in vivo regulation of mpf in xenopus oocytesfluorographs from a single stag 4e oocyte with a n...
TRANSCRIPT

Development 109, 149-156(1990)Printed in Great Britain © T h e Company of Biologists Limited 1990
149
In vivo regulation of MPF in Xenopus oocytes
ANDREW D. JOHNSON1* and L. DENNIS SMITH12
1 Department of Biological Sciences, Purdue University, West Lafayette, Indiana 47907, USA2 Department of Developmental and Cell Biology, Developmental Biology Center, University of California Irvine, California 92717, USA
* Present address: Department of Zoology, University of Texas, Austin Texas 78712, USA
Summary
Entry into M phase in the eukaryotic cell cycle iscontrolled by the oscillating activity of MPF. The activecomponent of MPF is now known to be the p34cdc2
protein kinase originally found in yeast. The p34cdc2
protein kinase displays a characteristic M-phase-specifichistone HI kinase activity when it interacts with cyclins,which are proteins that oscillate through the cell cycleand are thought to regulate p34cdc2 activity. Cyclins caninduce M phase when introduced into fully grownXenopus oocytes and cyclin may play a role in normaloocyte maturation. Small Xenopus oocytes do not ma-ture in response to the hormonal triggers which act on
stage 6 oocytes. We introduced cyclin into stage 4 (small)Xenopus oocytes and showed that it activates MPF inthese cells, probably by interacting with endogenousp34cdc2 kinase. We made labelled extracts from cyclin-mRNA-injected stage 4 oocytes and used them to showdifferentia] stability of clam cyclins A and B at oocytematuration. The relative stability of the two forms ofcyclin related directly to their ability to stabilize crudeMPF preparations from injected stage 6 oocytes.
Key words: Xenopus oocyte, in vivo regulation, MPF, cellcycle, cyclin.
Introduction
Full-grown amphibian oocytes arrested in late G2 ofmeiosis I, must mature to the second meiotic metaphasebefore fertilization is possible. Oocyte maturation isinduced by an action of progesterone at the oocytesurface. This leads to appearance of a cytoplasmicmaturation-promoting factor (MPF), which inducesnuclear membrane breakdown and the subsequentmeiotic events. MPF was first identified by injectingcytoplasm from maturing oocytes into oocytes not ex-posed to steroid hormone; recipients underwent matu-ration (Masui and Markert, 1971; Smith and Ecker,1971). Using this assay, MPF subsequently was ident-ified in mitotically cleaving embryonic cells (Wasser-man and Smith, 1978; Gerhart et al. 1984) as an activitythat peaked at each M phase and then disappeared.These and several additional studies on diverse celltypes have established MPF as a universal M-phase-inducing factor in eukaryotic cells (Gerhart et al. 1985).
In spite of intense effort for some time, MPF has onlyrecently been characterized. Lohka et al. (1988) ob-tained a highly purified egg fraction exhibiting MPFactivity which contained two predominant proteins, oneof 34 x 103 and one of 45 x 1CP Mr, respectively. The 34Kprotein has been identified as the Xenopus laevishomolog of a fission yeast protein encoded by the genecdc2+ which is required for the G2-M transition in theyeast mitotic cell cycle. This protein is a serine/
threonine that exhibits a strong preference for histoneHI as a substrate; histone HI kinase activity cyclesduring cell division coincident with MPF activity (re-view by Smith, 1989). The second protein has beenidentified as a Xenopus cyclin (Gautier et al. 1990).Cyclins were originally identified because of periodicsynthesis and degradation during mitotic divisions incleaving sea urchin and clam embryos (Evans et al.1983).
A number of studies have shown that p34cdc2 levelsremain constant throughout the mitotic cell cycle(Draetta et al. 1989; Labbe et al. 1989; Meijer et al.1989; Moreno et al. 1989), suggesting that the periodicsynthesis and turnover of cyclin regulates the periodicbursts of MPF activity. In support of this, cyclin isknown to associate with p34cdc2 to form active HIkinase (Draetta et al. 1989; Meijer et al. 1989), inhi-bition of cyclin synthesis prevents activation of MPFactivity in egg extracts (Minshull et al. 1989), andpreventing the destruction of cyclin prevents inacti-vation of MPF activity and maintains M-phase arrest(Murray et al. 1989; Luca and Ruderman, 1989).Further, Murray and Kirschner (1989) have showndirectly that synthesis of cyclin is the only requirementfor activation of MPF activity in a Xenopus egg extract.The activation of MPF during meiosis I appears to bemore complicated.
Full-grown (stage 6) Xenopus oocytes normally acti-vate MPF in response to progesterone by a process that

150 A. D. Johnson and L. D. Smith
requires protein synthesis (Wasserman and Masui,1975). Microinjection of cyclin mRNA into stage 6oocytes also induces entry into M phase (Swenson et al.1986; Pines and Hunt, 1987; Westendorf et al. 1989).One interpretation of these observations is that pro-gesterone induces the synthesis of cyclin, which thenleads to active MPF. However, it has been clear forsome time that stage 6 oocytes contain a precursor formof MPF, pre-MPF, which can be activated and ampli-fied post-translationally by small amounts of activeMPF (Gerhart et al. 1984). Furthermore, Cyert andKirschner (1988) have reported that pre-MPF canactivate spontaneously in an oocyte extract, requiringonly ATP for activation. These observations suggestthat stage 6 oocytes already contain both p34cdc^andcyclin in inactive form (see Barrett et al. 1990). In thisregard, Westendorf et al. (1989) have demonstrated thatclam oocytes contain a pool of sequestered cyclin,which is released by the stimulus (fertilization) thatinduces maturation.
