increased grain yield and biomass allocation in rice under cool night temperature
TRANSCRIPT

ORIGINAL ARTICLE
Increased grain yield and biomass allocation in rice under coolnight temperature
Keiichi KANNO and Amane MAKINOGraduate School of Agricultural Science, Tohoku University, Sendai 981-8555, Japan
Abstract
The effects of different night temperatures on grain yield were examined in rice (Oryza sativa L. Akita-63) dur-
ing the ripening period. Plants were grown under two different night temperatures (22 and 27�C) from anthesis
to harvesting. The day temperature was maintained at 27�C in both treatments. Although the final biomass at
harvest did not differ between the treatments, the dry weight of the panicles was significantly greater in the cool
night temperature treatment. This increase in panicle weight was associated with increases in the 1000-kernel
weight and the ratio of filled spikelets. Although panicle respiration in the high night temperature treatment
decreased to almost zero just after the cessation of grain filling, the net CO2 fixation rate per day in the whole
plant in this treatment tended to be higher, and this was associated with a higher level of starch accumulation.
After grain filling, the starch content decreased and the final dry weight of other plant parts, including dead
organs and new tillers, was greater in the high night temperature treatment. Thus, the noted decline in yield at
the high night temperature led to changes in carbon allocation to new sinks for vegetative reproduction at the
whole plant level. From these results, we considered the possibility that a cool night temperature tends to favor
carbon allocation to panicles, resulting in higher yield.
Key words: biomass, grain yield, night temperature, Oryza sativa L, respiration.
INTRODUCTION
Rice (Oryza sativa L.) provides more food for humans
than any other crop. More than 90% of the world’s pro-
duction of rice is in Asia. Demand for rice is growing fas-
ter than the demand for other crops because population
increases are greatest in the rice-consuming regions of
Asia and Africa. However, several researchers have
pointed out that an increase in night temperatures from
global warming may reduce rice yields throughout the
world. Seshu and Cady (1984) predicted that rice yields
will decrease by 0.41 ton ha)1 (41 g m)2) when the aver-
age minimum daily temperature rises from 22 to 23�C.
Peng et al. (2004) reported direct evidence of decreases in
rice yields resulting from increased night temperatures
associated with global warming, with night temperature
strongly negatively correlated with grain yield. Ziska and
Manalo (1996) also observed a decline in the grain yield
of rice grown at a high night temperature. Similar results
were found by other researchers, that is, a negative corre-
lation between night temperature and grain yield in rice
(Morita et al. 2002, 2004, 2005; Sato and Takahashi
1971).
As rice is a tropical crop, yield is always higher in north-
ern Japan than in southern Japan (Murata 1964).
Although the reason for this difference is unknown, it is
possible that a difference in night temperature leads to the
difference in yield (Suzuki and Moroyu 1962). One possi-
ble explanation that has been considered is a carbohydrate
deficit caused by increased respiratory loss at high temper-
ature during the ripening period (Arashi and Tateishi
1950; Suzuki and Moroyu 1962; Sato and Takahashi
1971). However, Morita and co-workers recently
reported that the main factor for the decline in yield under
high night temperature was not an increase in respiratory
loss because no correlation between final grain weight and
whole plant biomass was found under different day ⁄ night
temperatures (Morita et al. 2002, 2004). Although their
experimental conditions for the day ⁄ night temperatures
(i.e. a regime of 22 ⁄ 34�C) were unnatural, the results of
Morita et al. (2004) suggest that an unknown factor
besides respiratory loss is related to the relatively low yield
in southern Japan. However, it is not still clear why grain
weight is not correlated with the whole plant biomass at
Correspondence: A. MAKINO, Graduate School of AgriculturalScience, Tohoku University, 1-1 Tsutsumidori-Amamiyamachi,Sendai 981-8555, Japan. Email: [email protected]
Received 28 July 2009.Accepted for publication 2 March 2010.
