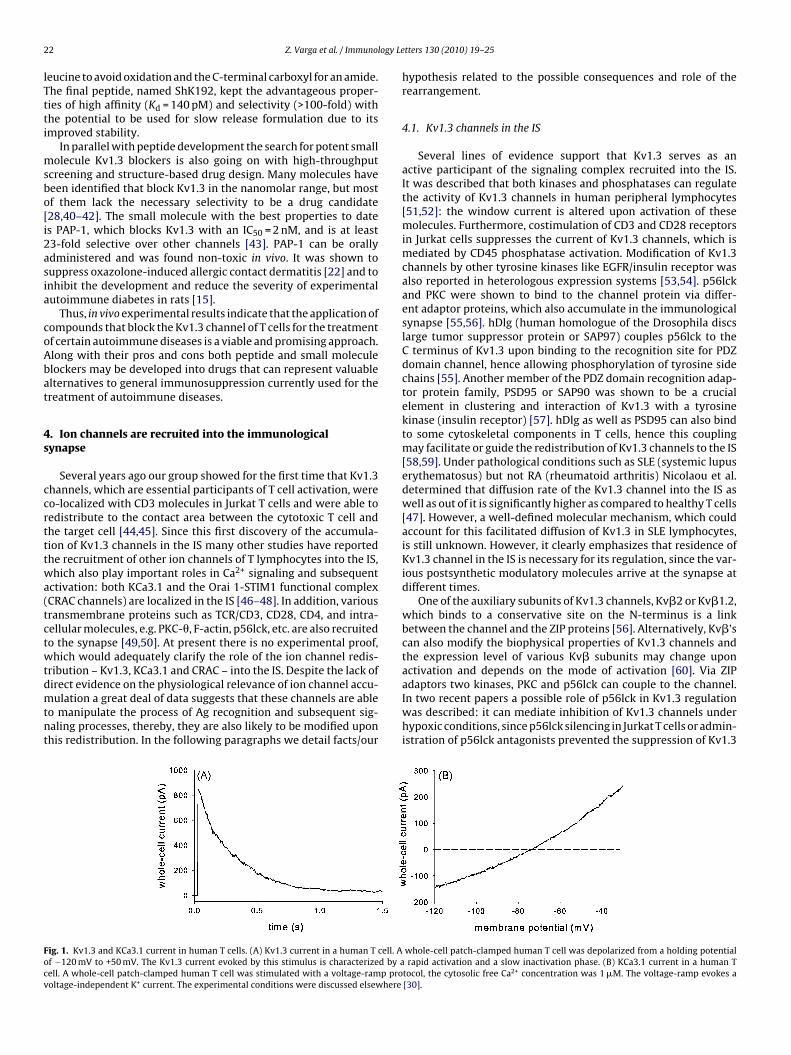
2 logy Le
lTtti
msbo[i2asia
coAbat
4s
ccrtttwa(tctwtdmtnt
Focv
2 Z. Varga et al. / Immuno
eucine to avoid oxidation and the C-terminal carboxyl for an amide.he final peptide, named ShK192, kept the advantageous proper-ies of high affinity (Kd = 140 pM) and selectivity (>100-fold) withhe potential to be used for slow release formulation due to itsmproved stability.
In parallel with peptide development the search for potent smallolecule Kv1.3 blockers is also going on with high-throughput
creening and structure-based drug design. Many molecules haveeen identified that block Kv1.3 in the nanomolar range, but mostf them lack the necessary selectivity to be a drug candidate28,40–42]. The small molecule with the best properties to dates PAP-1, which blocks Kv1.3 with an IC50 = 2 nM, and is at least3-fold selective over other channels [43]. PAP-1 can be orallydministered and was found non-toxic in vivo. It was shown touppress oxazolone-induced allergic contact dermatitis [22] and tonhibit the development and reduce the severity of experimentalutoimmune diabetes in rats [15].
