influence readily metabolizable carbon on ...aem.asm.org/content/54/10/2452.full.pdfactivity...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, OCt. 1988, p. 2452-2459 Vol. 54, No. 100099-2240/88/102452-08$02.00/0Copyright © 1988, American Society for Microbiology
Influence of Readily Metabolizable Carbon on PentachlorophenolMetabolism by a Pentachlorophenol-Degrading Flavobacterium sp.
EDWARD TOPP,lt* RONALD L. CRAWFORD,2 AND RICHARD S. HANSON1
Gray Freshwater Biological Institute and Department of Microbiology, University of Minnesota, P.O. Box 100, Navarre,Minnesota 55392,1 and Department ofBacteriology and Biochemistry, University of Idaho, Moscow, Idaho 838432
Received 28 March 1988/Accepted 13 July 1988
The influence of high concentrations of pentachlorophenol (PCP) and readily metabolizable carbon on theactivity and viability of a PCP-degrading Flavobacterium sp. was examined in a mineral salts medium. Lagspreceding PCP removal by glutamate-grown Flavobacterium cells were greatly attenuated by the addition ofglutamate, aspartate, succinate, acetate, glucose, or cellobiose. The effect of these supplementary carbonsources on the apparent lag was not mediated entirely through the stimulation of growth since PCP metabolismaccompanied the onset of growth. The specific activity of PCP-degrading cells in the absence of supplementarycarbon was 1.51 x ±-13± 0.08 x i0-'3 g of PCP per cell per h and in the presence of supplementary carbonwas 0.92 x 10-13 + 0.09 x 10-13 g of PCP per cell per h. Glutamate in combination with glucose or cellobiosepartially repressed PCP metabolism. PCP removal by PCP-induced, glutamate-grown cells suspended in thepresence of 4 g of sodium glutamate per liter was sensitive to shock loads of PCP, with a Kg of about 86.8 ,ug/ml. Subsequent removal rates, however, were more resistant to PCP. Optimal stimulation of PCP removal bysodium glutamate required 3.0 g/liter, about the same concentration as that which saturated growth in theabsence of PCP. PCP removal rates decayed within minutes following the transfer of PCP-induced,glutamate-grown cells to media containing PCP without supplementary carbon, and increasing PCP concen-trations accelerated the decay. The addition of glutamate to PCP-induced, glutamate-grown cells followingtheir incubation with PCP as the sole carbon source for 1 h promoted full recovery of activity. Theglutamate-mediated recovery was completely blocked by chloramphenicol. These data indicate that, regardlessof prior induction of PCP metabolism, the transfer of Flavobacterium cells from an environment containingreadily metabolizable carbon to one containing PCP as the sole carbon source resulted in cellular damage,death, and, consequently, long lags preceding PCP removal. We suggest that available carbon in pollutedenvironments could facilitate PCP removal by inoculated Flavobacterium cells by attenuating the toxicity ofPCP and contributing to the production and maintenance of PCP-degrading biomass.
A Flavobacterium sp. (20, 21) is one of several bacterialisolates that grow on and mineralize pentachlorophenol(PCP) (1, 4, 10, 23, 25, 30, 32). PCP is a recalcitrant biocideused primarily for wood preservation (7), is ubiquitous in theenvironment (22), and is considered a priority toxic pollutantby the U.S. Environmental Protection Agency (24). Theenhancement of biodegradation of toxic organic materialssuch as chlorinated aromatic compounds for the purpose ofdecontaminating polluted environments is of considerableinterest (26). For example, soils (6) and natural waters (20)containing high PCP concentrations are rapidly decontami-nated following inoculation with large numbers of this Fla-vobacterium sp. Similarly, a PCP-degrading Arthrobactersp. rapidly decontaminated PCP-containing soil (11), as didPseudomonas cepacia AC1100 when added to soil contain-ing 2,4,5-trichlorophenoxyacetic acid (17).There are numerous physical and chemical parameters
that influence the kinetics of PCP metabolism by this Flavo-bacterium sp. in soils and natural waters (6, 20). Oneparameter that may be of significance is the presence ofreadily metabolizable carbon. Through a variety of mecha-nisms, the metabolism of one compound may often beenhanced or inhibited by that of a second compound (14).Soil and water polluted with a toxic organic compound cansometimes contain appreciable concentrations of innocuous
* Corresponding author.t Present address: Land Resource Research Centre, Research
Branch, Agriculture Canada, Ottawa, Ontario KlA OC6, Canada.
carbon sources. PCP-contaminated wastewater streams pol-luted by wood-preserving operations contained 800 to 24,000mg per liter of biological oxygen demand (31). PCP degra-dation rates in several soils correlated with the organicmatter content (19). Degradation of 2,4-dichlorophenoxya-cetic acid or 3,5-dichlorobenzoic acid by acclimated aerobicenrichment cultures was stimulated by supplementation withnutrient broth (18). We have observed that the addition ofsupplementary carbon to soil heavily contaminated withPCP and inoculated with Flavobacterium cells can, in somecases, accelerate decontamination (data to be publishedelsewhere). We therefore examined the interactions of PCPand alternate carbon sources on the survival and activities ofFlavobacterium cells in axenic culture. We report here theeffects of supplementary carbon on the resistance to, andremoval of, high concentrations of PCP.