� 2010 Japanese Society of Soil Science and Plant Nutrition
Soil Science and Plant Nutrition (2010) 56, 412–417 doi: 10.1111/j.1747-0765.2010.00473.x

high night temperatures. Interestingly, Sato and Takah-
ashi (1971) observed a positive correlation between the
1000-kernel weight and the harvest index of rice growing
at different day ⁄ night temperatures. In addition, they
pointed out the possibility that a cool night temperature
increased the 1000-kernel weight. From their results,
however, it is difficult to evaluate the effects of night tem-
perature on grain yield because the grain yield depended
on both day and night temperatures.
The purpose of the present study was to elucidate the
effect of different night temperatures on grain yield and
biomass production of rice during the ripening period. We
grew rice plants hydroponically and transferred them to
temperature-controlled glasshouses under two different
night temperatures: 22 and 27�C. The day temperature
for all plants was maintained at 27�C. As previously
described (Kanno et al. 2009), the temperature of the
nutrient solution was also controlled using electric heaters
at the bottom of the plant pots. First we investigated the
allocation of biomass at the whole plant level at the
harvesting stage and examined the yield components of
the grains. Second we measured the rates of gas exchange
at the panicle and whole-plant levels. In addition, changes
in the amounts of carbohydrates were examined in the
stems during the ripening period. Finally, we discuss the
biological significance of a decline in rice yield under
elevated night temperatures.
MATERIALS AND METHODS
Plant culture and sampling
Rice (Oryza sativa L. cv Akita-63) plants were grown
hydroponically in a greenhouse following the procedure
used by Mae and Ohira (1981). Eight seedlings each were
transplanted to a 3.5-L plastic pot containing nutrient
solution. When the plants were heading, all plants were
moved into temperature-controlled greenhouses under
natural sunlight conditions (S-203A; Koito, Yokohama,
Japan) with two different night temperatures. One treat-
ment was set at 27�C during the day (05.00–19.00 hours)
and 22�C at night (cool night temperature [CNT]). The
other treatment was set at a constant 27�C (high night
temperature [HNT]). As reported previously (Kanno et al.
2009), because the water temperature of the nutrient
solution in the control in the morning is lower than the air
temperature, the solution was heated for 1 h from 04.00
to 05.00 hours with a 50-W heater attached to an
air-stone in the bottom of the pot. Thus, all plants were
grown under the same conditions during the daytime.
Four plants from each treatment were sampled between
11.00 and 14.00 hours 15, 30 and 45 days after anthesis.
Panicles, leaf blades, leaf sheaths, stems, roots and other
parts (containing dead parts and new tillers) were oven-
dried separately at 80�C for at least 3 days. The stems
were used to determine the sucrose and starch contents as
described previously (Nakano et al. 1995).
Yield components
Forty-five days after anthesis, the panicles from three pots
were collected and dried at room temperature for
3 weeks. Total panicle numbers per pot were in the range
of 21–26 (see Table 1). The panicles were hand-threshed
to determine the proportion of filled spikelets. The filled
spikelets were separated as they sunk in a NaCl solution
with a specific gravity of d = 1.06 (g cm)3). The sub-
merged spikelets were washed with water, hulled and then
oven-dried at 80�C to a constant weight to determine the
grain dry weight. The weight of the hulled grain was then
calculated as a moisture content of 15% to fresh weight.
