in situ localization and in vitro induction of plant copi ... · in situ localization and in vitro...
TRANSCRIPT

The Plant Cell, Vol. 12, 2219–2235, November 2000, www.plantcell.org © 2000 American Society of Plant Physiologists
In Situ
Localization and in Vitro
Induction of PlantCOPI-Coated Vesicles
Peter Pimpl,
a
Ali Movafeghi,
a
Sean Coughlan,
b
Jürgen Denecke,
c
Stefan Hillmer,
a,1
and David G. Robinson
a,1,2
a
Department of Structural Cell Physiology, Albrecht-von-Haller Institute of Plant Sciences, University of Göttingen, Germany
b
AG Biotechnology, DuPont Experimental Station, P.O. Box 0402, Wilmington, Delaware 19880-0402
c
Leeds Institute for Plant Biotechnology and Agriculture, University of Leeds, Leeds LS2 9JT, United Kingdom
Coat protein (COP)–coated vesicles have been shown to mediate protein transport through early steps of the secretorypathway in yeast and mammalian cells. Here, we attempt to elucidate their role in vesicular trafficking of plant cells, us-ing a combined biochemical and ultrastructural approach. Immunogold labeling of cryosections revealed that COPIproteins are localized to microvesicles surrounding or budding from the Golgi apparatus. COPI-coated buds primarilyreside on the
cis
-face of the Golgi stack. In addition, COPI and Arf1p show predominant labeling of the
cis
-Golgi stack,gradually diminishing toward the
trans
-Golgi stack. In vitro COPI-coated vesicle induction experiments demonstratedthat Arf1p as well as coatomer could be recruited from cauliflower cytosol onto mixed endoplasmic reticulum (ER)/Golgi membranes. Binding of Arf1p and coatomer is inhibited by brefeldin A, underlining the specificity of the recruitmentmechanism. In vitro vesicle budding was confirmed by identification of COPI-coated vesicles through immunogoldnegative staining in a fraction purified from isopycnic sucrose gradient centrifugation. Similar in vitro induction experi-
ments with tobacco ER/Golgi membranes prepared from transgenic plants overproducing barley
a
-amylase–HDELyielded a COPI-coated vesicle fraction that contained
a
-amylase as well as calreticulin.
INTRODUCTION
Protein trafficking in the secretory and endocytotic path-ways is facilitated by coated vesicles (Rothman andWieland, 1996; Robinson et al., 1998). Research on mam-malian and yeast cells has established that two differenttypes of non-clathrin-coated vesicles are responsible fortransport events occurring between the endoplasmic reticu-lum (ER) and the Golgi apparatus and for intra-Golgi trans-port. Whereas investigators generally agree that COPII–coated vesicles bud from the ER and transport proteins inthe anterograde direction (Barlowe et al., 1994; Bannykhand Balch, 1998), the function of COPI-coated vesicles re-mains controversial. Strong evidence suggests that COPI-coated vesicles are responsible for the recycling of escapedER-resident proteins from post-ER compartments (Letourneuret al., 1994), but data have also been presented supporting arole in anterograde transport
of proteins, especially within theGolgi stack (Orci et al., 1997; Lippincott-Schwartz et al., 1998).
The formation of both types of COP-coated vesicles in-volves the recruitment of a coat protein complex (coatomer
for COPI-coated vesicles, Sec23/24 and Sec13/31 dimersfor COPII-coated vesicles) by a membrane-associated GTPbinding protein (Arf1p and Sar1p for COPI and COPII, re-spectively; Schekman and Orci, 1996). Both Arf1p andSar1p exist in the cytosol as GDP-bound forms and becomeconverted to the GTP form through the action of a mem-brane-associated exchange factor, guanine nucleotide ex-change factor (Helms and Rothman, 1992). For Sar1p, this isSec12p, a transmembrane protein of the ER (Barlowe andSchekman, 1993); for Arf1p, the most likely candidates areGea1/2p and ARNO (ARF nucleotide binding site opener;Chardin et al., 1996; Peyroche et al., 1996). Regarding COPI-coated vesicles, coatomer also interacts with the cytoplasmicdomain of two types of transmembrane proteins: either a mem-ber of the p23/24 protein family (Bremser et al., 1999; Lavoie etal., 1999) or a dilysine motif present on type I membrane-span-ning ER resident proteins (Cosson and Letourneur, 1994).
Coatomer has been isolated from mammalian (Waters etal., 1991) and yeast (Duden et al., 1994) cells and consists of
seven polypeptides:
a
-COP (Ret1p), 135 kD;
b
-COP (Sec26p),107 kD;
b
’-COP (Sec27p), 102 kD;
g
-COP (Sec21p), 97 kD;
d
-COP (Ret2p), 57 kD;
«
-COP (Sec28p), 36 kD; and
z
-COP(Ret3p), 21 kD.
g
-COP contains a binding site for both dily-
sine retrieval motifs and the diphenylalanine motif of p23 (Harterand Wieland, 1998). Structural homologies but not func-tional similarities exist between
b
-COP and the
b
-adaptins
1
Current address: Zellenlehre, University of Heidelberg, Im Neuen-heimer Feld 230, D-69120 Heidelberg, Germany.
2
To whom correspondence should be addressed. E-mail [email protected]; fax 49-6221-5464.

2220 The Plant Cell
of clathrin-coated vesicles (Duden et al., 1991), between
d
-COP and the
m
-adaptins (Faulstich et al., 1996), and be-tween
z
-COP and the
s
-adaptins (Kuge et al., 1993).Coatomer is assembled in the cytosol from its subunit
polypeptides within 1 to 2 hr after their translation, is re-cruited onto membranes (Hara-Kuge et al., 1994), and is astable structure for
.
50 hr (Lowe and Kreis, 1996), whichsuggests that the protein can be used in many cycles ofvesicle formation. The initial binding of one coatomer to the
cytoplasmic domain of p23 (in a tetrapeptide conformation)apparently induces the attachment of further coatomers andis considered the underlying mechanism for COPI-coatedvesicle formation (Harter, 1999). As shown by immunogoldelectron microscopy, the membrane-bound coatomer inmammalian cells is localized mainly to the periphery of Golgicisternae (Duden et al., 1991; Oprins et al., 1993; Orci et al.,1997) and occasionally to special domains of the ER (Orci etal., 1994). In addition, coatomer is prominent on membranesof the so-called ER–Golgi intermediate compartment (ERGIC)(Oprins et al., 1993; Griffiths et al., 1995; Martinez-Menarguezet al., 1999), a compartment for which plants have no directstructural counterpart.
Plant
ARF1
,
Sar1
,
and
Sec12
genes and proteins havebeen identified (d’Enfert et al., 1992; Regad et al., 1993;Bar-Peled and Raikhel, 1997). In addition, knowledge on thetargeting and fusion machinery for transport vesicles inplants is rapidly expanding (reviewed in Robinson et al.,1998; Sanderfoot and Raikhel, 1999). Recently, we pre-sented evidence for the existence of homologs of
g
-COP(Sec21p) and Sec23p in cauliflower inflorescence andshowed that these polypeptides are part of cytosolic com-plexes having the properties of coatomer and the Sec23/24dimer (Movafeghi et al., 1999). Nevertheless, plant COPI-and COPII-coated vesicles have yet to be identified in situand isolated. Isolation of such vesicles would facilitate theanalysis of their biochemical composition, which in turnwould help resolve the controversy over cargo selection andbulk flow models (Vitale and Denecke, 1999). Here, in animportant step toward achieving this aim, we present bio-chemical and ultrastructural evidence for coatomer recruit-ment to donor membranes and COPI-coated vesiclebudding in vitro.
RESULTS
Identification of Plant
d
- and
«
-COP Homologs and Cross-Reactivity of Antisera
Maize homologs of the coatomer components
d
- and
«
-COPwere identified from the DuPont database of expressed se-quence tags by homology with known sequences in Gen-Bank. At the protein level, the 58-kD
d
-COP was 88%identical and 91% similar to the previously elucidated
d
-COPsequence for rice in GenBank (accession number E209222).The 31-kD maize
«
-COP was 77% identical and 90% similarto the Arabidopsis sequence (GenBank accession numberAX004238). Interestingly, both of the higher plant
d
-COP se-quences have greater similarity to the higher eukaryoticsequences (human
d
-COP, X81197; and bovine
d
-COP,X94265) than to the putative yeast
d
-COP (yfr05c). Similarly,the higher plant
«
-COP sequences were
z
45% identical to
Figure 1. Cross-Reactivities of Antisera Raised against Recombi-nant Proteins Prepared from cDNA Clones for Atg-, Zmd-, and «-COP,AtARF1p, and AtSar1p.
(A) Cytosolic proteins and total membranes from the organisms indi-cated were prepared as described in Methods and probed with theabove antisera. Equal amounts of protein were added to each lane(30 mg). c, cytosol; m, membrane.(B) Complete lanes and molecular masses of the Arabidopsis cyto-sol and membranes indicate specific recognition of a single antigenof the corresponding size for each of the above-listed antisera.
Plant Coatomer 2221
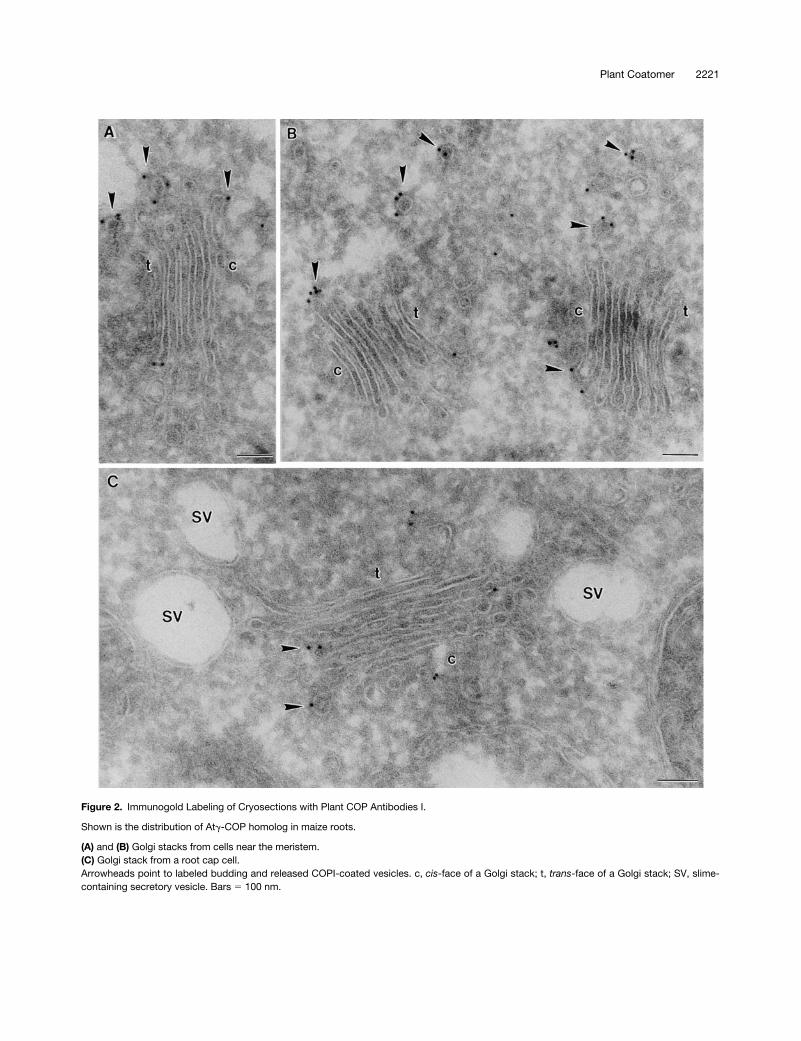
Figure 2. Immunogold Labeling of Cryosections with Plant COP Antibodies I.
