integrated pest management tactics for control of cabbage
TRANSCRIPT

Louisiana State UniversityLSU Digital Commons
LSU Historical Dissertations and Theses Graduate School
1997
Integrated Pest Management Tactics for Control ofCabbage Looper and Diamondback Moth inCabbage.Paul Wesley IveyLouisiana State University and Agricultural & Mechanical College
Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses
This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion inLSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please [email protected].
Recommended CitationIvey, Paul Wesley, "Integrated Pest Management Tactics for Control of Cabbage Looper and Diamondback Moth in Cabbage." (1997).LSU Historical Dissertations and Theses. 6541.https://digitalcommons.lsu.edu/gradschool_disstheses/6541

INFORMATION TO USERS
This manuscript has been reproduced from the microfilm master. LJMI
films the text directly from the original or copy submitted. Thus, some
thesis and dissertation copies are in typewriter face, while others may be
from any type o f computer printer.
The quality o f this reproduction is dependent upon the quality of the
copy submitted. Broken or indistinct print, colored or poor quality
illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction.
In the unlikely event that the author did not send UMI a complete
manuscript and there are missing pages, these will be noted. Also, if
unauthorized copyright material had to be removed, a note will indicate
the deletion.
Oversize materials (e.g., maps, drawings, charts) are reproduced by
sectioning the original, beginning at the upper left-hand comer and
continuing from left to right in equal sections with small overlaps. Each
original is also photographed in one exposure and is included in reduced
form at the back of the book.
Photographs included in the original manuscript have been reproduced
xerographically in this copy. Higher quality 6” x 9” black and white
photographic prints are available for any photographs or illustrations
appearing in this copy for an additional charge. Contact UMI directly to
order.
UMIA Beil &. Howell Infbnnation Company
300 NoithZeeb Road, Ann Arbor MI 48106-1346 USA 313/761-4700 800/321-0600
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

INTEGRATED PEST MANAGEMENT TACTICS FOR CONTROL OF CABBAGE LOOPER AND DIAMONDBACK MOTH IN CABBAGE
A DissertationSubmitted to the Graduate Faculty of the
Louisiana State University and Agricultural and Mechanical College
in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
in
The Department of Entomology
byPaul W. Ivey
A.Sc., College of Agriculture, Jamaica, 1987 B.Sc., University of the West Indies, Trinidad, 1989
M.S., Louisiana State University, 1991 December 1997
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

UMI Number: 9810829
UMI Microform 9810829 Copyright 1997, by UMI Company. All rights reserved.
This microform edition is protected against unauthorized copying under Title 17, United States Code.
UMI300 North Zeeb Road Ann Arbor, MI 48103
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

DedicationThe late Harold Ivey was a quiet man; a good husband; a good father. A man, though he did not accumulate any great wealth, who supported, protected, and cared for his family as best he could, and who placed a premium on education for his children; that is his enduring legacy. I dedicate this dissertation to the memory of Harold Ivey; a quiet man; a good husband; a good father. My father.
ii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

AcknowledgmentsOne of the many lessons that I have learned while
pursuing this degree is that there is no such thing as "a self-made man"; many persons have helped me to successfully complete this degree. My major professor. Dr. Seth James Johnson, has been very helpful. He has patiently, tactfully, and tenaciously guided me in the ways of science: generating ideas and testing them against facts, attention to details, critical thinking, scholarship, clarity of expression, and discipline; and sought to instill in me the higher levels of learning: analysis, evaluation, and synthesis. In addition, he was very instrumental in my getting a two-year tuition waiver award. It is difficult, indeed impossible, to exaggerate the gratitude that I extend to Dr. Johnson for his assistance and guidance in helping me make the best of my limited intellectual endowments.
I thank the members of my advisory committee - Professors Jerry B. Graves, Thomas J. Riley, Richard N. Story, and Paul H. Templet (Institute for Environmental Studies) - for their many constructive comments, suggestions, and reviews of my research work. Thanks also to Professor Abner M. Hammond for loaning me some of his pesticide application equipment. Mr. Lloyd Bailey and other members of the staff at the College of Agriculture, Science and
ill
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Education (formerly College of Agriculture), Jamaica, also are deserving of my thanks for their help with the portion of my work done in Jamaica. So too is Mr. Timothy Raiford and the field staff at Burden Research Plantation, Louisiana Agricultural Experiment Station, Baton Rouge, for their help with the part of my research conducted in Louisiana.
X am thankful to the Department of Entomology, Louisiana State University, and especially its head, Dr. Frank Guillot, for financial assistance in the form of an assistantship. The Environmental Foundation of Jamaica (EFJ) provided substantial financial support for the studies I did in Jamaica. I am very grateful to the former Executive Director of the EFJ, Dr. Terrence Thomas, for his help and encouragement. Mr. Lenworth Fulton, Managing Director of the College of Agriculture Jamaica Foundation, also contributed to my success by assisting with the cost of my accommodation for the duration of the studies I did in Jamaica.
Earlier in my life, Mrs. Marjorie Jackson provided me with guidance and direction for which I am very grateful. Also, Mr. Ralston Munn and Mrs. Floribel Morris-Barrett gave me opportunities which helped me to get on the path that led to where I am today; I thank them both.
Thanks also to my family members, especially my mother, Mrs. Enid Ivey, and friends who have provided moral support
iv
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

throughout the entire period of my work, study, reflection here at Louisiana State UniversityAgricultural and Mechanical College.
andand
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Table of ContentsDedication............................................ iiAcknowledgment*...................................... iiiList of Tables......................................viiiList of Figures....................................... xAbstract............. xiIntroduction........................................... 1
References Cited -Introduction..................... 4Literature Review........... 8
Cabbage Looper.................................... 8Biology and Life Cycle....................... 8Host Plants................................. 10Migration and Distribution................... 10Management of Cabbage Looper................. 12
Diamondback Moth.................. 13Biology and Life Cycle....................... 14Host Plants........... 16Migration and Distribution................... 17Management of Diamondback Moth............... 19
Pesticide Use Reduction: An Emerging Trend inSustainable Agricultural Practices................ 28The Integrated Pest Management Strategy and its Rolein Contemporary Agricultural Systems...............32
The Integrated Pest Management Concept........ 34IPM - A Definition........................... 35
Insect Migration and its Implications for IntegratedPest Management.................................. 36References Cited - Literature Review...............38
Chapter OneIntegration of Plant Resistance, Intercropping, and Microbial Control for Management of cabbage Looper and Diamondback Moth in Jamaica................... 53
Introduction.................................54Materials and Methods........................ 59Results..................................... 63Discussion.................................. 71References cited - Chapter One............... 77
vi
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Chapter TwoPotential of Plant Resistance and Bacillus thuringiensis for Control of Diamondback Moth in Janaica.......................................... 82
Introduction................................ 83Materials and Methods........................ 87Results................... ..................88Discussion.................................. 89References cited - Chapter Two................95
chapter ThreeToxicity of Three Fornulations of Bacillus thuringiensis Against Diamondback Moth..............98
Introduction....................... 99Materials and Methods....................... 102Results.................. .................104Discussion................................. 105References Cited - chapter Three............ 109
Chapter FourIntegration of Plant Resistance* Microbial Control* and an Action Threshold for cabbage Looper and Diamondback Management on cabbage in Louisiana 113
Introduction................................114Materials and Methods....................... 118Results.................................... 121Discussion................................. 134References Cited - Chapter Four............... 141
Chapter FiveInsecticide use Reduction in Management Programs for Cabbage Looper and Diamondback Moth...............146
Introduction .......................... 147Materials and Methods....................... 152Results. ................................ 155Discussion............ 160References Cited - Chapter Five.............167
summary and Conclusions...............................172Bibliography......................................... 176vita.................................................199
vii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

List of TablesTabls 1
Tablo l
Tabls 1
Tabls l.
Tabls 1.
Tabls 1.
Tabls 2.
Tabls 3.
Tabls 4.
Tabls 4.
. 1 Mean numbers + SEM of cabbage looper larvae per plant in all treatments compared with the full IPM treatment (Tropicana, companion crop, andB. thuringiensis)............................ 64
.2 Mean numbers + SEM of diamondback moth larvae per plant in all treatments compared with the full IPM treatment (Tropicana, companion crop, and B. thuringiensis)........................ 65
.3 Mean + SEM damage-free yield of cabbage (kg) per plant from all treatments compared with the full IPM treatment (Tropicana, companion crop, and B. thuringiensis)........................ 68
.4 Mean + SEM yield loss of cabbage (kg) per plant from all treatments compared with the full IPM treatment (Tropicana, companion crop, and B . thuringiensis)............................... 69
.5 Mean + SEM damage-free yield and value ofcabbage from all treatments.................. 70
.6 Damage-free yield and value of cabbage andtomato from all treatments................... 72
1 Mean numbers ± SEM of diamondback moth larvae per plant on three cabbage varieties treated with Agree 50 WP®............................ 90
1 Dose-Mortality response of a population of diamondback moth to three formulations of B. thuringiensis .............................. 106
1 Effect of insecticide treatment on abundance of cabbage looper, diamondback moth, lady beetles, ants, and spiders per plant on two cabbage varieties...................................123
2 Effect of cabbage variety on abundance of cabbage looper, diamondback moth, lady beetles, ants, and spiders per plant across all insecticide treatment regimes................124
viii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Table 4.3 Effect of insecticide treatment on damage-free yield, yield loss, damage rating per plant, and marketability of two cabbage varieties.......128
Table 4
Table 4
Table 4
Table 5
Table 5 *
Table 5,
Table 5.
.4 Effect of cabbage variety on damage-free yield, yield loss, damage rating per plant, and marketability of two cabbage varieties across all insecticide treatment regimes............130
.5 Effect of insecticide treatment on yieldparameters of two varieties of cabbage.......132
.6 Effect of variety on yield parameters of cabbage across all insecticide treatment regimes.....133
. 1 Effect of insecticide on abundance of cabbage looper, diamondback moth, lady beetles, ants, and spiders per plant across three application rates ..................................... 156
.2 Effect of application rate on abundance ofcabbage looper, diamondback moth, lady beetles, ants, and spiders per plant across three insecticides .............................. 158
3 Effect of insecticide on damage rating per plant and marketability of cabbage across three application rates...........................161
4 Effect of application rate on damage rating per plant and marketability of cabbage across three insecticides............................... 162
ix
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

List of FiguresFigurs 1.1 Total number of cabbage looper and
diamondback moth larvae per 400 plants on each sample date....................... 66
Figurs 2.1 Total number of diamondback moth larvae per240 plants on each sample date...........91
Figurs 4.1 Mean number of diamondback moth larvae perplant across insecticide treatments and cabbage varieties...................... 126
Figurs 4.2 Total number of cabbage looper anddiamondback moth larvae per 600 plants on each sample date....................... 127
Figurs 4.3 Mean marketability of cabbage acrossinsecticide treatments and cabbage varieties .............................131
Figurs 5.1 Total number of cabbage looper anddiamondback moth larvae per 600 plants on each sample date....................... 159
x
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

AbstractPopulation density estimates, yield data, damage rating, and marketability of cabbage were used to assess the effectiveness of integrating several control tactics - an action threshold, host plant resistance, intercropping (companion planting), microbial control, and pesticide use reduction - for managing the diamondback moth (DBM), Plutella xylostella (L.) , and the cabbage looper (CL), Trichoplusia ni (Hubner), in cabbage and in protecting beneficial arthropods. An action threshold of six DBM larvae per plant was inadequate for making insecticide application decisions because cabbage plots treated based on this action threshold had significantly higher densities of CL and DBM larvae, higher damage rating and yield loss, and lower damage-free yield and percentage of marketable cabbage compared with plots treated weekly. A new cabbage hybrid, Tropicana, was superior to two other varieties, KY Cross and Early Jersey, in resisting attack from DBM. The role of intercropping (companion planting) in insect control was inconclusive. Agree 50 WP® and WG®, microbial insecticides based on toxins of Bacillus thuringiensis Berliner, gave excellent results in controlling CL and DBM. Also, significant and favorable interactions between the bioinsecticide and the Tropicana cabbage variety indicated that they may be compatibly
xi
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

combined for DBM management. In addition, field results confirmed the potential of incorporating pesticide use reduction in management programs for lepidopteran pests of cabbage; plots treated at 25 and 50% of the recommended rates of Agree 50 WG®, Ambush 2 EC®, and Malathion 50® had essentially the same level of CL and DBM larval infestation, damage rating, and percentage of marketable cabbage as those treated at the recommended rates. Perhaps because only those actually in the canopy of plants were counted, population levels of generalist predators were generally low in treatment plots such that clear-cut treatment effects were not observed. These results indicate that, within the context of the pest management problems that have arisen because of the reliance on conventional insecticides as the sole control strategy for CL and DBM, IPM programs incorporating host plant resistance, microbial control, and pesticide use reduction may lead to a more sustainable management system for these pests.
xii
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

IntroductionThe world production of cruciferous vegetables is over
42.2 million tons. A complex of lepidopterous defoliators attack these crops, particularly cabbage, Brassica oleracea L. var. capitata L., broccoli, B. oleracea L. var. italica Plenck, and cauliflower, B. oleracea L. var. botrytis L., wherever they are grown. Of this insect pest complex, the diamondback moth (DBM), Plutella xylostella (L.) , is the most destructive (Talekar 1992 and references therein) . other insects such as the cabbage looper (CL) , Trichoplusia ni (Hubner), cabbage white butterfly, Pieris rapae (L.) and armyworms, Spodoptera spp., rarely reach population levels comparable with those of the DBM. In fact, DBM often represents greater than 75% of the insect pest population on cabbage and cause crop loss of up to 100%, making it the key insect pest from an economic standpoint (Magallona 1986) . Because the leaves are the marketable parts of the cabbage plant, the cumulative effects of DBM feeding damage are reductions in yield and quality of the produce. The DBM has had a marked impact on cabbage production in some areas; in Florida, for example, cabbage production declined from 19,600 acres in 1984-85 to 11,600 acres in 1991-92. This decline was attributed to the severity of attack from DBM and the concomitant high control costs for protecting cabbage, a
1
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

2
relatively low-value crop (USDA 1994). Talekar & Shelton (1993) estimated the annual cost worldwide for managing the DBM to be U.S. $1 billion. The cosmopolitan distribution and occurrence of DBM in economically damaging numbers results from its high rate of reproduction, adaptability to a wide range of climatic and environmental conditions, intensive cultivation of host plants, wide host range, absence of effective natural enemies in many areas, and migratory capability (Talekar & Shelton 1993) .
In most countries, use of synthetic insecticides is the main control tactic for DBM (Sun et al. 1986, Talekar & Shelton 1993). However, because of the heavy use of conventional insecticides and continuous production of crucifers, insecticide-resistant DBM biotypes have developed in many tropical and subtropical countries of the world. Resistance in DBM against a large number of formulations of synthetic insecticides from the carbamate, chlorinated hydrocarbon, organophosphate, and pyrethroid groups has been documented (Miyata et al. 1986, Fahmy & Miyata 1992, Sun 1992). In North America, the DBM was considered only a minor pest in the lepidopteran complex attacking crucifers (Marsh 1917, Harcourt 1958). However, in the last decade control failures and subsequent economic damage in crucifers have been reported in several states. Widespread resistance in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

3
DBM against several widely used insecticides was subsequently confirmed (Shelton & Wyman 1992, Shelton et al. 1993).
Because of the widespread development of insecticide- resistant populations of DBM and environmental, occupational, and public health problems associated with conventional insecticides (Pimentel et al. 1992, Cate & Hinkle 1994, NRC 1996) , a more rational approach to managing this pest is warranted. Integrated pest management (IPM) , "a pest management system that, in the context of the associated environment and the population dynamics of the pest species, utilizes all suitable techniques and methods in as compatible a manner as possible and maintains the pest populations at levels below those causing economic injury" (FAO 1968), is a viable option for managing DBM populations. Currently, some tactics which may be used in IPM programs for DBM management include biological control (Waage & Cherry 1992, Entwistle et al. 1993, USDA 1994), companion crops (Talekar et al. 1986, Srinivasan & Moorthy 1991), host plant resistance (Dickson & Eckenrode 1975; Adkisson & Dyck 1980; Dickson et al. 1984, 1986) , and reductions in the use of insecticide active ingredient (Trumble 1990, Hurst et al. 1992, Matteson 1995, Jacobsen 1996). Frequently, many of these different control components for DBM are investigated in isolation from one another. Such an approach is often necessary to assess the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

4
credibility of these potential management components, but the ultimate objective should be the integration of compatible control tactics for managing DBM populations. Therefore, the challenge facing entomologists is to successfully integrate potential management components into holistic, ecologically- sound, and economically feasible programs for managing DBM (and tangentially other insects in the crucifer pest complex such as CL) . Meeting this challenge was the objective of this study.
References Cited - IntroductionAdkisson, P. L. 8 V* A. Dyck. 1980. Resistant varieties in pest management systems, pp. 233-251. In F. G. Maxwell & P. R. Jennings (eds.), Breeding Plants Resistant to Insects. Wiley, New York.Cate, J. R. & M. R. Hinkle. 1994. Integrated Pest Management: The Path of a Paradigm. National AudubonSociety, Washington, D.C.Dickson, M. H. 8 C. j. Eckenrode. 1975. Variation in Brassica oleracea resistance to cabbage looper and imported cabbage worm in the greenhouse and fields. J. Econ. Entomol. 68: 757-760.Dickson, K. H., c. J. Eckenrode 8 A. E. Blamble. 1984. NYIR 9602, NYIR 9605, and NYIR 8329, lepidopterous pest-resistant cabbages. HortScience 19: 311-312.Dickson, M. H., C. J. Eckenrode 8 J. Lin. 1986. Breeding for diamondback moth resistance in Brassica oleracea, pp.137- 143. Jn N. S. Talekar 6 T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Entvistle, P. F, J. 8. Cory, M. J. Bailey 8 s. Higgs (eds.) 1993. Bacillus thuringiensis, an Environmental Biopesticide: Theory and Practice. Wiley, New York.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

5
Fahmy, A. R. £ T. Miyata. 1992. Development and reversion of chlorfluazuron resistance in diamondback moth, pp. 403- 410. In N. S. Talekar (ed.), Diamondback Moth and other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.FAO. 1968. Report of the first session of the FAO panel of experts on integrated pest control. FAO, Rome.Harcourt, D. 6. 1958. Biology of the diamondback moth ineastern Ontario. II. Life history, behavior and host relationships. Can. Entomol. 89: 554-564.Hurst, P. P., P. Beaumont, C. E. Jorgensen £ s. Winther. 1992. Pesticide reduction programmes in Denmark, The Netherlands, and Sweden. WWF International, Gland, Switzerland.Jacobsen, B. 1996. USDA IPM Initiative. United States Department of Agriculture. Beltsville, MD.MagaIlona, E. D. 1986. Developments in diamondback moth management in the Philippines, pp. 423-435. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Marsh, H. O. 1917. Life history of Plutella maculipennis, the diamondback moth. J. Agric. Res. 10: 1-10.Matteson, P. 1995. The "50% Pesticide Cuts" in Europe: A Glimpse of our Future? American Entomol. 41: 210-220.Miyata, T., T. Salto £ V. Noppun. 1986. Studies on the mechanism of diamondback moth resistance to insecticides, pp. 347-357. In N. S. Talekar £ T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.MRC. 1996. Ecologically Based Pest Management: NewSolutions for a New Century. National Research Council, National Academy Press. Washington, D.C.Pimentol, D., H. Acquay, M. Biltonen, P. Rice, M. Silva, J. Nelson, V. Lipner, 8. Giordano, A. Horowitz £ M. D'Amore. 1992. Environmental and economic costs of pesticide use. BioScience 42: 750-760.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

6
Shelton, A. M. & J. A. Wyman. 1992. Insecticide resistance of diamondback moth (Lepidoptera: Plutellidae) in NorthAmerica, pp. 447*454. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Shelton, A. K., J. A. wyaan, N. L Cushing, K. Apfelbeck, T. J Dennehy, S. E. R. Mahr & s. D. Eigenbrode. 1993.Insecticide resistance of diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae), in North America. J. Econ. Entomol. 86: 11-19.Srinivasan, R. £ P. N. K Moor thy. 1991. Indian mustard as a trap crop for management of major lepidopterous pests on cabbage. Tropical Pest Management. 37: 26-32.Sun, C. N. 1992. Insecticide resistance in diamondback moth, pp. 419-426. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Sun, C. N., T. K. wu, J. 8. Chen 6 W. T. Lee. 1986.Insecticide resistance in diamondback moth, pp.359-371. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Talekar, N. S. (ed). 1992. Diamondback Moth and otherCrucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Talekar, N. S., T. 8. Lee 6 S. W. Huang. 1986.Intercropping and modification of irrigation method for the control of diamondback moth, pp. 145-155. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Talekar, N. 8. £ A. M. Shelton. 1993. Biology, ecology and management of the diamondback moth. Annu. Rev. Entomol. 38: 275-301.Trumble, J. T. 1990. Vegetable insect control with minimal use of insecticides. HortScience 25: 159-164.USDA. 1994. Three-pronged attack on crop pests. Agricultural Research Service, Agricultural Research United States Department of Agriculture, Beltsville, MD.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

7
Waage, J. & A. Cherry. 1992. Quantifying the impact of parasitoids on diamondback moth, pp. 245-253. In N. s. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taivan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Literature R«vi«w cabbage Loopar
The common name "cabbage looper" (CL) of Trichoplusia. ni (Hubner), is somewhat of a misnomer because, far from being monophagous on cabbage, the adults oviposit and the larvae feed on a large number of plant species spanning 29 families. However, the CL is not equally stimulated or adapted to colonize all plant species, as evidenced by its reputation as a more serious pest of certain agricultural crops than of others. The leaves of cabbage and other cole crops appear to be a suitable substrate for adult oviposition and larval utilization by cabbage looper (Shorey 1963). Originally described in Europe, the CL was recognized in the U.S.A. before 1870 (Sutherland 1966) .Biology and Life Cycle
The adult CL is a mottled, grayish-brown moth with a wing span of about 40 mm. It has a small, silvery white figure 8 in the middle of each of the front wings. The moths are nocturnal fliers, but can be seen during the day resting on the underside of cabbage leaves. Female moths can lay 200-350 eggs, usually over a 10-12 day period. Eggs are white, round and about the size of a pinhead. Gravid moths usually lay eggs singly near the outer edges of lower leaves
8
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

9
and on plants not previously infested by loopers. Larvae hatch from the eggs three to six days after oviposit ion and immediately begin feeding on the underside of the leaf. The larvae will feed from two to four weeks. They are light- green with two white stripes on each side and two more stripes along the back. Young (early instars) larvae are yellowish to light-green and only 12.5 to 16 mm in length. Late instars (fifth-sixth) , grow to over 40 mm long, and inflict the most feeding damage. There are three pairs of slender thoracic legs and three pairs of thick prolegs on the abdomen. Unlike most Lepidoptera, there are no prolegs in the middle of the abdomen which is usually humped when the insect is moving. This characteristic is prevalent throughout all the larval stages and distinguishes loopers from other worms that feed on crucifers. On cabbage, young larvae feed between veins on the underside of the lower leaves. Large larvae make ragged holes in the foliage and generally move toward the center of the plant. They also can also burrow through three to six layers of tightly wrapped head leaves. Plants can be severely defoliated and stunted, producing no heads or becoming unfit for consumption. Large amounts of dark-green pellets excreted by the feeding looper may stain cauliflower heads, and the presence of looper
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

10
larvae in broccoli heads renders them unmarketable. The pupae are light-green initially but gradually turn dark-brown as they mature. They are about 20 mm in length and wrapped in a delicate cocoon of white silken threads. Pupae are usually found on the underside of lower leaves (shorey 1963, Bonnemaison 1965, Sutherland 1966, Robinson et al. 1995) . Host Plants
The CL exhibits polyphagia on at least 119 species, varieties, and cultivars of plants in 29 families, but favors cultivated crucifers, especially cabbage. Other common plants attacked include alfalfa, Medico sativa L.; beet, Beta vulgaris L./ celery, Apium graveolens L,; cotton, Gossypium hirsutum L.; lettuce; Lactuca sativa L.; parsley, Apium leptophyllum L.; potato, Solanum tuberosum L.; soybean, Glycine max (L.) Merr.; spinach, Spinacia oleracea L. ; and tobacco, Nicotina tobacum L. (Shorey et al. 1962, Sutherland 1966).Migration and Distribution
Migration in insects is a physiologically coordinated activity governed by environmental factors. From a physiological standpoint, migration is typified by an inhibition of responses to proximate stimuli generated by resources needed for "vegetative functions" (activities that
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

11
exploit available resources for growth and reproduction) . Migratory movement usually results from stimuli that forecast a shortage of resources and/or by the organism's ownendogenous rhythms. Migration can be differentiated from other types of movement because stimuli that would inhibit ranging or station keeping do not prevent an organism from migrating (Dingle 1996). The current thinking is that migration is an evolutionary adaptation which allows insects to displace themselves in time and space, therebycontributing to their fitness - survival and production of viable progeny. Migration essentially serves a specific ecological function of allowing insects to choose when and where to breed (Dingle 1986).
Many migratory insects of North America have been thesubject of much research on account of their economicimportance (Pashley et al. 1985; Johnson 1986, 1995; Johnson & Mason 1986, Gregory et al. 1990) . The CL is among the economically important noctuids that are highly mobile and cause damage to major food crops over wide areas. This insect cannot overwinter in the northern United States and moths annually migrate from winter habitats in the southern United states or Mexico to these areas and Canada from early July to late August, depending on the weather and airflow
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

12
patterns. Wolf et al. (1986) recovered the CL among the number of economically important species of Lepidoptera caught in light traps on oil platforms located in the Gulf of Mexico.
Undoubtedly, the CL's ability to migrate and disperse over long distances has contributed to the wide distribution of the CL; populations have been described in Europe, North and South America, the Hawaiian Islands, and Asia (Shorey 1963, Bonnemaison 1965, Sutherland 1966). It seems reasonable to believe that the distribution of CL mirrors to a great extent the distribution of cultivation of suitable host plants.Management of cabbage Looper
Late instars (fifth-sixth) of CL chew holes as large as 12.5 x 25 mm through the leaves of cabbage. This type of damage is cosmetically undesirable, resulting in significant economic losses (Sutherland 1966) . Typically, this insect has been controlled primarily with multiple applications of broad-spectrum insecticides. But this practice has led to the development of insecticide-resistant populations of DBM, an insect which hitherto the 1980s, was a minor pest in many cabbage growing areas (Marsh 1917, Harcourt 1954, Wyman 1992). The challenge therefore, is to find alternative, yet
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

13
effective control tactics, to conventional insecticides and weave them into integrated pest management approaches for successful, simultaneous management of these pests of crucifers.
Diamondback MothWorldwide, the most serious insect pest of cabbage and
other Brassica crops is the diamondback moth (DBM) , Plutella xylostella (L.) (Talekar 1992 and references therein). Crop losses caused by this insect may be as high as 50-100% (MagaIlona 1986, Sagenmuller & Rose 1986). All stages of DBM except the moth in flight may be found on host plants (Harcourt 1956).
The DBM was originally described in the genus Phalaena by Linnaeus in 1746, and later in 1758 by him as Phalaena Tinea xylostella. Curtis in 1832 changed its name to Cerostoma macnlipennis, and Zeller in 1843 named it Plutella crucifersurum. The DBM had several other genus- and species- name changes and it was not until 1966 that its name was changed by the International Commission on Zoological Nomenclature to Plutella xylostella (L.), the current widely accepted name (Bradley 1971, Moriuti 1986). There is still some lingering confusion regarding its family name because the literature is replete with papers placing the DBM in the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

14
family Plutellidae and other papers placing it in the family Yponomeutidae.Biology and Life Cycle
Adult DBHs are typically small and brownish-gray with a maximum wingspan of about 20 mm. Forewings are narrow, with a pale, wavy band along the posterior margin that forms three diamond-shaped patches when folded at rest. The hind wings are dark-gray. Females are usually lighter in color than males. The sexes can be positively identified by the anal segment; in males it is divided longitudinally on the ventral surface, whereas in females it is undivided, and females have a tuft of hair at the distal end of the abdomen. The moths are short-lived, nocturnal feeders on the nectar of host plants, but can be seen during the day resting on leaves. Females oviposit their eggs in the evenings, peaking on the day of emergence.
Oviposition lasts four days, during which the female lays 11-188 eggs. Eggs are minute, scale-like, oval, or cylindrical to oblong and vary from intense yellow to greenish-white to pale-green and are laid singly or in small groups on both sides of leaves of host plants. The ratio of eggs oviposited on the upper and lower leaf surfaces is 3:2; very few eggs are laid on stems and leaf petioles. Eggs are
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

15
laid preferentially in concavities of leaves rather than on smooth surfaces. Larvae hatch after an incubation period of between 3*8 days and are essentially leaf miners - they begin feeding in the spongy mesophyll layer just below the surface of the leaf tissue leaving the upper epidermis intact thus producing the characteristic "windowpane" effect. After several days (3-5) of feeding, they exit the leaf and all subsequent instars feed on the surface, where they produce larger complete holes. The entire plant may be riddled with holes under moderate to heavy populations. Larvae also feed in the developing heads of cabbage, causing deformed heads and encouraging soft rots.
Mature or fully grown fourth instars inflict the most damage. They are about 10-15 mm in length, light- to dull- green with a dark head capsule and short, scattered, and distinctly black setae. Pupae are of the obtect type (i.e., the appendages are glued to the body wall) , about 6 mm in length, and are yellowish to light-green initially. This color first changes to light-brown and later to dark-brown prior to adult eclosion. Pupation occurs in a netlike, silken cocoon on leaves or in soil debris (Marsh 1917, Hardy 1938, Robertson 1939, Harcourt 1956, Robinson et al. 1995).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

16
Host PlantsThe DBM exhibits monophagy on plants in the family
Cruciferae. Its food plant range coincides with the botanical distribution of the mustard oils (allyl isothiocyanates) and their glucosides (progoitrin, sinigrin, sinalbin, glucocheirolin, glucoconringlin, glucoerucin, glucotropaeolin, gluconasturtium, and gluconapin) (Gupta & Thorsteinson 1960). The significance of a mustard oil glucoside in the choice of food plants by an insect specific to Cruciferae was first observed by Verschaffelt (1910) . He showed that the gustatory effect of mustard oil glucosides in the host plants of DBM was to provide stimulus for ingestion, and that they were important determinants of host plant specificity. Host specificity in a monophagous insect is exhibited in two ways. The gravid female shows selectivity in its choice of a plant for oviposition, or the larvae will accept as food the leaves of only a restricted and often related number of plant species (Thorsteinson 1953). The DBM feeds on both cultivated plants and weeds from the family Cruciferae. Cultivated cruciferous plants attacked include, broccoli; Brassica oleracea L. var. italica Plenck; Brussells sprouts, B. oleracea var. L. gemmifera Zenker; cabbage, 3. oleracea L. var. capitata L.; Chinese cabbage, 3. chinensis
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

17
L.; cauliflower, B. oleracea L. var botrytis L. ; collards, B. oleracea L. var. virdis; mustard, B. juncea L. Czern; turnip, 3. rapa L.; and watercress, Rorippa aquaticum [L. ] (Thorsteinson 1953, Harcourt 1958, Talekar & Shelton 1993). Migration and Distribution
The evolution of wings and the ability to fly that resulted is one of the reasons for the success of the insects as a group (Borror et al. 1989). This ability enables them to increase their powers of movement by several orders of magnitude and to exploit habitats they could not otherwise use and greatly increases their environmental impact in addition to their response to environmental conditions (Rankin & Burchsted 1992). Migration in insects is a physiologically coordinated activity governed by environmental factors. As an evolutionary adaptation, migration allows insects to displace themselves temporally and spatially, thereby contributing to their survival and production of viable progeny by increasing their life history options. For the DBM, among several reasons responsible for its present cosmopolitan distribution is its ability to migrate and disperse over long distances. Mass migration of the DBM has been studied extensively in Britain, and its annual occurrence is attributed to migration by adult moths
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