Gas exchange rate of the whole plant
The gas exchange rate was determined using an environ-
mentally controlled growth chamber (Eyelatron FLI-
301NHL; Eyela, Tokyo, Japan) for the whole plant 15,
30 and 45 days after anthesis as described previously
(Kanno et al. 2009). The difference in the partial pressures
of CO2 entering and exiting the growth chamber was
measured continuously with an IRGA (LI-6252; Li-Cor,
Lincoln, NE, USA) and the absolute pressure of the CO2
was measured with another IRGA (ZRH; Fuji Electric,
Tokyo, Japan). Air was passed through soda lime to
remove the CO2 and mixed with a CO2 cylinder. The par-
tial pressure of CO2 in the chamber was adjusted to 37 Pa
and the flow rate through the chamber was maintained at
70 L min)1 (1.17 L s)1). Irradiance during the day was
800 lmol quanta m)2 s)1 at the center of the plant and
the temperature was set at the same temperature used
in the treatments. The respiratory rate of the panicles
was estimated by subtracting the dark respiration of the
Table 1 Yield components of rice grown at a night temperature of 22 or 27�C at the ripening stage (45 days after anthesis)
Temperature (�C)
Grain yield
(g pot)1)
1000-kernel
weight (g)
Filled
spikelets (%)
No. panicles
(no. pot)1)
No. spikelets
per panicle
CNT (27 ⁄ 22) 30.3 ± 0.7* 28.3 ± 0.1** 90.1 ± 1.6* 21.3 ± 0.3 55.8 ± 1.0
HNT (27 ⁄ 27) 25.4 ± 0.9* 26.0 ± 0.2** 80.9 ± 2.6* 23.7 ± 1.2 52.0 ± 1.0
Values are mean ± standard error (n = 3). *P < 0.05; **P < 0.01; CNT, cool night temperature; HNT, high night temperature.
� 2010 Japanese Society of Soil Science and Plant Nutrition
Rice yield under cool night temperature 413
panicle-removal plants from that of the whole plants.
Dark respiration of the whole plant was first measured for
3 h, and then all panicles were removed and dark respira-
tion was measured again on the remaining tissue for 3 h;
the difference was regarded as an estimate of panicle
respiration.
RESULTS
Although the final biomass at the harvesting stage did not
differ between the treatments, the dry weight of the pani-
cles was appreciably greater in the CNT plants than in the
HNT plants (Fig. 1). However, the dry weight of other
parts was greater in the HNT plants. These parts included
dead organs and new tillers. The dry weight of roots also
tended to be greater in the HNT plants. Thus, a mild
increase in night temperature stimulated the growth of
new sinks, and led to a relative decline in biomass alloca-
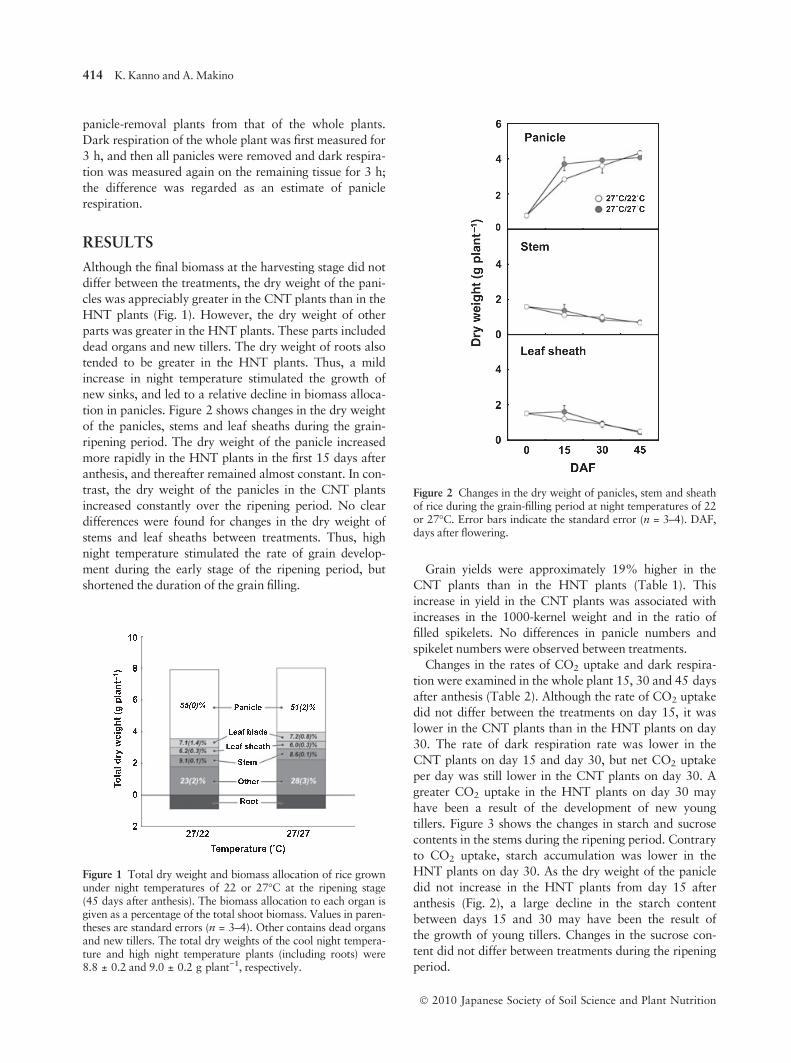
tion in panicles. Figure 2 shows changes in the dry weight
of the panicles, stems and leaf sheaths during the grain-
ripening period. The dry weight of the panicle increased
more rapidly in the HNT plants in the first 15 days after
anthesis, and thereafter remained almost constant. In con-
trast, the dry weight of the panicles in the CNT plants
increased constantly over the ripening period. No clear