Shown is the distribution of Atg-COP homolog in maize roots.
(A) and (B) Golgi stacks from cells near the meristem.(C) Golgi stack from a root cap cell.Arrowheads point to labeled budding and released COPI-coated vesicles. c, cis-face of a Golgi stack; t, trans-face of a Golgi stack; SV, slime-containing secretory vesicle. Bars 5 100 nm.
2222 The Plant Cell
the higher eukaryotic
«
-COP protein sequences in the publicdatabase.
The two plant COP-coated vesicle GTPases showed evengreater homology with their yeast and mammalian counter-parts. Thus, the Arabidopsis
ARF1
sequence (M95166) en-codes a protein that is 78% identical to yeast ARF1(B36167) and 88% identical to mouse (JC4945) and human(A33283) ARF1. Arabidopsis Sar1p (M95795) is somewhat lesshomologous, being 64% identical to Sar1p from yeast (A33619)and 59% identical to Sar1p from mouse (S39543).
Polyclonal antibodies raised against recombinant Zm
d
-and
«
-COPs recognize polypeptides of the expected molec-ular masses (58 and 31 kD, respectively) in total membraneand cytosolic extracts from roots of maize and Arabidopsis
as well as from cauliflower inflorescence (Figure 1A). In pro-tein gel blots with equal amounts of protein, the signal in thecytosolic fractions was always stronger than in the mem-brane fractions. With the GTPase antisera, however, the sig-nal for AtArf1p was roughly similar for both fractions, andthat for AtSar1p was even stronger in the membrane frac-tions. A similar distribution has been reported for Sar1p inyeast (Barlowe et al., 1993). Interestingly, overexpression ofSar1p in Arabidopsis alters this relationship, such that con-siderably more AtSar1p is present in the cytosol than on themembranes (Bar-Peled and Raikhel, 1997).
Although we had previously obtained weak cross-reac-tions with AtSec23p antibodies against polypeptides presentin membrane or cytosolic fractions prepared from yeast and
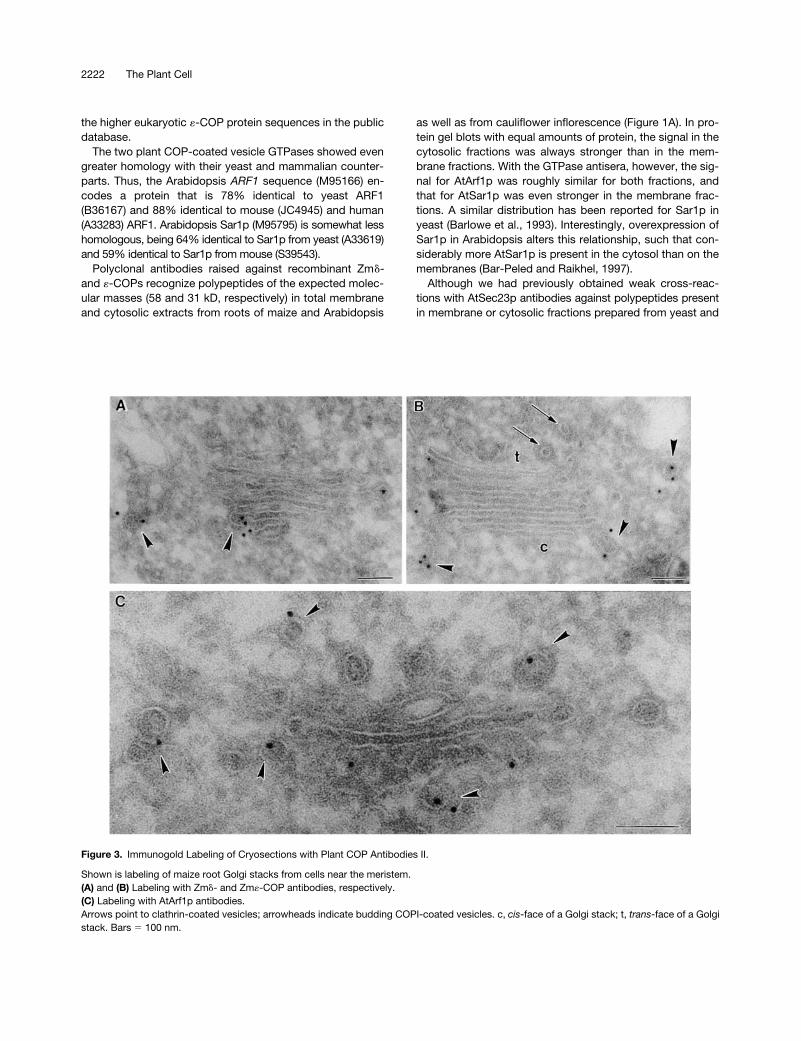
Figure 3. Immunogold Labeling of Cryosections with Plant COP Antibodies II.
Shown is labeling of maize root Golgi stacks from cells near the meristem.(A) and (B) Labeling with Zmd- and Zm«-COP antibodies, respectively.(C) Labeling with AtArf1p antibodies.Arrows point to clathrin-coated vesicles; arrowheads indicate budding COPI-coated vesicles. c, cis-face of a Golgi stack; t, trans-face of a Golgistack. Bars 5 100 nm.

Plant Coatomer 2223
pig brain (Movafeghi et al., 1999), none of the plant COPI orGTPase antisera reacted in this way (Figure 1A). This suggeststhat the major antigenic determinants are localized to nonho-mologous domains of COP-vesicle coat proteins. In addition,the antibodies recognized only one defined band (Figure 1B),confirming their suitability for immunocytochemistry.
Localization of Plant Coatomer
Because attempts to localize membrane-bound plant COPsby immunogold labeling of sections prepared from chemi-cally fixed and plastic-embedded material were repeatedlyunsuccessful, we applied the more sensitive Tokuyasu tech-nique (Tokuyasu, 1986). Here, we present localization dataon root tip cells from Arabidopsis and maize, the two organ-isms from which cDNAs were used to generate recombinantproteins and subsequently antibodies.
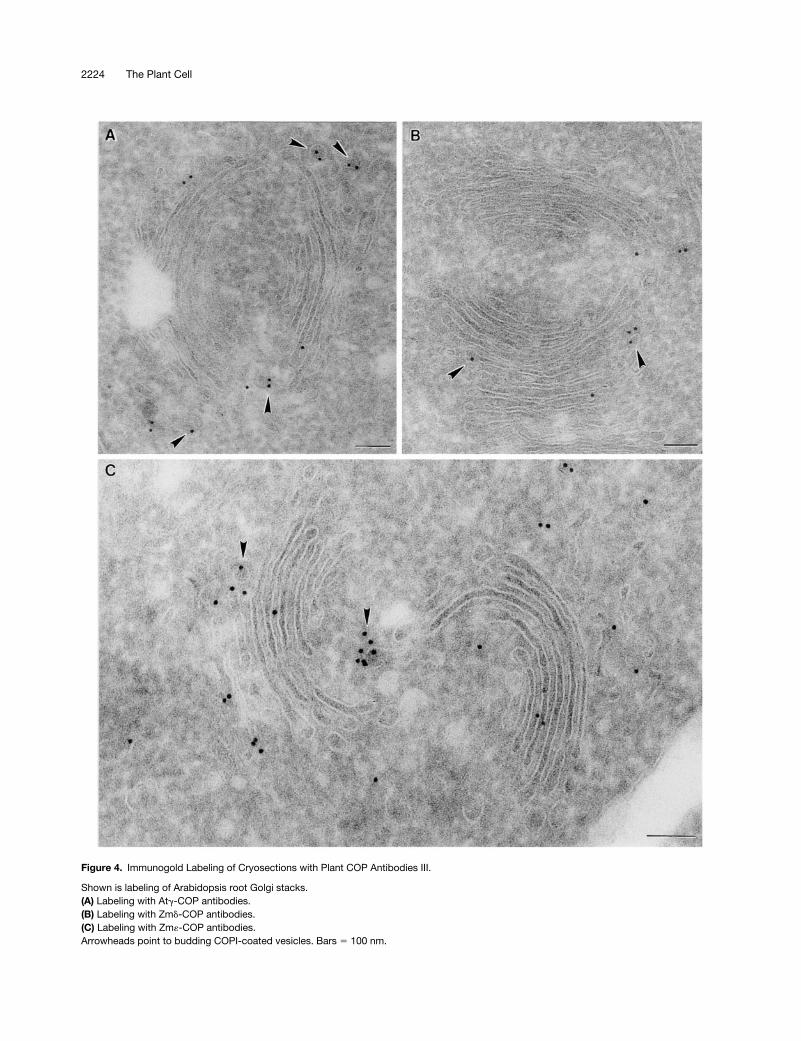
Immunogold labeling with all of the COP antisera (At
g
-COP, Zm
d
-COP, Zm
«
-COP) and by Arf1p antibodies on theroot tip cells of both species was highly specific: gold parti-cles were found only on, or in the immediate vicinity of,Golgi stacks (Figures 2 to 4). Gold label was generally ab-sent from the central portion of the cisternae, being insteadrestricted mostly to the rims. This peripheral labeling was of-ten very clearly directed to small (
z
60 nm in diameter) bud-ding vesicles (see Figures 2A, 2B, 3A to 3C, and 4A). Thesevesicles possessed a thin (15-nm thick), irregularly shapedcoat on their cytoplasmic surface and closely resembled theCOPI-coated vesicles depicted by this method in mamma-lian cells (see, e.g., Orci et al., 1997). Profiles of similarlycoated vesicles lying close to Golgi stacks were also labeledwith different COP antisera (Figures 2B and 3B). Those vesi-cles differed from clathrin-coated vesicles, which had athicker (25 to 30 nm) coat and a
trans
-Golgi position (Figure3A). Neither the clathrin-coated vesicles nor the large, slime-filled vesicles (Figure 2C) typical of the Golgi apparatus inmaize root cap cells (Mollenhauer and Morré, 1994) were la-beled by COPI-immunogold.
To ascertain the localization of coatomer within the plantGolgi stack, we statistically analyzed the immunogold-COPIlabeling on a large number of dictyosomes (Table 1). Forthis, we counted gold particles on budding vesicles still at-tached to cisternae as well as on putative COPI-coated ves-icles lying in the areas adjacent to the polar
trans
-face andon the respective
cis
- and
trans
-sectors around the Golgistack. It was immediately apparent that the majority of goldparticles are associated with coated vesicles with no obvi-ous cisternal attachment. However, we cannot exclude thepossibility that some of these vesicles may have connec-tions to cisternae of neighboring Golgi stacks that are out ofthe plane of the section. Nevertheless, such “released” ves-icles were rarely detected at the
trans-
face, and thosepresent in the
cis
-sector slightly outnumbered those in the
trans
-sector. On the other hand, the distribution of cisternal
rim COPI labeling was clearly weighted toward the
cis
sideof the Golgi stack, such that the most label was associatedwith the
cis
-most cisternae.