18
from the Baltic and southern Finland - a distance greater than 3,000 km. These studies show that moths can fly continuously for several days covering distances of 1,000 km per day (Mackenzie 1958) . Smith & Sears (1982) reported that in eastern Canada, yearly populations of DBM originate by adult migrants from the United States. And in Japan, the DBM is known to migrate from southwestern islands, that are mainly warm subtropical, to the cooler islands of Honshu and Hokaido (Honda 1992).
On account of its migratory capability and other factors, not surprisingly, the DBM is widely distributed. Diamondback moth has been reported in at least 128 countries and territories and is believed to be the most universally distributed of all Lepidoptera. The present cosmopolitan distribution and occurrence of DBM in economically damaging numbers results from its high rate of reproduction, adaptability to a wide range of climatic and environmental conditions, intensive cultivation of host plants, wide host range within the Cruciferae plant family, absence of effective natural enemies in many countries, and migratory behavior (CIE 1967, Salinas 1972, Talekar & Shelton 1993). Ooi (1986) reported that DBM populations were found past latitude 60°N in Iceland, in temperate regions, and in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

19
tropical areas of the world. Argentina marks the southernmost limit of DBM distribution in the New World (Salinas 1986). Europe, where crucifers have been grown for a long time, was suggested by Hardy (1938) as the original habitat of the DBM. However, Chu (1986) believed the DBM originated in Asia Minor and spread to other parts of the world along with the spread of cultivation of its host plants. Marsh (1917) and Harcourt (1956) gave early accounts of the occurrence of the DBM in North America.Management of Diamondback Moth
In most countries, use of synthetic insecticides is the main control tactic for DBM (Sun et al. 1986) . in developing countries of tropical and subtropical areas, cultivation of cruciferous vegetables is characterized by small farms on which land, labor, and pesticides are intensively used. In these countries, the growers rely on frequent use of insecticides, often applied with knapsack sprayers without due regard to hazard to the applicator. By contrast, in developed countries, production of crucifers is characterized by large-scale farming methods, including less intensive use of labor, increase of management and capital, and the consolidation of farms into larger holdings. Large-scale production of cruciferous vegetables is typically done in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

20
North America and Europe and is undergoing rapid expansion in Mexico and Central America. In these countries, pest management decisions tend to be uniform over large production areas. The main method of control of DBM is insecticides applied aerially or with ground equipment (Talekar & Shelton1993) . Because of the reliance on use of broad-spectrum insecticides for control of DBM, both in developing and developed countries, resistance against chemicals in all major groups of insecticides - carbamates, chlorinated hydrocarbons, organophosphates, and pyrethroids - has been documented (Miyata et al. 1986, Fahmy & Miyata 1992, Sun 1992) . Insecticide resistance is a genetically based decrease in susceptibility of an insect population to an insecticide (NRC 1986). Because resistance involves changes in gene frequencies in a population, it is, by definition, an evolutionary phenomenon (Roush & Tabashnik 1990). The development of resistance takes place over time via the selection of individuals best suited to survive exposure to an insecticide. Genes for resistance almost always pre-exist at low frequencies in most insect species; exposure to an insecticide merely increases the allelic frequencies of genes conferring resistance by removing insects without these genes from the breeding population. Hence the development of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

21
insecticide resistance is just a case of accelerated evolution (Graves et al. 1993) . An extensive suite of mechanisms conferring resistance in DBM against many conventional insecticides has been elucidated. These mechanisms include: reduced cuticular penetration ofinsecticides, manifested by tarsal autotomy (Moore and Tabashnik 1989); decreased sensitivity of acetyl cholinesterase (Sun et al. 1986); enhanced detoxification by esterases (Maa and Chuang 1983); oxidative detoxification (Yao et al. 1988) ; and glutathione S-transferase conjugation (Cheng et al. 1983, Kao et al. 1989).
The development of resistance in DBM against so many insecticides has served as a catalyst for numerous research projects worldwide aimed at discovering alternative control measures. Concern over burgeoning economic losses from DBM damage to crucifers has led to the convening of two international conferences, and has resulted in the publication of more than 1,400 papers dealing with DBM biology, ecology, and management (Talekar & Griggs 1986, Talekar et al. 1990, Talekar 1992). Another set of reasons behind the quest for alternatives to conventional insecticides for control of DBM is the need to reduce the environmental (Mineau & Collins 1988), and occupational and
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

22
public health risks (GAO 1986, Ecobichon et al. 1990) related to synthetic insecticides.
A more rational, comprehensive, and ecologically-based approach to managing DBM which would ameliorate the problems alluded to above is integrated pest management (IPM) . Integrated pest management is developed around knowledge of the ecology of pests and takes maximum advantage of the natural mechanisms of pest suppression. At present, some tactics which may be used in IPM programs for DBM management include biological control (Waage & Cherry 1992, Entwistle et al. 1993), companion crops (Talekar et al. 1986, Srinivasan & Moorthy 1991), host plant resistance (Dickson & Eckenrode 1975; Adkisson & Dyck 1980; Dickson et al. 1984, 1986), and reductions in the use of insecticide active ingredient (Trumble 1990, Hurst et al. 1992, Reus et al. 1994, Matteson 1995, Jacobsen 1996).
Biological control of DBM centers around the use of hymenopteran parasitoids and toxins of the soil bacterium Bacillus thuringiensis Berliner. The International Institute of Biological Control (IIBC) has been involved for many years in the discovery and distribution of effective natural enemies of DBM. All stages of the DBM are attacked by numerous parasitoids and predators with parasitoids being the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

23
most extensively studied. Also, larvae and adult DBM often are attacked by polyphagous predators such as birds and spiders. In 1928, IIBC entomologists began exploration for parasitoids of DBM in Europe, and in 1936 made an initial shipment of DBM parasitoids to New Zealand (Thomas & Ferguson 1989) . Even though more than 90 parasitoid species attack DBM, only about 67% of them seem to be important. Of these, six species are egg parasitoids, 38 attack larvae, and 13 attack pupae. Among the larval parasites, many of the effective species belong to two main genera, Cotesia (= Apanteles) and Diadegma. Several species from the genus Diadromus, most of which are pupal parasitoids, also exert important control (CIBC 1985, Chua & Ooi 1986, Alam 1992, Ooi 1992, Talekar et al. 1992, Talekar & Shelton 1993). Most of these species are native to Europe where the DBM is believed, by some authorities, to have originated. In Romania, 25 species of parasitoids occur and parasitize between 80-90% of DBM (Mustata 1992). In some countries, for example the Cape Verde Islands, complete control of DBM with parasitoids has been achieved eliminating the need for chemicals totally (Waage & Cherry 1992). In other countries however, the potential impact of parasitoids on DBM populations is being compromised by excessive use of broad
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

24
spectrum insecticides (Yaseem 1978, Lim 1986, Pollard 1991, Alam 1992) . It has been established that parasitoids, if conserved, can significantly contribute to suppression of DBM populations. Parasitoids may be the cornerstone in developing sound IPM programs for DBM management because not only do they constitute the main factor that is capable of regulating pests, but also a naturally-occurring resource that is usually self-perpetuating (Lim 1982, 1992; Waage & Cherry 1992). Predators have been little studied, but they have been suggested to be important mortality factors for DBM populations (Sivapragasam et al. 1988, Alam 1992, Robinson et al. 1995).
Use of microbial agents for controlling DBM has been most successful with toxins of the soil bacterium B. thuringiensis. This is an aerobic, gram-positive, spore- forming bacterium found commonly in nature. Discovered in 1911, B. thuringiensis produces several toxins, significant among which are proteinaceous parasporal crystalline inclusions (also called delta-endotoxins) that are formed during sporulation. These protein crystals are very toxic to early instars of DBM and commercial formulations, referred to as microbial insecticides, have been used for insect control for over two decades (Navon 1993) . Upon ingestion by
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

25
insects, the crystalline inclusion is solubilized in the midgut, releasing the delta-endotoxins which are then activated by midgut proteases. Following proteolytic activation, B. thuringiensis toxins bind to the brush border membrane of the midgut epithelium and initiate the formation of pores that cause cells to swell and lyse, thereby disrupting membrane integrity which ultimately results in insect death (Gill et al. 1992, May 1993). In addition, the toxins of B. thuringiensis are environmentally benign, cause little or no harm to humans, most beneficial insects (parasitoids and predators) , and other nontarget organisms (Hofte & Whiteley 1989, Entwhistle et al. 1993). The use of this kind of insecticide needs to be conserved indefinitely. However, resistance in DBM against toxins of B. thuringiensis subsp. kurstaki has been reported (Kirsch & Schmutterer 1988, Tabashnik et al. 1990, Syed 1992, Shelton et al. 1993, Perez & Shelton 1997, Tang et al. 1997). Reduced binding affinity of toxins to the brush border membrane of the midgut epithelium has been identified as the primary mechanism of resistance to B. thuringiensis (McGaughey 1985, Van Rie et al. 1990). Other potential mechanisms of resistance include reduced solubilization or activation of toxins in the midgut before binding takes place and decreased sensitivity to
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

26
events that occur after binding such as membrane insertion and formation of pores (Gill et al. 1992, McGaughey & Whalon1992). The DBM is the first insect to develop resistance against B. thuringiensis in open field populations (Tabashnik 1994). Preventing further development of resistance against B. thuringiensis in populations of DBM that are not yet resistant and reversing resistance in populations that are already resistant, to preserve the longevity and effectiveness of B. thuringiensis, must be prioritized. The sensible approach is to combine use of B. thuringiensis with other compatible control measures in integrated control programs for DBM management.
Multiple cropping systems constitute agricultural systems diversified in time and space. Agriculturists have been using traditional systems of multicropping (the growing more than one crop on the same piece of land) for centuries, and intercropping (the growing of two or more crops at the same time on the same piece of land) is one type of cultural system that is a common feature in developing countries of the tropical world (Coaker 1987) . Intercropping has received renewed attention in recent times, especially among organic farmers, for insect control. It is believed that companion crops, in addition to creating diversity by attracting and
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

27
maintaining beneficial insects (Pimentel 1961, Cromartie 1981, Risch et al. 1983), also produce powerful chemical deterrents to many pest species, or may mask the chemical cues of crops that attract pests (Zimmerman 1978; Altieri & Liebman 1985; McClure 1994a, 1994b). Intercropping ofcabbage with other plants has been reported to have potential for suppressing DBM populations. Cabbage plants intercropped with Indian mustard, Brassica juncea (L.) Czern., (Srinivasan & Moorthy 1991); barley, Hordeum vulgare L.; dill, Anethum gravBolens L.; garlic. Allium sativum L.; safflower, Carthamus tinctorius L.; and tomato, Lycopersicon esculentum Mill., (Buranday & Raros 1973, Talekar et al. 1986) supported lower DBM populations compared with cabbage grown in monoculture. As with other potential components for DBM management, intercropping should be evaluated and given consideration when designing IPM programs.
Beck (1965) defined plant resistance as, "the collective heritable characteristics by which a plant species, race, clone, or individual may reduce the probability of successful utilization of that plant as a host by an insect species, race, biotype, or individual." Host plant resistance to insects can be an effective pest management tactic when used alone or when used in combination with other pest management
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

28
tactics (Painter 1951, 1958; Creighton et al. 1975; Kogan1982). The potential for using resistant crucifer varietiesfor DBM management has been investigated. Plant Introduction(PI) 234599, a cauliflower cultivar from Australia was foundto be highly resistant in the field to DBM (Dickson &Eckenrode 1980) . Breeding programs aimed at transferringgenes for DBM resistance from PI 234599 to cabbage resultedin the release of four cabbage breeding lines possessingresistance: NYIR 8329, NYIR 9602, and NYIR 9605 (Dickson etal. 1984). Eigenbrode & Shelton (1992) reported that thesehybrids have performed well in advanced trials and productiontrials. Further investigations of the mechanisms ofresistance in these hybrids against DBM were undertaken byother investigators (Stoner 1990, Eigenbrode et al. 1991).However, DBM-resistant cabbage varieties are not generallyavailable commercially. Therefore, the advent of a newcabbage variety (Tropicana), reputed resistant to DBM, makesit worthwhile to investigate its potential for use in IPMprograms for DBM management.Pesticide Use Reduction: An Emerging Trend in SustainableAgricultural Practices
Pesticide use reduction (PUR) is an emerging trend which is fast gaining credibility in agricultural production systems to mitigate the adverse environmental, occupational
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

29
and public health effects of pesticides (Trumble 1990; Moore 1994; Matteson 1995; Jacobsen 1996). This tactic is currently being implemented in several European countries as well as in the United States of America (U.S.A.)* Denmark, Sweden, and The Netherlands, three northern European countries, have implemented PUR-related policies mainly in response to environmental concerns about agrichemical pollution. In the U.S.A., two bills regarding PUR were introduced in the congress in 1994. The "Federal Insecticide, Fungicide and Rodenticide Act (FIFRA) Amendments of 1994" contained "pilot programs designed to establish and implement PUR goals in selected ecosystems, in cooperation with agricultural producers, federal, state, and local officials, and other appropriate public and private entities". Also, the "Farm Viability and Pest Management Improvement Act of 1994", co-sponsored by a number of legislators sought to establish a national program which emphasized PUR in agriculture, and pursued alternatives to chemicals used for urban and landscape pest control.
In addition to the legislative approaches alluded to above, in 1993 the Environmental Protection Agency (EPA) and the United States Department of Agriculture (USDA) started a consultative process for establishing regional, commodity
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

30
specific pesticide use reduction targets to be met by the year 2000. As a result of that process, the Pesticide Environmental Stewardship Program (PESP), a partnership to promote environmental stewardship in the use of pesticides among the EPA, USDA, and the U.S. Food and Drug Administration (FDA) , and volunteer grower organizations, and utility companies, was announced. Aided by federal government policies and programs, the participants from the private sector will be encouraged to minimize pesticide use through increased research, field demonstrations, and training in integrated pest management (IPM) and efficient pesticide application techniques. Improved pest management strategies will also be an integral part of regional environmental stewardship plans, inclusive of the management of vegetation ear-marked as rights-of-way for utility companies (Matteson 1995) . Another policy initiative aimed at significantly reducing pesticide usage in agriculture was announced in December 1994. The USDA announced an initiative to help farmers implement IPM on 75% of total crop acreage by the year 2000. This initiative is based on needs identified by farmers and others through a comprehensive needs identification and priority setting process involving every state. A participatory process, the initiative engages, in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

31
a partnership mode, the Land Grant University System and seeks to make USDA programs more sensitive to the needs of producers by increasing state and federal involvement in establishing priorities for IPM research, education, and adoption for specific commodities, and in the evaluation of progress towards achieving these goals. The efforts of USDA agencies are to be coordinated at all levels to make IPM research and education programs more effective. Additionally, the initiative establishes a new program for research into substitutes for pesticides that may be phased out because of pesticides resistance or regulation (OTA 1995, Jacobsen 1996, NRC 1996). The adverse impacts of excessive pesticide usage on the environment, occupational and consumer health risks, development of insecticide-resistant insect biotypes, success of pesticide use reduction policies in Europe, and the intention of the EPA, USDA, FDA, and other federal agencies to pursue initiatives aimed at PUR in American agriculture make it timely to commence research on reduced pesticide use in cruciferous vegetables to combat the key insect pest attacking them - the DBM.
Research on DBM must be pursued holistically instead of focusing on different potential management components in isolation from one another. Continued development (study and
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

32
assessment of potential management components) and implementation of IPM programs to combat DBM are necessary. All available control tactics that are ecologically-sound and economically viable must be integrated for managing DBM, because history has shown that single technology solutions are futile in the long-run.The Integrated Pest Management Strategy and its Role in Contemporary Agricultural Systems
With the advent of synthetic insecticides in the 1940s, easy and complete control of insect pests appeared at hand. However, it soon became obvious that there were problems associated with the use of insecticides: some insect pests developed resistance, beneficial and nontarget organisms were adversely affected, and pest resurgence occurred. Additionally, environmental and occupational and public health concerns arose (Carson 1962, Brader 1979, Wearing 1988) .
Since the beginning of the "Age of Pesticides" which commenced with the commercial introduction of DDT into U.S. agriculture in 1946 (Metcalf 6 McKelvey 1976), Metcalf (1980) enumerates and describes three subsequent distinct phases of the changing role of insecticides in crop protection.
These are the Era of Optimism 1946-1962, the Era of Doubt 1962-1976, and the Era of Integrated Pest Management
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

33
(IPM) 1976. Today, the protection of food and fiber crops from insect, mite, disease, and weed pests in conventional agricultural systems still relies primarily on the use of chemical pesticides. However, continued reliance on conventional broad spectrum insecticides, the insect control strategy of the past four decades, is a no-win strategy. The combined interaction of insect pest resistance, pest resurgences, and outbreaks of secondary pests provide ample evidence of the futility of complete reliance on the strategy of chemical control (Luck et al. 1977).
In addition, the economics of chemical pest control, including dramatic increases in the costs of insecticides and the costs of developing new insecticides, dictate a shift from routine prophylactic applications of insecticides to a policy of judicious use where needed (Metcalf & Luckmann 1994) . Also, increasing public concerns and awareness about both acute and chronic hazards to human health and about the ultimate impacts of pesticide usage on nontarget organisms and environmental quality can be dealt with only by fundamental changes in both the quantity and the quality of pesticides employed for pest management in conventional agricultural systems (Wearing 1988, Pimentel et al. 1992).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

34
The Integrated Pest Management (IPM) ConceptWith respect to the three phases of the changing role of
insecticides in crop protection, we are now in the Era of Integrated Pest Management (Metcalf 1980). The misuse, overuse, and unnecessary use of insecticides have been the main catalysts behind the groundswell of interest in IPM. That this is the case is certainly appropriate because the IPM concept aims to minimize the disadvantages in the use of insecticides and to optimize the advantages (Davies 1978; Pimentel et al. 1980; Metcalf & Luckmann 1994; Sorenson1994).
The goal of IPM is not to eliminate all pests; some pests are tolerable and essential so that their natural enemies remain in the crop. Rather, the aim is to reduce pest populations to less them damaging numbers. The control tactics used in IPM include pest resistant or tolerant plants, and cultural, physical, mechanical, biological, and chemical control. By using multiple control tactics, the chances of insects adapting to any one tactic are minimized. Also, IPM usually results in decreased reliance on conventional insecticides and thus reduces their adverse impacts on the environment while simultaneously conserving insect pest susceptibility to insecticides by reducing
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

35
selection pressure. Integrated pest management requires an understanding of the ecology of the cropping system, including that of the pests, their natural enemies, and the surrounding environment because IPM is developed around knowledge of the ecology of pests and takes maximum advantage of the natural mechanisms of pest suppression (Corbet & Smith 1976; Hoy & Herzog 1985; NRC 1986; Brown 1993; Sorenson1993) .IPM - A Definition
There are numerous definitions of IPM. The Food and Agriculture Organization (FAO) of the United Nations defines IPM as "a population management system that, in the context of the associated environment and the population dynamics of the pest species, utilizes all suitable techniques and methods in as compatible a manner as possible and maintains pest populations below those causing economic injury" (FAO 1968) . And, the Office of Technology Assessment of the U.S. Congress, gives its definition of IPM as "the optimization of pest control in an economically and ecologically sound manner, accomplished by the coordinated use of multiple tactics to assure stable crop production and to maintain pest damage below the economic injury level while minimizing hazards to humans, animals, plants, and the environment" (OTA
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

36
1995) . Whereas definitions of IPM vary, the philosophy of IPM is consistent - IPM encompasses the desire to bring together a combination of control tactics which optimize pest control (in terms of cost/benefit) while at the same time minimizing negative environmental, occupational, and public health impacts.
Within the context of the pest management problems that have arisen as a result of the reliance on conventional insecticides as the sole control strategy for CL and DBM, the importance of and the incentives for adopting instead an IPM approach are manifest.Insect Migration and its Implications for Integrated PestManagement
It is recognized that insect migratory behavior has important implications for pest management because a large number of the world's key agricultural pests, CL and DBM included, are long-distance migrants. To find solutions to the problems they cause through economic losses, it is necessary to have a clear understanding of the role that migration plays in their life history (Dingle 1996). Most arable crop plants are ruderals (plants that are colonizers of bare ground) and occur early on the botanical succession continuum. These are ephemeral habitats, and therefore many pests of arable crops have, from their evolutionary history,
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

37
high inherent levels of migratory activity (Southwood 1962). Successful colonizing insect species have high rates of reproduction, short generation times, and good dispersal capability (Dingle 1972, Stinner et al. 1983).
The nexus between insect migration and IPM arises from the unintended negative impacts of pesticide use in modern agroecosystems and the development of insecticide-resistant biotypes of important insect pests which precipitated the "ERA of Integrated Pest Management" (Metcalf 1980) . To be successful, an IPM program must take pest mobility into account. The ability to accurately forecast insect populations before migrants arrive may allow farmers lead time to initiate control measures such as the planting of trap crops or alternate hosts to encourage the build-up of natural enemies prior to the arrival of economically damaging numbers of insects (Dingle 1996). Therefore, designing strategies for the deployment of the fundamental components of IPM - pest-resistant crop varieties, biological control, genetically engineered organisms, and management of pesticide use to thwart the development of resistance - and systems for pest and for crop-loss forecasting should, ideally, take into consideration insect migration (Johnson 1995) . In addition, particularly with respect to resistance management, pest
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

38
mobility limits the ability of individual fanners to manageresistance, and thus complicates the economics of resistancemanagement at the farm level (Young 1979). Often, thefailure to adequately consider migration is one of the manyreasons why IPM programs sometimes give less than desirableresults (Anonymous 1994).
References Cited - Literature ReviewAdkisson, P. L. 6 V. A. Dyck. 1980. Resistant varieties in pest management systems, pp. 233-251. In F. G. Maxwell & P. R. Jennings (eds.), Breeding Plants Resistant to Insects. Wiley, New York.Alam, M. M. 1992. Diamondback moth and its natural enemies in Jamaica and some other Caribbean Islands, pp. 233-243. In N.s. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Altieri, M. A. 8 M. Liebman. 1985. Insect, weed, and plant disease management in multiple cropping systems, pp. 183-218. In C. A. Francis (ed.), Multiple Cropping Systems. Macmillan Publishing Company, New York.Anonymous. 1994. Movement and dispersal and IPM, pp. 136- 137. In Proceedings, Second National Integrated Pest Management Symposium/ Workshop. ESCOP Pest Management Strategies Subcommittee and ES IPM Task Force. University Graphics, North Carolina State University, Raleigh, NC.Beck, S. D. 1965. Resistance of plants to insects. Annu. Rev. Entomol. 10: 207-232.Bonnemaison, L. 1965. Insect pests of crucifers and their control. Annu. Rev. Entomol. 10: 233-256.Borror, D. J., C. A. Triplehorn 6 N. F. Johnson. 1989. AnIntroduction to the Study of Insects. Saunders College Publishing. Philadelphia, PA.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

39
Brader, L. 1979. Integrated pest control in the developing world. Annu. Rev. Entomol. 24: 225-254.Bradley, J. D. 1971. The identity of Phalaena Tinea xylostella Linnaeus, 1758 (Insecta, Lepidoptera). Bull. Zool. Nomencl. 28: 11-13.Brown, W. M., Jr. 1993. The role of IPM in contemporary agriculture and environmental issues, pp. 171-180. In A. R. Leslie & 6. W. Cuperus (eds.), Successful Implementation of Integrated Pest Management for Agricultural Crops. Lewis Publishers, Boca Raton, FL.Buranday, R. P. 6R. 8. Raros. 1973. Effects of cabbage tomato intercropping on the incidence and oviposition of the diamondback moth, Plutella xylostella (L.). Philipp. Entomol. 2: 369-374.Carson, R. 1962. Silent Spring. Houghton Mifflin, Boston, MA.CIBC. 1985. Annual report of 1984-1985. Commonwealth Institute of Biological Control, Commonwealth Agricultural Bureaux. London.CIE. 1967. Distribution Maps of Pests. Map No. 12, Plutella xylostella (L.) (maculipennis Curt.). Commonwealth Institute of Entomology. London.Cheng, E. Y., T. M. Chou I C. H. Kao. 1983. Insecticide resistance study in Plutella xylostella (L.). IV. The activities of glutathione S-transferase in the organophosphorous-resistant strains. J. Agric. Res. China. 32: 373-378.Chu, Y. 1986. The migration of diamondback moth, pp. 77-81. In N. S. Talekar & T.D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Chua, T. H. & P. A. C. Ooi. 1986. Evaluation of three parasites in the biological control of diamondback moth in the Cameron Highlands, Malaysia, pp. 173-184. In N. s. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

40
Coaker, T. H. 1987. Cultural methods: The crop, pp. 79-85. In A. J. Burn, T. H. Coaker & P. C. Jepson (eds.), Integrated Pest Management. Academic Press, NY.Corbet, P. S. £ R. F. Smith. 1976. Integrated control: A realistic alternative to misuse of pesticides?, pp. 661-682. In C. B. Huffaker & p. s. Messenger (eds.), Theory and Practice of Biological Control. Academic Press, NY.Creighton, C. S., T. L. McFadden 6 M. L. Robbins. 1975.Complementary influence of host plant resistance and microbial-chemical control of cabbage caterpillars. HortScience 10: 487-488.Cromartie, W. J. 1981. The environmental control of insects using crop diversity, pp. 223-251. In D. Pimentel (ed.), CRC Handbook of Pest Management in Agriculture. CRC Press, Boca Raton, FL.Davies, J. E., R. F. Smith 6 V. Fred. 1978. Agromedical approach to pesticide management. Annu. Rev. Entomol. 23: 353-366.Dickson, M. H. and C. J. Eckenrode. 1975. Variation in Brassica oleracea resistance to cabbage looper and imported cabbage worm in the greenhouse and fields. J. Econ. Entomol. 68: 757-760.Dickson, M. H. 6 C. J. Eckenrode. 1980. Breeding for resistance in cabbage and cauliflower to cabbage looper, imported cabbage worm and diamondback moth. J. Am. Soc. Hort. Sci. 105: 782-785.Dickson, M. H., c. J. Eckenrode 6 A. E. Blamble. 1984. NYIR 9602, NYIR 9605, and NYIR 8329, lepidopterous pest-resistant cabbages. HortScience. 19: 311-312.Dickson, M. H., C. J. Eckenrode £ J. Lin. 1986. Breeding for diamondback moth resistance in Brassica oleracea, pp. 137- 143. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Dingle, H. 1972. Migration strategies of insects. Science 175: 1327-1335.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

41
Dingle/ H. 1986. Evolution and genetics of insect migration, pp 11-26. In W. Danthanarayana (ed.), Insect Flight: Dispersal and Migration. Springer-Verlag, NY.Dingle, H. 1996. Migration, The Biology of Life on the Move. Oxford University Press, New York, NY.Ecobichon, D. J., J. E. Davies, J. Doull, M. Ehrich, R. Joy, D. McMillan, R. MacPhail, L. «. Reiter, V. Slikker £ H. Tilson. 1990. Neurotoxic effects of pesticides, pp. 131- 199. In C. F. Wilkinson & S. R. Baker (eds.). The Effects of Pesticides on Human Health. Princeton Scientific Publishing Co., Inc. Princeton, NJ.Eigenbrode, s. D., K. A. Stoner, A. M Shelton, £ w. c. Kain.1991. Characteristics of glossy leaf waxes associated with resistance to diamondback moth (Lepidoptera: Plutellidae) in Brassica oleracea. J. Econ. Entomol. 84: 1609-1618.Eigenbrode, 8. D. £ A. M. Shelton. 1992. Resistance to diamondback moth in Brassica: Mechanisms and potential for resistant cultivars, pp. 65-74. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Entwistle, P. P., J. S. Cory, M. J. Bailey £ s. Higgs (eds.) 1993. Bacillus thuringiensis, an Environmental Biopesticide: Theory and Practice. Wiley, NY.Fahmy, A. R. £ T. Miyata. 1992. Development and reversion of chlorfluazuron resistance in diamondback moth, pp. 403- 410. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.FAO. 1968. Report of the first session of the FAO panel of experts on integrated pest control. FAO, Rome.GAO. 1986. Pesticides: EPA's Formidable Task to Assess and Regulate Their Risks. General Accounting Office.Washington, D.C.Gill, S. S., E. A. Cowles £ P. V. Pietrantonio. 1992. Themode of action of Bacillus thuringiensis endotoxins. Annu. Rev. Entomol. 37: 615-636.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

42
Graves, J. B., J. A. ottea, B. R. Leonard, E. Burris a s.Micinski. 1993. Insecticide resistance management: Anintegral part of IPM in cotton, pp. 3-5. In J. Tarver & J. Lord (eds.), Louisiana Agriculture 36, Winter 1993.Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station, Baton Rouge, LA.Gregory, B. M., Jr., S. J. Johnson, A. V. Lievens, A. M. Hammond, Jr. 6 A. Delgado-Salinas. 1990. A midlatitudesurvival model of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Environ. Entomol. 19: 1017-1023.Gupta, P. O. St A. J. Thorsteinson. I960. Food plant relationship of diamondback moth, Plutella xylostella (Curt.) . II. Sensory relationship of oviposition of the adult female. Entomol. Exp. Appl. 3: 305-314.Harcourt, D. G. 1954. The biology and ecology of thediamondback moth, Plutella maculipennis Curtis, in eastern Ontario. Ph.D. dissertation, Cornell University, Ithaca, NY.Harcourt, D. G. 1956. Biology of the diamondback moth in eastern Ontario. I. Distribution, economic history, synonymy, and general descriptions. Quebec Soc. Proct. Plants, pp. 155-160.Harcourt, D. G. 1958. Biology of the diamondback moth in eastern Ontario. II. Life history, behavior and host relationships. Can. Entomol. 89: 554-564.Hardy, J. E. 1938. Plutella maculipennis Curt., its natural and biological control in England. Bull. Entomol. Res. 29: 343-372.Hofte, H. St H. R. Hbiteley. 1989. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 53 242- 255.Honda, K. 1992. Hibernation and migration of the diamondback moth in north Japan, pp. 43-50. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Hoy, M. A. 6 D. C. Herzog. 1985. Biological Control in Agricultural IPM Systems. Academic Press, NY.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

43
Hurst, p .P., p. Beaumont, C. E. Jorgensen 6 8. Hint her. 1992. Pesticide reduction programmes in Denmark, The Netherlands, and Sweden. WWF International, Gland, Switzerland.Jacobsen, B. 1996. USDA IPM Initiative. United States Department of Agriculture. Beltsville, MD.Johnson, S. J. 1986. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere. Insect Sci. Applic. 8: 543-549.Johnson, 8. J. 1995. Insect migration in North America: Synoptic scale transport in a highly seasonal environment, pp. 31-66. In V. A. Drake & A. G. Gatehouse (eds.), Insect Migration: Tracking Resources Through Space and Time.Cambridge University Press.Johnson, S. J. 6 L. J. Mason. 1986. The noctuidae: A case history, pp. 421-433. In D. R. MacKenzie, C. S. Barfield, G.G. Kennedy & R. D. Berger (eds.), Movement and Dispersal of Agriculturally Important Biotic Agents. Claitors Publishing. Baton Rouge, LA.Kao, C. H., C. F. Hung 6 C. N. Sun. 1989. Parathion and methyl parathion resistance in diamondback moth (Lepidoptera: Plutellidae) larvae. J. Econ. Entomol. 85: 1299-1304.Kirscb, K. 6 J. Schmutterer. 1988. Low efficacy of a Bacillus thuringiensis (Ber.) formulation in controlling the diamondback moth, Plutella xylostella (L.), in the Philippines. J. Appl. Entomol. 105: 249-255.Kogan, M. 1982. Plant resistance in pest management, pp. 93-134. In R. L. Metcalf & W. H. Luckmann (eds.), Introduction to Pest Management. Wiley, NY.Lim, G. S. 1982. The biology and effects of parasites on the diamondback moth, Plutella xylostella (L.). Ph.D. dissertation, University of London.Lim, 6. 8. 1986. Biological control of diamondback moth,pp. 159-171. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