Thus, in vivo experimental results indicate that the application ofompounds that block the Kv1.3 channel of T cells for the treatmentf certain autoimmune diseases is a viable and promising approach.long with their pros and cons both peptide and small moleculelockers may be developed into drugs that can represent valuablelternatives to general immunosuppression currently used for thereatment of autoimmune diseases.
. Ion channels are recruited into the immunologicalynapse
Several years ago our group showed for the first time that Kv1.3hannels, which are essential participants of T cell activation, wereo-localized with CD3 molecules in Jurkat T cells and were able toedistribute to the contact area between the cytotoxic T cell andhe target cell [44,45]. Since this first discovery of the accumula-ion of Kv1.3 channels in the IS many other studies have reportedhe recruitment of other ion channels of T lymphocytes into the IS,hich also play important roles in Ca2+ signaling and subsequent
ctivation: both KCa3.1 and the Orai 1-STIM1 functional complexCRAC channels) are localized in the IS [46–48]. In addition, variousransmembrane proteins such as TCR/CD3, CD28, CD4, and intra-ellular molecules, e.g. PKC-�, F-actin, p56lck, etc. are also recruitedo the synapse [49,50]. At present there is no experimental proof,hich would adequately clarify the role of the ion channel redis-
ribution – Kv1.3, KCa3.1 and CRAC – into the IS. Despite the lack of
irect evidence on the physiological relevance of ion channel accu-ulation a great deal of data suggests that these channels are ableo manipulate the process of Ag recognition and subsequent sig-aling processes, thereby, they are also likely to be modified uponhis redistribution. In the following paragraphs we detail facts/our
ig. 1. Kv1.3 and KCa3.1 current in human T cells. (A) Kv1.3 current in a human T cell. Af −120 mV to +50 mV. The Kv1.3 current evoked by this stimulus is characterized by aell. A whole-cell patch-clamped human T cell was stimulated with a voltage-ramp prooltage-independent K+ current. The experimental conditions were discussed elsewhere
tters 130 (2010) 19–25
hypothesis related to the possible consequences and role of therearrangement.
4.1. Kv1.3 channels in the IS
Several lines of evidence support that Kv1.3 serves as anactive participant of the signaling complex recruited into the IS.It was described that both kinases and phosphatases can regulatethe activity of Kv1.3 channels in human peripheral lymphocytes[51,52]: the window current is altered upon activation of thesemolecules. Furthermore, costimulation of CD3 and CD28 receptorsin Jurkat cells suppresses the current of Kv1.3 channels, which ismediated by CD45 phosphatase activation. Modification of Kv1.3channels by other tyrosine kinases like EGFR/insulin receptor wasalso reported in heterologous expression systems [53,54]. p56lckand PKC were shown to bind to the channel protein via differ-ent adaptor proteins, which also accumulate in the immunologicalsynapse [55,56]. hDlg (human homologue of the Drosophila discslarge tumor suppressor protein or SAP97) couples p56lck to theC terminus of Kv1.3 upon binding to the recognition site for PDZdomain channel, hence allowing phosphorylation of tyrosine sidechains [55]. Another member of the PDZ domain recognition adap-tor protein family, PSD95 or SAP90 was shown to be a crucialelement in clustering and interaction of Kv1.3 with a tyrosinekinase (insulin receptor) [57]. hDlg as well as PSD95 can also bindto some cytoskeletal components in T cells, hence this couplingmay facilitate or guide the redistribution of Kv1.3 channels to the IS[58,59]. Under pathological conditions such as SLE (systemic lupuserythematosus) but not RA (rheumatoid arthritis) Nicolaou et al.determined that diffusion rate of the Kv1.3 channel into the IS aswell as out of it is significantly higher as compared to healthy T cells[47]. However, a well-defined molecular mechanism, which couldaccount for this facilitated diffusion of Kv1.3 in SLE lymphocytes,is still unknown. However, it clearly emphasizes that residence ofKv1.3 channel in the IS is necessary for its regulation, since the var-ious postsynthetic modulatory molecules arrive at the synapse atdifferent times.