Selective Removal of Membrane-AssociatedCOP-Vesicle Coat Proteins but Not AtArf1p Homolog
Because we had previously characterized At
g
-COP andAtSec23p homologs from cauliflower inflorescence, whichhas decided advantages as a potential source of plantcoatomer, we continued to use this tissue in our biochemi-cal work. To ascertain whether cytosolic nonclathrin coatprotein complexes from cauliflower florets can be recruitedonto endomembranes from the same tissue, we first had todetermine the conditions under which endogenous COP-vesicle coat proteins could be removed from the membranes.For this, we used the chaotrope KI, which has previouslybeen used successfully to gently dissociate nonintegral pro-teins from plant membranes (Hinz et al., 1997). As seen inFigure 5A, a 30-min treatment with 300 mM KI is sufficient toquantitatively dissociate both the At
g
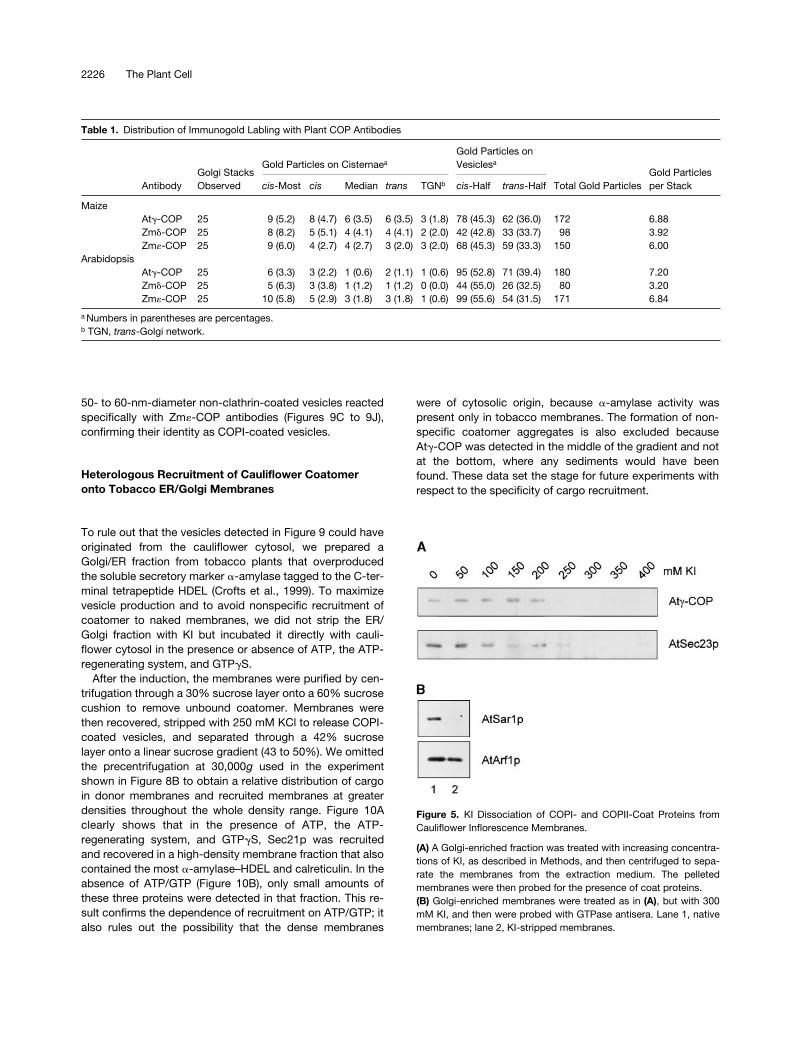
-COP and AtSec23p ho-molog proteins from cauliflower membranes. The antiseraprepared against these two COP-coat polypeptides recog-nize protein complexes in cauliflower cytosol correspondingto coatomer and the Sec23/24 dimer of the COPII coat(Movafeghi et al., 1999). The membrane fraction used was en-riched in Golgi membranes, as shown previously (Movafeghiet al., 1999). Although coatomer and the COPII coat were ef-fectively removed from cauliflower membranes through KItreatment, the two GTPases known to be involved in COP-vesicle coat protein recruitment were affected differently. Thecauliflower AtArf1p homolog was not dissociated by 300 mMKI, whereas the AtSar1p homolog was completely removedfrom the membranes (Figure 5B). This difference most proba-bly indicates that Arf1p, but not Sar1p, is anchored into themembrane by its myristoylated N terminus (Kahn et al., 1992).
Selective Recruitment of Coatomer but not ofAtSec23/24 Complex
To reconstitute the first step of vesicle budding, we incu-bated KI-stripped cauliflower membranes with concentratedcytosolic proteins in the presence of GTP
g
S and an ATP-regenerating system. Membrane recruitment was monitoredby centrifugation through a sucrose step gradient in whichmembranes and recruited proteins are obtained in the cen-trifuged pellet whereas unbound proteins remain in the su-pernatant. These two fractions were probed for GTPases,coatomer, and the Sec23/24 complex. Our results show thatboth the cauliflower Arf1p and Sar1p homologs and the Sec21homolog accumulated in the pellets as a result of the incubation(Figures 6A and 6B). In particular, all three cytosolic componentswere depleted from the cytosol (cf. Figure 1A cauliflower with
2224 The Plant Cell
Figure 4. Immunogold Labeling of Cryosections with Plant COP Antibodies III.
Shown is labeling of Arabidopsis root Golgi stacks.(A) Labeling with Atg-COP antibodies.(B) Labeling with Zmd-COP antibodies.(C) Labeling with Zm«-COP antibodies.Arrowheads point to budding COPI-coated vesicles. Bars 5 100 nm.

Plant Coatomer 2225
that in Figures 6A and 6B), suggesting a highly efficient recruit-ment process. In contrast to coatomer, however, the Sec23/24complex was not recruited and sedimented, even though Sar1pwas (Figures 6A and 6B). The substitution of GMP-PNP (5-guanylyl-imidodiphosphate), another nonhydrolyzable analog ofGTP, for GTP
g
S in the reaction mixture (Barlowe et al.,1994) also failed to induce recruitment of the AtSec23/24dimer (data not shown). The lack of Sec23p sedimentationagain suggests that recruitment of Sec21p is specific.
To rule out nonspecific sedimentation of aggregates as anexplanation for the intensifying effect on the bands ob-served for the membrane pellets, we repeated the experi-ment under the same conditions but omitting membranes.Figure 6C clearly shows that cytosol itself does not containsedimentable coatomer, Arf1p, or Sar1p, confirming thatsedimentation must have occurred as a result of membranerecruitment and sedimentation of the membrane.
To determine the saturability of the coatomer association,we performed recruitment assays with various ratios of cy-tosolic to membrane proteins. Even with fourfold more cyto-solic protein than membrane protein, the coatomer bindingwas still unsaturated (Figure 6C). This indicates that theconcentration of coat components in the cytosolic fractionwas probably a limiting factor in the recruitment assay.
Although cauliflower cytosol represents a nearly optimalsource of plant coatomer, easily transformed plants such astobacco will be more important for answering questions asto the nature of the cargo and the direction of COP-coatedvesicle transport. Accordingly, we attempted the heterolo-gous recruitment of coatomer from cauliflower cytosol ontomembranes isolated from tobacco mesophyll. We prepareda Golgi/ER fraction from tobacco leaves, stripped the en-dogenous coat proteins with KI, and performed an in vitrorecruitment with cauliflower cytosol, as described above.Figure 6D convincingly demonstrates the interchangeabilityof coatomer among dicotyledonous plants.
Inhibition of Coat Protein Recruitment by Brefeldin A
Treating plant and animal cells with the fungal metabolitebrefeldin A (BFA) is known to inhibit anterograde proteintransport and results in incorporation of the Golgi mem-branes and content into the ER (Klausner et al., 1992;Boevink et al., 1998). Studies on mammalian cells have es-tablished that BFA interacts with guanine nucleotide ex-change factor (Helms and Rothman, 1992), thereby inhibitingArf1p recruitment onto Golgi membranes and thus prevent-ing coatomer attachment (Dascher and Balch, 1994). For rea-sons not entirely understood, there is a discrepancy betweenthe concentrations of BFA that are effective in vivo
(
z
20
m
M)and in vitro
(200
m
M) in mammalian cells (Torii et al., 1995;Peyroche et al., 1999). In plant cells, the concentration ofBFA required to elicit a response in vivo
usually is between 2and 100
m
M (Satiat-Jeunemaitre et al., 1995). As shown in
Figure 7, concentrations
.200 mM are required to inhibit thein vitro recruitment of AtArf1p and coatomer. These resultsalso confirm the specificity of the recruitment reaction.
COPI-Coated Vesicle Formation in Vitro
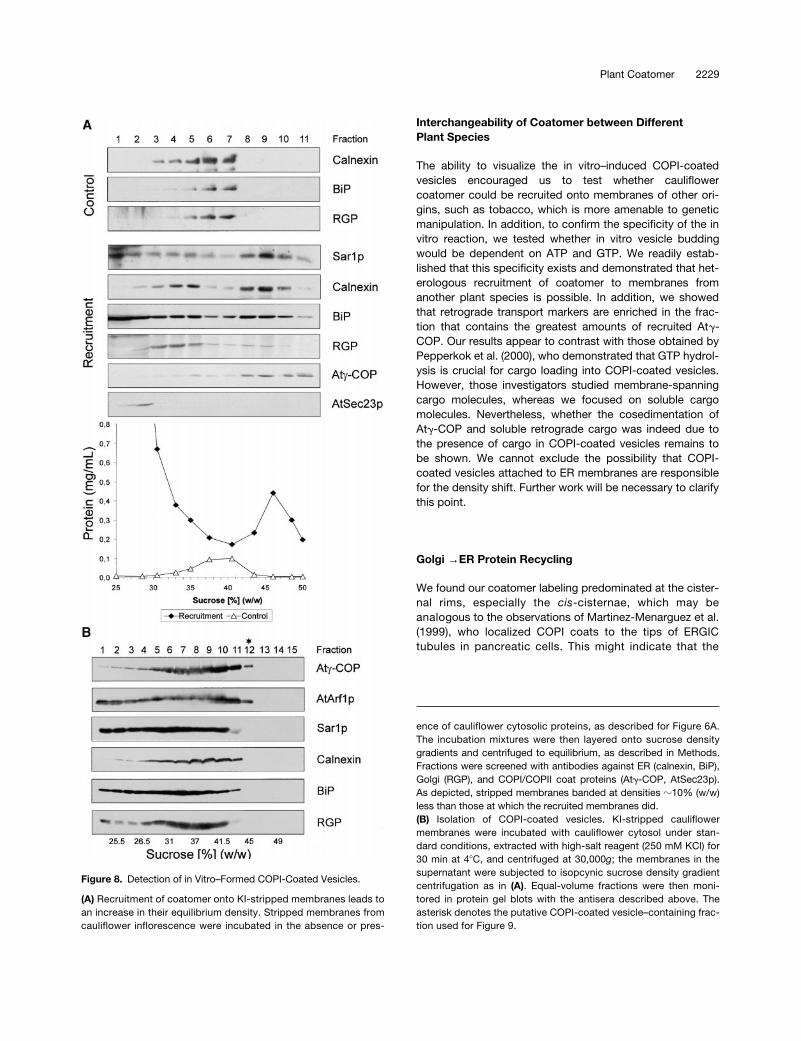
KI-stripped Golgi-enriched membrane fractions from cauli-flower inflorescence have a higher isopycnic density in su-crose after being incubated with cytosolic proteins in thepresence of GTPgS (Figure 8A). Compared with control mem-branes exposed to buffer and GTPgS, incubation with cytosolicproteins led to a density increase of 8 to 10% (sucrose equiva-lents). This shift in density could be attributed to the recruitmentof AtArf1p and coatomer onto Golgi membranes. Although theAtSec23p homolog stayed in the overlay, the attachment ofAtSar1p homolog to the ER could also result in such a shift. In-deed, protein gel blots with antisera against suitable markerproteins showed that Golgi membranes, as judged by the Golgimarker reversibly glycosylated protein (RGP) (Movafeghi et al.,1999) and the ER markers binding protein (BiP) and calnexin,shifted to greater densities. However, we considered recruit-ment of the AtSar1p homolog to be insufficient to cause such amassive increase in isopycnic density of the ER. Instead, wesurmised that this shift might have resulted from the attachmentof Golgi-derived COPI-coated vesicles, which, because oftheir Arf1p-bound GTPgS, cannot dissociate their coat and aretherefore unable to fuse. This, in fact, was the basic premisethat led to the first successful isolation of COPI-coated vesiclesfrom mammalian cells (Malhotra et al., 1989).