44
Lin, G. 8. 1992. Integrated pest management of diamondbackmoth: Practical realities, pp. 565-576. In N. S. Talekar (ed.) , Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Luck, R. F., R. Van den Bosch 6 R. Garcia. 1977. Chemical insect control - a troubled management strategy. BioScience 27: 606-611.Maa, W. C. & M. L. Cbuang. 1983. Esterase of diamondback moth, (Plutella xylostella) (L.). I. Enzymatic properties of larval esterases. Bull. Inst. Zool. Acad. Sinica 22: 123- 131.Mackenzie, J. M. B. 1958. Invasion of diamondback moth (Plutella maculipennis Curtis). J. Entomol. Sci. 91: 247- 250.Magallona, E. D. 1986. Developments in diamondback moth management in the Philippines, pp. 423-435. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.May, R. M. 1993. Resisting resistance. Mature 361: 593- 594.Marsh, H. 0. 1917. Life history of Plutella maculipennis, the diamondback moth. J. Agric. Res. 10: 1-10.Matteson, P. 1995. The "50% Pesticide Cuts" in Europe: A Glimpse of our Future? American Entomol. 41: 210-220.McClure, 8. 1994a. Companion planting for pest control, pp. 22-3 3. In S. McClure & S. Roth (eds.), Rodale's Successful Organic Gardening: Companion Planting. Rodale Press, Emmaus, PA.McClure, s. 1994b. Introduction and companion planting basics, pp. 9-21. In S. McClure & S. Roth (eds.), Rodale's Successful Organic Gardening: Companion Planting. Rodale Press, Emmaus, PA.McGaughey, W. H. 1985. Insect resistance to the biological insecticide Bacillus thuringiensis. Science 229: 193-195.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

45
KcGaughey, w. H. 6 K. B. Whalon. 1992. Managing insect resistance to Bacillus thuringiensis toxins. Science 258: 1451-1455.Metcalf, R. L. 1980. Changing role of insecticides in crop protection. Annu. Rev. Entomol. 25: 219-256.Metcalf, R. L. & J. J. McKelvey, Jr. 1976. The Future of Insecticides. John Wiley & Sons, NY.Metcalf, R. L. 6 W. H. Luckmann. 1994. Introduction to Insect Pest Management. John Wiley & Sons, Inc. NY.Mineau, P. 6 B. T. Collins. 1988. Avian mortality in agroecosystems: Methods of detection, pp. 13-27. In M. P. Greaves, B. D. Smith & P. W. Greig-Smith (eds.), Field Methods for the Study of Environmental Effects of Pesticides. Proceedings of the British Crop Protection Council. Cambridge, UK.Miyata, T., T. Saito, i V. Noppun. 1986. Studies on the mechanism of diamondback moth resistance to insecticides, pp. 347-357. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Moore, A. 6 B. E. Tabashnik. 1989. Leg autotomy of adult diamondback moth (Lepidoptera: Plutellidae) in response to tarsal contact with insecticide residues. J. Econ. Entomol. 82: 381-384.Moore, M. 1994. Pesticide use reduction: An emerging trend in state policy. Sustainable Agriculture Newsletter 6: 1-3.Moriuti, S. 1986. Taxonomic notes on the diamondback moth, pp. 83-88. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Mustata, G. 1992. Role of parasitoid complex in limiting the population of diamondback moth in Moldavia, Romania, pp. 203-211. In N .S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the second International Workshop. AVRDC, Tainan, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

46
Navon, A. 1993. Control of lepldopteran pests with Bacillus thuringiensis, pp. 125-146. In P. F. Entwistle, J. S. Cory, M. J. Bailey & S. Higgs (eds.), Bacillus thuringiensis. An Environmental Biopesticide: Theory and Practice. John Wiley and Sons Ltd., N Y.NRC. 1986. Pesticide Resistance: Strategies and Tactics for Management. National Research Council, National Academy Press. Washington, D.C.NRC. 1996. Ecologically Based Pest Management: NewSolutions for a New Century. National Research Council, National Academy Press. Washington, D.C.Ooi, P. A. C. 1986. Diamondback moth in Malaysia, pp. 25-34. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Ooi, P. A. C. 1992. Role of parasitoids in managing diamondback moth in the Cameron Highlands, Malaysia, pp. 255- 262. In N. S. Talekar (ed.), Diamondback Moth and other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.OTA, 1995. Biologically Based Technologies for Pest Control. OTA-ENV-636. Office of Technology Assessment. Washington, D.C.Painter, R. H. 1951. Insect Resistance in Crop Plants. Macmillan, New York.Painter, R. H. 1958. Resistance of plants to insects. Annu. Rev. Entomol. 3: 267-290.Pashley, D. P., S. J. Johnson 6 A. N. Sparks. 1985. Genetic population structure of migratory moths: The fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 78: 756- 762.Perez, C. J. 6 A. M. Shelton. 1997. Resistance of Plutella xylostella (Lepidoptera: Plutellidae) to Bacillusthuringiensis Berliner in Central America. J. Econ. Entomol. 90: 87-93.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

47
Pimentel, D. 1961. Species diversity and insect population outbreaks. Prot. Ecol. 2: 77-114.Pimentel, D., D. Andow, R. Dyson-Hudson, D. Gallahan, s. Jacobson, M. Irish, 8. Kroop, A. Moss, I. Schreiner, M. Shepard, T. Thompson 6 B. vinsant. 1980. Environmental and social costs of pesticides: a preliminary assessment. Oikos 34: 126-140.Pimentel, D., H. Acquay, M. Biltonen, P. Rice, M. Silva, J. Nelson, v. Lipner, s. Giordano, A. Horowitz 6 M. D'Amore.1992. Environmental and economic costs of pesticide use. BioScience. 42: 750-760.Pollard, G. ▼. 1991. Constraints to IPM development and astrategy for management of tomato and cabbage pests in Trinidad, West Indies. Tropical Pest Management 37: 59-62.Rankin, M. A. 6 C. A. Burchsted. 1992. The cost of migration in insects. Annu. Rev. Entomol. 37: 533-559.Reus, J. A. W. A., H. J. Weckseler 6 G. A. Pak. 1994.Towards a future EC pesticide policy: An inventory of risks of pesticide use, possible solutions and policy instruments. Center for Agriculture and Environment. Utrecht, The Netherlands.Risch, S. J., O. Adow 6 M. A. Altieri. 1983. Agroecosystem diversity and pest control: Data, tentative conclusions and new research directions. Environ. Entomol. 12: 625-629.Robertson, P. L. 1939. Diamondback moth investigation in New Zealand. N. Z. J. Sci. Technol. 20: 330-364.Robinson, D. E., K. M. Dalip fi A. Mansingh. 1995.Integrated Management of Pests and Pesticides in the Caribbean. Department of Zoology, University of the West Indies, Mona, Kingston. The Jamaica National Commission for UNESCO. Kingston.Roush, R. T. t B. E. Tabashnik (eds.) 1990. Pesticide Resistance in Arthropods. Chapman & Hall, NY.Sagenmuller, A. 6 E. Rose. 1986. Hoe 522 (CME 134), a new insect growth regulator for control of diamondback moth, pp. 271-278. In N. S. Talekar & T. D. Griggs (eds.), Diamondback
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

48
Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Salinas, P. J. 1972. Studies on the ecology and behavior of the larvae of Plutella xylostella (Linnaeus) (Lepidoptera: Plutellidae). Ph. D. Dissertation, University of London.Salinas, P. J. 1986. Studies on diamondback moth in Venezuela with reference to other Latin American countries, pp. 17—24. In N. S. Talekar & T. D. Griggs (eds.),Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Shelton, A. M., J. L. Robertson, J. D. Tang, c. Perez, S. D. Eigenbrode, H. K. Preisler, W. T. Wilsey 6 R. J. Cooley.1993. Resistance of diamondback moth (Lepidoptera: Plutellidae) to Bacillus thuringiensis subspecies in the field. J. Econ. Entomol. 86: 697-705.Shorey, H. H. 1963. The biology of Trichoplusia ni(Lepidoptera: Noctuidae). 111. Response to the oviposition substrate. Ann. Entomol. Soc. Am. 56: 165-170.Shorey, H. H., L. A. Andres & R. L. Hale. 1962. The biology of Trichoplusia ni. 1. Life history and behavior. Ann. Entomol. Soc. Am. 55: 592-597.Sivapragasam, A., Y. Ito 6 T. Saito. 1988. Population fluctuations of the diamondback moth, Plutella xylostella (L.) on cabbages in Bacillus thuringiensis sprayed and nonsprayed plots and factors affecting within-generation survival of immatures. Res. Popul. Ecol. 30: 329-342.Smith, D. B. St M. K. Sears. 1982. Evidence for dispersal of diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae), in southern Ontario. Proc. Entomol. Soc. Ont. 113: 21-28.Sorenson, A. A. 1993. IPM and growers: An evolution inthinking, pp. 129-150. In A. R. Leslie & G. W. Cuperus(eds.), Successful Implementation of Integrated Pest Management for Agricultural Crops. Lewis Publishers, Boca Raton, FL.Sorenson, A. A. 1994. The role of IPM in reducing environmental problems, pp. 17-24. In Proceedings, Second
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

49
National Integrated Pest Management Symposium/Workshop. ESCOP Pest Management Strategies Subcommittee and ES IPM Task Force. University Graphics, North Carolina State University, Raleigh, NC.Southwood, T. R. E. 1962. Migration of terrestrial arthropods in relation to habitat. Biol. Rev. 37: 171-214.Srinivasan, K. £ P. N. K Moor thy. 1991. Indian mustard as a trap crop for management of major lepidopterous pests on cabbage. Tropical Pest Management. 37: 26-32.Stinner, R. E., C. s. Barfield, J. L. Stimac £ L. Dohse. 1983. Dispersal and movement of insect pests. Annu. Rev. Entomol. 28: 319-335.Stoner, K. A. 1990. Glossy leaf wax and plant resistance to insects in Brassica oleracea under natural infestation. Environ. Entomol. 19: 730-739.Sun, C. N. 1992. Insecticide resistance in diamondbackmoth, pp. 419-426. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Sun, C. N., T. K. Wu, J. S. Chen £ W. T. Lee. 1986.Insecticide resistance in diamondback moth, pp. 359-371. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Sutherland, D. W. s. 1966. Biological investigations of Trichoplusia ni (Hubner) and other lepidoptera damaging cruciferous crops on Long Island, New York. Memoir 399, Cornell University Agricultural Experiment Station, New York State College of Agriculture, Ithaca, NY.Syed, A. R. 1992. Insecticide resistance in diamondbackmoth in Malaysia, pp. 437-442. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Tabashnik, B. E. 1994. Evolution of resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 39: 47-79.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

50
Tabashnik, B. S., N. L. Cushing, N. Pinson & M. w. Johnson. 1990. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae) . J. Econ. Entomol. 83: 1671-1676.Talekar, N. 8. (ed). 1992. Diamondback Moth and OtherCrucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Talekar, N. 8. & T. D. Griggs (eds.). 1986. DiamondbackMoth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Talekar, M. S., T. S. Lee 6 8. W. Huang. 1986.Intercropping and modification of irrigation method for the control of diamondback moth, pp. 145-155. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Talekar, N. S., J. C. Yang 6 S. T. Lee. 1990. Annotated bibliography of diamondback moth, vol. 2. AVRDC, Shanhua, Taiwan.Talekar, N. S., J. C. Yang 6 S. T. Lee. 1992. Introduction of Diadegma semiclausum to control diamondback moth in Taiwan, pp. 263-270. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Talekar, N. 8. 6 A. M. Shelton. 1993. Biology, ecology and management of the diamondback moth. Annu. Rev. Entomol. 38: 275-301.Tang, J. D., S. Gilboa, R. T. Roush 6 A. M. Shelton. 1997.Inheritance, stability, and lack-of-fitness costs of field- selected resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae) from Florida. J. Econ.Entomol. 90: 732-741.Thomas, W. P. 6 A. M. Ferguson. 1989. Plutella xylostella (L.), diamondback moth (Lepidoptera: Yponomeutidae), pp. 135-142. In P. J. Cameron, R. L. Hill, J. Bain & W. P. Thomas (eds.), A Review of Biological Control of Invertebrate Pests and Weeds in New Zealand 1874-1987. CAB International, Wallingford, UK.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

51
Thorsteinson, A. J. 1953. The chemotactic responses that determine host specificity in an oligophagous insect (Plutella maculipennis) [Curt.] Lepidoptera). Can. J. Zool. 31: 52-72.Trumble, J. T. 1990. Vegetable insect control with minimal use of insecticides. HortScience 25: 159-164.Van Rie, J., W. H. McGaughey, D. E. Johnson, B. D. Barnett 6H. Van Mellaert. 1990. Mechanisms of insect resistance to the microbial insecticide Bacillus thuringiensis. Science 247: 72-74.Verschaffelt, E. 1910. The cause determining the selection of food in some herbivorous insects. Proc. Amsterdam Acad. Sci. 13: 536-542.Waage, J. 6 A. Cherry. 1992. Quantifying the impact of parasitoids on diamondback moth, pp. 245-253. In N. S.Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Wearing, C. H. 1988. Evaluating the IPM implementation process. Annu. Rev. Entomol. 33: 17-38.Wolf, w. w., A. M. Sparks, s. D. Pair, J. K. Westbrook & F. M. Truesdale. 1986. Radar observations and collections of insects in the Gulf of Mexico, pp. 221-234. In w.Danthanarayana (ed.), Insect Flight: Dispersal andMigration. Springer-Verlag, New York.Wyman, J. A. 1992. Management approaches for cruciferous insect pests in central North America, pp. 503-509. Jn N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Yao, M. G., C. F. Hung 6 C. M. Sun. 1988. Fenvalerateresistance and aldrin epoxidation in larvae of thediamondback moth. Pestic. Biochem. Physiol. 30: 272-278.Yaseem, M. 1978. The establishment of two parasites of the diamondback moth Plutella xylostella (Lep: Plutellidae) in Trinidad, W.I. Entomophaga 23: 111-114.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

52
Young, J. R. 1979. Assessing the movement of the fall armyworm (Spodoptera furgiperda) using insecticide resistance and wind patterns, pp. 344-351. In R. L. Rabb & G. G. Kennedy (eds.), Movement of Highly Mobile Insects: Concepts and Methodology in Research. University Graphics, North Carolina State University, Raleigh.Zimmerman, L. 1978. Strange bedfellows: The voodoo and science of companion planting, pp. 73-77. In J. Lawrence (ed.) , The Harrowsmith Reader. Camden House Publishing Ltd., Canada.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Chapter Oaa
Integration of Plant Resistance, Intercropping, and Microbial control for Management of Cabbage Looper and Diamondback Moth in Jamaica
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

54
IntroductionThe diamondback moth (DBM), Plutella xylostella (L.) , is
the most serious insect pest of cabbage, Brassica oleracea var capitata L., and other cruciferous vegetables in the Caribbean (Salinas 1986, Alam 1992). In Jamaica, of 14 insect pests feeding on cabbage, armyworms Spodoptera spp., cabbage looper (CL), Trichoplusia ni (Hubner), cabbage white butterfly, Pieris rapae L., and DBM, cause the most damage. Collectively, crop damage from these insects is between 74 to 100% and generally occurs on the outer leaves, but frequently involve head injury, especially when populations are high. Of this insect pest complex, DBM alone accounts for greater than 75% of the insect pest populations and causes up to 90% crop loss, and is of equal economic importance on the plains, in sub-mountains, and hilly areas of the island (Alam 1992) . The level of DBM infestation varies with seasonal climatic conditions; populations are highest and damage most severe during the hot, humid periods of the year (June to mid-July) and during the hot, dry periods of July and August. By contrast, populations are lower and damage less severe during periods of high rainfall because rainfall is a major mortality factor in the life history of this insect (Harcourt 1963, 1986, Talekar et al. 1986, Sivapragasam et al. 1988).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

55
Cabbage production in Jamaica is concentrated in Douglas Castle and surrounding districts of Macknie, Kellits, and Murry Mountains, an area of about 12 km2 and 800 m above sea level in the north-central section of the island. Production is characterized by small farms (<0.40-2.0 ha), intensive use of land, labor, and pesticides and is geared toward meeting the demand for fresh vegetables by residents in urban centers such as May Pen, Mandeville, Kingston, and by the tourism industry, especially in the nearby resort town of Ocho Rios. Cultivation of crucifers, especially cabbage, is an important source of income for farmers, and so to meet quality standards which demand that cabbage heads and wrapper leaves be damage-free, they rely on and excessively use conventional insecticides for control of DBM. Over 18 different insecticides from the major insecticide groups have been used against DBM in Jamaica since the 1970s. However, because of its genetic plasticity, short generation time and high rate of reproduction, DBM populations have become less susceptible to many insecticides. Farmers often, in response to low efficacy of chemicals, use mixtures of several chemicals, increase doses of insecticides, and spray more frequently, sometimes once every other day. These practices simply exacerbate the problem and have caused the DBM to become resistant to practically all insecticides. In fact, the DBM
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

56
has several-fold resistance to most insecticides available in Jamaica (Robinson et al. 1995). Ineffective insecticides include diazinon, malathion, and profenofos (organophosphates); carbary1 and methomyl (carbamates); and deltamethrin and permethrin (pyrethroids) (Suah & Ellis 1974, Alam 1992) . The threat to crucifer production from DBM resistance against conventional insecticides and concern about their adverse environmental effects has led to increased interest in alternative control approaches. Integrated pest management (IPM) is one such alternative. The philosophy of IPM envisions the use of complementary strategies, especially where no single method seems to be effective, for insect pest control. Some control tactics which are complementary to each other, and which can be used in an IPM program for DBM management in Jamaica include host plant resistance, companion cropping, and biological control.
Plant Introduction (PI) 234599, a cauliflower cultivar with shiny green leaves, has been investigated for resistance to the major lepidopteran pests of crucifers, including DBM. It has been used in breeding programs aimed at transferring genes for resistance to other breeding lines of cauliflower and cabbage (Dickson & Eckenrode 1975, 1980, Eckenrode et al. 1986, Eigenbrode et al. 1991). These programs resulted in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

57
the release of four cabbage breeding lines possessing resistance: NYIR 8329, NYIR 9602, and NYIR 9605 (Dickson et al. 1984) . Eigenbrode & Shelton (1992) reported that these hybrids have performed well in advanced trials and production trials in New York and under pressure from high levels of infestation of insecticide-resistant DBM in Honduras. However, DBM-resistant cabbage varieties are not generally available commercially. But a new cabbage hybrid, Tropicana, reputed to be capable of resisting DBM, recently become available to cabbage growers in Jamaica in 1995. Therefore, the advent of this new cabbage hybrid creates an opportunity to investigate its potential for use in IPM programs for DBM management.
Intercropping, the practice of growing two or more crops together simultaneously on the same piece of land, is a common cultural practice in tropical countries where farms are small and land use is intensive. As an insect pest management tactic, there has been increased interest in this practice because for some crop-insect situations, intercropping has reduced pest populations. For cabbage, plants intercropped with Indian mustard, Brassica juncea (L) Czern., (Srinivasan & Moorthy 1991); barley, Hordeum vulgare L.; dill, Anethum graveolens Lgarlic, Allium sativum L.; safflower, Carthamus tinctorius L.; and tomato, Lycopersicon
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

58
esculentum Mill., (Burunday & Raros 1973, Talekar et al. 1986) , supported lower DBM populations compared with plants grown in pure stand. In Jamaica, intercropping is widely practiced, especially by small farmers seeking to hedge their bets against the failure of one crop and also to maximize output from their limited land holdings, but is not currently used as part of an IPM program for DBM management.
Biological control of DBM has focused on hymenopteran parasitoids (Lim et al. 1986, Wagge & Cherry 1992) and the microbial insecticide, B. thuringiensis Berliner (Shelton et al. 1993). Alam (1992) reported that a rich and diverse complex of parasitoids: Trichogramma sp., Diadegma insulare [Cresson], Cotesia plutellae [Kurdjumov], Oomyzus sokolovrskii [Kurdjumov], and Trichospilus diatraeae [Cherian and Margabandhu] and predators: coccinellids, chrysopids,syrphids, and staphylinids attacked DBM in Jamaica. Also, Robinson et al. (1995) reported that 15 species of spiders preyed on DBM larvae in Jamaica. Viewed ecologically, this natural enemy complex is an invaluable asset which needs to be conserved, since conservation permits natural enemies to operate to their full effectiveness. However, an IPM approach, employing several control measures simultaneously against DBM with the aim of conserving and complementing its natural enemy complex, has not been attempted in Jamaica. In
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

59
the 1970s, the Agriculture Ministry in Barbados and also the International Institute of Biological Control, Trinidad, West Indies, spearheaded research on the biological control of DBM in the Caribbean (Alam 1974, 1989, Bennett & Yaseem 1972). These efforts were subsequently expanded under the auspices of the Caribbean Agricultural Research and Development Institute (CARDI) to other islands including Jamaica. The main thrust of these efforts was the use of hymenopteran parasites, many of which were decimated by high levels of insecticide use by vegetable farmers (Pollard 1991, Jones 1985, Alam 1992) . Therefore, the objective of this study was to integrate several control tactics - host plant resistance, companion cropping, and foliar applications of B. thuringiensis - for control of DBM (and tangentially CL) on cabbage and additionally conserve beneficial arthropods in vegetable production areas in Jamaica.
Materials and Methods This experiment was initiated in May 1995 on the crop
farm of the College of Agriculture, Science, and Education, Port Antonio, Portland, Jamaica. Five treatments were replicated four times and randomly assigned to plots in a randomized complete block design. The treatments consisted of combinations of two cabbage varieties, Tropicana (a new hybrid reputed resistant to DBM) (Petoseed, Saticoy, CA) and
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

60
KY Cross (the standard variety widely grown by fanners in Jamaica) (Takii Seed, Kyoto, Japan); a companion crop (tomato); a commercial formulation of B. thuringiensis subspp. kurstaki and aizawai (Agree 50 WP® [wettable powder]) (Novartis, Greensboro, NC); and a pyre thro id insecticide (Karate 2.5 EC [emulsifiable concentrate]) (Zeneca Agrochemicals, Wilmington, DE) . Specific treatment combinations were:
■ Tropicana, companion crop plus Agree 50 WP® (the full IPM package),
■ Tropicana plus companion crop,■ Tropicana plus Agree 50 WP®,■ KY Cross, companion crop plus Agree 50 WP®, and■ KY Cross plus Karate 2.5 EC.
Individual treatment plots were 9.09 m x 9.09 m and wereseparated by a distance of 9.09 m. Plant rows were 9.09 mlong and 0.91 m wide. Four-week-old cabbage seedlings of Tropicana and KY Cross, grown in outdoor seedbeds, were planted 0.45 m apart on raised beds. Four week-old tomato plants (the companion crop, variety Roma) (Petoseed, Saticoy, CA), also grown in an outdoor seedbed, were planted four weeks earlier than the cabbage plants. These tomato seedlings were planted in two rows on the sides of beds (with cabbage seedlings planted down the middle) in appropriate
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

61
plots. The plots were sampled weekly by visual inspection of leaf surfaces for seven weeks between 29 August and 10 October. Counts of CL and DBM larvae and generalist predators (assassin bugs [Hemiptera: Reduviidae], groundbeetles [Coleoptera: Carabidae], lady beetles (Coleoptera: Coccinellidae], and ants [Hymenoptera: Formicidae]) within their canopy were made from 20 randomly selected cabbage plants in each plot.
Additionally, 20 third to fourth instar DBM were hand- collected from each treatment plot. These larvae were placed in plastic bags which were in turn placed in a cooler and transported to the laboratory where they were observed for parasitoid emergence. Larvae were reared until adult eclosion in petri dishes (9 cm diameter) on top of a filter paper moistened with water. Fresh leaves of cabbage were provided as needed.
The microbial insecticide, Agree 50 WP® [3.8% (AI) (25,000 IU per mg), was applied weekly (after sampling), at the recommended rate of 83 g in 15 liters of water, with a Solo 475 Knapsack Sprayer (Solo Incorporated, Newport News, VA) to appropriate plots based on the treatment combinations listed above.
Other plots, again based on the treatment combinations, also were treated at the same time with Karate 2.5 EC
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

62
(25g/liter lambda-cyhalothrin) at the recommended rate of 15 mis in 15 liters of water. At crop maturity, yield data were collected from 40 randomly selected cabbage plants in each plot. These plants were cut and taken to the edge of the field where the frame leaves were removed leaving four wrapper leaves as per the specifications for fresh market cabbage meeting the US No. 1 grade (USDA 1945) .
In order to get an index (in terms of weight) for quantifying the magnitude of the impact of CL and DBM damage to cabbage, the four wrapper leaves in addition to head leaves which showed damage by larvae were then removed, counted, and weighed after which the heads were weighed. The weight so obtained for the heads was regarded as damage-free yield. And the combined weight of the damaged wrapper and head leaves was regarded as yield loss. Data were subjected to analysis of variance (ANOVA) using PROC GLM of the SAS system (SAS Institute 1989).
Additionally, it was decided a priori, to use Dunnett's t-test (SAS Institute 1989) to compare all treatments with the full IPM treatment. Of particular interest was the comparison of the full IPM treatment with the KY Cross plus pyrethroid treatment because the latter is the treatment farmers currently use. Tomato fruits were harvested and weighed; these data however, were not analyzed statistically.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

63
ResultsThe overall ANOVA F-test indicated that there were
significant treatment effects for CL (F - 11.72; df = 4; P - 0.0004), DBM (F = 60.74; df « 4; P * 0.0001), damage-free yield (F = 52.73; df - 4; P = 0.0001), and yield loss (F = 29.70; df = 4; P = 0.0001). Comparisons of all treatments with the full IPM package, using Dunnett's fc-test (t = 2.807; df — 1; a = 0.05), showed that CL larval population density in the full IPM treated plots was significantly lower than in the Tropicana plus companion crop treated plots, but was not significantly different from that observed in plots receiving the other three treatments (Table 1.1). The population density of DBM larvae was significantly lower in the full IPM treated plots compared with the KY Cross plus pyrethroid treated plots, but was not significantly different from that observed in plots receiving the other three treatments (Table1.2) . Over the sampling period, numbers of CL larvae were initially low, followed by a peak in the middle and then plummeting at plant maturity and end of sampling at the end of the growing season. On the other hand, numbers of DBM larvae were low at the beginning of sampling but increased dramatically over the growing season and up to the end of the sampling at plant maturity (Figure 1.1). In the case of predators, the effects of the different treatments were
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 1.1. Mean numbers + SEM of cabbage looper larvae per plant in all treatments compared with the full IPM treatment (Tropicana, companion crop, and B. thuringiensis)
Treatment comparisons No. larvae Significance3T fopicanaV companiohc 3.
KY Cross, companion crop, & Bt. - IPM 3.55+0.08-2.14+0.56 NS
Tropicana & Bt. - IPM 5.59+1.78-2.14+0.56 NS
KY Cross & pyrethroid - IPM 1.06+0.59-2.14+0.56 NS
aDunnett's T = 2.807, df = 12, a = 0.05; S, significant; NS, not significant; Bt., Bacillus thuringiensis.
o\

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 1.2. Mean numbers + SEM of diamondback moth larvae per plant in all treatments compared with the full IPM treatment (Tropicana, companion crop, and B. thuringiensis)
Treatment comparisons No. larvae Significance8Tropicana, companion^ c IPM..........
KY Cross, companion crop, & Bt. - IPM 0.28+0.08-0.16+0.07 NS
Tropicana & Bt. - IPM 0.24+0.09-0.16+0.07 NS
KY Cross & pyrethroid - IPM 16.9 + 2.18 - 0.16 + 0.07 S
aDunnett's T = 2.807, df = 12, a = 0.05; S, significant; NS, not significant; Bt., Bacillus thuringiensis.
C\ui

No. o
l Lar
vae
300
200-
100
(•Sap 10-OctDate
Figure 1.1. Total number of cabbage looper and diamondback moth larvae per 400 plants on each sample date.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

67
significant in two treatments. Significantly higher numbers of assassin bugs (Triatoma sanguisuga [LeConte]) were present in the plots receiving the full IPM treatment than in plots receiving the KY Cross plus pyrethroid treatment. Also, the full IPM treated plots had significantly fewer ants (Solenopsis spp.) than the plots receiving the Tropicana plus companion crop treatment. No parasitoids emerged from field-collected DBM larvae reared in the laboratory. The effects of the different treatments on damage-free yield and yield loss per plant are shown in Tables 1.3 and 1.4, respectively. When all treatments were compared with the full IPM treatment, yield loss was significantly higher only in the KY Cross plus pyrethroid treatment. Regarding damage- free yield, that in the KY Cross plus pyrethroid treated plots was significantly lower than in the full IPM treated plots. The Tropicana plus Agree 50 WP® treatment resulted in significantly higher damage-free yield than the full IPM treatment, however the full IPM treatment had significantly higher damage-free yield than the Tropicana plus companion crop treatment.
A total of 165.5 kg of cabbage was harvested. This had a market value of $J5454.90 ($J33/kg) (Table 1.5) . The value of damage-free cabbage harvested was significantly higher for the Tropicana plus Agree 50 WP® treatment than for all
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 1.3. Mean + SEM damage-free yield of cabbage (kg) per plant from all treatments compared with the full IPM treatment (Tropicana, companion crop, and B.thuringiensis)Treat me n t c omparlsons MeanTsEM3 Significance^Tropicana, companion crop - IPM 0.61 + 0".b9 - 1.00 + 0.00 S
KY Cross, companion crop, & Bt. - IPM 0 . 9 3 +0.03-1.00+0.00 NS
Tropicana & Bt. - IPM 1.27 +0.03-1.00+0.00 S
KY Cross & pyrethroid - IPM 0.33+0.05-1.00+0.00 S
aSEM of zero due to rounding off; bDunnett's T = 2.807, df = 12, a = 0.05; S, significant; NS, not significant; Bt., Bacillus thuringiensis.
o\oo

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 1.4. Mean + SEM yield loss of cabbage (kg) per plant from all treatments compared with the full IPM treatment (Tropicana, companion crop, and B. thuringiensis)
Treatment comparisons Mean + SEMa Significance0Tropicana, iiorripanionc IPM......... 0.28'T'"or04 r'''o7oy'’’£'"oyoo............ NS....
KY Cross, companion crop, & Bt. - IPM 0 . 1 3+0.03-0.07+0.00 NS
Tropicana & Bt. - IPM 0. 0 8 +0.01-0.07+0.00 NS
KY Cross & pyrethroid - IPM 0. 80+0.12-0.07+0.00 S
aSEM of zero due to rounding off; bDunnett's T = 2.807, df = 12, a = 0.05; S, significant; NS, not significant; Bt., Bacillus thuringiensis.
o>w>

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 1.5. Mean + SEM damage-free yield and value of cabbage from all treatments
Treatments Marketable yield (kg) Value (J$33/kg)Tropicana, companion crop & Bt. 40.10 + 0.91b 1323.30 + 6.32b
Tropicana & companion crop 24.40 + 3.68c 805.20 + 121.46c
Tropicana & Bt. 50.60 + 1.33a 1669.80 + 43.94a
KY Cross, companion crop, & Bt. 37.10 + 1.22b 1224.30 + 40.10b
KY Cross & pyrethroid 13.10 + 2.19d 432.30 + 72.37d
Bt., Bacillus thuringiensis; Means within significantly different (Tukey1s Test, a :
a column followed by the = 0.05).
same letter are not
o

71
treatments. Comparing the full IPM and the KY Cross, companion crop plus Agree 50 WP® treatments, the values of damage-free cabbage were not significantly different. In addition, for the Tropicana plus companion crop treatment, the value of damage-free cabbage was significantly lower than that for all treatments except the KY Cross plus pyrethroid treatment, which had the lowest value. For tomatoes, total yield was 298 kg, i.e., an average yield of 24.83 kg per plot, the value of which was $J1092.50 ($J44/kg) per plot. The combined values of cabbage and tomato, for treatments which had both, are shown in Table 1.6.
DiscussionIntegrated Pest Management approaches for controlling
lepidopterous pests of crucifers have gained credibility in recent times. In this study, components of an IPM package had varying effects on CL and DBM larval infestation of cabbage, cabbage yield, and in protecting beneficial arthropods. For example, the number of CL larvae per plant was significantly higher in the Tropicana plus companion crop treatment when compared with the full IPM package. This may be attributed to the lack of any insecticide application in the former treatment. Furthermore, the number of DBM larvae per plant was significantly higher in the KY Cross plus pyrethroid treatment compared with the full IPM package.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 1.6. Damage-free yield and value of cabbage3 and tomatob from all treatments
Crop
Treatments
Cabbage
(kg)
Tomato
(kg)
Cabbage
(J$)
Tomato
(J$>
Total Value
(J$)Tropicana, companion crop & Bt. 40.10 24.83 1323.30 1092.50 2415.80
Tropicana & companion crop 24.40 24.83 805.20 1092.50 1897.70
Tropicana & Bt. 50.60 --- 1669.80 --- 1669.80
KY Cross, companion crop, & Bt. 37.10 24.83 1224.30 1092.50 2316.80
KY Cross & pyrethroid 13.10 --- 432.30 --- 432.30
• “value calculated at J$33/kg; bvailue calculated at J$44/kg.
-4M