One of the auxiliary subunits of Kv1.3 channels, Kv�2 or Kv�1.2,which binds to a conservative site on the N-terminus is a linkbetween the channel and the ZIP proteins [56]. Alternatively, Kv�’scan also modify the biophysical properties of Kv1.3 channels andthe expression level of various Kv� subunits may change uponactivation and depends on the mode of activation [60]. Via ZIP
adaptors two kinases, PKC and p56lck can couple to the channel.In two recent papers a possible role of p56lck in Kv1.3 regulationwas described: it can mediate inhibition of Kv1.3 channels underhypoxic conditions, since p56lck silencing in Jurkat T cells or admin-istration of p56lck antagonists prevented the suppression of Kv1.3whole-cell patch-clamped human T cell was depolarized from a holding potentialrapid activation and a slow inactivation phase. (B) KCa3.1 current in a human T
tocol, the cytosolic free Ca2+ concentration was 1 �M. The voltage-ramp evokes a[30].

Z. Varga et al. / Immunology Le
Fig. 2. Ion channels and Kv1.3-associated proteins in the immunological synapse(IS). Kv1.3, KCa3.1 Orai 1 and STIM1 is depicted schematically in the IS between anantigen presenting cell (APC) and a T cell. hDlg binds to the PDZ domain in the CtKs
cwitolmpvofo
4
CCsslivicfttctfit
atidqttb
[
[
erminus of Kv1.3, which is coupled to the p56lck. Kv�2 links the N terminus ofv1.3 channel to the ZIP1/2 protein, which can have several interacting partnersuch as p56lck, PKC, PPAR�, ras-GAP, p38MAPK, TRAF6.
urrent detected in hypoxia [61,62]. Moreover, ZIP/p62 can interactith several proteins, which induce gene transcription through var-
ous signaling pathways (NF-�B, PPAR�, p38MAPK); and are likelyo lead to T cell activation (see Fig. 2) [63–67]. The accumulationf Kv1.3 in the IS makes it possible to interact with the all proteinsinked to it. Recently, Toth et al. has demonstrated that rearrange-
ent of Kv1.3 channels into the IS modifies kinetic and equilibriumarameters of the activation and inactivation [68]. The slower acti-ation of Kv1.3 current may be an evidence for the redistributionf the channel into membrane domains with higher viscosity, theaster inactivation kinetics can be explained by dephosphorylationf the channel [69].
.2. Role of Ca2+ influx in the IS
Opening of CRAC channels, which is triggered by emptying of ERa2+ stores and formed upon STIM1-Orai 1 coupling, elevates thea2+ level in T cells. The Ca2+ concentration in the vicinity of theite of entry may go up to a concentration range, which in turn canwitch on KCa3.1 in a direct manner or Kv1.3 due to the local depo-arization. Moreover, the elements of the supramolecular complexn the IS can be Ca2+-dependent enzymes, which may become acti-ated, too. Due to the increased intracellular Ca2+ CRAC channelsnactivate, while the translocation of mitochondria to the synapseounteract this negative feedback and maintain the driving forceor the Ca2+ ions [70–72]. Interestingly, it has been reported latelyhat immobilization of Kv1.3 channels by crosslinking (accumula-ion in the IS is blocked) elevates Ca2+ response of Jurkat T cellshallenged by anti-CD3/anti-CD28 coated beads [73]. Nonetheless,he elevated intracellular Ca2+ level stabilizes and maintains therm contact between the T cell and the APC, but rearrangement ofhe CD3 molecules into the IS is not affected [74].