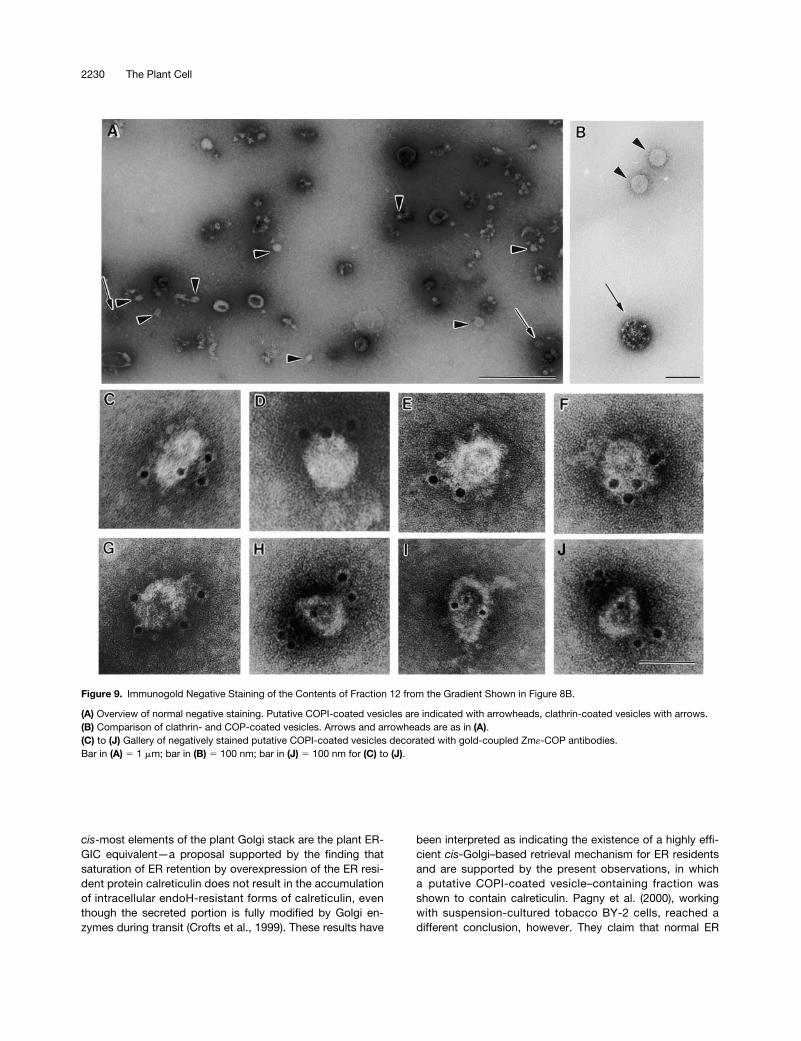
To release bound COPI-coated vesicles, we thereforetreated recruited cauliflower membranes with 250 mM KCl.The high-salt mixture was then centrifuged at 30,000g for30 min to remove most of the ER and Golgi membranes.Membranes in the 30,000g supernatant were subsequentlysubjected to an isopycnic sucrose density gradient centrifuga-tion. As shown in Figure 8B, despite the precentrifugation at30,000g, considerable amounts of Golgi and ER markers werestill carried over in the high-salt extract. Nevertheless, at leastone fraction (no. 12, at 45% sucrose) exhibited a clear signal inprotein gel blots with AtArf1p and Atg-COP antisera; this den-sity corresponded to that reported for mammalian COPI-coatedvesicles (42 to 45% [w/w] sucrose) (Malhotra et al., 1989). Frac-tion 12 did not, however, contain detectable amounts of BiP,calnexin, or Golgi (RGP) proteins. Therefore, we further ana-lyzed this fraction by negative staining as well as by immu-nogold negative staining because the very low amounts ofprotein precluded examination by sectioning procedures.
For the immunogold labeling, we used Zm«-COP antibod-ies, since «-COP is more exposed at the surface of coatomerthan are the other COPs (Duden et al., 1998). Using this methodof visualization, we found that the fraction was not homoge-neous. In addition to small numbers of 50- to 60-nm-diam-eter vesicles, fraction 12 contained undefined particleaggregates and small membrane fragments as well as clath-rin-coated vesicles (Figures 9A and 9B). However, only the
2226 The Plant Cell
50- to 60-nm-diameter non-clathrin-coated vesicles reactedspecifically with Zm«-COP antibodies (Figures 9C to 9J),confirming their identity as COPI-coated vesicles.
Heterologous Recruitment of Cauliflower Coatomer onto Tobacco ER/Golgi Membranes
To rule out that the vesicles detected in Figure 9 could haveoriginated from the cauliflower cytosol, we prepared aGolgi/ER fraction from tobacco plants that overproducedthe soluble secretory marker a-amylase tagged to the C-ter-minal tetrapeptide HDEL (Crofts et al., 1999). To maximizevesicle production and to avoid nonspecific recruitment ofcoatomer to naked membranes, we did not strip the ER/Golgi fraction with KI but incubated it directly with cauli-flower cytosol in the presence or absence of ATP, the ATP-regenerating system, and GTPgS.
After the induction, the membranes were purified by cen-trifugation through a 30% sucrose layer onto a 60% sucrosecushion to remove unbound coatomer. Membranes werethen recovered, stripped with 250 mM KCl to release COPI-coated vesicles, and separated through a 42% sucroselayer onto a linear sucrose gradient (43 to 50%). We omittedthe precentrifugation at 30,000g used in the experimentshown in Figure 8B to obtain a relative distribution of cargoin donor membranes and recruited membranes at greaterdensities throughout the whole density range. Figure 10Aclearly shows that in the presence of ATP, the ATP-regenerating system, and GTPgS, Sec21p was recruitedand recovered in a high-density membrane fraction that alsocontained the most a-amylase–HDEL and calreticulin. In theabsence of ATP/GTP (Figure 10B), only small amounts ofthese three proteins were detected in that fraction. This re-sult confirms the dependence of recruitment on ATP/GTP; italso rules out the possibility that the dense membranes
were of cytosolic origin, because a-amylase activity waspresent only in tobacco membranes. The formation of non-specific coatomer aggregates is also excluded becauseAtg-COP was detected in the middle of the gradient and notat the bottom, where any sediments would have beenfound. These data set the stage for future experiments withrespect to the specificity of cargo recruitment.
Table 1. Distribution of Immunogold Labling with Plant COP Antibodies
AntibodyGolgi StacksObserved
Gold Particles on Cisternaea
Gold Particles on Vesiclesa
Total Gold ParticlesGold Particlesper Stackcis-Most cis Median trans TGNb cis-Half trans-Half
MaizeAtg-COP 25 9 (5.2) 8 (4.7) 6 (3.5) 6 (3.5) 3 (1.8) 78 (45.3) 62 (36.0) 172 6.88Zmd-COP 25 8 (8.2) 5 (5.1) 4 (4.1) 4 (4.1) 2 (2.0) 42 (42.8) 33 (33.7) 98 3.92Zm«-COP 25 9 (6.0) 4 (2.7) 4 (2.7) 3 (2.0) 3 (2.0) 68 (45.3) 59 (33.3) 150 6.00
ArabidopsisAtg-COP 25 6 (3.3) 3 (2.2) 1 (0.6) 2 (1.1) 1 (0.6) 95 (52.8) 71 (39.4) 180 7.20Zmd-COP 25 5 (6.3) 3 (3.8) 1 (1.2) 1 (1.2) 0 (0.0) 44 (55.0) 26 (32.5) 80 3.20Zm«-COP 25 10 (5.8) 5 (2.9) 3 (1.8) 3 (1.8) 1 (0.6) 99 (55.6) 54 (31.5) 171 6.84
a Numbers in parentheses are percentages.b TGN, trans-Golgi network.
Figure 5. KI Dissociation of COPI- and COPII-Coat Proteins fromCauliflower Inflorescence Membranes.
(A) A Golgi-enriched fraction was treated with increasing concentra-tions of KI, as described in Methods, and then centrifuged to sepa-rate the membranes from the extraction medium. The pelletedmembranes were then probed for the presence of coat proteins.(B) Golgi-enriched membranes were treated as in (A), but with 300mM KI, and then were probed with GTPase antisera. Lane 1, nativemembranes; lane 2, KI-stripped membranes.

Plant Coatomer 2227
DISCUSSION
COPI-Coated Vesicles and the Plant Golgi Apparatus
Several morphological and biochemical characteristics dif-ferentiate the plant Golgi apparatus from its counterpart or-ganelle in mammalian cells and yeast (Andreeva et al., 1998;Dupree and Sherrier, 1998; Robinson and Hinz, 1999). Manyof these distinguishing features relate to the predominantfunction of the plant Golgi apparatus in synthesis of cell wallpolysaccharides, especially during cell division, and to thenecessity for directing proteins to two rather than one vacu-olar compartment (Robinson and Hinz, 1997, 1999). In addi-tion, the individual Golgi stacks of the plant cell appear to bein continuous movement throughout the cytoplasm along ananastomosing network of ER/actin filaments (Boevink et al.,1998) and are generally dispersed throughout the cyto-plasm. In mammalian cells, however, the Golgi is restrictedto a defined position near the nucleus. Despite these differ-ences, plant cells are generally assumed not to differ funda-mentally from other eukaryotes in terms of ER–Golgitransport, intra-Golgi transport, and recycling (Andreeva etal., 1998; Dupree and Sherrier, 1998).