73
This showed that the full IPM treatment was superior in reducing DBM population density compared with the current practice of treating the KY Cross cabbage variety with pyrethroids. In fact, the number of DBM larvae per plant in plots receiving the KY Cross plus pyrethroid treatment was 106 times that in plots receiving the full IPM treatment (Table 1.1). Resistance to the pyrethroid insecticide used (Karate 2.5 EC®) cannot be discounted because in Jamaica and other Caribbean islands, the DBM has several-fold resistance to most available insecticides (Suah & Ellis 1974, Robinson et al. 1995). Predators, though not statistically significant across all treatments, were absent or in very low numbers in the KY Cross plus pyrethroid treatment compared with the full IPM package. This was not unexpected because pyrethroids are non-selective insecticides. The lack of parasitism of DBM larvae may be attributed to the relatively short duration of the study which did not allow for the build-up of parasitoid populations. Diadegma insulare (Cresson) , Cotesia plutellae Kurdjumov, and other parasitoids are present in Douglas Castle and Castle Kelly, areas where cabbage has been grown continuously in commercial quantities since the 1970s, but their effect is being compromised because growers use excessively broad-spectrum insecticides such as carbamates, organophosphates, and pyrethroids.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

74
Between 20-22 applications of these insecticides over a growing season are not uncommon (Alam 1974, 1992). AtDouglas Castle and Castle Kelly B. thuringiensis would be ideally suited for control of DBM and conservation of predators and parasitoids.
The Tropicana plus Agree 50 WP18 treated plots had the highest yield of damage-free cabbage and the KY Cross plus pyrethroid treated plots had the lowest. The yield advantage of the Tropicana variety plus Agree 50 WP® treated plots may be attributed to the low numbers of DBM larvae per plant (0.24) that were present in these plots. Relatedly, yield loss was highest in the KY Cross plus pyrethroid treated plots. This was undoubtedly due to the very high numbers of DBM larvae per plant (16.9) that were present in these plots. Even though the number of CL larvae per plant in plots receiving the different treatments was relatively higher than the number of DBM larvae, the peak of the CL population occurred during the preheading stage of the cabbage plants. In addition, most of the CL larvae encountered were early instars and so the damage inflicted by them had less of an impact on yield than was the case with DBM whose population peaked when cabbage plants were in the heading stage (Figure 1.1). The effectiveness of Agree 50 WP® on damage-free cabbage yield was especially evident with the Tropicana
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

75
variety of cabbage. Agree 50 WP® contains both the kurstaki and aizawai strains of B . thuringiensis. The insecticidal properties of different subspp. of B. thuringiensis vary with the crystal proteins toxins which they produce (Hofte & Whiteley 1989) . The efficacy of Agree 50 WP® may have resulted from the combined toxicity of the two B. thuringiensis strains that it contained. The qualities of B. thuringiensis, especially its specificity to a small subset of insects and nontoxicity to beneficial organisms, make it suitable for use in IPM programs.
Regarding possible effects of intercropping on insect pest populations, it has been reported that companion crops, in addition to creating diversity by attracting and maintaining beneficial insects (Pimmental 1961, Cromartie 1981, Risch et al. 1983, Bach & Tabashnik 1990, Mitchell et al. 1997, Riggin-Bucci & Gold 1997), also produce powerful chemical deterrents to many pest species, or may mask the chemical cues of crops that attract pests (Zimmerman 1978, Altieri & Liebman 1985, McClure 1994a, 1994b). Thecharacteristic odor of tomato plants - the companion crop used in this study - is due to the presence of a-tomatine, a chemical that has been reported to have repellent effects on ovipositing DBMS (Burunday & Raros 1973, Talekar et al. 1986). Also, in a review of visual detection of plants by
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

76
herbivorous insects, Prokopy & Owens (1983) alluded to the utilization of visual stimuli in pest management to disrupt the host detection process. They suggested intermixing of host plants or planting host patches in particular spatial arrays - approaches not dissimilar to intercropping. In addition, Zimmerman (1978) drew comparisons between companion planting for pest management in agroecosystems and the interplanting observable in natural ecosystems. This idea ties in elegantly with the "plant apparency hypothesis" advanced by Price (1984), whereby in natural habitats the diversity of plants present makes each individual species "non-apparent." By contrast, in agricultural situations where large tracts of land are planted to a single crop (cabbage, for example), plants become "apparent" and therefore are "bound to be found" by herbivores. The role of tomato as a companion crop in controlling CL and DBM was inconclusive in this study and the author proposes that additional studies will be needed to determine if companion planting indeed works in reducing insect populations. However, an economic justification for planting the companion crop was demonstrated. The main additional inputs required for growing the companion crop were seeds, wooden stakes for supporting the plants, and increased labor. All the other inputs such as land preparation, fertilizer, insecticides,
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

77
etc. are done or applied simultaneously for both crops. Thatthe companion crop had a gross value of more than twice thatof the main crop is important. This is so because manyfarmers practice mixed cropping and the higher value mayserve as an incentive for them to choose to grow tomatoesalongside their lower value cabbage crop. The lower yield ofcabbage that occurred when it was grown with the companioncrop was offset by the higher value of the tomatoes.
The overall results of this study indicated that IPM ofCL and DBM may not necessarily require that all thecomponents investigated be used simultaneously. For example,it was shown that the new cabbage hybrid, Tropicana, whentreated with Agree 50 WP® gave the highest yield of damage-free cabbage compared with all the other treatments.
References Cited - Chapter OneAlam, M. M. 1974. Biological control of insect pests of horticultural crops in Barbados, pp. 253-261. In C. W. D Brathwaite, R. H. Phelps & F. D. Bennett (eds.), Proceedings of Protection of Horticultural Crops in the Caribbean. University of the West Indies, St. Augustine, Trinidad.Alam, M. M. 1989. Vegetable pests and their natural enemies in Barbados, pp. 295-302. In Proceedings of the Caribbean Food Crops Society.Alam, M. M. 1992. Diamondback moth and its natural enemies in Jamaica and some other Caribbean Islands, pp. 233-243. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Altiari, M. A. £ M. Liebman. 1985. Insect, weed, and plant disease management in multiple cropping systems, pp. 183-218.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

78
In C. A. Francis (ed.), Multiple Cropping Systems. Macmillan Publishing Company, New York.B&ch, C. E. £ B. E. Tabashnik. 1990. Effects of nonhost plant neighbors on population densities and parasitism rates of the diamondback moth (Lepidoptera: Plutellidae) . J. Econ. Entomol. 19: 987-994.Bennatt, F. D. 6 M. Yaaeem. 1972. Parasite introductions for the biological control of three insect pests in the lesser Antilles and British Honduras. PANS. 18: 468-474.Buranday, R. p. & R. s. Raros. 1973. Effects of cabbagetomato intercropping on the incidence and oviposition of the diamondback moth, Plutella xylostella (L.). Philipp. Entomol. 2: 369-374.Cromartie, W. J. 1981. The environmental control of insects using crop diversity, pp. 223-251. In D. Pimentel (ed.) , CRC Handbook of Pest Management in Agriculture. CRC Press, Boca Raton, FL.Dickson, M. H. & C. J. Eckenrode. 1975. Variation in Brassica oleracea resistance to cabbage looper and imported cabbage worm in the greenhouse and fields. J. Econ. Entomol. 68: 757-760.Dickson, M. H. 6 C. J. Eckenrode. 1980. Breeding for resistance in cabbage and cauliflower to cabbage looper, imported cabbage worm and diamondback moth. J. Am. Soc. Hort. Sci. 105: 782-785.Dickson, M. H., C. J. Eckenrode fi A. E. Blamble. 1984. NYIR 9602, NYIR 9605, and NYIR 8329, lepidopterous pest resistant cabbages. HortScience. 19: 311-312.Eckenrode, c. J., A. M. Shelton fi 6*. Lin. 1986. Resistance in crucifers to diamondback moth and other Lepidopterous pests, pp. 129-136. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Eigenbrode, s. D., k . a . Stoner, A. M. Shelton 6 w. c. Kain. 1991. Characteristics of glossy leaf waxes associated with resistance to diamondback moth (Lepidoptera: Plutellidae) in Brassica oleracea. J. Econ. Entomol. 84: 1609-1618.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

79
Eigenbrode, 8. D. 6 A. M. Shelton. 1992. Resistance to diamondback moth in Brassica: Mechanisms and potential for resistant cultivars, pp. 65-74. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Harcourt, D. G. 1963. Major mortality factors in the population dynamics of the diamondback moth, Plutella maculipennis (Curt.) (Lepidoptera: Plutellidae). Mem. Can. Entomol. Soc. 32: 55-66.Harcourt, D. G. 1986. Population dynamics of the diamondback moth in southern Ontario, pp. 3-16. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Ta iwan.Hofte, H. 6 H. R. Whitalay. 1989. Insecticidal crystal proteins of B. thuringiensis. Microbiol. Rev. 53: 242-255.Jones, M. T. 1985. Pest management and the vegetable farmer in Trinidad and Tobago, pp. 173-175. In Proceedings of the Caribbean Food Crops Society.Lim, G. S., A. Sivapragasam 6 M. Ruvaida. 1986. Impact assessment of Apanteles plutellae on diamondback moth using an insecticide-check method, pp. 195-205. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.McClure, S. 1994a. Companion planting for pest control, pp. 22-33. In S. McClure & S. Roth (eds.), Rodale's Successful Organic Gardening: Companion Planting. Rodale Press, Emmaus, PA.McClure, S. 1994b. Introduction and companion planting basics, pp. 9-21. In S. McClure & S. Roth (eds.), Rodale's Successful Organic Gardening: Companion Planting. Rodale Press, Emmaus, PA.Mitchell, E. R., G. Y. Hu 6 J. S. Okine. 1997. Diamondback moth (Lepidoptera: Plutellidae) infestation and parasistism by Diadegma insulare (Hymenoptera: Ichneumonidae) in collards and adjacent cabbage fields. Florida Entomol. 80: 54-62.Pimentel, D. 1961. Species diversity and insect population outbreaks. Prot. Ecol. 2: 77-114.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

80
Pollard# 6. V. 1991. Constraints to IPM development and a strategy for management of tomato and cabbage pests in Trinidad, West Indies. Tropical Pest Management 37: 59-62.Price# P. W. 1984. Insect Ecology. John Wiley & Sons, NY.Prokopy# R. J. fiB. D. Owens. 1983. visual detection of plants by herbivorous insects. Annu. Rev. Entomol. 28: 337- 364.Riggin-Bucci, T. 8 P. Gold. 1997. Impact of intraplot mixtures of toxic and nontoxic plants on population dynamics of diamondback moth (Lepidoptera: Plutellidae) and itsnatural enemies. J. Econ. Entomol. 90: 241-251.Risch# S. J.# D. Adov & M. A. Altieri. 1983. Agroecosystem diversity and pest control: Data, tentative conclusions and new research directions. Environ. Entomol. 12: 625-629.Robinson# D. £.# K. M. Da lip 8 A. Mansingh. 1995.Integrated Management of Pests and Pesticides in theCaribbean. Department of Zoology, University of the West Indies, Mona,Kingston. The Jamaica National Commission for UNESCO. Kingston.8alinas# P. J. 1986. Studies on diamondback moth inVenezuela with reference to other Latin American countries, pp. 17-24. In N. S. Talekar & T. D. Griggs (eds.),Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.SAS Institute. 1989. SAS user's guide: Statistics, 5th ed. SAS Institute, Cary, NC.Shelton# A. M.# c. W. Hoy, R. C. North# M. H. Dickson 8 J. Barnard. 1988. Analysis of resistance in cabbage varieties to damage by Lepidoptera and Thysanoptera. J. Econ. Entomol. 81: 634-640.Shelton# A. M., J. L. Robertson# J. D. Tang# C. Perez# s. D. Eigenbrode# H. K. Preisler# W. T. wilsey 8 R. J. Cooley. 1993. Resistance of diamondback moth (Lepidoptera: Plutellidae) to B. thuringiensis subspecies in the field. J. Econ. Entomol. 86: 697-705.Sivapragasam, A.# Y. Ito 8 T. saito. 1988. Population fluctuations of the diamondback moth, Plutella xylostella (L.) on cabbages in Bacillus thuringiensis sprayed and
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

81
nonsprayed plots and factors affecting within-generation survival of immatures. Res. Popul. Ecol. 30: 329-342.Srinvasan, K. & P. M. K Moor thy. 1991. Indian mustard as a trap crop for management of major lepidopterous pests on cabbage. Tropical Pest Management 37: 26-32.Suah, J. R. & D. W. Ellis. 1974. Insecticide control of cabbage caterpillars in Jamaica, pp. 263-266. In C. w. D Brathwaite, R. H. Phelps 6 F. 0. Bennett (eds), Proceedings of Protection of Horticultural Crops in the Caribbean. University of the West Indies, St. Augustine, Trinidad.Talekar, N. S., T. S. Lee £ 8. W. Huang. 1986.Intercropping and modification of irrigation method for the control of diamondback moth, pp. 145-155. Jn N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.USDA. 1945. United States standards for grades of cabbage.18 Federal Register 7084 (1953), redesignated 46 FR 63203 (1981). United States Department of Agriculture.Beltsville, MD.Wagge, J. £ A. Cherry. 1992. Quantifying the impact of parasitoids on diamondback moth, pp. 245-253. Jn N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Zimmerman, L. 1978. Strange bedfellows: The voodoo and science of companion planting, pp. 73-77. In The Harrowsmith Reader. Camden House Publishing Ltd., Canada.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Chapter Two
Potential of Plant Resistance and Bacillus thuringiensis for Control of Diamondback Moth in Jamaica.
82
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

83
IntroductionCabbage, Brassica oleracea var capitata L., is the most
widely grown cruciferous vegetable in Jamaica (C. W. Chambers, Ministry of Agriculture, personal communication). In the 1970s, high-yielding, tropically adapted cabbage hybrids were introduced, and from 1973 to 1986 cabbage production more than doubled, moving from 6,985 to 15,150 tons. Cabbage is grown intensively in some areas such as Douglas Castle and surrounding areas, and is attacked by a complex of lepidopteran insect pests - armyworms, Spodoptera spp., cabbage looper, Trichoplusia ni (Hubner) , cabbage white butterfly, Pieris rapae L., and diamondback moth (DBM), Plutella xylostella (L.). Control of these insects is a prerequisite for the production of cabbage which meet quality standards for damage- and pest-free produce. Under Jamaican conditions, DBM completes its development in about 17 days, giving rise to over 20 generations per year. This results in populations of the DBM frequently accounting for greater than 75% of the total insect pest population attacking cabbage and cause crop loss of up to 90%, making it the key insect pest from an economic standpoint (Alam 1992).
Climatic conditions in Jamaica are favorable to cabbage production throughout the year, a situation which in turn is conducive to continuous breeding of DBM. Historically,
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

84
farmers have relied primarily on multiple applications of broad-spectrum insecticides for control of this pest. Between 20-22 insecticide applications over a growing season are not uncommon. Because of this reliance on conventional insecticides, many from the organochlorine, organophosphate, carbamate, and pyrethroid groups are now ineffective (Alam 1974, Suah & Ellis 1974, Robinson et al. 1995). Reports of low efficacy of Biotrol® and Thuricide® - microbial insecticides containing the kurstaki strain of Bacillus thuringiensis Berliner - in controlling DBM, presumably due to insecticide resistance, led Alam (1992) to question their reliability. Additionally, high insecticide use has led to excessive residues on produce. In fact, a survey conducted in 1988 found that 54% of farmers in Douglas Castle and surrounding districts have used insecticides rates above the recommended rates, sometimes up to twice the recommended rate in their attempt to obtain effective control of DBM (Walton 1989) . Dueszeln (1995) reported that vegetables and fruits consumed locally or exported often have pesticide residues above tolerable levels.
Because of pest management problems, environmental degradation, and public health risks associated with conventional insecticides, it is imperative to find an integrated pest management (IPM) approach for DBM management
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

85
which utilizes tactics such as host plant resistance and microbial control.
The utility of host plant resistance as an insect pest management tactic is well established (Painter 1951). The potential for using resistant varieties of crucifers for DBH management has been investigated. Dickson et al. (1984, 1986) reported the release of four cabbage breeding lines possessing resistance to DBM. Results of genetic and other studies indicated that the resistance to DBM exhibited by these cabbage breeding lines was associated with the glossy dark-green leaf from an introduced cauliflower breeding line, Plant Introduction (PI) 234599 (Dickson et al. 1990, Stoner 1990, Eigenbrode & Shelton 1992). However, DBM-resistant cabbage varieties are not generally commercially available.
Use of microbial agents for controlling DBM has had the greatest success with insecticides based on toxins of the soil bacterium, B. thuringiensis. The kurstaki and aizawai strains of this bacterium produce a characteristic set of insecticidal crystalline proteins which are highly effective against early instars of DBM (Lim 1992). Furthermore, B. thuringiensis is environmentally benign, harmless to humans, and nontoxic to beneficial organisms (Hofte & Whiteley 1989), many of which are natural enemies of DBM. Because thekurstaki strain of B. thuringiensis is already of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

86
questionable reliability in Jamaica, and field resistance in DBM against B. thuringiensis subsp. kurstakl has been reported elsewhere (Tabashnik et al. 1990, Shelton et al. 1993, Perez & Shelton 1997), it is unwise for farmers to rely solely on this microbial insecticide for controlling DBM. The usefulness of B. thuringiensis is dependent on how long insect pests remain susceptible to its crystal protein toxins (McGaughey & Whalon 1992) . To thwart the development of insecticide resistance and preserve the longevity and effectiveness of B. thuringiensis against DBM, a more integrated approach for controlling this insect should be developed. Combining the use of foliar applications of B. thuringiensis and host plant resistance for DBM control is plausible. A new product, Agree 50 WP® (wettable powder), containing both the kurstaki and aizawai strains of B. thuringiensis and a new cabbage variety, Tropicana, reputed resistant to DBM, has recently become available to farmers. Before the advent of this new cabbage variety, two hybrids (KY and KK Cross) and an open-pollinated variety (Early Jersey) were available to growers in Jamaica. In 1995, marketing of Tropicana began by the leading distributor of agricultural inputs with claims of its ability to resist attack by DBM. However, the relative resistance of these cabbage varieties to DBM, under local conditions, has not
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

87
been empirically studied. Therefore, toward achieving the long range goal of IPM of DBM, the objective of this study was to determine the efficacy of Agree 50 WP® and evaluate the relative susceptibility of cabbage varieties grown in Jamaica against DBM.
Materials and Methods In September 1995, a field experiment was initiated on
the crop farm of the College of Agriculture, Science and, Education, Port Antonio, Portland, Jamaica. Three cabbage varieties, Tropicana, Early Jersey (Petoseed, Saticoy, CA) , and KY Cross (Takii Seed, Kyoto, Japan), were subjected to two different treatment regimes. These were Agree 50 WP® [3.8% (Al) (25,000 IU per ml) (B. thuringiensis subspecies kurstaki and aizawai); Novartis, Greensboro, NC] applied weekly and untreated controls. A split-plot experimental design with a 3 x 2 factorial treatment arrangement was used to partition cabbage variety and B. thuringiensis effects. Main plots were variety and subplots were Agree 50 WP®- treated and untreated controls. Each treatment was replicated four times. Individual plots were 2.73 m x 4.45 m and were separated by a distance of 1.52 m. Four-week-old seedlings of the three cabbage varieties, raised in outdoor seedbeds, were planted 0.45 m apart on raised beds spaced 0.91 m apart. Standard cultivation practices for cabbage
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

88
production were employed. Plots were sampled for DBM once per week for seven weeks between 18 November and 30 December by counting larvae on the leaves of 10 plants in each subplot. The microbial insecticide, Agree 50 WP® was applied to the appropriate plots once per week, at the recommended rate of 83 g in 15 liters of water using a Solo 475 Knapsack Sprayer (Solo Incorporated, Newport News, VA).
The data were subjected to analysis of variance (ANOVA) using PROC GLM of the SAS system (SAS Institute 1989) , and, where significant differences were found among cabbage varieties, the Waller-Duncan K-ratio t test was used for means separation (SAS Institute 1989).
ResultsDiamondback moth larval population density per plant was
significantly affected by cabbage variety (F - 20.94; df = 2; P = 0.0001) and insecticide treatment (F = 52; df = 1; P = 0.0001) . The ranking of cabbage varieties from most to least susceptible to DBM was KY Cross > Early Jersey > Tropicana. Plots treated weekly with Agree 50 WP® had significantly (F = 52; df = 1; P = 0.0001) fewer DBM larvae per plant compared with untreated plots (Table 2.1). The interaction between cabbage variety and insecticide treatment was marginally significant (F = 3.38; df = 2, P = 0.0567). Furtherexamination of this interaction was done using PROC PLOT of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

89
the SAS system (SAS Institute 1989). The cell means for the levels of the two factors, insecticide treatment and cabbage variety, as shown in Table 2.1, indicated that the Tropicana variety supported fewer larvae per plant across Agree 50 WP®- treated and untreated plots compared with the Early Jersey and KY Cross varieties, over the sampling period, the total number of DBM larvae remained relatively high in untreated plots but fell sharply in Agree 50 WP®-treated plots and was low at plant maturity (Figure 2.1).
DiscussionBased on larval population density estimates, it was
concluded that the Tropicana variety was superior to the other two varieties - KY Cross and Early Jersey - in resisting attack from DBM larvae. The author is not aware of any previous study in which cabbage varieties available to growers in Jamaica were evaluated under local conditions regarding their relative resistance to DBM. The existence of resistance to DBM in a commercially cultivated cabbage variety, such as Tropicana, is significant because of the economic importance of this insect pest stemming form the intractable problem of its widespread development of resistance to conventional insecticides. Several authors have reported on breeding, and the potential for using resistant cabbage varieties for DBM management (Dickson et
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 2.1. Mean numbers + SEM of diamondback moth larvae per plant on three cabbage varieties treated with Agree 50 WP®.
No. larvae per plantOverall variety means
+ sem2Variety Agree 50 WP® Untreated
Tropicana 0.30 + 0.21 1.55 + 0.39 0.93 + 0.31
Early Jersey 0.88 + 0.25 4.23 + 0.33 2.55 + 0.66
KY Cross 2.23 + 0.70 4.95 + 0.40 3.59 + 0.64Overall treatmentmeans + sem1 1.13 + 0.34 3.58 + 0.48
treatment means significantly different (ANOVA F test; a = 0.05). 2Variety means significantly different (Waller-Duncan K-ratio t test; a = 0.05).
VOo

100
70*I 00-J 00-6 40 •£ 30-
20-
10- ■O-23-Oec 30-Dec
Date
Figure 2.1. Total number of diamondback moth larvae per 240 plants on each sample date.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

92
al. 1984, 1986, 1990; Stoner 1990; Eigenbrode & Shelton,1992) but resistance against DBM in any commercially available cabbage variety in Jamaica has hitherto not been reported.
The ability of the Tropicana cabbage variety to resist DBM could possibly be due to morphological traits. The epidermal layer of its leaves is thicker, especially as plants approach the heading stage, and its heads are firmer and have tightly appressed leaves compared with those of the other two varieties. Because first instars of DBM are leafminers and must tunnel into the mesophyll cells of leaves to feed, the thicker epidermis of Tropicana may have been too great a challenge for the mandibles of neonate larvae. These neonates may starve to death, desiccate, drown or be washed from leaves, and be vulnerable to predators. In a review of plant resistance to insects, Beck (1965) noted that biophysical factors such as the physical form and structure of plant tissues influence their utilizability as suitable insect hosts. Eigenbrode and Shelton (1992) found that neonate DBM larvae had greater movement on resistant cabbage breeding lines than on susceptible ones. Also, Tanton (1962) found that leaf texture affects the number of nibbles and subsequent leaf area of Brassica rapa L. consumed by Phaedon cochleariae P. In addition, Iheagwam (1981) reported that
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

93
penetratiblility of the leaf tissue of Brassica oleracea L. influences the degree of exploitation by Aleyrodes brassicae Walker. And, Martin et al. (1975) found a negative correlation between internode rind hardness of Saccharwn officinarum L. and susceptibility to attack by neonate Diatraea saccharalis (F.) larvae.
Agree 50 WP® proved to be effective in controlling DBM. This was probably due primarily to the presence of the aizawai strain of B. thuringiensis. There are differences in the crystal protein toxin profiles of B. thuringiensis subsp. kurstaki and subsp. aizavai; the former produces Cry lA(a) , Cry lA(b), Cry lA(c) , Cry 11A, and Cry 11B, whereas the latter produces Cry lA(a), Cry lA(b), Cry 1C, Cry ID, and Cry 11D (Hofte & Whiteley 1989; Liu et al 1996). Shelton et al. (1993) found differential responses between DBM populations treated with two formulations containing B . thuringiensis subsp. kurstaki (Javelin WG® and Dipel 2X®) and populations treated with B. thuringiensis subsp. aizavrai (ZenTari®) . Commercial formulations of B. thuringiensis subsp. kurstaki, for example Biotrol® and Thuricide®, have been used in Jamaica for many years to control DBM but their reliability is now questionable (Alam 1992). However, formulations of B. thuringiensis subsp. aiaawai have recently begun to be more widely used. So, DBM populations in Jamaica have received no
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

94
prior exposure to the aizawai strain of B. thuringiensis. In fact, B. thuringiensis subsp. aizawai was first used against DBM in Jamaica in 1995, hence its effectiveness.
The existence of a significant interaction requires that caution be exercised when making statements about the main effects (cabbage variety and insecticide treatments), even though both were statistically significant with P values of 0.0001 (Freund & Wilson 1993). The consistently lower numbers of larvae on the Tropicana variety compared with Early Jersey and KY Cross, across Agree 50 WP®-treated and untreated plots, clearly showed that the Tropicana variety was superior to the other two varieties in resisting DBM. Also, from this interaction between cabbage variety and insecticide treatment, it may be inferred that the Tropicana variety may be compatibly combined with use of Agree 50 WP® for successful management of DBM.
Considering the low efficacy and tenuous reliability of commercial formulations of B. thuringiensis subsp. kurstaki against DBM in Jamaica (Alam 1992), the effectiveness of Agree 50 WP® (B. thuringiensis subspp. kurstaki and aizawai) seen in this study makes continued use of toxins of B. thuringiensis for controlling this insect a viable option, especially when combined with the Tropicana cabbage variety. However, the author recommends that formulations containing
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

95
both the kurstaki and aizawai strains of B . thuringiensis, such as Agree 50 WP®, should not be used extensively as this may allow for the development of resistance in the DBM to the aizawai strain without losing its resistance to the kurstaki strain.
References Cited - Chapter TwoAlan, M. K. 1974. Biological control of insect pests of horticultural crops in Barbados, pp. 253-261. In C. W. D. Brathwaite, R. H. Phelps & F. D. Bennett (eds.), Proceedings of Protection of Horticultural Crops in the Caribbean, university of the West Indies, St. Augustine, Trinidad.Alan, M. M. 1992. Diamondback moth and its natural enemies in Jamaica and some other Caribbean Islands, pp. 233-243. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Beck, S. D. 1965. Resistance of plants to insects. Annu. Rev. Entomol. 10: 207-232.Dickson, M. K., C. J. Eckenrode 6 A. E. Blamble. 1984. NYIR 9602, NYIR 9605, and NYIR 8329, lepidopterous pest resistant cabbages. HortScience 19: 311-312.Dickson, M. H., C. J. Eckenrode 6 J. Lin. 1986. Breeding for diamondback moth resistance in Brassica oleracea, pp. 137-143. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Dickson, M. H., A. M. Shelton, S. D. Eigenbrode, M. L. Vamosy 6 M. Mora. 1990. Breeding for resistance to diamondback moth, Plutella xylostella, in cabbage. HortScience. 25: 1643-1646.Duesslen, J. 1995. Pesticide residues in food, pp. 2-4. In The Agriculturist (April 1995). Caribbean Agricultural Communications Ltd., Kingston, Jamaica.Eigenbrode, S. D. 6 A. K. Shelton. 1992. Resistance to diamondback moth in Brassica: Mechanisms and potential for
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

96
resistant cultivars, pp. 65-74. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Freund, R. J. 6 W. J. Wilson. 1993. Statistical Methods. Academic Press. San Diego, CA.Hofte, K. & H. R. White ley. 1989. Insecticidal crystal proteins of S. thuringiensis. Microbiol. Rev. 53: 242-255.Iheagvam, E. U. 1981. The relationship between weight of insect, age, hardness and nitrogen content of cabbage leaves and fecundity of the cabbage whitefly, Aleyroides brassicae Walker (Homoptera: Aleyroididae) . Z. Ang. Ent. 91: 349-354.Lim, 6. 1992. Integrated pest management of diamondbackmoth: practical realities, pp. 565-576. In N. S Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Liu, Y. B., B. E. Tabashnik ( M. Pusstai-Carey. 1996.Field-evolved resistance to Bacillus thuringiensis Toxin CrylC in diamondback moth (Lepidoptera: Plutellidae) . J.Econ. Entomol. 89: 798-804.Martin, F. A., C. A. Richard 6 8. D. Hensley. 1975. Host resistance to Diatraea saccharalis (F.): Relationship ofsugarcane internode hardness to larval density. Environ. Entomol. 4: 687-688.McGaughey, W. H. £ M. E. Whalon. 1992. Managing insect resistance to Bacillus thuringiensis toxins. Science 258: 1451-1455.Painter, R. H. 1951. Insect Resistance in Crop Plants. Macmillan, New York.Peres, C. J. 6 A. M. Shelton. 1997. Resistance of Plutella xylostella (Lepidoptera: Plutellidae) to Bacillusthuringiensis Berliner in Central America. J. Econ. Entomol. 90: 87-93.Robinson, D. E., K. M. Dalip 6 A. Mansingh. 1995.Integrated Management of Pests and Pesticides in the Caribbean. Department of Zoology, University of the West Indies, Mona, Kingston. The Jamaica National Commission for UNESCO. Kingston.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

97
SAS Institute. 1989. SAS/STAT user's guide: Statistics, 5th ed. SAS Institute, Cary, NC.Shelton, A. M., J. L. Robertson, J. D. Teng, C. Peres, S. D. Eigenbrode, H. K. Preisler, W. T. Wilsey & R. J. Cooley. 1993. Resistance of diamondback moth (Lepidoptera: Plutellidae) to Bacillus thuringiensis subspecies in the field. J. Econ. Entomol. 86: 697-705.Stoner, K. A. 1990. Glossy leaf wax and plant resistance to insects in Brassica oleracea under natural infestation. Environ. Entomol. 19: 730-739.Suah, <J. R. t 0. W. Ellis. 1974. Insecticide control of cabbage caterpillars in Jamaica, pp. 263-266. In C. W. 0 Brathwaite, R. H. Phelps & F. D. Bennett (eds.), Proceedings of Protection of Horticultural Crops in the Caribbean. University of the West Indies, St. Augustine, Trinidad.Tabashnik, B. E., N. L. Cushing, M. Pinson 6 M.W. Johnson. 1990. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae) . J. Econ. Entomol. 83: 1671-1676.Tanton, M. 1962. The effect of leaf "toughness" on the feeding of larvae of the mustard beetle, Phaedon cochleriae F. Ent. Exp. & Appl. 5: 74-78.Walton, W. C. 1989. Problems associated with the control of diamondback moth in Douglas Castle, St. Ann. Department of Crop Science, Faculty of Agriculture, University of the West Indies, St. Augustine, Trinidad.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Chapter Three
Toxicity of Three Formulations of Bacillus thuringiensisAgainst Diamondback Moth.
98
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