Finally we should mention that in their recent paper Cahalannd Chandy suggested that changes of the ion concentrations inhe synaptic cleft (mainly the elimination of Ca2+ ions and increasen the K+ level) between the T cell and the APC may result in theepolarization of the T cell membrane [5]. The putative conse-
uences of this are the following: (1) attenuated Ca2+ flux intohe T lymphocytes, (2) increased negative surface charge due tohe polar head of lipid molecules, which can shift the transmem-rane potential toward depolarization, (3) stronger interaction[
[
tters 130 (2010) 19–25 23
between the interacting membrane proteins of the APC and lym-phocyte, (4) activation of voltage activated Ca2+ channels of the Tcell, and (5) changes in the biophysical properties of Kv1.3 chan-nels. These scenarious are certainly viable, but presently are stillassumptions with no solid experimental support. In summary,ion channels are important elements in efficient transmembranesignaling upon antigen recognition. Besides being responsible formaintaining the appropriate ion milieu during T cell activation, theyare also remarkable players in the crowd of molecules in signal-ing clusters. Thus, their role may be twofold: they may accomplishrequired steps in the signaling cascade (Ca2+-dependent enzymeactivation) or they may serve as a collectors of various interactingmolecular partners (as for Kv1.3).
5. Concluding remarks
Compounds influencing ion channel activity in nerve or musclecells to achieve certain therapeutic goals have already establishedthemselves as powerful pharmaceutical tools and the potential forsimilar successful immunological application was recognized about25 years ago. We are not quite yet at the end of this road, butthe wealth of information collected during this period about theintriguing and diverse function of T cell ion channels in one of thecritical steps of cellular immunity, along with major developmentsin specific channel blockers, took us a long way down on it. The suc-cess of in vivo experiments for the prevention of undesired actionsof the immune system via T cell ion channels predicts the avail-ability of a potent alternative to conventional immunosuppression,hopefully, in the near future.
Acknowledgements
Supported in part by grants: OTKA K 60740 and NK 61412,and TÁMOP-4.2.2/08/1 19 to G.P. Zoltan Varga and Peter Hajdu areBolyai fellows.
References
[1] Decoursey TE, Chandy KG, Gupta S, Cahalan MD. Voltage-gated K+ channels inhuman T lymphocytes: a role in mitogenesis? Nature 1984;307:465–8.
[2] Matteson DR, Deutsch C. K channels in T lymphocytes: a patch clamp studyusing monoclonal antibody adhesion. Nature 1984;307:468–71.
[3] Grissmer S, Nguyen AN, Cahalan MD. Calcium-activated potassium channels inresting and activated human T lymphocytes. Expression levels, calcium depen-dence, ion selectivity, and pharmacology. J Gen Physiol 1993;102:601–30.
[4] Fanger CM, Ghanshani S, Logsdon NJ, Rauer H, Kalman K, Zhou J, et al. Calmod-ulin mediates calcium-dependent activation of the intermediate conductanceKCa channel, IKCa1. J Biol Chem 1999;274:5746–54.
[5] Cahalan MD, Chandy KG. The functional network of ion channels in T lympho-cytes. Immunol Rev 2009;231:59–87.
[6] Zweifach A, Lewis RS. Mitogen-regulated Ca2+ current of T lymphocytes isactivated by depletion of intracellular Ca2+ stores. Proc Natl Acad Sci USA1993;90:6295–9.
[7] Partiseti M, Le Deist F, Hivroz C, Fischer A, Korn H, Choquet D. The calciumcurrent activated by T cell receptor and store depletion in human lymphocytesis absent in a primary immunodeficiency. J Biol Chem 1994;269:32327–35.
[8] Zhang SL, Yu Y, Roos J, Kozak JA, Deerinck TJ, Ellisman MH, et al. STIM1 is a Ca2+
sensor that activates CRAC channels and migrates from the Ca2+ store to theplasma membrane. Nature 2005;437:902–5.
[9] Liou J, Kim ML, Heo WD, Jones JT, Myers JW, Ferrell Jr JE, et al. STIM is aCa2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr Biol2005;15:1235–41.
10] Feske S, Gwack Y, Prakriya M, Srikanth S, Puppel SH, Tanasa B, et al. A muta-tion in Orai1 causes immune deficiency by abrogating CRAC channel function.Nature 2006;441:179–85.
11] Vig M, Peinelt C, Beck A, Koomoa DL, Rabah D, Koblan-Huberson M, et al.CRACM1 is a plasma membrane protein essential for store-operated Ca2+ entry.Science 2006;312:1220–3.