Reports that MPF could be continually amplified byserial transfers of cytoplasm containing MPF throughmultiple recipients initially implied that oocytes containa store of inactive MPF (Reynhout and Smith, 1974;Drury et al. 1975). Based on this biological assay, Taylorand Smith (1987) demonstrated that pre-MPF existed instage 4 oocytes as small as 750 ^m in diameter, but notin smaller oocytes. This suggests that one or morecomponents of MPF are not yet present in early stage 4oocytes. The present study was initiated to test thispossibility. The results show that p34cdc2 is present inoocytes as small as stage 3. Further, injection of cyclinmRNA into oocytes at all stages induces active MPF asassayed by the breakdown of the oocyte nucleus.Surprisingly, stable MPF activity can be extracted fromoocytes that overexpress cyclin protein by homogeniz-ation in simple saline buffers. These results are dis-cussed relative to the role of cyclin in regulating MPFactivity in vivo.
Materials and methods
Antibodies and cDNA clonesAntisera against p34cdc2 used in these studies was provided byDr G. Draetta of the Cold Spring Harbor Laboratory.Plasmids pAXH(+) carrying a clam cyclin A cDNA, andpCD102 encoding clam cyclin B were gifts from Dr J.Ruderman of Duke University.
Manipulation and injection of oocytesOocytes at all stages of oogenesis used in these studies weremanually defolliculated, and maintained in OR 2 medium(Wallace et al. 1973). When maturation was induced, oocyteswere incubated in 10/igml"1 progesterone in OR2. Proteinsynthesis was arrested by incubating oocytes in 10/igmP1
cycloheximide in OR2 for 30min prior to injection of ma-terials, and in the continuous presence of cycloheximidethereafter. The injection of either donor cytoplasm or cyto-solic extracts (50 nl) was performed in healing medium (Fordand Gurdon, 1977) with or without cycloheximide. Followinginjection, the oocytes were incubated in this medium for an
additional 30min to promote healing, and then transferred toOR 2.
Preparation of transcriptsTranscripts were synthesized with SP6 RNA polymeraseaccording to the conditions optimized by Krieg and Melton(1984) with the following modifications: 7mGpppG (Pharma-cia) was included in the reaction at 1 mM, the concentration ofGTP was 100 JJM, and that of SP6 polymerase was 120 units/100/il. The reaction was incubated at 40cC for 2h beforetreatment with DNAse. After extraction and ethanol precipi-tation, the reaction was passed over a NICK column (Pharma-cia) equilibrated with 0.3M sodium acetate, pH5.2, andprecipitated again with ethanol. These conditions routinelyyielded about 2 jig of translatable RNA/reaction. The RNApellet was finally resuspended in H2O at a concentration of 0.5to lmgml"1.
Western blotsA piece of ovary was removed from frogs anesthetized byhypothermia, and the ovary was digested in 2% collagenasein 0.1M NaPO4, pH7.4, at room temperature. This treatmenthas been shown to completely remove follicle cells fromoocytes (Horrell et al. 1987). The oocytes were rinsed exten-sively in OR2, and sized with an ocular micrometer. 5 to 10oocytes were homogenized in 0.5 to 1 ml buffer containing50 mM NaCl, and 0.5 mM PMSF. The homogenates werecentrifuged at 14 000 g and the supernatant was precipitatedwith 4 vol acetone. Pellets were dissolved by boiling inLaemmli buffer and electrophoresed on 10 % polyacrylamide.The gels were transferred to nitrocellulose overnight at 30volts in a Trans blotter (Bio-RAD). The blots were blocked in3 % BSA and probed with a 1000:1 dilution of antibody top34cdc2 in 3 % BSA in Tris-saline. The antibody was detectedwith [125]I protein A from ICN at a final specific activity of0.5/iCiml~^ in 1M NaCl, Tris pH8.0.
Translation of synthetic mRNAsOocytes incubated in OR2 were injected individually with 5 to15 nl delivering 5 to 10 ng of cyclin mRNA per oocyte. Theoocytes were transferred to OR2 containing lOO^CimP1
[35S]methionine (trans label) from ICN, and incubated for 6to 7h. The oocytes were processed as described above forelectrophoresis, or extracts were prepared as described be-low. Gels were impregnated with Amplify (Amersham), andused for fiuorography. It is usually possible to developfluorographs from a single stage 4 oocyte with an overnightexposure under these conditions. In vitro translation wasperformed with a nuclease-treated reticulocyte lysate (Pro-mega) using the recommended conditions, except that about1/10 of the recommended volume of extract was loaded ongels for electrophoresis.
Scoring for GVBDStage 6 oocytes were scored for GVBD by the appearance of awhite spot in the animal hemisphere, confirmed by dissectionafter fixation of oocytes in 0.5N perchloric acid (PCA). Insome cases, oocytes treated with cycloheximide failed todemonstrate a white spot, even if GVBD had occurred.Therefore, in these experiments, oocytes were incubated untila white spot had appeared in oocytes not incubated incycloheximide, usually 3 to 4h, after which they were fixed in0.5 N PCA and dissected. Stage 4 oocytes also do not exhibit awhite spot indicative of GVBD and these oocytes were alwaysdissected in PCA at a time equivalent to GVBD (white spot)in stage 6 oocytes injected with extracts.

In vivo regulation of MPF in Xenopus oocytes 151
Preparation of extractsFor the preparation of cyclin containing extracts from stage 4oocytes, 50 to 100 stage 4 oocytes of 650 to 700 j<m diameterwere injected with 5 to 10 ng of cyclin mRNA. These oocyteswere then incubated in trans label until companion unlabelledstage 4 oocytes had undergone GVBD (usually about 4 to6h). At this time the oocytes were rinsed once in extractionbuffer containing 50 ITIM NaCl, 10 mM Tris pH 7.4. The oocyteswere collected and allowed to settle to the bottom of amicrocentrifuge tube. As much excess extraction buffer aspossible was removed, and the oocyte's were homogenizedwith a heat-sealed micropipet tip. This homogenate was thencentrifuged for 5min at 14000g in a microcentrifuge. Thesupernatant was collected and used directly, or was frozen ondry ice and stored at -70°C. Before injection, an extract wasalways briefly centrifuged to remove debris. Extracts fromstage 6 oocytes were prepared using the same proceduresdescribed for stage 4 oocytes with the exception that only 10 to20 cells were used.