99
IntroductionThe diamondback moth (DBM), Plutella xylostella (L.) , is
a ubiquitous pest where cruciferous vegetables are grown. The annual worldwide control costs for this insect has been estimated to be US $1 billion (Talekar & Shelton 1993) . Like many other lepidopterous insect pests, the larval stage is the damaging stage of the DBM. All stages of the insect except the moth in flight can be found on host plants (Harcourt 1958). Third and fourth instars cause the most damage. Rosario and Cruz (1986) reported that an individual fourth instar may consume 0.63 cm2 of cabbage leaf per day and Ivey (1991) found that leaf area consumption rate increased as temperature increased within the range 10-30°c. Damage caused by DBM larvae consists of irregular "windows" in the leaves of host plants which subsequently coalesce and breakdown to form holes.
Approximately 857 acres of mustard, Brassica juncea [L. ] Czern, having a total farmgate value of $2.3 million were produced in Louisiana in 1995 (Picha & Hinson 1996). A complex of three lepidoptera larvae, the DBM, Plutella xylostella (L.), the cabbage looper, Trichoplusia ni (Hubner), and cabbage white butterfly, Pieris rapae (L.), attack mustard greens and other crucifers in the state. Of this insect pest complex, the DBM is the most damaging, and,
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

100
if not properly controlled, may cause serious losses of up to 100% (Calderan & Hare 1986, Magallona 1986). Control of DBM on crucifers in Louisiana has been based primarily on multiple, within-season applications of broad-spectrum insecticides. The Louisiana Cooperative Extension Service recommends 10 conventional insecticides and six formulations of Bacillus thuringiensis Berliner for control of this insect in the state (LCES 1997).
In North America (Shelton et al. 1993a) as veil as in many tropical countries (Georghiou 1986), insecticide- resistant strains of DBM have evolved in response to the selection pressure exerted by heavy insecticide use, resulting in control failures. Wyman (1992) and Shelton & Wyman (1992) reported that DBM resistance against conventional insecticides was endemic in several southern states including Florida, Georgia, North Carolina and Texas. Because of resistance to conventional insecticides attention has been focused on the use of the toxins of B. thuringiensis for controlling DBM and we are now well into our second generation of B. thuringiensis technology. B. thuringiensis is noted for its abundant production during sporulation of insecticidal proteins in the form of a crystal or crystal- complex. The insecticidal crystal proteins are commonly designated as "Cry" proteins and the genes encoding them as
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

101
"cry" genes. The Cry proteins typically undergo both solubilization and activation before becoming biologically active toxins. For most, solubilization takes place in the highly alkaline environment of lepidopteran insect midguts. Activation occurs through proteolysis by insect gut enzymes and may occur at the same time as solubilization. Once activated, the toxins recognize certain receptors on the surface of the insect midgut epithelial cells. A pore- complex forms through the cell membrane, resulting in loss of the insects' ability to regulate osmotic pressure. Ultimately the insect succumbs to massive water uptake (NBIAP 1995). Currently, industry has, at various levels of development, at least three distinct technologies for delivering B. thuringiensis endotoxins - foliar sprays, transgenic plants, and endophytic bacteria (Ely 1993, Kennedy & Whalon 1995). Commercial formulations of foliarly applied toxins of 5. thuringiensis have been used against the DBM since the 1970s. However, reports of resistance in DBM against toxins of B. thuringiensis subsp. kurstaki have appeared in the literature (Kirsh & Schmutterer 1988, Tabashnik et al. 1990, 1993, Syed 1992, Shelton et al. 1993b, Perez & Shelton 1997).
Because concerns are now being raised over the use of B . thuringiensis, with the demonstration of field resistance in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

102
DBM against its toxins and the increasing use of commercial formulations based on this bacterium for control of this insect, the resistance/ susceptibility status of DBM populations in Louisiana to commercial formulations of these microbial insecticides is worthy of investigation. An opportunity to do this presented itself in July 1996. A mustard grower in southern Louisiana experienced less than satisfactory control of DBM in fields treated with Thuricide®, a commercial formulation of B. thuringiensis subsp. kurstaki, and sought the assistance of personnel from the Department of Entomology at Louisiana State University. Larvae from this population were hand-collected, and in laboratory bioassays, their response to other formulations of B. thuringiensis strains was evaluated.
Materials and Methods A laboratory colony was started from DBM larvae hand-
collected on mustard (cv Florida Broadleaf) grown for commerce in a field in St. Francisville, East Baton Rouge Parish, Louisiana in July 1996. In the laboratory, the larvae were placed in groups of between 5-10 on top of a moistened filter paper (90 mm diameter) in petri dishes (100 x 15 mm) . Fresh leaves of the cabbage variety KY Cross (Takii Seed, Kyoto, Japan) collected from an untreated field plot, were provided as needed. At eclosion, adult moths were
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

103
released in screen cages (300 x 300 x 300 mm) containing a mustard seedling for oviposition. A cotton swab moistened with honey/water (a source of carbohydrates for energy) solution was placed in plastic cups in each cage. Eggs were allowed to hatch and once second instars appeared on the mustard seedling the larvae were transferred to petri dishes. The colony was reared for about three generations before bioassays were started.
Three commercial formulations of B. thuringiensis were tested: Agree 50 WP® (wettable powder) [3.8% (Al) (25,000 IU per mg) (B. thuringiensis subspp. kurstaki and aizawai); Novartis, Greensboro, NC], Dipel ES® (emulsifiable suspension) [3.4%(Al) (17,600 IU per mg) (B. thuringiensis subsp. kurstaki] and ZenTari AS® (aqueous suspension)[B. thuringiensis subsp. aizawai] (Abbott Laboratories, North Chicago, IL).
Concentrations ([AI]/L) were based on dilution series using the recommended "field" rates of the three formulations of B. thuringiensis. Dipel and ZenTari were tested at concentrations of 2.5, 0.25, 0.025, 0.0025 and 0.00025[ml(AI)/L); Agree was tested at concentrations of 3.0, 0.3, 0.03, 0.003 and 0.0003 [g(AI)/L]. All insecticides werediluted with distilled water. Tests with each concentration and the untreated control (distilled water) were replicated
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

104
five times. A leaf-dip assay similar to that described by Tabashnik et al. (1993) was used. Cabbage leaves (of the same variety fed to colony insects) were cut into discs of 20 mm diameter. Each disc was dipped into the test solution and hung vertically to dry at room temperature for about one hour. The discs were then placed in petri dishes (100 x 15 mm) on top of a moistened filter paper (90 mm diameter) . A camel's hair brush was used to transfer five third instars from the colony to the leaf disc in each petri dish. The petri dishes were then placed in plastic boxes (100 x 150 x 300 mm) with covers. Water (to maintain humidity) was placed to a depth of about one centimeter in the bottoms of the plastic boxes, above which a piece of cardboard supported by inverted petri dish covers was placed. Twenty five petri dishes were placed in each plastic box on top of the cardboard. The plastic boxes were held at room temperature for 96 h. After this time mortality was assessed by prodding the larvae with a forceps; a larva was considered dead if it did not move when prodded. Overall control mortality was 20%. Data were analyzed with a probit model using PROC PROBIT of the SAS system (SAS Institute 1989).
ResultsSlopes of the concentration-mortality lines were similar
for all three formulations of B. thuringiensis. Agree 50 WP®
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

105
and ZenTari AS® were equally toxic to DBM based on their LCS0.s and LCgo-s (95% confidence limits overlapped) . However, Dipel ES® appeared to be much less toxic to DBM compared with the other two formulations of B. thuringiensis (Table 3.1).
DiscussionThe results indicated that the DBM population tested was
susceptible to the three formulations of B. thuringiensis. This is relevant and important to cole crop producers in Louisiana as it is both an opportunity and a challenge. An opportunity for them to effectively control DBM and other insect pests of crucifers using insecticides based on toxins of B. thuringiensis, and a challenge to judiciously use these insecticides to prevent their loss to the development of resistance. At present, commercial formulations of insecticides based on toxins of the aizawai strain of B. thuringiensis are not widely used in Louisiana (J. Boudreaux & D. Pollet, Louisiana Cooperative Extension Service, personal communication).
Therefore, populations of DBM in the state had little exposure to this strain of B. thuringiensis. The susceptibility of DBM to toxins of the aizawai strain of B. thuringiensis means that microbial insecticides such as Agree 50 WP® and ZenTari AS® represent additional pest control options that are available to crucifer growers in Louisiana.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission
Table 3.1. Dose-mortality response of a population of diamondback moth to three formulations of B. thuringiensis
Parameter
Insecticide Slope + SE LCso (95% Cl) LC95 (95% CL)
Agree 50 WP® 0.39 + 0.16 1.53 x 10"6 (5.06 x 10'13 - 6.9 x 10'5) 0.03 (0.003 - 78.68)g [Al]/L g [AI]/L
ZenTari AS® 0.53 + 0.18 1.54 x 10-5 (7.28 x 10'10 - 1.53 x 10“4) 0.02 (0.004 - 3.03)ml [Al]/L ml [AI]/L
Dipel ES®““S ^ r o .. i j
0.48 + 0.21 0.01a ml [AI]/L 39.86a ml [AI]/L
s95%Cl"'could"'not be"'estimated""'because"of"̂ poor fIF of the probit r^
Hoo>

107
The usage pattern of B . thuringiensis in the field from which the DBM larvae tested were collected was alarming, and is strongly discouraged. On this farm, Thuricide®, a commercial formulation of B . thuringiensis subsp. kurstaki, was usually tank-mixed with pyrethroids and applied with ground equipment weekly or even as often as two-day intervals. In addition, mustard (a preferred host for DBM oviposition) (Srinivasan & Moorthy 1991) was grown continuously without any host-free period. These conditions were conducive to continuous generations and high populations of DBM. Initial and subsequent cases of resistance against B. thuringiensis in DBM were caused by repeated foliar sprays of commercial formulations of B. thuringiensis subsp. kurstaki. Tabashnik et al. (1990) reported that DBM collected from commercial watercress, cabbage, and broccoli fields in Hawaii that had been subjected to frequent applications of B. thuringiensis subsp. kurstaki had LC5Qs and LCgSs that were 25 and 33 times, respectively, greater than those of susceptible laboratory colonies. Also, Shelton et al (1993b) found that resistance against B. thuringiensis subsp. kurstaki in DBM occurred in Florida under circumstances where it was applied as a tank-mix with pyrethroids, organophosphates, or carbamates for six years. In addition, Shelton et al. (1993b) used a leaf-dip bioassay
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

108
and concluded that field populations of DBM exhibiting LCs0s of B. thuringiensis subsp. kurstaki >0.05 mg (Al) /liter of water were less susceptible in field studies and proposed this as a discriminating dose for control failures. Determination of discriminating doses of insecticides is of paramount importance in resistance monitoring programs and are also extremely desirable in correlating field control failures with the development of resistance (Graves et al.1993) .
Although the DBM population tested exhibited susceptibility to all three formulations of B. thuringiensis, the reduced toxicity of Dipel®, compared with the other two formulations, may be attributed to the past exposure of the population to Thuricide®. Perhaps the population has developed tolerance to B. thuringiensis subsp. kurstaki, the harbinger of future development of resistance. But it is possible too that the failure to control DBM by the mustard grower in St. Francisville was due to unfavorable environmental conditions, poor insecticide coverage, or other factors. Bacillus thuringiensis toxins must be ingested by insects to be effective. The feeding activity of insects is influenced by environmental factors such as relative humidity, light, temperature, and food quality. If not enough toxin is ingested larvae may recover from the gut
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

109
lesions and start feeding again in 48-72 hours (Luthy et al. 1982). In addition, DBM feeds on the undersides of leaves (Harcourt 1958) and so it is important that the nozzles of ground sprayers are adjusted to ensure that the undersides of leaves are covered with insecticides.
The durability of insecticides containing toxins of B. thuringiensis is being threatened by the frequency with which these products are applied over a growing season to produce damage-free produce as demanded by current cosmetic standards for crucifers destined for the fresh market. History has shown that reliance on a single component strategy for DBM control is futile (Talekar & Shelton 1993). However, there is an apparent extreme localization of resistance in DBM against B. thuringiensis, a situation that gives hope for resistance management (Waage 1992). Therefore, a more rational approach to DBM management in Louisiana that utilizes insecticides based on toxins of B. thuringiensis as only one tactic among several tactics is warranted.
References Cited - Chapter ThreeCalderan, J. I. t C. J. Hare. 1986. Control of diamondback moth in Southeast Asia by profenofos, pp. 289-295. In N. s. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Ely, S. 1993. The engineering of plants to express Bacillus thuringiensis 5-endotoxins, pp. 105-124. In P. F. Entwistle, J. S. Cory, M. J. Bailey & S. Higgs (eds.), Bacillus
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

110
thuringiensis. An Environmental Biopesticide: Theory andPractice. John Wiley £ Sons Ltd., NY.Georghiou, 8. P. 1986. The magnitude of the resistance problem, In Pesticide Resistance: Strategies and Tactics for Management. National Research Council. Academic Press. Washington, D.C.Graves, J. B., J. A. Ottea, B. R. Leonard, E. Burris £ 8. Micinski. 1993. Insecticide resistance management: Anintegral part of IPM in cotton, pp. 3-5. In J. Tarver & J. Lord (eds.), Louisiana Agriculture 36, Winter 1993. Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station, Baton Rouge, LA.Harcourt, D. 6. 1958. Biology of the diamondback moth ineastern Ontario. II. Life history, behavior and host relationships. Can. Entomol. 93: 820-831.Ivey, P. W. 1991. Diamondback moth feeding dynamics on cabbage and collards. M.S. Thesis, Louisiana State University, Baton Rouge.Kennedy, 6. 6. £ M. A. Whalon. 1995. Managing pestresistance to Bacillus thuringiensis endotoxins: Constraints and incentives to implementation. J. Econ. Entomol. 88: 454- 460.Kirsch, K. £ J. Schautterer. 1988. Low efficacy of a Bacillus thuringiensis (Ber.) formulation in controlling the diamondback moth, Plutella xylostella (L.), in thePhilippines. J. Appl. Entomol. 105: 249-255.LCES. 1997. Insect Control Guide 1997. Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station. Baton Rouge, LA.Luthy, P., J. Cordier £ H. Fisher. 1982. Bacillus thuringiensis as a bacterial insecticide: Basicconsiderations and application, pp. 35-208. In E. Kurstak (ed.), Microbial and Viral Pesticides. Marcel Dekker, Inc., New York.Magallona, E. D. 1986. Developments in diamondbackmanagement in the Philippines, pp. 423-435. In N. S. Talekar& T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Ta iwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Ill
NBIAP. 1995. Special issue on Bt. National Biological Impact Assessment Program, News Report, December 1995.Peres, C. J. 5 A. K. Shelton. 1997. Resistance of Plutella xylostella (Lepidoptera: Plutellidae) to Bacillusthuringiensis Berliner in Central America. J. Econ. Entomol. 90: 87-93.Picha, D. H. 5 R. A. Hinson. 1996. The commercial vegetable industry in Louisiana, pp. 6-8. In J. Tarver & J. Lord (Eds.), Louisiana Agriculture. Vol. 39, No. 3.Rosario, C. 6 C. Crus. 1986. Life cycle of diamondback moth, Plutella xylostella (L.), (Lepidoptera: Plutellidae) in Puerto Rico. J. Agric. of the University of Puerto Rico. 70: 229-234.SAS Institute. 1989. SAS/STAT user's guide: statistics, 5ch ed. SAS Institute, Cary, NC.Shelton, A. H. 6 J. A. Wyman. 1992. Insecticide resistance of diamondback moth (Lepidoptera: Plutellidae) in NorthAmerica, pp. 447- 454. In N. S. Talekar (ed.) Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Shelton, A. M., J. A. Wyman, N. L. Cushing, K. Apfelbeck, T.J Dennehy, s. E. R. Mahr 6 8. D. Eigenbrode. 1993a.Insecticide resistance of diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae) , in North America. J. Econ. Entomol. 86: 11-19.Shelton, A. M., J. L. Robertson, J. D. Tang, c. Perez, s. D.Eigenbrode, K. K. Preisler, w. T. Wilsey 6 R. J. Cooley.1993b. Resistance of diamondback moth (Lepidoptera: Plutellidae) to Bacillus thuringiensis subspecies in the field. J. Econ. Entomol. 86: 697-705.Srinivasan, K. 6 P. N. K. Moor thy. 1991. Indian mustard as a trap crop for management of major lepidopterous pests on cabbage. Tropical Pest Management. 37: 26-32.Syed, A. R. 1992. Insecticide resistance in diamondback moth in Malaysia, pp. 437-442. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

112
Tabashnik, B. E., N. L. Cushing, N. Finson £ M. w. Johnson. 1990. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae) . J. Econ. Entomol. 83: 1671-1676.Tabashnik, B. E., N. Finson , C. F. Chilcut, H. L. Cushing £ M. W. Johnson. 1993. Increasing efficiency of bioassays: Evaluating resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J. Econ.Entomol. 86: 635-644.Talekar, N. S. £ A. N. Shelton. 1993. Biology, ecology and management of the diamondback moth. Annu. Rev. Entomol. 38: 275-301.Waage, J. 1992. Summary, pp. 577-579. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Wyman, J. A. 1992. Management approaches for cruciferous insect pests in central North America, pp. 503-509. In N. s. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Chapter Four
Integration of Plant Resistance, Microbial Control, and an Action Threshold for Cabbage Looper and Diamondbacfc Moth Management on Cabbage in Louisiana.
113
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

114
IntroductionApproximately 2,700 acres of cruciferous vegetables with
a total farmgate value of $4,894,686 were grown in Louisiana in 1995. Cabbage, Brassica oleracea var. capi tat a L., occupied 1,393 acres followed by mustard, B. juncea [L. ] Czern., (857 acres), collards, B. oleracea var. virdis L., (172 acres), turnips, B. campestris var. rapa L., (314acres), and radish, Raphanus sativus L., (2 acres) (Picha & Hinson 1996). in Louisiana and other states in North America, cruciferous crops are susceptible to attack from a complex of lepidopteran insect leaf skeletonizers and defoliators. This insect pest complex consists of armyworms, Spodoptera spp., the cabbage looper (CL), Trichoplusia ni (Hubner), the cabbage white butterfly, Pieris rapae (L.) , and the diamondback moth (DBM), Plutella xylostella (L.). Populations of DBM frequently account for greater than 75% of the insect pest population on cabbage and cause crop loss of up to 100%, making DBM the key insect pest from an economic standpoint (Mallagona 1986). To satisfy cosmetic standards and ensure blemish-free produce, farmers use frequent and multiple applications of synthetic insecticides to control these insects. In 1987, control failures of DBM were reported in a number of southern (Florida, Georgia, North Carolina, Texas) and northern (New York, Wisconsin) states.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

115
These control failures catapulted the DBM, which had hitherto been only a minor pest (Har court 1954) of cruciferous vegetables, into becoming the most destructive (Wyman 1992). The source of insecticide-resistant populations of DBM in New York and Wisconsin was traced to larvae transported on cabbage seedlings imported from the southern states. During 1988-1989, 44 populations of DBM from 19 states were tested for resistance. Many exhibited resistance to one or more of three commonly used synthetic insecticides (methomyl, permethrin, and methamidophos) representing, respectively, the carbamate, pyrethroid, and organophosphate classes of insecticides (Shelton & Wyman 1992, Shelton et al. 1993a).
In addition to North America, insecticide-resistant populations of DBM have been reported from several other countries around the world (Miyata et al. 1986, Hama 1987) . Georghiou (1981) reported resistance in DBM against 36 insecticides in 14 countries. The development of insecticide-resistant insect biotypes result from intense selection pressure exerted by frequent applications of insecticides (Roush & Tabashnik 1990) . Because of the widespread development of resistance in DBM to conventional insecticides, a lot of attention has focused in recent times on use of the microbial insecticide Bacillus thuringiensis Berliner. However, reliance on the use of B . thuringiensis
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

116
as a single component for DBM control is ill advised. Indeed, resistance against B. thuringiensis subsp. kurstaki in DBM has been documented in the Philippines (Kirsch & Schmutterer 1988), Hawaii (Tabashnik et al. 1990), Malaysia (Syed 1992), in Florida in mainland USA (Shelton et al. 1993b), and in Central America (Perez & Shelton 1997). A recurring theme in cases where resistance in DBM against B. thuringiensis subsp. kursteUci has developed is intensive and repeated foliar applications of spore-crystal formulations of its toxins as a single component approach for DBM control (Liu et al. 1996). In Louisiana, farmers apply insecticides - including commercial formulations of B . thuringiensis subsp. kurstaki - at intervals as frequently as every five days for controlling DBM on crucifers (LCES 1997) . Past experiences with DBM provide ample evidence that singlecomponent strategies for its control are futile (Talekar & Shelton 1993) . Clearly then, given the genetic plasticity of the DBM and the necessity of preserving its insecticide susceptibility as a resource, an integrated approach should be developed for managing this insect in Louisiana.
Integrated pest management (IPM) is "a population management system that utilizes all suitable techniques in a compatible manner to reduce pest populations and maintain them at levels below those causing economic injury" (Smith &
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

117
Reynolds 1966). This is the approach needed for DBM management (Lim 1992). Two tactics which can be combined with use of toxins of £. thuringiensis for DBM management on cabbage in Louisiana are host plant resistance and an action threshold for objectively making insecticide application decisions.
Story and Black (Departments of Entomology & Plant Pathology, Louisiana State University; unpublished) screened 189 breeding lines of cabbage for resistance against DBM at two sites in Louisiana in 1987. Several lines exhibited some degree of resistance but no subsequent breeding work followed. In addition, Yandia (1989) evaluated eight commercial cabbage varieties available to growers in Louisiana for resistance against DBM but no significant differences among varieties were detected. A new hybrid cabbage variety, Tropicana, bred for tropical conditions, and reputed resistant to DBM, became available in 1995 to growers in the Caribbean. Because Louisiana experiences subtropical climatic conditions, especially in early summer, it was decided to evaluate the Tropicana cabbage variety in Baton Rouge for resistance against DBM.
Consistent with the IPM concept, insecticides should only be applied based on objective criteria. Action thresholds used for DBM management have either been based on
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

118
visual damage or larval density estimates (Chalfant et al. 1979, Workman et al. 1980, Alam et al. 1987, Maltais et al.1994). Within the context of the pest management problems alluded to above, and the recognition that IPM provides the most viable alternative to the heavy reliance on pesticides, the aim of this study was to integrate host plant resistance, microbial control, and an action threshold for DBM (and tangentially CL) management in Louisiana.
Materials and Methods In April 1996, a field experiment was initiated at
Burden Research Plantation, Louisiana Agricultural Experiment Station, Baton Rouge, LA to study the integration of an action threshold, Bacillus thuringiensis, and host plant resistance for DBM and CL management. Two cabbage varieties, Tropicana (a new hybrid reputed resistant to DBM) (Petoseed, saticoy, CA) and KY Cross (a standard variety) (Takii Seed, Kyoto, Japan), were subjected to three treatments. Treatments were: Agree 50 WP® (wettable powder) (Bacillus thuringiensis subspp. kurstaki and aizawai; Novartis, Greensboro, NC) applied weekly, Agree 50 WP® applied based on an action threshold, and untreated controls. Cabbage varieties and insecticide treatments were replicated four times and assigned randomly to plots in a randomized complete block design. Individual plots were 9.8 x 9.8 m and were
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

119
separated by a distance of 1.5 m. Plant rows were 9.8 m long, 0.9 m wide, and were 1.2 m from center to center. Six- week-old cabbage seedlings, raised in outdoor seedbeds and watered weekly with Miracle Grow"’, were planted 0.46 m apart on raised beds. Mustard (Brassica juncea [L.] Czern.), a preferred host for DBM oviposition (Srinivasan & Moorthy 1991) , was directly seeded on skip rows between cabbage plots approximately four weeks before the cabbage seedlings were transplanted to encourage the build-up of DBM population. Standard cultivation practices for cabbage production were followed (LCES 1996).
Plots were sampled by visual inspection of leaf surfaces weekly for five weeks between 29 May and 26 June. Counts of CL and DBM larvae and generalist predators (ants[Hyraenoptera: Formicidae], lady beetles [Coleoptera:Coccinellidae], and spiders [Acarina spp.]) within their canopy were made from 25 randomly selected cabbage plants in each plot. The microbial insecticide, Agree 50 WP® [3.8%(Al) (25,000 IU per mg), was applied weekly (after sampling)to the appropriate plots or when an action threshold of six DBM larvae per plant was exceeded. A similar actionthreshold was recommended to farmers by Alam et al. (1987) . Because the DBM is the key pest of cabbage in Louisiana in the spring, and the fact that past experience showed that
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

120
successful control of DBM simultaneously took care of CL populations, no threshold for CL was considered. Insecticide applications were made at the recommended rate of 83 g in 15 liters of water using a Solo 425 Knapsack Sprayer (Solo Incorporated, Newport News, VA).
At crop maturity, damage rating and yield data were recorded. Twenty-five randomly selected plants in each plot were rated for larval damage as described by Wyman (1992). Heads were rated for damage on a 1-6 scale of increasing damage with one representing no larval feeding on the head or four wrapper leaves; two and three representing increasing but slight damage to wrapper leaves; and four, five, and six representing increasing head deunage. All heads with a damage rating of three or less were considered marketable according to fresh market standards. And on this basis, for each insecticide treatment and cabbage variety, the percentage of marketable cabbage was calculated. Yield data were recorded from 40 randomly selected cabbage plants in each plot. These plants were cut and taken to the edge of the field where the frame leaves were removed leaving four wrapper leaves as per the specifications for cabbage meeting the US No. 1 grade for fresh market cabbage (USDA 1945) . To get an index for quantifying (in terms of weight) the magnitude of the effect of CL and DBM damage on cabbage yield, the four wrapper
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

121
leaves in addition to head leaves which showed damage by CL and DBM larvae were then removed, counted, and weighed after which the heads were weighed. The weight so derived for the heads was regarded as deunage-free yield; the combined weight of the damaged wrapper and head leaves was regarded as yield loss; and deunage-free yield plus yield loss was regarded as potential yield. Also, ratios (expressed as percent) of damage-free yield and yield loss to potential yield were calculated.
Data were subjected to analysis of variance (ANOVA) using PROC GLM of the SAS system, and, where significant differences among insecticide treatments and cabbage varieties were detected, Tukey's test (SAS Institute 1989) was used for means separation. Additionally, Pearson's product correlations were determined (using PROC CORR of the SAS system) (SAS Institute 1989) among damage-free yield, yield loss, and damage rating of cabbage.
ResultsThe main effects, insecticide treatment and cabbage
variety, had varying impacts on the population densities of CL, DBM, and predators. Insecticide treatment had a marginally significant (F - 2.94; df = 2; P = 0.0571) effect on CL population density per plant, a highly significant (F = 21; df = 2/ P = 0.0001) effect on DBM population density
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

122
per plant, but no significant effect on the population densities of lady beetles (F = 0.49; df = 2; P — 0.6133), ants (F - 1.26; df - 2; P = 0.2878), and spiders (F = 0.27; df = 2; P = 0.7621) per plant. The number of CL and DBM larvae per plant was lower in plots treated weekly with Agree 50 WP® compared with plots receiving the other two insecticide treatments. However, numbers of lady beetles, ants, and spiders per plant were not different among plots receiving all three insecticide treatments (Table 4.1).
Regarding the effect of cabbage variety, there was no significant effect on the population densities of lady beetles (F = 3.20; df - 1; P = 0.0762) and ants (F = 0.01; df = 1; P = 0.9288) by the KY Cross variety compared with the Tropicana variety. However, cabbage variety had a significant effect on the number of CL (F = 27; df = 1; P = 0.0001) , and DBM larvae (F = 54.65; df - 1; P - 0.0001) , and spiders (F = 5.87; df = 1; P = 0.0170) per plant; their numbers were higher on the KY Cross variety compared with the Tropicana variety (Table 4.2). The action threshold of six DBM larvae per plant was exceeded twice, on 12 June and 19 June, resulting in two applications of Agree 50 WP®, only in plots of the KY Cross cabbage variety. The interaction between cabbage variety and insecticide treatment was significant (F = 4.96; df = 2; P = 0.0086) only for DBM.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission
Table 4.1. Effect of insecticide treatment on abundance of cabbage looper, diamondback moth, lady beetles, ants, and spiders per plant on two cabbage varieties
Treatments
ArthropodsB. thuringiensis
(weekly)
S + SEM
B. thuringiensis (threshold)
>< + SEM
Untreated
x + SEM
Cabbage looper 0.24 + 0.05a 0.42 + 0.07b 0.44 + 0.08b
Diamondback moth 0.60 + 0.11a 2.04 + 0.26b 2.27 + 0.31b
Lady beetles 0.03 + 0.01a 0.04 + 0.01a 0.04 + 0.01a
Ants 0.00 + 0.00a 0.02 + 0.01a 0.02 + 0.01a
Spiders 0.11 + 0.02a 0.13 + 0.02a 0.13 + 0.03a
x + SEMs within a row followed by the same letter are not significantly different (Tukey's Test; a = 0.05).
MCJ

124Table 4.2. Effect of cabbage variety on abundance of
cabbage looper, diamondback moth, lady beetles, ants, and spiders across all insecticide treatment regimes
Cabbage varietyArthropods
KY Cross Tropicana
X ± SEM x ± SEM
Cabbage looper 0.55 + 0.07a 0.18 + 0.07b
Diamondback moth 2.48 + 0.26a 0.80 + 0.26b
Lady beetles 0.05 + 0.00a 0.03 + 0.01a
Ants 0.01 + 0.00a 0.01 + 0.01a
Spiders 0.15 + 0.01a 0.09 + 0.02b
x ± SEMs within a row followed by the same letterare not significantly different (Anova F-Test; a = 0.05).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

125
Further examination of this interaction was done using PROC PLOT of the SAS system (SAS Institute 1989) . The cell means for the levels of the two factors, insecticide treatment and cabbage variety, were plotted and are shown in Figure 4.1. The KY Cross cabbage variety supported 0.96, 3.03, and 3.46, while the Tropicana variety supported 0.26, 1.06, and 1.09 DBM larvae per plant in cabbage plots treated weekly with Agree 50 WP®, in plots treated with Agree 50 WP® based on the action threshold of six DBM larvae per plant, and in untreated plots, respectively. Over the sampling period, DBM maintained higher population levels compared with cabbage looper (Figure 4.2) .
Damage-free yield, yield loss, deunage rating per plant, and marketability were significantly impacted by insecticide treatment and cabbage variety. Plots treated weekly with Agree 50 WP® had significantly (F = 7.70; df = 2; P = 0.0038) higher damage-free yield and marketable cabbage (F = 27.72; df = 2; P - 0.0001) but significantly lower yield loss (F - 14.47; df - 2; P = 0.0002) and damage rating (F = 36.39; df = 2; P = 0.0001) per plant compared with plots receiving the other two treatments (Table 4.3).
Concerning the effect of cabbage variety, yield loss (F = 31.97; df = 1; P - 0.0001) and damage rating (F = 119.15; df - 1; P - 0.0001) were significantly lower, but damage-free
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