12] Penna A, Demuro A, Yeromin AV, Zhang SL, Safrina O, Parker I, et al. The CRACchannel consists of a tetramer formed by Stim-induced dimerization of Oraidimers. Nature 2008;456:116–20.
13] Wulff H, Calabresi PA, Allie R, Yun S, Pennington M, Beeton C, et al. The voltage-gated Kv1.3 K+ channel in effector memory T cells as new target for MS. J ClinInvest 2003;111:1703–13.

2 logy Le
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
4 Z. Varga et al. / Immuno
14] Sallusto F, Geginat J, Lanzavecchia A. Central memory and effector memoryT cell subsets: function, generation, and maintenance. Annu Rev Immunol2004;22:745–63.
15] Beeton C, Wulff H, Standifer NE, Azam P, Mullen KM, Pennington MW, et al.Kv1.3 channels are a therapeutic target for T cell-mediated autoimmune dis-eases. Proc Natl Acad Sci USA 2006;103:17414–9.
16] Beeton C, Wulff H, Barbaria J, Clot-Faybesse O, Pennington M, Bernard D, etal. Selective blockade of T lymphocyte K+ channels ameliorates experimentalautoimmune encephalomyelitis, a model for multiple sclerosis. Proc Natl AcadSci USA 2001;98:13942–7.
17] Ghanshani S, Wulff H, Miller MJ, Rohm H, Neben A, Gutman GA, et al.Up-regulation of the IKCa1 potassium channel during T-cell activation. Molec-ular mechanism and functional consequences. J Biol Chem 2000;275:37137–49.
18] Rus H, Pardo CA, Hu L, Darrah E, Cudrici C, Niculescu T, et al. The voltage-gatedpotassium channel Kv1.3 is highly expressed on inflammatory infiltrates inmultiple sclerosis brain. Proc Natl Acad Sci USA 2005;102:11094–9.
19] Beeton C, Barbaria J, Giraud P, Devaux J, Benoliel AM, Gola M, et al. Selectiveblocking of voltage-gated K+ channels improves experimental autoimmuneencephalomyelitis and inhibits T cell activation. J Immunol 2001;166:936–44.
20] Beeton C, Pennington MW, Wulff H, Singh S, Nugent D, Crossley G, et al. Target-ing effector memory T cells with a selective peptide inhibitor of Kv1.3 channelsfor therapy of autoimmune diseases. Mol Pharmacol 2005;67:1369–81.
21] Pennington MW, Beeton C, Galea CA, Smith BJ, Chi V, Monaghan KP, et al.Engineering a stable and selective peptide blocker of the Kv1.3 channel in Tlymphocytes. Mol Pharmacol 2009;75:762–73.
22] Azam P, Sankaranarayanan A, Homerick D, Griffey S, Wulff H. Targeting effectormemory T cells with the small molecule Kv1.3 blocker PAP-1 suppresses allergiccontact dermatitis. J Invest Dermatol 2007;127:1419–29.
23] Grgic I, Wulff H, Eichler I, Flothmann C, Kohler R, Hoyer J. Blockade of T-lymphocyte KCa3.1 and Kv1.3 channels as novel immunosuppression strategyto prevent kidney allograft rejection. Transplant Proc 2009;41:2601–6.
24] Koo GC, Blake JT, Talento A, Nguyen M, Lin S, Sirotina A, et al. Blockade of thevoltage-gated potassium channel Kv1.3 inhibits immune responses in vivo. JImmunol 1997;158:5120–8.
25] Valverde P, Kawai T, Taubman MA. Selective blockade of voltage-gatedpotassium channels reduces inflammatory bone resorption in experimentalperiodontal disease. J Bone Miner Res 2004;19:155–64.
26] Panyi G, Possani LD, Rodriguez de la Vega RC, Gaspar R, Varga Z. K+ channelblockers: novel tools to inhibit T cell activation leading to specific immuno-suppression. Curr Pharm Des 2006;12:2199–220.