Here, we have established that COPI-coated vesicles inplants are predominantly formed by the cis- and medialGolgi cisternae, although they are also detected at the mar-gins of trans-cisternae. Coatomer is also localized to ERGICin mammalian cells (Oprins et al., 1993; Griffiths et al., 1995;Martinez-Menarguez et al., 1999). This post-ER, pre-Golgicompartment has been described morphologically as con-sisting of vesicular tubular clusters (Bannykh and Balch,1998) and is considered to be formed by the homotypic fu-sion of COPII-coated vesicles (Aridor et al., 1995; Scales etal., 1997). The COPI-coated vesicles formed from this com-partment are believed to concentrate membrane proteinsdestined for recycling to the ER (Martinez-Menarguez et al.,1999). Although plant cells have no obvious structural ho-molog to ERGIC, it is perhaps significant that the cis-mostelements of the plant Golgi apparatus often appear as a dis-continuous cisterna; this may reflect a fusion site for ER-derived COPII-coated vesicles.
Coatomer Recruitment and COPI-CoatedVesicle Formation
COPI-coated vesicle formation in mammalian cells dependson two cytosolic factors: the GTPase Arf1p and coatomer (Orciet al., 1993; Palmer et al., 1993). For coatomer recruitment,Arf1p is in a GTP-bound form, but it is reconverted to the GDPform by a GTPase-activating protein, GAP (Ostermann et al.,1993), at which point coatomer is released. Interestingly,Arf-GAP activity is stimulated by coatomer (Goldberg, 1999).GTP hydrolysis is required for coat protein dissociation(Tanigawa et al., 1993), and the inhibition of GTP hydrolysis
Figure 6. In Vitro Recruitment of Cytosolic COPI-Vesicle Coat Pro-teins onto KI-Stripped Membranes.
(A) Recruitment of GTPases onto cauliflower membranes. KI-stripped membranes were incubated with cytosol, ATP-regeneratingsystem, and GTPgS for 25 min at 208C. Membranes with bound pro-teins were separated from unbound proteins by centrifugation.Small GTPases in each fraction were detected in a protein gel blotwith specific antibodies against AtArf1p and AtSar1p. Lane 1 con-tains KI-stripped membranes before incubation with cytosolic pro-teins; lane 2, the supernatant after recruitment; and lane 3, thepelleted membranes after recruitment.(B) Selective recruitment of coatomer onto cauliflower membranes.KI-stripped membranes were incubated as described in (A), and thesediment and supernatant proteins were screened by protein gelblotting with Atg-COP and AtSec23p antibodies. Lane 1 containsnative membranes; lane 2, KI-stripped membranes; lane 3, cytosolicproteins; lane 4, supernatant after recruitment; and lane 5, pelletedmembranes after recruitment.(C) Absence of precipitable Atg-COP, AtArf1p, AtSec23p, andAtSar1p in cytosol only (left). Nonsaturation of coat protein recruit-ments onto cauliflower membranes (right). The ratio of cytosolic:mem-brane proteins in the recruitment assay was increased by 30% (lanes1 and 4), 100% (lanes 2 and 5), and 200% (lanes 3 and 6) over thestandard values (6 mg mL21:0.2 mg L21). Lanes 1 to 3 represent thesupernatant fractions after recruitment; lanes 4 to 6, the correspond-ing pelleted membrane fractions. S represents the supernatant, P thepellet after recruitment in the absence of membranes. Membranes,medium, incubation, and other conditions were as described in (A).Antibody screening was as described in (A) and (B).(D) Heterologous recruitment of COPI-vesicle coat proteins fromcauliflower cytosol onto Golgi/ER membranes from tobacco meso-phyll. Tobacco membranes were isolated as described in Methods,incubated with cauliflower cytosol, and subsequently screened withantibodies under the conditions used in (A). Lane 1 contains KI-stripped membranes; lane 2, pelleted membranes after recruitment.

2228 The Plant Cell
has been completed. Because 80% of Sar1p is membranebound (Figure 1; for yeast, see also Barlowe et al., 1993),the amount of Sar1p homolog in the cauliflower cytosolmay be insufficient to support recruitment of Sec23/24dimer.
The shift in equilibrium density of KI-stripped cauliflowermembranes after incubation with cytosol and GTPgS pointsto a massive recruitment of coat proteins. This will increasethe density of the Golgi membranes but perhaps also indi-rectly that of the ER. Golgi-derived COPI-coated vesiclesformed in the presence of GTPgS may dock onto their targetmembranes (in part, the ER), but they cannot fuse with thatmembrane because of their inability to uncoat. As shown byMalhotra et al. (1989), high-salt treatment causes the re-lease of attached, unfused COPI-coated vesicles, which canthen be separated from the donor/acceptor membranes bya combination of sequential differential and sucrose densitycentrifugations. We used conditions similar to those ofMalhotra et al. (1989) and were able to visualize COPI-coated vesicles, as defined by labeling with anti–Zm«-COPantibodies. However, we could not obtain a homogeneousCOPI-coated vesicle fraction as reported by Malhotra et al.(1989), who used a relatively pure rabbit liver Golgi fraction.The cisternal elements of the latter were easily sedimented atlow-speed centrifugation (microfuge), whereas in our experi-ments, centrifugation at 30,000g did not achieve this. Be-cause of their dispersed nature, plant Golgi membranes aredifficult to isolate in large quantities, a property that remainsan obstacle for future research with purified components.
The interaction between coatomer and dilysine motifs inthe cytosolic domain of type I ER membrane proteins hasformed the basis for the notion that COPI-coated vesiclesare involved in retrograde transport from the Golgi appara-tus (Cosson and Letourneur, 1994; Letourneur et al., 1994).Our in vitro data neither confirm nor exclude this possibility,and we point out that the identification of potential cargomolecules in COPI-coated vesicles induced in vitro is madeinherently difficult because of a peculiarity of the conditionsunder which they are formed. Nickel et al. (1998) andMalsam et al. (1999) showed that the uptake of antero-grade and retrograde integral membrane reporter mole-cules into COPI-coated vesicles was markedly diminishedwhen they were formed in the presence of GTPgS ratherthan GTP. Indeed, Malsam et al. (1999) went so far as tosuggest that, when GTP hydrolysis is prevented, coatomerbinds instantaneously, leading to the release of cargo-freeCOPI-coated vesicles. In addition, in vivo experiments witha GTPase-deficient ARFI mutant suggest that when GTPhydrolysis is prevented, cargo-free COPI-coated vesiclesare produced (Pepperkok et al., 2000). This would meanthat the in vitro approach to determining the cargo contentof plant COPI-coated vesicles can be of value only whenassays of the induction of COPI-coated vesicles are per-formed in the presence of GTP. However, such vesiclesare inherently more unstable than those produced in thepresence of GTPgS.
Figure 7. BFA Inhibits Recruitment of Coatomer and AtArf1p inCauliflower Recruitment Assay.
Bound coatomer/Arf1p in control and BFA-treated samples wasquantified by densitometry of protein gel blots of samples of equalvolume. Shown are the results of four separate experiments whoseaverage is given as columns; the error bars represent the extent ofthe variation between experiments. (1) denotes the presence of 300mM BFA and (2) the absence of BFA in the incubation media.
by GTPgS blocks uncoating and leads to the accumulationof COPI-coated vesicles in vitro (Melançon et al., 1987;Malhotra et al., 1989). Importantly, high-salt treatment ofGolgi membranes does not dissociate anchored Arf1p butdoes remove Arf-GAP and is thought to enhance even moreaccumulation of COPI-coated vesicles (Ostermann et al.,1993). For this reason, we used KI-stripped instead of nativemembranes in our first recruitment experiments.
KI-stripped cauliflower membranes still possessed anAtArf1p homolog but lacked coatomer, and incubation ofthese membranes with concentrated cauliflower cytosol inthe presence of GTPgS led to the binding of coatomer. Thisrecruitment was selective: under the same conditions, theAtSec23/24 dimer homolog of COPII-coated vesicles, alsoshown to be present in cauliflower cytosol (Movafeghi et al.,1999), did not attach. Recruitment was also specific, asjudged by the BFA inhibition of both Arf1p and coatomer re-cruitment.
Why the AtSec23/24p homolog was not recruited is un-clear. Although the cauliflower Golgi fraction used mighthave had insufficient COPII budding sites in the ER mem-branes to support a COPII coat recruitment, we think it morelikely that the explanation involves an aspect of the crucialrole Sar1p plays in the production of COPII-coated vesicles(Schekman and Orci, 1996). As with COPI-coated vesicles,binding of Sar1p precludes the recruitment of the Sec23/24dimer (Barlowe et al., 1994; Aridor et al., 1998); unlike Arf1p,however, Sar1p does not become anchored in the mem-brane and is released after the assembly of the COPII coat
Plant Coatomer 2229
Interchangeability of Coatomer between DifferentPlant Species
The ability to visualize the in vitro–induced COPI-coatedvesicles encouraged us to test whether cauliflowercoatomer could be recruited onto membranes of other ori-gins, such as tobacco, which is more amenable to geneticmanipulation. In addition, to confirm the specificity of the invitro reaction, we tested whether in vitro vesicle buddingwould be dependent on ATP and GTP. We readily estab-lished that this specificity exists and demonstrated that het-erologous recruitment of coatomer to membranes fromanother plant species is possible. In addition, we showedthat retrograde transport markers are enriched in the frac-tion that contains the greatest amounts of recruited Atg-COP. Our results appear to contrast with those obtained byPepperkok et al. (2000), who demonstrated that GTP hydrol-ysis is crucial for cargo loading into COPI-coated vesicles.However, those investigators studied membrane-spanningcargo molecules, whereas we focused on soluble cargomolecules. Nevertheless, whether the cosedimentation ofAtg-COP and soluble retrograde cargo was indeed due tothe presence of cargo in COPI-coated vesicles remains tobe shown. We cannot exclude the possibility that COPI-coated vesicles attached to ER membranes are responsiblefor the density shift. Further work will be necessary to clarifythis point.
Golgi →ER Protein Recycling
We found our coatomer labeling predominated at the cister-nal rims, especially the cis-cisternae, which may beanalogous to the observations of Martinez-Menarguez et al.(1999), who localized COPI coats to the tips of ERGICtubules in pancreatic cells. This might indicate that the
Figure 8. Detection of in Vitro–Formed COPI-Coated Vesicles.
(A) Recruitment of coatomer onto KI-stripped membranes leads toan increase in their equilibrium density. Stripped membranes fromcauliflower inflorescence were incubated in the absence or pres-
ence of cauliflower cytosolic proteins, as described for Figure 6A.The incubation mixtures were then layered onto sucrose densitygradients and centrifuged to equilibrium, as described in Methods.Fractions were screened with antibodies against ER (calnexin, BiP),Golgi (RGP), and COPI/COPII coat proteins (Atg-COP, AtSec23p).As depicted, stripped membranes banded at densities z10% (w/w)less than those at which the recruited membranes did.(B) Isolation of COPI-coated vesicles. KI-stripped cauliflowermembranes were incubated with cauliflower cytosol under stan-dard conditions, extracted with high-salt reagent (250 mM KCl) for30 min at 48C, and centrifuged at 30,000g; the membranes in thesupernatant were subjected to isopcynic sucrose density gradientcentrifugation as in (A). Equal-volume fractions were then moni-tored in protein gel blots with the antisera described above. Theasterisk denotes the putative COPI-coated vesicle–containing frac-tion used for Figure 9.