Results
Cyclin activates MPF in small oocytesCyclins are characterized as being either type A or typeB depending on their degree of sequence homology tothe two cyclins identified in clams (Swenson et al. 1989).Either type A or B cyclins are reported to inducematuration when the respective mRNAs are micro-injected into stage 6 Xenopus oocytes (Swenson et al.1986; Pines and Hunt, 1987; Westendorf et al. 1989).Initial studies to test the effects of cyclin on MPFactivation in stage 4 oocytes showed that both mRNAs(5ng/oocyte) induced GVBD when injected intooocytes of about 750 ̂ m in diameter. Additional exper-iments were performed in which cyclin A mRNA onlywas injected into oocytes as small as 650 ^m in diameter.In this case also, GVBD was observed in 98% of therecipient oocytes. In order to verify that the injectedmRNA actually resulted in active MPF, cyclin AmRNA was injected into the smallest oocytes and, afterGVBD had occurred, cytoplasm from these oocyteswas injected into stage 6 oocytes treated with cyclohexi-mide. The induction of GVBD in the absence of proteinsynthesis is considered diagnostic for the presence ofMPF activity (Gerhart et al. 1984, 1985). As shown inTable 1, a high percentage of the stage 6 recipientsunderwent GVBD in response to the injected cyto-plasm.
Cyclin alone is not known to induce GVBD in theabsence of p34cdc2. Thus, the simplest explanation toexplain the results above is that stage 4 oocytes alreadycontain p34cdc2 protein. To test this directly, proteinisolated from stage 3 to 6 oocytes was electrophoresedon polyacrylamide gels, transferred to nitrocelluloseand reacted with antibody to p34cdc2. Fig. 1 demon-strates that the protein is present in oocytes of all stagesexamined. Based on the relative intensity of the bands,the p34cdc2 protein appears to accumulate throughoogenesis.
Extracts from cydin-A-mRNA-injected oocytes induceGVBD in stage 6 recipientsMurray and Kirschner (1989) demonstrated that cyclin
Table 1. Cyclin mRNA induction of MPF in oocytes
TYpfltmpnt ofrecipients
ProgesteroneCyclin A mRNAInjection of cytoplasm from:
Stage 6Stage 6+prog.Stage 6+cyclin A mRNAStage 4+cyclin A mRNA
Injection of extract from:Stage 4Stage 4+cyclin A mRNAStage 6Stage 6+prog.Stage 6+cyclin A mRNA
% GVBC) in recipientsincubated in:
OR2
93(30)100(30)
0(20)100 (20)100 (10)96(23)
0(20)92(48)0(10)0(10)
100 (10)
OR2+Cyclohex.
0(20)0(20)
-95 (21)
100 (32)77(39)
-94(32)0(10)0(20)
95 (20)
Stage 6 and stage 4 oocytes (650-700 ̂ m in diameter) wereinjected individually with 5ng cyclin A mRNA. For cytoplasmictransfers, recipient stage 6 oocytes were injected with 10 nlcytoplasm from donor oocytes. Stage 6+progesterone refers tooocytes incubated in progesterone (10/<gml~') and exhibiting awhite spot (usually 6-7 hours after steroid exposure). Stage6+cyclin mRNA injected refers to oocytes taken at the time ofGVBD (about 3-4 hours after injection). Stage 6 refers to controloocytes (no steroid exposure). Stage 4+cyclin mRNA refers todonors used at a time when companion mRNA-injected oocyteshad undergone GVBD as determined by dissection (about 6 hoursafter injection). For injection of extracts, stage 4 or stage 6 oocytespreviously injected with 5 ng cyclin A mRNA were homogenized atthe time of GVBD and the extracts (see Materials and methods)were frozen on dry ice for use the next day. Recipients (stage 6)were injected with 50nl of the thawed extract. The total number ofrecipient oocytes assayed in each experiment is indicated inparentheses.
protein translated in a reticulocyte lysate was sufficientto induce MPF activity when added to a cell-free eggextract. However, our attempts to induce GVBD instage 6 oocytes by injecting cyclin-mRNA-directedreticulocyte lysates have been unsuccessful (data notshown). One possibility for the failure of cyclin-mRNA-directed lysates to activate MPF in vivo might be that aninsufficient mass of cyclin was produced in the lysates.Alternatively, it is possible that these lysates do notdirect appropriate post-translational modification of theprotein required for interaction with p34cdc2 in vivo. Totest this possibility, we prepared an extract from stage 4oocytes previously injected with cyclin mRNA.
Stage 4 oocytes are known to contain 'spare trans-lational capacity' (Taylor et al. 1985a) and, as shown inFig. 2, translate injected message very well. Fig. 2 (lane2) shows a unique protein of 56xlO3Mr in cyclin-A-mRNA-injected stage 4 oocytes that is also present inreticulocyte lysates containing the same transcript (lane1) but is absent from uninjected oocytes (lane 3). Wehave assumed this protein to be cyclin A. In indepen-dent experiments, we have estimated the rate of cyclinA synthesis in these oocytes to be as high as 7ngoocyte"1. Furthermore, since cyclin mRNA generatesMPF activity in these cells, it seems likely that any pre-requisite post-translational modifications have oc-cutTed.