27] Chandy KG, Wulff H, Beeton C, Pennington M, Gutman GA, Cahalan MD. K+
channels as targets for specific immunomodulation. Trends Pharmacol Sci2004;25:280–9.
28] Vennekamp J, Wulff H, Beeton C, Calabresi PA, Grissmer S, Hansel W, et al.Kv1.3-blocking 5-phenylalkoxypsoralens: a new class of immunomodulators.Mol Pharmacol 2004;65:1364–74.
29] Goldstein SA, Pheasant DJ, Miller C. The charybdotoxin receptor of a ShakerK+ channel: peptide and channel residues mediating molecular recognition.Neuron 1994;12:1377–88.
30] Corzo G, Papp F, Varga Z, Barraza O, Espino-Solis PG, Rodriguez de la VegaRC, et al. A selective blocker of Kv1.2 and Kv1.3 potassium channels from thevenom of the scorpion Centruroides suffusus suffusus. Biochem Pharmacol2008;76:1142–54.
31] Aiyar J, Withka JM, Rizzi JP, Singleton DH, Andrews GC, Lin W, et al. Topologyof the pore-region of a K+ channel revealed by the NMR-derived structures ofscorpion toxins. Neuron 1995;15:1169–81.
32] Doyle DA, Morais CJ, Pfuetzner RA, Kuo A, Gulbis JM, Cohen SL, et al. The struc-ture of the potassium channel: molecular basis of K+ conduction and selectivity.Science 1998;280:69–77.
33] Rauer H, Lanigan MD, Pennington MW, Aiyar J, Ghanshani S, Cahalan MD, etal. Structure-guided transformation of charybdotoxin yields an analog thatselectively targets Ca2+-activated over voltage-gated K+ channels. J Biol Chem2000;275:1201–8.
34] Mouhat S, Visan V, Ananthakrishnan S, Wulff H, Andreotti N, Grissmer S, etal. K+ channel types targeted by synthetic OSK1, a toxin from Orthochirusscrobiculosus scorpion venom. Biochem J 2005;385:95–104.
35] Castaneda O, Sotolongo V, Amor AM, Stocklin R, Anderson AJ, Harvey AL, et al.Characterization of a potassium channel toxin from the Caribbean Sea anemoneStichodactyla helianthus. Toxicon 1995;33:603–13.
36] Kalman K, Pennington MW, Lanigan MD, Nguyen A, Rauer H, Mahnir V, etal. ShK-Dap22, a potent Kv1.3-specific immunosuppressive polypeptide. J BiolChem 1998;273:32697–707.
37] Yan L, Herrington J, Goldberg E, Dulski PM, Bugianesi RM, Slaughter RS, etal. Stichodactyla helianthus peptide, a pharmacological tool for studying Kv3.2channels. Mol Pharmacol 2005;67:1513–21.
38] Middleton RE, Sanchez M, Linde AR, Bugianesi RM, Dai G, Felix JP, et al. Substitu-tion of a single residue in Stichodactyla helianthus peptide, ShK-Dap22, revealsa novel pharmacological profile. Biochemistry 2003;42:13698–707.
39] Beeton C, Wulff H, Singh S, Botsko S, Crossley G, Gutman GA, et al. A novelfluorescent toxin to detect and investigate Kv1.3 channel up-regulation inchronically activated T lymphocytes. J Biol Chem 2003;278:9928–37.
40] Nguyen AN, Kath JC, Hanson DC, Biggers MS, Cannif PC, Donovan CB, et al. Novelnonpeptide agents potently block the C-type inactivated conformation of Kv1.3and suppress T cell activation. Mol Pharmacol 1996;50:1672–9.
[
[
tters 130 (2010) 19–25
41] Koo GC, Blake JT, Shah K, Staruch MJ, Dumont F, Wunderler D, et al. Correolideand derivatives are novel immunosuppressants blocking the lymphocyte Kv1.3potassium channels. Cell Immunol 1999;197:99–107.