MATERIALS AND METHODS
Growth of Flavobacterium cells in batch culture. The organ-ism used was that isolated by Pignatello et al. (21), describedby Martinson et al. (20), and used in the studies of Steiertand Crawford (27) and Steiert et al. (28). Unless otherwiseindicated, cells were routinely grown with agitation in Er-lenmeyer flasks at 30°C in mineral salts (MS) medium with 4g of L-sodium glutamate per liter as the carbon source andinduced for PCP metabolism by the addition of 50 ,ug of PCPper ml in the mid-logarithmic phase of growth (27). Cells
2452
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

PCP-DEGRADING FLAVOBACTERIUM SP. 2453
were harvested by centrifugation (12,000 x g, 10 min), andthe pellet was washed and suspended in the appropriatemedium. Contamination was routinely checked for by plat-ing on nutrient agar. The MS medium was modified by theaddition of 0.25 g of NH4Cl per liter (MS-NH4+ medium)when the carbon source was not an amino acid.Growth of Flavobacterium cells on PCP in continuous cul-
ture. Cells were grown in a Bioflo model C30 chemostat(New Brunswick Scientific Co., Inc., Edison, N.J.); themedium feed pump was replaced with a Minipuls II peristal-tic pump (Gilson Medical Electronics, Inc., Middleton,Wis.). The pH was held at 7.1 by means of a New Brunswickmodel pH-40 pH controller and pump module delivering 0.2N NaOH. The working volume was 365 ml, the temperaturewas held at 30°C, the agitation rate was 300 rpm, the air flowrate was 120 ml/h, and the dilution rate was 0.026 h-1. Forcarbon-limited growth with PCP as the sole carbon source,the medium composition was as follows (in milligrams perliter): KH2PO4, 630; K2HPO4, 190; MgSO4 7H20, 200;FeSO4 6H20, 5; (NH4)2SO4, 250; and PCP, 250. The me-dium in the carboy was shielded with aluminum foil. Mediumwas prepared with ultrapure H20 (Millipore Q system). Allinorganic constituents were autoclaved together, except theFe salt, which was added in the form of a filter-sterilizedsolution to the cooled medium. PCP was added from afilter-sterilized stock solution.
Experimental incubations. Unless otherwise indicated, allincubations were in MS-NH4+ medium incubated with agi-tation in Erlenmeyer flasks at 30°C. Supplementary carbonsources were added from filter-sterilized stock solutions orwere autoclaved directly in MS medium, with the exceptionof carbohydrates, which were autoclaved separately. Or-ganic acids were neutralized with NaOH when required.Chlorinated aromatic compounds were added from filter-sterilized stock solutions (Tuffryn polysulfone membranes,Acrodisc; Gelman Sciences, Inc., Ann Arbor, Mich.). Sig-nificant binding of PCP to nylon filters was observed.Growth was monitored by A60 or colony counts of seriallydiluted samples plated on full-strength nutrient agar. Con-centrations of PCP and 2,3,5,6-tetrachlorophenol were de-termined by A318; concentrations of 2,4,6-trichlorophenoland 2,3,4,6-tetrachlorophenol were determined by A312. Cellsuspensions were diluted 1:1 with 0.1 N NaOH immediatelyfollowing sampling so that all samples had a uniformly highpH and to stop metabolic activity. The diluted samples werethen centrifuged (Eppendorf microcentrifuge; 5 min at15,600 x g), and the absorbance of the supernatant wasdetermined. Chloramphenicol was added from a concen-trated stock solution prepared in 95% ethanol. Where re-quired, spectrophotometric PCP determinations were cor-rected for the absorbance of chloramphenicol. Allincubations were done in duplicate, and all experimentswere repeated at least twice.
Calculation of the substrate inhibition constant for PCP.The substrate inhibition constant (Ki) for PCP was deter-mined on the basis of the Haldane function by using amodification of the method of Jones et al. (16). The relation-ship between PCP concentration and initial metabolism ratewas derived by plotting the reciprocal of the initial velocity(0 to 30 min) against the mean PCP concentration (0 and 30min determinations) and determining the x intercept whichcorresponds to -Ki (16).
Calculation of the specific activities of PCP metabolism.Per-cell specific activities ofPCP metabolism were estimatedwith data from several batch culture experiments. The initialPCP concentrations were in the range of 100 to 150 ,ug/ml,
150
100
50
0
A
I zA
UA
U
0
-60 0 60 120
S
S
A
S U
0 21
HOURS
FIG. 1. Removal of PCP by induced Flavobacterium cells sus-pended in the presence of 4 g of sodium glutamate per liter plusvarious PCP concentrations. Linear regression analysis of the datain the inset revealed the Ki in this experiment to be 93.2 p.g/ml. Cellswere suspended to an initial optical density of 0.88 and incubatedwith sodium glutamate 10 min prior to the addition of PCP.
and the initial cell densities were in the range of 107 to 108cells per ml. The rate of PCP metabolism within a specifiedincubation period (e.g., 5 to 15 h) was divided by the mean ofthe viable cell count determined at the beginning and end ofthe incubation period.