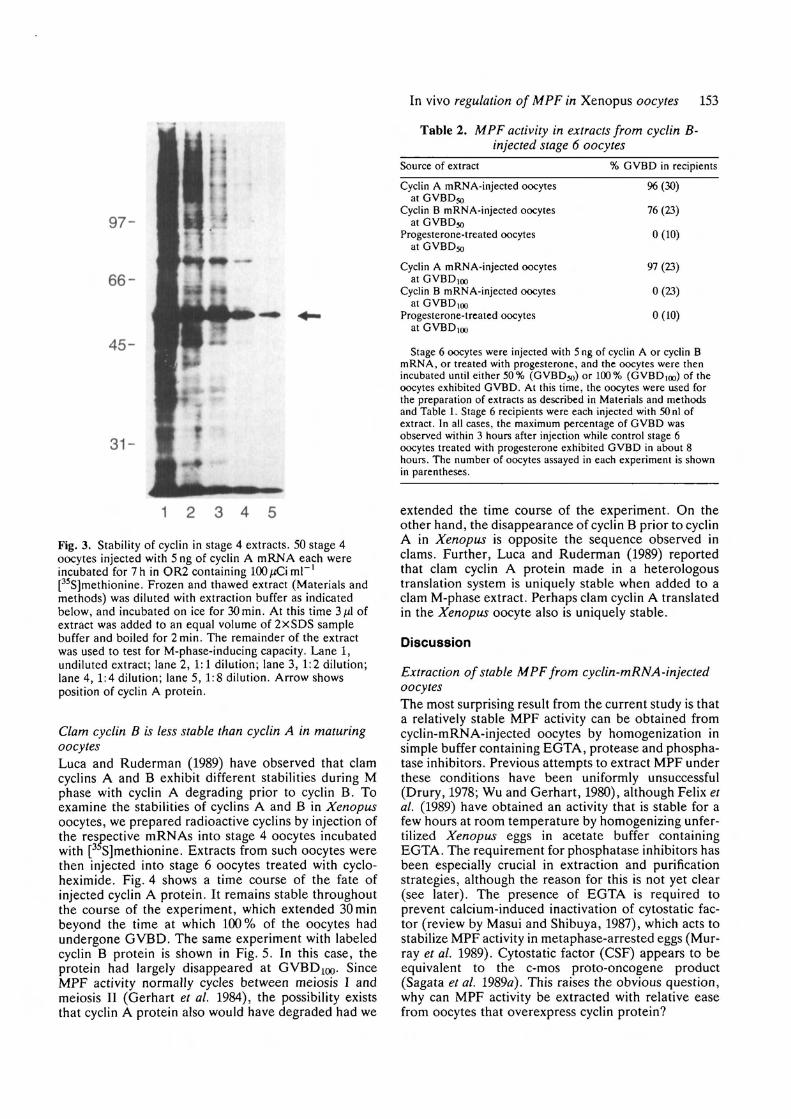
Fig. 3 shows a series of dilutions of a radioactive

152 A. D. Johnson and L. D. Smith
97-
66--97
-66
45-
-45
31-
1 2 3 4 5
Fig. 1. Identification of p34cdc2 in oocytes at different stagesof oogenesis. Each lane of the blot represents the proteinisolated from 5 oocytes at the indicated stages. Lane 1,1300 jan oocytes; lane 2, 1000 fan oocytes; lane 3, 850^moocytes; lane 4, 700^m oocytes; lane 5, 500^m oocytes.Arrow designates position of pM"*02 kinase. Relativemobilities of relative molecular mass markers in this andsubsequent figures are indicated by the numbers in the leftmargin (jWrxl0~3).
extract from cyclin-A-mRNA-injected stage 4 oocytes;the extract had been frozen and thawed and incubatedon ice for 30 min (the average duration of experiments).The cyclin A protein appears to be completely stableunder these conditions. When the extract was injectedinto cycloheximide-treated stage 6 oocytes, GVBD wasinduced in 3 to 4 h (Table 1), while progesterone-treated control oocytes exhibited GVBD in about 7h.The precocious induction of GVBD by cyclin comparedto progesterone has been observed previously in exper-iments involving injection of different amounts of cyclinmRNA (Westendorf et al. 1989) and presumably is afunction of cyclin protein concentration. In support ofthis, the stage 4 extract retained full activity (GVBD at3h after injection) at a dilution of 1:4, but then began todecrease. At a 1:8 dilution, GVBD occurred in 25 % ofthe oocytes at 6 h after injection, in 50 % of the oocytesat 8.5 h with a 1:16 dilution, and in 12.5 h in 16 % of theoocytes after a 1:32 dilution. Further dilutions resultedin loss of activity.
The extraction of relatively stable MPF activity fromcyclin-mRNA-injected stage 4 oocytes by homogeniz-ation in a simple buffer was unexpected since similar
-31
1 2 3*Ffg.̂ 2. Translation of "synthetic cyclin A m'R*NA in stage 4oocytes and reticulocyte lysate. Stage 4 oocytes 650-700 /onin diameter were injected with 5 ng of synthetic cyclin AmRNA, and incubated for 7h in OR2 containinglOO^CimF1 [35S]methionine (trans-label). Controluninjected oocytes were incubated under the sameconditions. Protein was isolated as described in Materialsand methods. For translation in vitro, 0.5 ̂ (g of syntheticcyclin A mRNA was added to mRNA-dependentreticulocyte lysate (Promega), to which [ S]methionineTranslabel (ICN) was added at lj/Cij/1 . The lysates wereincubated for 30 min at 30°C, then boiled in SDS samplebuffer to terminate translation. Lane 1, in vitro translation;lane 2, stage 4 oocytes injected with cyclin A mRNA; lane3, uninjected stage 4 oocytes. Arrow shows position ofcyclin A protein.
approaches with maturing (non-injected) stage 6oocytes have not been successful. To determine if thiswas due to the overexpression of cyclin protein and notsome other difference between stage 4 and stage 6oocytes, stage 6 oocytes injected with cyclin A mRNAwere homogenized and the crude extracts were testedfor MPF activity. The data from these experiments arepresented in Tables 1 and 2. Extracts prepared fromcyclin-A-mRNA-injected oocytes when 50%(GVBD50) and 100% (GVBD]0o) of the oocytesexhibited GVBD was capable of inducing GVBD inrecipients, even when the recipient oocytes had beentreated with cycloheximide. Similar results were ob-tained when stage 6 oocytes injected with cyclin BmRNA were homogenized at GVBDSQ, but no MPFactivity was present in extracts prepared at GVBD 100(Table 2).