2230 The Plant Cell
been interpreted as indicating the existence of a highly effi-cient cis-Golgi–based retrieval mechanism for ER residentsand are supported by the present observations, in whicha putative COPI-coated vesicle–containing fraction wasshown to contain calreticulin. Pagny et al. (2000), workingwith suspension-cultured tobacco BY-2 cells, reached adifferent conclusion, however. They claim that normal ER
cis-most elements of the plant Golgi stack are the plant ER-GIC equivalent—a proposal supported by the finding thatsaturation of ER retention by overexpression of the ER resi-dent protein calreticulin does not result in the accumulationof intracellular endoH-resistant forms of calreticulin, eventhough the secreted portion is fully modified by Golgi en-zymes during transit (Crofts et al., 1999). These results have
Figure 9. Immunogold Negative Staining of the Contents of Fraction 12 from the Gradient Shown in Figure 8B.
(A) Overview of normal negative staining. Putative COPI-coated vesicles are indicated with arrowheads, clathrin-coated vesicles with arrows.(B) Comparison of clathrin- and COP-coated vesicles. Arrows and arrowheads are as in (A).(C) to (J) Gallery of negatively stained putative COPI-coated vesicles decorated with gold-coupled Zm«-COP antibodies.Bar in (A) 5 1 mm; bar in (B) 5 100 nm; bar in (J) 5 100 nm for (C) to (J).

Plant Coatomer 2231
residents such as calreticulin are actively excluded from ERexport and are therefore not recycled from the Golgi appa-ratus. On the other hand, foreign proteins expressed with anHDEL C-terminal tetrapeptide do leave the ER and may beretrieved even from later Golgi compartments than the cis-cisternae.
Relevant to this controversy is the subcellular distributionof the plant H/KDEL receptor ERD2p. According to the dataof Boevink et al. (1998), an ERD2-GFP construct in tobaccomesophyll is distributed throughout the Golgi stack and isdetectable in both the central and peripheral portions of thecisternae. Those results were obtained from cells overex-pressing ERD2-GFP, however, and such overexpression orthe attachment of GFP, or both, might influence the localiza-tion of the receptor. Localization of ERD2 in wild-type cellsmay well be different, possibly even restricted to the cis-cis-ternae. In the normal physiological state, recycling solubleER residents by way of the ERD2 pathway may play only aminor role in retention. Hence, retrograde transport of solu-ble cargo in COPI-coated vesicles may be detectable only inartificial situations in which recycling is increased, that is,during ER stress or in transgenic plants overproducing ret-rograde cargo molecules such as a-amylase–HDEL or cal-reticulin.
METHODS
Material
Cauliflower (Brassica oleracea var botrytis) inflorescences (10 to 15cm in diameter) were excised from greenhouse-grown plants, andthe woody stalk tissue was removed. Arabidopsis thaliana var Co-lumbia seeds were surface sterilized, transferred to Erlenmyer flaskscontaining Murashige and Skoog basal medium (no. 5519; Sigma),and shaken at 120 rpm under weak light at 258C. Seven- to 10-day-old seedlings were harvested. After 24 hr of imbibition under runningtap water, maize (Zea mays var mutin) seeds were sown on moist fil-ter paper and incubated at room temperature in the dark for 4 to 6days. Commercially obtained bakers’ yeast (Saccharomyces cere-viseae) was cultured in Erlenmeyer flasks, as described byMovafeghi et al. (1999), and the cells were removed by low-speedcentrifugation. Pig brain was purchased at a local slaughterhouse,transported on ice, frozen in liquid nitrogen, and stored at 2808Cuntil required.
Generation of Recombinant Proteins and Preparation of Antisera
COPI Proteins
The open reading frames of maize d- and «-COPs (coat proteins)(GenBank accession numbers AF216582 and AF216583, respec-tively) were cloned by standard procedures into the NdeI/BamHIsite of the pET-15b vector (Novagen, Madison, WI) and expressed
Figure 10. Heterologous in Vitro Induction of COPI-Coated Vesiclesfrom Cauliflower Cytosol and Tobacco Membranes from Plants Ex-pressing a-Amylase–HDEL.
(A) Incubation performed in the presence (1) of ATP/GTPgS. Thehistogram represents the profile of a-amylase activity. The proteingel blots of the corresponding fractions document the distributionsof calreticulin and Atg-COP in relation to a-amylase activity.(B) Incubation performed in the absence (2) of ATP/GTPgS.All incubations and membrane separations were as given in Fig-ure 8B. Activity measurements were performed in triplicate; errorbars represent the extent of variation.

2232 The Plant Cell
dium B (25 mM Hepes/KOH, pH 7.2, 50 mM KCl, 1 mM DTT, and10% [v/v] glycerin). Subsequently, the protein solution was centri-fuged at 150,000g for 1 hr before subdivision into 2-mL aliquots thatwere then frozen with liquid nitrogen and stored at 2808C before use.Ordinarily, a standard cytosol preparation had a protein concentra-tion of 17 to 20 mg mL21.
Preparation of a Golgi-Enriched Fraction and KI Dissociation of Coat Proteins
Thirty-five grams of cauliflower floret tissue was homogenized in 35mL of prechilled medium A (but at pH 7.5 and minus the KCl andEDTA) by hand-grinding for 10 min with a mortar and pestle. Thehomogenate was passed through two layers of Miracloth and fourlayers of gauze and precentrifuged at 10,000g for 10 min. The super-natant (25 mL) was then layered onto a sucrose step gradient (5 mLof 30% [w/w] sucrose and 5 mL of 35% [w/w] sucrose) made up inmedium C (25 mM Hepes/KOH, pH 7.2, 20 mM KCl). After centrifu-gation at 100,000g for 3 hr in a swinging bucket rotor, the mem-branes collecting at the 30/35% interphase were removed, dilutedwith an equal volume of medium C, and recentrifuged at 70,000g for20 min. (Previous measurements of marker enzymes and proteinshad indicated that cauliflower Golgi membranes equilibrated at su-crose densities between 30 and 35% [Movafeghi et al., 1999].) Mem-brane pellets were resuspended in 2 to 4 mL of 300 mM KI (dissolvedin medium C containing 200 mM sucrose) and rotated for 45 min at48C before being recentrifuged at 70,000g for 20 min. The strippedmembranes were resuspended in medium C. The same procedurewas used to isolate Golgi membranes from tobacco (Nicotiana ta-bacum var SR1) mesophyll, except that 10 mM ascorbic acid, 2 mMsodium metabisulfite, and 1 g L21 polyvinylpyrrolidone were addedto medium C.
Recruitment Assays
A standard recruitment assay contained in a total volume of 4 mL thefollowing components: 0.8 mg of membranes, 24 mg of cytosolicproteins, 50 mM ATP, 250 mM UTP, 20 mM GTPgS, 200 mM DTT, 28units of creatine kinase, 2 mM creatine phosphate, and medium Ccontaining 200 mM sucrose. The recruitment was started by addingthe membranes, and the assay tubes were gently shaken at 208C for25 min. At the end of the recruitment, the contents of each assaytube were layered onto sucrose step gradients (4 mL of 20% [w/w]sucrose and 4 mL of 25% [w/w] sucrose) and centrifuged at100,000g for 3 hr. The soluble overlay and the pellet were separated,and the soluble proteins were recovered by precipitation with chloro-form/methanol according to Wessel and Flügge (1984).
Isolation of COPI-Coated Vesicles
The contents of an 8-mL recruitment mixture (see above) was lay-ered onto two sucrose step gradients (4 mL of recruitment mixtureonto 7 mL of 20% [w/w] sucrose and 1 mL of 55% [w/w] sucrose)and centrifuged at 100,000g for 3 hr. The membranes collecting atthe interface were diluted with an equal volume of a solution of 500mM KCl, 300 mM sucrose, and 25 mM Hepes-KOH, pH 7.2, andshaken for 45 min at 48C. The high-salt mixture was then centrifugedat 30,000g for 30 min, and the supernatant (4 mL) was layered over a
in Escherichia coli (strain BL21DE3; 200-mL cultures, 1 hr, 308C).The His-fusion proteins were recovered in the insoluble pellet. Theinclusion bodies were washed and solubilized as described bySambrook et al. (1989). The recombinant proteins were purified bybinding and eluting from a 1-mL HisBand resin column (Novagen)under denaturing conditions, according to the manufacturer’s in-structions. The identities of the purified proteins were confirmed byN-terminal sequencing after separation by SDS-PAGE and trans-ferring to polyvinylidenedifluoride, as given by Matsudaira (1990).Polyclonal antibodies in rabbits were prepared commercially (Co-vance, Denver, PA).
GTPases
The AtSar1p encoding region (GenBank accession numberM95795) was removed from pJPT1 by cutting with BamHI, subject-ing to Klenow treatment, cutting with BglII, and purifying by agarosegel electrophoresis. pGEX-3X was cut with EcoRI, Klenow treated,and cut with BamHI, and the excised fragment ligated onto it. Thiswas followed by dephosphorylation with calf intestine alkalinephosphatase, yielding plasmid pPP2. To specifically amplify theArf1p coding region (GenBank accession number M95166), we pre-pared total first-strand cDNA from 2-week-old Arabidopsis seed-lings, as described previously (Denecke et al., 1995). To amplifythis cDNA by the polymerase chain reaction, we used GATCAG-GGATCCGGTTGTCATTCGG as the sense primer and GCT-AGAATTCCATCTATGCCTTGCTTGCGAT as antisense primer in 59
to 39 direction. This allowed generation of a BamHI site at the Nterminus and an EcoRI site at the C terminus. The amplified frag-ment obtained was cut with these two restriction enzymes, purifiedon gel, and ligated into pGEX-3x that had been previously cut withBamHI and EcoRI; this yielded pPP4. Production and purificationof recombinant Sar1 and ARF1 recombinant glutathione S-trans-ferase fusion proteins as well as the commercial preparation of an-tisera were performed as previously described (Movafeghi et al.,1999).
Purification of IgGs
IgGs were separated from crude serum on a protein A–SepharoseCL-4B (no. P3391; Sigma) column and eluted with 100 mM glycine-HCl, pH 2.9, as given by Harlow and Lane (1988).