RESULTS
PCP was inhibitory to its metabolism by PCP-inducedcells. Induced cells suspended in MS medium containing 4 gof sodium glutamate per liter and various PCP concentra-tions ranging from 70 to 150 ,ug/ml removed PCP rapidly(Fig. 1). The initial rate of PCP removal was inhibited byincreasing PCP concentrations with a Ki of 86.8 + 4.5 ,ug/ml(mean + standard error of two experiments). The rateaccelerated afterwards until PCP concentrations fell to un-detectable levels. Addition of chloramphenicol (100 Ru/ml) toinduced cells 10 min prior to the start of an incubation withPCP and glutamate did not significantly influence the initialrate of PCP removal but prevented its acceleration duringthe remainder of the incubation (Fig. 2). PCP removal rateswere influenced by the concentrations of both PCP andglutamate (Fig. 1 to 3). The concentration of sodium gluta-mate that was required for optimal stimulation was about thesame as that concentration required for optimal growth inthe absence of PCP, namely, 3 g/liter (16.0 mM) (Fig. 3).PCP removal rates by glutamate-grown, induced cells
decayed rapidly when these cells were shifted from theglutamate-containing medium to one containing only PCP(Fig. 2). The initial rate was more strongly inhibited byhigher PCP concentrations. This decay was reversed by the
VOL. 54, 1988
I
A
A
n
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
0 1 2 3HOURS
FIG. 2. Removal of PCP by glutamate-grown, induced Flavobac-terium cells in the presence of 4 g of sodium glutamate per liter (0),in the presence of sodium glutamate and 100 ,ug of chloramphenicolper ml (0), or without any additional supplements (0). Cells weresuspended to an optical density (600 nm) of 0.92 and incubated for 10min prior to the addition of glutamate and PCP. Treatments notreceiving chloramphenicol were supplemented with the sameamount of 95% ethanol to a final concentration of 4 ,ug/ml.
addition of supplementary carbon (Fig. 4). Full activity wasrecovered about 1.5 h after the addition of 4 g of sodiumglutamate per liter to cells incubated for 1 h with 70 p.g ofPCP per ml as the sole carbon source. Addition of chloram-phenicol at 100 ,ug/ml 10 min before the addition of sodiumglutamate completely blocked recovery.At lower cell densities and temperatures (21°C), supple-
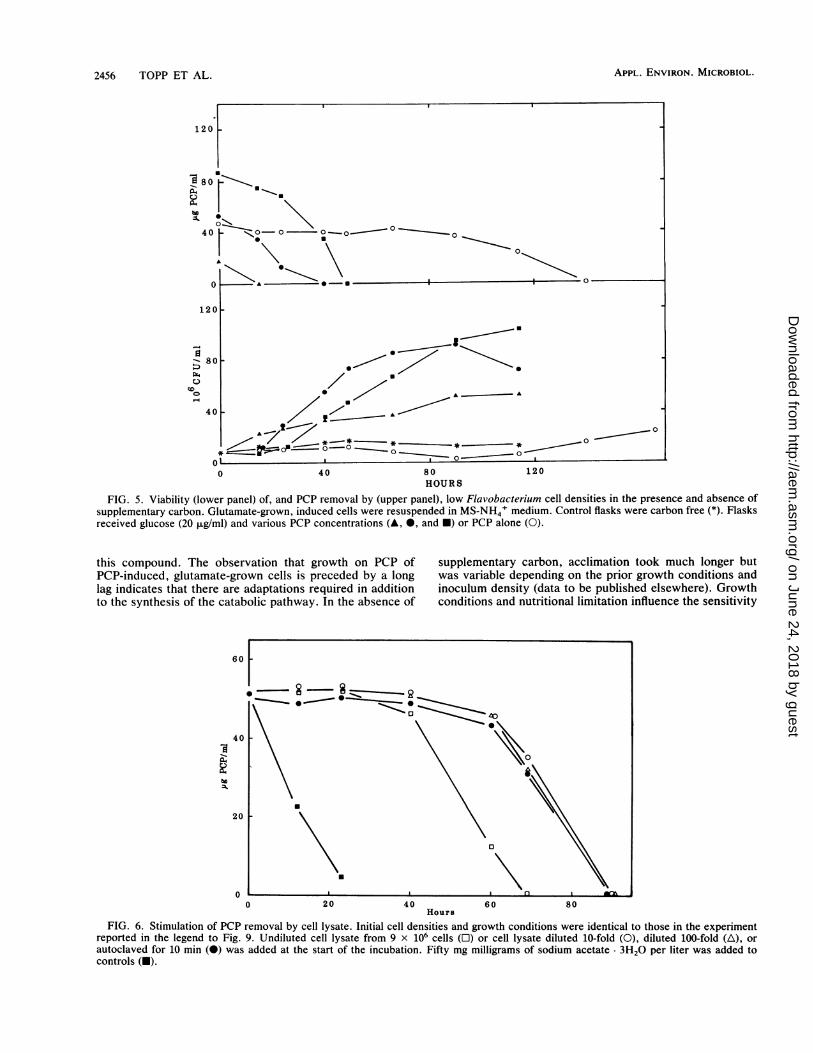
mentary carbon dramatically influenced the kinetics of PCPremoval. Induced, glutamate-grown cells suspended to 107CFU/ml with PCP at 50 ,ug/ml as the only carbon sourcemetabolized PCP but only after a lag of about 90 h, duringwhich about 90% of the cells lost viability (Fig. 5). Growthaccompanied PCP removal after 90 h. The viability of cellssuspended in carbon-free medium remained relatively con-stant during the incubation. In the presence of glucose at 20,ug/ml, PCP at 50 ,ug/ml was removed without any apparentlag. Increasing PCP concentrations caused a longer lag ingrowth, and PCP at 90 ,ug/ml reduced the initial rate of PCPremoval. In both the presence and absence of glucose,growth did not occur until significant PCP removal activitywas effected. As expected, yields were higher when cellsreceived more carbon. Essentially equivalent results werefound when acetate at 20 ,ug/ml was used instead of glucose(data not shown). Prior to acclimation to PCP as the solecarbon source, the viable population was reduced by 90%and the possible stimulatory role of the carbon released bycell lysis was examined. French-pressed, filter-sterilized(0.45-,um pore size; Acrodisc, Gelman) cell lysate from 9 x106 cells, corresponding in number to those dying during the90-h lag period, was added at the start of an incubation withPCP as the sole carbon source (Fig. 6). This additionsignificantly reduced the lag period prior to PCP metabolism.However, the lag period was not reduced if the lysate wasdiluted 10-fold or 100-fold or if it was autoclaved prior tobeing added.