In vivo regulation of MPF in Xenopus oocytes 153
97-
66-
45-
Table 2. MPF activity in extracts from cyclin B-injected stage 6 oocytes
31-
1 2 3 4 5
Fig. 3. Stability of cyclin in stage 4 extracts. 50 stage 4oocytes injected with 5 ng of cyclin A mRNA each wereincubated for 7 h in OR2 containing 100 JJC\ ml"'[35S]methionine. Frozen and thawed extract (Materials andmethods) was diluted with extraction buffer as indicatedbelow, and incubated on ice for 30min. At this time 3 fA ofextract was added to an equal volume of 2xSDS samplebuffer and boiled for 2min. The remainder of the extractwas used to test for M-phase-inducing capacity. Lane 1,undiluted extract; lane 2, 1:1 dilution; lane 3, 1:2 dilution;lane 4, 1:4 dilution; lane 5, 1:8 dilution. Arrow showsposition of cyclin A protein.
Clam cyclin B is less stable than cyclin A in maturingoocytesLuca and Ruderman (1989) have observed that clamcyclins A and B exhibit different stabilities during Mphase with cyclin A degrading prior to cyclin B. Toexamine the stabilities of cyclins A and B in Xenopusoocytes, we prepared radioactive cyclins by injection ofthe respective mRNAs into stage 4 oocytes incubatedwith [ S]methionine. Extracts from such oocytes werethen injected into stage 6 oocytes treated with cyclo-heximide. Fig. 4 shows a time course of the fate ofinjected cyclin A protein. It remains stable throughoutthe course of the experiment, which extended 30minbeyond the time at which 100% of the oocytes hadundergone GVBD. The same experiment with labeledcyclin B protein is shown in Fig. 5. In this case, theprotein had largely disappeared at GVBD1Oo- SinceMPF activity normally cycles between meiosis I andmeiosis II (Gerhart et al. 1984), the possibility existsthat cyclin A protein also would have degraded had we
Source of extract
Cyclin A mRNA-injected oocytesat GVBD50
Cyclin B mRNA-injected oocytesat GVBD50
Progesterone-treated oocytesat GVBDW
Cyclin A mRNA-injected oocytesat GVBD100
Cyclin B mRNA-injected oocytesat GVBDioo
Progesterone-treated oocytesat GVBD.nn
% GVBD in recipients
96(30)
76(23)
0(10)
97(23)
0(23)
0(10)
Stage 6 oocytes were injected with 5 ng of cyclin A or cyclin BmRNA, or treated with progesterone, and the oocytes were thenincubated until either 50% (GVBD50) or 100% (GVBDioo) of theoocytes exhibited GVBD. At this time, the oocytes were used forthe preparation of extracts as described in Materials and methodsand Table 1. Stage 6 recipients were each injected with 50 nl ofextract. In all cases, the maximum percentage of GVBD wasobserved within 3 hours after injection while control stage 6oocytes treated with progesterone exhibited GVBD in about 8hours. The number of oocytes assayed in each experiment is shownin parentheses.
extended the time course of the experiment. On theother hand, the disappearance of cyclin B prior to cyclinA in Xenopus is opposite the sequence observed inclams. Further, Luca and Ruderman (1989) reportedthat clam cyclin A protein made in a heterologoustranslation system is uniquely stable when added to aclam M-phase extract. Perhaps clam cyclin A translatedin the Xenopus oocyte also is uniquely stable.
Discussion
Extraction of stable MPF from cyclin-mRNA-injectedoocytesThe most surprising result from the current study is thata relatively stable MPF activity can be obtained fromcyclin-mRNA-injected oocytes by homogenization insimple buffer containing EGTA, protease and phospha-tase inhibitors. Previous attempts to extract MPF underthese conditions have been uniformly unsuccessful(Drury, 1978; Wu and Gerhart, 1980), although Felix etal. (1989) have obtained an activity that is stable for afew hours at room temperature by homogenizing unfer-tilized Xenopus eggs in acetate buffer containingEGTA. The requirement for phosphatase inhibitors hasbeen especially crucial in extraction and purificationstrategies, although the reason for this is not yet clear(see later). The presence of EGTA is required toprevent calcium-induced inactivation of cytostatic fac-tor (review by Masui and Shibuya, 1987), which acts tostabilize MPF activity in metaphase-arrested eggs (Mur-ray et al. 1989). Cytostatic factor (CSF) appears to beequivalent to the c-mos proto-oncogene product(Sagata et al. 1989a). This raises the obvious question,why can MPF activity be extracted with relative easefrom oocytes that overexpress cyclin protein?

154 A. D. Johnson and L. D. Smith
97-
66-
45-
3 1 -
1 2 3 4 5 6
Fig. 4. Cyclin A is stable after injection into oocytes. Anextract was prepared from stage 4 oocytes (700 ̂ mdiameter) previously injected with 5 ng/oocyte cyclin AmRNA and 50nl/oocyte was injected into stage 6 recipientsincubated in cycloheximide. Each lane represents theequivalent of protein from 4 oocytes; equal radioactivitywas loaded in each lane. Variability of label recovered fromeach group of oocytes was about 7%. Lane 1, ljul ofreticulocyte lysate to which cyclin A mRNA was added (seeFig. 3); lane 2, 200nl of original extract (4 oocyteequivalents); lane 3, oocytes processed at time 0; lane 4,oocytes processed at lh post-injection; lane 5, oocytesprocessed at 2h post-injection; lane 6, oocytes processed at3.5 h post-injection (30min after 100% GVBD wasobserved in this population of oocytes). Controlprogesterone-treated oocytes reached 100 % GVBD inabout 9h. Arrow shows position of cyclin A protein.