No. o
f La
rvae
14
3
2.1
2
1J
1 OJ
0BL (weekly) Threshold Untreated
Insecticide Treatment
Figure 4.1. Mean number of diamondback moth larvae per plant across insecticide treatments and cabbage varieties.
126
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.
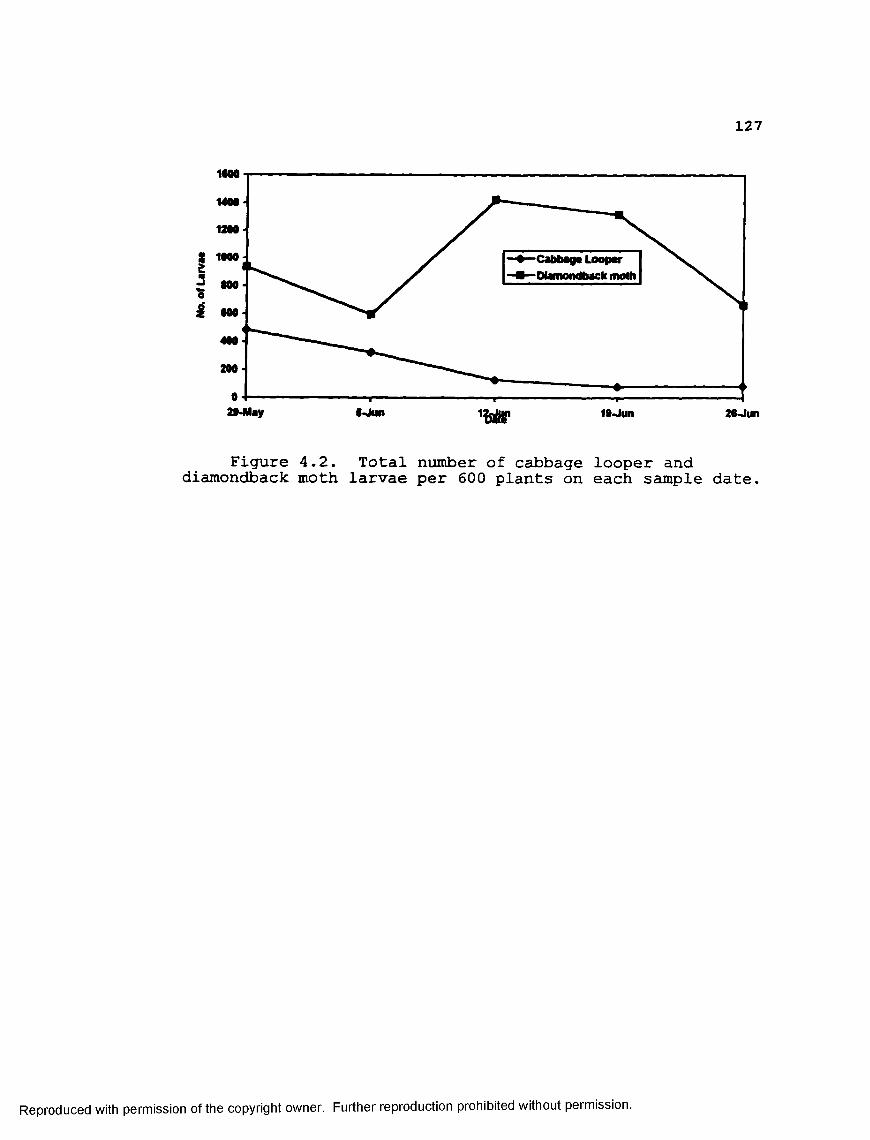
127
Diamondback moth
200
19-Jun 28-Jun
Figure 4.2. Total number of cabbage looper and diamondback moth larvae per 600 plants on each sample date.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission
Table 4.3. Effect of insecticide treatment on damage-free yield, yield loss, damage rating per plant, and marketability of two cabbage varieties
Treatments
ParameterB. thuringiensis
(weekly)
x + SEM
B. thuringiensis (threshold)
S + SEM
Untreated
x + SEM
Damage-free yield 1.51 + 0.06a 1.25 + 0.06b 1.24 + 0.09b(kg)
Yield loss (kg) 0.09 + 0.02a 0.24 + 0.04b 0.25 + 0.05b
Damage rating 1.96 + 0.34a 3.35 + 0.41b 3.56 + 0.39b
Marketability (%) 86.50 + 7.25a 49.50 + 16.40b 48.50 + 16.62b
x + SEMs within a Test; a = 0.05).
row followed by the same letter are not significantly different (Tukey's
HM00

129
yield (F = 18.33; df - 1; P = 0.0004) and marketable cabbage (F ~ 195.08; df - 1; P - 0.0001) were significantly higher for the Tropicana variety compared with the KY Cross variety (Table 4.4). The interaction between insecticide treatment and cabbage variety was significant (F = 17.16; df = 2; P = 0.0001) only for marketability of cabbage. Further examination of this interaction was done using PROC PLOT of the SAS system (SAS Institute 1989) . The cell means for the levels of the two factors, insecticide treatment and cabbage variety, were plotted and are shown in Figure 4.3. The marketability of cabbage heads of the KY Cross variety was 73%, 7%, and 5%, while the marketability of those of the Tropicana variety was 100%, 92%, and 92% in plots treated weekly with Agree 50 WP®, in plots treated with Agree 50 WP® based on the action threshold, and in untreated plots, respectively.
Potential yield of cabbage was not significantly (F = 2.02; df -2; P - 0.1610) different in plots receiving the three insecticide treatments (Table 4.5) but was significantly (F = 5.28; df — 1; P — 0.0338) higher in plots of the Tropicana variety compared with KY Cross (Table 4.6) . The ratios of deunage-free yield and yield loss to potential yield of cabbage was significantly affected by insecticide treatment and cabbage variety. The ratio of damage-free
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

130
Table 4.4. Effect of cabbage variety on damage-free yield, yield loss, damage rating per plant, and marketability of two cabbage varieties across all insecticide treatment regimes
ParameterCabbage variety
KY
X
Cross
± SEM
Tropicana
x ± SEM
Damage-free yield (kg)
1.19 + 0.07a 1.47 + 0.05b
Yield loss (kg) 0.27 + 0.04a 0.12 + 0.02b
Damage rating 3.93 + 0.28a 2.05 + 0.21b
Marketability
(%)
28.33 + 10.28a 94.67 + 2.22b
x ± SEMs within a row followed by the same letterare not significantly different (Anova F-Test; a = 0.05).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Mar
ketab
le ca
bbag
e (%
)
100 ♦00-
•0 ■
70 ;00- Tropicana
XY Cross00 •40-00 •20 -
UntreatedInecticlde Treatment
Figure 4.3. Mean marketability of cabbage across insecticide treatments and cabbage varieties.
131
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 4.5. Effect of insecticide treatment on yield parameters of two varieties of cabbage
Treatments
ParameterB. thuringiensis
(weekly)B. thuringiensis
(threshold)Untreated
x + SEM x + SEM * + SEM
Potential yield (kg) 1.60 + 0.05a 1.49 + 0.05a 1.49 + 0.06a
Damage-free yield/Potential yield <%)
93.75 + 1.49a 83.50 + 2.73b 82.38 + 4.20b
Yield loss/Potential yield (%)
6.25 + 1.49a 16.50 + 2.73b 17.62 + 4.20b
x + SEMs within a row followed by the same letter are not significantly different (Tukey's Test; a = 0.05).
utu

133
Table 4.6. Effect of variety on yield parameters of cabbage across all insecticide treatment regimes
ParameterCabbage variety
KY Cross
x ± SEM
Tropicana
x ± SEM
Potential yield 1.46 + 0.04a 1.59 + 0.04b(kg)
Damage-freeyield/Potential 80.75 + 2.99a 92.30 + 1.14byield (%)
Yieldloss/Potential 19.25 + 2.99a 7.67 + 1.14byield (%)
5? ± SEMs within a row followed by the same letterare not significantly different (Anova F-Test; a = 0.05).
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

134
yield of cabbage to potential yield was significantly (F = 8.57; df = 2; P = 0.0024) higher, but the ratio of yield loss to potential yield was significantly (F = 8.57; df - 2; P = 0.0024) lower in plots treated weekly with Agree 50 WP® compared with plots receiving the other two insecticide treatments (Table 4.5). Regarding the effect of cabbage variety, the ratio of damage-free yield of cabbage to potential yield was significantly (F = 21.95; df - l; P = 0.0002) higher but the ratio of yield loss to potential yield significantly (F = 21.95; df = 1; P = 0.0002) lower in plots of the Tropicana variety compared with plots of KY Cross (Table 4.6). There were no significant interactions between insecticide treatment and cabbage variety relative to the potential yield and ratios of damage-free yield and yield loss to potential yield of cabbage. There were significant and high negative correlations between marketable yield of cabbage and marketable yield loss (r = - 0.85566; Pr > r = 0.0001) and between damage rating and marketable yield (r = - 0.78644; Pr > r — 0.0001). The correlation between damage rating and marketable yield loss was significant and positive (r = 0.87142; Pr > r = 0.0001).
DiscussionThe significantly lower number of DBM larvae per plant
in plots treated weekly with Agree 50 WP® compared with the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

135
other two treatments is indicative of the effectiveness of the microbial insecticide. The efficacy of the parasporal crystal protein toxins of B. thuringiensis against DBM is well documented (Hofte & Whiteley 1989). Therefore, the level of CL and DBM control achieved with Agree 50 WP® indicated that these Louisiana populations were susceptible to toxins of B. thuringiensis. Ivey et al. (1996) reported that a DBM population collected in St. Francisville, Louisiana was susceptible to two other commercial formulations of B. thuringiensis (Dipel ES® [B. thuringiensis subsp. kurstaki] and ZenTari AS® [B. thuringiensis subsp. aizawai]. That the population densities per plant of all predators were not significantly different in all three treatments was not surprising because their overall population levels were generally low relative to that of DBM over the sampling period. But the presence of predators in the plots underscores the safety of B. thuringiensis to beneficial arthropods. Preservation of predators is highly desirable because they, unlike parasitoids, immediately kill or disable larvae, thereby preventing further cosmetic injury to cabbage plants. Although beneficial predators may have only a minor impact by themselves, they contribute to overall pest mortality (Hoffmann & Frodsham 1993) . The existence of a significant interaction makes it necessary to exercise
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

136
caution when making statements about the main effects (cabbage variety and insecticide treatments), even though both were statistically significant with P values of 0.0001. Graphical representation of the cell means is very useful in interpreting the consequences of interaction (Freund & Wilson1993) . Examination, by graphical representation, of the significant interaction between insecticide treatment and cabbage variety, relative to DBM larval density, showed that the resistant variety, Tropicana, consistently supported fewer DBM larvae per plant across all three insecticide treatments compared with the susceptible variety, KY Cross (Figure 4.1). Also, Figure 4.3, which represents the interaction between insecticide treatment and cabbage variety relative to the marketability of cabbage, showed that the Tropicana cabbage variety consistently produced a higher percentage of marketable cabbage across all three insecticide treatments compared with the KY Cross variety. These plots, Figures 4.1 and 4.3, clearly indicated that the Tropicana cabbage variety was superior to the KY Cross variety in resisting DBM and that the Tropicana variety may be compatibly combined with use of B. thuringiensis for successful DBM management. Also, this was consistent with previous findings (see chapters one & two). Further evidence of the superiority of the Tropicana variety over the KY Cross
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

137
variety was seen when marketable yield, marketable yield loss (and their ratios to potential yield), and damage rating of cabbage also were considered (Tables 4.4 & 4.6). The damage ratings of cabbage heads harvested from plots treated weekly with Agree 50 WP® and those of the Tropicana cabbage variety were 1.96 and 2.05 respectively and, therefore, met fresh market standards. However, cabbage heads from plots which received the other two treatments and those of the KY Cross variety had damage ratings of 3.45, 3.56, and 3.93,respectively, and so, did not meet these standards (Tables 4.3 & 4.4) . This is consistent with the damage rating system used by Wyman (1992) where cabbage heads with damage ratings of three or less satisfied the criteria for meeting fresh- market standards, and further underscores the effectiveness of Agree 50 WP® and the ability of the Tropicana variety to resist DBM compared with the KY Cross variety.
The significant and high negative correlations between marketable yield of cabbage and marketable yield loss, and between damage rating and marketable yield were not surprising. Both Tropicana and KY Cross cabbage varieties are in the same maturity class and, on average, have heads of similar size and weight. Intuitively, therefore, one would expect that as marketable yield of cabbage increased then marketable yield loss would decrease, and as damage rating
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

138
increased then marketable yield would decrease and vice versa. Also, the significant and high positive correlation between deunage rating and yield loss was, again, not unexpected because, if deunage rating increased then yield loss would, in tandem, increase. Clearly then, it would be advantageous to the Tropicana cabbage variety that supported fewer CL and DBM larvae, produced higher damage-free yield and marketable cabbage, and had lower yield loss and damage rating compared with the KY Cross variety. The advantages of the Tropicana variety may translate into increased gross returns for cabbage growers.
The number of CL and DBM larvae, predators, damage-free yield, yield loss, damage rating, and marketability of cabbage were not significantly different in plots treated based on the action threshold of an average of six DBM larvae per plant compared with plots that were untreated. In fact, the action threshold was exceeded only twice over the sampling period, resulting in two applications of Agree 50 WP®. That the damage rating of plots treated based on this action threshold was significantly higher (3.45) compared with a damage rating of 1.96 for plots treated weekly with Agree 50 WP®, suggests that this action threshold, recommended by Alam et al. (1987), was inadequate for making insecticide application decisions for controlling DBM on
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

139
cabbage. Several action thresholds for lepidopterous pests of cabbage have been used. Chalfant et al. (1979) proposed a visual damage threshold of one to two new feeding holes per plant, whereas Workman et al. (1980) proposed one to two initial feeding sites (windows) per plant to obtain cabbage meeting fresh market standards. Additionally, Shelton et al. (1982) proposed an action threshold of 0.5 cabbage looper equivalents (CLE) (1 CLE - 20 DBM) per plant at the heading stage to obtain damage-free cabbage. In a study comparing the above-mentioned action thresholds, Shelton et al. (1983) found that the action threshold proposed by Chalfant et al. (1979) took less time to assess, required the fewest insecticide applications, but its use led to plants sustaining the most injury. Whereas use of action thresholds has resulted in reductions in the number of insecticide applications, there have been concomitant reductions in marketable yield of cabbage that meets current standards. The results of this study agree with this finding. It seems that as long as quality standards demand that harvested cabbage heads, and the adjacent four wrapper leaves, be damage free (as is required for US No. 1 grade) (USDA 1945), action thresholds will have little practical value. Indeed, based on these exacting cosmetic standards cabbage may be regarded as having nonthresholds, a situation typical of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

140
vegetable and fruit crops with extreme cosmetic requirements (Luckmann & Metcalf 1994).
Consumers exert a significant influence on insect pest control for cruciferous vegetables. By demandingcosmetically perfect produce they stimulate increased usage of insecticides. Collins et al. (1992) alluded to the contradiction whereby consumers desire perfect, blemish-free produce yet do not want produce subjected to heavy pesticide application. Fresh market cabbage provides a classical example of this contradiction and also highlights the fact that socio-economic factors, such as cosmetic standards, can greatly influence insect pest management. Perhaps it is time for consumers to accept cabbage with minor insect damage because good control is achievable with safe insecticides containing toxins of B. thuringiensis. However, the toxins have to be ingested to be effective, hence some feeding damage is inevitable. Market research to evaluate programs designed to offer consumers choices of products with different production practices is needed. So too is public education regarding the relationship between cosmetic appearance and pesticide use (Powers & Heifner 1993). Lowering current quality standards for fresh market cabbage may be warranted to increase the durability of B. thuringiensis-based insecticides and prolong the longevity of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

141
DBM susceptibility to their toxins. This is necessary inlight of the documented cases of resistance in DBM againstfoliar applications of B. thuringiensis subsp. kurstaki inthe Philippines (Kirsch & Schmutterer 1988), Hawaii(Tabashnik et al. 1990, 1991, 1992), Malaysia (Syed 1992),Florida in mainland USA (Shelton et al. 1993b), and CentralAmerica (Perez & Shelton 1997).
References Cited - Chapter FourAlam, M. M«, J. Reid 8 I. H. Gibbs. 1987. Diamondback moth and its control. Caribbean Agricultural Research and Development Institute, Factsheet # PP-F/5.87.Chalfant, R. B., W. H. Denton, D. J. Schuster 6 R. B. Workman. 1979. Management of cabbage caterpillars in Florida and Georgia by using visual damage thresholds. J. Econ. Entomol. 72: 411-413.Collins, J. K., G. W. cuperus, B. Cartwright, J. A. Stark 8 L. L. Ebro. 1992. Consumer attitudes on production systems for fresh produce. J. Sustain. Agric. 3: 67-77.Freund, R. J. & W. J. Wilson. 1993. Statistical Methods. Academic Press. San Diego, CA.Georghiou, G. P. 1981. The occurrence of resistance to pesticides in arthropods. An index of cases reported through 1980. FAO, RomeHama, H. 1987. Development of pyrethroid resistance in the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae). Appl. Entomol. Zool. 22: 166-175.Harcourt, D. G. 1954. The biology and ecology of the diamondback moth, Plutella maculipennis Curtis, in eastern Ontario. Ph.D. Dissertation, Cornell University, Ithaca, NY.Hoffmann, M. P. 8 A. C. Frodsham. 1993. Natural enemies of vegetable insect pests. Cooperative Extension, Cornell University, Ithaca, NY.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

142
Hofte, H. £ H. R. Whiteley. 1989. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 53: 242- 255.Ivey, P .w., R. N. Story £ s. J. Johnson. 1998. Response of a Louisiana population of diamondback moth to Bacillus thuringiensis strains, pp. 77-80. In H. Y Hanna (ed.), Vegetable Research Report 1996. Louisiana Agricultural Experiment station Mimeo Series 122.Kirseh, K. £ J. Schmutterer. 1988. Low efficacy of a Bacillus thuringiensis (Ber.) formulation in controlling the diamondback moth, Plutella xylostella (L.) , in the Philippines. J. Appl. Entomol. 105: 249-255.LCES. 1996. Commercial vegetable production recomendations. Louisiana Cooperative Extension Service, Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station, Baton Rouge, LA.LCES. 1997. Insect control guide. Louisiana Cooperative Extension Service, Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station, Baton Rouge, LA.Lim, 6. 1992. Integrated pest management of diamondbackmoth: Practical realities, pp. 565-576. In N. S. Talekar (ed.) Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Liu, Y., B. E. Tabashnik £ M. Pusztai-Carey. 1996. Field- evolved resistance to Bacillus thuringiensis toxin CrylC in diamondback moth (Lepidoptera: Plutellidae). J. Econ.Entomol. 89: 798-804.Luckmann, W. H. £ R. L. Metcalf. 1994. The pest-management concept, pp. 1-31. In R. L. Metcalf & W. H. Luckmann (eds.), Introduction to Insect Pest Management. John Wiley £ Sons, New York.Magallona, E. D. 1986. Developments in diamondback moth management in the Philippines. pp. 423-435. In N. S. Talekar £ T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International workshop. AVRDC, Shanhua, Taiwan.Maltais, P. M., J. R. Nuckle £ P. V. LeBlanc. 1994.Economic threshold for management of lepidoptera larvae on
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

143
broccoli in southeastern New Brunswick. J. Econ. Entomol. 87: 766-774.Miyata, T., T. Saito a V. Noppun. 1986. studies on themechanism of diamondback moth resistance to insecticides, pp. 347-357. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Peres, C. J. and A. M. Shelton. 1997. Resistance ofPlutella xylostella (Lepidoptera: Plutellidae) to Bacillus thuringiensis Berliner in Central America. J. Econ. Entomol. 90: 87-93.Picha, D. H. 6 R. A. Hinson. 1996. The commercial vegetable industry in Louisiana, pp. 6-8. In J. Tarver & J. Lord (eds.), Louisiana Agriculture. Vol. 39, No. 3.Powers, N. J. 6 R. G. Heifner. 1993. Federal gradestandards for fresh produce: Linkages to pesticide use.Agriculture Information Bulletin No. 675, August 1993. United States Department of Agriculture, Beltsville, MD.Roush, R. T. 6 B. E. Tabashnik (ads.). 1990. PesticideResistance in Arthropods. Chapman & Hall, New York.SAS Institute. 1989. SAS/STAT user's guide: Statistics, 5th edition. SAS Institute, Cary, NC.Shelton, A. M., J. T. Andaloro £ <7. Barnard. 1982. Effects of cabbage looper, imported cabbage worm, and diamondback moth on fresh market and processing cabbage. J. Econ. Entomol. 75: 742-745.Shelton, A. M., M. K. Sears, J. A. Hyman £ T. C. Quick. 1983. Comparison of action thresholds for lepidopterous larvae on fresh market cabbage. J. Econ. Entomol. 76: 196- 199.Shelton, A. M. £ J. A. Hyman. 1992. Insecticide resistance of diamondback moth (Lepidoptera: Plutellidae) in NorthAmerica, pp. 447-454. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Shelton, A. M., J. A. Hyman, N. L. Cushing, K. Apfelbeck, T.J Dennehy, S. E. R. Mahr £ s. D. Eigenbrode. 1993a.Insecticide resistance of diamondback moth, Plutella
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

144
xylostella (Lepidoptera: Plutellidae), in North America. J. Econ. Entomol. 86: 11-19.Shelton, A. M., J. L. Robertson, J. D. Tang, c. Perez, s. D. Eigenbrode, H. K. Preisler, W. T. Wilsey & R. j. Cooley. 1993b. Resistance of diamondback moth (Lepidoptera: Plutellidae) to Bacillus thuringiensis subspecies in the field. J. Econ. Entomol. 86: 697-705.Smith, R. F 6 H. T. Reynolds. 1966. Principles, definitions and scope of integrated pest control, pp. 11-17. in Proceedings, Symposium on Integrated Pest Control, Food & Agriculture Organization, Rome.Sriniv&san, K. 6 P. M. K. Moor thy. 1991. Indian mustard as a trap crop for management of major lepidopterous pests on cabbage. Tropical Pest Management. 37: 26-32.Syed, A. R. 1992. Insecticide resistance in diamondback moth in Malaysia, pp. 437-442. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Tabashnik, B. E., N. L. Cushing, N. Finson 6 M. w. Johnson. 1990. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae) . J. Econ. Entomol. 83: 1671-1676.Tabashnik, B. E, N. Finson 6 M. V. Johnson. 1991. Managing resistance to Bacillus thuringiensis: Lessons from thediamondback moth (Lepidoptera: Plutellidae). J. Econ.Entomol. 84: 49-55.Tabashnik, B. E., J. M. Schwartz, N. Finson 6 M. W. Johnson. 1992. Inheritance of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J. Econ.Entomol. 85: 1046-1055.Talekar, N. S. 6 A. M. Shelton. 1993. Biology, ecology and management of the diamondback moth. Annu. Rev. Entomol. 38: 275-301.USDA. 1945. United States standards for grades of cabbage. 18 Federal Register 7084 (1953), redesignated 46 FR 63203 (1981) . United States Department of Agriculture, Beltsville, MD.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

145
Workman, R. B., R. B. chalfant & D. j. Schuster. 1980.Management, of the cabbage looper and diamondback moth on cabbage using two damage thresholds and five insecticide treatments. J. Econ. Entomol. 73: 757-758.Wyman# J. A. 1992. Management approaches for cruciferous insect pests in central North America# pp. 503-509. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Yandia, A. 1989. Host plant resistance to the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae) , M.S. Thesis, Louisiana State University, Baton Rouge.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Chapter Five
Insecticide Use Reduction in Management Programs for Cabbage Looper And Diamondback Moth
146
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

147
IntroductionGlobally, approximately 2.5 million tons of pesticides
costing $20 billion are used annually (PN 1990} . In the United States alone, an estimated 500,000 tons of 600 different kinds of pesticides are used each year at a cost (purchase price plus application) of $4.1 billion (Pimentel et al. 1992) . However, even with this high level of pesticide usage, pests - mainly weeds, plant pathogens, and insects - still are responsible for destroying 37% of food and fiber crops (Pimentel & Lehman 1993). Without pesticides, Pimentel et al. (1978) estimated that losses to pests would increase by 10%, but specific crop losses would be between zero and 100%. Undoubtedly therefore, pesticides play a pivotal role in the protection and maintenance of world food production.
The conventional wisdom is that pesticide usage has a cost/benefit ratio of 1:4, i.e., each dollar spent onpesticides returns $4 in crops saved (Carrasco-Tauber 1989, Metcalf & Luckmann 1994). Most benefits derived from using pesticides are predicated on direct crop returns. These assessments do not take into account the environmental impacts and economic costs related to pesticides. In other words, these costs are externalized, i.e., foisted upon
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

148
society and ecosystems, and include: impacts on human health (EPA 1977, Ecobichon et al. 1990); livestock and livestock product losses (Beasley & Trammel 1989); pest resistance, pest population resurgence, outbreaks of secondary pest populations, destruction of beneficial organisms (Georghiou 1990, Brown et al. 1994) ; crop pollination problems and honeybee losses (Mussen 1990); crop, crop product, fish, wildlife, and microorganism losses (Stinger & Lyons 1974, Mineau & Collins 1988, Clark 1989); ecological disruptions; and government spending to reduce the environmental and social costs of pesticide use (GAO 1986, USBC 1990). Clearly, the serious nature of these negative environmental and social impacts of pesticides dictates that current and future pest control programs be re-evaluated to include ecologically-sound and sustainable pest management practices.
Reduced usage is a tactic which is currently being implemented in several European countries to mitigate the adverse effects of pesticides (Hurst et al. 1992, Reus et al.1994), and may contribute to building and maintaining a more sustainable agricultural system. Denmark, Sweden, and The Netherlands have implemented pesticide use reduction (PUR) policies mainly in response to environmental concerns about agrichemical pollution. In the United States, legislation
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

149
regarding reduced pesticide usage has been debated in the U.S. Congress. Two bills regarding pesticide use reduction were introduced in 1994. The "Federal Insecticide, Fungicide and Rodenticide Act (FIFRA) Amendments of 1994" contained "pilot programs designed to establish and implement pesticide use reduction goals in selected ecosystems, in cooperation with agricultural producers, federal, state, and local officials, and other appropriate public and private entities". Also, the "Farm Viability and Pest Management Improvement Act of 1994" was introduced. This act has the backing of a coalition of consumer, environmental, and sustainable agriculture organizations and seeks to bring about a national program which would place emphasis on pesticide use reduction in agriculture. In addition to the legislative approaches alluded to above, in 1993 the Environmental Protection Agency (EPA) and the United States Department of Agriculture (USDA) started a consultative process (the Pesticide Environmental Stewardship Program (PESP)) for establishing regional, commodity-specific pesticide use reduction targets to be met by the year 2000 (Matteson 1995). Another policy initiative aimed at significantly reducing pesticide usage was announced in December 1994. The USDA announced an initiative to help
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

150
farmers implement IPM on 75% of total crop acreage by the year 2000 (OTA 1995, Jacobsen 1996, NRC 1996).
The most destructive insect pest of cruciferous vegetables worldwide is the diamondback moth (DBM), Plutella xylostella (L.) (Talekar & Shelton (1993). These vegetables, members of the plant family Cruciferae, occur in temperate and tropical regions and represent a disparate, widely distributed, and important group of plants that include broccoli, cabbage, cauliflower, collards, and mustard. Because these vegetables usually have high cosmetic standards, effective control of the DBM is necessary. Historically, the primary method of control of DBM involves insecticides applied by air or ground equipment, or with knapsack sprayers. However, as a direct consequence of this dependence on synthetic insecticides, in North America (Shelton & Wyman 1992, Shelton et al. 1993), as well as in many developing countries (Suah & Ellis 1974, Georghiou 1986, Miyata et al. 1986) , insecticide-resistant populations of DBM have developed.
Armyworms, Spodoptera spp., cabbage looper (CL), Trichoplusia ni [Hubner], and the cabbage white butterfly, Pierls rapae (L.) also are leaf skeletonizers of crucifers. However, their population densities rarely, if ever, approach
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

151
those of DBM during a growing season. In addition, these insects are relatively susceptible to most insecticides and no cases of insecticide resistance have been reported.
In Louisiana, the Cooperative Extension Service of Louisiana State University's Agricultural Center recommends several insecticides at specified rates for controlling CL and DBM. These insecticides include organophosphates, carbamates, pyrethroids, and products containing toxins of the soil bacterium, Bacillus thuringiensis Berliner (LCES 1997) .
The magnitude of insecticide resistance by the DBM, the importance of crucifers in the diet of many people, the adverse impacts that excessive use of pesticides have on the environment, the success of pesticide use reduction policies in Europe, and the intention of the EPA, USDA, FDA, and other federal agencies to pursue policy initiatives aimed at pesticide use reduction in American agriculture, make it timely to commence research on reduced pesticide usage for managing the major insect pests of cruciferous vegetables. Therefore, within the context of developing a rational and sustainable management system for CL and DBM, the objective of this study was to test the effectiveness of using reduced rates of insecticides for controlling these insects and
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

152
determine the impact of these reduced insecticide rates on beneficial arthropods.
Materials and MethodsIn May-June 1997, a field experiment was conducted at
Burden Research Plantation, Louisiana Agricultural Experiment Station, Baton Rouge, LA. Three insecticides recommended by the Louisiana Cooperative Extension Service (LCES 1997), an organophosphate (Malathion 50® [emulsifiable concentrate]; Riverside/Terra Corp., Sioux City, IA), a pyrethroid (Ambush 2 EC® [emulsifiable concentrate]; ICI Americas Inc., Wilmington, DE) , and a microbial insecticide (Agree 50 WG® [wettable granule]; Novartis, Greensboro, NC) were applied weekly to cabbage plots using Solo 425 Knapsack Sprayers (Solo Incorporated, Newport News, VA) . All three insecticides were applied to randomly chosen plots at rates corresponding to 25, 50, and 100% of the highest recommended rates. Untreated plots served as controls and all treatments were replicated four times in a randomized completed block design.
The cabbage variety Early Round Dutch (Alexandria Seed Inc., Alexandria, LA) was planted in plots that were 9.8 x9.8 m and separated by a distance of 1.5 m. Plant rows were9.8 m in length, 0.9 m wide, and were 1.2 m from center to
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

153
center. Five-week-old cabbage seedlings, raised in an outdoor seedbed and watered weekly with Miracle Grow8, were planted 0.4 m apart on raised beds. Mustard, Brassica juncea [L. ] Czern, a preferred host for DBM oviposition (Srinivasan & Moorthy 1991), was directly seeded on skip rows between cabbage plots four weeks before the cabbage seedlings were transplanted to attract and encourage the build-up of feral DBM populations. Standard cultivation practices for cabbage production were followed (LCES 1996).
Plots were sampled by visual inspection of leaf surfaces weekly for five weeks between 28 May and 25 June. Counts of CL and DBM larvae and generalist predators (ants [Hymenoptera: Formicidae], lady beetles [Coleoptera:Coccinellidae], and spiders (Acarina spp.]) within their canopy were made from 15 randomly selected cabbage plants in each plot. The three rates of the three insecticides were applied weekly (after sampling) using dedicated Solo 425 Knapsack Sprayers. For Agree 50 WG® (Bacillus thuringiensis subspp. kurstaki and aizavrai; 3.8% (Al) [25,000 IU per mg]), the quantities used in 15 liters of water were 20 g (25%), 40 g (50%), and 80 g (100%). In the case of Ambush 2 EC® ([3- phenoxyphenyl] methyl [+] cis, trans-3-[2,2-dichloroethenyl]- 2,2-dimethyl cyclopropanecar boxy late) , the quantities used in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

154
15 liters of water were 3.75 mis (25%), 7.5 mis (50%), and 15 mis (100%). For Malathion 50® (o,o- dimethyl-phosphorodithioate of diethyl mercaptosuccinate), the quantities used in 15 liters of water were 8.5 mis (25%), 17 mis (50%), and 34 mis (100%) . For each insecticide, spraying commenced with the lowest rate and ended with the highest rate. At the end of spraying, sprayers were washed with a solution of household ammonia and then triple rinsed with water.
At crop maturity, damage rating data were recorded; fifteen randomly selected plants in each plot were rated for larval deunage on 1-6 scale of increasing damage. On this scale, one represented no larval feeding on the head or four wrapper leaves; two and three represented increasing but slight damage to wrapper leaves; and four, five, and six represented increasing head damage. Heads with a damage rating of three or less were considered marketable according to fresh market standards (Wyman 1992). For each insecticide and application rate, the percentage of marketable cabbage was calculated.
Data were subjected to analysis of variance (ANOVA) using PROC GLM of the SAS system, and, where significant differences among insecticides and application rates were
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