0 2.0 4.0 6.0gNa -GIu/I
FIG. 3. Effect of sodium glutamate concentration on enhance-ment of PCP removal (upper panel) and growth rate in the absenceof PCP (lower panel). The rate of PCP removal catalyzed in the first30 min of incubation by cells incubated with PCP (65 p.g/ml) andvarious concentrations of sodium glutamate was determined. Theinitial growth rates of cells incubated with various concentrations ofsodium glutamate was determined on the basis of optical density.
The specific activities for PCP removal, estimated fromseveral batch experiments, were 1.51 x 10-13 + 0.08 xlo-13 g of PCP per cell per h (mean + standard error of sixdeterminations) in the absence of supplementary carbon and0.92 x i0-'3 ± 0.09 x i0'1 g of PCP per cell per h (mean+ standard error of three determinations) in the presence ofsupplementary carbon.Apparent lags in PCP metabolism by initially uninduced
cells were greatly reduced by supplementation with readilymetabolizable carbon sources (Table 1). Uninduced Flavo-bacterium cells were resuspended to 109 cells per ml inMS-MH4+ medium containing 51.5 ,ug of PCP per ml eitheralone or in combination with glucose, cellobiose, succinate,aspartate, or glutamate and were incubated for 24 h. In theabsence of supplementary carbon, no significant amount ofPCP was removed during the incubation. In the presence ofsupplementary carbon, however, PCP was removed after avariable period of time. The removal of PCP was accompa-nied by significant growth on all the supplementary com-pounds tested, with the exception of succinate. Succinate isa very poor substrate for the Flavobacterium sp.When uninduced cells were suspended in the presence of
sodium glutamate with various PCP concentrations, increas-ing PCP concentrations did not significantly influence the lagin PCP removal but did reduce the initial rate of removalthereafter (data not shown). Uninduced cells did not imme-diately reduce the aqueous PCP concentration, indicatingthat nonspecific binding to viable cells was not significant.
2454 TOPP ET AL.
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

PCP-DEGRADING FLAVOBACTERIUM SP. 2455
60 [
40
oo
20 [
00 1.0 2.0 3.0 4.0
HOURS
FIG. 4. Protein synthesis requirement for recovery from PCP-mediated damage. Glutamate-grown, induced cells were resuspended inMS-NH4' medium containing PCP (70 ,ug/ml). Batch cultures were incubated in the absence of supplementary carbon, except for one (A)which received 4 g of sodium glutamate per liter at the start of the incubation. At the time indicated by the second arrow from the left, somecultures (open symbols) received 4 g of sodium glutamate per liter. One of these (A) also received chloramphenicol (100 ,ug/ml) 10 min priorto the addition of sodium glutamate (indicated by the first arrow from the left). A control flask (c) did not receive any supplements duringthe incubation.
Under some conditions, supplementary carbon partiallyrepressed PCP metabolism. Incubation of glutamate-growncells with glucose and glutamate together significantly re-
pressed removal of PCP, 2,4,6-trichlorophenol, 2,4,5,6-tetra-chlorophenol, and 2,3,5,6-tetrachlorophenol, all of which are
metabolized by PCP-induced cells (28) (Table 2 and Fig. 7).Nevertheless, repressed cells did significantly metabolizeeach of the chlorinated phenols, whereas, in the absence ofsupplementary carbon, there was no significant metabolism.Cellobiose-grown cells, but not glutamate-grown cells, were
also repressed by cellobiose in combination with sodiumglutamate (data not shown). Minimum concentrations ofabout 0.25 g/liter for glucose and about 4.0 g/liter for sodiumglutamate were required to repress (Table 3). The growth ofrepressed cells was inhibited by PCP to the same extent as
that of nonrepressed cells (Fig. 7). Glutamate-grown, PCP-induced cells incubated in the presence of both glucose andsodium glutamate removed PCP with essentially linear ki-netics during a 3-h incubation (Fig. 8). PCP was removedmore rapidly in all treatments in which supplementary car-
bon was added. Neither acetate nor citrate repressed PCPmetabolism (data not shown).There was no evidence that acclimation to growth on PCP
as the sole carbon source was the result of genetic selection.Flavobacterium cells grown in continuous culture with PCPas the sole C source for 20 days were plated onto nutrientagar and subsequently grown on sodium glutamate in MSmedium. Induced cells suspended in MS medium containingPCP or PCP plus sodium glutamate responded in a manner
similar to that of glutamate-grown cells that had not previ-ously been exposed to PCP (data not shown).