42] Hanson DC, Nguyen A, Mather RJ, Rauer H, Koch K, Burgess LE, et al. UK-78,282,a novel piperidine compound that potently blocks the Kv1.3 voltage-gatedpotassium channel and inhibits human T cell activation. Br J Pharmacol1999;126:1707–16.
43] Schmitz A, Sankaranarayanan A, Azam P, Schmidt-Lassen K, Homerick D, HanselW, et al. Design of PAP-1, a selective small molecule Kv1.3 blocker, for thesuppression of effector memory T cells in autoimmune diseases. Mol Pharmacol2005;68:1254–70.
44] Panyi G, Vamosi G, Bacso Z, Bagdany M, Bodnar A, Varga Z, et al. Kv1.3 potassiumchannels are localized in the immunological synapse formed between cytotoxicand target cells. Proc Natl Acad Sci USA 2004;101:1285–90.
45] Panyi G, Bagdany M, Bodnar A, Vamosi G, Szentesi G, Jenei A, et al. Colocalizationand nonrandom distribution of Kv1.3 potassium channels and CD3 moleculesin the plasma membrane of human T lymphocytes. Proc Natl Acad Sci USA2003;100:2592–7.
46] Lioudyno MI, Kozak JA, Penna A, Safrina O, Zhang SL, Sen D, et al. Orai1 andSTIM1 move to the immunological synapse and are up-regulated during T cellactivation. Proc Natl Acad Sci USA 2008;105:2011–6.
47] Nicolaou SA, Szigligeti P, Neumeier L, Lee SM, Duncan HJ, Kant SK, et al.Altered dynamics of Kv1.3 channel compartmentalization in the immunologi-cal synapse in systemic lupus erythematosus. J Immunol 2007;179:346–56.
48] Nicolaou SA, Neumeier L, Peng Y, Devor DC, Conforti L. The Ca2+-activated K+
channel KCa3.1 compartmentalizes in the immunological synapse of human Tlymphocytes. Am J Physiol Cell Physiol 2007;292:C1431–9.
49] van der Merwe PA. Formation and function of the immunological synapse. CurrOpin Immunol 2002;14:293–8.
50] Bromley SK, Burack WR, Johnson KG, Somersalo K, Sims TN, Sumen C, et al. Theimmunological synapse. Annu Rev Immunol 2001;19:375–96.
51] Chung I, Schlichter LC. Native Kv1.3 channels are upregulated by protein kinaseC. J Membr Biol 1997;156:73–85.
52] Chung I, Schlichter LC. Regulation of native Kv1.3 channels by cAMP-dependentprotein phosphorylation. Am J Physiol 1997;273:C622–33.
53] Holmes TC, Fadool DA, Levitan IB. Tyrosine phosphorylation of the Kv1.3 potas-sium channel. J Neurosci 1996;16:1581–90.
54] Bowlby MR, Fadool DA, Holmes TC, Levitan IB. Modulation of the Kv1.3 potas-sium channel by receptor tyrosine kinases. J Gen Physiol 1997;110:601–10.
55] Hanada T, Lin L, Chandy KG, Oh SS, Chishti AH. Human homologue of theDrosophila discs large tumor suppressor binds to p56lck tyrosine kinaseand Shaker type Kv1.3 potassium channel in T lymphocytes. J Biol Chem1997;272:26899–904.
56] Gong J, Xu J, Bezanilla M, van Huizen R, Derin R, Li M. Differential stimula-tion of PKC phosphorylation of potassium channels by ZIP1 and ZIP2. Science1999;285:1565–9.
57] Marks DR, Fadool DA. Post-synaptic density perturbs insulin-induced Kv1.3channel modulation via a clustering mechanism involving the SH3 domain. JNeurochem 2007;103:1608–27.
58] Xavier R, Rabizadeh S, Ishiguro K, Andre N, Ortiz JB, Wachtel H, et al. Discs large(Dlg1) complexes in lymphocyte activation. J Cell Biol 2004;166:173–8.