One explanation is that oocytes that have completedmaturation normally contain relatively small amountsof cyclin, and these are reduced further during themultiple and time-consuming chromatographic stepsused for purification (Lohka et al. 1988). This mightexplain why recovery of MPF has been about 1 % of thestarting activity. There are several reasons for believingthis. For example, Meijer et al. (1989) have reportedthat the ratio of p34cdc2 to cyclin in cleaving sea urchinembryos is about 15:1. Gautier et al. (1989) havereported further that only about 10% of the p34cdc2
kinase actually functions as MPF during Xenopusoocyte maturation, suggesting insufficient levels ofcyclin to interact with all p34cdc2. In addition, MPFpurified from starfish oocytes by a procedure requiring
B 1 2 3
31-
1 2 3 4 5
Fig. 5. Translation of cyclin B mRNA in stage 4 oocytesand stability after injection into stage 6 oocytes. Stage 4oocytes (700/«n diameter) were injected with 5 ng each ofcyclin B mRNA and the oocytes were incubated for 7 h in[ S]methionine. The oocytes then were homogenized andthe extract was injected into stage 6 oocytes (Materials andmethods; Figs 3-5). (A) Stability of cyclin B in stage 6oocytes. Lane 1, cyclin B translated in reticulocyte lysate;lane 2, stage 6 oocytes processed at time=0; lane 3, stage 6oocytes processed 1 h after injection of extract; lane 4, stage6 oocytes processed 2h after injection; lane 5, stage 6oocytes processed at 2.5 h after injection. This timerepresents GVBD in 100% of the oocytes; GVBD inuninjected but progesterone-exposed oocytes was 9h.Arrow shows the position of cyclin B protein.(B) Translation in stage 4 oocytes. Lane 1, cyclin B mRNAdirected reticulocycte lysate; lane 2, cyclin mRNA-injectedoocytes; lane 3, control oocytes. Arrow indicates position ofcyclin B protein.
multiple chromatographic steps (recovery about 1%)contained only p34c but not cyclin, while use of arapid purification procedure on maturing starfishoocytes yielded about a 50% recovery and the MPFcontains both the cdc2 protein as well as cyclin B in a1:1 stoichiometric ratio (Labbe et al. 1989).
The above discussion suggests that maintenance ofhigh cyclin levels during extraction and purificationprocedures is a major prerequisite for the recovery ofmaximal p34cdc2 kinase, hence MPF activity. By thesame token, we suggest that the overproduction ofcyclin protein in mRNA-injected oocytes generates ahigh cyclin to p34cdc2 ratio, facilitating the relativelysimple extraction of MPF activity from small numbersof oocytes. The mechanism by which cyclin functions inthis regard is not clear. However, several studies havenow demonstrated that p34cdc2 kinase is active as MPFonly when it is dephosphorylated (Gautier et al. 1989;

In vivo regulation of MPF in Xenopus oocytes 155
Dunphy and Newport, 1989; Labbe et al. 1989). Thus,the simplest suggestion is that interaction of cyclin withactive p34cdc2 kinase prevents phosphorylation andinactivation of the kinase.
The role of cyclin in MPF activationThere appears to be little doubt that the periodicsynthesis and degradation and cyclin regulates MPFactivity during mitosis as well as during the progressionfrom meiosis I to meiosis II. Since the induction ofmeiosis I in Xenopus oocytes requires protein synthesis,it is reasonable to suggest that progesterone stimulationleads to the synthesis of cyclin, which then interactswith stored p34cdc2 to produce active MPF. However,while newly synthesized cyclin has been detected incleaving embryos (Murray and Kirschner, 1989), it hasnot been observed prior to completion of the firstmeiotic division (Minshull et al. 1989). On the otherhand, recent evidence indicates that Xenopus oocytescontain a store of cyclin B already complexed withp34cdc2 (see Barrett et al. 1990), analagous to thesituation previously reported in clam oocytes (Westen-dorf et al. 1989). In this case, the protein synthesisrequirement for MPF activation during meiosis I pre-sumably would involve synthesis of a protein thatsomehow activates the stored p34cdc2-cyclin complex. Alogical candidate for this activator is the c-mos protein;it is synthesized during oocyte maturation prior to MPFactivation and injection of synthetic c-mos RNA in-duces oocyte maturation (Sagata et al. 1989£>; Watanabeet al. 1989). How, then, does injection of cyclin mRNA(or cyclin protein) also activate MPF and induce oocytematuration?
An answer to this question depends to some extenton the answer to another question, what constitutesMPF activity? Biochemically, MPF is viewed as acomplex of p34cdc2 kinase and cyclin, with the formeractually exhibiting MPF activity and the latter acting toregulate that activity (Draetta et al. 1989; Meijer et al.1989; Murray and Kirschner, 1989; Westendorf et al.1989). Biologically, MPF is defined as an activity thatinduces GVBD in the absence of progesterone and inthe absence of protein synthesis (Cyert and Kirschner,1988). Thus, in a system in which both components pre-exist, injection of either individually might be expectedto exhibit MPF activity provided not all of the storedcomponents exist in a stable complex.
Stage 4 oocytes contain pre-MPF, which can beactivated by injection of small amounts of active MPFand which can be amplified by repeated serial transfersof cytoplasm (Taylor and Smith, 1987). Presumably,this means that stage 4 oocytes contain both p34c
(this study) and some cyclin, as appears to be the case instage 6 oocytes. Thus, assuming that the level of p34cdc2
in both stage 4 and stage 6 oocytes is in excess relative tocyclin, the protein translated from injected cyclinmRNA would interact with p34cdc2, initiating oocytematuration. As pointed out by Westendorf et al. (1989),the cytoplasmic environment is modified in a number ofways during maturation and these changes could resultin the activation of stored pre-MPF. The practical
implication of such a suggestion is that amplification ofMPF as assayed biologically would be a consequence ofthe induction of maturation as opposed to a directaction of small amounts of active MPF or stored pre-MPF.