DISCUSSION
The metabolism of PCP by the Flavobacterium sp. was
facilitated by, but did not require, supplementary carbon.We had considered the possibility that PCP metabolismmight be reductant limited since the pathway includes, at a
minimum, two reductive dehalogenation steps (27). If thepathway is analogous to that in Rhodococcus chloropheno-licus, there may also be at least a third reductive dehaloge-nation (2). If the Flavobacterium sp. was starved for reduc-tant when grown on PCP, provision of an additional electrondonor would be expected to increase the specific activity ofPCP removal. The somewhat higher specific activities in theabsence of additional carbon indicate that additional carbonis not necessary for PCP metabolism. Furthermore, theFlavobacterium sp. grows in continuous culture on PCP as
the sole carbon source (data not shown).When uninduced glutamate-grown Flavobacterium cells
were inoculated into a medium containing PCP and a sup-plementary carbon source, the time required for the onset ofmeasurable PCP removal corresponded to that required foradaptation to, and growth on, the supplementary compound(Table 1). In the absence of cell-free activity, we have no
measurement of enzyme synthesis other than whole-cellactivity. Therefore, we were unable to determine whetherthe reduction of apparent lag effected by supplementarycarbon was through a reduction in induction time, throughstimulation of the activity of an induced population, or both.Since non-PCP-metabolizing mutants grow on sodium gluta-mate in the presence of PCP at 100 jig/ml (data not shown),PCP metabolism is not required for growth in the presence of
I IAI I
A
0
VOL. 54, 1988
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
2456 TOPP ET AL.
1200
e 280 -
I.-
040 A o _ 0
O.0 \N-., 0\
120 -
- 81
CD0o
41o~~~~ A-X/ A
ol.-/S *~~~
0 40 80HOURS
120
FIG. 5. Viability (lower panel) of, and PCP removal by (upper panel), low Flavobacterium cell densities in the presence and absence ofsupplementary carbon. Glutamate-grown, induced cells were resuspended in MS-NH4' medium. Control flasks were carbon free (*). Flasksreceived glucose (20 pg/ml) and various PCP concentrations (A, 0, and *) or PCP alone (0).
this compound. The observation that growth on PCP ofPCP-induced, glutamate-grown cells is preceded by a longlag indicates that there are adaptations required in additionto the synthesis of the catabolic pathway. In the absence of
supplementary carbon, acclimation took much longer butwas variable depending on the prior growth conditions andinoculum density (data to be published elsewhere). Growthconditions and nutritional limitation influence the sensitivity
60
401I
to
20 -
00
U
U
20 40
0
60 80Hours
FIG. 6. Stimulation of PCP removal by cell lysate. Initial cell densities and growth conditions were identical to those in the experimentreported in the legend to Fig. 9. Undiluted cell lysate from 9 x 106 cells (l) or cell lysate diluted 10-fold (0), diluted 100-fold (A), orautoclaved for 10 min (0) was added at the start of the incubation. Fifty mg milligrams of sodium acetate 3H20 per liter was added tocontrols (O).
APPL. ENVIRON. MICROBIOL.
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

PCP-DEGRADING FLAVOBACTERIUM SP. 2457
TABLE 1. Growth and PCP metabolism in the presence andabsence of various supplementary carbon sourcesa
Residual ,ug of PCP per ml (growth [A6w]) ata:Treatment
3 h 7 h 24 h
PCP alone 50.0 48.8 (0.33) 48.2 (0.33)PCP+glutamate 2.5 ND (1.30) ND (1.90)PCP+aspartate 3.2 ND (0.94) ND (1.70)PCP+succinate 34.0 10.4 (0.37) 4.1 (0.42)PCP+glucose 3.4 ND (1.50) ND (1.65)PCP+cellobiose 47.4 41.8 (0.30) 4.1 (0.68)
a Supplementary substrate concentrations were adjusted to give the samecarbon concentration as that contained in 4 g of sodium glutamate per liter.PCP and other carbon sources were added together at the start of theincubation. Growth was measured by A6w. At 0 h, for all treatments, 51.5 ,ugof PCP per ml the A6w for all treatments at 0 h was 0.43. ND, Not done.