155
detected, Tukey's HSD test (SAS Institute 1989) was used for the separation of means.
ResultsThe main effects, insecticide and application rate, had
varying impacts on the population densities of CL, DBM, and predators. Insecticide had highly significant effects on CL (F = 8.57; df =3/ P = 0.0001), DBM (F = 69.41; df = 3; P = 0.0001), lady beetle (F = 4.27; df = 3; P = 0.0060), and spider (F - 12.20; df ~ 3; P — 0.0001) population densities per plant , but no significant effect on the population density of ants (F = 1.81; df - 3; P = 0.1461) per plant. The number of CL larvae per plant was lower in plots treated with Agree 50 WG® and Ambush 2 EC® compared with Malathion 50®-treated and untreated plots (Table 5.1). For DBM, larval density per plant was higher in untreated plots compared with plots treated with Agree 50 WG®, Ambush 2 EC®, and Malathion 50®. Ants, lady beetles, and spiders had relatively low population levels in plots treated with all three insecticides and also in untreated plots (Table 5.1).
Concerning the effect of application rate, there was a marginally significant (F = 3.10; df = 2; P = 0.0474) effect on the population density of CL per plant, but no significant effect on the population densities of DBM (F = 1.09; df = 2;
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 5.1. Effect of insecticide on abundance of cabbage looper, diamondback moth, lady beetles, ants, and spiders per plant across three application rates
ArthropodsInsecticide
Malathion 50® Ambush 2 EC® Agree 50 WG® Untreated
Cabbage looper 0.45 + 0.08a 0.18 + 0.04b 0.15 + 0.04b 0.58 + 0.15a
Diamondback moth 0.61 + 0.07b 0.33 + 0.05b 0.25 + 0.05b 2.55 + 0.36a
Lady beetles 0.01 + O.OOab 0.00 + 0.00b 0.01 + O.OOab 0.03 + 0.01a
Ants 0.09 + 0.03ab 0.06 + 0.03b 0.07 + 0.02ab 0.20 + 0.10a
Spiders 0.07 + O.Olbc 0.04 + 0.01c 0.08 + 0.01b 0.16 + 0.02a
x + SEMs in rows followed by the same letter(s) are not different (Tukey's Test;a = 0.05)uia>

157
P = 0.3387), lady beetles (F = 2.29; df = 2; P = 0.1044), ants (F = 0.40; df — 2; P — 0.6677), and spiders (F = 0.22; df - 2; P — 0.8046) . The number of CL larvae per plant was not different in plots treated at all three rates of the three insecticides, but was higher in untreated plots compared with plots treated at 25, 50 and 100% of therecommended rates of the three insecticides (Table 5.2). Similarly, for DBM, larval density per plant was not different in plots treated at 25, 50, and 100% of therecommended rates of the three insecticides, but was higher in untreated plots (Table 5.2). The interaction between insecticide and application rate was not significant for CL (F = 0.15; df = 4; P = 0.9620), DBM (F = 0.36; df = 4; P = 0.8380) , lady beetles (F = 0.72; df = 4; P = 0.5803) , ants (F = 1.88; df = 4; P - 0.1162), and spiders (F = 1.15; df = 4; P - 0.3360). Diamondback moth maintained higher population levels than Cl over the sampling period (Figure 5.1). Regarding damage rating and marketability of cabbage, the main effects, insecticide and application rate, had significant and non significant effects, respectively. Insecticide had highly significant effects on damage rating (F = 211.45; df = 3; P = 0.0001) and marketability of cabbage (F = 77.99; df - 3; P = 0.0001). The damage rating of
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 5.2. Effect of application rate on abundance of cabbage looper/ diamondback moth/ lady beetles, ants, and spiders per plant across three insecticides
ArthropodsApplication Rate
25% 50% 100% Untreated
Cabbage looper 0.32 + 0.06b 0.32 + 0.07b 0.15 + 0.03b 0.58 + 0.15a
Diamondback moth 0.49 + 0.07b 0.39 + 0.05b 0.32 + 0.05b 2.55 + 0.36a
Lady beetles 0.01 + 0.00b 0.01 + O.OOab 0.00 + 0.00b 0.03 + 0.01a
Ants 0.09 + 0.03ab 0.08 + 0.03ab 0.05 + 0.02b 0.20 + 0.10a
Spiders 0.07 + 0.01b 0.07 + 0.01b 0.06 + 0.01b 0.16 + 0.02a
x + SEMs in rows followed by the same! letter(s) are not different (Tukey's Test;a = 0.05)»-•U1

W/UniO'ON
MODiamondback Motti Cabbag* Looper
3M -
300 ■
2M ■
200 •
1M -
100 ■
M ■
4-Jun 19-JunData
Figure 5.1. Total number of cabbage looper and diamondback larvae per 600 plants on each sample date.
159
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

160
cabbage was lower and the percent marketability higher in plots treated with Agree 50 WG® and Ambush 2 EC® compared with Malathion 50®-treated and untreated plots (Table 5.3).
Application rate had no significant effects on deunage rating (F = 0.59; df - 2; P = 0.5626) and marketability of cabbage (F — 0.40; df -2; P = 0.6757) . The damage rating and percentage of marketable cabbage were not different in plots treated at 25, 50, and 100% of the recommended rates of the three insecticides, but damage rating was higher and percentage of marketable cabbage lower in untreated plots (Table 5.4). The interaction between insecticide and application rate was not significant for damage rating (F = 2.21; df - 4; P — 0.0921) and marketability of cabbage (F = 0.61; df- 4; P = 0.6600).
DiscussionThe significantly higher number of CL larvae per plant
in plots treated with Malathion 50® (organophosphate) compared with plots treated with Agree 50 WG® (B. thuringiensis subspp. kurstaki and aizavrai) and Ambush 2 EC® (pyrethroid) indicates that the organophosphate insecticide was less effective in controlling this insect (Table 5.1). In fact, the number of CL larvae in Malathion 50®-treated plots was comparable with that in untreated plots. In the
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission.
Table 5.3. Effect of insecticide on damage rating per plant and marketability of cabbage across three application rates
Yield ParameterInsecticide
Malathion 50® Ambush 2 EC® Agree 50 WG® Untreated
Damage Rating 3.47 + 0.15b 1.27 + 0.08c 1.24 + 0.05c 4.78 + 0.17a
Marketability(%) 51.50 + 5.78b 99.42 + 0.58a 100.00 + 0.00a 18.25 + 5.59c
x + SEMs in rows followed by the same letter(s) are not different (Tukey's Test;a » 0.05)
t-*ON

Reproduced
with perm
ission of the
copyright ow
ner. Further
reproduction prohibited
without
permission
Table 5.4. Effect of application rate on damage rating per plant and marketability of cabbage across three insecticides
Yield Parameter
Application Rate
25% 50% 100% Untreated
Damage Rating 2.05 + 0.27a 2.02 + 0.34a 1.91 + 0.37a 4.78 + 0.17b
Marketability(%) 86.50 + 6.24a 82.50 + 7.96a 82.25 + 8.45a 18.25 + 5.59b
x + SEMs in rows followed by the same letter(s) are not different (Tukey's Test;a = 0.05)
H<J\fo

163
case of DBM, the number of larvae was higher (though not significantly so) in Malathion 50®-treated plots, again suggesting the greater efficacy of the microbial and pyrethroid insecticides. Also, with respect to damage rating and marketability of cabbage, the efficacy of Ambush 2 EC® and Agree 50 WG® was evident; clearly Malathion 50® did not satisfactorily control CL and DBM thereby accounting for the reduced percentage of marketable cabbage and increased damage rating (Table 5.3). The low efficacy of organophosphates in controlling DBM, and DBM's ability to develop resistance against this class of insecticides is well documented in the tropics and subtropics (Sun et al. 1978; Georghiou 1981, 1986, 1990; Miyata et al. 1986; Sun 1992), and in several southern states in North America including Florida (Leibee & Savage 1992),Georgia (Adams 1992), North Carolina (Shelton & Wyman 1992, Shelton et al. 1993) , and Texas (Magaro & Edelson 1990, Plapp et al. 1992).
The effect of application rate on CL and DBM larval infestation of cabbage and on damage rating and marketability clearly showed that the three rates of the three insecticides gave essentially the same results (Tables 5.2 & 5.4). This is an important finding because it indicates that insecticide use reduction is a distinct possibility in management
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

164
programs for the lepidopteran pests of cabbage. From a practical standpoint, if a farmer uses a quarter or one half the recommended rate of an insecticide and gets satisfactory control of Cl and DBM, then it would uneconomical to apply 100% of the recommended rate. Also, if lower rates are effective and higher ones are used, this increases the selection pressure on the treated population and may thus precipitate the development of insecticide resistance. Because the development of insecticide resistance is related to selection pressure (Roush & Tabashnik 1990), fundamental to resistance management is the need to reduce selection pressure. Therefore, control strategies that minimize exposure of DBM (an insect notorious for its ability to quickly develop resistance) to insecticides may be helpful in managing resistance by reducing the intensity of selection (Beck & Cameron 1992, McHugh & Foster 1995). In addition, using lower rates of pesticides also reduces the potential environmental impacts of leachate on groundwater, improves food safety through lowered levels of residues on produce, lessens hazard to applicators, and conserves beneficial organisms. In this study, the population levels of generalist predators were generally low across insecticides and application rates such that clear-cut treatment effects
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

165
were not seen (Tables 5.1 & 5.2). Perhaps this was due to the fact that only those actually in the canopy of plants were counted. However, it is not unreasonable to assume that reduced rates of pesticides would be relatively less detrimental to them. In the case of the microbial insecticide Agree 50 WG®, the safety of B. thuringiensis to humans and non-target organisms is well known (Entwistle et al. 1993); using reduced rates of B. thuringiensis-based insecticides would be doubly safe. And the pyrethroids as a group, because of the low rates (0.125 to 0.225 kg a.i./ha) of active ingredient which are effective against most pests, compared with the rates for organophosphates (1.25 to 2.25 kg a.i./ha) (Ware 1988), are known to be less detrimental to beneficial arthropods.
Testing insecticides at rates lower than those suggested by manufacturers and then making crop-pest-dosage recommendations is not unprecedented. In Denmark, for example, pesticides are tested at dosage rates lower than those requested by manufacturing companies and crop-pest- dosage combinations found effective by the Danish Institute of Plant and Soil Sciences are the only ones recommended to farmers (Reus et al. 1994) . This is being done within the context of the European Union mandate which seeks reductions
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

166
of 50% or more in the use of agricultural pesticides (fungicides, herbicides, insecticides, soil sterilants, and others) by the year 2000. To date, pesticide use reduction policies implemented in several countries in Europe have significantly lowered the use of pesticide active ingredients. In the Netherlands, for instance, pesticide use reduction targets for the year 2000 have already been met on model research farms (that develop new IPM technologies) without yield or revenue losses; in fact, most of the model farms have reported increased profits (Matteson 1995). In the United States, the "Federal Insecticide, Fungicide, and Rodenticide Act Amendments of 1994", the "Farm Viability and Pest Management Improvement Act of 1994", and the "Pesticide Environmental Stewardship Program", together with the USDA's IPM Initiative seem to be a good policy mix aimed at pesticide use reduction in American agriculture.
In light of the pest management problems associated with lepidopteran pests of cabbage, especially the DBM, the negative externalities related to the heavy use of insecticides for their control, and the need to develop sustainable pest management practices, the IPM approach - which seeks to minimize the disadvantages in the use of insecticides and to maximize the advantages - is the only
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

167
practical solution to the dilemma of the "pesticide treadmill". Therefore, tactics such as pesticide use reduction should be incorporated into IPM programs for these pests.
References cited - Chapter FiveAdams, D. B. 1992. On-farm components of diamondback moth management in Georgia, USA, pp. 499-501. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Beasley, V. R. 6 H. Trammel. 1989. Insecticides, pp. 97- 107. In R. W. Kirk (ed.), Current Veterinary Therapy: Small Animal Practice. W. B. Saunders Publishing Co.,Philadelphia, PA.Beck, N. G. 8 P. J. Cameron. 1992. Developing a reduced spray program for Brassicas in New Zealand, pp. 341-350. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Brown, L. R., H. Kane 8 D. M. Roodman. 1994. Vital Signs: The Trends That are Shaping our Future, WorldWatch Institute, Washington, DC. W. W. Norton & Company, NY.Carrasco-Tauber, C. 1989. Pesticide Productivity Revisited. M.S. Thesis, University of Massachusetts, Amherst, MA.Clark, R. B. 1989. Marine Pollution. Oxford: Clarendon Press.Ecobichon, D. J., J. B. Davies, J. Doull, M. Ehrich, R. Joy, D. McMillan, R. MacPhail, L. w. Reiter, W. Slikker & H. Tilson. 1990. Neurotoxic effects of Pesticides, pp. 131- 199. In C. F. Wilkinson & S.R. Baker (eds.), The Effects of Pesticides on Human Health. Princeton Scientific Publishing Co., Inc. Princeton, NJ.Entvistle, P. 7., J. 8. Cory, M. J. Bailey & 8. Higgs (eds.) 1993. Bacillus thuringiensis, An Environmental Biopesticide: Theory and Practice. Wiley, NY.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

168
EPA. 1977. Minutes of Administrator's Pesticide Policy Advisory Committee. U.S. Environmental Protection Agency. March 1977. Washington, D.C.GAO. 1986. Pesticides: EPA's Formidable Task to Assess andRegulate Their Risks. General Accounting Office.Washington, D.C.Georghiou, G. P. 1981. The occurrence of resistance to pesticides in arthropods: An index of cases reported through 1980. FAO, Rome.Georghiou, G. P. 1986. The magnitude of the resistance problem, pp. 14-43. In Pesticide Resistance, Strategies and Tactics for Management. National Academy of Sciences.Washington, D.C.Georghiou, G. P. 1990. Overview of insecticide resistance, pp. 18-41. In M. B. Green, H. M. LeBaron & W. K. Moberg (eds.), Managing Resistance to Agrochemicals: FromFundamental Research to Practical Strategies. American Chemical Society. Washington, D.C.Hurst, P. P., P. Beaumont, C. E. Jorgensen & S. Winther. 1992. Pesticide reduction programmes in Denmark, TheNetherlands, and Sweden. WWF International, Gland, Switzerland.Jacobsen, B. 1996. USDA IPM Initiative. United States Department of Agriculture. Beltsville, MD.LCES. 1996. Commercial vegetable productionrecommendations. Louisiana Cooperative Extension Service, Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station, Baton Rouge, LA.LCES. 1997. Insect control guide. Louisiana Cooperative Extension Service, Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station, Baton Rouge, LA.Leibee, G. L. 6 K. E. Savage. 1992. Insecticide resistance in diamondback moth in Florida, pp. 427-435. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

169
Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Magaro, j. j. & j. V. Edelson. 1990. Diamondback moth (Lepidoptera: Plutellidae) in south Texas: A technique for resistance monitoring in the field. J. Econ. Entomol. 83: 1201- 1206.Matteson, P. 1995. The "50% Pesticide Cuts" in Europe: A glimpse of our future? American Entomologist. 41: 210-220.McHugh, J. J. 5 R. E. Foster. 1995. Reduction of diamondback moth (Lepidoptera: Plutellidae) infestation in head cabbage by overhead irrigation. J. Econ. Entomol. 88: 162-168.Metcalf, R. L. 5 W. H. Luckmann. 1994. Introduction to Insect Pest Management. John Wiley & Sons, Inc., NY.Mineau, P. 6 B. T. Collins. 1988. Avian mortality in agroecosystems: Methods of detection, pp. 3-12. Jn M. P. Greaves, B. D. Smith & P. W. Greig-Smith (eds.), Field Methods for the Study of Environmental Effects of Pesticides. Proceedings of the British Crop Protection Council. Cambridge, UK.Miyata, T., T. Saito % v. Noppun. 1986. Studies on the mechanism of diamondback moth resistance to insecticides, pp. 347-357. In Talekar, N. S. & T. D. Griggs (eds.), Diamondback Moth Management. Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Mussen, E. 1990. California crop pollination. Gleanings in Bee Culture. 118: 646-647.NRC. 1996. Ecologically Based Pest Management: NewSolutions for a New Century. National Research Council, National Academy Press. Washington, D.C.OTA. 1995. Biologically Based Technologies for Pest Control. OTA—ENV-636. Office of Technology Assessment. Washington, D.C.Pimentel, D., J. Krummel, d . Gallahan, J. Hough, A. Merrill, I. Schreiner, P. Vittum, F. Koziol, E. Back, D. Yen 6 S.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

170
Fiance. 1978. Benefits and costs of pesticide use in U.S. food production. BioScience 28: 778-784.Pimental, D., H. Acquay, M. Biltonen, P. Rice, M. Silva, J. Nelson, V. Lipner, S. Giordano, A. Borovits & M. O'Anore. 1992. Environmental and economic costs of pesticide use. BioScience 42: 750-760.Pimentel, D. & B. Lehman (eds.). 1993. The PesticideQuestion: Environment, Economics and Ethics. Chapman & Hall. NY.Plapp, F. «. jr., J. J. Magaro £ J. v. Edelson. 1992.Diamondback moth in south Texas: A technique for resistance monitoring in the field, pp. 443-446. In N. S. Talekar (ed.) , Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.PN. 1990. Towards a reduction in pesticide use. Pesticide News, March 1990.Reus, J. A. W. A., H. J. Weckseler 6 G. A. Pak. 1994.Towards a future EC pesticide policy: An inventory of risks of pesticide use, possible solutions and policy instruments. Center for Agriculture and Environment. Utrecht, The Netherlands.Roush, R. T. & B. E. Tabashnik (ads.). 1990. PesticideResistance in Arthropods. Chapman & Hall, NY.SAS Institute. 1989. SAS/STAT user's guide: Statistics, 5th edition. SAS Institute, Cary, NC.Shelton, A. M. £ J. A. Hyman. 1992. Insecticide resistance of diamondback moth (Lepidoptera: Plutellidae) in NorthAmerica, pp. 447-454. In Talekar, N.S. (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Shelton, A. M., J. A. Hyman, N. L. Cushing, K. Apfelbeck, T. J Dennehy, S. B. R. Mahr £ s. D. Eigenbrode. 1993.Insecticide resistance of diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae), in North America. J. Econ. Entomol. 86: 11-19.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

171
Srinivasan, K. 8 P. N. K. Moor thy. 1991. Indian mustard as a trap crop for management of major lepidopterous pests on cabbage. Tropical Pest Management. 37: 26-32.Stringer, A. fi C. Lyons. 1974. The effect of benomyl and thiophanate-methyl on earthworm populations in apple orchards. Pesti. Sci. 5: 189-196.Suah, J. R. & D. V. Ellis. 1974. Insecticide control of cabbage caterpillars in Jamaica, pp. 263-266. In C. w. D. Brathvaite, R. H. Phelps & F. D. Bennett (eds.), Proceedings of Protection of Horticultural Crops in the Caribbean, University of the West Indies, St. Augustine, Trinidad.Sun, C. N. 1992. Insecticide resistance in diamondback moth, pp. 419-426. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Sun, C. N., H. Chi t H. T. Feng. 1978. Diamondback moth resistance to diazinon and methomyl in Taiwan. J. Econ. Entomol. 71: 551-554.Talekar, M. 8. & A. M. Shelton. 1993. Biology, ecology and management of the diamondback moth. Annu. Rev. Entomol. 38: 275-301.USBC. 1990. Statistical Abstract of the United States, U.S. Bureau of Census, U.S. Department of Commerce. Washington, D.C.Ware, 6. W. 1988. Complete Guide to Pest Control. Thompson Publications, Fresno, CA.Wyman, J. A. 1992. Management approaches for cruciferous insect pests in central North America, pp. 503-509. Jn N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

Summary and ConclusionsThe objectives of this study were to investigate the
efficacy of integrating various control tactics for managing the diamondback moth (DBM), Plutella xylostella. (L.), the most destructive pest of crucifers worldwide; and tangentially its accomplice, the cabbage looper (CL), Trichoplusia ni (Hubner) . Specifically, the study focused on five IPM tactics - an action threshold, host plant resistance, intercropping (companion planting), microbial control, and pesticide use reduction.
Consistent with the Integrated pest management concept, insecticides should only be applied based on objective criteria such as action thresholds. Results indicate that an action threshold of six DBM larvae per plant was inadequate for making insecticide application decisions because cabbage plots treated weekly had significantly fewer CL and DBM larvae, lower damage rating and yield loss, and higher damage-free yield and percentage of marketable cabbage compared with plots treated based on the action threshold.
It seems that as long as quality standards demand that harvested cabbage heads, and the adjacent four wrapper leaves be damage free, action thresholds will have little practical value. Also, relatedly, by demanding cosmetically perfect
172
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

173
produce, consumers stimulate increased usage of insecticides; yet, paradoxically, they do not want produce subjected to heavy pesticide application.
The potential of plant resistance was made clear specifically through the evaluation of a new cabbage hybrid, Tropicana. Data showed that it was superior to two other varieties (KY Cross and Early Jersey) widely grown in Jamaica in resisting attack from DBM. Another tactic, intercropping (companion cropping), which has received renewed attention for insect control, especially among organic gardeners, also was investigated. By increasing diversity by attracting and maintaining beneficial arthropods, producing powerful chemical deterrents to many pest species, or masking the chemical cues that attract pests, companion crops have been reported to reduce insect populations. In this study, the role of tomato - the companion crop used - in insect control was inconclusive.
Concerns are now being raised over the use of microbial insecticides based on the delta-endotoxins of B. thuringiensis because of the documentation of resistance in the DBM against the kurstaki strain in Central America, Hawaii, Malaysia, the Philippines, and mainland USA. However, encouragingly, the B. thuringiensis-based products used in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

174
this study (Agree 50 WP® & WG®; B. thuringiensis subspp. kurstaki and aizawai) gave excellent results in controlling CL and DBM. It also was shown, based on favorable interactions, that the new cabbage hybrid, Tropicana, could be compatibly combined with use of Agree 50 WP® for DBM management.
Regarding pesticide use reduction, an IPM tactic that is fast gaining credibility, results conclusively confirmed its potential for incorporation in management programs for lepidopteran pests of cabbage; plots treated at 25, 50, and 100% of the recommended rates of the three insecticides had essentially the same level of CL and DBM larval infestation, damage rating, and percentage of marketable cabbage.
At the present time, resistance in the DBM against insecticides is widespread in many tropical and subtropical countries and in several states in North America. Control strategies that minimize exposure of the DBM to pesticides may be helpful in managing insecticide resistance by reducing selection pressure. Integrated pest management programs incorporating host plant resistance, microbial control, and pesticide use reduction - tactics found to be effective in this study - and other tactics such as conservation of biological control agents, host-free periods, trap crops,
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

destruction of crop residues, methods must be implemented to
175
and other cultural control achieve this objective.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

BibliographyAdams, 0. B. 1992. On-Farm components of diamondback moth management in Georgia, USA, pp. 499-501. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Adkisson, P. L. & V. A. Dyck. 1980. Resistant varieties in pest management systems, pp. 233-251. In F. G. Maxwell & P. R. Jennings (eds.), Breeding Plants Resistant to Insects. Wiley, New York.Alam, M. M. 1974. Biological control of insect pests of horticultural crops in Barbados, pp. 253-261. In C. w. D. Brathwaite, R. H. Phelps & F. D. Bennett (eds.), Proceedings of Protection of Horticultural Crops irf the Caribbean. University of the West Indies, St. Augustine, Trinidad.Alam, M. M. 1989. Vegetable pests and their natural enemies in Brbados, pp. 295-302. In Proceedings of the Caribbean Food Crops Society.Alam, M. M. 1992. Diamondback moth and its natural enemies in Jamaica and some other Caribbean Islands, pp. 233-243. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Alam, M. M., J. Reid S I. H. Gibbs. 1987. Diamondback moth and its control. Caribbean Agricultural Research and Development Institute, Factsheet # PP-F/5.87.Altieri, M. A. & M. Liebman. 1985. Insect, weed, and plant disease management in multiple cropping systems, pp. 183-218. In C. A. Francis (ed.), Multiple Cropping Systems. Macmillan Publishing Company, NY.Anonymous. 1994. Movement and dispersal and IPM, pp. 136- 137. In Proceedings, Second National Integrated Pest Management Symposium/Workshop. ESCOP Pest Management Strategies Subcommittee and ES IPM Task Force. University Graphics, North Carolina state University, Raleigh, NC.
176
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

177
Bach, C. E. I B. E. Tabashnik. 1990. Effects of nonhost plant neighbors on population densities and parasitism rates of the diamondback moth (Lepidoptera: Plutellidae) . J. Econ. Entomol. 19: 987-994.Beasley, V. R. £ H. Trammel. 1989. Insecticides, pp. 97- 107. In R. W. Kirk (ed.). Current Veterinary Therapy: Small Animal Practice. W. B. Saunders Publishing Co.Philadelphia, PA.Beck, S. D. 1965. Resistance of plants to insects. Annu. Rev. Entomol. 10: 207-232.Beck, M. 6. 6 P. J. Cameron. 1992. Developing a reduced spray program for Brassicas in New Zealand, pp. 341-350.In K. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Bennett, F. D. 6 M. Yaseem. 1972. Parasite introductions for the biological control of three insect pests in the lesser Antilles and British Honduras. PANS. 18: 468-474.Bonnemaison, L. 1965. Insect pests of crucifers and their control. Annu. Rev. Entomol. 10: 233-256.Borror, D. J., c. A. Triplehorn 6 N. F. Johnson. 1989. AnIntroduction to the Study of Insects. Saunders College Publishing. Philadelphia, PA.Brader, L. 1979. Integrated pest control in the developing world. Annu. Rev. Entomol. 24: 225-254.Bradley, J. D. 1971. The identity of Phalaena Tinea xylostella Linnaeus, 1758 (Insecta, Lepidoptera). Bull. Zool. Nomencl. 28: 11-13.Brown, W. M., Jr. 1993. The role of IPM in contemporary agriculture and environmental issues, pp. 171-180. In A.R. Leslie & G. W. Cuperus (eds.). Successful Implementation of Integrated Pest Management for Agricultural Crops.Lewis Publishers, Boca Raton, FL.Brown, L. R., H. Kane £ D. M. Roodman. 1994. Vital Signs: The Trends that are Shaping our Future. WorldWatchInstitute, Washington, D.C. W. w. Norton & Company. NY.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

178
Buranday# R. P. 6 R. 8. Raroa. 1973. Effects of cabbage tomato intercropping on the incidence and oviposition of the diamondback moth, Plutella xylostella (L.). Phillip. Entomol. 2: 369-374.Calderan, J. I. & C. J. Hare. 1986. Control of diamondback moth in Southeast Asia by profenofos, pp. 289-295. Jn N. S. Talekar 6 T. 0. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop, AVRDC, Shanhua, Taiwan.Carson# R. 1962. Silent Spring. Houghton Mifflin, Boston, MA.Carrasco-Tauber, C. 1989. Pesticide Productivity Revisited. M.S. Thesis, University of Massachusetts, Amherst.Cate# J. R. 6 M. K. Hinkle. 1994. Integrated PestManagement: The Path of a Paradigm. National AudubonSociety, Washington, D.C.Chalfant# R. B.# W. K. Denton# D. J. Schuster 6 R. B. Workman. 1979. Management of cabbage caterpillars in Florida and Georgia by using visual damage thresholds. J. Econ. Entomol. 72: 41-413.Cheng# E. Y., T. M. Chou £ C. H. Kao. 1983. Insecticide resistance study in Plutella xylostella (L.). IV. The activities of glutathione S-transferase in the organophosphorous-resistant strains. J. Agric. Res. China. 32: 373-378.Chu# Y. 1986. The migration of diamondback moth, pp. 77-81. Jn N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop, AVRDC, Shanhua, Taiwan.Chua# T. H. 6 P. A. C. Ooi. 1986. Evaluation of three parasites in the biological control of diamondback moth in the Cameron Highlands, Malaysia, pp. 173-184. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop, AVRDC, Shanhua, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

179
CZBC. 1985. Annual report of 1984-1985. Commonwealth Institute of Biological Control, Commonwealth Agricultural Bureaux. London.CIS. 1967. Distribution maps of Pests. Map No. 12, Plutella xylostella (L.) (maculipennis Curt.). Commonwealth Institute of Entomology. London.Clark, R. B. 1989. Marine Pollution. Oxford: Clarendon Press.Coaker, T. H. 1987. Cultural methods: The crop, pp. 79-85. In A. J. Burn, T. H. Coaker & P. C. Jepson (eds.), Integrated Pest Management. Academic Press, NY.Collins, J. K., a. W. Cuperus, B. Cartwright, J. A. Stark 6 L. L. Ebro. 1992. Consumer attitudes on production systems for fresh produce. J. Sustain. Agric. 3: 67-77.Corbet, P. 8. & R. F. Smith. 1976. Integrated control: A realistic alternative to misuse of pesticides?, pp. 661-682. In C. B. Huffaker & P. S. Messenger (eds.), Theory and practice of Biological Control. Academic Press, NY.Creighton, C. S., T. L. McFadden 6 M. L. Robbins. 1975.Complementary influence of host plant resistance and microbial-chemical control of cabbage caterpilars. HortScience 10: 487-488.cromartie, V. J. 1981. The environmental control of insects using crop diversity, pp. 223-251. In D. Pimentel (ed.), CRC Handbook of Pest Management in Agriculture. CRC Press, Boca Raton, FL.Davies, <J. E., R. F. Smith 6 V. Fred. 1978. Agromedicalapproach to pesticide management. Annu. Rev. Entomol. 23:353-366.Dickson, M. H. 6 C. J. Eckenrode. 1975. Variation inBrassica oleracea resistance to cabbage looper and imported cabbage worm in the greenhouse and fields. J. Econ. Entomol. 68: 757-760.Dickson, M. H. 6 c. J. Eckenrode. 1980. Breeding forresistance in cabbage and cauliflower to cabbage looper,
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

180
imported cabbage worm and diamondback moth. J. Am. Soc. Hort. Sci. 105: 782-785.Dickson, M.H., C.J. Eckenrode & A.E. Blaable. 1984. NYIR 9602, NYIR 9605, and NYIR 8329, lepidopterous pest-resistant cabbages. HortScience. 19: 311-312.Dickson, M. H., c. J. Eckenrode t J. Lin. 1986. Breeding for diamondback moth resistance in Brassica oleracea, pp. 137-143. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Dickson, M. H., A. M. Shelton, S. D. Eigenbrode, M. L. vamosy 6 M. Mora. 1990. Breeding for resistance to diamondback moth, Plutella xylostella, in cabbage. HortScience 25: 1643- 1646.Dingle, H. 1972. Migration strategies of insects. Science 175: 1327-1335.Dingle, H. 1986. Evolution and genetics of insect migration, pp. 11-26. In W. Danthanarayana (ed.), Insect Flight: Dispersal and Migration. Springer-Verlag, New York.Dingle, H. 1996. Migration, The Biology of Life on the Move. Oxford University Press, New York, NY.Dueszlen, J. 1995. Pesticides residues in food, pp. 2-4. In The Agriculturist (April 1995). Caribbean Agricultural Communications Ltd., Kingston, Jamaica.Ecobichon, D. J., J. B. Davies, J. Doull, M. Ehrich, R. Joy, D. McMillan, R. MacPhail, L. W. Reiter, W. slikker 6 H. Tilson. 1990. Neurotoxic effects of pesticides, pp. 131- 199. In C. F. Wilkinson & S. R. Baker (eds.), The Effects of Pesticides on Human Health. Princeton Scientific Publishing Co., Inc. Princeton, NJ.Eigenbrode, S. D., K. A. Stoner, A. M. Shelton 6 w. C. Kain.1991. Characteristics of glossy leaf waxes associated with resistance to diamondback moth (Lepidoptera: Plutellidae) in Brassica oleracea, J. Econ. Entomol. 84:1609-1618.
OEigenbrode, s. D. 6 A. M. Shelton. 1992. Resistance to diamondback moth in Brassicai Mechanisms and potential for
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

181
resistant cultivars, pp. 65-74. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Ely, S. 1993. The engineering of plants to express Bacillus thuringiensis 5-endotoxins, pp. 105-124. Jn P. F. Entwistle, J. S. Cory, M. J. Bailey & S. Higgs (eds.), Bacillus thuringiensis, An Environmental Biopesticide: Theory and Practice. John Wiley & Sons Ltd., NY.Entvistle, P. P., J. S. Cory, M. J. Bailey & 8. Higgs (eds.) 1993. Bacillus thuringiensis, An Environmental Biopesticide: Theory and Practice. Wiley, New York.EPA. 1977. Minutes of Administrator's Pesticide Policy Advisory Committee. U.S. Environmental Protection Agency. March 1977. Washington, D.C.Fahmy, A. R. & T. Miyata. 1992. Development and reversion of chlorfluazuron resistance in diamondback moth, pp. 403- 410. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.FAO. 1968. Report of the first session of the FAO panel of experts on integrated pest control. FAO, Rome.Freund, R. J. 6 W. J. Wilson. 1993. Statistical Methods. Academic Press, San Diego, CA.GAO. 1986. Pesticides: EPA's Formidable Task to Assess and Regulate Their Risks. General Accounting Office.Washington, D.C.Georghiou, G. P. 1981. The occurrence of resistance to pesticides in arthropods: An index of cases reported through 1980. FAO, Rome.Georghiou, G. P. 1986. The magnitude of the resistance problem, pp.14-43. In Pesticide Resistance, Strategies and Tactics for Management. National Academy of Sciences. Washington, D.C.Georghiou, G. P. 1990. Overview of insecticide resistance, pp. 18-41. In M. B. Green, H. M. LeBaron & W. K. Moberg (eds.), Managing Resistance to Agrochemicals: From
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