Preparation of a Cytosol Fraction
Five hundred grams of cauliflower floret tissue was homogenized in500 mL of prechilled medium A (25 mM Hepes/KOH, pH 8.0, 300 mMsucrose, 500 mM KCl, 3 mM EDTA, 1 mM DTT, 2 mM o-phenanthro-line, 1.4 mg L21 pepstatin, 0.5 mg L21 leupeptin, 2 mg L21 aprotinin, and1 mg mL21 trans-epoxysuccinyl-L-leucylamido-(4-guanido)-butane[E-64]) by using a Waring Blendor in three 15-sec bursts. The slurrywas then passed through two layers of Miracloth (Calbiochem) andfour layers of gauze. After precentrifugation at 10,000g for 15 min,cell membranes were removed by a further centrifugation at150,000g for 1 hr. Ammonium sulfate (final concentration, 65% satu-rated) was then added to the supernatant, and the precipitated pro-teins were recovered by centrifugation at 10,000g for 15 min. Afterthe pellet was redissolved in 90 mL of medium A, the protein mixturewas subjected to dialysis five times (25 min each) against 2 L of me-

Plant Coatomer 2233
sucrose step gradient (8 mL, consisting of five 1.6-mL steps of 20,30, 40, 50, and 60% [w/w] sucrose dissolved in medium C containing2.5 mM magnesium acetate), which was then centrifuged at100,000g for 10 hr. The 0.5-mL fractions collected were analyzed byprotein gel blots and negative staining (see below).
Gel Electrophoresis, Protein Gel Blotting, andProtein Determination
Membrane and cytosolic proteins were separated on 10 or 12%minigels by standard procedures, electrophoretically transferred tonitrocellulose by using a semidry blotting apparatus, and visualizedwith an ECL kit (Amersham Life Sciences, Braunschweig, Germany).The primary antibodies were used at the following dilutions: Atg-COP, 1:2500; AtSec23p, 1:2500; Zmd-COP, 1:5000; Zm«-COP,1:10,000; AtARF1p, 1:2500; AtSar1p, 1:2500; binding protein (BiP),1:10,000; calnexin, 1:10,000; and RGP, 1:20,000 (supplied by Dr. K.Dhugga, Pioneer Hi-Bred International, Johnston, IA). Bands on theECL film were quantified by using the BASYS gel-analyzing system(BioTec Fisher, Reiskirchen, Germany). Protein concentrations weredetermined according to Bradford (1976).
Cryosectioning and Immunogold Labeling
Root tips 2 mm long were excised, immersed in a primary fixative,and infiltrated with sucrose, as described previously (Hinz et al.,1999). The root tips were then mounted onto specimen stubs, frozenin liquid nitrogen, and cut into ultrathin sections at 21208C with a LeicaUCT microtome (Leica, Bensheim, Germany). Frozen sections werepicked up as described by Liou et al. (1996) and transferred to Form-var-coated nickel grids stabilized with carbon.
Immunogold labeling consisted of the following steps: three 5-minwashes in 20 mM glycine/PBS; blocking for 5 min in 1% (w/v) BSA inPBS; 1-hr incubation at room temperature with primary antibody in1% BSA in PBS; three 5-min washes in 0.1% BSA in PBS; 1-hr incu-bation at room temperature with gold-coupled secondary antibody in1% PBS; one 5-min wash in 0.1% BSA; three 5-min washes in PBS;and five 2-min washes in double-distilled water. This was followed bya staining procedure consisting of a 5-min incubation with uranyl ac-etate/oxalate (in 150 mM oxalic acid, adjusted to pH 7.0 with ammo-nium hydroxide), two 2-min washes in double-distilled water, and a5-min incubation in cold 1.8% (w/v) methyl cellulose containingaqueous 0.4% (w/v) uranyl acetate, pH 4.0.
The primary antibodies and their dilutions were as follows: Atg-COP, 1:200; Zmd-COP, 1:100; Zm«-COP, 1:200; Arf1p, 1:500. Sec-ondary antibodies (goat anti–rabbit IgG) coupled to 10-nm-diameterparticles of gold were obtained from BioCell (Cardiff, UK) and diluted1:30 in PBS containing 1% BSA.
Negative Staining and Immunogold Negative Staining
Negative staining was performed on carbon-coated mica, as de-scribed previously (Robinson et al., 1987). Immunogold negativestaining was performed as described by Drucker et al. (1995). Themembranes in a 10-mL aliquot were allowed to attach to the surfaceof a carbon-coated Formvar grid and were then stabilized by a 10-secfixation in 2.5% glutaraldehyde in phosphate buffer, pH 7.0, before ex-posure to the primary antibody solution (Zm«-COP diluted 1:1000) for1 hr. After washing and blocking, the samples were exposed for 45
min to rabbit IgGs (diluted 1:100) coupled to 10-nm-diameter gold par-ticles before staining for 5 sec in aqueous 3% uranyl acetate.
ACKNOWLEDGMENTS
We thank Christel Rühling for her technical support in cryosectioning.This work was supported by the Deutsche Forschungsgemeinschaft(SFB 523 and GK “Signalvermittelter Transport von Proteinen undVesikeln”) and the Biotechnology and Biological Sciences ResearchCouncil. A.M. was the recipient of a scholarship from the Ministry of Cul-ture and Higher Education of Iran.
Received June 6, 2000; accepted September 8, 2000.
REFERENCES
Andreeva, A.V., Kutuzov, M.A., Evans, D.E., and Hawes, C.R.(1998). The structure and function of the Golgi apparatus: A hun-dred years of questions. J. Exp. Bot. 49, 1281–1291.
Aridor, M., Bannykh, S.I., Rowe, T., and Balch, W.E. (1995).Sequential coupling between COPII and COPI vesicle coats in endo-plasmic reticulum to Golgi transport. J. Cell Biol. 131, 875–893.
Aridor, M., Weissman, J., Bannykh, S.I., Nuoffer, C., and Balch,W.E. (1998). Cargo selection by the COPII budding machineryduring export from the endoplasmic reticulum. J. Cell Biol. 141,61–70.
Bannykh, S.I., and Balch, W.E. (1998). Selective transport of cargobetween the endoplasmic reticulum and Golgi compartments.Histochem. Cell Biol. 109, 463–475.
Barlowe, C., and Schekman, R. (1993). SEC12 encodes a guaninenucleotide exchange factor essential for transport vesicle forma-tion from the ER. Nature 365, 347–349.
Barlowe, C., d’Enfert, C., and Schekman, R. (1993). Purificationand characterization of SAR1p, a small GTP-binding proteinrequired for transport vesicle formation from the endoplasmicreticulum. J. Biol. Chem. 268, 873–879.
Barlowe, C., Orci, L., Yeong, T., Hosobuchi, M., Hamamoto, S.,Salama, N., Rexach, M.F., Ravazzola, M., Amherdt, M., andSchekman, R. (1994). COP II: A membrane coat formed by secproteins that drives vesicle budding from the endoplasmic reticu-lum. Cell 77, 895–907.
Bar-Peled, M., and Raikhel, N.V. (1997). Characterization ofAtSec12 and AtSar1, proteins likely involved in endoplasmic retic-ulum and Golgi transport. Plant Physiol. 114, 315–324.
Boevink, P., Oparka, K., Santa Cruz, S., Martin, B., Betteridge,A., and Hawes, C. (1998). Stacks on tracks: The plant Golgiapparatus traffics on an actin/ER network. Plant J. 15, 441–447.
Bradford, M.M. (1976). A rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principle ofprotein–dye binding. Anal. Biochem. 72, 248–254.
Bremser, M., Nickel, W., Schweikert, M., Ravazzola, M.,Amherdt, M., Hughes, C.A., Söllner, T.H., Rothman, J.E., and

2234 The Plant Cell
Wieland, F.T. (1999). Coupling of coat assembly and vesicle bud-ding to packaging of putative cargo receptors. Cell 96, 495–506.
Chardin, P., Paris, S., Antonny, B., Robineau, S., Béraud-Dufuor,S., Jackson, C.L., and Chabre, M. (1996). A human exchangefactor for ARF contains Sec7- and pleckstrin-homology domains.Nature 384, 481–484.
Cosson, P., and Letourneur, F. (1994). Coatomer interaction withdi-lysine endoplasmic reticulum motifs. Science 263, 1629–1631.
Crofts, A.J., Leborgne-Castel, N., Hillmer, S., Robinson, D.G.,Phillipson, B., Carlsson, L., Ashford, D.A., and Denecke, J.(1999). Saturation of the endoplasmic reticulum retention machin-ery reveals anterograde bulk flow. Plant Cell 11, 2233–2248.
Dascher, C., and Balch, W.E. (1994). Dominant inhibitory mutantsof ARF1 block endoplasmic reticulum to Golgi transport and trig-ger disassembly of the Golgi apparatus. J. Biol. Chem. 269, 1437–1448.
Denecke, J., Carlsson, L., Vidal, S., Ek, B., Höglund, A.-S., VanZeijl, M., Sinjorgo, K.M.C., and Palva, E.T. (1995). The tobaccohomolog of mammalian calreticulin is present in protein com-plexes in vivo. Plant Cell 7, 391–406.
d’Enfert, C., Geusse, M., and Gaillardin, C. (1992). Fission yeastand a plant have functional homologues of the Sar1 and Sec12proteins involved in ER to Golgi traffic in budding yeast. EMBO J.11, 4205–4210.
Drucker, M., Herkt, B., and Robinson, D.G. (1995). Demonstrationof a b-type adaptin at the plant plasma membrane. Cell Biol. Int.19, 191–201.
Duden, R., Griffiths, G., Frank, R., Argos, P., and Kreis, T.E.(1991). b-COP, a 110 kDa protein associated with non-clathrin-coated vesicles and the Golgi complex, shows homology to b-adap-tin. Cell 64, 649–665.
Duden, R., Hosobuchi, M., Hamamoto, S., Winey, M., Byers, B.,and Schekman, R. (1994). Yeast beta- and beta’-coat proteins(COP): Two coatomer subunits essential for endoplasmic reticu-lum–to–Golgi protein traffic. J. Biol. Chem. 269, 24486–24495.
Duden, R., Kajikawa, L., Wuestehube, L., and Schekman, R.(1998). «-COP is a structural component of coatomer that func-tions to stabilize a-COP. EMBO J. 17, 985–995.
Dupree, P., and Sherrier, D.J. (1998). The plant Golgi apparatus.Biochim. Biophys. Acta 1404, 259–270.
Faulstich, D., Auerbach, S., Orci, L., Ravazzola, M., Wegehingel,S., Lottspeich, F., Stenbeck, G., Harter, C., Wieland, F.T., andTschochner, H. (1996). Architecture of coatomer: Molecular char-acterization of delta COP and protein interactions within the com-plex. J. Cell Biol. 135, 53–61.
Goldberg, J. (1999). Structural and functional analysis of the ARF1–ARFGAP complex reveals a role for coatomer in GTP hydrolysis.Cell 96, 893–902.
Griffiths, G., Pepperkok, R., Krijnse-Locker, J., and Kreis, T.E.(1995). Immunocytochemical localization of b-COP to the ER–Golgi boundary and the TGN. J. Cell Sci. 108, 2839–2856.
Hara-Kuge, S., Kuge, O., Orci, L., Amherdt, M., Ravazzola, M.,Wieland, F.T., and Rothman, J.E. (1994). En bloc incorporationof coatomer subunits during the assembly of COP-coated vesi-cles. J. Cell Biol. 124, 883–892.
Harlow, E., and Lane, D. (1988). Antibodies: A Laboratory Manual.
(Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).pp. 309–312.