of bacteria to chlorinated phenols and other antimitcrobialagents (3, 13).The concentration of sodium glutamate required to opti-
mally stimulate PCP removal was similar to that required tosaturate growth. It may be that the metabolic step that limitsgrowth on glutamate also limited glutamate-dependent en-hancement of PCP metabolism.When glutamate-grown induced cells were incubated with
4 g of sodium glutamate per liter and PCP at variousconcentrations, the initial rate of PCP removal, measuredduring the first 30 min of incubation, was inhibited by PCPwith a Ki of 86.8 p.g/ml. PCP metabolism rates acceleratedduring the incubation; however, in the presence of chloram-phenicol, rates remained linear and independent of the PCPconcentrations within the range of 70 to 145 ,ug/ml (data notshown). Therefore, there was a difference in the sensitivityof induced cells upon initial exposure to high PCP concen-trations and thereafter. Several conclusions can be drawnfrom these results. In the presence of chloramphenicol, thedamage effected upon initial exposure to PCP remained withthe population throughout the incubation and, therefore,could not be repaired in the absence of protein synthesis. On
TABLE 2. Repression of metabolism of chlorinated phenols byglucose and glutamatea
Repression induced by treatment with:%o SubstrateSubstrateb Sodium glutamate Glucose removed
(4 g/liter) (5 g/liter)
PCP - 2.5+ - 61.9+ + 15.5
2,4,6-TCP - - 0.8+ - 100.0+ + 53.0
2,3,4,6-TeCP - - 0.0+ - 100.0+ + 31.9
2,3,5,6-TeCP - - 2.5+ - 100.0+ + 22.0
a Uninduced glutamate-grown cells were resuspended to an optical densityof 0.8 in MS-NH4+ medium, and 15-ml portions were dispensed into 50-miErlenmeyer flasks. Cell suspensions were incubated for 3 h. Chlorinatedsubstrates were present at an initial concentration of 25 ,ug/ml.
b 2,4,6-TCP, 2,4,6-Trichlorophenol; 2,3,4,6-TeCP, 2,3,4,6-tetrachlorophe-nol; 2,3,5,6-TeCP, 2,3,5,6-tetrachlorophenol.
the other hand, the linear kinetics throughout the incubationin the presence of chloramphenicol indicate that no damageoccurred beyond that suffered initially and that proteinsynthesis was not required for maintenance of activity. If,relative to concentration, PCP had retained its initial toxicitythroughout the incubation, the kinetics would have decayedbecause of accumulating cellular damage. These data indi-cate that cells were sensitive to shock loads of PCP but that,following the initial exposure, the cells were more resistant.We suggest that PCP-metabolizing cells are afforded signif-icant protection by the destruction of the compound as itenters the cell. Higher bulk concentrations are tolerated byinduced, PCP-degrading cells since more external PCP isrequired to raise the internal concentration to the toxicthreshold. The initial damage caused by shock loads of PCPis presumably due to the rapid entry of PCP prior to theonset of metabolism. In the absence of chloramphenicol, therates accelerated somewhat until the PCP concentration wasreduced to undetectable levels. Repair of the damage ini-tially effected by PCP as well as an increase in the PCP-metabolizing enzyme content above the initial complementmay have contributed to the increase in activity.When PCP-induced cells were shifted from a medium
containing PCP and sodium glutamate to one containing PCPas the only carbon source, PCP removal activity decayedwithin minutes. As expected, the loss of activity was morepronounced at higher PCP concentrations (Fig. 2). Thedecay in PCP degradation rate was accompanied by cellulardamage, the repair of which required protein synthesis. At aminimum, PCP binds to and changes the configuration ofproteins (33) and intercalates into and disrupts membranefluidity (9), and it may competitively inhibit enzymes havingadenine-containing cofactors or utilizing adenine-containingsubstrates (8, 29). Although cellular damage became appar-ent quite rapidly, reduction in viability of cells recovered onnutrient agar took longer (data not shown). Survivorsadapted to PCP as the sole carbon source and metabolizedthe compound with specific activities somewhat higher thanthose of cells growing under mixed-substrate conditions. Wedo not know what specifically causes the decay in activityfollowing the shift. It may be that some adaptation inaddition to the PCP pathway is necessary for the Flavobac-terium sp. to grow on PCP alone and that supplementarycarbon represses the required function. There may also be ametabolic downshift and depletion of intracellular pools, therecovery of which takes significant time. We hypothesizethat events such as metabolic shifts result in transientcessation of PCP metabolism and leave the cells morevulnerable to PCP-mediated damage and that this situationcan result in a fraction of the population being killed.The sensitivity of this Flavobacterium sp. falls within the
range expected of gram-negative bacteria. For example, theMIC (growth in nutrient broth) of PCP for Escherichia coliKH434 was 20 ptg/ml; that for a Pseudomonas sp. was 500,ug/ml (15).The apparent lag prior to measurable PCP removal is a
function of the time required for acclimation and the accu-mulation of sufficient biomass to get measurable rates. Whenthe initial cell density was in the range of 107 CFU/ml,material released from lysed cells apparently shortened thelag phase. We do not know why the stimulatory property ofthe lysis products was destroyed by autoclaving.
Partial repression of the metabolism of chlorinated phe-nols was effected by glucose and sodium glutamate whenthese carbon sources were present together (Table 2). Inter-estingly, synthesis of citrate synthase and aconitate hydra-
VOL. 54, 1988
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

APPL. ENVIRON. MICROBIOL.