differences were found for changes in the dry weight of
stems and leaf sheaths between treatments. Thus, high
night temperature stimulated the rate of grain develop-
ment during the early stage of the ripening period, but
shortened the duration of the grain filling.
Grain yields were approximately 19% higher in the
CNT plants than in the HNT plants (Table 1). This
increase in yield in the CNT plants was associated with
increases in the 1000-kernel weight and in the ratio of
filled spikelets. No differences in panicle numbers and
spikelet numbers were observed between treatments.
Changes in the rates of CO2 uptake and dark respira-
tion were examined in the whole plant 15, 30 and 45 days
after anthesis (Table 2). Although the rate of CO2 uptake
did not differ between the treatments on day 15, it was
lower in the CNT plants than in the HNT plants on day
30. The rate of dark respiration rate was lower in the
CNT plants on day 15 and day 30, but net CO2 uptake
per day was still lower in the CNT plants on day 30. A
greater CO2 uptake in the HNT plants on day 30 may
have been a result of the development of new young
tillers. Figure 3 shows the changes in starch and sucrose
contents in the stems during the ripening period. Contrary
to CO2 uptake, starch accumulation was lower in the
HNT plants on day 30. As the dry weight of the panicle
did not increase in the HNT plants from day 15 after
anthesis (Fig. 2), a large decline in the starch content
between days 15 and 30 may have been the result of
the growth of young tillers. Changes in the sucrose con-
tent did not differ between treatments during the ripening
period.
Figure 1 Total dry weight and biomass allocation of rice grownunder night temperatures of 22 or 27�C at the ripening stage(45 days after anthesis). The biomass allocation to each organ isgiven as a percentage of the total shoot biomass. Values in paren-theses are standard errors (n = 3–4). Other contains dead organsand new tillers. The total dry weights of the cool night tempera-ture and high night temperature plants (including roots) were8.8 ± 0.2 and 9.0 ± 0.2 g plant)1, respectively.
Figure 2 Changes in the dry weight of panicles, stem and sheathof rice during the grain-filling period at night temperatures of 22or 27�C. Error bars indicate the standard error (n = 3–4). DAF,days after flowering.
� 2010 Japanese Society of Soil Science and Plant Nutrition
414 K. Kanno and A. Makino

Finally, panicle respiration was determined. As it was
difficult to directly measure the respiratory rate of the
panicles, the rate was estimated by subtracting the dark
respiration of panicle-removal plants from that of the
whole plants. Panicle respiration did not differ between
the treatments on day 15, but decreased to almost zero
in the HNT plants on day 30 (Table 3). This means
that panicle respiration had no effect on the dry weight of
panicles in the HNT plants.