182
Fundamental Research to Practical Strategies. American Chemical Society. Washington, D.C.Gill, S. S., E. A. Cowles 6 P. ▼. Pietrantonio. 1992. Themode of action of Bacillus thuringiensis endotoxins. Annu. Rev. Entomol. 37: 615-636.Graves, J. B., J. A. Ottea, B. R. Leonard, E. Burris 6 S. Micinski. 1993. Insecticide resistance management: Anintegral part of IPM in cotton, pp. 3-5. In J. Tarver & J. Lord (eds.), Louisiana Agriculture 36, Winter 1993. Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station,Baton Rouge, LA.Gregory, B. M., Jr., S. J. Johnson, A. w. Lievens, A. M. Hammond, Jr. & A. Delgado-salinas. 1990. A midlatitudesurvival model of Anticarsia gemmatalis (Lepidoptera: Noctuidae). Environ. Entomol. 19: 1017-1023.Gupta, P. D. £ A. J. Thorsteinson. i960. Food plant relationship of diamondback moth, Plutella xylostella (Curt.) . II. Sensory relationship of oviposition of the adult female. Entomol. Exp. Appl. 3: 305-314.Hama, H. 1987. Development of pyrethroid resistance in the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae). Appl. Entomol Zool. 22: 166-175.Harcourt, D. G. 1954. The biology and ecology of thediamondback moth, Plutella maculipennis Curtis, in eastern Ontario. Ph.D. Dissertation, Cornell University, Ithaca, NY.Harcourt, D. G. 1956. Biology of the diamondback moth in eastern Ontario. I. Distribution, economic history, synonymy, and general descriptions. Quebec Soc. Proct. Plants, pp. 155-160.Harcourt, D. G. 1958. Biology of the diamondback moth in eastern Ontario. II. Life history, behavior and host relationships. Can. Entomol. 89: 554-564.Harcourt, D. G. 1963. Major mortality factors in thepopulation dynamics of the diamondback moth, Plutellamaculipennis (Curt.) (Lepidoptera: Plutellidae). Mem. Can. Entomol. Soc. 32: 55-66.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

183
Harcourt, D. a. 1986. Population dynamics of the diamondback moth in southern Ontario, pp. 3-16. In N. S. Talekar & T. 0. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Hardy, J. E. 1938. Plutella maculipennis Curt., its natural and biological control in England. Bull. Entomol. Res. 29: 343-372.Hoffman, M. P. fi A. C. Fordshaa. 1993. Natural enemies of vegetable insect pests. Cooperative Extension, Cornell University, Ithaca, NY.Hofte, H. 6 H. R. Whiteley. 1989. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 53: 242- 255.Honda, K. 1992. Hibernation and migration of the diamondback moth in north Japan, pp. 43-50. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Hoy, M. A. 6 D. C. Herzog. 1985. Biological Control in Agricultural IPM Systems. Academic Press, NY.Hurst, P. P., P. Beaumont, c. E. Jorgensen t S. winther. 1992. Pesticide reduction programmes in Denmark, The Netherlands, and Sweden. WWF International, Gland, Switzerland.Iheagvam, E. U. 1981. The relationship between weight of insect, age, hardness and nitrogen content of cabbage leaves and fecundity of the cabbage whitefly, Aleyroides brassicae Walker (Homoptera: Aleyroididae) . Z. Ang. Ent. 91: 349-354.Ivey, P. W. 1991. Diamondback moth feeding dynamics on cabbage and collards. M.S. Thesis, Louisiana State University, Baton Rouge.Ivey, P. W., R. N. Story 6 S. J. Johnson. 1996. Response of a Louisiana population of diamondback moth to Bacillus thuringiensis strains, pp. 77-80. In H. Y. Hanna (ed.), Vegetable Research Report 1996. Louisiana Agricultural Experiment Station Mimeo Series 122.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

184
Jacobsen, B. 1996. USDA IPM Initiative. United States Department of Agriculture. Beltsville, MD.Johnson, 8. J. 1986. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere. Insect Sci. Applic. 8: 543*549.Johnson, 8. J. 1995. Insect migration in North America: Synoptic scale transport in a highly seasonal environment, pp. 31—66. In V. A. Drake & A. 6. Gatehouse (eds.), Insect Migration: Tracking Resources Through Space and Time.Cambridge University Press.Johnson, S. J. 6 L. J. Mason. 1986. The noctuidae: A case history, pp. 421-433. In D. R. Mackenzie, c. S. Barfield,G.G. Kennedy & R. D. Berger (eds.), Movement and Dispersal of Agriculturally Important Biotic Agents. Claitors Publishing. Baton Rouge, LA.Jones, M. T. 1985. Pest management and the vegetable farmer in Trinidad and Tobago, pp. 173-175. In Proceedings of the Caribbean Food Crops Society.Kao, c. H., C. F. Hung 6 C. M. Sun. 1989. Parathion and methyl parathion resistance in diamondback moth (Lepidoptera: Plutellidae) larvae. J. Econ. Entomol. 85: 1299-1304.Kennedy, G. G. 6 M. A. Whalon. 1995. Managing pest resistance to Bacillus thuringiensis endotoxins: Constraints and incentives to implementation. J. Econ. Entomol. 88: 454- 460.Kirsch, K. 6 J. Schmutterer. 1988. Low efficacy of a Bacillus thuringiensis (Ber.) formulation in controlling the diamondback moth, Plutella xylostella (L.), in the Philippines. J. Appl. Entomol. 105: 249-255.Kogan, M. 1982. Plant resistance in pest management, pp. 93-134. In R. L. Metcalf & W. H. Luckmann (eds.), Introduction to Pest Management. Wiley, NY.LCES. 1996. Commercial vegetable production recommendations. Louisiana Cooperative Extension Service, Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station. Baton Rouge, LA.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

185
LCES. 1997. Insect control guide. Louisiana Cooperative Extension Service, Louisiana State University Agricultural Center, Louisiana Agricultural Experiment Station. Baton Rouge, LA.Leibee, 6. L. S K. E. Savage. 1992. Insecticide resistance in diamondback moth in Florida, pp. 427-435. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Lim, 6. S. 1982. The biology and effects of parasites on the diamondback moth, Plutella xylostella (L.). Ph.D. Dissertation, University of London.Lim, 6. S. 1986. Biological control of diamondback moth, pp. 159-171. In N. S. Talekar & T. D.. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Lim, G. S. 1992. Integrated pest management of diamondback moth: Practical realities, pp. 565-576. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Lim, G. S., A. Sivapragasam 8 M. Ruvaida. 1986. Impact assessment of Apanteles plutellae on diamondback moth using an insecticide-check method, pp. 195-205. Jn N. s. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Liu, Y. B., B. E. Tabashnik 6 M. Pusztai-Carey. 1996.Field-evolved resistance to Bacillus thuringiensis toxin CrylC in diamondback moth (Lepidoptera: Plutellidae). J. Econ. Entomol. 89: 798-804.Luck, R. F., R. Van dan Bosch 6 R. Garcia. 1977. Chemical insect control - a troubled management strategy. BioScience 27: 606-611.Luckmann, W. H. 6 R. L. Metcalf. 1994. The pest-management concept, pp. 1-31. In R. L. Metcalf & W. H. Luckmann (eds.), Introduction to Insect Pest Management. John Wiley & Sons, Inc., NY.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

186
Luthy, P., J. Cordier & H. Fisher. 1982. Bacillus thurincfiensis as a bacterial insecticide: Basicconsiderations and application, pp. 35-208. In E. Kurstak (ed.), Microbial and Viral Pesticides. Marcel Dekker, Inc., NY.Maa, V. C. i M. L. Chuang. 1983. Esterase of diamondback moth, (Plutella xylostella) (L.). I. Enzymatic properties of larval esterases. Bull. Inst. Zool. Acad. Sinica. 22: 123- 131.Mackenzie, J. M. B. 1958. Invasion of diamondback moth (Plutella maculipennis Curtis). J. Entomol. Sci. 91: 247- 250.Magallona, E. D. 1986. Developments in diamondback moth management in the Philippines, pp. 423-435. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Magaro, J. J. & J. V. Edelson. 1990. Diamondback moth (Lepidoptera: Plutellidae in south Texas: A technique for resistance monitoring in the field. J. Econ. Entomol. 83: 1201-1206.Maltais, P. M., J. R. Muckle 6 P. ▼. LeBlanc. 1994.Economic threshold for management of lepidoptera larvae on broccoli in southeastern New Brunswick. J. Econ. Entomol. 87: 766-774.Marsh, H. O. 1917. Life history of Plutella maculipennis, the diamondback moth. J. Agric. Res. 10: 1-10.Martin, F. A., C. A. Richard 6 8. D. Hensley. 1975. Host resistance to Diatraea saccharalis (F.): Relationship ofsugarcane internode hardness to larval density. Environ. Entomol. 4: 687-688.Matteson, P. 1995. The "50% Pesticide Cuts" in Europe: A glimpse of our future? American Entmologist 41-210-220.May, R. M. 1993. Resisting resistance. Nature 361: 593-594.McClure, 8. 1994a. Companion planting for pest control, pp.22-33. In S. McClure & S. Roth (eds.), Rodale's Successful
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

187
Organic Gardening: Companion Planting. Rodale Press, Emmaus, PA.McClure, 8. 1994b. Introduction and companion plantingbasics, pp. 9-21. In S. McClure £ S. Roth (eds.), Rodale's Successful Organic Gardening: Companion Planting. Rodale Press, Emmaus, PA.McGaughey, W. H. 1985. Insect resistance to the biological insecticide Bacillus thuringiensis. Science 229: 193-195.McGaughey, W. H. £ M. E. Whalon. 1992. Managing insectresistance to Bacillus thuringiensis toxins. Science 258: 1451-1455.McHugh, J. J. £ R. E. Foster. 1995. Reduction of diamondback moth (Lepidoptera: Plutellidae) infestation in head cabbage by overhead irrigation. J. Econ. Entomol. 88: 162-168.Metcalf, R. L. 1980. Changing role of insecticides in crop protection. Annu. Rev. Entomol. 25: 219-256.Metcalf, R. L. £ V. H. Luckmann. 1994. Introduction toInsect Pest Management. John Wiley & Sons, Inc., New York.Metcalf, R. L. £ J. J. McKelvey, Jr. 1976. The Future of Insecticides. John Wiley & Sons, Inc., NY.Mineau, P. £ B. T. Collins. 1988. Avian mortality inagroecosystems: Methods of detection, pp. 3-12. Iji M. P. Greaves, B. D. Smith & P. W. Greig-Smith (eds.), FieldMethods for the Study of Environmental Effects of Pesticides. Proceedings of the British Crop Protection Council.Cambridge, UK.Mitchell, E. R., G. Y. Hu £ J. 8. Okine. 1997. Diamondback moth (Lepidoptera: Plutellidae) infestation and parasitism by Diadegma insulare (Hymenoptera: Ichneumonidae) in collards and adjacent cabbage fields. Florida Entomol. 80: 54-62.Miyata, T., T. Saito £ V. Noppun. 1986. Studies on the mechanism of diamondback moth resistance to insecticides, pp. 347-357. In N. S. Talekar £ T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

188
Moore, A. SB. B. Tabashnik. 1989. Leg autonomy of adult diamondback moth (Lepidoptera: Plutellidae) in reponse to tarsal contact with insecticide residues. J. Econ. Entomol. 82: 381—384.Moore, M. 1994. Pesticide use reduction: An emerging trend in state policy. Sustainable Newsletter 6: 1-3.Moriuti, S. 1986. Taxonomic notes on the diamondback moth, pp. 83-88. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Mussen, E. 1990. California crop pollination. Gleanings in Bee Culture. 118: 646-647.Mustata, G. 1992. Role of parasitoid complex in limiting the population of diamondback moth in Moldavia, Romania, pp. 203-211. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Navon, A. 1993. Control of lepidopteran pests with Bacillus thuringiensis, pp. 125-146. In P. F. Entwistle, J. S. Cory, M. J. Bailey & S. Higgs (eds.), Bacillus thuringiensis, an Environmental Biopesticide: Theory and Practice. John Wiley & Sons, Inc., New York.NBIAP. 1995. Special issue on Bt. National Biological Impact Assessment Program, News Report, December 1995.NRC. 1986. Pesticide Resistance: Strategies and Tactics for Management. National Research Council, National Academy Press. Washington, D.C.NRC. 1996. Ecologically Based Pest Management: NewSolutions for a New Century. National Research Council, National Academy Press. Washington, D.C.Ooi, P. A. C. 1986. Diamondback moth in Malaysia, pp. 25- 35. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Ooi, P. A. C. 1992. Role of parasitoids in managing diamondback moth in the Cameron Highlands, Malaysia, pp. 255-
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

189
262. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.OTA. 1995. Biologically Based Technologies for Pest Control. OTA-ENV-636. Office of Technology Assessment. Washington, D.C.Painter, R. H. 1951. Insect Resistance in Crop Plants.Macmillan, New York.Painter, R. H. 1958. Resistance of plants to insects.Annu. Rev. Entomol. 3: 267-290.Pashley, D. P., S. J. Johnson & A. N. Sparks. 1985. Genetic population structure of migratory moths: The fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 78: 756- 762.Peres, C. J. 8 A. M. Shelton. 1997. Resistance of Plutella xylostella (Lepidoptera: Plutellidae) to Bacillusthuringiensis Berliner in Central America. J. Econ. Entomol. 90: 87-93.Picha, D. H. 5 R. A. Hinson. 1996. The commercial vegetableindustry in Louisiana, pp. 6-8. In J. Tarver & J. Lord(eds.), Louisiana Agriculture. Vol. 39, No. 3.Pimentel, D. 1961. Species diversity and insect population outbreaks. Prot. Ecol. 2: 77-114.Pimentel, D., J. Krummel, D. Gallahan, J. Hough, A. Merrill, I. Schreiner, P. Vittum, F. Kosiol, E. Beck, D. Yen 6 s. Fiancd. 1978. Benefits and costs of pesticide use in U.S. food production. BioScience 28: 778-784.Pimentel, D., D. Andow, R. Dyson-Hudson, D. Gallahan, S. Jacobsen, M. Irish, S. Kroop, A. Moss, I. Schreiner, M. Shepard, T. Thompson 6 B. Vinzant. 1980. Environmental and social costs of pesticides: A preliminary assessment. Oikos 34: 126-40.Pimentel, D., H. Acquay, M. Biltonen, P. rice, M. Silva, J. Nelson, V. Lipner, S. Giordano, A. Horowitz 6 M. D'Amore.1992. Environmental and economic costs of pesticide use. BioScience 42: 750-760.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

190
Pimentel, D. 8 H. Lehman («ds.). 1993. The pesticidequestion: Environment, economics and ethics. Chapman & Hall.NY.Plapp, F. W., Jr., J. J. Magaro 8 j. v. Edelson. 1992.Diamondback moth in south Texas: A technique for resistance monitoring in the field, pp. 443-446. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.PN. 1990. Towards a reduction in pesticide use. Pesticide News, March 1990.Pollard, 6. V. 1991. Constraints to IPM development and a strategy for management of tomato and cabbage pests in Trinidad, West Indies. Tropical Pest Management 37: 59-62.Powers, N. J. & R. 6. Heifner. 1993. Federal grade standards for fresh produce: Linkages to pesticide use.Agriculture Information Bulletin No. 675, August 1993. United States Department of Agriculture, Beltsville, MD.Price, P. W. 1984. Insect Ecology. John Wiley & Sons, New York.Prokopy, R. J. & E. D. Ovens. 1983. Visual detection of plants by herbivorous insects. Annu. Rev. Entomol. 28: 337- 364.Rankin, M. A. 8 C. A. Burchsted. 1992. The cost of migration in insects. Annu. Rev. entomol. 37: 533-559.Reus, J. A. W. A., H. J. Wekseler 8 6. A. Pak. 1994.Towards a future EC pesticide policy: An inventory of risks of pesticide use, possible solutions and policy instruments. Center for Agriculture and Environment. Utrecht, The Netherlands.Riggin-Bucci, T. 8 F. Gould. 1997. Impact of intraplot mixtures of toxic and nontoxic plants on population dynamics of diamondback moth (Lepidoptera: Plutellidae) and itsnatural enemies. J. Econ. Entomol. 90: 241-251.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

191
Rischf S. J. D. Adov £ M. A. Altiari. 1983. Agroecosystem diversity and pest control: Data, tentative conclusions and new research directions. Environ. Entomol. 12: 625-629.Robertson/ P. L. 1939. Diamondback moth investigation in New Zealand. N. Z. J. Sci. Technol. 20: 330-364.Robinson, D. E., K. K. Dalip £ A. Mansingh. 1995.Integrated Management of Pests and Pesticides in theCaribbean. Department of Zoology, University of the WestIndies, Mona, Kingston. The Jamaica National Commission forUNESCO. Kingston.Rosario, C. £ C. crus. 1986. Life cycle of diamondback, Plutella xylostella (L.), (Lepidoptera: Plutellidae) inPuerto Rico. J. Agric. Of the University of Puerto Rico. 70: 229-234.Roush, R. T. £ B. E. Tabashnik (eds.). 1990. PesticideResistance in Arthropods. Chapman £ Hall, NY.Sagenmuller, A. £ E. Rose. 1986. Hoe 522 (CME 134) , a new insect growth regulator for control of diamondback moth, pp. 271-278. In N. S. Talekar £ T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Salinas, P. J. 1972. Studies on the ecology and behavior of the larvae of Plutella xylostella (Linnaeus) (Lepidoptera: Plutellidae). Ph.D. Dissertation, University of London.Salinas, P. J. 1986. Studies on diamondback moth in Venezuela with reference to other Latin American countries, pp. 17-24. In N. S. Talekar £ T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.SAS Institute. 1989. SAS/STAT user's guide: Statistics, 5ch edition. SAS Institute, Cary, NC.
Shelton, A. M., J. T. Andaloro fi j. Barnard. 1982. Effects of cabbage looper, imported cabbage worm, and diamondback moth on fresh market and processing cabbage. J. Econ. Entomol. 75: 742-745.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

192
Shelton, A. M., M. K. Sears, J. A. Wynan & T. C. Quick. 1983. Comparison of action thresholds for lepidopterous larvae on fresh market cabbage. J. Econ. Entomol. 76: 196- 199.Shelton, A. M., C. W. Soy, R. C. North, M. H. Dickson, 8 J. Barnard. 1988. Analysis of resistance in cabbage varieties to damage by Lepidoptera and Thysanoptera. J. Econ. Entomol. 81: 634-640.Shelton, A. M. 8 J. A. Wynan. 1992. Insecticide resistance of diamondback moth (Lepidoptera: Plutellidae) in NorthAmerica, pp. 447-454. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Shelton, A. M., J. A. Wyman, N. L. Cushing, K. Apfelbeck, T. J. Dennehy, S. E. R. Mahr, 8 s. D. Eigenbrode. 1993.Insecticide resistance of diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae), in North America. J.Econ. Entomol. 86: 11-19.Shelton, A. M., J. L. Robertson, J. D. Tang, c. Perez, s. D. Eigenbrode, H. K. Preisler, w. T. wilsey 8 R. j. Cooley.1993. Resistance of diamondback moth (Lepidoptera:Plutellidae) to Bacillus thuringiensis subspecies in the field. J. Econ. Entomol. 86: 697-705.Shorey, H. H., L. A. Andres 6 R. L. Hale. 1962. The biology of Trihoplusia ni. 1. Life history and behavior. Ann. Entomol. Soc. Am. 5: 592-597.Shorey, H. H. 1963. The biology of Trichoplusia ni(Lepidoptera: Noctuidae) . 111. Response to the ovipositionsubstrate. Ann. Entomol. Soc. Am. 56: 165-170.Sivapragasaa, A. Y. Ito 8 T. Saito. 1988. Population fluctuations of the diamondback moth, Plutella xylostella (L.) on cabbages in Bacillus thuringiensis sprayed and nonsprayed plots and factors affecting within-generation survival of immatures. Res. Popul. Ecol. 30: 329-342.Smith, R. F. 8 H. T. Reynolds. 1966. Principles,definitions and scope of integrated pest control, pp. 11-17. In Proceedings, Symposium on Integrated Pest Control, Food & Agrculture Organization, Rome.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

193
Smith, D. B. t M. K. Sears. 1982. Evidence for dispersal of diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae) , in southern Ontario. Proc. Entomol. Soc. Ont. 113: 21-28.Sorenson, X. X. 1993. IPM and growers: An evolution in thinking, pp. 129-150. In A. R. Leslie & 6. W. Cuperus (eds.), Successful Implementation of Integrated Pest Management for Agricultural Crops. Lewis Publishers, Boca Raton, FL.Sorenson, X. X. 1994. The role of IPM in reducingenvironmental problems, pp. 17-24. In Proceedings, Second National Integrated Pest Management Symposium/Workshop. ESCOP Pest Management Strategies Subcommittee and ES IPM TaskForce. University Graphics, North Carolina State University, Raleigh, NC.Southvood, T. R. E. 1962. Migration of terrestrial arthropods in relation to habitat. Biol. Rev. 37: 26-32.Srinvasan, K. t P. N. K. Moor thy. 1991. Indian mustard asa trap crop for management of major lepidopterous pests on cabbage. Tropical pest Management 37: 189-196.Stinner, R. E., c. S. Barfield, J. L. Stimac 6 L. Dohse. 1983. Dispersal and movement of insect pests. Annu. Rev. Entomol. 28: 319-335.Stoner, K. X. 1990. Glossy leaf wax and plant resistance to insects in Brassica oleracea under natural infestation.Environ. Entomol. 19: 730-739.Stringer, X. 6 C. Lyons. 1974. The effect of benomyl and thiophanate-methyl on earthworm populations in apple orchard. Pesti. Sci. 5: 189-196.Suah, J. R. & D. W. Ellis. 1974. Insecticide control ofcabbage caterpillars in Jamaica, pp. 263-66. In c. W. D.Braithwaite, R. H. Phelps & F. D. Bennett (eds.), Proceedings of Protection of Horticultural Crops in the Caribbean, University of the West Indies, St. Augustine, Trinidad & Tobago.Sun, C. N. 1992. Insecticide resistance in diamondback moth, pp. 419-426. In N. S. Talekar (ed.), Diamondback Moth
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

194
and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Sun, C. N., H. Chi & H. T. Feng. 1978. Diamondback resistance to diazinon and methomyl in Taiwan. J. Econ. Entomol. 71: 551-554.Sun, C. N., T. K. Wu, J. S. Chen 8 W. T. Lee. 1986.Insecticide resistance in diamondback moth, pp. 359-371. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Sutherland, D. W. s. 1966. Biological investigations of Trichoplusia ni (Hubner) and other lepidoptera damaging cruciferous crops on Long Island, New York. Memoir 399, Cornell University Agricultural Experiment Station, New York State College of Agriculture, Ithaca, NY.Syed, A. R. 1992. Insecticide resistance in diamondback moth in Malaysia, pp. 437-442. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests. Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Tabashnik, B. E. 1994. Evolution of resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 39: 47-79.Tabashnik, B. E., M. L. Cushing, N. Finson 6 M. W. Johnson. 1990. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae) . J. Econ. Entomol. 83: 1671-1676.Tabashnik, B. S., N. Finson 6 M. W. Johnson. 1991. Managing resistance to Bacillus thuringiensis: Lessons from thediamondback moth (Lepidoptera: Plutellidae). J. Econ.Entomol. 84: 49-55.Tabashnik, B. E., J. M. Schwarts, N. Finson 6 M. w. Johnson. 1992. Inheritance of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J. Econ.Entomol. 85: 1046-1055.Tabashnik, B. E., M. Finson, C. F. Chi1cut, N. L. Cushing 6 M. W. Johnson. 1993. Increasing efficiency of bioassays: Evaluating resistance to Bacillus thuringiensis in
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

195
diamondback moth (Lepidoptera: Plutellidae). J. Econ.Entomol. 86: 635-644.Talekar, N. 8. (ed.). 1992. Diamondback Moth and otherCrucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Talekar, N. 8. fi T. D. Griggs (eds). 1986. Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Talekar, N. 8., T. 8. Lee 6 8. W. Huang. 1986.Intercropping and modification of irrigation method for the control of diamondback moth, pp. 145-155. In N. S. Talekar & T. D. Griggs (eds.), Diamondback Moth Management, Proceedings of the First International Workshop. AVRDC, Shanhua, Taiwan.Talekar, N. S., J. C. Yang 6 S. T. Lee. 1990. Annotated Bibliography of Diamondback Moth, vol 2. AVRDC, Shanhua, Taiwan.Talekar, N. S., J. C. Yang 6 8. T. Lee. 1992. Introduction of Diadegma semiclausum to control diamondback moth in Taiwan, pp. 263-270. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Talekar, N. S. 6 A. M. Shelton. 1993. Biology, ecology and management of the diamondback moth. Annu. Rev. Entomol. 38: 275-301.Tang, J. D., 8. Gilboa, R. T. Roush 6 A. M. Shelton. 1997.Inheritance, stability, and lack-of-fitness costs of field- selected resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae) from Florida. J. Econ.Entomol. 90: 732-741.Tanton, M. 1962. The effect of leaf "toughness" on the feeding of larvae of the mustard beetle, Phaedon cochleriae F. Ent. Exp. &Appl. 5: 74-78.Thomas, V. P. 6 A. M. Ferguson. 1989. Plutella xylostella (L.), diamondback moth (Lepidoptera: Yponomeutidae), pp. 135-
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

196
142. In P. J. Cameron, R. L. Hill, J. Bain & W. P. Thomas (eds.), A Review of Biological Control of Invertebrate Pests and Heeds in New Zealand 1874-1987. CAB International, Wallingford, OK.Thorsteinson, A. J. 1953. The chemotactic responses that determine host specificity in an oligophagous insect (Plutella maculipennis) [Curt.] Lepidoptera). Can. J. Zool. 31: 52-72.Trumble, J. T. 1990. Vegetable insect control with minimal use of insecticides. HortScience 25: 159-164.USBC. 1990. Statistical Abstract of the United States, U.S. Bureau of Census, U.S. Department of Commerce. Washington, D.C.USDA. 1945. United States standards for grades of cabbage. 18 Federal Register 7084 (1953), redesignated 46 FR 63203(1981). United States Department of Agriculture, Beltsville, MD.USDA. 1994. Three-pronged attack on crop pests.Agricultural Research Service, Agricultural Research, July1994. United states Department of Agriculture, Beltsville, MD.Van Rie, J., W. H. McGaughey, D. E. Johnson, B. D. Barnett 5H. Van Mellaert. 1990. Mechanisms of insect resistance to the microbial insecticide Bacillus thuringiensis. Science 247: 72-74.Verschaffelt, E. 1910. The cause determining the selection of food in some herbivorous insects. Proc. Amsterdam Acad. Sci. 13: 536-542.Waage, J. 1992. Summary, pp. 577-579. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Waage, J. £ A. Cherry. 1992. Quantifying the impact of parasitoids on diamondback moth, pp. 245-253. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

197
Walton# W. C. 1989. Problems associated with the control of diamondback moth in Douglas Castle, St. Ann. Department of Crop Science, Faculty of Agriculture, University of the West Indies, St. Augustine, Trinidad.Ware# G. W. 1988. Complete Guide to Pest Control. Thompson Publications, Fresno, CA.Wearing# C. H. 1988. Evaluating the IPM implementation process. Annu. Rev. Entomol. 33: 17-38.Wolf# W. W.# A. N. Sparks, S. D. Pair# J. K. Westbrook 8 F. M. Truesdale. 1986. Radar observations and collections of insects in the Gulf of Mexico, pp. 221-234. In W. Danthanarayana (ed.), Insect Flight: Dispersal and Migration. Springer-Verlag, New York.Workman# R. B. R. B. chalfant 6 D. j. Schuster. 1980.Management of the cabbage looper and diamondback moth on cabbage using two damage thresholds and five insecticide treatments. J. Econ. Entomol. 73: 757-758.Wyman# J. A. 1992. Management approaches for cruciferous insect pests in central North America, pp. 503-509. In N. S. Talekar (ed.), Diamondback Moth and Other Crucifer Pests, Proceedings of the Second International Workshop. AVRDC, Tainan, Taiwan.Yandia# A. 1989. Host plant resistance to the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae), M.S. Thesis, Louisiana State University, Baton Rouge.Yao# M. G.# C. F. Hung 6 C. N. Sun. 1988. Fenvalerate resistance and aldrin epoxidation in larvae of the diamondback moth. Pestic. Biochem. Physiol. 30: 272-278.Yaseem# M. 1978. The establishment of two parasites of the diamondback moth Plutella xylostella (Lep: Plutellidae) in Trinidad, W.I. Entomophaga 23: 111-114.Young# J. R. 1979. Assessing the movement of the fall armyworm (Spodoptera frugiperda) using insecticide resistance and wind patterns, pp. 344-351. In R. L Rabb & G. G. Kennedy (eds.), Movement of Highly Mobile Insects: Concepts andMethodology in Research. University Graphics, North Carolina State University, Raleigh. NC.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

198
Zimmerman, L. 1978. Strange bedfellows: The voodoo and science of companion planting, pp. 73-77. In J. Lawrence (ed.), The Harrowsmith Reader. Camden Publishing Ltd., Canada.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

VitaPaul W. Ivey was born in Ewarton, St. Catherine,
Jamaica, to Harold and Enid Ivey on May 20, in the early 1960s. He graduated from Ewarton Secondary School in June 1980, and then participated in the National Youth Service Corps until August 1981. He then entered Passley Gardens Agricultural High School in Port Antonio, Portland, Jamaica. In 1982, he transferred to Elim Agricultural School where he graduated in 1983. After a short stint at Ewarton Secondary School as an Agricultural Science Teacher, he enrolled at the College of Agriculture, Port Antonio, Portland, Jamaica, in 1984; he graduated in 1987 with an Associate of Science degree in Agriculture. He then went on to the University of the West Indies, St. Augustine, Trinidad, where he obtained a Bachelor of Science degree, with honors, in Agriculture in 1989. He next entered the Department of Entomology at Louisiana State University in the fall semester of 1989; he received a Master of Science degree in Entomology from L.S.U. in 1991. From October 1991 to August 1994, he was employed as a Lecturer in Entomology and Crop Production, and later Head, Department of Plant, Soil Sciences and Engineering at the College of Agriculture. He reentered the Department of Entomology at Louisiana State University on a graduate
199
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

200
assistantship in the fall of 1994. Since that time, to obtain his doctoral degree, he has been conducting research on integrated pest management tactics for managing the diamondback moth, Plutella xylostella (L.) and the cabbage looper, Trichoplusia ni (Hubner) - the major lepidopteran pests of cabbage - under the able supervision of Professor Seth J. Johnson. He will receive his degree of Doctor of Philosophy in December 1997.
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.

DOCTORAL EXAMINATION AND DISSERTATION REPORT
C a n d i d a t e i
M a j o r F i e l d :
T i t l e o f D i s s e r t a t i o n :
D a t e o f E x a m i n a t i o n :
July 31. 1997
P a u l W . I v e y
E n to m o lo g y
I n t e g r a t e d P e s t M a n a g e m e n t T a c t i c s f o r C o n t r o l o f C a b b a g e L o o p e r a n d D ia m o n d b a c k M o th i n C a b b a g e
A p p r o v e d :
M a j o r Pro:
ite S c h o o l
EXAMINING COMMITTEE:
Reproduced with permission of the copyright owner. Further reproduction prohibited without permission.