1.4
1.00
80.6
0.2
00 2 4 6 8
140
100
&PL60 bD
20
0
HOURSFIG. 7. Repression of PCP removal by glucose. Several batch cultures of Flavobacterium cells growing on 4 g of sodium glutamate per
liter in MS medium were inoculated at the start of the incubation. At the time indicated by the first arrow from the left, glucose was addedto half of the batch cultures (H and OI). Thirty minutes later, PCP was added to cell suspensions supplemented with sodium glutamate only(a) or with sodium glutamate + glucose (O). Controls did not receive PCP (O and O). Residual PCP concentrations in glucose-amended (A)and unamended flasks (A) were monitored.
tase is repressed under similar conditions (12). The concen-trations required to do so were high and would not beencountered in situ. Repressed cells were not inhibited anymore by PCP than were nonrepressed cells; this suggeststhat the repressed rate was sufficient to optimally protect thecells (Fig. 7).
In summary, supplementary carbon decreased acclimationtime and facilitated PCP metabolism by glucose- or gluta-mate-grown Flavobacterium cells. Once cells were accli-mated to growth on PCP alone, their specific activity was
TABLE 3. Glucose and glutamate concentrations required forrepression of PCP metabolisma
Concn (g/liter) Amt of PCP removed
Sodium glutamate Glucose (,ugml)
4.0 0.0 52.84.0 0.075 52.04.0 0.125 59.54.0 0.250 19.24.0 0.500 17.64.0 1.00 20.7
0.0 0.5 61.01.0 0.5 46.34.0 0.5 11.68.0 0.5 11.7
16.0 0.5 6.7a Glutamate-grown cells were resuspended to an optical density of 0.57 in
MS medium and dispensed into Erlenmeyer flasks, and the indicated sub-strates were added. Cells were preincubated for 30 min prior to the addition of61.0 jig of PCP per ml. Cell suspensions were then indubated for 2.75 h.
somewhat higher than that of cells in the presence ofsupplementary carbon. PCP metabolism was subject torepression in the presence of significant concentrations ofglutamate plus either glucose or cellobiose. We suggest that
HOURSFIG. 8. Effect of glucose on PCP removal by preinduced cells.
Flavobacterium cells were grown on sodium-glutamate and inducedwith PCP. Cells were harvested and resuspended in MS-NH4'medium containing PCP (100 ,u.g/ml) and 4 g of sodium glutamate perliter (A), 4 g of glucose per liter (O), both glucose and sodium-glutamate (0), or no supplementary carbon (0).
2458 TOPP ET AL.
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

PCP-DEGRADING FLAVOBACTERIUM SP. 2459
the metabolizable carbon in polluted environments andwaste streams may hasten decontamination should theseattenuate the effects of PCP and contribute to the growth ofPCP-degrading biomass. From the perspective of commer-cial application, readily metabolizable carbon might increasethe survivability in inoculants grown in vitro and thenexposed to high PCP concentrations in situ.
ACKNOWLEDGMENTS
E. Topp was the recipient of a Natural Science and EngineeringCouncil of Canada Postgraduate Scholarship prior to receivingsupport from Agriculture Canada Research Branch. This work waspartially supported by Biotrol, Inc., of Chaska, Minn.
LITERATURE CITED1. Apajalahti, J. H. A., P. Karpanoja, and M. S. Salkinoja-Salonen.
1986. Rhodococcus chlorophenolicus sp. nov., a chlorophenol-mineralizing actinomycete. Int. J. Syst. Bacteriol. 36:246-251.
2. Apajalahti, J. H. A., and M. S. Salkinoja-Salonen. 1987. Com-plete dechlorination of tetrahydroquinone by cell extracts ofpentachlorophenol-induced Rhodococcus chlorophenolicus. J.Bacteriol. 169:5125-5130.
3. Brown, M. R. W., and P. Williams. 1985. Influence of substratelimitation and growth phase on sensitivity to antimicrobialagents. J. Antimicrob. Chemother. 15(Suppl. A):7-14.
4. Chu, J. P., and E. J. Kirsch. 1972. Metabolism of pentachloro-phenol by an axenic bacterial culture. Appl. Microbiol. 23:1033-1035.
5. Chu, J. P., and E. J. Kirsch. 1973. Utilization of halophenyls bya pentachlorophenol metabolizing bacterium. Dev. Ind. Micro-biol. 14:264-273.
6. Crawford, R. L., and W. W. Mohn. 1985. Microbiologicalremoval of pentachlorophenol from soil using a Flavobacte-rium. Enzyme Microb. Technol. 7:617-620.
7. Crosby, D. G. 1981. Environmental chemistry of pentachloro-phenol. Pure Appl. Chem. 53:1051-1080.
8. DalI-Larsen, T., H. Kryvi, and L. Klunysoyr. 1976. Dinitrophe-nol, dicoumarol and perntachlorophenol as inhibitors and para-site substrates in the ATP phosphoribosyltransferase reaction.Eur. J. Biochem. 66:443-446.
9. Duxbury, C. L., and J. E. Thompson. 1987. Pentachlorophenolalters the molecular organization of membranes in mammaliancells. Arch. Environ. Contam. Toxicol. 16:367-373.