DISCUSSION
In agreement with previous studies, our results indicated
that a mild increase in night temperature during the ripen-
ing period led to a significant decline in grain yield (Fig. 1;
Table 1). This decline in yield was caused by declines in
both grain weight and the ratio of filled spikelets
(Table 1). Similar results were observed by Morita et al.
(2004). Conversely, Peng et al. (2004) attributed the
decline in yield at high night temperature to a reduction in
the total number of spikelets per plant. In the case of
Morita’s experiments and ours, because the temperature
treatments were imposed after anthesis, no differences in
spikelet number per panicle and panicle number per plant
were observed. Thus, when the plants were grown under
different night temperatures from the vegetative stages
through to anthesis, a decline in yield at high night tem-
perature might be caused by lower grain maturity as well
as a decrease in the spikelet number.
Morita et al. (2005) reported that high night tempera-
ture shortens the duration of grain development and
consequently results in a decline in final grain weight.
They also found a reduction in cell size between the
central point and the surface of the endosperm of the
grain, and considered the possibility of lower assimilate
supply to the grains at high night temperature. A shorten-
ing of the duration of grain development at high night
temperature may be related to an accelerated rate of
increase in grain weight. Kobata and Uemuki (2004)
reported that an early cessation of grain development is
likely to be caused by a shortage of the assimilate supply
to meet the requirements for increases in the grain growth
rate. In fact, early cessation of grain development was also
observed when a high night temperature was applied to
only the panicles (Morita et al. 2004; Nagato and Ebata
1960). However, Morita et al. (2004) found that the
decline in grain weight was greater when the whole plant
was exposed to a high night temperature rather than the
panicles alone. This suggests that the noted decline in
Figure 3 Changes in the starch and sucrose contents in thestems of rice during the grain-filling period at night temperaturesof 22 or 27�C. Error bars indicate the standard error(n = 3). *P < 0.05; **P < 0.01. DW, dry weight. DAF, daysafter flowering.
Table 2 Dark respiration and gross and net CO2 uptake inwhole plants grown at a night temperature of 22 or 27�C fromanthesis, 15, 30 and 45 days after anthesis
Temperature
(�C)
Dark
respiration
(nmol CO2
plant)1 s)1)
Gross CO2
uptake
(nmol CO2
plant)1 s)1)†
Net CO2
uptake
(nmol CO2
plant)1 s)1)‡
On anthesis
– 34 102 68
Day 15
CNT (27 ⁄ 22) 14 89 75
HNT (27 ⁄ 27) 21 91 70
Day 30
CNT (27 ⁄ 22) 4.6 21 16
HNT (27 ⁄ 27) 7.7 41 34
Day 45
CNT (27 ⁄ 22) 8.6 3.5 )5.1
HNT (27 ⁄ 27) 4.1 5.0 0.9
†Gross CO2 uptake is defined as the CO2 uptake actually measured dur-ing the day. ‡Net CO2 uptake is defined as the gross CO2 uptake minusdark respiration. Values are means (n = 2). CNT, cool night temperature;HNT, high night temperature.
Table 3 Dark respiration of the panicles and whole plantsgrown at night temperatures of 22 or 27�C 15 and 30 days afteranthesis
Temperature
(�C)
Panicle
(nmol CO2
plant)1 s)1)
Whole plant
(nmol CO2
plant)1 s)1) Ratio (%)†
Day 15
CNT (27 ⁄ 22) 10.6 25.2 42.1
HNT (27 ⁄ 27) 10.0 23.2 42.9
Day 30
CNT (27 ⁄ 22) 2.64 5.10 51.9
HNT (27 ⁄ 27) 0 13.9 0
†Ratio of respiration of the panicle to the whole plant. Values are means(n = 2).