The above suggestion that injected cyclin mRNAleads to active MPF by interaction with free p34cdc2
raises certain questions regarding the role of c-mosprotein in both the activation and stabilization of MPFactivity. It is not clear that stage 6 oocytes induced tomature by the product of injected cyclin mRNA everdevelop CSF since it is not certain such oocytes progressto metaphase II (Swenson et al. 1986). However,oocytes induced to mature by injection of smallamounts of active MPF do arrest at metaphase II.Presumably, if c-mos synthesis is required for sucharrest, then injected MPF must also induce the trans-lation of endogenous c-mos mRNA. On the other hand,c-mos protein presumably is not present in stage 4oocytes (Watanabe et al. 1989) and we have observedthat injection of c-mos mRNA into stage 4 oocytes doesnot activate MPF (Xu and Smith, unpublished data).Nevertheless, a relatively stable MPF activity can beobtained from stage 4 oocytes injected with cyclinmRNA. This implies that c-mos protein does not actdirectly on p34cdc2 or cyclin. Perhaps c-mos acts on theXenopus homolog of another gene product known to beinvolved in p34 function in yeast such as weel + orniml+ (Russell and Nurse, I987a,b). To be consistentwith the present study, we need postulate that suchproteins are present only in stage 6 oocytes. In such acase, the stage 4 oocyte may turn out to be a usefulsystem to address many of these kinds of questions sinceevents leading to MPF activation can be separated fromother progesterone-induced changes leading to matu-ration in stage 6 oocytes.
We are indebted to Dr J. Ruderman of Duke University forthe generous gift of plasmids, and the communication ofunpublished data. We are also grateful to Dr G. Draetta ofCold Spring Harbor for the gift of anti cdc2 antibody. A. D. J.would like to thank the members of our laboratory for theirhelp, particularly during the completion of this project, andalso Dr Rob Steele, and Tom Unger of UCI for many helpfuldiscussions during the course of this work. A.D.J. is verygrateful to Dr Paul Krieg for his help and generosity duringthe preparation of this manuscript. This research was sup-ported by NIH grant HD 04229 awarded to L.D.S.
References
BARRETT, C. B., SCHROETKE, R. M., VAN DER HOORN, F. A.,NORDEEN, S. K. AND MALLER, J. L. (1990). Ha-rasv""2-IhrW
activated S6 kinase and p34cdc2 kinase in Xenopus oocytes:Evidence for c-mosxe-dependent and independent pathways.Molec. cell. Biol. 10, 310-315.
CYERT, M. S. AND KIRSCHNER, M. W. (1988). Regulation of MPFactivity in vitro. Cell 53, 185-195.
DRAETTA, G., LUCA, F., WESTENDORF, J., BRIZUELA, L.,RUDERMAN, J. AND BEACH, D. (1989). cdc2 is complexed withboth cyclin A and B: evidence for inactivation of MPF byproteolysis. Cell 56, 829-838.
DRURY, K. C. (1978). Method for the preparation of maturation

156 A. D. Johnson and L. D. Smith
promoting factor (MPF) from in vitro matured oocytes ofXenopus laevis. Differentiation 10, 181-186.
DRURY, K. C. AND SCHORDERET-SLATKINE, S. (1975). Effects ofcycloheximide on the "autocatalytic" nature of the maturationpromoting factor in oocytes of Xenopus laevis. Cell 4, 269-274.
DUNPHY, W. G. AND NEWPORT, J. W. (1989). Fission yeast pl3blocks mitotic activation and tyrosine dephophorylation of theXenopus cdc2 protein kinase. Ceil 58, 181-191.
EVANS, T., ROSENTHAL, E. T., YOUNGBLOOM, J., DISTEL, D. AND
HUNT, T. (1983). Cyclin: a protein specified by maternal mRNAin sea urchin eggs that is destroyed at each cell division. Cell 33,389-396.
FEUX, M.-A., PINES, J., HUNT, T. AND KARSENTI, E. (1989). A
post-ribosomal supernatant from activated Xenopus eggs thatdisplays post-translationally regulated oscillation of its cdc2+mitotic kinase activity. EMBO J. 8, 3059-3069.
FORD, C. C. AND GURDON, J. B. (1977). A method for enucleatingoocytes of Xenopus laevis. J. Embryol. exp. Morph. 37, 203-209.
GAUTIER, J., MATSUKAWA, T., NURSE, P. AND MALLER, J. (1989).
Dephosphorylation and activation of Xenopus p34 cdc2 proteinkinase during the cell cycle. Nature 339, 626-629.
GAUTIER, J., MINSHULL, J., LOHKA, M., GLOTZER, M., HUNT, T.
AND MALLER, J. L. (1990). Cyclin is a component of MPF fromXenopus. Cell (in press).
GERHART, J., W U , M., CYERT, M. AND KIRSCHNER, M. (1985). M-
phase promoting factors from eggs of Xenopus laevis. Cytobios43, 335-347.
GERHART, J., Wu, M. AND KIRSCHNER, M. (1984). Cell cycledynamics of an M-phase-specific cytoplasmic factor in Xenopuslaevis oocytes and eggs. / . Cell Biol. 98, 1247-1255.
HORRELL, A . , SHUTTLEWORTH, J. AND COLMAN, A . (1987).
Transcript levels and translational control of hsp70 synthesis inXenopus oocytes. Genes and Dev. 1, 433-444.
KRIEG, P. A. AND MELTON, D. A. (1984). Functional messengerRNAs are produced by SP6 transcription of cloned cDNAs.Nucleic Acids Res. 12, 7057-7071.