10. Edgehill, R. V., and R. K. Finn. 1982. Isolation, characterizationand growth kinetics of bacteria metabolizing pentachlorophe-nol. Eur. J. Appl. Microbiol. Biotechnol. 16:179-184.
11. Edgehill, R. V., and R. K. Finn. 1983. Microbial treatment ofsoil to remove pentachlorophenol. Appl. Environ. Microbiol.45:1122-1125.
12. Flechtner, V. R., and R. S. Hanson. 1969. Coarse and finecontrol of citrate synthase from Bacillus subtilis. Biochim.Biophys. Acta 184:252-262.
13. Gilbert, P., and M. R. W. Brown. 1978. Influence of growth rateand nutrient limitation on the gross cellular composition ofPseudomonas aeruginosa and its resistance to 3- and 4-chloro-phenol. J. Bacteriol. 133:1066-1072.
14. Harder, W., and L. Dikhuizen. 1982. Strategies of mixedsubstrate utilization in microorganisms. Philos. Trans. R. Soc.
Lond. B Biol. Sci. 197:459-480.15. Izaki, K., M. Takahashi, Y. Sato, Y. Sasugawa, K. Sato, and C.
Furusaka. 1981. Some properties of pentachlorophenol-resistantgram-negative bacteria. Agric. Biol. Chem. 45:765-767.
16. Jones, G. L., F. Jansen, and A. J. McKay. 1973. Substrateinhibition of the growth of bacterium NCIB 8250 by phenol. J.Gen. Microbiol. 74:139-148.
17. Kilbane, J. J., D. K. Chatterlee, and A. M. Chakrabarty. 1983.Detoxification of 2,4,5-trichlorophenoxyacetic acid from con-taminated soil by Pseudomonas cepacia. Appl. Environ. Micro-biol. 45:1697-1700.
18. Kim, C. J., and W. J. Maier. 1986. Acclimation and biodegra-dation of chlorinated organic compounds in the presence ofalternate substrates. J. Water Pollut. Control Fed. 58:157-164.
19. Kuwatsuka, S., and M. Igarashi. 1975. Degradation of PCP insoils. II. The relationship between the degradation of PCP andthe properties of soils, and the identification of the degradationproducts of PCP. Soil Sci. Plant Nutr. 21:405-414.
20. Martinson, M. M., J. G. Steiert, D. L. Saber, W. W. Mohn, andR. L. Crawford. 1986. Microbiological decontamination of pen-tachlorophenol-contaminated natural waters, p. 529-534. InG. C. Llewellyn and C. E. O'Rear (ed.), Biodeterioration 6.C.A.B. International, Slough, United Kingdom.
21. Pignatello, J. J., M. M. Martinson, J. G. Steiert, R. E. Carlson,and R. L. Crawford. 1983. Biodegradation and photolysis ofpentachlorophenol in artificial freshwater streams. Appl. Envi-ron. Microbiol. 46:1024-1031.
22. Rao, K. R. 1978. Pentachlorophenol. Chemistry, pharmacology,and environmental toxicology. Plenum Publishing Corp., NewYork.
23. Saber, D. L., and R. L. Crawford. 1985. Isolation and charac-terization of Flavobacterium strains that degrade pentachloro-phenol. Appl. Environ. Microbiol. 50:1512-1518.
24. Sittig, M. 1981. Handbook of toxic and hazardous chemicals.Noyes Publications, Park Ridge, N.J.
25. Stanlake, G. J., and R. K. Finn. 1982. Isolation and character-ization of a pentachlorophenol-degrading bacterium. Appl. En-viron. Microbiol. 44:1412-1427.
26. Steiert, J. G., and R. L. Crawford. 1985. Microbial degradationof chlorinated phenols. Trends Biotechnol. 3:300-305.
27. Steiert, J. G., and R. L. Crawford. 1986. Catabolism of pentach-lorophenol by a Flavobacterium sp. Biochem. Biophys. Res.Commun. 141:825-830.
28. Steiert, J. G., J. J. Pignatello, and R. L. Crawford. 1987.Degradation of chlorinated phenols by a pentachlorophenol-degrading bacterium. Appl. Environ. Microbiol. 53:907-910.
29. Stockdale, M., and M. J. Selwyn. 1971. Influence of ringsubstituents on the action of phenols on some dehydrogenases,phosphokinases, and the soluble ATPase from mitochondria.Eur. J. Biochem. 21:416-423.
30. Suzuki, T. 1977. Metabolism of pentachlorophenol by a soilmicrobe. J. Environ. Sci. Health Part B 12(2):113-127.
31. Thompson, W. S. 1970. Contribution of the wood-preservingindustry to water pollution, p. 50-77. In W. S. Thompson (ed.),Pollution abatement and control in the wood preserving indus-try. Mississippi State University Press, Jackson.
32. Watanabe, I. 1973. Isolation of pentachlorophenol-decomposingbacteria from soil. Soil Sci. Plant Nutr. 19:109-116.
33. Weinbach, E. C., and J. Garbus. 1965. The interaction ofuncoupling phenols with mitochondria and with mitochondrialproteins. J. Biol. Chem. 240:1811-1819.
VOL. 54, 1988
on June 24, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from