� 2010 Japanese Society of Soil Science and Plant Nutrition
Rice yield under cool night temperature 415

grain weight under high night temperature conditions is
the result of events at the whole plant level. Morita et al.
(2004) pointed out the possibility of increased panicle
respiratory losses under high night temperature condi-
tions. Chowdhury and Wardlow (1978) reported a
decline in grain weight with increased seed respiratory
losses at high temperatures in cereals including wheat,
barley and sorghum. However, our results clearly indicate
that panicle respiration decreased to almost zero just after
the early cessation of grain development (Table 3). This
indicates that a decline in grain weight at high night tem-
perature is not caused by panicle respiratory losses.
Our results indicated that the final total biomass at the
harvest stages did not differ between the treatments
(Fig. 1). These results suggest that the decline in yield at
high night temperature was not caused by a promotion of
respiratory loss. Similarly, Morita et al. (2004) observed
no correlation between grain weight and whole plant
biomass under varying day ⁄ night temperatures, and
considered that the decline in grain weight at high temper-
ature was not caused by a carbohydrate deficit in sources
as a result of increased respiration loss. We found greater
accumulation of starch in the stems in the high night
temperature treatment just after the cessation of grain
development (Fig. 3), and net CO2 uptake per day tended
to be higher in this treatment (Table 2). In addition, the
starch content decreased after the cessation of grain devel-
opment (Fig. 3) and the dry weight of other plant parts,
including dead organs and new tillers, was greater at the
final stages (Fig. 1). From these results, we consider the
possibility that a decline in yield at high night temperature
is the result of changes in carbon allocation to new sinks
for vegetative reproduction at the whole plant level. How-
ever, we cannot rule out another possibility that an early
cessation of grain development by high night temperature
gives rise to carbon reallocation to new tillers. In fact,
Nakano et al. (1995) observed that trimming of the pani-
cles stimulates the development of late tillers and roots. In
their experiments, the photosynthate after trimming of the
panicles was efficiently translocated to other organs, such
as new tillers and roots. Similar results were observed by
Sato and Takahashi (1971) in rice growing under different
day ⁄ night temperatures. Thus, further experiments are
needed to determine changes in source and sink regulation
under high night temperature conditions.
Rice yield is always higher in northern Japan than in
southern Japan (Murata 1964). Our results clearly indi-
cate that a cool night temperature tends to favor carbon
allocation to panicles, resulting in higher yield. The rice
cultivar Oryza sativa originates from Asian wild strains of
Oryza rupifigon, with continuous variation between the
annual and perennial types (for a review see Morishita
1984). Therefore, perenniality associated with the ances-
tral species may have a strong influence on the enormous
diversity of O. sativa. As O. sativa can ratoon, individu-
als are essentially perennial (Oka and Morishita 1967).
Previously, Sano and Morishita (1982) highlighted that
O. sativa shows higher reproductive (annual) effort in
suboptimal environments than in optimal environments.
Thus, under high night temperature conditions individuals
may partially switch from an annual form to a perennial
form because rice is a tropical plant.
ACKNOWLEDGMENTS
This work was supported by a grant from the Ministry
of Agriculture, Forestry and Fisheries of Japan (Genomics
for Agricultural Innovation, GPN-0007), Scientific
Research B from the Japan Society for the Promotion of
Science (No. 20380041) and Research in a Proposed
Research Area (Planned Research No. 21114006) from
the Ministry of Education, Culture, Sports, Science and
Technology. We wish to thank Professor Emeritus Tada-
hiko Mae for his valuable advice and comments during
the course of this study. We also thank Professors Yukio
Watanabe and Hikaru Goto for their encouragement on
this work and Dr Louis J. Irving for critical reading of the
manuscript.