59] Round JL, Tomassian T, Zhang M, Patel V, Schoenberger SP, Miceli MC. Dlgh1coordinates actin polymerization, synaptic T cell receptor and lipid raft aggre-gation, and effector function in T cells. J Exp Med 2005;201:419–30.
60] Vicente R, Escalada A, Soler C, Grande M, Celada A, Tamkun MM, et al. Patternof Kv� subunit expression in macrophages depends upon proliferation and themode of activation. J Immunol 2005;174:4736–44.
61] Robbins JR, Lee SM, Filipovich AH, Szigligeti P, Neumeier L, Petrovic M, etal. Hypoxia modulates early events in T cell receptor-mediated activation inhuman T lymphocytes via Kv1.3 channels. J Physiol 2005;564:131–43.
62] Szigligeti P, Neumeier L, Duke E, Chougnet C, Takimoto K, Lee SM, et al.Signalling during hypoxia in human T lymphocytes—critical role of the src pro-tein tyrosine kinase p56Lck in the O2 sensitivity of Kv1.3 channels. J Physiol2006;573:357–70.
63] Joung I, Kim T, Stolz LA, Payne G, Winkler DG, Walsh CT, et al. Modificationof Ser59 in the unique N-terminal region of tyrosine kinase p56lck regulatesspecificity of its Src homology 2 domain. Proc Natl Acad Sci USA 1995;92:5778–82.
64] Park I, Chung J, Walsh CT, Yun Y, Strominger JL, Shin J. Phosphotyrosine-independent binding of a 62-kDa protein to the src homology 2 (SH2) domainof p56lck and its regulation by phosphorylation of Ser-59 in the lck uniqueN-terminal region. Proc Natl Acad Sci USA 1995;92:12338–42.
65] Vadlamudi RK, Joung I, Strominger JL, Shin J. p62, a phosphotyrosine-independent ligand of the SH2 domain of p56lck, belongs to a new class ofubiquitin-binding proteins. J Biol Chem 1996;271:20235–7.
66] Diradourian C, Le May C, Cauzac M, Girard J, Burnol AF, Pegorier JP. Involvementof ZIP/p62 in the regulation of PPAR� transcriptional activity by p38-MAPK.Biochim Biophys Acta 2008;1781:239–44.
67] Kim JY, Ozato K. The sequestosome 1/p62 attenuates cytokine gene expres-sion in activated macrophages by inhibiting IFN regulatory factor 8 and TNF
receptor-associated factor 6/NF-�B activity. J Immunol 2009;182:2131–40.68] Toth A, Szilagyi O, Krasznai Z, Panyi G, Hajdu P. Functional consequences ofKv1.3 ion channel rearrangement into the immunological synapse. ImmunolLett 2009;125:15–21.
69] Hajdu P, Varga Z, Pieri C, Panyi G, Gaspar Jr R. Cholesterol modifies the gatingof Kv1.3 in human T lymphocytes. Pflugers Arch 2003;445:674–82.

ogy Le
[
[
[
[73] Nicolaou SA, Neumeier L, Steckly A, Kucher V, Takimoto K, Conforti L.Localization of Kv1.3 channels in the immunological synapse modulates
Z. Varga et al. / Immunol
70] Zweifach A, Lewis RS. Rapid inactivation of depletion-activated calcium currentICRAC due to local calcium feedback. J Gen Physiol 1995;105:209–26.
71] Zweifach A, Lewis RS. Slow calcium-dependent inactivation of depletion-activated calcium current. Store-dependent and -independent mechanisms. JBiol Chem 1995;270:14445–51.
72] Malli R, Naghdi S, Romanin C, Graier WF. Cytosolic Ca2+ prevents the subplas-malemmal clustering of STIM1: an intrinsic mechanism to avoid Ca2+ overload.J Cell Sci 2008;121:3133–9.
[
tters 130 (2010) 19–25 25
the calcium response to antigen stimulation in T lymphocytes. J Immunol2009;183:6296–302.
74] Krummel MF, Sjaastad MD, Wulfing C, Davis MM. Differential clustering of CD4and CD3� during T cell recognition. Science 2000;289:1349–52.