LABBE, J . -C , CAPONY, J.-P., CAPUT, D., CAVADORE, J . -C ,
DERANCOURT, J., KAGHAD, M., LELIAS, J.-M., PICARD, A. AND
DOREE, M. (1989). MPF from starfish oocytes at first meioticmetaphase is a heterodimer containing one molecule of cdc2 andone molecule of cyclin B. EMBO J. 8, 3053-3058.
LOHKA, M. J., HAYES, M. K. AND MALLER, J. L. (1988).
Purification of maturation-promoting factor, an intracellularregulator of early mitotic events. Proc. natn. Acad. Sci. U.S.A.85, 3009-3013.
LUCA, F. C. AND RUDERMAN, J. V. (1989). Control of programmedcyclin destruction in a cell free system. J. Cell Biol. 109,1895-1910.
MASUI, Y. AND MARKERT, C. (1971). Cytoplasmic control of nuclearbehavior during meiotic maturation of frog oocytes. J. exp. Zool.177, 129-146.
MASUI, Y. AND SHIBUYA, E. K. (1987). Development of cytoplasmicactivities that control chromosome cycles during maturation ofamphibian oocytes. In Molecular Regulation of Nuclear Events inMitosis and Meiosis. (ed. R. A. Schlegal, M. S. Halleck, and P.N. Rao), pp. 1-42. Academic Press, Orlando, Fl.
MEJLIER, L., ARION, D., GOLSTEYN, R., PINES, J., BRIZUELA, L.,
HUNT, T. AND BEACH, D. (1989). Cyclin is a component of thesea urchin egg M-phase specific histone HI kinase. EMBO J. 8,2275-2282.
MINSHULL, J., BLOW, J. J. AND HUNT, T. (1989). Translation of
cyclin mRNA is necessary for extracts of activated Xenopus eggsto enter mitosis. Cell 56, 947-956.
MORENO, S., HAYLES, J. AND NURSE, P. (1989). Regulation of
p34cdc2 protein kinase during mitosis. Cell 58, 361-372.MURRAY, A. W. AND KIRSCHNER, M. W. (1989). Cyclin synthesis
drives the early embryonic cell cycle. Nature 339, 275-280.MURRAY, A. W., SOLOMON, M. J. AND KIRSCHNER, M. W. (1989).
The role of cyclin synthesis and degradation in the control ofMPF activity. Nature 339, 280-286.
PINES, J. AND HUNT, T. (1987). Molecular cloning andcharacterization of the mRNA for cyclin from sea urchin eggs.EMBOJ. 6, 2987-2995.
REYNHOUT, J. K. AND SMITH, L. D. (1974). Studies on theappearance and nature of a maturation-inducing factor in thecytoplasm of amphibian oocytes exposed to progesterone. DeviBiol. 38, 394-400.
RUSSELL, P. AND NURSE, P. (1987n). Negative regulation of mitosisby weel+ a gene encoding a protein kinase homolog. Cell 49,559-567.
RUSSELL, P. AND NURSE, P. (19876). The mitotic inducer niml+functions in a regulatory network of protein kinase homologscontrolling the initiation of mitosis. Cell 49, 569-576.
SAGATA, N., DAAR, I., OSKARSSON, M., SHOWALTER, S. AND VANDE
WOUDE, G. F. (19896). The product of the mos proto-oncogeneas a candidate "initiator" for oocyte maturation. Science 245,643-646.
SAGATA, N., WATANABE, N., VANDE WOUDE, G. F. AND IKAWA, Y.
(1989a). The c-mos proto-oncogene product is a cytostatic factorresponsible for meiotic arrest in vertebrate eggs. Nature 342,512-518.
SMITH, L. D. (1989). The induction of oocyte maturation:Transmembrane signaling events and regulation of the cell cycle.Development 107, 685-699.
SMITH, L. D. AND ECKER, R. E. (1971). The interaction of steroidswith Rana pipiens oocytes in the induction of maturation. DeviBiol. 25, 233-247.
SWENSON, K. I., FARRELL, K. M. AND RUDERMAN, J. V. (1986).
The clam embryo protein cyclin A induces entry into M phaseand the resumption of meiosis in Xenopus oocytes. Cell 47,861-870.
TAYLOR, M. A., JOHNSON, A. D. AND SMITH, L. D. (1985).
Growing Xenopus oocytes have spare translational capacity.Proc. natn. Acad Sci U.S.A 82, 6586-6589.
TAYLOR, M. A. AND SMITH, L. D. (1987). Induction of maturationin small Xenopus laevis oocytes. Devi Biol. 121, 111-118.
WALLACE, R. A., JARED, D. W., DUMONT, J. N. AND SEGA, M. W.
(1973). Protein incorporation by isolated amphibian oocytes. III.Optimum incubation conditions. J. exp. Zool. 18, 321-334.
WASSERMAN, W. J. AND MASUI, Y. (1975). Effects of cycloheximideon a cytoplasmic factor initiating meiotic maturation in Xenopusoocytes. Expl Cell Res. 91, 381-388.
WASSERMAN, W. J. AND SMITH, L. D. (1978). The cyclic behavior ofa cytoplasmic factor controlling nuclear membrane breakdown. J.Cell Biol. 78, R15-R22.
WATANABE, N., VANDE WOUDE, G. F., IKAWA, Y. AND SAGATA, N.(1989). Specific proteolysis of the c-mos proto-oncogene productby calpain on fertilization of Xenopus eggs. Nature 342, 505-511.
WESTENDORF, J. M., SWENSON, K. I. AND RUDERMAN, J. V. (1989).
The role of cyclin B in meiosis I. J. Cell Biol. 108, 1431-1444.Wu, M. AND GERHART, J. C. (1980). Partial purification and
characterization of the maturation-promoting factor from eggs ofXenopus laevis. Devi Biol. 79, 465-477.
(Accepted 8 February 1990)