REFERENCES
Arashi K, Tateishi S 1950: Effect of meteorological conditions on
growth and yield of rice plant in Kyushu district with special
reference to the consideration from the stand point of ‘‘Aki-
ochi’’ phenomena. I. Air temperature. Kyushu Agric. Res.,
6, 95–98.
Chowdhury SI, Wardlow IF 1978: The effect of temperature on
kernel development in cereals. Aust. J. Agric. Res., 29, 205–
223.
Kanno K, Mae T, Makino A 2009: High night temperature stim-
ulates photosynthesis, biomass production and growth dur-
ing the vegetative stage of rice plants. Soil Sci. Plant Nutr.,
55, 124–131.
Kobata T, Uemuki N 2004: High temperature during the grain-
filling period do not reduce the potential grain dry matter
increase of rice. Agron J., 96, 406–414.
Mae T, Ohira K 1981: Remobilization of nitrogen related to leaf
growth and senescence in rice plants (Oryza sativa L.). Plant
Cell Physiol., 22, 1067–1074.
Morishita H 1984: Wild plants and domestication. In Biology of
Rice, Ed S Tsunoda and N Takahashi, pp. 3–30, Elsevier,
Amsterdam Oxford New York Tokyo.
Morita S, Shiratsuchi H, Takanashi J, Fujita K 2002: Effects of
high temperature on ripening in rice plants – comparison of
the effects of high night temperatures and high day tempera-
tures. Jpn. J. Crop Sci., 71, 102–109.
Morita S, Shiratsuchi H, Takanashi J, Fujita K 2004: Effects of
high temperature on ripening in rice plants – analysis of the
effects of high night and high day temperatures applied to
� 2010 Japanese Society of Soil Science and Plant Nutrition
416 K. Kanno and A. Makino

the panicle and other parts of the plant. Jpn. J. Crop Sci.,
73, 77–83.
Morita S, Yonemura J, Takahashi J 2005: Grain growth and
endosperm cell size under high night temperatures in rice
(Oryza sativa L.). Ann Bot., 95, 695–701.
Murata Y 1964: On the influence of solar radiation and air tem-
perature upon the local difference in the productivity of
paddy rice in Japan. Proc. Crop Sci. Soc. Jpn., 33, 59–66.
Nagato K, Ebata M 1960: Effects of temperature in the ripening
periods upon the development and qualities of lowland rice
kernels. Proc. Crop Sci. Soc. Jpn., 28, 275–278.
Nakano H, Makino A, Mae T 1995: Effects of panicle removal
on the photosynthetic characteristics of the flag leaf of rice
plants during the ripening stage. Plant Cell Physiol., 36,
653–659.
Oka HI, Morishita H 1967: Variations in the breeding systems
of a wild rice Oryza perenis. Evolution, 21, 249–258.
Peng S, Huang J, Sheehy JE et al. 2004: Rice yields decline with
higher night temperature from grobal warming. Proc. Natl
Acad. Sci. USA, 101, 9971–9975.
Sano Y, Morishita H 1982: Variation in resource allocation and
adaptive strategy of a wild rice Oryza perennis Moench.
Bot. Gaz., 143, 518–523.
Sato K, Takahashi M 1971: The development of rice
grains under controlled environment. I. The effects of
temperature, its daily range and photoperiod during rip-
ening on grain development. Tohoku J Agric Res, 22,
57–68.
Seshu DV, Cady FB 1984: Response of rice to solar radiation and
temperature estimated from international yield trials. Crop
Sci., 24, 649–654.
Suzuki S, Moroyu H 1962: Effect of high-night-temperature
treatment on the growth of rice plants with special reference
to its nutritional condition. Bull. Chugoku Agr. Exp. Stn., 8,
269–290.
Ziska LH, Manalo PA 1996: Increasing night temperature can
reduce seed set and potential yield of tropical rice. Aust. J.
Plant Physiol., 23, 791–794.
� 2010 Japanese Society of Soil Science and Plant Nutrition
Rice yield under cool night temperature 417