Harter, C. (1999). COPI proteins: A model for their role in vesiclebudding. Protoplasma 207, 125–132.
Harter, C., and Wieland, F.T. (1998). A single binding site for di-lysine retrieval motifs and p23 within the g-subunit of coatomer.Proc. Natl. Acad. Sci. USA 95, 11649–11654.
Helms, J.B., and Rothman, J.E. (1992). Inhibition by brefeldin A ofa Golgi membrane enzyme that catalyzes exchange of guaninenucleotide bound to ARF. Nature 360, 352–354.
Hinz, G., Menze, A., Hohl, I., and Vaux, D. (1997). Isolation of pro-legumin from developing pea seeds: Its binding to endomem-branes and assembly into prolegumin hexamers in the proteinstorage vacuole. J. Exp. Bot. 48, 139–149.
Hinz, G., Hillmer, S., Bäumer, M., and Hohl, I. (1999). Vacuolarstorage proteins and the putative vacuolar sorting receptor BP-80exit the Golgi apparatus of developing pea cotyledons in differenttransport vesicles. Plant Cell 11, 1509–1524.
Kahn, R.A., Randazzo, P., Serafini, T., Weiss, O., Rulka, C., Clark,J., Amherdt, M., Roller, P., Orci, L., and Rothman, J.E. (1992).The amino terminus of ADP- ribosylation factor (ARF) is a criticaldeterminant of ARF activities and is a potent and specific inhibitorof protein transport. J. Biol. Chem. 267, 13039–13046.
Klausner, R.D., Donaldson, J.G., and Lippincott-Schwartz, J.(1992). Brefeldin A: Insights into the control of membrane trafficand organelle structure. J. Cell Biol. 116, 1071–1080.
Kuge, O., Hara-Kuge, S., Orci, L., Ravazzola, M., Amherdt, M.,Tanigawa, G., Wieland, F.T., and Rothman, J.E. (1993). Zeta-COP, a subunit of coatomer, is required for COP-coated vesicleassembly. J. Cell Biol. 123, 1727–1734.
Lavoie, C., Paiement, J., Dominguez, M., Roy, L., Dahan, S.,Gushue, J.N., and Bergeron, J.J.M. (1999). Roles for a2p24 andCOPI in endoplasmic reticulum cargo exit site formation. J. CellBiol. 146, 285–299.
Letourneur, F., Gaynor, E.C., Henneke, S., Demolliere, C.,Duden, R., Emr, S.D., Riezman, H., and Cosson, P. (1994).Coatomer is essential for retrieval of dilysine-tagged proteins tothe endoplasmic reticulum. Cell 79, 1199–1207.
Liou, W., Geuze, H.J., and Slot, J.W. (1996). Improving structuralintegrity of cryosections for immunogold labeling. Histochem. CellBiol. 106, 41–58.
Lippincott-Schwartz, J., Cole, N.B., and Donaldson, J.G. (1998).Building a secretory apparatus: Role of ARF1/COPI in Golgi bio-genesis and maintenance. Histochem. Cell Biol. 109, 449–462.
Lowe, M., and Kreis, T.E. (1996). In vivo assembly of coatomer, theCOP I coat precursor. J. Biol. Chem. 271, 30725–30730.
Malhotra, V., Serafini, T., Orci, L., Shepherd, J.C., and Rothman,J.E. (1989). Purification of a novel class of coated vesicles medi-ating biosynthetic protein transport through the Golgi stack. Cell58, 329–336.
Malsam, J., Gommel, D., Wieland, F.T., and Nickel, W. (1999). Arole for ADP-ribosylation factor in the control of cargo uptake dur-ing COPI-coated vesicle biogenesis. FEBS Lett. 462, 267–272.
Martinez-Menarguez, J.A., Geuze, H.J., Slot, J.W., andKlumpermann, J.A. (1999). Vesicular tubular clusters betweenthe ER and Golgi mediate concentration of soluble secretory pro-teins by exclusion from COPI-coated vesicles. Cell 98, 81–90.

Plant Coatomer 2235
Matsudaira, P. (1990). Limited N-terminal sequence analysis. Meth-ods Enzymol. 182, 602–613.
Melançon, P., Glick, B.S., Malhotra, V., Weidman, P.J., Serafini,T., Gleason, M.L., Orci, L., and Rothman, J.E. (1987). Involve-ment of GTP-binding “G“ proteins in transport through the Golgistack. Cell 51, 1053–1062.
Mollenhauer, H.H., and Morré, D.J. (1994). Structure of the Golgiapparatus. Protoplasma 180, 14–28.
Movafeghi, A., Happel, N., Pimpl, P., Tai, G.-H., and Robinson,D.G. (1999). Arabidopsis Sec21p and Sec23p homologs. Proba-ble coat proteins of plant COP-coated vesicles. Plant Physiol.119, 1437–1445.
Nickel, W., Malsam, J., Gorgas, K., Ravazzola, M., Jenne, N.,Helms, J.B., and Wieland, F.T. (1998). Uptake by COPI-coatedvesicles of both anterograde and retrograde cargo is inhibited byGTPgS in vitro. J. Cell Sci. 111, 3081–3090.
Oprins, A., Duden, R., Kreis, T.E., Geuze, H.J., and Slot, J.W.(1993). b-COP localizes mainly to the cis-Golgi side in exocrinepancreas. J. Cell Biol. 121, 49–59.
Orci, L., Palmer, D.J., Amherdt, M., and Rothman, J.E. (1993).Coated vesicle assembly in the Golgi requires only coatomer andARF proteins from the cytosol. Nature 364, 732–734.
Orci, L., Perrelet, A., Ravazzola, M., Amherdt, M., Rothman, J.E.,and Schekman, R. (1994). Coatomer-rich endoplasmic reticulum.Proc. Natl. Acad. Sci. USA 88, 8611–8615.
Orci, L., Stamnes, M., Ravazzola, M., Amherdt, M., Perrelet, A.,Söllner, T.H., and Rothman, J.E. (1997). Bidirectional transport bydistinct populations of COPI-coated vesicles. Cell 90, 335–349.
Ostermann, J., Orci, L., Tani, K., Amherdt, M., Ravazzola, M.,Elazar, Z., and Rothman, J.E. (1993). Stepwise assembly offunctionally active transport vesicles. Cell 75, 1015–1025.
Pagny, S., Cabanes-Macheteau, M., Gillikin, J.W., Leborgne-Castel, N., Lerouge, P., Boston, R.S., Faye, L., and Gomord, V.(2000). Protein recycling from the Golgi apparatus to the endo-plasmic reticulum in plants and its minor contribution to calreticu-lin retention. Plant Cell 12, 739–755.
Palmer, D.J., Helms, J.B., Beckers, C.J., Orci, L., and Rothman,J.E. (1993). Binding of coatomer to Golgi membranes requiresADP-ribosylation factor. J. Biol. Chem. 268, 12083–12089.
Pepperkok, R., Whitney, J.A., Gomez, M., and Kreis, T.E. (2000).COPI vesicles accumulating in the presence of a GTP restrictedArf1 mutant are depleted of anterograde and retrograde cargo. J.Cell Sci. 113, 135–144.
Peyroche, A., Paris, S., and Jackson, C.L. (1996). Nucleotideexchange on ARF mediated by yeast Gea1 protein. Nature 384,479–481.
Peyroche, A., Antonny, B., Robineau, S., Acker, J., Cherfils, J.,and Jackson, C.L. (1999). Brefeldin A acts to stabilize an abortiveARF–GDP–Sec7 domain protein complex: Involvement of specificresidues of the Sec7 domain. Mol. Cell 3, 275–285.
Regad, F., Bardet, C., Tremousaygue, D., Moisan, A., Lescure,B., and Axelos, M. (1993). cDNA cloning and expression of anArabidopsis GTP-binding protein of the ARF family. FEBS Lett.316, 133–136.
Robinson, D.G., and Hinz, G. (1997). Vacuolar biogenesis and pro-tein transport to the plant vacuole: A comparison with the yeastvacuole and the mammalian lysosome. Protoplasma 197, 1–25.
Robinson, D.G., and Hinz, G. (1999). Golgi-mediated transport ofseed storage proteins. Seed Sci. Res. 9, 263–283.
Robinson, D.G., Ehlers, U., Herken, R., Herrmann, B., Mayer, F.,and Schürmann, F.-W. (1987). Methods of Preparation for Elec-tron Microscopy. (Berlin: Springer-Verlag). pp. 1–190.
Robinson, D.G., Hinz, G., and Holstein, S.E.H. (1998). The molecularcharacterization of transport vesicles. Plant Mol. Biol. 38, 49–76.
Rothman, J.E., and Wieland, F.T. (1996). Protein sorting by trans-port vesicles. Science 272, 227–234.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). MolecularCloning: A Laboratory Manual. (Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press).
Sanderfoot, A., and Raikhel, N.V. (1999). The specificity of vesicletrafficking: Coat proteins and SNAREs. Plant Cell 11, 629–641.
Satiat-Jeunemaitre, B., Cole, L., Bourett, T., Howard, R., andHawes, C. (1995). Brefeldin A effects in plant and fungal cells:Something new about vesicle trafficking? J. Microsc. 181, 162–177.
Scales, S.J., Pepperkok, R., and Kreis, T.E. (1997). Visualization ofER-to-Golgi transport in living cells reveals a sequential mode ofaction for COPII and COPI. Cell 90, 1137–1148.
Schekman, R., and Orci, L. (1996). Coat proteins and vesicle bud-ding. Science 271, 1526–1533.
Tanigawa, G., Orci, L., Amherdt, M., Ravazzola, M., Helms, J.B.,and Rothman, J.E. (1993). Hydrolysis of bound GTP by ARF pro-tein triggers uncoating of Golgi-derived COP-coated vesicles. J.Cell Biol. 123, 1365–1371.
Tokuyasu, K.T. (1986). Application of cryoultramicrotomy to immu-nocytochemistry. J. Microsc. 143, 139–149.
Torii, S., Banno, T., Watanabe, T., Ikehara, Y., Murakami, K., andNakayama, K. (1995). Cytotoxicity of brefeldin A correlates withits inhibitory effect on membrane binding of COP coat proteins. J.Biol. Chem. 270, 11574–11580.
Vitale, A., and Denecke, J. (1999). The endoplasmic reticulum—Gateway of the secretory pathway. Plant Cell 11, 615–628.
Waters, M.G., Serafini, T., and Rothman, J.E. (1991). “Coatomer”:A cytosolic protein complex containing subunits of non-clathrin-coated Golgi-transport vesicles. Nature 349, 248–251.
Wessel, D., and Flügge, U.I. (1984). A method for the quantitativerecovery of proteins in dilute solutions in the presence of deter-gents and lipids. Anal. Biochem. 138, 141–143.

DOI 10.1105/tpc.12.11.2219 2000;12;2219-2235Plant Cell
Peter Pimpl, Ali Movafeghi, Sean Coughlan, Jürgen Denecke, Stefan Hillmer and David G. RobinsonIn Situ Localization and in Vitro Induction of Plant COPI-Coated Vesicles
This information is current as of October 6, 2018
References /content/12/11/2219.full.html#ref-list-1
This article cites 70 articles, 33 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists